

Abstract
Abstract
TNF is highly expressed in synovial tissue of rheumatoid arthritis (RA) patients, where it induces proinflammatory cytokine secretion. However, in other cases, TNF will cause cell death. Considering the abnormal proliferation and activation of rheumatoid arthritis synovioblasts, the proper rate of synovioblast apoptosis could possibly relieve arthritis. However, the mechanism mediating TNF-induced synovioblast survival versus cell death in RA is not fully understood. Our objective was to study the role of CD147 in TNF downstream pathway preference in RA synovioblasts. We found that overexpressing TNF in synovial tissue did not increase the apoptotic level and, in vitro, TNF-induced mild synovioblast apoptosis and promoted IL-6 secretion. CD147, which was highly expressed in rheumatoid arthritis synovial fibroblasts (RASFs), increased the resistance of synovioblasts to apoptosis under TNF stimulation. Downregulating CD147 both increased the apoptotic rate and inhibited IκB kinase (IKK)/IκB/NF-κB pathway-dependent proinflammatory cytokine secretion. Further, we determined that it was the extracellular portion of CD147 and not the intracellular portion that was responsible for synovioblast apoptosis resistance. CD147 monoclonal antibody inhibited TNF-induced proinflammatory cytokine production but had no effect on apoptotic rates. Thus, our study indicates that CD147 is resistant to TNF-induced apoptosis by promoting IKK/IκB/NF-κB pathway, and the extracellular portion of CD147 is the functional region.
Key messages
CD147 inhibits TNF-stimulated RASF apoptosis.
CD147 knockdown decreases IKK expression and inhibits NF-κB-related cytokine secretion.
CD147’s extracellular portion is responsible for apoptosis resistance.
CD147 antibody inhibits TNF-related cytokine secretion without additional apoptosis.
Keywords: Rheumatoid arthritis synovial fibroblast, TNF, NF-κB, CD147, Apoptosis
Introduction
Rheumatoid arthritis (RA) is a chronic autoimmune inflammatory disease with severe inflammatory symptoms in joints, mostly in hands, feet, and knees [1]. RA affects approximately 1 % of the population, with high incidence in women and old people [2]. Abnormal accumulation of fibroblasts, macrophages, and lymphocytes in synovial joints finally causes the destruction of the bone and cartilage [3]. Rheumatoid arthritis synovial fibroblasts (RASFs) are regarded as direct inducers of such damage. They receive molecular signals from macrophages and lymphocytes, and they secrete metalloproteinases (MMPs), IL-6, IL-8, and granulocyte–macrophage colony-stimulating factor (GM-CSF) among others [4]. One of the most important upstream signals for RASFs is TNF, a highly expressed cytokine in both serum and synovium of RA patients [5].
TNF is a type II transmembrane homotrimer that is released in a soluble form through proteolytic cleavage by TNF-converting enzyme (TACE) [6]. Mainly secreted by synovial macrophages, TNF is considered to be an important factor in the pathogenesis of rheumatoid arthritis, which causes intractable inflammation in joints. Infliximab, a TNF antibody, has already proved its effectiveness in treating RA patients [7]. However, with different cell types, various activation states, and a variety of microenvironment factors, TNF can contribute to either proinflammatory cytokine secretion or cell death [8]. The TNF receptor is known as one of the death receptors, and it can receive death signals given by TNF ligands and further induce caspase-dependent apoptosis or caspase-independent necroptosis (a type of programmed necrosis) [9]. The apoptotic rate of RASFs under TNF stimulation could be significant, because it could both decrease the number of effective RASFs and inhibit TNF-related immune activation. With two different downstream signaling pathways, NF-κB-dependent or NF-κB-independent cell survival versus caspase-related apoptosis [10], TNF signaling could be influenced by certain other important molecules that help to ‘switch’ between different patterns [11]. Although TNF has been studied for a long time, the mechanism underlying downstream-pattern preference is still unclear. Elucidating the TNF-related cell death pathway in synovioblasts should increase our understanding of RA and may suggest treatments.
CD147, also called extracellular matrix metalloproteinase inducer (EMMPRIN) and leukocyte activation-associated M6 antigen or HAb18G, is a transmembrane glycoprotein [12]. CD147 is a receptor for cyclophilin A (CypA) and PfRh5, and it contributes to tumor metastasis, inflammation, and pathogen (plasmodium falciparum and coronavirus) infection [13–20]. Our previous works have proven that CD147 is upregulated in RA synoviocytes, including RASFs and macrophage-like cells [21]. CD147 overexpression can increase MMP production and invasiveness in synoviocytes. In addition, CD147 on RASFs induces the upregulation of VEGF and HIF-1, which further promotes angiogenesis in synovium and causes persistence of synovitis [22]. CD147/HAb18 monoclonal antibody can inhibit cartilage erosion and synovitis in the SCID mouse model of rheumatoid arthritis [23]. However, so far, the role of CD147 in TNF-induced RASF apoptosis has not been studied. It was reported that in hepatocellular carcinoma (HCC), CD147 inhibits HCC cell apoptosis under endoplasmic reticulum (ER) stress [24]. Others have shown that TNF is the key in the cross talk between ER stress and oxidative stress [25]. These data highlight the possibility that CD147 plays a negative role in TNF-related apoptosis.
In our report, we investigated the function of CD147 in TNF signaling pathway preference in RA synovioblasts, and further, we attempted to identify the functional region of CD147 that is responsible for this preference.
Materials and methods
Patients and tissue preparation
Twenty-five RA patients and 14 osteoarthritis (OA) patients were involved in the experiments (informed consent had been obtained). All the patients with RA and OA satisfied the diagnostic criteria of the American College of Rheumatology [26]. The mean age of the RA patients was 48 years, ranging from 39 to 57 years (female/male = 1.78); the mean age of the OA patients was 53 years, ranging from 43 to 61 years (female/male = 0.75). Synovial tissues were obtained from the RA and OA patients during joint arthroscopy in Xijing Hospital (Xi’an, China). The ethics approval for this study was granted by Ethical Committee of Fourth Military Medical University.
Cell culture
Primary RA synovial fibroblasts (synoviocyte B) were isolated from synovium of RA patients following a manner previously described [22]. To have pure and biologically functional fibroblasts, the primary fibroblasts used here were at third to fifth passage. RASFs cultured in DMEM high-glucose medium (with 10 % fetal bovine serum) in regular culture conditions.
shRNA lentivirus
The Trans-Lentiviral pLKO System was used to construct the stable CD147 knockdown synovioblasts RASF A6, and RASF NC was RASF transfected with lentivirus without the CD147 short hairpin RNA (shRNA) sequence. The construction of the lentivirus carrying CD147 shRNA was described previously [27]. Namely, the plasmids of Trans-Lentiviral pLKO System (psPAX2 1.125 μg/mL, pMD2G 0.125 μg/mL, pLKO A6/NC 1.25 μg/mL) were transfected into HEK 293T cells, and the medium (containing lentivirus) of HEK 293T cells was collected and then cultured with RASFs. Puromycin at 4 μg/mL (Sigma, Buchs, Switzerland) was added to the medium for further selection (10 days), and the successfully transfected RASF A6 and RASF NC cells were resistant to puromycin, while the blank control group RASF CTL cells were killed by puromycin at this concentration. To obtain stable cell clones, RASF A6 and RASF NC cells were maintained in medium containing 1 μg/mL puromycin for at least 2 weeks.
Transfection of CD147 extracellular and intracellular portions into RASF A6
The vector pcDNA 3.1 was used to carry the full length, extracellular portion, and intracellular portion of CD147. The exogenous CD147 has 269 amino acids, in which 0–229 is the extracellular portion and 206–269 is the intracellular portion (both containing transmembrane region). The CD147 shRNA sequence A6 targeted 3′-UTR region of CD147 messenger RNA (mRNA) and will not inhibit the exogenous extracellular portion and intracellular portion of CD147 expression which do not contain 3′-UTR sequence. Transfection followed the instructions for Lipofectamine 2000 (Invitrogen, Basel, Switzerland), and concentration of vectors was 5 μg/mL. Cells were tested and used 36 h after the transfection.
Immunohistochemistry assay
Synovial tissue immunohistochemical staining was performed using a streptavidin–peroxidase kit (Zymed, San Diego, USA). The primary antibodies were anti-TNF mouse mAb (Proteintech, Wuhan, China, 1:100), anti-CD147 mouse mAb (prepared in our laboratory, 1:200, final concentration 5 μg/mL), and anti-cleaved-caspase 3 (CC3) rabbit polyclonal Ab (Cell Signaling, Danvers, USA, 1:200). After primary antibody incubation (4 °C overnight), sections were reacted with biotin-labeled goat anti-mouse/rabbit IgG, horseradish peroxidase-labeled streptavidin, and diaminobenzidine (Zymed, San Diego, USA), respectively. Then, nuclei were restained with hematoxylin. Irrelevant mouse anti-human IgG (Biolegend, San Diego, USA) and rabbit serum were used as negative controls. The TNF and cleaved-caspase 3 quantitative analysis was performed by Image-Pro Plus Version 6.0 software. Three random fields of each section were visualized. The average number of cleaved-caspase 3-positive cells in three fields was considered to be the value of CC3 positive, and the average integral optical density summary (IOD SUM) was used to measure TNF expression.
Immunofluorescence microscopy
Cells were attached to glass coverslips for 24 h, and then the coverslips were placed in a 6-well plate. After TNF (R&D system, Abingdon, UK) treatment for 24 h, the cells were fixed in 4 % formaldehyde in PBS, permeabilized with 0.1 % Triton X-100 for 20 min and blocked with goat serum in PBS for 1 h. The coverslips were then incubated with the CD147 (prepared in our laboratory, 1:200, final concentration 5 μg/mL) and cleaved-caspase 3 (Cell Signaling, Danvers, USA, 1:300) antibodies for 12 h. The cells were washed in PBS, incubated with their corresponding FITC and APC secondary antibodies (Thermo scientific, Rockford, USA, 1:200) for 1 h, and nuclei were stained with DAPI. Finally, the cells were detected with a Nikon fluorescence microscope.
Flow cytometry assay
CD147 expression in the RASFs was detected with flow cytometry as previously described [21]. To assess apoptosis, TNF-stimulated and unstimulated cells were collected and washed in PBS and stained for annexin V and propidium iodide (PI) with the Annexin V-FITC Apoptosis Detection Kit (Millipore, Boston, USA) following the instructions. The cells were analyzed by FACSCalibur flow cytometry (BD Pharmingen, San Diego, USA). The data were processed using FlowJo software.
Real-time PCR assay
After different treatments, mRNAs of RASFs were isolated by E.Z.N.A. Total RNA Kit II (OMEGA BioTek, Norcross, USA) in RNase-free conditions. Then, reverse transcription of the mRNA used the TaKaRa PrimeScript® RT reagent kit following the manufacturer’s instructions. The real-time quantitative RT-PCR 25 μL system included the following: SYBR 10 μL (TaKaRa, Otsu, Japan), primers 1 μL each (forward and reverse primers), cDNA template 3 μL, and ddH2O 10 μL. GAPDH mRNA was used to normalize the target mRNA. The relative quantity in each experiment was automatically calculated by comparative quantitation mode of MxPro qPCR system software. At least three independent experiments were performed and analyzed. Relative expression is the ratio to control mRNA. Primer sequences: huGAPDH forward 5′- GCACCGTCAAGGCTGAGAAC-3′, huGAPDH reverse 5′-TGGTGAAGACGCCAGTGGA-3′; huCD147 forward 5′-ACTCCTCACCTGCTCCTTGA-3′, huCD147 reverse 5′-GCCTCCATGTTCAGGTTCTC-3′; huIL-6 forward 5′- CAATAACCACCCCTGACCCAA-3′, huIL-6 reverse 5′-TCTGAGGTGCCCATGCTACA-3′; huIL-1β forward 5′-AAACAGATGAAGTGCTCCTTCCAGG-3′, huIL-1β reverse 5′- TGGAGAACACCACTTGTTGCTCCA-3′; huIL-8 forward 5′-CATACTCCAAACCTTTCCACCCC-3′, huIL-8 reverse 5′-TCAGCCCTCTTCAAAAACTTCTCCA-3′; huCD147EP forward 5′- CTTGAATGACAGCGCCACAG-3′, huCD147EP reverse 5′- GGAGTACTCTCCCCACTGGT-3′; and huCD147IP forward 5′- GACGTCCTGGATGATGACGA-3′, huCD147IP reverse 5′- CGGACGTTCTTGCCTTTGTC-3′, huIKK forward 5′-GGAAGTACCTGAACCAGTTTGAG-3′, huIKK reverse 5′-GCAGGACGATGTTTTCTGGCT-3′.
Cytokine ELISA
Serum-free medium from RASF samples was collected and centrifuged at 10,000g for 5 min. IL-6 release from RASFs into the culture medium was directly measured with a Human IL-6 ELISA Kit and Human IL-1β ELISA Kit (DAKEWE, Shenzhen, China) according to the manufacturer’s instruction. Optical density was determined with a Bio-Rad Microplate reader, and absorption was measured at 450 nm. A standard curve for each measurement was established using a cytokine standard provided by the ELISA Kit.
Western blot assay
RASF CTL, RASF NC, and RASF A6 cells treated with TNF or TPCK were collected using 100 μL RIPA (Biotime, Shanghai, China) with protease inhibitor and phenylmethanesulfonyl fluoride (PMSF, Biotime, Shanghai, China) on ice. After 20 min 13,000g centrifugation at 4 °C, supernatant protein concentrations were determined by bicinchoninic acid (Thermo scientific, Rockford, USA) protein assay. A 10 % SDS-PAGE gel was used to separate proteins, and the proteins were transferred to PVDF membrane. Blots were probed for 12 h at 4 °C with primary antibodies diluted in Tris-buffered saline (TBS) containing 5 % nonfat milk (1:2000 dilution for CD147 antibody (final concentration 1 μg/mL), 1:50 dilution for β-tubulin antibody (final concentration 1 μg/mL), 1:1000 dilution for RelA, NF-κBI (p50), IκB, and IKK antibodies (Proteintech, Wuhan, China). Then, secondary antibodies (goat anti-mouse antibody, goat anti-rabbit antibody, 1:3000, Thermo scientific, Rockford, USA) were incubated with the PVDF membrane for 1 h at room temperature. The visualization was realized with an enhanced chemiluminescence (ECL)/Western blotting detection system.
Statistical analysis
The experiments were performed independently at least three times. The data were expressed as the mean value ± SD. Unpaired and paired t tests or one-way analysis of variance followed by Dunnett’s posttest (for subgroup analyses) was used. All of the analyses were made using the SPSS 19 statistical software package, and the statistical images were processed by GraphPad Prism 5.0 software. Bonferroni’s correction was applied for the number of hypotheses made (n), and differences were considered to be statistically significant when p < 0.05/n.
Results
Cells in RA synovium are resistant to TNF-induced apoptosis
To study apoptosis in RA patients’ synovium, we first monitored TNF expression in RA synovium tissue. HE staining displayed pathologic changes in RA and OA synovial tissues (Fig. 1a). For immunohistochemistry, irrelevant mouse anti-human IgG and rabbit serum were used as negative controls (Fig. 1b). Sections from different patients showed that TNF was overexpressed in RA synovium but remained relatively low level in OA (Fig. 1c, d). Staining for cleaved-caspase 3 (CC3) revealed no obvious differences between RA and OA synovium tissues (Fig. 1c, d), indicating that overexpressed TNF in RA synovium did not cause extra apoptosis.
Fig. 1.
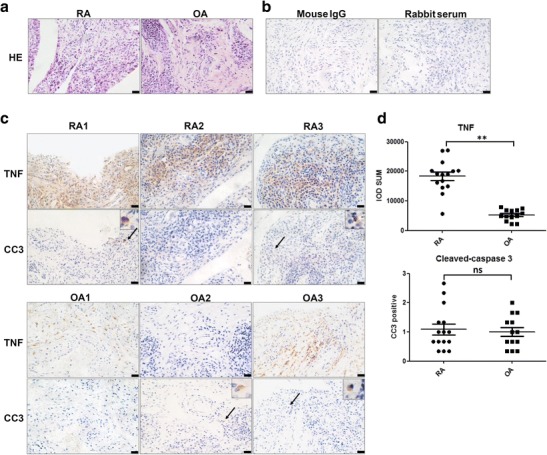
TNF and cleaved-caspase 3 expression in synovial tissues of patients with rheumatoid arthritis (RA) and osteoarthritis (OA). a HE staining of RA and OA synovial sections. b Mouse IgG and rabbit serum were used as isotype controls. c Representative sections of RA1, RA2, RA3, OA1, OA2, and OA3 synovial tissue specimens stained for TNF and cleaved-caspase 3. Positive cleaved-caspase 3 staining (black arrow) indicated apoptotic cells. d Statistical results of TNF and cleaved-caspase 3 stains. Scale bar = 20 μm. **p < 0.01/2. ns not significant
TNF induces mild RASF apoptosis
In vitro, we examined TNF-induced synovioblast apoptosis. After treating with TNF (10 ng/mL) for 24 h, apoptotic RASFs (annexin V positive) increased to 1.5 times their normal level (Fig. 2a, b). Then, we assessed apoptosis in RASFs treated with TNF over a time course. The rates of apoptotic RASFs fluctuated at the beginning and reached their maximum value at 24 h (Fig. 2c). As the TNF concentration increased (from 0, 5, 10, 20, 40, to 80 ng/mL), the apoptotic RASFs accordingly increased (Fig. 2d). Real-time PCR results demonstrated that CD147 mRNA expression changed after TNF treatment and were highest at 24 h (Fig. 2e, f).
Fig. 2.
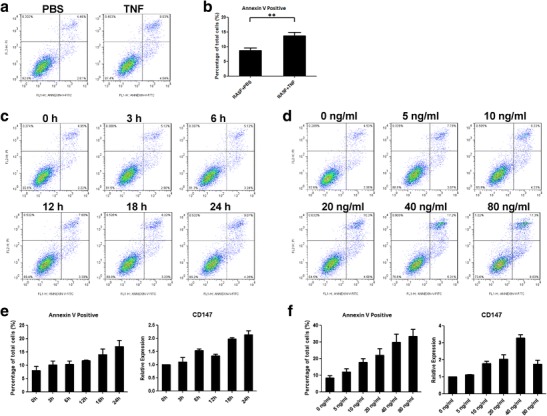
TNF could induce apoptosis in synovioblasts. a After treating synovioblasts with 10 ng/mL TNF for 24 h, the annexin V-positive apoptotic rate increased slightly. b Statistical results of TNF-induced apoptosis. c Representative results indicating apoptosis under TNF stimulation at different time points (0, 3, 6, 12, 18, and 24 h). d Representative results indicating RASF apoptosis with different concentration of TNF stimulation (0, 5, 10, 20, 40, and 80 ng/mL). e Statistical results of annexin V positivity rates and CD147 expression after treating with TNF for different times. f Statistical results of annexin V positivity rates and CD147 expression with different concentration of TNF stimulation. **p < 0.01/2
CD147 assists synovioblast apoptosis resistance under TNF stimulation
To further analyze the mechanism of TNF-related synovioblast apoptosis resistance, we detected CD147 expression in RA and OA synovial tissue (Fig. 3a). Immunohistochemical staining revealed upregulated CD147 in RA synovium. We used lentivirus to knockdown CD147 expression in the RASFs. Western blot and flow cytometry results indicated that the lentivirus disabled CD147 protein expression (RASF A6) and real-time PCR showed the downregulation of CD147 mRNA (RASF A6) (Fig. 3a). RASF NC refers to lentivirus transfection without inhibition of CD147 expression, and RASF CTL represents RASF without lentivirus transfection.
Fig. 3.
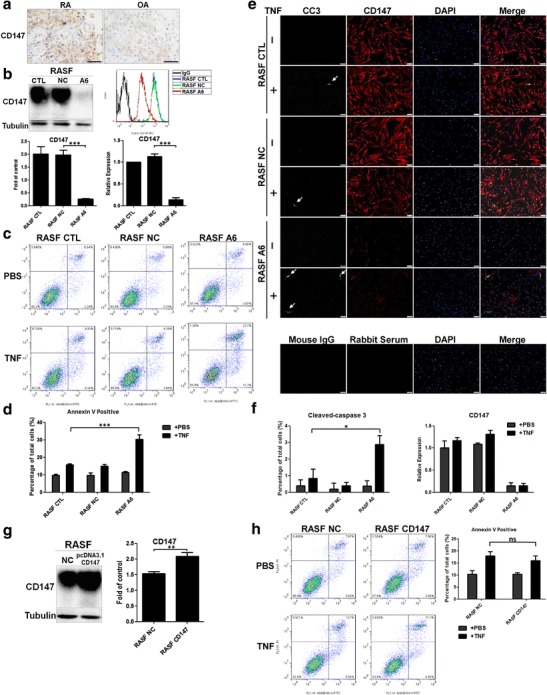
CD147 downregulation facilitated TNF-induced synovioblast apoptosis. a CD147 was overexpressed on RA synoviocytes (RA, n = 4; OA, n = 4), scale bar = 20 μm. b Lentivirus transfection successfully knocked down CD147 mRNA and protein expression (approximately 10 % of original expression) as proven by real-time PCR, western blot, and flow cytometry. c, d Representative flow cytometry results and statistical analysis of TNF-induced (10 ng/mL, 24 h) apoptosis in RASF CTL, RASF NC, and RASF A6 cells. Annexin V-positive dots indicating that CD147 assisted synovioblast apoptosis resistance under TNF stimulation. e, f Representative immunofluorescence pictures and statistical analysis of TNF-induced (10 ng/mL, 24 h) apoptosis (CC3 positive, white arrow) and CD147 expression, scale bar = 100 μm. g After transfection of full-length CD147, the expression of CD147 in RASFs was measured by western blot. h Representative flow cytometry results and statistical analysis of TNF-induced (10 ng/mL, 24 h) apoptosis in RASF NC and RASF CD147. *p < 0.05/n; **p < 0.01/n; ***p < 0.001/n. ns not significant
We next treated RASF CTL, NC, and A6 cells with TNF at 10 ng/mL for 24 h. CD147 downregulation (RASF A6) significantly enhanced TNF-induced apoptosis, whereas for RASF CTL and RASF NC cells, apoptosis remained at a relatively low rate (Fig. 3c, d). An immunofluorescence assay indicated that CD147 fluorescence was markedly lower in RASF A6 and CC3 stain illustrated that CD147 downregulation significantly enhanced TNF-induced apoptosis (Fig. 3e, f).
What’s more, we transfected full-length CD147 pcDNA 3.1 (RASF CD147) and negative control (RASF NC) plasmids to primary RASFs, and western blot results indicated the successful CD147 overexpression (Fig. 3g). After transfection, TNF-induced apoptosis was not significantly different between RASF NC and RASF CD147 (Fig. 3h).
Downregulated CD147 inhibits NF-κB pathway-dependent proinflammatory cytokine secretion
After testing CD147’s contribution to TNF-induced apoptosis resistance, we further examined the relationship between cytokine expression and CD147 expression in TNF-treated and N-p-Tosyl-l-phenylalanine chloromethyl ketone (TPCK, a NF-κB inhibitor, final concentration 50 μΜ, Sigma, Buchs, Switzerland)-treated synovioblasts. Real-time PCR results demonstrated that CD147 expression decreased after lentivirus transfection, and CD147 downregulation (A6 group) inhibited IL-6, IL-8, and IL-1β mRNA expression after TNF treatment while revealed no obvious difference when treating with both TNF and the NF-κB inhibitor TPCK (Fig. 4a). Western blot assay demonstrated that downregulated CD147 decreased IκB kinase (IKK) protein level. Meanwhile, using RelA antibody, we found three main bands around 65 kD (RelA), 115 kD (RelA-NFκB I complex), and 160 kD (IκB-RelA-NFκB I complex). In RASF A6, 160 kD (IκB-RelA-NFκB I complex) band was much stronger than RASF CTL and RASF NC, which indicated that CD147 probably played a role in collapse of IκB-RelA-NFκB I complex (Fig. 4b, c). In addition, CD147 downregulation (RASF A6) significantly enhanced TNF-induced apoptosis; however, downregulated CD147 made no difference in apoptotic rate when using both TNF and the NF-κB inhibitor TPCK (Fig. 4d, e). IKK mRNA expression was decreased in A6 group, which illustrated that CD147 positively regulated IKK transcription (Fig. 4f).
Fig. 4.
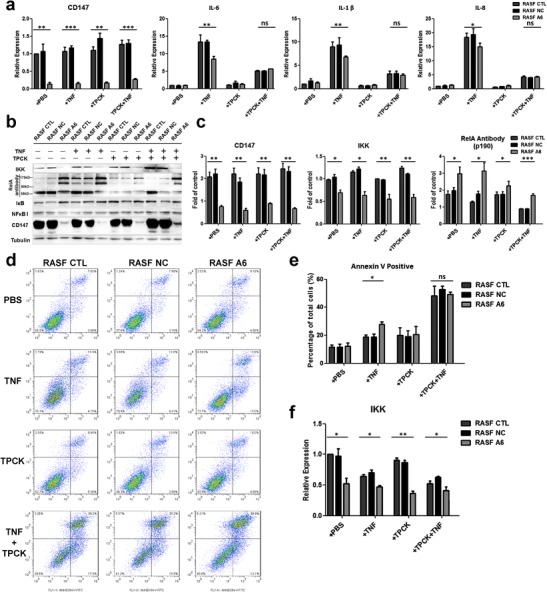
CD147 knockdown inhibited NF-κB pathway activation under TNF stimulation. a Relative CD147, IL-6, IL-1β, and IL-8 mRNA expression was measured by real-time PCR under TNF and/or TPCK stimulation. b, c Representative results and statistical analysis of IKK, NF-κB (p65, p50, and relative complex), IκB, and CD147 protein expression by western blot assay under TNF and/or TPCK stimulation. d, e Representative results and statistical analysis of apoptosis induced by TNF and/or TPCK. f Relative IKK mRNA expression was measured by real-time PCR under TNF and/or TPCK stimulation. *p < 0.05/n; **p < 0.01/n; ***p < 0.001/n. ns not significant
The extracellular portion of CD147 is responsible for synovioblast apoptosis resistance
After transfecting the extracellular or intracellular portion (EP or IP) of CD147 into the RASF A6 (CD147 knockdown) cells (Fig. 5a), real-time PCR results indicated that compared to the RASF A6 group, RASF A6EP cells only expressed the CD147 extracellular portion mRNA, whereas the RASF A6IP group only expressed the CD147 intracellular portion mRNA. Western bolt and flow cytometry results indicated that the extracellular portion of CD147 successfully expressed on cell membrane (Fig. 5b); however, owing to the lack of proper antibody, we did not get the proof of intracellular portion of CD147 protein in RASF A6IP. TNF-induced apoptosis was reduced in the RASF A6EP group but not in RASF A6IP group (Fig. 5c, d), which suggested that the extracellular portion of CD147 was responsible for the TNF-induced apoptosis resistance of RASFs.
Fig. 5.
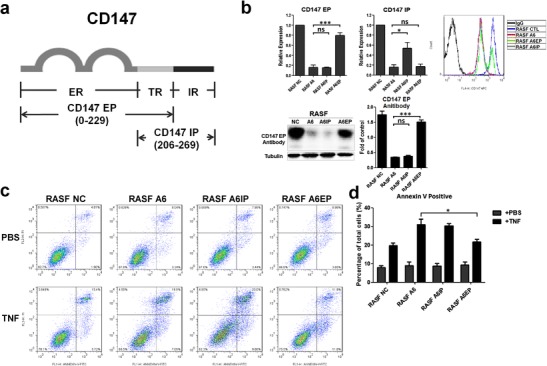
The extracellular portion of CD147 is responsible for synovioblast resistance to TNF-induced apoptosis. a Diagram of exogenously expressed CD147. ER extracellular region (0–206), TR transmembrane region (206–229), IR intracellular region (229–269). CD147 EP denotes the extracellular portion of CD147 (including transmembrane region); CD147 IP denotes the intracellular portion of CD147 (including transmembrane region). Genes for the exogenous proteins were carried by pcDNA 3.1 vectors. b Successful expression of CD147 EP and CD147 IP mRNAs in the RASF A6 cell line was verified by real-time PCR. RASF A6EP denotes the extracellular portion of CD147 expressed in RASF A6; RASF A6IP denotes the intracellular portion of CD147 expressed in RASF A6. Western blot and flow cytometry results for CD147 EP protein expression. c, d Representative results and statistical analysis flow cytometry assay illustrated that only the extracellular portion of CD147 could reverse the resistance to TNF-stimulated apoptosis of RASF A6 cells. *p < 0.05/n; **p < 0.01/n; ***p < 0.001/n. ns not significant
Decreased level of proinflammatory cytokines in CD147 antibody-treated RASFs
The CD147 monoclonal antibody 5A12 (prepared in our laboratory) was previously shown to inhibit proinflammatory cytokine secretion in T lymphocytes. We added 5A12 mAb (20 μg/mL) to TNF-treated RASFs to assess the role of the antibody in both apoptosis and cytokine secretion. Apoptotic RASFs increased after the TNF treatment, but there was no obvious change after the 5A12 mAb was added (Fig. 6a, b). Real-time PCR data indicated a negative role of 5A12 mAb in TNF-induced IL-6, IL-1β, and IL-8 mRNA expression (Fig. 6c), and ELISA results supported the inhibitory effect of 5A12 on IL-6 and IL-1β secretion (Fig. 6d).
Fig. 6.
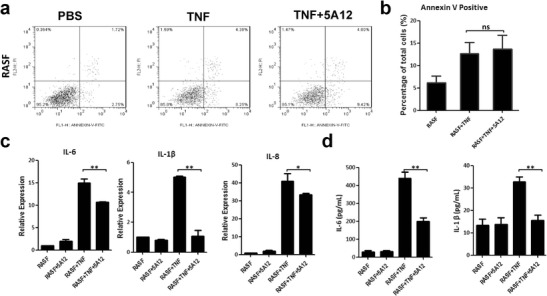
In RASFs, CD147 monoclonal antibody 5A12 inhibits TNF-induced proinflammatory cytokine production but does not influence apoptotic rates. a Representative data of TNF-induced apoptosis, which could not be amplified by 5A12 mAb. b Statistic analysis of TNF-induced apoptosis illustrating that 5A12 mAb did not affect TNF-induced cell apoptosis. c Inhibitory role of 5A12 mAb in TNF-related cytokine production. d ELISA results showed that IL-6 and IL-1β secretion was lower in the TNF+5A12 group than in the TNF group. *p < 0.05/n; **p < 0.01/n. ns not significant
Discussion
TNF overexpression is a significant signature of RA. Detected in serum and synovial fluid, TNF is responsible for the immune activation of lymphocytes (both B and T lymphocytes), fibroblasts, endothelium cells, and macrophages through promoting the secretion of cytokines, such as IL-6 and IL-1β, which in turn cause intractable inflammation [28]. Infliximab, a TNF antibody, has already proven its effectiveness in treating RA patients. However, in other cases, TNF is also known as an inducer of apoptosis and necroptosis. Apoptosis is dominant in physiological cell death, which plays an important role in individual development [29] and in responding to mild stress. In contrast, necroptosis, a programmed type of necrosis, is more common under pathological conditions [30]. Unlike necrosis, apoptotic cells produce apoptotic bodies that can be removed quickly without further damage.
In the synovium of RA patients, high expression of TNF greatly enhances the potential for TNF-related synovioblast apoptosis. The proper degree of RASF apoptosis, for one thing, can alleviate the bone and cartilage erosion by reducing the number of fibroblasts; for another, it can consume more TNF, which would otherwise cause extra immune responses. In our study, we found that TNF expression was not correlated with apoptotic rates between RA and OA synovial tissues, both of which remained at a relatively low level, and TNF only caused mild apoptosis in vitro. The key question is which TNF signaling pattern is preferred in RA synovium, the NF-κB-dependent (or NF-κB-independent) cell survival pathway or the caspase-related apoptosis pathway. The factors contributing to TNF signaling pattern preference include the concentration of TNF, the time course of TNF stimulation [31], and the expression of other important molecules. The high level and long duration of TNF expression in serum and synovium of RA patients have been proven in clinical studies [32–34]; nevertheless, little is known about the vital molecules that can switch TNF signaling between the different patterns.
CD147 is a transmembrane protein originally found in hepatocytes. As a cancer biomarker, CD147 facilitates MMP production and increases the invasiveness of hepatocellular carcinoma [35–37]. Similarly, in RA synoviocytes, CD147 promotes the erosion of the bone and cartilage. Moreover, the overexpression of CD147 in RASFs results in angiogenesis in synovium. Even though there is no indication of CD147 in TNF-stimulated RASFs, it has been confirmed that CD147 inhibits HCC cell apoptosis under ER stress. On the other hand, TNF is significant in ER stress, suggesting that CD147 may be vital to protecting cells from TNF-stimulated apoptosis.
Our results indicate for the first time that CD147 is one of the molecules causing the NF-κB-dependent cell survival pathway to be preferred over the apoptosis pathway under TNF activation, which finally causes the apoptosis resistance of RASF. Though previous study had testified the NF-κB activating role of recombinant human CD147 (as an exogenous stimulation) in cardiomyocytes [38], we for the first time revealed that CD147 downregulation in RASF could inhibit the IKK transcription and protein expression, which demonstrated that CD147 could be a direct transcription factor of IKK or a regulator of other IKK transcription factors. The possible mechanism of such inflammatory activation and apoptosis resistance is CD147 that increases IKK transcription and protein expression, so that IκB-RelA-NFκB I complex is easier to be collapsed under TNF stimulation. As a result, in RA synovium, where there is a high level of TNF, the NF-κB pathway of RASF is activated; meanwhile, the apoptosis pathway is blocked.
As for the region of CD147 that functions in RASFs TNF-related inflammatory activation and apoptosis resistance, we find that the extracellular portion plays a major role. Blocking the function of CD147’s extracellular portion by using the CD147 monoclonal antibody 5A12 failed to alleviate TNF-related apoptosis resistance, indicating that it is not the antigen-antibody binding site that contributes to the resistance. However, the direct molecule interacting with CD147 extracellular portion and mechanism of IKK transcriptional activation is still unknown. In our research, the intracellular portion of CD147 is not related to synovioblasts apoptosis resistance. Although CD147 intracellular portion is detected in mRNA level, we could not exclude the possibility of unsuccessful protein expression. CD147 IP is a 7-kD exogenous protein, which can be easily degraded and difficult to be detected. Therefore, the concrete mechanism of intracellular portion of CD147 still needs further exploration.
In summary, for the first time, CD147 has been proven to be responsible for the apoptosis resistance of RASFs under TNF stimulation. Our study clarifies one of the mechanisms of TNF-related RA pathogenesis, and it sheds light on a potential RA treatment targeting CD147-related inflammatory activation and apoptosis resistance.
Acknowledgments
We thank Ling Li for generously giving CD147 shRNA-related lentiviral plasmids. This research was supported by the National Basic Research Program of China (2015CB553704) and the National Science and Technology Major Projects of New Drugs (2012ZX09103301-026).
Conflict of interest
The authors declare no conflict of interest.
Abbreviations
- RA
Rheumatoid arthritis
- OA
Osteoarthritis
- TNF
Tumor necrosis factor
- IκB
NF-kappa-B inhibitor
- IKK
IκB kinase
- CTL
Blank control
- NC
Negative control
- A6
With CD147 shRNA lentivirus transfection
- PI
Propidium iodide
- EP
Extracellular portion
- IP
Intracellular portion
Contributor Information
Ping Zhu, Phone: 86-29-84773951, Email: zhuping@fmmu.edu.cn.
Zhi-nan Chen, Phone: 86-29-84774547, Email: zhinanchen@fmmu.edu.cn.
References
- 1.Epstein FH, Harris ED., Jr Rheumatoid arthritis: pathophysiology and implications for therapy. N Engl J Med. 1990;322:1277–1289. doi: 10.1056/NEJM199005033221805. [DOI] [PubMed] [Google Scholar]
- 2.Pope RM. Apoptosis as a therapeutic tool in rheumatoid arthritis. Nat Rev Immunol. 2002;2:527–535. doi: 10.1038/nri846. [DOI] [PubMed] [Google Scholar]
- 3.Korb-Pap A, Stratis A, Muhlenberg K, Niederreiter B, Hayer S, Echtermeyer F, Stange R, Zwerina J, Pap T, Pavenstadt H, et al. Early structural changes in cartilage and bone are required for the attachment and invasion of inflamed synovial tissue during destructive inflammatory arthritis. Ann Rheum Dis. 2012;71:1004–1011. doi: 10.1136/annrheumdis-2011-200386. [DOI] [PubMed] [Google Scholar]
- 4.Izquierdo E, Cañete JD, Celis R, Del Rey MJ, Usategui A, Marsal S, Sanmartí R, Criado G, Pablos JL. Synovial fibroblast hyperplasia in rheumatoid arthritis: clinicopathologic correlations and partial reversal by anti–tumor necrosis factor therapy. Arthritis Rheumatol. 2011;63:2575–2583. doi: 10.1002/art.30433. [DOI] [PubMed] [Google Scholar]
- 5.Feldmann M. Development of anti-TNF therapy for rheumatoid arthritis. Nat Rev Immunol. 2002;2:364–371. doi: 10.1038/nri802. [DOI] [PubMed] [Google Scholar]
- 6.Wajant H, Pfizenmaier K, Scheurich P. Tumor necrosis factor signaling. Cell Death & Differentiation. 2003;10:45–65. doi: 10.1038/sj.cdd.4401189. [DOI] [PubMed] [Google Scholar]
- 7.Genovese MC, Fleischmann R, Furst D, Janssen N, Carter J, Dasgupta B, Bryson J, Duncan B, Zhu W, Pitzalis C, et al. Efficacy and safety of olokizumab in patients with rheumatoid arthritis with an inadequate response to TNF inhibitor therapy: outcomes of a randomised phase IIb study. Ann Rheum Dis. 2014;73:1607–1615. doi: 10.1136/annrheumdis-2013-204760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Aggarwal BB. Signalling pathways of the TNF superfamily: a double-edged sword. Nat Rev Immunol. 2003;3:745–756. doi: 10.1038/nri1184. [DOI] [PubMed] [Google Scholar]
- 9.Vandenabeele P, Galluzzi L, Vanden Berghe T, Kroemer G. Molecular mechanisms of necroptosis: an ordered cellular explosion. Nat Rev Mol Cell Biol. 2010;11:700–714. doi: 10.1038/nrm2970. [DOI] [PubMed] [Google Scholar]
- 10.Yan J, Xiang J, Lin Y, Ma J, Zhang J, Zhang H, Sun J, Danial NN, Liu J, Lin A. Inactivation of BAD by IKK inhibits TNFalpha-induced apoptosis independently of NF-kappaB activation. Cell. 2013;152:304–315. doi: 10.1016/j.cell.2012.12.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dragovich T, Rudin CM, Thompson CB. Signal transduction pathways that regulate cell survival and cell death. Oncogene. 1998;17:3207–3213. doi: 10.1038/sj.onc.1202587. [DOI] [PubMed] [Google Scholar]
- 12.Jiang JL, Tang J. CD147 and its interacting proteins in cellular functions. Sheng Li Xue Bao. 2007;59:517–523. [PubMed] [Google Scholar]
- 13.Chen Z, Mi L, Xu J, Yu J, Wang X, Jiang J, Xing J, Shang P, Qian A, Li Y. Function of HAb18G/CD147 in invasion of host cells by severe acute respiratory syndrome coronavirus. J Infect Dis. 2005;191:755–760. doi: 10.1086/427811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Crosnier C, Bustamante LY, Bartholdson SJ, Bei AK, Theron M, Uchikawa M, Mboup S, Ndir O, Kwiatkowski DP, Duraisingh MT. Basigin is a receptor essential for erythrocyte invasion by Plasmodium falciparum. Nature. 2011;480:534–537. doi: 10.1038/nature10606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dai L, Bai L, Lu Y, Xu Z, Reiss K, Valle LD, Kaleeba J, Toole BP, Parsons C, Qin Z. Emmprin and KSHV: new partners in viral cancer pathogenesis. Cancer Lett. 2013;337:161–166. doi: 10.1016/j.canlet.2013.05.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Seizer P, Klingel K, Sauter M, Westermann D, Ochmann C, Schönberger T, Schleicher R, Stellos K, Schmidt E-M, Borst O. Cyclophilin A affects inflammation, virus elimination and myocardial fibrosis in coxsackievirus B3-induced myocarditis. J Mol Cell Cardiol. 2012;53:6–14. doi: 10.1016/j.yjmcc.2012.03.004. [DOI] [PubMed] [Google Scholar]
- 17.van Ooij C. Parasitology: Basigin opens the door to malaria. Nat Rev Microbiol. 2011;10:3–3. doi: 10.1038/nrmicro2723. [DOI] [PubMed] [Google Scholar]
- 18.Yang Y, Lu N, Zhou J, Z-n C, Zhu P. Cyclophilin A up-regulates MMP-9 expression and adhesion of monocytes/macrophages via CD147 signalling pathway in rheumatoid arthritis. Rheumatology. 2008;47:1299–1310. doi: 10.1093/rheumatology/ken225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yurchenko V, Constant S, Eisenmesser E, Bukrinsky M. Cyclophilin–CD147 interactions: a new target for anti‐inflammatory therapeutics. Clin Exp Immunol. 2010;160:305–317. doi: 10.1111/j.1365-2249.2010.04115.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hu J, Dang N, Yao H, Li Y, Zhang H, Yang X, Xu J, Bian H, Xing J, Zhu P, et al. Involvement of HAb18G/CD147 in T cell activation and immunological synapse formation. J Cell Mol Med. 2010;14:2132–2143. doi: 10.1111/j.1582-4934.2010.01012.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhu P, Lu N, Z-g S, Zhou J, Wu Z-b, Yang Y, Ding J, Chen Z. CD147 overexpression on synoviocytes in rheumatoid arthritis enhances matrix metalloproteinase production and invasiveness of synoviocytes. Arthritis Res Ther. 2006;8:R44. doi: 10.1186/ar1899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang C, Yao H, Ln C, Jf J, Wang L, Jy D, Zheng Z, Zn C, Zhu P. CD147 induces angiogenesis through a vascular endothelial growth factor and hypoxia‐inducible transcription factor 1α–mediated pathway in rheumatoid arthritis. Arthritis Rheum. 2012;64:1818–1827. doi: 10.1002/art.34341. [DOI] [PubMed] [Google Scholar]
- 23.Jia J, Wang C, Shi Z, Zhao J, Jia Y, Zhao-Hui Z, Li X, Chen Z, Zhu P. Inhibitory effect of CD147/HAb18 monoclonal antibody on cartilage erosion and synovitis in the SCID mouse model for rheumatoid arthritis. Rheumatology (Oxford) 2009;48:721–726. doi: 10.1093/rheumatology/kep099. [DOI] [PubMed] [Google Scholar]
- 24.Tang J, Guo YS, Zhang Y, Yu XL, Li L, Huang W, Li Y, Chen B, Jiang JL, Chen ZN. CD147 induces UPR to inhibit apoptosis and chemosensitivity by increasing the transcription of Bip in hepatocellular carcinoma. Cell Death Differ. 2012;19:1779–1790. doi: 10.1038/cdd.2012.60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Xue X, Piao J-H, Nakajima A, Sakon-Komazawa S, Kojima Y, Mori K, Yagita H, Okumura K, Harding H, Nakano H. Tumor necrosis factor α (TNFα) induces the unfolded protein response (UPR) in a reactive oxygen species (ROS)-dependent fashion, and the UPR counteracts ROS accumulation by TNFα. Journal of Biological Chemistry. 2005;280:33917–33925. doi: 10.1074/jbc.M505818200. [DOI] [PubMed] [Google Scholar]
- 26.Aletaha D, Neogi T, Silman AJ, Funovits J, Felson DT, Bingham CO, Birnbaum NS, Burmester GR, Bykerk VP, Cohen MD. 2010 rheumatoid arthritis classification criteria: an American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheumatol. 2010;62:2569–2581. doi: 10.1002/art.27584. [DOI] [PubMed] [Google Scholar]
- 27.Guo N, Zhang K, Lv M, Miao J, Chen Z, Zhu P. CD147 and CD98 complex-mediated homotypic aggregation attenuates the CypA-induced chemotactic effect on Jurkat T cells. Mol Immunol. 2015;63:253–263. doi: 10.1016/j.molimm.2014.07.005. [DOI] [PubMed] [Google Scholar]
- 28.Feldmann M, Steinman L. Design of effective immunotherapy for human autoimmunity. Nature. 2005;435:612–619. doi: 10.1038/nature03727. [DOI] [PubMed] [Google Scholar]
- 29.Bobba A, Amadoro G, La Piana G, Calissano P, Atlante A. Glycolytic enzyme upregulation and numbness of mitochondrial activity characterize the early phase of apoptosis in cerebellar granule cells. Apoptosis. 2015;20:10–28. doi: 10.1007/s10495-014-1049-1. [DOI] [PubMed] [Google Scholar]
- 30.Christofferson DE, Yuan J. Necroptosis as an alternative form of programmed cell death. Curr Opin Cell Biol. 2010;22:263–268. doi: 10.1016/j.ceb.2009.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Baud V, Karin M. Signal transduction by tumor necrosis factor and its relatives. Trends Cell Biol. 2001;11:372–377. doi: 10.1016/S0962-8924(01)02064-5. [DOI] [PubMed] [Google Scholar]
- 32.Buckland J. Rheumatoid arthritis: anti-TNF and anti-IL-17 antibodies—better together! Nat Rev Rheumatol. 2014;10:699. doi: 10.1038/nrrheum.2014.183. [DOI] [PubMed] [Google Scholar]
- 33.van Schie KA, Hart MH, de Groot ER, Kruithof S, Aarden LA, Wolbink GJ, Rispens T. The antibody response against human and chimeric anti-TNF therapeutic antibodies primarily targets the TNF binding region. Ann Rheum Dis. 2015;74:311–314. doi: 10.1136/annrheumdis-2014-206237. [DOI] [PubMed] [Google Scholar]
- 34.Feldmann M, Brennan FM, Williams RO, Cope AP, Gibbons DL, Katsikis PD, Maini RN. Evaluation of the role of cytokines in autoimmune disease: the importance of TNF alpha in rheumatoid arthritis. Prog Growth Factor Res. 1992;4:247–255. doi: 10.1016/0955-2235(92)90022-A. [DOI] [PubMed] [Google Scholar]
- 35.Sun J, Hemler ME. Regulation of MMP-1 and MMP-2 production through CD147/extracellular matrix metalloproteinase inducer interactions. Cancer Res. 2001;61:2276–2281. [PubMed] [Google Scholar]
- 36.Wu J, Ru N, Zhang Y, Li Y, Wei D, Ren Z, Huang X, Chen Z, Bian H. HAb18G/CD147 promotes epithelial–mesenchymal transition through TGF-β signaling and is transcriptionally regulated by Slug. Oncogene. 2011;30:4410–4427. doi: 10.1038/onc.2011.149. [DOI] [PubMed] [Google Scholar]
- 37.Zhang D-W, Zhao Y-X, Wei D, Li Y-L, Zhang Y, Wu J, Xu J, Chen C, Tang H, Zhang W. HAb18G/CD147 promotes activation of hepatic stellate cells and is a target for antibody therapy of liver fibrosis. J Hepatol. 2012;57:1283–1291. doi: 10.1016/j.jhep.2012.07.042. [DOI] [PubMed] [Google Scholar]
- 38.Venkatesan B, Valente AJ, Prabhu SD, Shanmugam P, Delafontaine P, Chandrasekar B. EMMPRIN activates multiple transcription factors in cardiomyocytes, and induces interleukin-18 expression via Rac1-dependent PI3K/Akt/IKK/NF-kappaB andMKK7/JNK/AP-1 signaling. J Mol Cell Cardiol. 2010;49:655–663. doi: 10.1016/j.yjmcc.2010.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]