

Abstract
The present study aimed to investigate the involvement of long intergenic non-coding RNA for kinase activation (LINK-A) long non-coding RNA (lncRNA) in osteosarcoma. Plasma levels of LINK-A lncRNA and transforming growth factor β1 (TGF-β1) were measured by reverse transcription-quantitative polymerase chain reaction and ELISA, respectively. Correlation between LINK-A lncRNA and TGF-β1 was analyzed by Pearson correlation coefficient. LINK-A lncRNA and TGF-β1 were upregulated in patients with osteosarcoma compared with healthy controls. LINK-A lncRNA and TGF-β1 were positively correlated in the two groups. LINK-A lncRNA short hairpin RNAs (shRNAs) were transfected into osteosarcoma cell lines and Transwell migration assay, Matrigel invasion assay and flow cytometry were used to evaluate cell migration, invasion and stemness, respectively. Effects of LINK-A lncRNA silencing and overexpression on TGF-β1 expression were analyzed by western blotting. LINK-A lncRNA shRNA silencing inhibited, whereas TGF-β1 treatment promoted cell migration, invasion and stemness. LINK-A lncRNA silencing inhibited TGF-β1 expression, whereas TGF-β1 treatment had no effects on LINK-A lncRNA expression. TGF-β1 reduced the inhibitory effects of LINK-A lncRNA knockdown on cancer cell migration, invasion and stemness. These data indicated that LINK-A lncRNA is upregulated in osteosarcoma and may regulate migration, invasion and stemness of osteosarcoma cells through TGF-β1.
Keywords: osteosarcoma, long intergenic non-coding RNA for kinase activation, long non-coding RNA, transforming growth factor β, migration, invasion, stemness
Introduction
Osteosarcoma is a type of primary bone tumor that affects approximately four out of one million people in some countries, such as Argentina (1). With the efforts made in treatment of osteosarcoma, survival rate and time of patients with this disease has been markedly improved (2). However, no further significant improvement has been achieved since the 1990s and the overall survival is still poor (3,4). Patients with early stage osteosarcoma usually exhibit satisfactory treatment outcomes, following radical resection. Once distant tumor metastasis occurs surgical resection is not applicable and survival is extremely low (5). At present, active treatment after early diagnosis is still the key to treatment of patients with osteosarcoma.
Transforming growth factor β (TGF-β) is a multifunctional cytokine that regulates the activation of different regulatory proteins and downstream substrates that are involved in cell differentiation, proliferation, chemotaxis and activation of immune cells (6,7). TGF-β is generally considered as a double-edged sword in the development of most types of human malignancies (8); activation of TGF-β signaling inhibits cancer cell proliferation at the initiation of tumorigenesis but promotes tumor cell migration and invasion by mediating epithelial-mesenchymal transition (9). A growing body of literature has revealed that TGF-β signaling achieves signal transduction in cancer biology not only by interacting with regulatory proteins, but also through crosstalk with non-coding RNAs, such as long non-coding RNAs (lncRNAs) (10,11). Long intergenic non-coding RNA for kinase activation (LINK-A) lncRNA is a recently characterized oncogenic lncRNA in triple-negative breast cancer and ovarian carcinoma (12,13). The present study demonstrated that LINK-A lncRNA was upregulated in osteosarcoma and may regulate migration, invasion and stemness of osteosarcoma cells through TGF-β1.
Materials and methods
Human tissues and cell lines
Plasma samples were collected from 66 patients with osteosarcoma and 54 healthy volunteers who were admitted by Jining First People's Hospital (Shizhong, China) between January 2014 and April 2018. The 66 patients with osteosarcoma included 30 males and 26 females, aged between 16 and 32 years (mean age 23.1±2.1 SD, years). There were 12 cases in stage I, 14 cases in stage II, 16 cases in stage III and 24 cases in stage IV. The following inclusion criteria were used: i) Patients diagnosed by biopsies; ii) patients who were diagnosed for the first time; iii) patients who fully understood the experimental protocol and signed informed consent. The exclusion criteria were: i) Patients who were diagnosed with multiple diseases; ii) patients who were treated within 6 months before sample collection. The 54 healthy volunteers included 29 males and 25 females, aged between 17 and 30 years (mean age, 22.9±2.4 years). The two groups had similar age and sex distributions.
MG-63 and U2OS osteosarcoma cell lines were purchased from the American Type Tissue Collection (ATCC; Manassas, VA, USA). Eagle's minimum essential medium (MEM; cat. no. 30-2003; ATCC) supplemented with 10% heat-inactivated fetal bovine serum (FBS, Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was used to cultivate cancer cells under normal conditions (37°C; 5% CO2). In cases of TGF-β1 treatment, cells were treated in medium containing exogenous TGF-β1 (Sigma-Aldrich; Merck KGaA) at doses of 10, 20, 30, 40 and 50 ng/ml for 24 h prior to further experiments.
Cell transfection
Vectors (pcDNA3.1) containing LINK-A lncRNA short hairpin RNA (shRNA 5′-AAAAGCTTCTCTCACCCTTCAAATTGGATCCAATTTGAAGGGTGAGAGAAGC-3′) were designed and synthesized by Shanghai GeneChem Co., Ltd. (Shanghai, China). MISSION shRNA Control Vector was purchased from Sigma-Aldrich (cat. no. SHC001; Merck KGaA). LINK-A (NCBI ID, NR_015407.1) lncRNA overexpression vectors (pcDNA3.1) and empty vectors (pcDNA3.1) were provided by Shanghai Sangong Pharmaceutical Co., Ltd. (Shanghai, China). Lipofectamine® 2000 reagent (cat. no. 11668-019; Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) was used to transfect 15 nM vectors into 5×105 MG-63 and U2OS cells. Transfections were performed at 37°C for 6 h. Non-transfected cells were used as control cells. Cells transfected with empty vectors were used as negative transfection control cells. Transfection efficiency was determined by reverse transcription-quantitative polymerase chain reaction (RT-qPCR). Cells were harvested 24 h post-transfection prior to subsequent experiments.
RT-qPCR
Total RNA extraction from 0.3 ml plasma and in vitro cultivated cells (1×105) was performed using Monarch® Total RNA Miniprep kit (New England Biolabs, Inc., Ipswich, MA, USA). Reverse transcription was performed using High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Inc.) to synthesize cDNA. PCR reaction systems were prepared using Luna® Universal One-Step RT-qPCR kit (SYBR; New England Biolabs, Inc.). CFX96 Touch™ Real-Time PCR detection system (Bio-Rad Laboratories, Inc., Hercules, CA, USA) was used to perform all qPCR reactions. The qPCR thermocycling conditions were as follows: Initial denaturation at 95°C for 52 sec, followed by 40 cycles of 95°C for 14 sec and 58.5°C for 26 sec. Sequences of primers used were as follows: Human LINK-A, forward 5′-TTCCCCCATTTTTCCTTTTC-3′, reverse 5′-CTCTGGTTGGGTGACTGGTT-3′; β-actin, forward 5′-GACCTCTATGCCAACACAGT-3′, reverse 5′-AGTACTTGCGCTCAGGAGGA3′. This experiment was performed in triplicate, and all quantitation cycle values were normalized to β-actin and relative expression was quantified by the 2−ΔΔCq method (14).
ELISA
TGF-β1 in 0.3 ml plasma was detected using Human TGF-β1 Quantikine ELISA Kit (cat. no. DB100B; R&D Systems, Inc.). All operations were performed following manufacturer's protocol. Optical density values were detected at 540 nm.
Transwell migration and invasion assays
QCM Chemotaxis Cell Migration assay, 24-well (8 µm), colorimetric (cat. no. ECM508; Sigma-Aldrich; Merck KGaA), and QCM ECMatrix Cell Invasion assay, 24-well (8 µm), colorimetric (cat. no. ECM550; Sigma-Aldrich; Merck KGaA), were used. In cases of TGF-β1 treatment, 10 ng/ml TGF-β1 (Sigma-Aldrich; Merck KGaA) was added to the MEM according to manufacturer's instructions. Briefly, serum-free cell suspensions (3×104 cells/ml) were made and 0.1 ml of a cell suspension was transferred to the upper chamber of the Transwell plates. Culture medium containing 20% FBS was added into the lower chamber. Cells were cultivated for 24 h at 37°C. Membranes were cleaned using a cotton swab, followed by staining with 0.5% crystal violet (Sigma-Aldrich; Merck KGaA) for 20 min at room temperature. This protocol was used for both invasion and migration assays; prior to the invasion assay the upper chamber was coated with Matrigel (cat. no. 356234; EMD Millipore, Billerica, MA, USA). The experiments were performed in triplicate. Cells were observed and counted under Olympus CX43 light microscope (×40 magnification).
Flow cytometry
MG-63 and U2OS cells (3×105) were trypsinized, harvested, and incubated with phycoerythrin (PE)-conjugated CD133 (1:1,500; cat. no. 566593; BD Biosciences, San Jose, CA, USA) or immunoglobulin (Ig) G1-PE antibody (1:1,500; cat. no. 130-093-193; Miltenyi Biotec, Bergisch Gladbach, Germany) in buffer (1X PBS + 0.5% BSA, Sigma-Aldrich; Merck KGaA) for 20 min at 4°C. Signals were detected using a FACS Aria flow cytometry system (BD Immunocytometry Systems, San Jose, CA, USA) and processed by Cell Quest software v5.1 (BD Biosciences). This experiment was performed in triplicate.
Western blotting
Total protein was extracted from in vitro cultivated cells (1×105) using the Total Protein Extraction kit provided by Merck KGaA (cat. no. 2140). Protein concentrations were measured using BCA assay (Sangon Biotech Co., Ltd., Shanghai, China). Proteins (30 µg per lane) were separated by 12% SDS-PAGE and gel transferred to PVDF membranes. Blocking was performed using PBS containing 5% skimmed milk for 1 h at room temperature. Primary antibodies against TGF-β1 (cat. no. ab9758; rabbit anti-human; 1:1,200; Abcam, Shanghai, China) and GAPDH (ab9485; rabbit anti-human; 1:1,400; Abcam) were used at 4°C for 18 h. The secondary antibody was horseradish peroxidase-conjugated goat anti-rabbit IgG (cat. no. MBS435036; 1:1,000; MyBioSource, Inc., San Diego, CA, USA). ECL™ western blotting reagents (Sigma-Aldrich; Merck KGaA) were added to develop signals. ImageJ software v.1.46 (National Institutes of Health, Bethesda, MD, USA) was used for densitometry analysis. TGF-β1 expression was normalized to GAPDH. This experiment was performed in triplicate.
Statistical analysis
Data are presented as the mean ± standard deviation; values were calculated and processed using GraphPad Prism 6 software (GraphPad Software, Inc., La Jolla, CA, USA). Pearson correlation coefficient was used for correlation analysis. Student's t-test was used for comparisons between two groups. One-way analysis of variance followed by Tukey test was used for comparisons among multiple groups. Diagnostic analysis was performed by receiver operating characteristics (ROC) curve. P<0.05 was considered to indicate a statistically significant difference.
Results
LINK-A lncRNA and TGF-β1 are upregulated in patients with osteosarcoma compared with healthy controls
RT-qPCR results revealed that, compared with healthy controls, plasma levels of LINK-A lncRNA were significantly increased in patients with osteosarcoma (Fig. 1A). Similarly, ELISA results demonstrated that plasma levels of TGF-β1 were significantly higher in patients with osteosarcoma compared with levels in healthy controls (Fig. 1B).
Figure 1.

LINK-A lncRNA and TGF-β1 are upregulated in patients with osteosarcoma compared with healthy controls. (A and B) Compared with healthy controls, plasma levels of (A) LINK-A lncRNA (B) and TGF-β1 were significantly increased in patients with osteosarcoma. *P<0.05. LINK-A, long intergenic non-coding RNA for kinase activation; lncRNA, long non-coding RNA; TGF-β1, transforming growth factor β1.
Plasma LINK-A lncRNA and TGF-β1 are positively correlated in patients with osteosarcoma and in healthy controls
Pearson correlation coefficient analysis demonstrated that plasma levels of LINK-A lncRNA and TGF-β1 were positively correlated in both osteosarcoma patients (Fig. 2A) and in healthy controls (Fig. 2B).
Figure 2.

Plasma LINK-A lncRNA and TGF-β1 are positively correlated in both patients with osteosarcoma and healthy controls. (A and B) Plasma levels of LINK-A lncRNA and TGF-β1 were positively correlated in (A) patients with osteosarcoma and (B) healthy controls. LINK-A, long intergenic non-coding RNA for kinase activation; lncRNA, long non-coding RNA; TGF-β1, transforming growth factor β1.
Plasma LINK-A lncRNA has diagnostic potentials for early stage osteosarcoma
Among the 66 patients with osteosarcoma, there were 26 cases in stage I and II, which are considered as the early stages of cancer development. ROC curve analysis was performed to evaluate the diagnostic value of plasma LINK-A lncRNA for early stage osteosarcoma. The area under the curve was 0.8877, with standard error of 0.03549 and 95% confidence interval of 0.8182–0.9673 (P<0.0001; Fig. 3).
Figure 3.
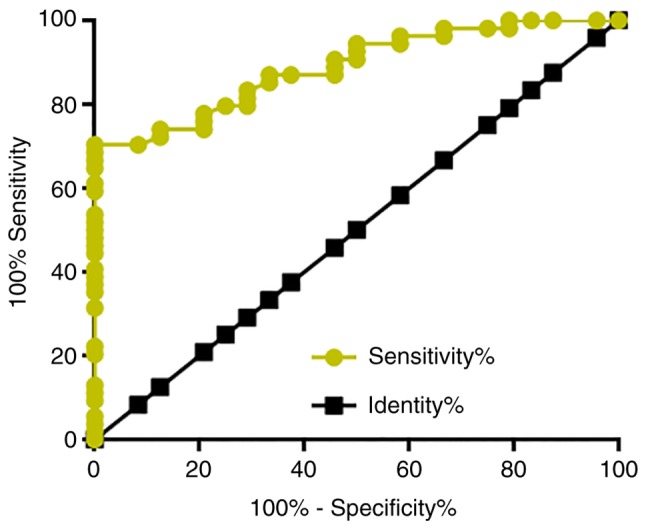
Plasma LINK-A lncRNA has diagnostic potentials for early stage osteosarcoma. ROC curve was performed to analyze the diagnostic value of LINK-A for osteosarcoma. Patients with early stage osteosarcoma were true positive cases and the healthy controls were true negative cases. LINK-A, long intergenic non-coding RNA for kinase activation; lncRNA, long non-coding RNA.
LINK-A lncRNA positively regulates TGF-β1 in MG-63 and U2OS osteosarcoma cell lines
RT-qPCR analysis was used to confirm that LINK-A lncRNA expression was decreased or increased following shRNA or overexpression vector transfection, respectively, in MG-63 and U2OS osteosarcoma cell lines compared with the control groups (Fig. 4A). LINK-A lncRNA shRNA mediated the downregulation, whereas LINK-A lncRNA overexpression mediated the upregulation of TGF-β1 expression in the two osteosarcoma cell lines (Fig. 4B). By contrast, exogenous TGF-β1 treatment at doses of 10, 20, 30, 40 and 50 ng/ml failed to significantly alter the expression of LINK-A lncRNA in these cells (Fig. 4C).
Figure 4.

LINK-A lncRNA knockdown mediates TGF-β1 inhibition in MG-63 and U2OS osteosarcoma cell lines. (A) LINK-A lncRNA knockdown and overexpression were achieved in MG-63 and U2OS osteosarcoma cell lines following transfection. (B) LINK-A lncRNA shRNA mediated the downregulation, whereas LINK-A lncRNA overexpression mediated the upregulation of TGF-β1 in MG-63 and U2OS osteosarcoma cell lines. (C) TGF-β1 treatment at doses of 10, 20, 30, 40 and 50 ng/ml failed to significantly alter the expression of LINK-A lncRNA. *P<0.05. C, control; LINK-A, long intergenic non-coding RNA for kinase activation; lncRNA, long non-coding RNA; NC, negative control; TGF-β1, transforming growth factor β1.
LINK-A lncRNA knockdown inhibits migration, invasion and stemness of MG-63 and U2OS cells
Compared with the control groups, LINK-A lncRNA knockdown significantly inhibited, whereas treatment with TGF-β1 at a dose of 10 ng/ml significantly promoted the migration (Fig. 5A), invasion (Fig. 5B) and stemness (reflected by percentage of CD133+ cells; Fig. 5C) of cells of MG-63 and U2OS osteosarcoma cell lines. In addition, exogenous TGF-β1 significantly attenuated the effects of LINK-A lncRNA knockdown (Fig. 5).
Figure 5.

LINK-A lncRNA knockdown mediates the inhibition of migration, invasion and stemness of MG-63 and U2OS osteosarcoma cell lines through TGF-β1. (A-C) LINK-A lncRNA knockdown significantly inhibited, whereas treatment with TGF-β1 (10 ng/ml) significantly promoted the (A) migration, (B) invasion and (C) stemness of cells of MG-63 and U2OS osteosarcoma cell lines. In addition, exogenous TGF-β1 co-treatment significantly attenuated these effects of LINK-A lncRNA knockdown. For invasion and migration data, C group was set to ‘100’ and all other groups were normalized to C. *P<0.05. C, control; LINK-A, long intergenic non-coding RNA for kinase activation; lncRNA, long non-coding RNA; NC, negative control; TGF-β1, transforming growth factor β1.
Discussion
LINK-A lncRNA is a recently characterized lncRNA with oncogenic functionality in triple-negative breast cancer and ovarian carcinoma (12,13). The key finding of the present study was that LINK-A lncRNA may also be an oncogenic lncRNA in osteosarcoma and may participate in cell migration, invasion and stemness. The actions of LINK-A lncRNA were at least partially mediated by TGF-β1.
It has been reported that the development of osteosarcoma affects the expression of a large set of genes in the human body, including lncRNAs (15,16). In the present study LINK-A lncRNA was significantly upregulated in plasma from patients with osteosarcoma compared with healthy controls. A recent study has demonstrated that LINK-A lncRNA is involved in the regulation of migration and invasion of ovarian carcinoma cells (13). The present study revealed that inhibition of LINK-A lncRNA inhibited migration and invasion and reduced stemness of cancer cells in osteosarcoma in vitro. Therefore, inhibition of LINK-A lncRNA may serve as a potential therapeutic target for osteosarcoma. It is also worth mentioning that inhibition of LINK-A lncRNA exhibited no significant effects on osteosarcoma cell proliferation (data not shown), which indicated that LINK-A lncRNA is unlikely to be involved in the regulation of proliferation of osteosarcoma, which is consistent with the previous study (13).
Early diagnosis is crucial for the survival of patients with osteosarcoma. The present study enrolled 26 patients with osteosarcoma at stages I and II. ROC curve analysis revealed that the upregulation of LINK-A lncRNA distinguished patients with early stage osteosarcoma from healthy controls. Therefore, circulating LINK-A lncRNA may be a potential marker for the early diagnosis of osteosarcoma. However, clinical trials are needed to further test the applicability.
Activation of TGF-β is frequently observed in cancer development (17,18). Inhibition of TGF-β signaling is considered a promising therapeutic target for cancers (19). The present study also demonstrated significantly higher plasma levels of TGF-β1 in patients with osteosarcoma compared with healthy controls. It is known that TGF-β signals in cancer by interacting with lncRNAs (10,11). The present study indicated that LINK-A lncRNA may be upstream of TGF-β1 in the regulation of migration, invasion and stemness of osteosarcoma. As the plasma levels of LINK-A lncRNA and TGF-β1 were revealed to be positively correlated in both osteosarcoma and healthy controls, the interaction between LINK-A lncRNA and TGF-β1 is unlikely to be disease-specific. However, whether this interaction is direct or indirect is still unknown. LINK-A lncRNA overexpression and knockdown failed to affect TGF-β1 expression at the mRNA level (data not shown); therefore, LINK-A lncRNA may affect TGF-β1 accumulation or degradation, but not gene transcription.
In conclusion, LINK-A lncRNA and TGF-β1 were both upregulated in osteosarcoma. LINK-A lncRNA may regulate TGF-β1 to participate in the regulation of migration, invasion and stemness of cancer cells in osteosarcoma.
Acknowledgements
Not applicable.
Funding
No funding was received
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Authors' contributions
YK did the experiments, analyzed the data and was a major contributor in writing the manuscript. ZN, CM and HG performed the experiments and literature reviews, and analyzed data. All authors read and approved the final manuscript and all authors should confirm its accuracy.
Ethics approval and consent to participate
Ethical approval was obtained from the Ethics Committee of Jining First People's Hospital, Shizhong, Jining, China. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and national research committees, the 1964 Declaration of Helsinki and its later amendments. Informed written consent was obtained from all patients and controls following an explanation of the nature and possible consequences of the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Moreno F, Cacciavillano W, Cipolla M, Coirini M, Streitenberger P, López Martí J, Palladino M, Morici M, Onoratelli M, Drago G, et al. Childhood osteosarcoma: Incidence and survival in Argentina. Report from the national pediatric cancer registry, ROHA network 2000–2013. Pediatr Blood Cancer. 2017;64 doi: 10.1002/pbc.26533. [DOI] [PubMed] [Google Scholar]
- 2.Durfee RA, Mohammed M, Luu HH. Review of osteosarcoma and current management. Rheumatol Ther. 2016;3:221–243. doi: 10.1007/s40744-016-0046-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Anderson ME. Update on survival in osteosarcoma. Orthop Clin North Am. 2016;47:283–292. doi: 10.1016/j.ocl.2015.08.022. [DOI] [PubMed] [Google Scholar]
- 4.Berner K, Johannesen TB, Berner A, Haugland HK, Bjerkehagen B, Bøhler PJ, Bruland ØS. Time-trends on incidence and survival in a nationwide and unselected cohort of patients with skeletal osteosarcoma. Acta Oncol. 2015;54:25–33. doi: 10.3109/0284186X.2014.923934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Slade AD, Warneke CL, Hughes DP, Lally PA, Lally KP, Hayes-Jordan AA, Austin MT. Effect of concurrent metastatic disease on survival in children and adolescents undergoing lung resection for metastatic osteosarcoma. J Pediatr Surg. 2015;50:157–160. doi: 10.1016/j.jpedsurg.2014.10.038. [DOI] [PubMed] [Google Scholar]
- 6.Nakao A, Afrakhte M, Morén A, Nakayama T, Christian JL, Heuchel R, Itoh S, Kawabata M, Heldin NE, Heldin CH, ten Dijke P. Identification of Smad7, a TGFbeta-inducible antagonist of TGF-beta signalling. Nature. 1997;389:631–635. doi: 10.1038/39369. [DOI] [PubMed] [Google Scholar]
- 7.Soderberg SS, Karlsson G, Karlsson S. Complex and context dependent regulation of hematopoiesis by TGF-beta superfamily signaling. Ann N Y Acad Sci. 2009;1176:55–69. doi: 10.1111/j.1749-6632.2009.04569.x. [DOI] [PubMed] [Google Scholar]
- 8.Akhurst RJ, Derynck R. TGF-beta signaling in cancer-a double-edged sword. Trends Cell Biol. 2001;11:S44–S51. doi: 10.1016/S0962-8924(01)02130-4. [DOI] [PubMed] [Google Scholar]
- 9.Willis BC, Borok Z. TGF-beta-induced EMT: Mechanisms and implications for fibrotic lung disease. Am J Physiol Lung Cell Mol Physiol. 2007;293:L525–L534. doi: 10.1152/ajplung.00163.2007. [DOI] [PubMed] [Google Scholar]
- 10.Mondal T, Subhash S, Vaid R, Enroth S, Uday S, Reinius B, Mitra S, Mohammed A, James AR, Hoberg E, et al. MEG3 long noncoding RNA regulates the TGF-β pathway genes through formation of RNA-DNA triplex structures. Nat Commun. 2015;6:7743. doi: 10.1038/ncomms8743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li Z, Dong M, Fan D, Hou P, Li H, Liu L, Lin C, Liu J, Su L, Wu L, et al. lncRNA ANCR down-regulation promotes TGF-β-induced EMT and metastasis in breast cancer. Oncotarget. 2017;8:67329–67343. doi: 10.18632/oncotarget.18622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lin A, Li C, Xing Z, Hu Q, Liang K, Han L, Wang C, Hawke DH, Wang S, Zhang Y, et al. The LINK-A lncRNA activates normoxic HIF1α signalling in triple-negative breast cancer. Nat Cell Biol. 2016;18:213–224. doi: 10.1038/ncb3295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ma J, Xue M. LINK-A lncRNA promotes migration and invasion of ovarian carcinoma cells by activating TGF-β pathway. Biosci Rep. 2018;38(pii):BSR20180936. doi: 10.1042/BSR20180936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 15.Atiye J, Wolf M, Kaur S, Monni O, Böhling T, Kivioja A, Tas E, Serra M, Tarkkanen M, Knuutila S. Gene amplifications in osteosarcoma-CGH microarray analysis. Genes Chromosomes Cancer. 2005;42:158–163. doi: 10.1002/gcc.20120. [DOI] [PubMed] [Google Scholar]
- 16.Li JP, Liu LH, Li J, Chen Y, Jiang XW, Ouyang YR, Liu YQ, Zhong H, Li H, Xiao T. Microarray expression profile of long noncoding RNAs in human osteosarcoma. Biochem Biophys Res Commun. 2013;433:200–206. doi: 10.1016/j.bbrc.2013.02.083. [DOI] [PubMed] [Google Scholar]
- 17.Solar M, Paul W. Chain relaxation in thin polymer films: Turning a dielectric type-B polymer into a type-A' one. Soft Matter. 2017;13:1646–1653. doi: 10.1039/C6SM02557A. [DOI] [PubMed] [Google Scholar]
- 18.Hawinkels LJ, Paauwe M, Verspaget HW, Wiercinska E, van der Zon JM, van der Ploeg K, Koelink PJ, Lindeman JH, Mesker W, ten Dijke P, Sier CF. Interaction with colon cancer cells hyperactivates TGF-β signaling in cancer-associated fibroblasts. Oncogene. 2014;33:97–107. doi: 10.1038/onc.2012.536. [DOI] [PubMed] [Google Scholar]
- 19.Colak S, Ten Dijke P. Targeting TGF-β signaling in cancer. Trends Cancer. 2017;3:56–71. doi: 10.1016/j.trecan.2016.11.008. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.