

Abstract
Parkinson’s disease is the second most common neurodegenerative disorder; it affects 1% of the population over the age of 65. The number of people with Parkinson’s disease is set to rapidly increase due to changing demographics and there is an unmet clinical need for disease-modifying therapies. The pathological hallmarks of Parkinson’s disease are the progressive degeneration of dopaminergic neurons in the substantia nigra and their axons which project to the striatum, and the aggregation of α-synuclein; these result in a range of debilitating motor and non-motor symptoms. The application of neurotrophic factors to protect and potentially regenerate the remaining dopaminergic neurons is a major area of research interest. However, this strategy has had limited success to date. Clinical trials of two well-known neurotrophic factors, glial cell line-derived neurotrophic factor and neurturin, have reported limited efficacy in Parkinson’s disease patients, despite these factors showing potent neurotrophic actions in animal studies. There is therefore a need to identify other neurotrophic factors that can protect against α-synuclein-induced degeneration of dopaminergic neurons. The bone morphogenetic protein (BMP) family is the largest subgroup of the transforming growth factor-β superfamily of proteins. BMPs are naturally secreted proteins that play crucial roles throughout the developing nervous system. Importantly, many BMPs have been shown to be potent neurotrophic factors for dopaminergic neurons. Here we discuss recent work showing that transcripts for the BMP receptors and BMP2 are co-expressed with several key markers of dopaminergic neurons in the human substantia nigra, and evidence for downregulation of BMP2 expression at distinct stages of Parkinson’s disease. We also discuss studies that explored the effects of BMP2 treatment, in in vitro and in vivo models of Parkinson’s disease. These studies found potent effects of BMP2 on dopaminergic neurites, which is important given that axon degeneration is increasingly recognized as a key early event in Parkinson’s disease. Thus, the aim of this mini-review is to give an overview of the BMP family and the BMP-Smad signalling pathway, in addition to reviewing the available evidence demonstrating the potential of BMP2 for Parkinson’s disease therapy.
Keywords: axon growth, BMP2, dopaminergic neurons, neuroprotection, neurotrophic factor, neuroregeneration, Parkinson's disease, a-synuclein
Introduction
Parkinson’s disease (PD) is a common neurodegenerative disorder, the pathological hallmarks of which are the progressive degeneration of nigrostriatal midbrain dopaminergic (mDA) neurons and intraneuronal inclusions of α-synuclein, known as Lewy bodies and Lewy neurites (Spillantini et al., 1997; O’Keeffe and Sullivan, 2018). There is increasing evidence that axonal degeneration, potentially mediated by α-synuclein, may be a key early event in the pathogenesis of PD (O’Keeffe and Sullivan, 2018). Since there is a lack of disease-modifying therapies for PD, two experimental therapies focusing on neuroprotection and neurorepair have been trialed. These are: (i) administration of neurotrophic factors to protect the remaining mDA neurons from degeneration, and (ii) grafting of fetal midbrain neurons to replace the mDA neurons that have been lost (Hegarty et al., 2014a). For the purpose of this review, we will focus on the former.
Search Strategy and Selection Criteria
For this review we searched NCBI and Science Direct for literature published from inception to September 28, 2019 for articles referencing one or more of the following: BMP2; BMP; Neurotrophic Factor; Dopaminergic neurons; Parkinson’s disease.
The Bone Morphogenetic Protein Family
Neurotrophic factors (NTFs) are small secreted proteins that regulate the development, survival, growth, differentiation and maturation of discrete neuronal populations (Sullivan and O’Keeffe, 2016). In PD, two of the most widely studied dopaminergic NTFs are glial cell line-derived neurotrophic factor (GDNF) and neurturin, both of which are members of the transforming growth factor-β (TGF-β) superfamily of cytokines. GDNF and neurturin showed potent neurotrophic effects in in vitro and in vivo models of PD, which led to their subsequent clinical testing in PD patients (Nutt et al., 2003; Patel et al., 2005; Slevin et al., 2005; Lang et al., 2006; Decressac et al., 2011; Warren Olanow et al., 2015; Whone et al., 2019a, b). However, these clinical trials have thus far failed to meet their primary endpoints. Consequently, there is an ongoing need to identify alternative NTFs for dopaminergic neuroprotection in PD. Bone morphogenetic proteins (BMPs) are being increasingly studied in this regard.
The BMP family is the largest subgroup of the TGF-β superfamily of proteins and is comprised of at least 20 growth factors, including the growth and differentiation factor (GDF) family (Kawabata et al., 1998). BMPs were first identified by their presence in extracts of demineralized bone (Wozney, 1992) and have since been shown to induce a sequential developmental cascade of cartilage and bone formation, as well as playing roles in various other developmental processes (Chen et al., 2004). Experimental evidence has demonstrated that BMPs are involved in many biological processes across various cell types, including monocytes, epithelial cells, mesenchymal cells, and importantly, neuronal cells (Miyazono and Shimanuki, 2008; Wang et al., 2014). Within these cells, BMPs are known to regulate growth, differentiation, chemotaxis and apoptosis, while also playing vital roles in the morphogenesis of almost all organs and tissues throughout embryogenesis (Hogan, 1996; Wang et al., 2014). Additionally, BMPs are crucial regulators of axonal growth in many distinct neuronal populations (Gratacos et al., 2002; Parikh et al., 2011; Hegarty et al., 2013a).
Bone Morphogenetic Protein-Smad Signaling
BMPs signal through a canonical pathway that involves transcription factors known as Smads. In this pathway, BMPs bind to a heterotetrameric complex of cell surface transmembrane type I and type II serine/threonine kinase receptors, causing the subsequent activation of Smad signaling (Weiss and Attisano, 2013) (Figure 1). Depending on the cellular context, BMP signaling can also occur via non-canonical pathways involving the activation of many intracellular pathways, including GTPases, mitogen activated protein kinase, and phosphoinositide 3-kinase pathways (Zhang, 2009). To date, there are seven identified type I receptors, the activin receptor-like kinases (ALK) 1–7, and five type II receptors (Hegarty et al., 2013a). There are eight distinct Smad proteins, which constitute three separate functional groupings. These include the receptor-regulated Smads (R-Smads)-1, -2, -3, -5, and -8/9, the common-mediator Smad (Co-Smad) 4 and the inhibitory Smads (I-Smads) 6 and 7 (Wrana and Attisano, 2000). Following ligand binding to the heterotetrameric receptor complex, which is composed of a type I and type II receptor dimer, the constitutively active type II receptors intracellularly transphosphorylate the type I receptors at serine and threonine residues via a highly conserved glycine/serine domain (Wrana and Attisano, 2000). The phosphorylation of type I receptors leads to the recruitment and subsequent phosphorylation of the R-Smads, which can then form a heterotrimer complex with Co-Smad 4. The I-Smads function as major negative regulators of BMP-Smad signaling and compete with the R-Smads for both receptor binding and Co-Smad 4 binding. The I-Smads also target the receptors for degradation via the proteasome (Shi and Massague, 2003). The signaling pathway culminates with the R-Smads and Co-Smad 4 complex translocating to the nucleus, where it binds to DNA and regulates the transcription of target genes (Wrana and Attisano, 2000). However, although there are many ligands of the TGF-β superfamily, there are very few receptors available to accommodate ligand binding. Thus, the type I and type II receptors are required to bind to more than one ligand type (Shi and Massague, 2003). There are two known type I BMP receptors (BMPRs), BMPR1A (also known as ALK3) and BMPR1B (also known as ALK6); however, BMPs are also capable of binding to ALK1 and 2. Furthermore, BMPs can bind to three type II receptors: BMPR2, activin type 2A receptor and activin type 2B receptor (Hegarty et al., 2013a). Recent studies have identified dynamin-dependent endocytosis to be a crucial regulator of the BMPRs at the surface of the cell membrane and to play a central role in the activation, temporal kinetics and magnitude of BMP signal transduction (Hegarty et al., 2017a).
Figure 1.
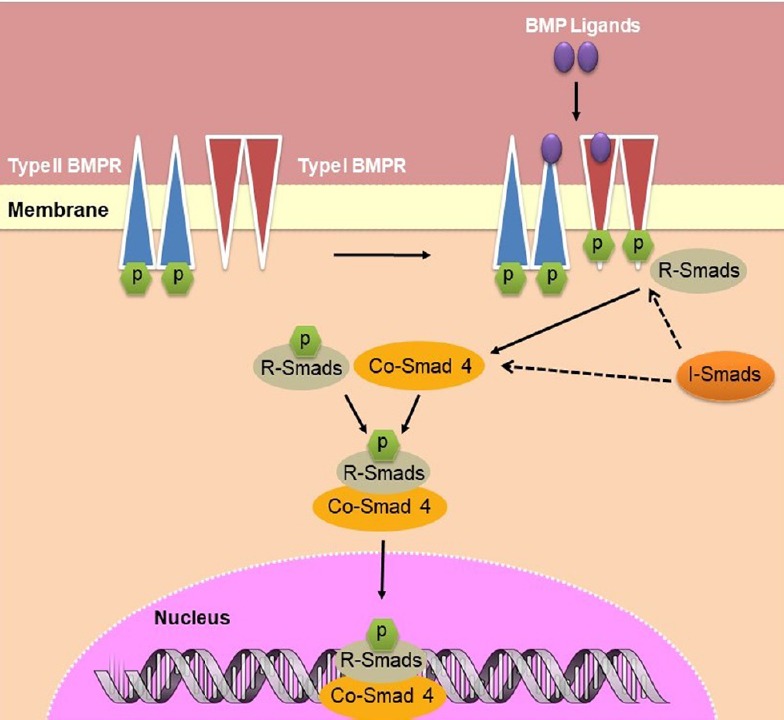
Schematic of BMP-Smad signalling from cell membrane to nucleus.
BMP ligands bind to a heterotetrameric complex of type I and type II serine/threonine kinase cell surface receptors, causing the constitutively active type II receptor to intracellularly phosphorylate the type I receptor. R-Smads and I-Smads compete for binding at the receptor complex for phosphorylation and subsequent complex formation with Co-Smad 4. This complex translocates to the nucleus, where it binds to the DNA and alters transcription. BMP: Bone morphogenetic protein. BMPR: BMP receptor; Co-Smad 4: common-mediator Smad 4; I-Smads: inhibitory Smads; R-Smads: receptor-regulated Smads.
The type I and II BMPRs can form the heterotetrameric complex required for Smad activation in various ways; consequently, BMP ligands have varying affinities for these combinations. For example, BMP2 binds to BMPR1A and BMPR1B with a higher affinity than it does to BMPR2. Similarly, GDF5 binds to BMPR2 and BMPR1B with greater affinity than to BMPR1A. In fact, the affinity of GDF5 for BMPR1B is at least 15-fold higher than it is for BMPR1A, signifying the fundamental importance of BMPR1B for GDF5 signaling (Nickel et al., 2005). Such varying affinities for multiple receptor complexes is relevant when studying the effects of potential NTFs in different cell lines. In addition, various studies on the structural and functional features of the BMPRs have highlighted a cytoplasmic loop segment, known as the L45 loop, located adjacent to the glycine/serine domain on type I receptors, which is involved in R-Smad binding. A corresponding loop, the L3 loop, is present on the R-Smads and is involved in determining the specific Smad substrates for the type I receptor kinases. Smads 1, 5 and 8 are the pivotal effectors that are primarily linked to members of the BMP/GDF subfamily, while Smad 2 and 3 are typically associated with TGF-βs, Activins and Nodals (Mueller and Nickel, 2012).
Bone Morphogenetic Protein Expression in the Nervous System
Several studies have characterized the temporal expression profiles of the various BMPs and their receptors in the developing nervous system. BMPR1 and BMPR2 are stably expressed in the brain throughout adulthood, in several regions including the cortex, striatum, hippocampus and substantia nigra (SN) (Chen et al., 2003; Miyagi et al., 2011). The expression of BMPR1B and BMPR2 receptors in the rat ventral mesencephalon (VM) is present from embryonic day (E) 14 onwards, after which it is continuous through to at least post-natal day (P) 90 (for review see Hegarty et al., 2014b). Similarly, BMPR1B and BMPR2 transcripts are expressed in the mouse VM from E12 onwards (Hegarty et al., 2017b). Notably, this BMPR temporal expression profile parallels the growth of nigrostriatal dopaminergic projections. While multiple BMPs have been examined as potential NTFs for mDA neurons (for review see Hegarty et al., 2014b), given recent data showing that BMP2 is expressed in the human SN, and that it can protect against neurotoxin and α-synuclein-induced degeneration (Goulding et al., 2019), we will focus on BMP2 for the purposes of this article.
Neurotrophic Effects of Bone Morphogenetic Protein 2 in Primary Cultures of Rodent Midbrain
Within the BMP family, BMP2 is one of the most extensively studied BMPs. An early paper reported that recombinant human (rh)BMP2 exerted neurotrophic effects on cultured E13–15 rat VM mDA neurons (Jordan et al., 1997). Specifically, treatment with 10 ng/mL rhBMP2 increased the survival of tyrosine hydroxylase-positive (TH+) mDA neurons, by 1.5-fold compared to that in controls, after 8 days in vitro (DIV). The same study demonstrated that BMP2 treatment of mDA cultures resulted in increased numbers of 5-bromodeoxyuridine-positive and glial fibrillary acidic protein-positive cells, markers of cell proliferation and astrocytes, respectively. These early findings indicated not only does BMP2 promote the survival of mDA neurons, but that it also appears to play a selective role in the proliferation and differentiation of neural progenitors.
A further study also demonstrated positive effects of BMP2 treatment on mDA neurons in E14 VM cultures. Specifically, cultures treated with BMP2 displayed a concentration-dependent increase in the number of mDA neurons, reaching a maximum increase of 237% after treatment with 10 ng/mL, compared to vehicle-treated controls after 7 DIV (Reiriz et al., 1999). In addition, BMP2 treatment increased the number of primary neurites and neurite branching in TH+ cells in a concentration-dependent manner, reaching a maximal effect with 1 ng/mL rhBMP2 (Reiriz et al., 1999). Similarly, other studies have shown a significant increase in the neurite length of TH+ neurons in E14 rat VM primary cultures after treatment with 50 ng/mL or 200 ng/mL rhBMP2 (Hegarty et al., 2014b; Goulding et al., 2019). One study investigated whether BMP2-induced effects were mediated by astroglial cells and reported that addition of 10 ng/mL BMP2 had no effect on the number or morphology of astroglial cells in E14 VM cultures (Reiriz et al, 1999). They further showed that BMP2-induced increases in TH+ cell numbers were not altered in the presence of 30 µM α-aminoadipic acid, a gliotoxin that eliminates glial fibrillary acidic protein-immunoreactive astrocytes (Reiriz et al., 1999). Interestingly however, this study did report that treatment with 100 ng/mL BMP2 led to significant increases in both the number and morphology of astrocytes (Reiriz et al., 1999). These findings suggest that, at low doses, BMP2 treatment promotes both the number and morphological differentiation of mDA neurons, while at higher doses, it also influences the number and phenotype of astroglial cells (Reiriz et al., 1999).
Several studies have reported that the neurotrophic effects of BMP2 and BMP signalling often occur in tandem with, and are amplified by, other NTFs. For example, BMP2-induced signalling in E19 rat striatal neurons was found to be facilitated by co-administration with either BDNF or neurotrophin-3 (Gratacos et al., 2001). However, there is limited data available on the interactions of BMPs with other NTFs in the context of dopaminergic neurons, and this will be an important point to consider in future research.
Neurotrophic Effects of Bone Morphogenetic Protein 2 in Cell Line Models of Human Dopaminergic Neurons
In addition to the neurotrophic effects of BMP2 reported in studies on primary VM cultures, similar effects have been demonstrated in SH-SY5Y cells, a cell line which, despite some limitations, is a widely-used in vitro model of human mDA neurons (Xie et al., 2010; Hegarty et al., 2016; Xicoy et al., 2017; Goulding et al., 2019). Specifically, SH-SY5Y cells treated with 50 ng/mL or 200 ng/mL rhBMP2 for 3 or 4 DIV respectively, displayed an increase in total neurite length (Hegarty et al., 2013b; Goulding et al., 2019). This supported the previous studies which showed that BMP2 treatment induced neuronal differentiation of mDA neurons (Reiriz et al., 1999; Hegarty et al., 2014b; Goulding et al., 2019). Treatment with BMP2 was also found to activate the canonical Smad 1/5/8 signaling pathway in SH-SY5Y cells to induce neurite growth (Hegarty et al., 2013b). A causal link between neurite growth and BMP signaling was demonstrated when it was shown that pretreatment with dorsomorphin, a selective small molecule inhibitor of BMPR1 receptors (Yu et al., 2008; Hegarty et al., 2013b), completely prevented BMP2-induced differentiation of SHSY5Y cells (Hegarty et al., 2013b). In addition, dorsomorphin treatment completely blocked BMP2-induced Smad activation. Similar experiments on E14 VM cultures showed that dorsomorphin blocked both morphological differentiation of mDA neurons and activation of the Smad pathway, which had been induced by BMP2 treatment (Hegarty et al., 2013b, 2014b). Collectively, these studies show that the phenotypic effects exerted by BMP2 are directly dependent on the activation of the Smad 1/5/8 signaling pathway in mDA neurons (Hegarty et al., 2013b, 2014b).
Expression of Bone Morphogenetic Protein 2 and Its Receptors in the Human Substantia Nigra
We have recently shown, using unbiased co-expression analysis, that the expression of BMP2 and of its receptors is positively correlated with the expression of five key dopaminergic markers in the human SN, that is ALDH1A1, Nurr1, Girk2, TH and LMX1B (Goulding et al., 2019). Furthermore, we have shown that BMP2 is expressed at significantly higher levels in the SN than in the putamen or the cerebellum, areas that are known to be especially responsive to BMPs. Interestingly, we have also analyzed open source gene expression data from SN of age- and gender-matched samples at various Braak stages of PD and found that transcripts for BMP2 were significantly downregulated at later stages (Goulding et al., 2019). These data suggest a functional role for both BMP2 and the BMPRs in dopaminergic neurons within the human SN.
Protective Effects of Bone Morphogenetic Protein 2 against Neurotoxin- and α-Synuclein-Induced Neurodegeneration In Vitro
On investigating the effects of rhBMP2 in cellular in vitro models of PD, we found that treatment with 50 ng/mL rhBMP2 for 72 hours increased neurite length by 1.5-fold in SH-SY5Y cells and 1.3-fold in E14 VM cultures, compared to controls (Figure 2). Additionally, BMP2 treatment protected the survival of TH+ neurons in these mDA neuronal cultures against reductions induced by either 1-methyl-4-phenylpyridinium iodide or 6-hydroxydopamine (6-OHDA). Furthermore, treatment with 50 ng/mL rhBMP2 resulted in significant increases in both neurite length and somal area, in cultures treated with either 1-methyl-4-phenylpyridinium iodide or 6-OHDA for 72 hours (Goulding et al., 2019). Additionally, treatment with 50 ng/mL rhBMP2 protected against wild-type-α-synuclein- and mutant A53T-α-synuclein-induced neurite degeneration in both SH-SY5Y cells and in mDA neurons after 3 DIV (Goulding et al., 2019). This is important given that neurite degeneration is an early pathological feature of PD pathogenesis, which may be as a result of α-synuclein accumulation (O’Keeffe and Sullivan, 2018).
Figure 2.
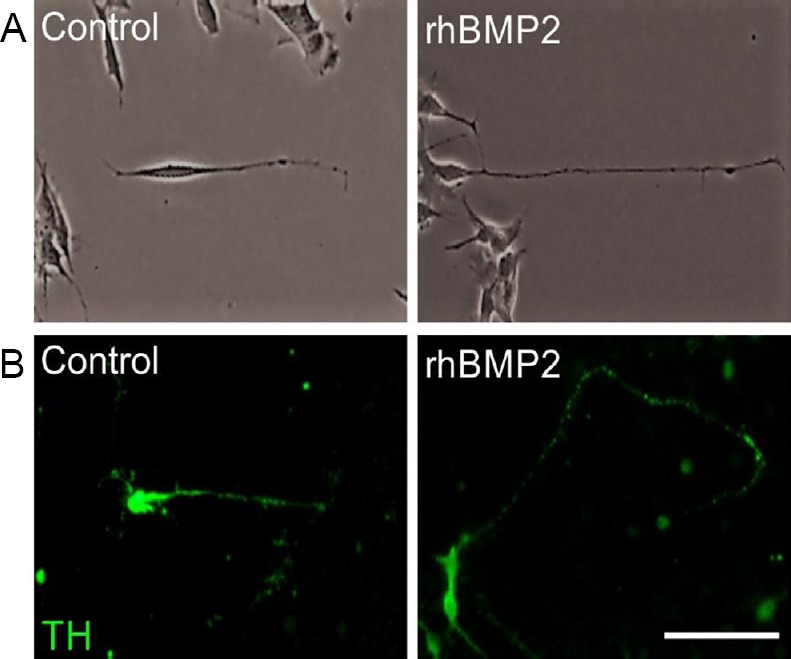
rhBMP2 promotes neurite length in SH-SY5Y cells and in primary cultures of dopaminergic neurons.
(A) SH-SY5Y cells and (B) TH+ dopaminergic neurons in primary cultures of E14 rat VM, after treatment with 50 ng/mL rhBMP2 for 72 hours compared to untreated controls. Scale bar: 50 µm. rhBMP2: Recombinant human bone morphogenetic protein 2; TH+: tyrosine hydroxylase-positive; VM: ventral mesencephalon.
Neurotrophic effects of Bone Morphogenetic Protein 2 In Vivo
To support the positive neuroprotective effects of BMP2 seen in the in vitro studies, there are some promising data regarding its in vivo effects. One study reported that pretreatment with 50 ng/mL rhBMP2 could maintain cell viability in cultures of E14 VM for up to 6 hours, compared to untreated control cultures which demonstrated a 25% decrease in cell viability over the same period (Espejo et al., 1999). A subsequent in vivo study showed that treatment of E14 rat VM with 50 ng/mL rhBMP2 prior to grafting into the striatum of 6-OHDA-lesioned rats resulted in a significant increase in the survival of the grafted dopaminergic neurons (Espejo et al., 1999). This neuroprotective effect of BMP2 pretreatment in vivo was substantial at 1 week after grafting, and remained, although at a lower level, after 4 weeks (Espejo et al., 1999). After 1 week, the rats that had received BMP2-treated VM grafts showed no difference in motor impairment, as measured by apomorphine-induced rotations, than those that had received untreated VM grafts; however after four weeks, rats with BMP2-treated VM grafts displayed significant reductions in rotations, compared to those with untreated grafts. Furthermore, the morphology of the grafted dopaminergic neurons was notably more differentiated in the rhBMP2 pre-treated group than in the untreated graft group at 4 weeks post-grafting (Espejo et al., 1999). In light of these studies showing the in vivo potential of BMP2 treatment, it will be important to test both recombinant BMP2 and viral vectors carrying the BMP2 transgene in neurotoxin and α-synuclein in vivo models of PD.
As well as the neurotrophic effects of BMP2 described above, another study has highlighted the potential of BMP2 to induce differentiation of neural stem cells (NSCs) in vitro (Yan et al., 2016). This is an important finding, given the low survival rate, and considering the ethical and logistical difficulties involved, of human fetal VM tissue grafts for PD treatment (Olanow et al., 2003; Ishii and Eto, 2014). Specifically, Yan et al. (2016) employed five doses of BMP2 (that is 0.1, 1, 10, 20 and 100 ng/mL) over a 14-day period, to treat NSCs harvested from E14 VM cells. This study found that 20 and 100 ng/mL doses of BMP2 induced the highest increases in the number of dopaminergic neurons, while also increasing their neurite length (Yan et al., 2016). This effect of BMP2 on NSC differentiation was induced via miR-145-mediated upregulation of Nurr1, a key player in the maintenance of DA neurons (Yan et al., 2016). NSCs offer a viable therapeutic approach to PD therapy as the cells are self-renewing and can be differentiated into all neural lineage cells, therefore showing promising potential to be used for replacement of those mDA neurons that are lost from the PD brain.
Conclusion
Here we have reviewed the available evidence that supports BMP2 as a factor with potential to be used in neurotrophic therapy for PD. It will be important to determine whether this BMP ligand is effective in protecting dopaminergic neurons against α-synuclein-induced degeneration in vivo, in translational models of PD, such as the α-synuclein rat model, as the next stage in rationalizing its further study as a potential therapeutic agent for PD.
Footnotes
Conflicts of interest: The authors declare no conflicts of interest.
Financial support: This work was supported by a RISAM PhD Scholarship from Cork Institute of Technology (R00094948) and a research grant from Science Foundation Ireland (SFI) under the grant number 15/CDA/3498 (to GWOK).
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Funding: This work was supported by a RISAM PhD Scholarship from Cork Institute of Technology (R00094948) and a research grant from Science Foundation Ireland (SFI) under the grant number 15/CDA/3498 (to GWOK).
C-Editors: Zhao M, Li JY; T-Editor: Jia Y
References
- 1.Chen D, Zhao M, Mundy GR. Bone morphogenetic proteins. Growth Factors. 2004;22:233–241. doi: 10.1080/08977190412331279890. [DOI] [PubMed] [Google Scholar]
- 2.Chen HL, Lein PJ, Wang JY, Gash D, Hoffer BJ, Chiang YH. Expression of bone morphogenetic proteins in the brain during normal aging and in 6-hydroxydopamine-lesioned animals. Brain Res. 2003;994:81–90. doi: 10.1016/j.brainres.2003.09.020. [DOI] [PubMed] [Google Scholar]
- 3.Decressac M, Ulusoy A, Mattsson B, Georgievska B, Romero-Ramos M, Kirik D, Bjorklund A. GDNF fails to exert neuroprotection in a rat alpha-synuclein model of Parkinson’s disease. Brain. 2011;134:2302–2311. doi: 10.1093/brain/awr149. [DOI] [PubMed] [Google Scholar]
- 4.Espejo M, Cutillas B, Ventura F, Ambrosio S. Exposure of foetal mesencephalic cells to bone morphogenetic protein-2 enhances the survival of dopaminergic neurones in rat striatal grafts. Neurosci Lett. 1999;275:13–16. doi: 10.1016/s0304-3940(99)00708-9. [DOI] [PubMed] [Google Scholar]
- 5.Goulding SR, Sullivan AM, O’Keeffe GW, Collins LM. Gene co-expression analysis of the human substantia nigra identifies BMP2 as a neurotrophic factor that can promote neurite growth in cells overexpressing wild-type or A53T α-synuclein. Parkinsonism Relat Disord. 2019;64:194–201. doi: 10.1016/j.parkreldis.2019.04.008. [DOI] [PubMed] [Google Scholar]
- 6.Gratacos E, Checa N, Perez-Navarro E, Alberch J. Brain-derived neurotrophic factor (BDNF) mediates bone morphogenetic protein-2 (BMP-2) effects on cultured striatal neurones. J Neurochem. 2001;79:747–755. doi: 10.1046/j.1471-4159.2001.00570.x. [DOI] [PubMed] [Google Scholar]
- 7.Gratacos E, Gavalda N, Alberch J. Bone morphogenetic protein-6 is a neurotrophic factor for calbindin-positive striatal neurons. J Neurosci Res. 2002;70:638–644. doi: 10.1002/jnr.10438. [DOI] [PubMed] [Google Scholar]
- 8.Hegarty SV, Collins LM, Gavin AM, Roche SL, Wyatt SL, Sullivan AM, O’Keeffe GW. Canonical BMP-Smad signalling promotes neurite growth in rat midbrain dopaminergic neurons. Neuromolecular Med. 2014a;16:473–489. doi: 10.1007/s12017-014-8299-5. [DOI] [PubMed] [Google Scholar]
- 9.Hegarty SV, O’Keeffe GW, Sullivan AM. BMP-Smad 1/5/8 signalling in the development of the nervous system. Prog Neurobiol. 2013a;109:28–41. doi: 10.1016/j.pneurobio.2013.07.002. [DOI] [PubMed] [Google Scholar]
- 10.Hegarty SV, O’Keeffe GW, Sullivan AM. Neurotrophic factors: from neurodevelopmental regulators to novel therapies for Parkinson’s disease. Neural Regen Res. 2014b;9:1708–1711. doi: 10.4103/1673-5374.143410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hegarty SV, Sullivan AM, O’Keeffe GW. BMP2 and GDF5 induce neuronal differentiation through a Smad dependant pathway in a model of human midbrain dopaminergic neurons. Mol Cell Neurosci. 2013b;56:263–271. doi: 10.1016/j.mcn.2013.06.006. [DOI] [PubMed] [Google Scholar]
- 12.Hegarty SV, Sullivan AM, O’Keeffe GW. Protocol for evaluation of neurotrophic strategies in Parkinson’s disease-related dopaminergic and sympathetic neurons in vitro. J Biol Methods. 2016;3:e50. doi: 10.14440/jbm.2016.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hegarty SV, Sullivan AM, O’Keeffe GW. Endocytosis contributes to BMP2-induced Smad signalling and neuronal growth. Neurosci Lett. 2017a;643:32–37. doi: 10.1016/j.neulet.2017.02.013. [DOI] [PubMed] [Google Scholar]
- 14.Hegarty SV, Wyatt SL, Howard L, Stappers E, Huylebroeck D, Sullivan AM, O’Keeffe GW. Zeb2 is a negative regulator of midbrain dopaminergic axon growth and target innervation. Sci Rep. 2017b;7:8568. doi: 10.1038/s41598-017-08900-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hogan BL. Bone morphogenetic proteins: multifunctional regulators of vertebrate development. Genes Dev. 1996;10:1580–1594. doi: 10.1101/gad.10.13.1580. [DOI] [PubMed] [Google Scholar]
- 16.Ishii T, Eto K. Fetal stem cell transplantation: Past present and future. World J Stem Cells. 2014;6:404–420. doi: 10.4252/wjsc.v6.i4.404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jordan J, Bottner M, Schluesener HJ, Unsicker K, Krieglstein K. Bone morphogenetic proteins: neurotrophic roles for midbrain dopaminergic neurons and implications of astroglial cells. Eur J Neurosci. 1997;9:1699–1709. doi: 10.1111/j.1460-9568.1997.tb01527.x. [DOI] [PubMed] [Google Scholar]
- 18.Kawabata M, Imamura T, Miyazono K. Signal transduction by bone morphogenetic proteins. Cytokine Growth Factor Rev. 1998;9:49–61. doi: 10.1016/s1359-6101(97)00036-1. [DOI] [PubMed] [Google Scholar]
- 19.Lang AE, Gill S, Patel NK, Lozano A, Nutt JG, Penn R, Brooks DJ, Hotton G, Moro E, Heywood P, Brodsky MA, Burchiel K, Kelly P, Dalvi A, Scott B, Stacy M, Turner D, Wooten VG, Elias WJ, Laws ER, et al. Randomized controlled trial of intraputamenal glial cell line-derived neurotrophic factor infusion in Parkinson disease. Ann Neurol. 2006;59:459–466. doi: 10.1002/ana.20737. [DOI] [PubMed] [Google Scholar]
- 20.Miyagi M, Mikawa S, Hasegawa T, Kobayashi S, Matsuyama Y, Sato K. Bone morphogenetic protein receptor expressions in the adult rat brain. Neuroscience. 2011;176:93–109. doi: 10.1016/j.neuroscience.2010.12.027. [DOI] [PubMed] [Google Scholar]
- 21.Miyazono K, Shimanuki T. Bone morphogenetic protein receptors and actions A2 - Bilezikian, John P. In: Raisz LG, Martin TJ, editors. Principles of bone biology. Third Edition. San Diego: Academic Press; 2008. pp. 1177–1196. [Google Scholar]
- 22.Mueller TD, Nickel J. Promiscuity and specificity in BMP receptor activation. FEBS Lett. 2012;586:1846–1859. doi: 10.1016/j.febslet.2012.02.043. [DOI] [PubMed] [Google Scholar]
- 23.Nickel J, Kotzsch A, Sebald W, Mueller TD. A single residue of GDF-5 defines binding specificity to BMP receptor IB. J Mol Biol. 2005;349:933–947. doi: 10.1016/j.jmb.2005.04.015. [DOI] [PubMed] [Google Scholar]
- 24.Nutt JG, Burchiel KJ, Comella CL, Jankovic J, Lang AE, Laws ER, Jr, Lozano AM, Penn RD, Simpson RK, Jr, Stacy M, Wooten GF. factor IGSGIiGcl-dn (2003) Randomized, double-blind trial of glial cell line-derived neurotrophic factor (GDNF) in PD. Neurology. 60:69–73. doi: 10.1212/wnl.60.1.69. [DOI] [PubMed] [Google Scholar]
- 25.O’Keeffe GW, Sullivan AM. Evidence for dopaminergic axonal degeneration as an early pathological process in Parkinson’s disease. Parkinsonism Relat Disord. 2018;56:9–15. doi: 10.1016/j.parkreldis.2018.06.025. [DOI] [PubMed] [Google Scholar]
- 26.Olanow CW, Goetz CG, Kordower JH, Stoessl AJ, Sossi V, Brin MF, Shannon KM, Nauert GM, Perl DP, Godb J, Freeman TB. A double-blind controlled trial of bilateral fetal nigral transplantation in Parkinson’s disease. Ann Neurol. 2003;54:403–414. doi: 10.1002/ana.10720. [DOI] [PubMed] [Google Scholar]
- 27.Parikh P, Hao Y, Hosseinkhani M, Patil SB, Huntley GW, Tessier-Lavigne M, Zou H. Regeneration of axons in injured spinal cord by activation of bone morphogenetic protein/Smad1 signaling pathway in adult neurons. Proc Natl Acad Sci U S A. 2011;108:E99–107. doi: 10.1073/pnas.1100426108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Patel NK, Bunnage M, Plaha P, Svendsen CN, Heywood P, Gill SS. Intraputamenal infusion of glial cell line–derived neurotrophic factor in PD: A two-year outcome study. Ann Neurol. 2005;57:298–302. doi: 10.1002/ana.20374. [DOI] [PubMed] [Google Scholar]
- 29.Reiriz J, Espejo M, Ventura F, Ambrosio S, Alberch J. Bone morphogenetic protein-2 promotes dissociated effects on the number and differentiation of cultured ventral mesencephalic dopaminergic neurons. J Neurobiol. 1999;38:161–170. [PubMed] [Google Scholar]
- 30.Shi Y, Massague J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. 2003;113:685–700. doi: 10.1016/s0092-8674(03)00432-x. [DOI] [PubMed] [Google Scholar]
- 31.Slevin JT, Gerhardt GA, Smith CD, Gash DM, Kryscio R, Young B. Improvement of bilateral motor functions in patients with Parkinson disease through the unilateral intraputaminal infusion of glial cell line-derived neurotrophic factor. J Neurosurg. 2005;102:216–222. doi: 10.3171/jns.2005.102.2.0216. [DOI] [PubMed] [Google Scholar]
- 32.Spillantini MG, Schmidt ML, Lee VM, Trojanowski JQ, Jakes R, Goedert M. Alpha-synuclein in Lewy bodies. Nature. 1997;388:839–840. doi: 10.1038/42166. [DOI] [PubMed] [Google Scholar]
- 33.Sullivan AM, O’Keeffe GW. Neurotrophic factor therapy for Parkinson’s disease: past present and future. Neural Regen Res. 2016;11:205–207. doi: 10.4103/1673-5374.177710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wang RN, Green J, Wang Z, Deng Y, Qiao M, Peabody M, Zhang Q, Ye J, Yan Z, Denduluri S, Idowu O, Li M, Shen C, Hu A, Haydon RC, Kang R, Mok J, Lee MJ, Luu HL, Shi LL. Bone morphogenetic protein (BMP) signaling in development and human diseases. Genes Dis. 2014;1:87–105. doi: 10.1016/j.gendis.2014.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Warren Olanow C, Bartus RT, Baumann TL, Factor S, Boulis N, Stacy M, Turner DA, Marks W, Larson P, Starr PA, Jankovic J, Simpson R, Watts R, Guthrie B, Poston K, Henderson JM, Stern M, Baltuch G, Goetz CG, Herzog C, Kordower JH, Alterman R, Lozano AM, et al. Gene delivery of neurturin to putamen and substantia nigra in Parkinson disease: A double-blind, randomized, controlled trial. Ann Neurol. 2015;78:248–257. doi: 10.1002/ana.24436. [DOI] [PubMed] [Google Scholar]
- 36.Weiss A, Attisano L. The TGFbeta superfamily signaling pathway. Wiley Interdiscip Rev Dev Biol. 2013;2:47–63. doi: 10.1002/wdev.86. [DOI] [PubMed] [Google Scholar]
- 37.Whone A, Luz M, Boca M, Woolley M, Mooney L, Dharia S, Broadfoot J, Cronin D, Schroers C, Barua NU, Longpre L, Barclay CL, Boiko C, Johnson GA, Fibiger HC, Harrison R, Lewis O, Pritchard G, Howell M, Irving C, et al. Randomized trial of intermittent intraputamenal glial cell line-derived neurotrophic factor in Parkinson’s disease. Brain. 2019a;142:512–525. doi: 10.1093/brain/awz023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Whone AL, Boca M, Luz M, Woolley M, Mooney L, Dharia S, Broadfoot J, Cronin D, Schroers C, Barua NU, Longpre L, Barclay CL, Boiko C, Johnson GA, Fibiger HC, Harrison R, Lewis O, Pritchard G, Howell M, Irving C, et al. Extended treatment with glial cell line-derived neurotrophic factor in Parkinson’s disease. J Parkinsons Dis. 2019b;9:301–313. doi: 10.3233/JPD-191576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wozney JM. The bone morphogenetic protein family and osteogenesis. Mol Reprod Dev. 1992;32:160–167. doi: 10.1002/mrd.1080320212. [DOI] [PubMed] [Google Scholar]
- 40.Wrana JL, Attisano L. The Smad pathway. Cytokine Growth Factor Rev. 2000;11:5–13. doi: 10.1016/s1359-6101(99)00024-6. [DOI] [PubMed] [Google Scholar]
- 41.Xicoy H, Wieringa B, Martens GJM. The SH-SY5Y cell line in Parkinson’s disease research: a systematic review. Mol Neurodegener. 2017:12–10. doi: 10.1186/s13024-017-0149-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xie HR, Hu LS, Li GY. SH-SY5Y human neuroblastoma cell line: in vitro cell model of dopaminergic neurons in Parkinson’s disease. Chin Med J (Engl) 2010;123:1086–1092. [PubMed] [Google Scholar]
- 43.Yan W, Chen ZY, Chen JQ, Chen HM. BMP2 promotes the differentiation of neural stem cells into dopaminergic neurons in vitro via miR-145-mediated upregulation of Nurr1 expression. Am J Transl Res. 2016;8:3689–3699. [PMC free article] [PubMed] [Google Scholar]
- 44.Yu PB, Hong CC, Sachidanandan C, Babitt JL, Deng DY, Hoyng SA, Lin HY, Bloch KD, Peterson RT. Dorsomorphin inhibits BMP signals required for embryogenesis and iron metabolism. Nat Chem Biol. 2008;4:33–41. doi: 10.1038/nchembio.2007.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang YE. Non-Smad pathways in TGF-beta signaling. Cell Res. 2009;19:128–139. doi: 10.1038/cr.2008.328. [DOI] [PMC free article] [PubMed] [Google Scholar]