

Abstract
GtxA, a leukotoxic RTX-toxin, has been proposed a main virulence factor of Gallibacterium anatis. To evaluate the impact of GtxA during infection, we experimentally infected laying hens with a G. anatis wild-type (WT) strain and its isogenic gtxA deletion mutant (ΔgtxA), respectively, and monitored the birds during a 6 day period. Birds inoculated with ΔgtxA had significantly reduced gross lesions and microscopic changes compared to the birds inoculated with the WT strain. To assess the host response further, we quantified the expression of pro-inflammatory cytokines and apoptosis genes by RT-qPCR. In the ovarian tissue, the expression levels of IL-4 and TNF-α were significantly lower in the ΔgtxA group compared to the WT group, while IL-6 and IL-10 levels appeared similar in the two groups. In the spleen tissue of ΔgtxA infected chickens, IL-4 expression was also lower compared to the WT infected chickens. The results indicated that GtxA plays a key role in an acute cytokine-mediated Th2-like response against G. anatis infection in the ovary tissue. The pro-inflammatory response in the ovary tissue of birds inoculated with ΔgtxA mutant was thus significantly lower than the wild-type response. This was, at least partly, supported by the apoptosis gene expression levels, which were significantly higher in the ΔgtxA mutant compared to the wild-type infected chickens. In conclusion, GtxA clearly plays an important role in the pathogenesis of G. anatis infections in laying hens. Further investigations into the specific factors regulating the host response is however needed to provide a more complete understanding of the bacteria-host interaction.
Introduction
Gallibacterium anatis is a member of the Pasteurellaceae family colonizing the upper respiratory tract and lower genital tract of chickens [1]. Experimental infection studies have indicated that G. anatis has a pathogenic potential with a special affinity for the reproductive tract [2–4], and accumulating evidence points at G. anatis having an important role as a cause of salpingitis and peritonitis in laying hens worldwide, significantly compromising animal welfare and decreasing egg-yield [5–7].
Although some knowledge has been generated on specific factors important in the pathogenesis of G. anatis infections, several questions remain [8]. “Repeats in Toxins” (RTX-toxins) are potent cytolytic toxins produced by many Gram-negative bacteria including a wide range of species within the Pasteurellaceae family [9]. A G. anatis specific RTX-toxin (GtxA; Gallibacterium toxin) was identified by Kristensen et al. [10]. Gallibacterium anatis strains lacking parts of gtxA lost their haemolytic and cytolytic activity [11, 12]. Furthermore, GtxA has been shown to be immunogenic, and thus a promising vaccine candidate [13], which was partly confirmed by a study showing that the three recombinant proteins, GtxA-N, GtxA-C, and FlfA, could induce protection against G. anatis in an in vivo challenge model [14]. Several questions however remain regarding the role of GtxA in the pathogenesis of G. anatis [10].
The innate immune system utilizes pathogen-associated molecular patterns, such as Toll-like receptors (TLRs), which are expressed in chicken reproductive organs [15, 16] and play an essential role in the host defense mainly through immune recognition by the chicken [17]. Subsequently, a series of downstream cytokines, such as interferons (IFN) and interleukins (IL) are induced via cellular signaling pathways. Pathogenic G. anatis adhering to primary chicken oviduct epithelial cells have been shown to induce a strong inflammatory response and secretion of various cytokines, such as IL-6, TNF-α, and IFN-γ, which may lead to cell damage [18]. To what extend pathogenic G. anatis induces inflammatory cytokines in vivo has however not been reported.
Regulation of programmed cell death or apoptosis also plays an important role during bacteria-host interactions [19–21]. Many bacterial pathogens including Escherichia coli and Salmonella excrete pore-forming toxins that induce apoptosis, which may allow the pathogen to engage or circumvent the host’s efforts at limiting infections [22–24]. The importance of pore-forming toxins in G. anatis-induced apoptosis and their role in the pathogenesis has however not yet been clarified.
To investigate the role of GtxA during experimental infection in the natural host, we aimed at comparing lesions in laying hens infected with a G. anatis wildt-ype (WT) strain and its isogenic gtxA deletion mutant strain, which is unable to express GtxA. To assess the specific impact of GtxA on selected host immune factors, expression levels of genes encoding apoptosis and pro-inflammatory cytokines were assessed.
Materials and methods
Experimental animals and housing facilities
Twenty-four Lohmann Brown layer hens, 29-week old, were purchased from a commercial breeder with high biosecurity standards. At arrival, a swab from the cloacal mucosa was obtained from each chicken and immediately streaked on blood agar (BA) plate (Blood agar base, Difco, Heidelberg, Germany added 5% citrated bovine blood) The plates were incubated in a sealed plastic bag at 37 °C for 18 h. All plates were examined and presumptive G. anatis colonies were confirmed by PCR [25]. One colony of confirmed G. anatis growth from each bird was picked and prepared for storage at −80 °C for later use. The hens were kept under free indoor housing conditions with controlled ventilation, humidity and temperature, and were provided with fresh water and feed ad libitum. The hens were allowed to acclimatize for 1 week prior to the trial. All work with the chickens was carried out with the approval of the Danish Animal Experiments Inspectorate (license number: 2012-15-2934-00339).
Bacterial strains and experimental infection
A GtxA deletion mutant, G. anatis ΔgtxA, constructed as previously reported by Kristensen et al. [10] and its virulent parent wild-type (WT) strain G. anatis (strain no. 12656-12) were used for the inoculations. The WT strain was originally isolated from a bird with septicaemia. The strain has been characterized in detail by phenotypic and genotypic methods and has previously been used for in vivo studies of pathogenicity [2, 26]. Both strains were stored at −80 °C and cultivated overnight on BA in a sealed plastic bag to obtain single colonies, which were subsequently incubated in Brain Heart Infusion (BHI) broth (Oxoid, Basingstoke, UK) with agitation at 37 °C overnight. Prior to inoculation, an overnight culture of each strain was added to fresh BHI (in dilution ratio 1:2) followed by incubation at 37 °C for approximately three hours to reach the late log-phase and a concentration of approximately 109 CFU/mL. The bacterial concentration in each inoculum was verified by plate counts of tenfold serial dilutions of inocula on BA plates in duplicate. For the infection study, birds were randomly assigned to one of three groups (Table 1). The G. anatis ΔgtxA infection group was inoculated with a single dose of 4.9 × 108 CFU (in 1 mL BHI). The G. anatis WT infection group received a single dose of 5 × 108 CFU (in 1 mL BHI). The inocula were injected into the peritoneal cavity, just ventral for the spine and caudal to the last rib, using a 25 G cannula with a 30 mm-length. Chickens in the control group served as uninfected controls and were inoculated with 1 mL of sterile BHI. The unequal distribution of chickens in the gtxA mutant and wild-type groups (16 vs 4, respectively) was based on several previous studies done with the G. anatis WT (strain 12656-12), which made infections with this strain highly predictable and thus allowed fewer birds to be included [2, 14, 27].
Table 1.
Design of the study. A total of 24 birds were inoculated withG. anatisΔgtxA(16 chickens),G. anatis12656-12 (4 chickens) or BHI (4 chickens)
| Group | Inoculum | Dose/CFU | Chickens examined post-inoculation (pi) | |
|---|---|---|---|---|
| 2 days pi | 6 days pi | |||
| G. anatis ΔgtxA | ΔgtxA | 4.9 × 108 | 8 | 8 |
| G. anatis wild-type | 12656-12 | 5.0 × 108 | 2 | 2 |
| Control | BHI | – | 2 | 2 |
Post-mortem examination
The chickens were euthanized and subjected to post-mortem examination at either 2 or 6 days post-infection (pi) (Table 1). Recording of gross lesions and tissue sampling for histology was done for selected organs including the spleen, liver, ovary and oviduct. Tissue samples from the spleen and ovary were stored in RNAlater (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Bacterial isolates from the spleen, liver, ovary and salpinx were obtained by streaking tissue swabs onto BA. Isolates of the G. anatis WT strain were identified as G. anatis when greyish, semi-transparent colonies surrounded by a haemolytic zone of 1 to 2 mm and had phenotypic characters resembling those previously reported [28]. Gallibacterium anatis ΔgtxA showed identical characteristics but lacked a haemolytic zone [10]. Tissue samples from the ovary and oviduct were fixed in 10% neutral buffered formalin for 24 h, processed by standard histological procedures through graded concentrations of ethanol and xylene, and finally embedded in paraffin wax, sectioned 3–5 µm thick, stained with haematoxylin and eosin, and evaluated by light microscopy (Olympus BX45). Samples from all birds were examined for histological lesions together with the controls and the inflammatory reactions, including cellular infiltration, edema, exudation and necrosis, were evaluated according to extent and distribution. The lesions were differentiated using semi-quantitative grading system where scored 0 (no lesions), + (few scattered lesions), ++ (moderate number lesions), and +++ (severe diffuse lesions).
Pulsed-field gel electrophoresis (PFGE)
To verify that the re-isolated G. anatis from lesions were identical to the inoculum and different from the resident cloacal strains, pairs of G. anatis obtained from the cloaca and from lesions from eleven birds were characterized by pulsed-field gel electrophoresis (PFGE). The isolates had been kept at −80 °C in BHI broth with 10% glycerol and were plated on BA. Inoculated plates were incubated at 37 °C in sealed plastic bags for 18 h. From pure cultures, a single colony typical of G. anatis was picked and incubated overnight in 10 mL BHI broth at 37 °C with shaking. The preparation of bacterial DNA and separation of fragments was done as described [30] and using SalI (R0114, New England BioLabs) and XbaI (R0145, New England BioLabs) as restriction enzymes.
Serum sampling and enzyme-linked immunosorbent assay (ELISA)
GtxA specific antibody levels were quantified in serum samples by ELISA. Blood, 1–2 mL, was taken from vena brachialis from all birds at the day of inoculation (Day 0) and at 2 and 6 days after inoculation, respectively, and left at 4 °C overnight. Serum was collected after centrifugation at 1800 g for 10 min and stored at −20 °C. Two microtiter plate wells (Nunc-Immuno™ MicroWell™ 96-Well Plates, Thermo Scientific) were coated overnight at 4 °C with 0.5 µg GtxA recombinant protein diluted in carbonate-bicarbonate buffer (pH 9.6) (Sigma-Aldrich). Each well was then washed; this and all subsequent washing steps consisted of three washes in 350 µL wash buffer (PBS + 0.05% Tween 20) [13]. The wells were blocked for 2 h at room temperature in 200 µL blocking solution (PBS containing 0.05% Tween 20 and 2% bovine serum albumin (BSA)) and washed. The antibody titers were assayed by serial threefold dilutions of chicken serum ranging from 1:200 to 1:48 600. All dilutions were prepared in triplicates in a dilution buffer (PBS containing 0.05% Tween 20 and 0.1% BSA), 100 µL was added to each well and plates were incubated for 1 h at 37 °C. For each assay, 12 control wells were included, which contained pure dilution buffer; secondary antibody was added to 6 of these wells as a measure of background, and the other 6 wells remained blank as a negative control for the ELISA. Following incubation, the wells were washed and 100 µL polyclonal goat anti-chicken IgG (Fc): HRP (AbD Serotec) diluted 1:4000 in diluting buffer were added to each well and the plates incubated for a 1 h at 37 °C and then washed. To detect binding, 100 µL of 3,3′,5,5′-Tetramethylbenzidine liquid substrate (Sigma) were added to each well. The plates were incubated for 2 min and then the reaction was stopped by addition of 100 µL 1 M HCl. The absorbance was read immediately at 450 in a PowerWave XS spectrophotometry (BioTek Instruments).
Differential gene expression analysis by Real-Time Quantitative PCR (RT-qPCR)
To examine the mRNA expression profiles in the spleen and ovary tissue samples obtained during necropsy and placed in RNA later and stored according to the manufacturer’s instructions (Merck Life Science A/S, Søborg, Denmark). For RNA extraction a total of 20 mg tissue from each organ was placed in 1.5 mL tube containing beads. Each sample was then homogenized in 1 mL lysate buffer RLT using a Vortex adapter (Mo Bio, Carlsbad, CA, USA) and centrifugated at 1800 g. After homogenization and centrifugation for 5 min, the upper phase of total RNA was collected and purified with an RNeasy mini kit (Qiagen, Hilden, Germany) in accordance with the manufacturer’s instructions. The concentration and purity of RNA were determined by spectrophotometer (Nanodrop 1000, Thermo Scientific). 5 µg of RNA extracted from each tissues sample was reverse-transcribed into cDNA using M-MLV reverse transcriptase kit (Invitrogen) primed with Invitrogen’s protocol. The amount of cDNA corresponding to 20 ng of reverse-transcribed RNA was amplified by RT-qPCR, using specific primers (Tables 2 and 3) RT-qPCR was performed in 25 μL volumes in 96-well microplates using FastStart Essential DNA Green Master Mix (Roche, Germany) The 3-step amplification and signal detection were performed using a LightCycler R® 96 (Roche) with an initial pre-incubation at 95 °C for 10 min followed by 40 cycles of 95 °C for 15 s, 56 °C for the 30 s and 72 °C for 30 s. The β-actin gene was used as a housekeeping gene, to correct for differences in template RNA levels between samples during the experiment. Each RT-qPCR experiment thus included triplicates of 16 test samples, one no-template-control, and a log10 dilution series. The mean Ct value was used for subsequent calculations. Expression of the IL-4, IL-6, IL-10, TNF-α, casp-3, casp-8, casp-9, bax and bcl-2 genes was quantified according to Hangalapura et al. [29] (Table 2). First, the Difference Factor for each sample was calculated by dividing the mean Ct value for the β-actin gene of each individual sample by the mean Ct value for the β-actin gene of all samples. Secondly, the adjusted cytokine mRNA amount per sample was calculated using the following formulae:
Table 2.
List of the primers used in RT-qPCR analysis of mRNA expression of selected host genes
| Name | Primer | Primer sequence(5′–3′) | Accession | Conc. (mM) |
|---|---|---|---|---|
| bcl-2 | Forward | GATGACCGAGTACCTGAACC | NM205339 | 0.2 |
| Reverse | CAGGAGAAATCGAACAAAGGC | |||
| bax | Forward | TCCTCATCGCCATGCTCAT | XM422067 | 0.4 |
| Reverse | CCTTGGTCTGGAAGCAGAAGA | |||
| caspase-8 | Forward | TGGCCCTCTTGAACTGAAAG | AY057940 | 0.4 |
| Reverse | TCCACTGTCTGCTTCAATACC | |||
| caspase-9 | Forward | CGAAGGAGCAAGCACGACAG | AY057940 | 0.2 |
| Reverse | CCGCAGCCCTCATCTAGCAT | |||
| caspase-3 | Forward | TGGCCCTCTTGAACTGAAAG | AY057940 | 0.4 |
| Reverse | TCCACTGTCTGCTTCAATACC | |||
| IL-6 | Forward | GCTCGCCGGCTTCGA | AJ250838 | 0.2 |
| Reverse | GGTAGGTCTGAAAGGCGAACAG | |||
| IL-4 | Forward | AACATGCGTCAGCTCCTGAAT | AJ621735 | 0.4 |
| Reverse | TCTGCTAGGAACTTCTCCATTGAA | |||
| IL-10 | Forward | CATGCTGCTGGGCCTGAA | J621614 | 0.4 |
| Reverse | CGTCTCCTTGATCTGCTTGATG | |||
| TNF-α | Forward | GCCCTTCCTGTAACCAGATG | NM_204267 | 0.2 |
| Reverse | ACACGACAGCCAAGTCAACG | |||
| β-actin | Forward | CCGCTCTATGAAGGCTACGC | L08165 | 0.4 |
| Reverse | CTCTCGGCTGTGGTGGTGAA |
Table 3.
Gross pathology and re-isolation rates ofG. anatisfrom different organs following experimental infection with a wild-type strain (G. anatis12656-12) (4 chickens) or its isogenicgtxAdeletion mutant (ΔgtxA) (16 chickens)
| G. anatis | Peritoneum | Ovary | Oviduct | |||
|---|---|---|---|---|---|---|
| Purulent peritonitis | Re-isolation of G. anatis | Purulent oophoritis | Re-isolation of G. anatis | Salpingitis | Re-isolation of G. anatis | |
| ΔgtxA | 2/16 (13%) | 2/16 (13%) | 5/16 (31%) | 10/16 (63%) | 0/16 (0%) | 7/16 (44%) |
| Wild-type | 3/4 (75%) | 3/4 (75%) | 3/4 (75%) | 3/4 (75%) | 3/4 (75%) | 3/4 (75%) |
Statistics
Fisher’s Exact test was used for comparison of differences in the extent of the gross lesions and bacterial re-isolation rates. ANOVA was used to compare antibody-titers. The Kruskal–Wallis test was used for comparisons of histological lesion scores. Differences in gene expression levels were compared by Students t-test using GraphPad Prism 7® for Windows (GraphPad, San Diego, USA). P-values ≤ 0.05 were considered statistically significant.
Results
Pathology induced by G. anatis WT strain and its isogenetic ΔgtxA mutant
No clinical signs of infection were seen after the bacterial inoculations. At the necropsy, ten out of the 16 birds having received G. anatis ΔgtxA demonstrated mild macroscopic lesions whereas the remaining chickens had no lesions. Of the chickens infected with G. anatis WT, three had severe lesions and one had no lesions. No lesions were found in the birds in the BHI inoculated control group. The number of organs with lesions in birds from the ΔgtxA group, was significantly lower compared to the number of organs with lesions in WT group (P = 0.003). No difference was found between number of lesions when comparing birds examined at 2 or 6 days pi from the ΔgtxA group (P = 0.2). The histological lesions in both the ΔgtxA and WT infection groups supported the observations noted from the gross lesions (Figure 1). No histological lesions were found in birds without gross lesions. In the ΔgtxA group, few focal infiltrations of inflammatory cells, predominantly lymphocytes, were found in the oviduct and in one bird a necrotic focus was found. The ovarian stromal tissue was edematous with blood in the vascular system. Additionally, only light infiltration of inflammatory cells and degeneration of follicles were seen in birds with purulent oophoritis (Figure 1C). In the group infected with the WT strain, multifocal infiltration with inflammatory cells was present in the oviduct and several foci with bacterial colonization were also evident. In the ovary, infiltration with inflammatory cells including lymphocytes and heterophils was identified. In some areas, granuloma formation surrounded by epithelioid cells and necrosis was found (Figure 1D). A significant difference in histological lesion scores of both ovarian (P < 0.001) and oviduct tissues (P < 0.001) was found when comparing the two groups (Figure 2).
Figure 1.
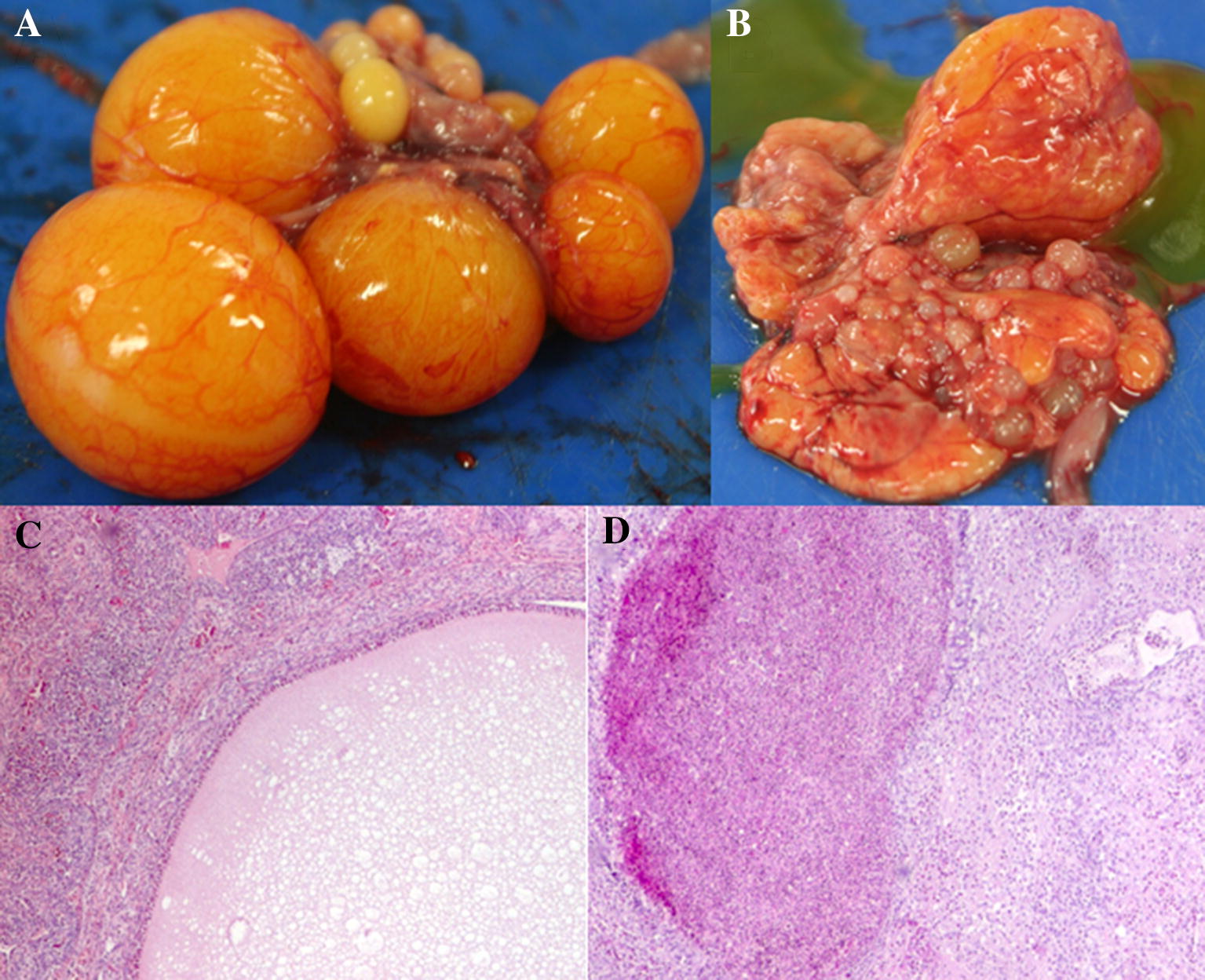
Gross lesions, histopathology and apoptotic cells found in chickens infected withG. anatisΔgtxAmutant orG. anatiswild-type (WT) strain. Ovaries from 29-week old laying hens. Two days post-inoculation with either G. anatis ΔgtxA or G. anatis WT. AG. anatis ΔgtxA infection group. Oophoritis with vascular engorgement and slight edema. BG. anatis wild-type. Diffuse purulent oophoritis and folliculitis with ruptures follicles. C Hematoxylin and eosin (HE)-straining of the follicle in ovarian tissue from the G. anatis ΔgtxA infection group. Focal oophoritis with the presence of inflammatory cells in the stroma and vascular engorgement. Infiltrates of few heterophilic granulocytes and mononuclear cells. D HE-straining of the ovary of the G. anatis WT infection group. Purulent oophoritis with heavy infiltration of inflammatory cells and granuloma formation (black arrows) in the follicle.
Figure 2.
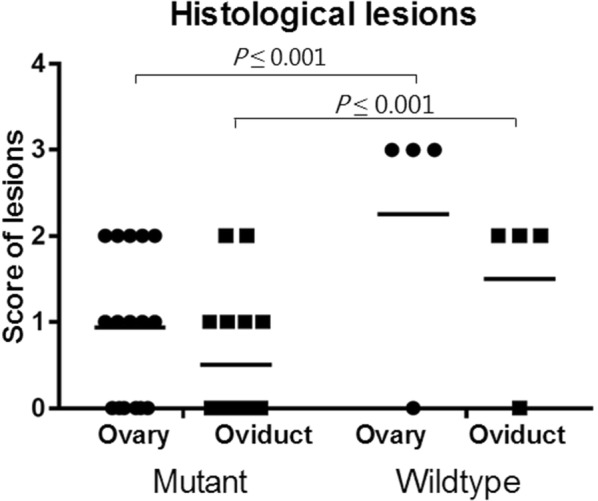
Score of histological lesions in the ovary and oviduct of chickens infected withG. anatisΔgtxAorG. anatiswild-type strain. No clinical signs of infection were observed after inoculation. At the necropsy, 10 out of the 16 birds having received G. anatis ΔgtxA demonstrated mild lesions, whereas remaining birds did not have any lesions. In birds infected with G. anatis WT strain, three of four birds had severe lesions and one was without lesions. No lesions were found in the control bird group. The number of organs with lesions in birds from the ΔgtxA group was significantly lower compared to the number of organs with lesions in WT group (P = 0.003). No difference was found between a number of lesions when comparing birds examined at 2 or 6 days pi from the ΔgtxA group (P = 0.2). In the ΔgtxA infection group, a mild non-purulent oophoritis characterized by vascular congestion and enlargement of the stromal tissue of the ovary (Figure 1A) was observed in five birds. In three birds, the oophoritis was purulent and in two birds, both examined 6 days pi, focal purulent oophoritis and localized peritonitis was found. Salpingitis was not observed in any of the birds from the ΔgtxA mutant infection group. The three out of four birds in the WT strain group had gross lesions, all exhibiting diffuse purulent peritonitis, diffuse purulent oophoritis and salpingitis (Figure 1B). No lesions were found in the spleen or liver of any birds. * indicates a statistically significant difference between groups (P ≤ 0.05).
Re-isolation of G. anatis
Resident cloacal G. anatis was isolated from 17 of the 24 birds prior to the experimental infections. At the necropsy G. anatis was re-isolated in pure culture from all gross lesions in the infected groups. Additionally, G. anatis was isolated from the oviduct mucosa of hens of the ΔgtxA infected group even in cases where no gross lesions were evident (Table 3). No bacteria were isolated from the internal organs of birds in the control group. To compare the isolates obtained from lesions and from the cloacal mucosa, pairs of isolates were characterized from eleven individual birds. All eleven isolates associated lesions had identical band patterns corresponding to the inoculated strains, whereas all of the cloacal isolates clearly had different PFGE profiles (Additional file 1).
The titer of GtxA specific antibodies was quantified at the time of inoculation and found at background levels in both groups (Figure 3).
Figure 3.
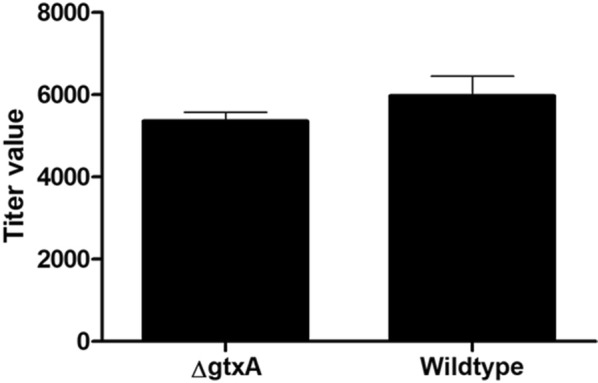
Titers for GtxA-specific IgG in serum from groups of birds. Blood drawn at the time of inoculation with either G. anatis ΔgtxA mutant (n = 4) or G. anatis wild-type (n = 8). No significant difference was found between the groups (P = 0.2).
Gene differential expression analysis by real-time quantitative PCR
To assess the ability of GtxA in mounting a host response during the G. anatis infection, the expression levels of cytokines IL-4, IL-6 IL-10 and TNF-α were quantified in the ovary and spleen tissues. No differences in the expression of the pro-inflammatory cytokines IL-6 and IL-10 were found in the ovary at 2 days pi between WT strain and ΔgtxA (Figure 4A), whereas significantly lower expression of IL-4 and TNF-α, respectively, was found in the ΔgtxA group compared to the WT group. At 6 days pi, expression of IL-4 remained significantly lower in the ΔgtxA group compared to the WT strain group (Figure 4B), while the ovarian expression of IL-6, TNF-α and IL-10 did not differ between the two infection groups (Figure 4B).
Figure 4.
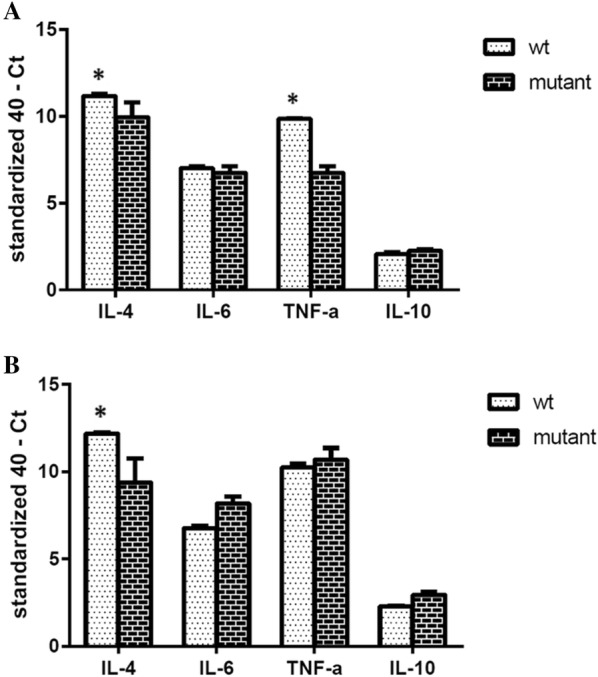
Changes of the cytokine mRNA expression levels in the ovary. Cytokine mRNA was isolated from culture positive ovary tissue at 2 days pi (A) and 6 days pi (B). As values are subtracted from the negative endpoint, higher values represent higher levels of cytokine mRNA levels. Error bars are S.E. for each treatment group (*P < 0.05, **P < 0.01) between WT (white bars) and ΔgtxA mutant groups (black bars).
In the spleen, the TNF-α expression was significantly higher in the ΔgtxA infected hens compared to the WT group at 2 days pi, while for IL-4, IL-6 and IL-10 no statistically significant difference was found between the two groups (Figure 5A). At 6 days pi, the expression of IL-4 and IL-10 in the spleen tissue was significantly lower in the ΔgtxA group compared to the WT group, while TNF-α expression was significantly higher in the ΔgtxA group (Figure 5B). In all tissue samples from WT group and the ΔgtxA group the level of IFN-γ was not detectable (data not shown).
Figure 5.
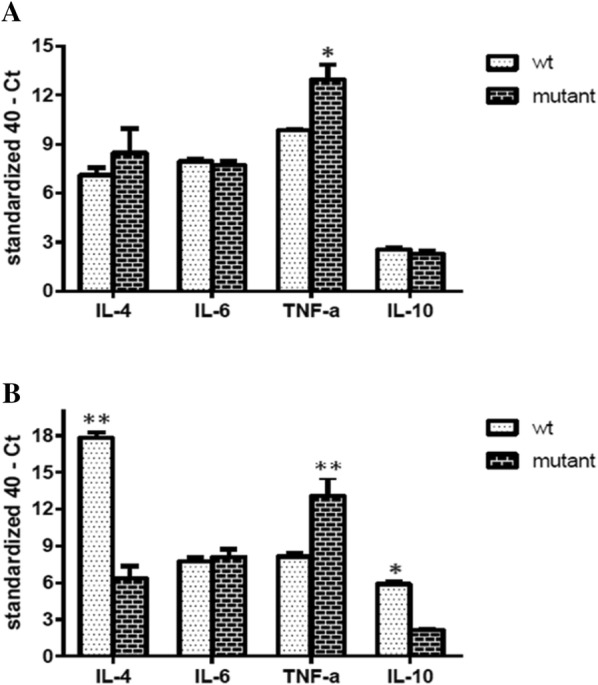
Changes of the cytokine mRNA expression levels in the spleen. Cytokine mRNA was isolated from culture positive spleen tissue at 2 days pi (A) and 6 days pi. (B). Error bars are S.E. for each treatment group (*P < 0.05, **P < 0.01) between WT (white bars) and ΔgtxA mutant groups (black bars).
Apoptosis-related gene expression in the ovary
The expression of selected apoptosis genes was evaluated in the ovarian tissue where the expression of bax was significantly increased in the ΔgtxA infection group compared to the WT group at 2 days pi (Figure 6). At 6 days pi, the expression the apoptosis genes was not different between the ΔgtxA and WT group, respectively (Figure 6).
Figure 6.

The relative expression levels of apoptosis genes in ovary tissue at 2 days and 6 days post-infection ofgtxAmutant compared with WT strain. Error bars are S.E. for each treatment group The figure shows the gene expression profile of apoptosis-related (Bcl-2), caspase-8, caspase-3, caspase-9 differed significantly (*P < 0.05, **P < 0.01) between the WT strain and ΔgtxA mutant groups, respectively.
Apoptosis-related gene expression in the spleen
The expression of apoptosis genes in the spleen tissue was assessed at 2 days and 6 days pi (Figure 7). The expression of bax, bcl-2, casp-3, casp-8 and casp-9 were all numerically lower at 2 days pi in the gtxA mutant group compared with WT group, yet this was only significant for casp-3 and casp-9 (Figure 7). At 6 days pi, no statistically significant difference in the expression was observed between the ΔgtxA infected group and the WT group (Figure 7).
Figure 7.

The relative expression changes of apoptosis genes in the spleen at 2 days and 6 days pi with thegtxAmutant compared with WT strain, respectively. Error bars are S.E. for each treatment group The gene expression profiles of apoptosis-related mRNA of B cell lymphoma-2 (Bcl-2), caspase-8, caspase-3, caspase-9 differed significantly (*P < 0.05, **P < 0.01, ***P < 0.001) between WT and ΔgtxA mutant groups, respectively.
Discussion
The results of the present study showed that the virulence of G. anatis was severely decreased in vivo when expression of the gtxA gene was impaired. Loss of virulence was indicated by a significantly lower lesion score in birds infected with the ΔgtxA strain compared to those infected with the GtxA-producing strain (Figures 1 and 2). The lesions induced by the G. anatis WT strain corresponded closely with observations made from previous in vivo studies [2, 14, 27], and thus justified the limited number of chickens used in the WT group. One WT infected hen had no apparent lesions and the inoculum had seemingly been deposited and trapped in a lump of abdominal fat. The observed differences in severity of lesions are in accordance with in vivo studies investigating strains of Actinobacillus pleuropneumoniae lacking the ability to produce different RTX-toxins either due to inactivation or deletion of the toxin genes, which all showed a sharp decline in virulence including lower mortality, morbidity and diminished lesions in pigs [31–33]. Experimental infection with a lktA (a leukotoxin gene) deleted mutant of Mannheimia haemolytica also led to a decrease in clinical signs and lung lesions in calves when compared to the isogenic WT strain [34]. Hence our findings support previous observations of RTX-toxins produced by other members of Pasteurellaceae and stress the profound role of GtxA in the pathogenesis of G. anatis.
Most RTX-toxins seem to enforce a strong leukotoxic effect and thereby debilitating the host immune response, which seems particularly aimed at specific host cells and thereby a likely cause of the host-specific pathogenicity commonly observed among Pasteurellaceae bacteria [9, 35]. In the present study, the inflammatory response promoted by the ΔgtxA mutant was significantly reduced whereas the WT induced severe purulent oophoritis with a high number of heterophilic granulocytes present and granuloma formation, indicating that GtxA plays an important role at activating heterophils during the immediate inflammatory response against G. anatis. Previous reports have indicated a similar role of the heterophils during other bacterial infections [36–38].
The highly decreased cellular infiltration and inflammatory reaction found in the tissues of the birds infected with the ΔgtxA mutant (Figures 1A and C) indicated less tissue destruction in the absence of GtxA, subsequently leading to a weaker host response. Virulence factors, including RTX-toxins, of other species of Pasteurellaceae have been shown to boost a strong pro-inflammatory response in vitro [36] and in vivo [2] and the lack of GtxA expression might similarly, lead to a diminished inflammatory response. This is, at least partly, supported by the re-isolation of the ΔgtxA mutant from the oviduct without a concurrent inflammatory response. However, it also suggests that G. anatis unability to produce GtxA withstands the capacity to effectively colonize the chicken oviduct and thereby points at other bacterial factors e.g. the F17-like fimbria that may take part in this process [12, 40, 41].
Due to the widespread occurrence of G. anatis among laying hens, it was not surprising that G. anatis could be isolated in the cloacae of most of the chickens entering the experiment. However, the strains isolated from the lesions associated with the experimental infections all showed to be identical to the strains used for the inoculation and different from the G. anatis isolates from the cloaca (Additional file 1). The factors permitting a commensal lifestyle of G. anatis are not well understood, but the present results indicate that the bird’s immune system generally have not been stimulated to initiate an adaptive response in healthy carriers as the GtxA specific titers were at or below the background level, as previously reported [14, 27]. Overall, the G. anatis ΔgtxA mutant was strongly attenuated upon experimental infection in its natural host suggesting that GtxA contributes significantly to the severity of infections in chickens.
Previous studies have clearly indicated GtxA to be a virulence factor while having the ability to induce protective immunity against G. anatis [27]. However, the specific host immune responses, here represented by cytokine and apoptosis gene expression profiles, in poultry have not previously been reported. In the current study we investigated pro-inflammatory cytokines involved in systemic inflammation and stimulation of the acute phase reaction [29, 36, 41]. We observed a high level of IL-4 and TNF-α mRNA expression in the ovary at 2 days and 6 days pi in the WT group, but a relatively low level in the spleen tissue at 2 days pi (Figures 4 and 5). The results are consistent with those reported by Zhang et al. [18], who revealed that G. anatis could induce marked inflammatory responses in primary chicken oviduct epithelial cells. The results indicate that GtxA may be involved in the stimulation of an IL-4 and TNF-α and promotes Th2-like response particularly in the ovary tissue, which previously has been suggested a main target for G. anatis [39].
As a pro-inflammatory cytokine, IL-6 is involved in the recruitment of immune cells, including lymphocytes and circulating monocytes, to the site of infection [17]. In our study, there were no significant differences in the IL-6 mRNA expression when comparing the WT and the ΔgtxA mutant in the ovary or the spleen tissues, respectively (Figures 4 and 5). It may be that in chickens, GtxA modulates the adaptive rather than innate immune response.
The regulation of the Th1-Th2 cytokine ratio plays an important role in balancing the host immune response [42, 43]. Our results indicated that GtxA expression primarily initiated a Th2-like response, as indicated by the increased IL-4 expression at 2 days and 6 days in the ovary tissue and similar but slightly delayed response (only at day 6) in the spleen tissue. On the contrary, in the ΔgtxA infected group, TNF-α was highly expressed, particularly in the spleen tissue at 2 days and 6 days whereas the level of IL-4 was low. These results indicate that the ΔgtxA mutant is able to inhibit or simply unable to induce secretion of IL-4. IL-4 is produced by Th2 cells that take part in the regulation of humoral immunity in poultry [44]. The cytokine expression levels are in good accordance with our pathological findings, showing a highly decreased cellular infiltration and inflammatory reaction in the tissues of birds infected with ΔgtxA (Figures 1A and 1C). The results also indicated that the ΔgtxA mutant induced less tissue and cellular destruction, subsequently leading to a weaker host response possibly through partial impairment of the Th2-like pathway. IL-10 is an anti-inflammatory cytokine [45, 46], negatively affecting the expression of Th1 cytokines [41]. We found that tissues from the WT infected group also had increased expression of IL-4 and partly IL-10 but not IFN-γ (which was below the detection level in all samples examined), supporting the Th2 bias suggested.
We hypothesized that GtxA was involved in regulating the apoptotic response in the host. Two major forms of apoptosis are commonly recognized, one involving activation of casp-8 (external induction) through the “death receptor” in the plasma membrane and the other involving disruption of the host cell homeostasis (intrinsic induction) by casp-9 regulation [47]. Both initiator caspases can recruit the executioner casp-3, which degrades cellular targets during apoptosis [47]. RTX toxins, such as Staphylococcus aureus toxin (alpha) and Actinobacillus leukotoxin, at low concentrations, have been suggested to cause the formation of small pores in the host cell plasma membrane allowing influx of Ca2+, which activates the intrinsic apoptotic pathways [48–50]. In our study, casp-3, -8, -9, bax and bcl-2 expression was significantly lower in the spleen tissue at 2 days pi in the chickens challenged with the ΔgtxA mutant compared to the WT (Figure 7), while the level of TNF-α was elevated (Figure 5A). This may reflect a dose–response relation where some but only a limited number of WT bacterial cells reached the spleen as opposed to the ΔgtxA mutant where very few if any bacterial cells made it to the spleen. On the contrary, in the ovary tissue, ΔgtxA induced higher levels of some of the pro-apoptosis genes (casp-8,-9 and bax) at 2 days, whereas apoptosis suppression, induced by increased bcl-2 expression, seemed apparent at 6 days (Figure 6B). The results are consistent with the fact that we observed only mild or no macro- and microscopic lesions in the ovary of the ΔgtxA infected chickens despite presence of bacteria in 10 out of 16 birds (Table 3). Based on these observations the ΔgtxA mutant seem able of inducing an apoptotic host response, which may allow prolonged survival in the host. At a high exposure of ovary tissue to the WT and GtxA a clear inflammatory response was observed. GtxA presumably forms large pores and rapid cytolysis of phagocytic cells [10], allowing the bacteria to escape the host defenses, which may prolong bacterial survival and increase the severity of the disease. The possible RTX toxin-induced pro-inflammatory death pathway, is a mechanism employed by Salmonella enterica, serovar Typhimurium and some Shigella species by initiating casp-1 expression through the lymphocyte function-associated antigen-1 (LFA-1) pathway [50–52]. Further research is however needed on how TNF-α is induced by non-GtxA expressing bacterial cell and how that may be involved in the regulation of apoptosis in cells exposed to G. anatis. Host cells may exploit the apoptosis process as a primitive defence mechanism against bacterial infections. On the other hand, several bacteria seem to have evolved mechanisms to prevent or delay apoptosis of host cells to permit successful bacterial replication [45]. GtxA may thus be involved in apoptosis suppression to facilitate G. anatis multiplication in the ovary at an early stage, which is considered a preferred site of this bacterium [39].
This study is the first investigation to assign a specific role of GtxA in the outcome of an infection of the natural host. It was documented that GtxA is important for the severity of lesions in an biologically relevant in vivo experiment and that GtxA contributes to stimulating both the innate and parts of the adaptive cellular immune system through primarily a Th2 response. Finally, our results indicated GtxA to be involved in the induction of apoptosis-related genes in spleen tissue.
Supplementary information
Additional file 1. Pulsed-Field Gel Electrophoresis typing of residentG. anatisisolates and strains used in the inoculum. The 12656-12 wild-type (WT) strain and gtxA mutant (M) strains. Cloacal isolates (C) and isolates obtained post-inoculation from organs with lesions (L). WT, M and L strains have identical genotypes whereas the cloacal isolates belong to a genetically different group.
Acknowledgements
Katrine Madsen is acknowledged for her technical assistance.
Authors’ contributions
BT, SEP, RJS, BMK and AMB designed the experiments, conducted the analyses, and drafted the manuscript; BT, SEP, RJS, RHO, BMK, AMB conducted the study and supervised all analyses; BT and AMB critically revised the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by the Faculty of Health and Medical Sciences at the University of Copenhagen and The Chinese Scholarship Council.
Availability of data and materials
The datasets generated and analysed during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Bo Tang, Email: botang@sund.ku.dk.
Susanne E. Pors, Email: gnz219@alumni.ku.dk
Bodil M. Kristensen, Email: bodilmk@hotmail.com
Ragnhild Bager J. Skjerning, Email: ragnhildbager@hotmail.com
Rikke H. Olsen, Email: cava@sund.ku.dk
Anders M. Bojesen, Email: miki@sund.ku.dk
Supplementary information
Supplementary information accompanies this paper at 10.1186/s13567-020-00764-2.
References
- 1.Bojesen AM, Nielsen SS, Bisgaard M. Prevalence and transmission of haemolytic Gallibacterium species in chicken production systems with different biosecurity levels. Avian Pathol. 2003;32:503–510. doi: 10.1080/0307945031000154107. [DOI] [PubMed] [Google Scholar]
- 2.Bojesen AM, Nielsen OL, Christensen JP, Bisgaard M. In vivo studies of Gallibacterium anatis infection in chickens. Avian Pathol. 2004;33:145–152. doi: 10.1080/03079450310001652059. [DOI] [PubMed] [Google Scholar]
- 3.Paudel S, Alispahic M, Liebhart D, Hess M, Hess C. Assessing pathogenicity of Gallibacterium anatis in a natural infection model: the respiratory and reproductive tracts of chickens are targets for bacterial colonization. Avian Pathol. 2013;42:527–535. doi: 10.1080/03079457.2013.843160. [DOI] [PubMed] [Google Scholar]
- 4.Paudel S, Liebhart D, Hess M, Hess C. Pathogenesis of Gallibacterium anatis in a natural infection model fulfils Koch’s postulates: 1. Folliculitis and drop in egg production are the predominant effects in specific pathogen free layers. Avian Pathol. 2014;43:443–449. doi: 10.1080/03079457.2014.955782. [DOI] [PubMed] [Google Scholar]
- 5.Mirle C. Studies into incidence of Pasteurella haemolytica infections and their relevance to hens, with particular reference to diseases of the egg-laying apparatus. Monatsh Vet Med. 1991;46:545–549. [Google Scholar]
- 6.Neubauer C, De Souza-Pilz M, Bojesen AM, Bisgaard M, Hess M. Tissue distribution of haemolytic Gallibacterium anatis isolates in laying birds with reproductive disorders. Avian Pathol. 2009;38:1–7. doi: 10.1080/03079450802577848. [DOI] [PubMed] [Google Scholar]
- 7.Wang C, Pors SE, Christensen JP, Bojesen AM, Thøfner ICN. Comparison and assessment of necropsy lesions in end-of-lay laying hens from different housing systems in Denmark. Poult Sci. 2019 doi: 10.3382/ps/pez569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Persson G, Bojesen AM. Bacterial determinants of importance in the virulence of Gallibacterium anatis in poultry. Vet Res. 2015;46:57. doi: 10.1186/s13567-015-0206-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Frey J. The role of RTX toxins in host specificity of animal pathogenic Pasteurellaceae. Vet Microbiol. 2011;153:51–58. doi: 10.1016/j.vetmic.2011.05.018. [DOI] [PubMed] [Google Scholar]
- 10.Kristensen BM, Frees D, Bojesen AM. GtxA from Gallibacterium anatis, a cytolytic RTX-toxin with a novel domain organisation. Vet Res. 2010;41:25. doi: 10.1051/vetres/2009073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kristensen BM, Frees D, Bojesen AM. Expression and secretion of the RTX-toxin GtxA among members of the genus Gallibacterium. Vet Microbiol. 2011;153:116–123. doi: 10.1016/j.vetmic.2011.05.019. [DOI] [PubMed] [Google Scholar]
- 12.Johnson TJ, Danzeisen JL, Trampel D, Nolan LK, Seemann T, Bager RJ, Bojesen AM. Genome analysis and phylogenetic relatedness of Gallibacterium anatis strains from poultry. PLoS One. 2013;8:e54844. doi: 10.1371/journal.pone.0054844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bager RJ, Kudirkiene E, Da Piedade I, Seemann T, Nielsen TK, Pors SE, Mattsson AH, Boyce JD, Adler B, Bojesen AM. In silico prediction of Gallibacterium anatis pan-immunogens. Vet Res. 2014;45:80. doi: 10.1186/s13567-014-0080-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pors SE, Skjerning RB, Flachs EM, Bojesen AM. Recombinant proteins from Gallibacterium anatis induces partial protection against heterologous challenge in egg-laying hens. Vet Res. 2016;47:36. doi: 10.1186/s13567-016-0320-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ozoe A, Isobe N, Yoshimura Y. Expression of Toll-like receptors (TLRs) and TLR4 response to lipopolysaccharide in hen oviduct. Vet Immunol Immunopathol. 2009;127:259–268. doi: 10.1016/j.vetimm.2008.10.325. [DOI] [PubMed] [Google Scholar]
- 16.Michailidis G, Theodoridis A, Avdi M. Transcriptional profiling of Toll-like receptors in chicken embryos and in the ovary during sexual maturation and in response to Salmonella Enteritidis infection. Anim Reprod Sci. 2010;122:294–302. doi: 10.1016/j.anireprosci.2010.09.004. [DOI] [PubMed] [Google Scholar]
- 17.Nawab A, An L, Wu J, Li G, Liu W, Zhao Y, Wu Q, Xiao M. Chicken toll-like receptors and their significance in immune response and disease resistance. Int Rev Immunol. 2019;38:284–306. doi: 10.1080/08830185.2019.1659258. [DOI] [PubMed] [Google Scholar]
- 18.Zhang XP, Lu CJ, Li YT, Yang X, Wang XW, Chang HT, Liu HY, Chen L, Zhao J, Wang CQ. In vitro adherence and invasion of primary chicken oviduct epithelial cells by Gallibacterium anatis. Vet Microbiol. 2017;203:136–142. doi: 10.1016/j.vetmic.2017.02.009. [DOI] [PubMed] [Google Scholar]
- 19.Weinrauch Y, Zychlinsky A. The induction of apoptosis by bacterial pathogens. Ann Rev Microbiol. 1999;53:155–187. doi: 10.1146/annurev.micro.53.1.155. [DOI] [PubMed] [Google Scholar]
- 20.Fan TJ, Han LH, Cong RS, Liang J. Caspase family proteases and apoptosis. Acta Biochim Biophys Sin. 2005;37:719–727. doi: 10.1111/j.1745-7270.2005.00108.x. [DOI] [PubMed] [Google Scholar]
- 21.Elmore S. Apoptosis: a review of programmed cell death. Tox Path. 2007;35:495–516. doi: 10.1080/01926230701320337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Czuprynski CJ, Welch RA. Biological effects of RTX toxins: the possible role of lipopolysaccharide. Trends Microbiol. 1995;3:480–483. doi: 10.1016/S0966-842X(00)89016-2. [DOI] [PubMed] [Google Scholar]
- 23.Fernandez-Prada C, Tall BD, Elliott SE, Hoover DL, Nataro JP, Venkatesan MM. Hemolysin-positive enteroaggregative and cell-detaching Escherichia coli strains cause oncosis of human monocyte-derived macrophages and apoptosis of murine J774 cells. Infect Immun. 1998;66:3918–3924. doi: 10.1128/IAI.66.8.3918-3924.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Moss JE, Aliprantis AO, Zychlinsky A. The regulation of apoptosis by microbial pathogens. Int Rev Cytol. 1999;187:203–259. doi: 10.1016/S0074-7696(08)62419-5. [DOI] [PubMed] [Google Scholar]
- 25.Bojesen AM, Vazquez ME, Robles F, Gonzalez C, Soriano EV, Olsen JE, Christensen H. Specific identification of Gallibacterium by a PCR using primers targeting the 16S rRNA and 23S rRNA genes. Vet Microbiol. 2007;123:262–268. doi: 10.1016/j.vetmic.2007.02.013. [DOI] [PubMed] [Google Scholar]
- 26.Bojesen AM, Torpdahl M, Christensen H, Olsen JE, Bisgaard M. Genetic diversity of Gallibacterium anatis isolates from different chicken flocks. J Clin Microbiol. 2003;41:2737–2740. doi: 10.1128/JCM.41.6.2737-2740.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pedersen IJ, Pors SE, Bager Skjerning RJ, Nielsen SS, Bojesen AM. Immunogenic and protective efficacy of recombinant protein GtxA-N against Gallibacterium anatis challenge in chickens. Avian Pathol. 2015;44:386–391. doi: 10.1080/03079457.2015.1069259. [DOI] [PubMed] [Google Scholar]
- 28.Christensen H, Bisgaard M, Bojesen AM, Mutters R, Olsen JE. Genetic relationships among avian isolates classified as Pasteurella haemolytica, ‘Actinobacillus salpingitidis’ or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov. and description of additional genomospecies within Gallibacterium gen. nov. Intl J Syst Evol Microbiol. 2003;53:275–287. doi: 10.1099/ijs.0.02330-0. [DOI] [PubMed] [Google Scholar]
- 29.Hangalapura BN, Kaiser MG, van der Poel JJ, Parmentier HK, Lamont SJ. Cold stress equally enhances in vivo pro-inflammatory cytokine gene expression in chicken lines divergently selected for antibody responses. Dev Comp Immunol. 2006;30:503–511. doi: 10.1016/j.dci.2005.07.001. [DOI] [PubMed] [Google Scholar]
- 30.Pors SE, Hansen MS, Christensen H, Jensen HE, Petersen A, Bisgaard M. Genetic diversity and associated pathology of Pasteurella multocida isolated from porcine pneumonia. Vet Microbiol. 2011;150:354–361. doi: 10.1016/j.vetmic.2011.02.050. [DOI] [PubMed] [Google Scholar]
- 31.Reimer D, Frey J, Jansen R, Veit HP, Inzana TJ. Molecular investigation of the role of Apxl and Apxll in the virulence of Actinobacillus pleuropneumoniae serotype 5. Microb Pathog. 1995;18:197–209. doi: 10.1016/S0882-4010(95)90049-7. [DOI] [PubMed] [Google Scholar]
- 32.Kamp EM, Stockhofe-Zurwieden N, Van Leengoed L, Smits MA. Endobronchial inoculation with Apx toxins of Actinobacillus pleuropneumoniae leads to pleuropneumonia in pigs. Infect Immun. 1997;65:4350–4354. doi: 10.1128/IAI.65.10.4350-4354.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liu J, Chen X, Tan C, Guo Y, Chen Y, Fu S, Bei W, Chen H. In vivo induced RTX toxin ApxIVA is essential for the full virulence of Actinobacillus pleuropneumoniae. Vet Microbiol. 2009;137:282–289. doi: 10.1016/j.vetmic.2009.01.011. [DOI] [PubMed] [Google Scholar]
- 34.Tatum FM, Briggs RE, Sreevatsan SS, Zehr ES, Hsuan SL, Whiteley LO, Ames TR, Maheswaran SK. Construction of an isogenic leukotoxin deletion mutant of Pasteurella haemolytica serotype 1: characterization and virulence. Microb Pathog. 1998;24:37–46. doi: 10.1006/mpat.1997.0181. [DOI] [PubMed] [Google Scholar]
- 35.Frey J, Kuhnert P. RTX toxins in Pasteurellaceae. Int J Med Microbiol. 2002;292:149–158. doi: 10.1078/1438-4221-00200. [DOI] [PubMed] [Google Scholar]
- 36.Chen ZW, Chien MS, Chang NY, Chen TH, Wu CM, Huang C, Lee WC, Hsuan SL. Mechanisms underlying Actinobacillus pleuropneumoniae exotoxin ApxI induced expression of IL-1β, IL-8 and TNF-α in porcine alveolar macrophages. Vet Res. 2011;42:25. doi: 10.1186/1297-9716-42-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bojesen AM, Petersen KD, Nielsen OL, Christensen JP, Bisgaard M. Pasteurella multocida infection in heterophil-depleted chickens. Avian Dis. 2004;48:463–470. doi: 10.1637/7151. [DOI] [PubMed] [Google Scholar]
- 38.Nakamura K, Maeda M, Imada Y, Imada T, Sato K. Pathology of spontaneous colibacillosis in a broiler flock. Vet Pathol. 1985;22:592–597. doi: 10.1177/030098588502200614. [DOI] [PubMed] [Google Scholar]
- 39.Bager RJ, Nesta B, Pors SE, Soriani M, Serino L, Boyce JD, Adler B, Bojesen AM. The fimbrial protein FlfA from Gallibacterium anatis is a virulence factor and vaccine candidate. Infect Immun. 2013;81:1964–1973. doi: 10.1128/IAI.00059-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kudirkienė E, Bager RJ, Johnson TJ, Bojesen AM. Chaperone-usher fimbriae in a diverse selection of Gallibacterium genomes. BMC Genomics. 2014;15:1093. doi: 10.1186/1471-2164-15-1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Iqbal M, Philbin VJ, Withanage G, Wigley P, Beal RK, Goodchild MJ, Barrow P, McConnell I, Maskell DJ, Young J. Identification and functional characterization of chicken toll-like receptor 5 reveals a fundamental role in the biology of infection with Salmonella enterica serovar Typhimurium. Infect Immun. 2005;73:2344–2350. doi: 10.1128/IAI.73.4.2344-2350.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mosmann TR, Coffman R. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Ann Rev Immunol. 1989;7:145–173. doi: 10.1146/annurev.iy.07.040189.001045. [DOI] [PubMed] [Google Scholar]
- 43.Wu QJ, Zheng XC, Wang T, Zhang TY. Effects of oridonin on immune cells, Th1/Th2 balance and the expression of BLys in the spleens of broiler chickens challenged with Salmonella pullorum. Res Vet Sci. 2018;119:262–267. doi: 10.1016/j.rvsc.2018.07.008. [DOI] [PubMed] [Google Scholar]
- 44.Chtanova T, Mackay CR. T cell effector subsets: extending the Th1/Th2 paradigm. Adv Immunol. 2001;78:233–266. doi: 10.1016/S0065-2776(01)78005-4. [DOI] [PubMed] [Google Scholar]
- 45.Arsenault RJ, Genovese KJ, He H, Wu H, Neish AS, Kogut MH. Wild-type and mutant AvrA—Salmonella induce broadly similar immune pathways in the chicken ceca with key differences in signaling intermediates and inflammation. Poult Sci. 2015;95:354–363. doi: 10.3382/ps/pev344. [DOI] [PubMed] [Google Scholar]
- 46.Duell BL, Tan CK, Carey AJ, Wu F, Cripps AW, Ulett GC. Recent insights into microbial triggers of interleukin-10 production in the host and the impact on infectious disease pathogenesis. FEMS Immunol Med Microbiol. 2012;64:295–313. doi: 10.1111/j.1574-695X.2012.00931.x. [DOI] [PubMed] [Google Scholar]
- 47.Thumbikat P, Dileepan T, Kannan MS, Maheswaran SK. Mechanisms underlying Mannheimia haemolytica leukotoxin-induced oncosis and apoptosis of bovine alveolar macrophages. Microb Pathol. 2005;38:161–172. doi: 10.1016/j.micpath.2005.01.002. [DOI] [PubMed] [Google Scholar]
- 48.Khelef N, Zychlinsky A, Guiso N. Bordetella pertussis induces apoptosis in macrophages: role of adenylate cyclase-hemolysin. Infect Immun. 1993;61:4064–4071. doi: 10.1128/IAI.61.10.4064-4071.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Jonas D, Walev I, Berger T, Liebetrau M, Palmer M, Bhakdi S. Novel path to apoptosis: small transmembrane pores created by staphylococcal alpha-toxin in T lymphocytes evoke internucleosomal DNA degradation. Infect Immun. 1994;62:1304–1312. doi: 10.1128/IAI.62.4.1304-1312.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lally ET, Hill RB, Kieba IR, Korostoff J. The interaction between RTX toxins and target cells. Trends Microbiol. 1999;7:356–361. doi: 10.1016/S0966-842X(99)01530-9. [DOI] [PubMed] [Google Scholar]
- 51.Hilbi H, Moss JE, Hersh D, Chen Y, Arondel J, Banerjee S, Flavell RA, Yuan J, Sansonetti PJ, Zychlinsky A. Shigella-induced apoptosis is dependent on caspase-1 which binds to IpaB. J Biol Chem. 1998;273:32895–32900. doi: 10.1074/jbc.273.49.32895. [DOI] [PubMed] [Google Scholar]
- 52.Boise LH, Collins CM. Salmonella-induced cell death: apoptosis, necrosis or programmed cell death? Trends Microbiol. 2001;9:64–67. doi: 10.1016/S0966-842X(00)01937-5. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1. Pulsed-Field Gel Electrophoresis typing of residentG. anatisisolates and strains used in the inoculum. The 12656-12 wild-type (WT) strain and gtxA mutant (M) strains. Cloacal isolates (C) and isolates obtained post-inoculation from organs with lesions (L). WT, M and L strains have identical genotypes whereas the cloacal isolates belong to a genetically different group.
Data Availability Statement
The datasets generated and analysed during the current study are available from the corresponding author on reasonable request.