

Abstract
Objective
To investigate the impact of JTXK granule on the miRNA expression profiles in hepatic tissue of diabetic mice, and to explore the molecular targets and associated signaling pathways of JTXK granule in its anti-diabetic effect.
Methods
Eight mice were randomly selected as normal group fed with chow diet. Then high fat diet was used to induce diabetic model, and the mice were subsequently divided into JTXK-treated group (J group, n = 6) and model group (M group, n = 6). After 8 weeks’ intervention we examined the fasting blood glucose and observed the histopathologic changes in hepatic tissue between these two groups. Next we screened the differentially expressed miRNAs between the two groups using microRNA sequencing analysis. Finally, miRNA target gene prediction, GO and KEGG analysis were applied to explore the function of DEMs.
Results
The blood glucose level in J group was significantly lower than M group (P < 0.05). The results from H&E staining showed that the arrangement and structure of hepatocytes from J group were basically normal with fewer ballooning degeneration and less inflammatory cell infiltration. Furthermore, a total of 33 significantly differentiated miRNAs were detected in comparison between the two groups (| log2(fold change) | >0.3, P < 0.05). MiRNA–mRNA analysis showed that mmu-miR-30a-5p, mmu-miR-23b-5p, mmu-miR-199a-5p, mmu-miR-425-5p, and mmu-miR-214-3p are closely related to inflammatory response, histological changes and insulin signal transduction in liver. In addition, KEGG analysis showed that the DEMs were closely related to Ras and insulin signaling pathway.
Conclusion
JTXK granule exerts anti-diabetic effect in hepatic tissue of diabetic mice by modulating miRNAs and mRNAs network.
Keywords: Jiang Tang Xiao Ke Granule, bioinformatic analysis, microRNAs, anti-diabetic effect, diabetes mellitus
Graphical Abstract
JTXK granule protects diabetic liver by modulating miRNA-mRNA network.

Highlights
-
-
JTXK granule ameliorated hyperglycemia in diabetic mice.
-
-
JTXK granule elicited protective effect against diabetes-induced histopathological changes in liver.
-
-
JTXK granule significantly changed the miRNA–mRNA expression profile in liver tissue of diabetic mice.
-
-
JTXK granule exerted anti-hyperglycemia and anti-inflammatory effect in liver tissue of diabetic mice by modulating miRNAs and mRNAs associated with Ras and insulin signaling.
Introduction
The prevalence of diabetes mellitus (DM), a metabolic disorder, is rapidly increasing worldwide (Cho et al., 2018). Diabetes is characterized by hyperglycemia which is caused by either insulin deficiency, or impaired biological function of insulin (Manrique et al., 2014). Long-term hyperglycemia in diabetes leads to various tissue damage and organ dysfunction (Forbes and Cooper, 2013). Furthermore, diabetes is known to cause liver disorder, and contribute to chronic liver injury, including fatty liver, hepatic steatosis, liver fibrosis and cirrhosis (Fukuda et al., 2016). Epidemiological studies show that the risks of non-alcoholic fatty liver disease (NAFLD) as well as hepatocellular carcinoma (HC) are much higher among diabetic patients than non-diabetic population (Liu et al., 2017). In diabetic liver injury, hepatic steatosis and inflammatory cell infiltration could be observed, and insulin resistance and glucolipid metabolic disorder could emerge (Tian et al., 2017).
MicroRNAs are a series of non-coding single strand small RNAs, usually with a length of about 22 nucleotides, which are derived from RNA transcript regions that fold to hairpin structures (Shukla et al., 2011). MicroRNAs can identify binding sites on 3′-UTR of target genes and their function is mainly to inhibit downstream gene expression through transcriptional inhibition, mRNA cleavage or degradation (Wilczynska and Bushell, 2015). Growing evidence shows that microRNAs are involved in gene regulations in many pathophysiological processes, including glycolipid metabolism, antioxidant stress and inflammation, and thereby participate and exert significant impact on diabetes and its associated complications (Iacomino and Siani, 2017). Furthermore, studies have demonstrated that a variety of microRNAs (miRNAs) can regulate multiple target genes associated with insulin and Ras signaling pathways, thus forming a complex network and playing vital role in the regulation of glucolipid metabolism and inflammatory response (Kasinski and Slack, 2010; Mor et al., 2011). As a result, these miRNAs may serve as important regulators and therapeutic targets of diabetes associated liver injury.
In recent years, significant clinical breakthroughs have been made in the study of traditional Chinese medicine formula. It has been found that Chinese medicine formula play a major role in the treatment of diabetes through multiple targets with various mechanisms (Zhijun et al., 2013). Jiang Tang Xiao Ke (JTXK) Granule is a traditional Chinese herbal medicine compound for diabetes mellitus, which is composed of Rehmannia glutinosa (Gaertn.) DC. (Dihuang), Cornus officinalis Siebold & Zucc. (Shanzhuyu), Panax ginseng C.A.Mey. (Renshen), Coptis chinensis Franch. (Huanglian), Salvia miltiorrhiza Bunge (Danshen) and Pueraria montana var. lobata (Gegen) and so on. Our previous data have shown that JTXK Granule plays the anti-diabetic role by regulating glycolipid metabolism, resisting oxidative stress, and also protecting islet cells, which act through regulating phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT)-related microRNAs and mRNAs expressions in pancreatic tissues (Zhao et al., 2014; Mo et al., 2017; Yu et al., 2017). Another study also suggested that JTXK granule can regulate glycolipid metabolism in the liver tissue of diabetic mice (Yi et al., 2016). In addition to the amelioration of glucolipid metabolic disorder, we also found that JTXK granule exerted anti-inflammatory effect and ameliorated diabetic liver injury in high fat diet (HFD)-induced diabetic mice. Therefore, this study aims to investigate the potential role of JTXK granule in modulating the miRNAs and mRNAs network, and its related signaling pathways in liver tissue, and to reveal the underlying molecular targets of this classic herbal formula.
Materials and Methods
Preparation of JTXK Granule
JTXK formula is composed of Rehmannia glutinosa (Gaertn.) DC. (Dihuang), Cornus officinalis Siebold & Zucc. (Shanzhuyu), Panax ginseng C.A.Mey. (Renshen), Coptis chinensis Franch. (Huanglian), Salvia miltiorrhiza Bunge (Danshen), and Pueraria montana var. lobata (Gegen) etc (Please see Supplementary Table S1 for detail). All the herbal materials (prepared herbal medicine in small pieces ready for decoction) were purchased from Tongrentang Chinese Medicine Pharmacy (Beijing, China). The authenticity of the herbal medicine was verified by Lec. Xiaoxu Dong from Science and Technology Developmental Department of Traditional Chinese Materia Medica. Then all the herbs were mixed, processed and the concentrated extracts were made into granules (3.26 grams of herbs/gram of granules, extraction ratio 30.67%). The batch number was 20140518. Fingerprint of this granule that illustrated the main ingredients were shown in Figure 1 and in accordance with our previous study (Mo et al., 2017). Finally, the granules were stored in −4°C refrigerator and prepared suspension of corresponding concentration with distilled water before use.
FIGURE 1.

Fingerprint of JTXK granule. Paeonol (22), Salvia acid B (18), Berberine (19), Coptisine (16), Puerarin (10).
Animal Models and Experimental Procedure
All the animal experimental procedures were approved by the Animal Care Committee of Beijing University of Chinese Medicine (Beijing, China). In this study, 8-week-old ICR mice were purchased from Beijing Sibeifu Bioscience, Co. Ltd. (Beijing, China). All the mice were fed in the barrier environment of Beijing University of Chinese Medicine. The light–dark cycle was 12 h/12 h, the room temperature was controlled at 23°C and the relative humidity was ∼45%. After 1 week of adaptive feeding, 8 mice were randomly selected as normal group fed with standard diet (AIN-96G feed from Sibeifu Bioscience, Co., Ltd,. Beijing, China), and the remaining mice were fed with high-fat diet (HFD, MD12032, 45%fat Kcal%, Jiangsu Medicine, Co., Ltd., Jiangsu, China) during the entire experiment. After 12 weeks’ feeding, mice were divided into model group (M group, n = 6) and JTXK-treated group (J group, n = 6), randomly. Then, J group mice were treated with JTXK granule dissolved in sterilized water (1.75 g/kg) by gavage for 8 consecutive weeks. While the mice in normal and M group received the same amount of sterile water. Finally, all mice were sacrificed after 8 weeks of intervention, and the blood samples and livers were collected for further experiments.
Liver Histopathologic Analysis
The livers were immediately removed from the experimental mice at sacrifice, fixed by 10% neutral PBS formaldehyde and then embedded in paraffin. Next, the maximum sagittal sections of each lobe were cut into 4-μm sections and stained with hematoxylin and eosin (H&E) according to a set routine. An optical microscope (Olympus, Tokyo, Japan) was used to observe the histopathological changes in liver tissue.
MicroRNA Sequencing Analysis
We selected three mice from either M group or J group for miRNA Sequencing analysis. Total RNA was isolated for library preparation using multiplex small RNA library prep set and multiplex oligos set (NEBNext®, United States). In brief, the RNA was spliced at the 3′ end, hybridized with RT primer, spliced at the 5′ end, then reverse transcribed. Next, PCR amplification was carried out to obtain stable library. Following, quality assessment and quantification were accomplished using Qubit Flurometer (Thermo Fisher Scientific) and Qseq100 DNA Analyzer (Bioptic, Inc.). Then after mixing and denaturing, the libraries were added to Illumina HiSeq (HiSeq X Ten System, Illumina, Inc.) platform for sequencing in accordance with specific instructions. Next, raw data were filtered to get clean data. Number and distribution of all reads were analyzed. And unique reads were compared with Rfam, miRBase to identify the known and novel miRNAs. The expression of microRNAs in each sample was calculated using reads per million (RPM, RPM = each miRNA reads/all miRNA reads), and then the differential microRNAs were screened out (| log2(fold change) | >0.3, P < 0.05, average expression > 1 in at least one group). Finally, bar chart and hierarchical clustering figure were generated to illustrate the differential miRNA expression between M group and J group. The raw data of the miRNA-Seq has been deposited in Gene Expression Omnibus (GEO) database1 for public access (GEO Series accession number GSE145234).
Differential MiRNA Target Gene Prediction and Enrichment Analysis
The target genes of differentially expressed microRNAs (including both known and novel microRNAs) were predicted by Miranda and RNAhybrid, respectively. The intersection part of the results was considered as the final target gene prediction results, and then it was divided into target genes corresponding to either up-regulated or down-regulated miRNAs. Then gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses were carried out using Fisher’s exact test to identify the significantly related GO terms and KEGG pathways (P < 0.05).
Quantitative Real-Time PCR Verification of Selected miRNA
In order to validate the sequencing results, qRT-PCR analysis was carried out using selected miRNAs. We extracted total RNA from mouse liver tissue, and reverse-transcripted total RNA into cDNA with miRNA First-Strand cDNA Synthesis Kit (TIANGEN, Beijing, China). Then the miRNA expression measurement was done by qRT-PCR with miRNA qPCR Detection Kit (SYBR Green) (TIANGEN, Beijing, China) and Roche LightCycler® 96 fluorescence quantification PCR instrument (Roche, Basel, Switzerland). GAPDH was amplified as internal reference and the relative expression of miRNA to GAPDH was calculated. Primers used in this study were summarized and shown in Table 1.
TABLE 1.
MiRNA and mRNA primers for quantitative PCR analysis.
| Primier name | Sequence |
| U6 | F: GCTTCGGCAGCACATATACTAAAAT |
| R: CGCTTCACGAATTTGCGTGTCAT | |
| mmu-miR-23b-5p | RT: GTCGTATCCAGTGCAGGGTCCGAG |
| GTATTCGCACTGGATACGACAAATCAGC | |
| F: ATGGTTCGTGCGTGGAGAGAAAGGCAGTTC | |
| R: GCAGGGTCCGAGGTATTC | |
| Mmu-miR-185-5p | RT: GTCGTATCCAGTGCAGGGT |
| CCGAGGTATTCGCACTGGATACGACAGGAACTG | |
| F: ATGGTTCGTGCGTGGAGAGAAAGGCAGTTC | |
| R: GCAGGGTCCGAGGTATTC | |
| Gapdh ms | F: TGCCCCCATGTTTGTGATG |
| R: TGTGGTCATGAGCCCTTCC |
Statistical Analysis
In present study, we used SPSS 20.0 as a tool for statistical analysis. The results were presented as mean ± SEM (standard error). One-way ANOVA was applied in comparison of more than two group means. P < 0.05 was considered as the difference was statistically significant.
Results
JTXK Granule Decreased Fasting Blood Glucose Level in Diabetic Mice
To investigate JTXK granule’s effect on diabetes, the fasting blood glucose (FBG) level was measured by collecting blood samples after 8 weeks’ intervention. As shown in Figure 2, the level of FBG was significantly higher in M group than J group, indicating that the JTXK granule had anti-diabetic effect in HFD induced diabetic mice.
FIGURE 2.

JTXK granule reduced fasting blood glucose level. Results are shown as mean ± SEM. *P < 0.05 compared with model group. ##P < 0.01compared with normal group. Normal group, n = 8; Model group, n = 6; JTXK group, n = 6.
JTXK Granule Exerted Protective Effect on Liver Tissue in Diabetic Mice
To detect the role of JTXK granule in diabetic liver tissue, histopathologic changes in different groups were observed, as shown in Figure 3. In normal group, the hepatic lobules of liver tissue were structurally intact, and the hepatic cell cords were arranged neatly. The morphology of hepatocytes was normal, and the nuclei were located in the center of hepatocytes. However, in M group (HFD-induced diabetic mice, Model), the hepatocytes were swelling and abnormally arranged with obvious steatosis. There were lots of lipid vacuoles in the cytoplasm and ballooning degeneration of hepatocytes, as well as inflammatory cell infiltration in liver tissue (smaller cells with dark stained nuclear). In contrast, the arrangement and structure of hepatocytes from liver samples in J group were basically normal with fewer ballooning degeneration and less inflammatory cell infiltration, which supports that JTXK granule may preserve hepatocytes and histological structure of liver tissue.
FIGURE 3.
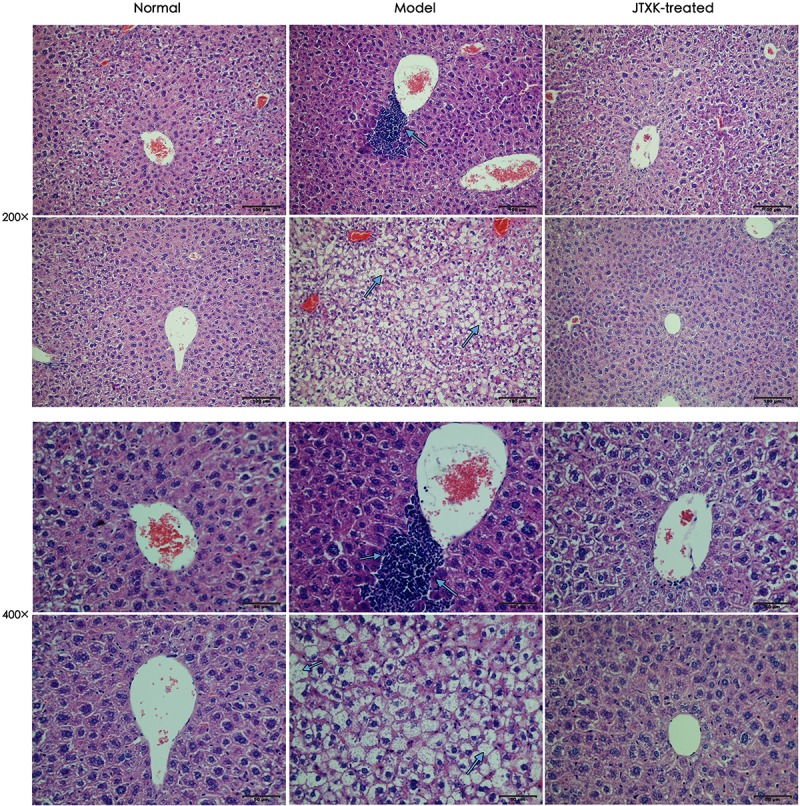
H&E Staining of hepatic tissue. Representative histology images in each group were exhibited (200, 400×). Normal: normal group, mice were fed with standard diet; Model: M group, mice were fed with 45% HFD; JTXK-treated: J group, mice were fed with 45%HFD and administrated with JTXK granule.
JTXK Granule Administration Altered the MiRNA Expression Profile in Diabetic Mice
Using miRNA sequencing analysis, we identified 33 differentially expressed miRNAs (DEMs) in liver tissue (| log2(fold change) | >0.3, P < 0.05, average expression >1 in at least one group) in total, of which 5 miRNAs were upregulated and 28 miRNAs were downregulated in JTXK-treated group (Tables 2, 3). Information including RPM, base and read length distribution of all miRNAs in this sequencing is available in the Supplementary Material (Supplementary Figures S1–S3). According to the hierarchical clustering plot and bar chart, the differentially expressed mRNAs could be well-categorized into J group and M group (Figure 4). The upregulated mmu-miR-23b-5p and down-regulated mmu-miR-185-5p were selected for qRT-PCR validation and the results were consistent with the sequencing data (Figure 5). The difference between M group and J group was significant, indicating that JTXK granule was capable of modulating the miRNA expression profile in liver tissue of HFD-induced diabetic mice.
TABLE 2.
Upregulated miRNAs between diabetic and JTXK-treated mice in liver tissue.
| ID | miR_name | fold_change | P_value |
| 387225 | mmu-miR-30a-5p* | 1.5344 | 0.0102 |
| 102466970 | mmu-miR-1957b* | 1.6572 | 0.0163 |
| 100124441 | mmu-miR-465c-3p* | 2.6407 | 0.0338 |
| 387217 | mmu-miR-23b-5p* | 1.5081 | 0.0360 |
| 723895 | mmu-miR-31-3p* | 1.7527 | 0.0427 |
*Upregulated DEMs are listed in this table (| log2(Fold change) | > 0.3, P < 0.05). The novel microRNAs are not listed.
TABLE 3.
Downregulated miRNAs between diabetic and JTXK-treated mice in liver tissue.
| ID | miR_name | fold_change | P_value |
| 723821 | mmu-miR-199a-5p* | 0.6483 | 0.0004 |
| 723864 | mmu-miR-425-5p* | 0.6212 | 0.0028 |
| 387210 | mmu-miR-214-3p* | 0.6718 | 0.0032 |
| 724064 | mmu-miR-148b-3p* | 0.6657 | 0.0039 |
| 387247 | mmu-let-7d-5p* | 0.6407 | 0.0047 |
| 387181 | mmu-miR-186-5p* | 0.5131 | 0.0061 |
| 723966 | mmu-let-7c-5p* | 0.7903 | 0.0071 |
| 387244 | mmu-let-7a-5p* | 0.6874 | 0.0081 |
| 387163 | mmu-miR-145a-3p* | 0.6626 | 0.0093 |
| 387180 | mmu-miR-185-5p* | 0.7616 | 0.0108 |
| 751550 | mmu-miR-146b-5p* | 0.3446 | 0.0124 |
| 751519 | mmu-miR-423-5p* | 0.6898 | 0.0178 |
| 100124452 | mmu-miR-598-3p* | 0.6630 | 0.0190 |
| 387186 | mmu-miR-191-5p* | 0.4280 | 0.0220 |
| 723897 | mmu-miR-33-3p* | 0.6995 | 0.0237 |
| 387249 | mmu-let-7g-5p* | 0.7338 | 0.0272 |
| 751545 | mmu-miR-505-3p* | 0.5765 | 0.0288 |
| 387245 | mmu-let-7b-5p* | 0.7833 | 0.0311 |
| 723909 | mmu-miR-342-3p* | 0.3192 | 0.0319 |
| 723827 | mmu-miR-221-5p* | 0.6531 | 0.0324 |
| 102467000 | mmu-miR-292b-5p* | 0.3451 | 0.0336 |
| 723944 | mmu-miR-200c-3p* | 0.8019 | 0.0348 |
| 387163 | mmu-miR-145a-5p* | 0.7240 | 0.0368 |
| 723902 | mmu-miR-7a-1-3p* | 0.6903 | 0.0372 |
| 751522 | mmu-miR-495-3p* | 0.2111 | 0.0387 |
| 723837 | mmu-miR-32-5p* | 0.7937 | 0.0474 |
| 723910 | mmu-miR-351-3p* | 0.6937 | 0.0475 |
| 723823 | mmu-miR-219a-1-3p* | 0.5572 | 0.0490 |
*Downregulated DEMs are listed in this form (| log2(Fold change) | > 0.3, P < 0.05). The novel microRNAs are not listed.
FIGURE 4.

Bar chart (A) and Hierarchical clustering (B) of DEMs in Model group and JTXK-treated group. (A) The abscissa of bar chart is log2(fold change) of DEMs. Red bars represent miRNAs significantly increased in JTXK-treated group, while the green bars represent the downregulated miRNAs (only the top 20 was illustrated). (B) Hierarchical clustering was generated according to the expression levels of miRNA, and a total of six samples were taken from either J group (JTXK-treated group) or M group (Model group). Green, significantly decreased; Red significantly increased.
FIGURE 5.

qRT-PCR validation for miRNA sequencing data. (A) Comparison of mmu-miR-23b-5p and mmu-miR-185-5p data between two groups. (B) Comparison between qRT-PCR and microarray analyses for both mmu-miR-23b-5p and mmu-miR185-5p. Results from these two analyses showed consistency.
Target Gene Prediction and Co-expression Network of Differentially Expressed MiRNAs
In order to identify target genes of DEMs, MiRanda and RNAhybrid were used for target genes prediction, and the intersection of the prediction was considered as the final result. In total, 798 upregulated miRNAs and 3229 downregulated miRNAs were identified. One miRNA was able to target multiple genes, while a single gene was also related to multiple miRNAs. In this way, several relevant miRNAs can construct a network to connect the key genes in a signaling pathway. In present study, in order to better understand the underlying mechanism of JTXK granule, we chose five representative miRNAs to build up the network (Figure 6). Among these five DEMs, two were upregulated (mmu-miR-30a-5p and mmu-miR-23b-5p) and three were downregulated (mmu-miR-199a-5p, mmu-miR-425-5p, mmu-miR-214-3p). The upregulated miRNA mmu-miR-30a-5p was associated with three mRNAs, and mmu-miR-23b-5p linked to 13 mRNAs. Meanwhile, the downregulated miRNAs, including mmu-miR-199a-5p, mmu-miR-425-5p and mmu-miR-214-3p, were associated with 17, 3, and 9 mRNAs, respectively.
FIGURE 6.

The miRNA–mRNA network. Red nodes represent upregulated miRNAs (miR-23b, miR-30a), while green nodes represent downregulated miRNAs (miR-199a, miR-214, miR-425). Blue dots represent mRNAs, while gray solid lines illustrate correlations between miRNAs and mRNAs.
Gene Ontology (GO) Enrichment Analysis
Gene ontology (GO) analysis is a well-known gene function classification system, which can be used to describe the features of genes and its products in organisms. GO enrichment analysis of target genes was shown in Figure 7. To be more specific, Chart A represented target gene number and their functional terms of upregulated miRNAs, and chart B showed the target gene number and functional terms of downregulated miRNAs. All the terms were categorized into biological process (BP), cellular component (CC), and molecular function (MF). As shown in Figure 6, biological regulation (BP), cell (CC) and binding (MF) were the mostly enriched terms of upregulated miRNAs. Meanwhile, the down-regulated miRNAs were mainly enriched in biological regulation (BP), cell and cell part (CC), and binding (MF).
FIGURE 7.

GO enrichment analysis results. Image (A) represents the gene number and GO terms of upregulated miRNAs while the image (B) shows the results from downregulated miRNAs. The abscissa is GO classification which includes biological process, cellular component and molecular function, the secondary function of GO. The ordinate on the left side is the percentage of target genes, while on the right side is the number of target genes.
Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analysis
KEGG enrichment analysis, presented in the form of KEGG terms, could identify the vital biochemical metabolic pathways as well as cellular signaling transduction pathways in which differentially expressed genes (DEGs) participate. In this study, there were 17 upregulated pathways and 41 downregulated pathways in J group compared with M group. The pathways obtained by enrichment analysis were illustrated by bubble diagram (Figure 8). If more than 30 pathways revealed, only the first 30 pathways were included for plotting. According to KEGG enrichment analysis, we confirmed that insulin signaling was significantly upregulated while Ras signaling was inhibited (Figure 9).
FIGURE 8.

KEGG enrichment bubble diagram. (A) Shows the enrichment analysis of upregulated pathways, while (B) shows the results of downregulated pathways. The abscissa is the enrichment factor (the number of differentially enriched target genes of microRNAs/the number of background genes of the pathway). The ordinate is the description of the corresponding pathway. The bubble size is the number of differentially enriched genes, and the bubble color represents the P-value of the enrichment significance.
FIGURE 9.

Insulin signaling and Ras signaling pathways. (A) Identified insulin signaling. Red, target genes of up-regulated miRNA; Green, target genes of down-regulated miRNA; Yellow, target genes of both up and down regulated miRNAs. (B) Identified Ras signaling. Red, target genes of up-regulated miRNA; Green, target genes of down-regulated miRNA; Yellow, target genes of both up and down regulated miRNAs.
Discussion
Diabetes mellitus is a complex metabolic disease, which is affected by both genetic and environmental factors (American Diabetes Association [ADA], 2019). Diabetes can cause dysfunction and destruction in a variety of tissues and organs, eventually leading to different diabetic complications. Liver is an important organ in the process of glycolipid metabolism, and it is also one of the peripheral target tissues of insulin (Dongiovanni et al., 2016). Diabetes or insulin resistance causes excessive accumulation of lipids in liver, abnormal glycogen synthesis and gluconeogenesis, resulting in fatty liver and abnormal glycolipid levels (Garcia-Compean et al., 2009; Sun and Lazar, 2013; Rines et al., 2016). High fat diet induced metabolic disorder will lead to insulin resistance and hyperglycemia. And the ascending of blood glucose level is usually mild, which could be taken as early diabetes (Peyot et al., 2010; Kleinert et al., 2018). Our previous studies demonstrated that JTXK granules can effectively regulate blood glucose and lipid levels, protect islet β-cells, promote insulin secretion, and improve insulin sensitivity. In this study, we found that JTXK granules exert protective effect on liver tissue as well. In order to elucidate the mechanism of this protective effect, we explored the effect of JTXK granules on miRNA profile in hepatic tissue of diabetic mice by miRNA sequencing.
Substantial studies have confirmed that miRNAs exert critical impact on the pathogenesis of obesity, insulin resistance, and diabetes mellitus (Chakraborty et al., 2014; Brandao et al., 2017; Castano et al., 2018). MicroRNAs regulate their target genes and have a great effect on different cellular processes and signaling pathways. Because the changes of miRNAs are earlier than target genes and their expressions are tissue-specific, they are suitable to serve as biomarkers for the diagnosis and treatment of certain diseases (Sud et al., 2017). In this study, JTXK granules significantly affected 33 microRNAs, 5 of which were upregulated and 28 were downregulated. Plenty of differentially expressed genes were obtained by associating with differentially expressed miRNAs. Our results suggested that there were 798 target genes of upregulated miRNAs and 3229 genes of downregulated miRNAs. Through sequencing analysis, we preliminary discovered the action of JTXK granule in regulating miRNA and mRNA expression in diabetic liver. The differentially expressed miRNAs and mRNAs form an interacting feedback network through a series of signal transductions and protein regulations, resulting in the protection of liver physiological structure and modulation of glucose metabolism. To further clarify the key molecular mechanism of JTXK granules, we selected 5 most significantly expressed DEMs (miR-23b, miR-30a, miR-199a, miR-214, miR-425) to construct a miRNA–mRNA co-expression network.
MiR-23b is a microRNA related to immune response. It can inhibit the inflammatory response induced by IL-17, TNF-α or IL-1β, block nuclear Factor-Kappa-B (NF-κB) signaling, and reduce the expression of inflammatory factors. It can also block Ras/mitogen-activated protein kinase (MAPK) signal transduction via targeting G3BP2 in Ras signaling network (Zhao et al., 2016; Wang et al., 2018). In this study, the expression of miR-23b was upregulated, which inhibited the inflammatory response in liver tissue. MiR-30a expression was also increased after treatment with JTXK granule. MiR-30a is known to be actively involved in insulin signaling and is recognized as one of the potential signature miRNAs in diabetes (Karolina et al., 2011). One study showed that miR-30a suppressed Ras/Raf/MEK/ERK signal transduction and thus delayed the progression of HC (Zhou et al., 2017). In fact, approximately 80% of HC are closely related to inflammation (Nakamura et al., 2018). This is due to inflammation stimulated reactive oxygen species over production (Lucy et al., 2014), which may lead to DNA damage and oncogene mutation (Coussens and Werb, 2002; Alessandro et al., 2010). The authors believe that the blockade of Ras signal is mainly associated with the high expression of miR-30a. Furthermore, Li et al. (2018) claimed that miR-199a decreased insulin sensitivity in liver tissue, which was mainly related to the inhibition of autophagy mediated by autophagy-related protein 14 (Atg14). Convincing evidences have demonstrated that autophagy as a regulator of immunity response can inhibit inflammatory reactions (Yang et al., 2013), and is suppressed under the circumstance of insulin resistance and hyperinsulinemia. Therefore, it is obvious that defective autophagy may cause inflammation activation and accelerate the progression of insulin resistance and diabetes (Hui-Yu et al., 2009; Yang et al., 2010). In addition, miR-425 has been shown to be upregulated in liver tissue of HFD-induced obese mice. On the one hand, mir-425 impairs insulin/PI3K/AKT signal transduction; on the other hand, this miRNA is actively involved in the inflammatory response mediated by NF-κB (Kwon et al., 2014; Arora et al., 2016). MiR-214 had been demonstrated to aggravate insulin resistance by inhibiting Akt and Rock1 expression, which indicated its negative impact on insulin signaling (Honardoost et al., 2018). Moreover, NF-κB can promote the expression of miR-214, while the latter can inhibit the expressions of anti-inflammatory factors and promote the release of inflammatory factors such as TNF-α and IL-6, thus aggravating the inflammatory response (Zhao et al., 2015). In the present study, JTXK granules upregulated the expressions of miR-23b, miR-30a, as well as downregulated the expressions of miR-199a, miR-425 and miR-125, which together can improve insulin signaling as well as inhibit Ras signaling. These results provide a strong proof that JTXK granule showed anti-diabetic effect through regulation of these potential targets, which are all closely related to insulin resistance and inflammation.
KEGG analysis showed that JTXK granules improved insulin signaling and inhibited Ras signaling pathway. Insulin signaling is of great significance in glucose metabolism and is rather complicated since it involves many important process such as β-cell development, insulin secretion, downstream signal conduction, and so on. Representative genes or mRNAs include insulin, IRS1, PI3K, Akt, etc. Studies have shown that many miRNAs are capable of modulating protein cascades in insulin signaling, thereby exerting important effect in diabetes (Chakraborty et al., 2014). Ras is a small GTPase that plays an important part in cell growth, proliferation as well as differentiation. After Ras activation, signal can be transmitted downstream via Ras/Raf/MEK/ERK pathway to regulate the expression of various genes and proteins. Ras signal also interacts with insulin and PI3K signal pathway (Chuang et al., 1994; Yonezawa et al., 1994), making it important in cellular metabolism. Experimental data show that inhibition of Ras signal enhances insulin sensitivity and glucose uptake (Mor et al., 2011). Other studies also showed that Ras is closely related to several diabetic complications such as diabetic nephropathy, diabetic erectile dysfunction and diabetic retinopathy, which suggest that Ras signal contributes to the pathogenesis of diabetic complications (Carney, 2016; Fu et al., 2018; Zhang et al., 2018). In addition, Ras signal is able to activate NF-κB signaling pathway, which is considered to promote inflammatory reaction and eventually contribute to hyperglycemia and various diabetic complications (Mcclelland and Phillip, 2014). Taken together, crosstalk among this miRNA–mRNA network played an important role in regulation of insulin and Ras signaling, which shed light on JTXK granule’s anti-diabetic effect.
Conclusion
Our study first suggests that JTXK granule exerts anti-diabetic effect in liver tissue of diabetic mice through multiple molecular targets and signaling pathways by modulating miRNA–mRNA expression profile, especially associated with insulin and Ras signaling. This study improved our understanding on the molecular mechanism of this classic Chinese herbal formula in the management of type 2 diabetes. Meanwhile, miRNAs highlighted in this study may serve as potential biomarkers of diabetes which are useful for diagnosis, prognostic and pharmaceutical design.
Data Availability Statement
The data generated for this study can be found in the Gene Expression Omnibus (GEO) database using accession number GSE145234.
Ethics Statement
The animal study was reviewed and approved by the Animal Care Committee of Beijing University of Chinese Medicine.
Author Contributions
FM, GJ, and SG designed the study. YB and JH prepared all samples for analysis. DZ, YZ, and RD collected and analyzed the data. YB, XB, DG, WH, NY, and FM interpreted the results and wrote the manuscript. All authors gave final approval for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the Science and Technology Developmental Department of Traditional Chinese Materia Medica for the assistance with the herbal medicine authenticity verification.
Funding. This research project was funded by the National Natural Science Foundation of China (No. NSFC81703971), the Graduate Construction project of Beijing Municipal Education Commission (No. 1000062520181) as well as the Key Drug Development Program (No. 2012ZX09103201-005).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2020.00173/full#supplementary-material
RPM distribution of all miRNAs. The abscissa represents the six samples, and the ordinate is log10(RPM + 1). Each section of the abscissa shows 5 statistics: from the top to the bottom are the maximum, the upper quartile, the median, the lower quartile, and the minimum.
Base distribution of all miRNA sequences. The abscissa shows the position, and the ordinate is the percentage of base distribution corresponding to the position.
Read length distribution of all miRNAs. The abscissa is the read length of miRNA, and the ordinate is the number of miRNAs corresponding to the read length in each sample.
Table of plant scientific full name. Plants used in JTXK granule are listed in this table. The left column is the common name and the right column is the scientific full name from Kew MPNS.
References
- Alessandro F., Floriana M., Concetta T., Fortunato C., Carmela L. (2010). Chronic inflammation and oxidative stress in human carcinogenesis. Int. J. Cancer 121 2381–2386. 10.1002/ijc.23192 [DOI] [PubMed] [Google Scholar]
- American Diabetes Association [ADA] (2019). 2. Classification and diagnosis of diabetes: standards of medical care in diabetes-2019. Diabetes Care 42(Suppl. 1), S13–S28. 10.2337/dc19-S002 [DOI] [PubMed] [Google Scholar]
- Arora P., Wu C., Hamid T., Arora G., Agha O., Allen K., et al. (2016). Acute metabolic influences on the natriuretic peptide system in humans. J. Am. Collog. Cardiol. 67 804–812. 10.1016/j.jacc.2015.11.049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandao B. B., Guerra B. A., Mori M. A. (2017). Shortcuts to a functional adipose tissue: the role of small non-coding RNAs. Redox Biol. 12 82–102. 10.1016/j.redox.2017.01.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carney E. F. (2016). Diabetic nephropathy: MiR-23b protects against fibrosis in diabetic nephropathy. Nat. Rev. Nephrol. 12:197. 10.1038/nrneph.2016.22 [DOI] [PubMed] [Google Scholar]
- Castano C., Kalko S., Novials A., Parrizas M. (2018). Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc. Natl. Acad. Sci. U.S.A. 115 12158–12163. 10.1073/pnas.1808855115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakraborty C., Doss C. G., Bandyopadhyay S., Agoramoorthy G. (2014). Influence of miRNA in insulin signaling pathway and insulin resistance: micro-molecules with a major role in type-2 diabetes. Wiley Interdiscip. Rev. RNA 5 697–712. 10.1002/wrna.1240 [DOI] [PubMed] [Google Scholar]
- Cho N. H., Shaw J. E., Karuranga S., Huang Y., da Rocha Fernandes J. D., Ohlrogge A. W., et al. (2018). IDF Diabetes Atlas: global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes. Res. Clin. Pract. 138 271–281. 10.1016/j.diabres.2018.02.023 [DOI] [PubMed] [Google Scholar]
- Chuang L. M., Hausdorff S. F., Myers M. G., Jr., White M. F., Birnbaum M. J., Kahn C. R. (1994). Interactive roles of Ras, insulin receptor substrate-1, and proteins with Src homology-2 domains in insulin signaling in Xenopus oocytes. J. Biol. Chem. 269 27645–27649. [PubMed] [Google Scholar]
- Coussens L. M., Werb Z. (2002). The role of inflammation and liver cancer. Nature 420 860–867. 10.1007/978-3-0348-0837-8_16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dongiovanni P., Rametta R., Meroni M., Valenti L. (2016). The role of insulin resistance in nonalcoholic steatohepatitis and liver disease development–a potential therapeutic target? Expert Rev. Gastroenterol. Hepatol. 10 229–242. 10.1586/17474124.2016.1110018 [DOI] [PubMed] [Google Scholar]
- Forbes J. M., Cooper M. E. (2013). Mechanisms of diabetic complications. Physiol. Rev. 93 137–188. 10.1152/physrev.00045.2011 [DOI] [PubMed] [Google Scholar]
- Fu C. J., Ling Y. J., Yan J. C., Zhao H. Q., Qin Y. H. (2018). Effect of Shuangdan Mingmu capsule on the Ras-Raf-1-MEK-ERK pathway in retinal tissue in rats with diabetic retinopathy. J. Hum. Univ. Chin. Med. 38 728–731. 10.3969/j.issn.1674-070X.2018.07.003 [DOI] [Google Scholar]
- Fukuda T., Hamaguchi M., Kojima T., Hashimoto Y., Ohbora A., Kato T., et al. (2016). The impact of non-alcoholic fatty liver disease on incident type 2 diabetes mellitus in non-overweight individuals. Liver Int. 36 275–283. 10.1111/liv.12912 [DOI] [PubMed] [Google Scholar]
- Garcia-Compean D., Jaquez-Quintana J. O., Gonzalez-Gonzalez J. A., Maldonado-Garza H. (2009). Liver cirrhosis and diabetes: risk factors, pathophysiology, clinical implications and management. World J. Gastroenterol. 15 280–288. 10.3748/wjg.15.280 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honardoost M., Keramati F., Arefian E., Mohammadi Yeganeh S., Soleimani M. (2018). Network of three specific microRNAs influence type 2 diabetes through inducing insulin resistance in muscle cell lines. J. Cell Biochem. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Hui-Yu L., Jianmin H., Cao S. Y., Tao H., Degen Z., Jianbo S., et al. (2009). Hepatic autophagy is suppressed in the presence of insulin resistance and hyperinsulinemia: inhibition of FoxO1-dependent expression of key autophagy genes by insulin. J. Biol. Chem. 284 31484–31492. 10.1074/jbc.M109.033936 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iacomino G., Siani A. (2017). Role of microRNAs in obesity and obesity-related diseases. Genes Nutr. 12:23. 10.1186/s12263-017-0577-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karolina D. S., Armugam A., Tavintharan S., Wong M. T., Lim S. C., Sum C. F., et al. (2011). MicroRNA 144 impairs insulin signaling by inhibiting the expression of insulin receptor substrate 1 in type 2 diabetes mellitus. PLoS One 6:e22839. 10.1371/journal.pone.0022839 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasinski A. L., Slack F. J. (2010). Potential microRNA therapies targeting Ras, NFkappaB and p53 signaling. Curr. Opin. Mol. Ther. 12 147–157. [PubMed] [Google Scholar]
- Kleinert M., Clemmensen C., Hofmann S. M., Moore M. C., Renner S., Woods S. C., et al. (2018). Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 14 140–162. 10.1038/nrendo.2017.161 [DOI] [PubMed] [Google Scholar]
- Kwon D. N., Chang B. S., Kim J. H. (2014). MicroRNA dysregulation in liver and pancreas of CMP-Neu5Ac hydroxylase null mice disrupts insulin/PI3K-AKT signaling. Biomed. Res. Int. 2014:236385. 10.1155/2014/236385 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li B., Wu X. S., Chen H. B., Zhuang C. L., Zhang Z. G., Yao S. S., et al. (2018). miR199a-5p inhibits hepatic insulin sensitivity via suppression of ATG14-mediated autophagy. Cell Death Dis. 9:405 10.1038/s41419-018-0439-437 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu M., Wang J., Zeng J., Cao X., He Y. (2017). Association of NAFLD with diabetes and the impact of BMI changes: a 5-year cohort study based on 18,507 elderly. J. Clin. Endocrinol. Metab. 102:1309. 10.1210/jc.2016-3440 [DOI] [PubMed] [Google Scholar]
- Lucy B. R., Karin W., Harm D. W., Behiye O., Adri V. O., Fernando S., et al. (2014). Decreased serum level of miR-146a as sign of chronic inflammation in type 2 diabetic patients. PLoS One 9:e115209. 10.1371/journal.pone.0115209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manrique C., Lastra G., Sowers J. R. (2014). New insights into insulin action and resistance in the vasculature. Ann. N. Y. Acad. Sci. 1311 138–150. 10.1111/nyas.12395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mcclelland A. D., Phillip K. (2014). microRNA in the development of diabetic complications. Clin. Sci. 126 95–110. 10.1042/CS20130079 [DOI] [PubMed] [Google Scholar]
- Mo F. F., An T., Zhang Z. J., Liu Y. F., Liu H. X., Pan Y. Y., et al. (2017). Jiang Tang Xiao Ke granule play an anti-diabetic role in diabetic mice pancreatic tissue by regulating the mRNAs and MicroRNAs associated with PI3K-Akt signaling pathway. Front. Pharmacol. 8:795 10.3389/fphar.2017.00795 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mor A., Aizman E., George J., Kloog Y. (2011). Ras inhibition induces insulin sensitivity and glucose uptake. PLoS One 6:e21712. 10.1371/journal.pone.0021712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura N., Tatsuo K., Hitomi N., Shunichi M., Tadatoshi T., Masahiko S., et al. (2018). Persistent hepatic inflammation plays a role in hepatocellular carcinoma after sustained virological response in patients with HCV infection. Int. J. Med. Sci. 15 466–474. 10.7150/ijms.23147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peyot M. L., Pepin E., Lamontagne J., Latour M. G., Zarrouki B., Lussier R., et al. (2010). Beta-cell failure in diet-induced obese mice stratified according to body weight gain: secretory dysfunction and altered islet lipid metabolism without steatosis or reduced beta-cell mass. Diabetes Metab. Res. Rev. 59 2178–2187. 10.2337/db09-1452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rines A. K., Sharabi K., Tavares C. D., Puigserver P. (2016). Targeting hepatic glucose metabolism in the treatment of type 2 diabetes. Nat. Rev. Drug Discov. 15 786–804. 10.1038/nrd.2016.151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shukla G. C., Singh J., Barik S. (2011). MicroRNAs: processing, maturation, target recognition and regulatory functions. Mol. Cell Pharmacol. 3 83–92. [PMC free article] [PubMed] [Google Scholar]
- Sud N., Zhang H., Pan K., Cheng X., Cui J., Su Q. (2017). Aberrant expression of microRNA induced by high-fructose diet: implications in the pathogenesis of hyperlipidemia and hepatic insulin resistance. J. Nutr. Biochem. 43:125. 10.1016/j.jnutbio.2017.02.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun Z., Lazar M. A. (2013). Dissociating fatty liver and diabetes. Trends Endocrinol. Metab. 24 4–12. 10.1016/j.tem.2012.09.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian W., Chen L., Zhang L., Wang B., Li X. B., Fan K. R., et al. (2017). Effects of ginsenoside Rg1 on glucose metabolism and liver injury in streptozotocin-induced type 2 diabetic rats. Genet. Mol. Res. 16: gmr16019463. 10.4238/gmr16019463 [DOI] [PubMed] [Google Scholar]
- Wang W., Wang Y., Liu W., van Wijnen A. J. (2018). Regulation and biological roles of the multifaceted miRNA-23b (MIR23B). Gene 642 103–109. 10.1016/j.gene.2017.10.085 [DOI] [PubMed] [Google Scholar]
- Wilczynska A., Bushell M. (2015). The complexity of miRNA-mediated repression. Cell Death Differ. 22 22–33. 10.1038/cdd.2014.112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang L., Li P., Fu S., Calay E. S., Hotamisligil G. S. (2010). Defective Hepatic Autophagy in Obesity Promotes ER Stress and Causes Insulin Resistance. Cell Metab. 11 467–478. 10.1016/j.cmet.2010.04.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang W., Yan-Bo L., Jia-Jing Y., Ying W., Li-Bo Z., Guang-Ying X., et al. (2013). Autophagy regulates inflammation following oxidative injury in diabetes. Autophagy 9 272–277. 10.4161/auto.23628 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yi Z., Mu Q., Na Y., Zhao D., Zuo J., Hong A., et al. (2016). Effect of Jiangtang Xiaoke Granule on hepatic function and oxidative stress in type 2 Diabetic KKAy mice. World Chin. Med. 34 2626–2629. [Google Scholar]
- Yonezawa K., Ando A., Kaburagi Y., Yamamoto-Honda R., Kitamura T., Hara K., et al. (1994). Signal transduction pathways from insulin receptors to Ras. Analysis by mutant insulin receptors. J. Biol. Chem. 269 4634–4640. [PubMed] [Google Scholar]
- Yu N., Fang X., Zhao D. D., Mu Q. Q., Zuo J. C., Ma Y., et al. (2017). Anti-Diabetic Effects of Jiang Tang Xiao Ke Granule via PI3K/Akt Signalling Pathway in Type 2 Diabetes KKAy Mice. PLoS One 12:e0168980. 10.1371/journal.pone.0168980 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Z., Zhang H. Y., Zhang Y., Li H. (2018). Inactivation of the Ras/MAPK/PPARgamma signaling axis alleviates diabetic mellitus-induced erectile dysfunction through suppression of corpus cavernosal endothelial cell apoptosis by inhibiting HMGCS2 expression. Endocrine 63 615–631. 10.1007/s12020-018-1810-1812 [DOI] [PubMed] [Google Scholar]
- Zhao B., Li H., Liu J., Han P., Zhang C., Bai H., et al. (2016). MicroRNA-23b targets Ras GTPase-Activating Protein SH3 domain-binding protein 2 to alleviate Fibrosis and Albuminuria in Diabetic Nephropathy. J. Am. Soc. Nephrol. 27:ASN.2015030300. 10.1681/ASN.2015030300 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao D. D., Yu N., Li X. K., Fang X., Mu Q., Qin P. J., et al. (2014). Antidiabetic and Antioxidative Effect of Jiang Tang Xiao Ke Granule in High-Fat Diet and Low-Dose Streptozotocin Induced Diabetic Rats. Evid. Based Complement. Alternat. Med. 2014:475192. 10.1155/2014/475192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao L., Liu Y. W., Yang T., Gan L., Yang N., Dai S. S., et al. (2015). The mutual regulation between miR-214 and A2AR signaling plays an important role in inflammatory response. Cell. Signal. 27 2026–2034. 10.1016/j.cellsig.2015.07.007 [DOI] [PubMed] [Google Scholar]
- Zhijun W., Jeffrey W., Patrick C. (2013). Treating type 2 diabetes mellitus with traditional chinese and Indian medicinal herbs. Evid. Based Complement. Alternat. Med. 2013:343594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou K., Luo X., Wang Y., Cao D., Sun G. (2017). MicroRNA-30a suppresses tumor progression by blocking Ras/Raf/MEK/ERK signaling pathway in hepatocellular carcinoma. Biomed. Pharmacother. 93 1025–1032. 10.1016/j.biopha.2017.07.029 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
RPM distribution of all miRNAs. The abscissa represents the six samples, and the ordinate is log10(RPM + 1). Each section of the abscissa shows 5 statistics: from the top to the bottom are the maximum, the upper quartile, the median, the lower quartile, and the minimum.
Base distribution of all miRNA sequences. The abscissa shows the position, and the ordinate is the percentage of base distribution corresponding to the position.
Read length distribution of all miRNAs. The abscissa is the read length of miRNA, and the ordinate is the number of miRNAs corresponding to the read length in each sample.
Table of plant scientific full name. Plants used in JTXK granule are listed in this table. The left column is the common name and the right column is the scientific full name from Kew MPNS.
Data Availability Statement
The data generated for this study can be found in the Gene Expression Omnibus (GEO) database using accession number GSE145234.