

Abstract
The observations that Lymphopenia is common in severe acute respiratory syndrome (SARS) patients and that peripheral blood mononuclear cell (PBMC) could be infected by SARS-CoV indicate that PBMC could be useful in identifying the gene expression profile in convalescent patients and tracing the host response to SARS-CoV infection. In this study, the altered genes expressions in the PBMC of convalescent SARS patients were investigated with suppression subtractive hybridization (SSH). We found that genes encoded by mitochondrial DNA (mtDNA) were obviously upregulated, while mitochondria were now found to be closely connected with antiviral immunity. The identification of a viral gene, M, in SSH cDNA library shows the long-term existence of SARS-CoV in vivo. In addition, some oxidative stress sensitive genes, heat shock proteins, transcription factors, and cytokines showed remarkable elevation. Thin-section electron microscope shows increased lysosome-like granule and mitochondria in PBMC of patients. These results provide important intracellular clue for tracing host response to SARS-CoV infection and suggest a role of mitochondria in that process.
KEY WORDS: Severe acute respiratory syndrome, peripheral blood mononuclear cell, suppression subtractive hybridization, mitochondria
INTRODUCTION
Severe acute respiratory syndrome (SARS) is caused by a novel coronavirus (1, 2), called SARS-CoV. Since its outbreak in 2003, some important issues about this novel virus have been clarified. Many questions, especially how this virus causes disease and how the host responds to it, however, remain to be answered. Unlike the other known human coronaviruses such as HCoV-229E, HCoV-OC43, HCoV-NL63, and HCoV-HKU1, which are generally associated with relatively mild respiratory tract infection, occasionally with pneumonia (3–5), SARS-CoV can cause fatal infectious pneumonia. Remarkably, SARS-CoV and HCoV-NL63 employ a co-receptor, ACE2, for cellular entry (6, 7). Furthermore, clinical observations show that there is a significant correlation between age and prognosis. Children have a good prognosis (8), while elderly patients with chronic illness fare badly (9). These findings indicate that both the uniqueness of this virus and the host factor are important for this disease.
It was reported that PBMC could be infected by SARS-CoV (10). Lymphopenia is also common in SARS patients (11). These observations indicate that PBMC could be useful in identifying the gene expression profile in convalescent patients and tracing the host response to SARS-CoV infection. In this study, we design to detect the alteration of genes expression in the PBMC from 10 convalescent SARS patients with suppression subtractive hybridization (SSH). We found that genes encoded by mitochondrial DNA (mtDNA) were obviously upregulated, while mitochondria were now found to be closely connected with antiviral immunity (12, 13). The identification of a viral gene, M, in SSH cDNA library shows the long-term existence of SARS-CoV in vivo, which is consistent with the reports by two groups (14, 15). In addition, some oxidative stress sensitive genes, heat shock proteins, transcription factors, and cytokines showed remarkable elevation.
PATIENTS AND METHODS
Patients and Controls
Ten convalescent SARS patients diagnosed by the WHO definition of SARS were admitted to the Second Affiliated Hospital of Sun Yat-sen University in Guangzhou during a major SARS outbreak in February 2003 and selected for analyzing the altered genes expression in peripheral blood mononuclear cells (PBMC). These patients are all female with the age range from 20 to 38 (at mean of 26). Ten healthy females with similar age were used as control.
Isolation of PBMC
Peripheral blood samples from 10 convalescent SARS patients were collected in June 19 2003, 17–19 weeks from the onset of SARS in these patients. PBMC were isolated using Ficoll-PaqueTM PLUS (Amersham Biosciences) according to manufacturer's protocol and suspended in stock medium (10% DMSO plus 90% autologous serum) and frozen in liquid nitrogen for the study. PBMC of 10 healthy controls were conducted in the same manner as mentioned above.
RNA Preparation
Total RNA was separately extracted from PBMC of 10 convalescent SARS patients using TRIzol® Reagent (Invitrogen), and the concentration was determined with the UV absorption at 260 nm. The total RNA samples from 10 convalescent SARS patients were equally combined to reduce the individual difference in following subtractive hybridization. Before it was used in the following assay, the mixed total RNA was treated with DNase I (RNase free). Total RNA samples of 10 controls were treated in the same way. PolyA+ RNA was isolated from total RNA using the Oligotex mRNA Mini Kit (Qiagen) according to manufacturer's protocol.
Suppression Subtractive Hybridization (SSH) and Cloning of cDNA
As the quantitative insufficiency of RNA, we performed first-strand cDNA and double-strand cDNA synthesis with SMARTTM PCR cDNA Synthesis Kit (Clontech). Total RNA and polyA+ RNA were respectively used as starting materials to synthesize cDNA. To eliminate the artificial impact, two separate SSH processes were performed with PCR-Select cDNA Subtraction Kit (Clontech) according to manufacturer's manual. The cDNAs derived from total RNA were subject to the first suppression subtractive hybridization (SSH-I); the cDNAs derived from polyA+ RNA were subject to the second suppression subtractive hybridization (SSH-II). To evaluate the subtraction efficiency, the abundance of GAPDH gene was investigated in subtracted products by PCR.
The subtracted products were purified by QIAquick PCR Purificaton Kit (Qiagen) and linked with pGEM-T Easy Vector (Promega). The recombinants were transformed into E. coli TG1 with Gene Pluser (Bio-Rad). After PCR confirmation, the positive transformants were selected for sequence analysis.
Sequence Analysis
Sequencing of transformants from subtractive cDNA was performed on ABI3730 sequencer (Applied Biosystem). Sequences were jointly analyzed for homology with BLASTN program at NCBI (http://www.ncbi.nlm.nih.gov/BLAST) and BLAT program at UCSC Genome Bioinformatics (http://www.ucsc.edu). Genes were classified by gene annotation given by Gene Ontology (GO) Consortium (http://www.geneontology.org/).
Expression Analysis by Reverse Transcription- Polymerase Chain Reaction (RT-PCR)
Genes of interest were selected for RT-PCR analysis, and the first-strand cDNA derived from total RNA was used for template. With Oligo(dT) as primer, the reverse transcription was performed using SuperScriptTM First-Strand Synthesis System (Invitrogen) according to manufacturer's protocol. For PCR, the amplification parameters were 2 min at 94°C, 30 cycles of 30 s at 94°C, 50 s at 53–65°C (change temperature for different primer pairs), and 1.5 min at 72°C, followed by 5 min at 72°C. GAPDH and β-actin were used as quantification references. The primers were shown in Table I. The amplified products were subject to electrophoresis, the gels were screened with Gel Doc XR documentation system (BIO-RAD), and the bands were analyzed with Quantity One analysis software (BIO-RAD).
Table I.
The Sequences of Primers of Selected Genes for RT-PCR
| Primer sequence (5′-3′) | |||
|---|---|---|---|
| Gene | Sense | Antisense | Product size (bp) |
| 16s rRNA | CATGACACAGCAAGACGAGAAGAC | CAGGGAGGAATTTGAAGTAGATAG | 420 |
| ND1 | GGCTACTACAACCCTTCGCTGACG | TTGTGTTGTGATAAGGGTGGAGAG | 354 |
| COX1 | GCTTCCTAGGGTTTATCGTGTGAG | ATGGATTTTGGCGTAGGTTTGGTC | 398 |
| ZNF331 | TGGGTTACGCAATGATTTCTTAGT | ACCTCGCTCCCTTATCCTCCTTCT | 438 |
| FOS | TGTTCCCAGCATCATCCAG | TGCGTTTTGCTACATCTCC | 543 |
| IFRD1 | CTACTGCTGACCATATGCCCAATC | AAAATCCCGTTCCTCCACTG | 306 |
| DNAJB1 | GTGAGAGGCAGACCAGCAG | CCAGACCATCAAGGGAGAGG | 343 |
| IL1B | TCCTGCGTGTTGAAAGATGATAAG | TCTGCCAGCCCTAGGGATTGAGT | 315 |
| FTH1 | TCACTACTGGAACTGCACAAACTG | CCAAGACCTCAAAGACAACACCTG | 397 |
| PRDX1 | ACGGAGATCATTGCTTTCAGTG | CTTCCCCATGTTTGTCAGTG | 355 |
| BCL11B | AGTGGTGGTCTTTTGGATGAGG | CTTGTGCTTTGGGATGGCTTAGTC | 418 |
| HBB | AGTGCTCGGTGCCTTTAGTG | AATATCCCCCAGTTTAGTAGTTG | 320 |
| SARS-CoV M | GCTTGTTTTCCTCTGGCTCTTG | CTACACGCTGCGACGCTCCTAAT | 412 |
| GAPDH | ACCACAGTCCATGCCATCAC | TCCACCACCCTGTTGCTGTA | 452 |
| ACTIN | TTCCAGCCTTCCTTCCTGGG | TTGCGCTCAGGAGGAGCAAT | 224 |
Thin-Section Electron Microscope
PBMC from convalescent SARS patients and healthy controls were washed twice with 1× PBS and were respectively mixed together and subject to centrifugation. Deposited cells (about 5×106) were fixed with 1 ml 2.5% glutaraldehyde in 0.1 M phosphate buffer overnight at 4°C. After washing with 0.1 M phosphate buffer, the fixed cells were post-fixed with 1% osmium tetroxide in 0.1 M phosphate buffer, dehydrated with gradient alcohol, embedded with spur resin, double-stained with uranyl acetate and lead citrate, and observed under a JEM_100CX II transmission electron microscope. Specimens were observed with 19,000× magnification, and PBMC were divided into three groups according to the number of mitochondria appearing in each intact cross sections of PBMC (<5, PBMC containing less than five mitochondria; 5–10, PBMC containing 5–10 mitochondria; >10, PBMC containing more than 10 mitochondria). Total 150 intact cross sections of PBMC from patients or controls were respectively counted.
RESULTS
Gene Profile in SSH cDNA Libraries
There are about 300 available sequences obtained in each SSH cDNA library. These sequences were subsequently subject to homology alignment against human genome by jointly using BLASTN program and BLAT program, and a total of 411 individual genes/ sequences were identified. The classification of these genes/sequences is shown in Fig. 1.
Fig. 1.
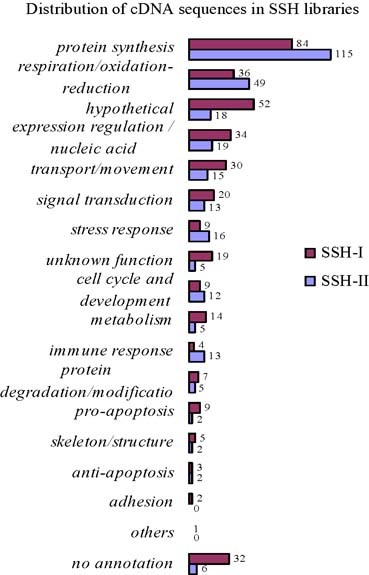
Genes profile in SSH cDNA libraries.
Unexpectedly, we found that the genes encoded by mtDNA occupied a considerable proportion, ∼30%, in both cDNA libraries (Fig. 2). Genes that appeared thrice or more together in two cDNA libraries are shown (Table II). These genes occupy about 40% (264 of 667) of the entire EST sequences, while more than 70% of them are mitochondrial genes. Interestingly, three cDNA molecules representing the mitochondrial control region were identified in a SSH cDNA library (Table II). Mitochondrial control region also called displacement-loop (D-loop) regulatory region is the non-coding region in mtDNA and also the initial site of mtDNA transcription. Both H-strand and L-strand of mtDNA initiate transcription at this region. The primary transcripts are the polycistronic precursor RNA molecules and are subsequently cleaved and processed to produce the individual tRNA and mRNA molecules (16). RNA molecules matching to D-loop region were regarded as transcription/replication primers (17, 18). Furthermore, owing to the annular mtDNA, they may also be the results of strand extension during transcription process. Our finding that two of these three cDNA molecules contain the polyA tail supports this speculation (data not shown). In both cases, however, the repeated appearances of these RNA molecules in SSH cDNA library imply the upregulated transcription of mtDNA. As a whole, the abundant mitochondrial genes appearing in SSH cDNA libraries reflect the unusual activation of mitochondria in PBMC of convalescent SARS patients.
Fig. 2.
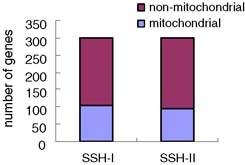
The proportion of mitochondrial genes in two SSH libraries.
Table II.
Genes Appeared Thrice or More in Suppression Subtractive Hybridization (SSH) cDNA Libraries
| Redundancya | |||||
|---|---|---|---|---|---|
| Gene symbol/name | SSH-I | SSH-II | Total | Locationb | Category |
| RNR2 (16s rRNA) | 53 | 27 | 80 | M | Protein synthesis |
| RNR1 (12s rRNA) | 16 | 22 | 38 | M | Protein synthesis |
| ATP6 (ATP synthase F0 subunit 6) | 5 | 13 | 18 | M | Respiration/redox |
| COX2 (cytochrome c oxidase subunit II) | 6 | 7 | 13 | M | Respiration/redox |
| ND1 (NADH dehydrogenase, subunit 1) | 7 | 3 | 10 | M | Respiration/redox |
| COX1 (cytochrome c oxidase subunit I) | 8 | 8 | M | Respiration/redox | |
| ND5 (NADH dehydrogenase, subunit 5) | 4 | 4 | 8 | M | Respiration/redox |
| ATP8 (ATP synthase F0 subunit 8) | 1 | 4 | 5 | M | Respiration/redox |
| ND4 (NADH dehydrogenase, subunit 4) | 2 | 3 | 5 | M | Respiration/redox |
| CYTB (cytochrome b) | 4 | 4 | M | Respiration/redox | |
| mitochondrial control region | 3 | 3 | M | Expression regulation/ nucleic acid modification | |
| HBB (hemoglobin beta) | 10 | 3 | 13 | — | Transport/motility |
| DNAJB1 (DnaJ (Hsp40) homolog, subfamily B, member 1) | 4 | 4 | 8 | — | Stress response |
| RPL10 (ribosomal protein L10) | 1 | 6 | 7 | — | Protein synthesis |
| 28S rRNA (28S ribosomal RNA) | 6 | 6 | — | Protein synthesis | |
| RPS4X (ribosomal protein S4, X-linked X isoform) | 6 | 6 | — | Protein synthesis | |
| RPS12 (ribosomal protein S12) | 5 | 5 | — | Protein synthesis | |
| EEF1A1 (eukaryotic translation elongation factor 1 alpha) | 4 | 4 | — | Protein synthesis | |
| HSPA8 (heat shock 70 kDa protein 8 isoform 2) | 4 | 4 | — | Stress response | |
| ZNF331 (zinc finger protein 331) | 4 | 4 | — | Expression regulation/ nucleic acid modification | |
| BCL11B (B-cell CLL/lymphoma 11B (zinc finger protein)) | 3 | 3 | — | Cell cycle and development | |
| FOS (v-fos FBJ murine osteosarcoma viral oncogene homolog) | 1 | 2 | 3 | — | Expression regulation/ nucleic acid modification |
| FTH1 (ferritin, heavy polypeptide 1) | 1 | 2 | 3 | — | Transport/motility |
| HERPUD1 (homocysteine-inducible, endoplasmic reticulum stress-inducible, ubiquitin-like domain member 1) | 1 | 2 | 3 | — | Stress response |
| RPS15A (ribosomal protein S15a) | 3 | 3 | — | Protein synthesis | |
| SARS-CoV Mc | 1 | 1 | |||
aRedundancy, the number of individual cDNA clones appearing in SSH cDNA libraries.bWhere proteins are located in or genes are from: M, mitochondria; (–) nuclear.cGene appeared in SSH cDNA libraries was encoded by SARS-CoV.
Remarkably, the M gene of SARS-CoV was found in one cDNA library (Table II). Liu et al. (14) reported that the duration of virus excretion in stools of four SARS patients was more than 100 days after onset of symptoms, and suggested that coexisting illness or conditions were associated with longer viral excretion in stools. Chu et al. (15) also reported the long-term duration of RT-PCR positivity in SARS patients after discharge. We collected the PBMC at around 17–19 weeks from the onset of symptoms, and the 10 selected patients were all female. Does this means there is a correlation between long-term persistence and female? Considering the clinical observation of slight predominance of female patients (19), further analysis will be necessary.
Many other genes, such as those related to expression regulation/nucleic acid modification, transport/movement, signal transduction, stress response, cell cycle/development, and so on, were also present in cDNA libraries (Fig. 1). In addition, there were some genes with unknown function and sequences encoding hypothetical proteins.
RT-PCR Analysis of Selected Genes
Some genes of interest were selected for RT-PCR confirmation. The altered expression of these genes is shown (Fig. 3). Expectedly, similar upregulated extent of mitochondrial genes, 16S rRNA, NADH dehydrogenase subunit 1 (ND1), and cytochrome c oxidase subunit I (COX1) has been found, corresponding to the special mitochondrial transcription manner, by which the mitochondrial genes are jointly transcribed. The evident upregulation (almost double) of mitochondrial genes confirms the abundant appearance of that in SSH cDNA libraries.
Fig. 3.
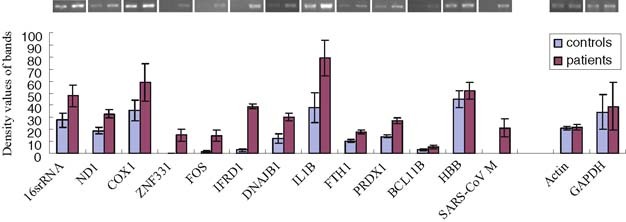
RT-PCR analysis for selected genes. The altered expressions of selected genes in convalescent SARS patients and healthy controls were shown. The existence of SARS-CoV M gene was also shown. GAPDH and β-actin were used as quantificational reference.
In addition to mitochondrial genes, there are other genes that show prominent increase in expression. Two transcription factors, zinc finger protein 331 (ZNF331) and FBJ murine osteosarcoma viral oncogene homolog (FOS), also called c-fos, display distinct upregulation. The stress response protein DnaJ (Hsp40) homolog subfamily B member 1 (DNAJB1), the differentiation-associated gene (IFRD1) and the cytokine (IL-1B), also show intensely increased expression. Furthermore, two oxidative stress sensitive genes, peroxiredoxin 1 (PRDX1) and ferritin heavy polypeptide 1 (FTH1), show obvious elevation. The upregulation of these genes is consistent with the activated immune status in patients. The presence of M gene of SARS-CoV also reveals its substantial persistence in PBMC of convalescent SARS patients (Fig. 3).
Thin-Section Electron Microscope Analysis of PBMC
The PBMC of convalescent SARS patients and controls were subject to thin-section electron microscope analysis. We have respectively observed 150 intact cross sections of PBMC from patients and controls. The statistical result is shown in Fig. 4. It is found that the difference in the number of mitochondria in PBMC between patients and controls was substantial (P<0.01). We have also found that there were prominently increased lysosome-like granules in PBMC from convalescent SARS patients (Fig. 5). The increase of lysosome-like granules could reflect the activated status of these PBMC.
Fig. 4.
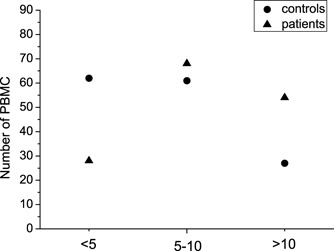
The statistic result of PBMC from convalescent SARS patients and healthy controls. PBMC were divided into three groups based on the number of mitochondria in each cell: <5, PBMC containing less than five mitochondria; 5–10, PBMC containing 5–10 mitochondria; >10, PBMC containing more than 10 mitochondria.
Fig. 5.
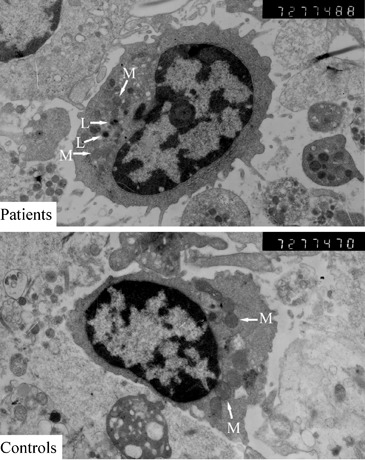
Thin-section electron microscope analysis of PBMC from convalescent SARS patients and healthy controls (original magnification 7200×). M, mitochondria; L, lysosome-like granule.
DISCUSSION
In the present study, we identified that genes coded in mitochondria, a main origin of intracellular free radicals, also an organelle now being found closely connected with antiviral immunity (12, 13), were obviously upregulated in PBMC of convalescent SARS patients. Some of these genes such as PRDX1, FTH1, and FOS responding to oxidative stress also showed remarkable elevation. In addition, the intense upregulations of stress response protein DNAJB1, differentiation-associated gene IFRD1, cytokine IL-1B, and other genes have been observed. In conjunction with the increase of lysosome-like granules and the persistence of viral gene, these results reflect the activated status of the PBMC from convalescent SARS patients.
The active transcription of mitochondrial genes seems to imply that the mitochondria of PBMC have been deeply affected by SARS-CoV infection and/or clinical therapy in some unclear ways. Many factors including virus infection, oxidative stress, nucleoside analogues, and some hormones are able to impact mitochondria and mitochondrial gene transcription. Often, the factors are able not only to influence each other but also to jointly act on mitochondria. It was widely reported that nucleoside analogues, especially nucleoside reverse transcriptase inhibitors (NRTI), could induce mtDNA depletion (20), and subsequently lead to compensatory upregulation of mitochondrial transcription (21). However, none of the patients studied in this trial have been treated with NRTI. Some hormones such as steroid and thyroid can increase mitochondrial gene transcription and respiratory enzyme biosynthesis (22). Four of these patients have ever accepted short-term glucocorticoid treatment (5–11 days). However, the difference of mitochondrial gene expression between them and the other patients is not significant at the time of study (data not shown). These could suggest that there are additional triggers are required for the significant upregulation of mitochondrial transcription.
It was reported that HIV infection was associated with mitochondrial DNA (mtDNA) depletion, extensive mitochondrial respiratory chain (MRC) disturbances and increased oxidative damage (23). This gives us a hint whether SARS-CoV can also affect mitochondria. Interestingly, two proteins of SARS-CoV, 3b and non-structural protein 10 (nsp10), show a close connection with mitochondria. Yuan et al. (24) reported the mitochondrial location of SARS-CoV 3b, and Li et al. (25) found that SARS-CoV nsp10 can specifically interact with the NADH 4L subunit and cytochrome oxidase II. These provide support for our speculation that SARS-CoV could affect the mitochondria in infected cell. Recently, an observation that mitochondrial antiviral signaling (MAVS) (26), also called virus-induced siganling adaptor (VISA) (27), IFN-β promoter stimulator 1 (IPS-1) (28), or CARD adaptor inducing IFN-β (CARDIF) (29), an important protein required for activating type I interferon expression during virus infection requires that it be localized to the mitochondria for its function, establishes a link between mitochondria and innate immunity (12). Furthermore, increasing studies uncover the connections between mitochondria and viruses (30–33). These investigations shed new light on the role of mitochondria in host antiviral immune response, and inspire our further investigation on how SARS-CoV affects infected cell and induces disease.
Despite the inconsistent reports, oxidative stress caused by mitochondria is shown to be involved in the regulation of mitochondrial transcription and proliferation with complicated compensatory mechanism (34–36). In SARS, both pathogen and host factors are considered to be important for the pathogenesis (9). As one kind of important immune factors, free radicals, responsible for oxidative stress, including reactive oxygen species (ROS), and reactive nitrogen species (RNS) produced by mitochondria are essential not only for defense mechanism of cell but also for tissue injury. Activated phagocytes, e.g., neutrophil, monocyte, and macrophage, can rapidly generate a mass of free radicals, so-called respiratory burst, that kill both infected and normal cell at sites of infection. One of primary pathology of SARS is the tissues infiltration of monocyte, neutrophil, and lymphocyte, especially in lung (37, 38), so it is reasonable to speculate that PBMC respond actively to the SARS-CoV infection, and that free radicals with cytotoxic properties released by these immune cells and infected cell participate in the tissues injury in SARS. Furthermore, the extreme manifestation of SARS is acute respiratory distress syndrome (ARDS), while many investigations have showed that oxidant stress contributes to the development of ARDS (39). Some recent evidences also suggest that these reactive species at subtoxic levels can serve as second messengers and redox-signaling molecules (39). This assigns reactive species profound biological roles in infection-induced immune response.
In addition to mitochondrial gene, some other genes responding to oxidative stress display prominent expression increase in the present investigation (Fig. 3). Their increases support the opinion that oxidative stress participates in the pathogenesis of SARS. The obvious augment of PRDX1, a member of antioxidant enzymes, behaving as a dam upon oxidative stress (40), suggests the oxidant/antioxidant imbalance and the demand of antioxidant for eliminating excessive oxidant in PBMC of SARS patients. As a subunit of the ferritin protein, an iron-binding protein, the elevated expression of FTH1 implies the increase of ferritin. Ferritin is an important antioxidant, and it can suppress ROS accumulation through iron sequestration (41). Another iron-binding protein, lactoferrin was also highly upregulated in patients with SARS reported by Reghunathan et al. (42). FOS is a component of redox-sensitive transcription factor complex activator protein 1 (AP-1), and can be activated by various stimuli including ROS (43). Angiotensin II (Ang-II) is a component of renin–angiotensin system, an important factor known for maintaining blood pressure, and was recently considered contributing to severe acute lung injury in SARS (44). Interestingly, Ang-II can elicit the activation of the redox-sensitive AP-1, while the mediator is mitochondria-derived ROS (45). Does this imply mitochondria-derived ROS induced by Ang-II plays a role in host response to SARS-CoV infection? Moreover, it was reported that the nucleocapsid (N) protein of SARS-CoV could selectively activate the AP-1 pathway (46). Is this related with the induction of reactive species? These questions are interesting and require further investigation. Furthermore, the remarkable upregulation of ZNF331 is noticeable. Knowledge about ZNF331 is limited. It was found highly upregulated in PBMC of benzene-exposed workers, while the question of why benzene should so markedly affect ZNF331 expression was still unclear (47). Recently, it was reported that the adverse effects of benzene were dependent on its metabolites and associated with free radical formation (48, 49). This may suggest that ZNF331 is also a probable redox-sensitive transcription factor.
The epidemic of SARS has come to its end. Many questions, particularly how this virus causes disease and how the host responds to it, however, remain unanswered. In this study, we identified the obvious upregulation of mitochondrial genes and genes responding to oxidative stress in PBMC of convalescent SARS patient. These results provide an intracellular clue for tracing host response to SARS-CoV infection and suggest a role of mitochondria in that process. Detailed analysis of these genes could provide more information in understanding this new emerging disease.
ACKNOWLEDGMENTS
We thank the SARS affected staffs of the second affiliated hospital of Sun Yat-Sen University for their cooperation with this study. This work was partially supported by the ‘973’ National Key Program for Developing Basic Research (No. 2003CB514110), by the anti-SARS grant from Guangdong Province, and by the European Commission's Sixth Framework Programme under contract number 511060—Development of Intervention Strategies against SARS in a European-Chinese Taskforce (DISSECT).
Footnotes
Both authors contributed equally to the work, China
REFERENCES
- 1.Peiris JSM, Lai ST, Poon LLM, Guan Y, Yam LYC, Lim W, Nicholls J, Yee WKS, Yan WW, Cheung MT, Cheng VCC, Chan KH, Tsang DNC, Yung RWH, Ng TK. Yuen KY, and members of the SARS study group: Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet. 2003;361:1319–1325. doi: 10.1016/S0140-6736(03)13077-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ksiazek TG, Erdman D, Goldsmith CS, Zaki SR, Peret T, Emery S, Tong S, Urbani C, Corner JA, Lim W, Rollin PE, Dowell SF, Ling AE, Humphrey CD, Shieh WJ, Guarner J, Paddock CD, Rota P, Fields B, DeRisi J, Yang JY, Cox N, Hughes JM, LeDuc JW, Bellini WJ, Anderson LJ. SARS Working Group: A novel coronavirus associated with severe acute respiratory syndrome. N Engl J Med. 2003;348:1953–1966. doi: 10.1056/NEJMoa030781. [DOI] [PubMed] [Google Scholar]
- 3.El-Sahly HM, Atmar RL, Glezen WP, Greenberg SB. Spectrum of clinical illness in hospitalized patients with “Common Cold” virus infections. Clin Infect Dis. 2000;31:96–100. doi: 10.1086/313937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Falsey AR, Walsh EE, Hayden FG. Rhinovirus and coronavirus infection associated hospitalizations among older adults. J Infect Dis. 2002;185:1338–1341. doi: 10.1086/339881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bastien N, Robinson JL, Tse A, Lee BE, Hart L, Li Y. Human coronavirus NL-63 infections in children: A 1-year study. J Clin Microbiol. 2005;43:4567–4573. doi: 10.1128/JCM.43.9.4567-4573.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li WH, Moore MJ, Vasilieva N, Sui JH, Wong SK, Berne MA, Somasundaran M, Sullivan JL, Luzuriaga K, Greenough TC, Choe H, Farzan M. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426:450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hofmann H, Pyrc K, Van Der Hoek L, Geier M, Berkhout B, Pohlmann S. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci USA. 2005;102:7988–7993. doi: 10.1073/pnas.0409465102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chiu WK, Cheung PC, Ng KL, Ip PL, Sugunan VK, Luk DC, Ma LC, Chan BH, Lo KL, Lai WM. Severe acute respiratory syndrome in children: Experience in a regional hospital in Hong Kong. Pediatr Crit Care Med. 2003;4:279–283. doi: 10.1097/01.PCC.0000077079.42302.81. [DOI] [PubMed] [Google Scholar]
- 9.Lo AW, Tang NL, To KF. How the SARS coronavirus causes disease: Host or organism? J Pathol. 2006;208(2):142–151. doi: 10.1002/path.1897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Li L, Wo J, Shao J, Zhu H, Wu N, Li M, Yao H, Hu M, Dennin RH. SARS-coronavirus replicates in mononuclear cells of peripheral blood (PBMC) from SARS patients. J Clin Virol. 2003;28:239–244. doi: 10.1016/S1386-6532(03)00195-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li TS, Qiu ZF, Zhang LQ, Han Y, He W, Liu ZY, Ma XJ, Fan HW, Lu W, Xie J, Wang H, Deng G, Wang A. Significant changes of peripheral T lymphocyte subsets in patients with severe acute respiratory syndrome. J Infect Dis. 2004;189:648–651. doi: 10.1086/381535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.McWhirter SM, Tenoever BR, Maniatis T. Connecting mitochondria and innate immunity. Cell. 2005;122(5):645–647. doi: 10.1016/j.cell.2005.08.026. [DOI] [PubMed] [Google Scholar]
- 13.Seth RB, Sun L, Chen ZJ. Antiviral innate immunity pathways. Cell Res. 2006;16(2):141–147. doi: 10.1038/sj.cr.7310019. [DOI] [PubMed] [Google Scholar]
- 14.Liu W, Tang F, Fontanet A, Zhan L, Zhao QM, Zhang PH, Wu XM, Zuo SQ, Baril L, Vabret A, Xin ZT, Shao YM, Yang H, Cao WC. Long-term SARS coronavirus excretion from patient cohort, China. Emerg Infect Dis. 2004;10(10):1841–1843. doi: 10.3201/eid1010.040297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chu CM, Leung WS, Cheng VC, Chan KH, Lin AW, Chan VL, Lam JY, Chan KS, Yuen KY. Duration of RT-PCR positivity in severe acute respiratory syndrome. Eur Respir J. 2005;25(1):12–14. doi: 10.1183/09031936.04.00057804. [DOI] [PubMed] [Google Scholar]
- 16.Clayton DA. Replication and transcription of vertebrate mitochondrial DNA. Annu Rev Cell Biol. 1991;7:453–478. doi: 10.1146/annurev.cb.07.110191.002321. [DOI] [PubMed] [Google Scholar]
- 17.Ojala D, Crews S, Montoya J, Gelfand R, Attardi G. A small polyadenylated RNA (7 S RNA), containing a putative ribosome attachment site, maps near the origin of human mitochondrial DNA replication. J Mol Biol. 1981;150(2):303–314. doi: 10.1016/0022-2836(81)90454-X. [DOI] [PubMed] [Google Scholar]
- 18.Lee DY, Clayton DA. RNase mitochondrial RNA processing correctly cleaves a novel R loop at the mitochondrial DNA leading-strand origin of replication. Genes Dev. 1997;11(5):582–592. doi: 10.1101/gad.11.5.582. [DOI] [PubMed] [Google Scholar]
- 19.Donnelly CA, Ghani AC, Leung GM, Hedley AJ, Fraser C, Riley S, Abu-Raddad LJ, Ho LM, Thach TQ, Chau P, Chan KP, Lam TH, Tse LY, Tsang T, Liu SH, Kong JH, Lau EM, Ferguson NM, Anderson RM. Epidemiological determinants of spread of causal agent of severe acute respiratory syndrome in Hong Kong. Lancet. 2003;361(9371):1761–1766. doi: 10.1016/S0140-6736(03)13410-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kakuda TN. Pharmacology of nucleoside and nucleotide reverse transcriptase inhibitor-induced mitochondrial toxicity. Clin Ther. 2000;22:685–708. doi: 10.1016/S0149-2918(00)90004-3. [DOI] [PubMed] [Google Scholar]
- 21.Miro O, Lopez S, Rodriguez de la Concepcion M, Martinez E, Pedrol E, Garrabou G, Giralt M, Cardellach F, Gatell JM, Vilarroya F, Casademont J. Upregulatory mechanisms compensate for mitochondrial DNA depletion in asymptomatic individuals receiving stavudine plus didanosine. J Acquir Immune Defic Syndr. 2004;37(5):1550–1555. doi: 10.1097/00126334-200412150-00002. [DOI] [PubMed] [Google Scholar]
- 22.Van Itallie CM. Thyroid hormone and dexamethasone increase the levels of a messenger ribonucleic acid for a mitochondrially encoded subunit but not for a nuclear-encoded subunit of cytochrome c oxidase. Endocrinology. 1990;127(1):55–62. doi: 10.1210/endo-127-1-55. [DOI] [PubMed] [Google Scholar]
- 23.Miro O, Lopez S, Martinez E, Pedrol E, Milinkovic A, Deig E, Garrabou G, Casademont J, Gatell JM, Cardellach F. Mitochondrial effects of HIV infection on the peripheral blood mononuclear cells of HIV-infected patients who were never treated with antiretrovirals. Clin Infect Dis. 2004;39(5):710–716. doi: 10.1086/423176. [DOI] [PubMed] [Google Scholar]
- 24.Yuan X, Shan Y, Yao Z, Li J, Zhao Z, Chen J, Cong Y. Mitochondrial location of severe acute respiratory syndrome coronavirus 3b protein. Mol Cells. 2006;21(2):186–191. [PubMed] [Google Scholar]
- 25.Li Q, Wang L, Dong C, Che Y, Jiang L, Liu L, Zhao H, Liao Y, Sheng Y, Dong S, Ma S. The interaction of the SARS coronavirus non-structural protein 10 with the cellular oxido-reductase system causes an extensive cytopathic effect. J Clin Virol. 2005;34(2):133–139. doi: 10.1016/j.jcv.2004.12.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Seth RB, Sun L, Ea CK, Chen ZJ. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-kappaB and IRF3. Cell. 2005;122(5):669–682. doi: 10.1016/j.cell.2005.08.012. [DOI] [PubMed] [Google Scholar]
- 27.Xu LG, Wang YY, Han KJ, Li LY, Zhai Z, Shu HB. VISA is an adapter protein required for virus-triggered IFN-beta signaling. Mol Cell. 2005;19(6):727–740. doi: 10.1016/j.molcel.2005.08.014. [DOI] [PubMed] [Google Scholar]
- 28.Kawai T, Takahashi K, Sato S, Coban C, Kumar H, Kato H, Ishii KJ, Takeuchi O, Akira S. IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat Immunol. 2005;6(10):981–988. doi: 10.1038/ni1243. [DOI] [PubMed] [Google Scholar]
- 29.Meylan E, Curran J, Hofmann K, Moradpour D, Binder M, Bartenschlager R, Tschopp J. Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus. Nature. 2005;437(7062):1167–1172. doi: 10.1038/nature04193. [DOI] [PubMed] [Google Scholar]
- 30.Yedavalli VS, Shih HM, Chiang YP, Lu CY, Chang LY, Chen MY, Chuang CY, Dayton AI, Jeang KT, Huang LM. Human immunodeficiency virus type 1 Vpr interacts with antiapoptotic mitochondrial protein HAX-1. J Virol. 2005;79(21):13735–13746. doi: 10.1128/JVI.79.21.13735-13746.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Carr SM, Carnero E, Garcia-Sastre A, Brownlee GG, Fodor E. Characterization of a mitochondrial-targeting signal in the PB2 protein of influenza viruses. Virology. 2006;344(2):492–508. doi: 10.1016/j.virol.2005.08.041. [DOI] [PubMed] [Google Scholar]
- 32.Su J, Wang G, Barrett JW, Irvine TS, Gao X, McFadden G. Myxoma virus M11L blocks apoptosis through inhibition of conformational activation of Bax at the mitochondria. J Virol. 2006;80(3):1140–1151. doi: 10.1128/JVI.80.3.1140-1151.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hiraragi H, Kim SJ, Phipps AJ, Silic-Benussi M, Ciminale V, Ratner L, Green PL, Lairmore MD. Human T-lymphotropic virus type 1 mitochondrion-localizing protein p13(II) is required for viral infectivity in vivo. J Virol. 2006;80(7):3469–3476. doi: 10.1128/JVI.80.7.3469-3476.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Miranda S, Foncea R, Guerrero J, Leighton F. Oxidative stress and upregulation of mitochondrial biogenesis genes in mitochondrial DNA-depleted HeLa cells. Biochem Biophys Res Commun. 1999;258(1):44–49. doi: 10.1006/bbrc.1999.0580. [DOI] [PubMed] [Google Scholar]
- 35.Lee HC, Yin PH, Lu CY, Chi CW, Wei YH. Increase of mitochondria and mitochondrial DNA in response to oxidative stress in human cells. Biochem J. 2000;348(Pt 2):425–432. doi: 10.1042/0264-6021:3480425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Suliman HB, Carraway MS, Welty-Wolf KE, Whorton AR, Piantados CA. Lipopolysaccharide stimulates mitochondrial biogenesis via activation of nuclear respiratory factor-1. J Biol Chem. 2003;278:41510–41518. doi: 10.1074/jbc.M304719200. [DOI] [PubMed] [Google Scholar]
- 37.Nicholls JM, Poon LL, Lee KC, Ng WF, Lai ST, Leung CY, Chu CM, Hui PK, Mak KL, Lim W, Yan KW, Chan KH, Tsang NC, Guan Y, Yuen KY, Peiris JS. Lung pathology of fatal severe acute respiratory syndrome. Lancet. 2003;361:1773–1778. doi: 10.1016/S0140-6736(03)13413-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ding Y, Wang H, Shen H, Li Z, Geng J, Han H, Cai J, Li X, Kang W, Weng D, Lu Y, Wu D, He L, Yao K. The clinical pathology of severe acute respiratory syndrome (SARS): A report from China. J Pathol. 2003;200:282–289. doi: 10.1002/path.1440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Quinlan G, Upton R: Oxidant–antioxidant balance in acute respiratory distress syndrome. In: European Respiratory Monograph: ARDS, T Evans, M Griffiths, B. Keogh (eds). European Respiratory Journals Ltd., 2002, pp 33–46
- 40.Rabilloud T, Heller M, Gasnier F, Luche S, Rey C, Aebersold R, Benahmed M, Louisot P, Lunardi J. Proteomics analysis of cellular response to oxidative stress. Evidence for in vivo overoxidation of peroxiredoxins at their active site. J Biol Chem. 2002;277(22):19396–19401. doi: 10.1074/jbc.M106585200. [DOI] [PubMed] [Google Scholar]
- 41.Pham CG, Bubici C, Zazzeroni F, Papa S, Jones J, Alvarez K, Jayawardena S, De Smaele E, Cong R, Beaumont C, Torti FM, Torti SV, Franzoso G. Ferritin heavy chain upregulation by NF-kappaB inhibits TNFalpha-induced apoptosis by suppressing reactive oxygen species. Cell. 2004;119(4):529–542. doi: 10.1016/j.cell.2004.10.017. [DOI] [PubMed] [Google Scholar]
- 42.Reghunathan R, Jayapal M, Hsu LY, Chng HH, Tai D, Leung BP, Melendez AJ. Expression profile of immune response genes in patients with Severe Acute Respiratory Syndrome. BMC Immunol. 2005;6:2. doi: 10.1186/1471-2172-6-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Rao GN, Berk BC. Active oxygen species stimulate vascular smooth muscle cell growth and proto-oncogene expression. Circ Res. 1992;70(3):593–599. doi: 10.1161/01.res.70.3.593. [DOI] [PubMed] [Google Scholar]
- 44.Kuba K, Imai Y, Rao S, Gao H, Guo F, Guan B, Huan Y, Yang P, Zhang Y, Deng W, Bao L, Zhang B, Liu G, Wang Z, Chappell M, Liu Y, Zheng D, Leibbrandt A, Wada T, Slutsky AS, Liu D, Qin C, Jiang C, Penninger JM. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat Med. 2005;11:875–879. doi: 10.1038/nm1267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wu S, Gao J, Ohlemeyer C, Roos D, Niessen H, Kottgen E, Gessner R. Activation of AP-1 through reactive oxygen species by angiotensin II in rat cardiomyocytes. Free Radic Biol Med. 2005;39(12):1601–1610. doi: 10.1016/j.freeradbiomed.2005.08.006. [DOI] [PubMed] [Google Scholar]
- 46.He R, Leeson A, Andonov A, Li Y, Bastien N, Cao J, Osiowy C, Dobie F, Cutts T, Ballantine M, Li X. Activation of AP-1 signal transduction pathway by SARS coronavirus nucleocapsid protein. Biochem Biophys Res Commun. 2003;311:870–876. doi: 10.1016/j.bbrc.2003.10.075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Forrest MS, Lan Q, Hubbard AE, Zhang L, Vermeulen R, Zhao X, Li G, Wu YY, Shen M, Yin S, Chanock SJ, Rothman N, Smith MT. Discovery of novel biomarkers by microarray analysis of peripheral blood mononuclear cell gene expression in benzene-exposed workers. Environ Health Perspect. 2005;113(6):801–807. doi: 10.1289/ehp.7635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sheets P, Carlson G. Kinetic factors involved in the metabolism of benzene in mouse lung and liver. J Toxicol Environ Health A. 2004;67(5):421–430. doi: 10.1080/15287390490273488. [DOI] [PubMed] [Google Scholar]
- 49.Rana SV, Verma Y. Biochemical toxicity of benzene. J Environ Biol. 2005;26(2):157–168. [PubMed] [Google Scholar]