

Abstract
Porcine epidemic diarrhea virus (PEDV) causes significant loss to the swine industry. The emergence of novel PEDV strains in recent years has decreased the effectiveness of PEDV vaccines. We have developed a live recombinant vaccine, a swinepox virus vector that expresses a truncated S protein (rSPV-St) from a recent PEDV strain, SQ2014, and evaluated its immunogenicity and effectiveness in a swine model. Vaccination of swine with rSPV-St elicited a robust antibody response specific for the homologous PEDV SQ2014. Serum IgA titers in rSPV-St-vaccinated animals were significantly higher than in those immunized with inactivated vaccines. The effectiveness of antibodies induced by the rSPV-St vaccine in protection against PEDV was tested in a passive-transfer model in which piglets were challenged with the homologous virus SQ2014 and the heterologous strain CV777. When challenged with the homologous virus, sera from rSPV-St vaccination provided complete protection. However, sera from rSPV-St vaccination did not provide any protection against the heterologous virus challenge. Amino acid sequence differences in the S proteins of the two viruses were identified within neutralizing epitopes, which might have contributed to the divergent clinical results. Our data suggest that rSPV-St is potentially an effective vaccine against infection with emerging PEDV strains.
Introduction
Porcine epidemic diarrhea virus (PEDV) causes vomiting, diarrhea, and dehydration, and the mortality rate in newborn pigs can be up to 100% [1, 2]. The virus is highly contagious and affects pigs of all ages and breeds. PEDV has been reported in many countries since it was first identified in Britain and Belgium [3]. In China, PEDV is more prevalent than porcine rotavirus and transmissible gastroenteritis virus, which also cause viral diarrhea in pigs [4]. The most common practice for preventing PEDV infection is transfer of passive immunity from sows to nursing piglets [5]. Pregnant sows are vaccinated with inactivated or attenuated vaccines and antibodies are passively transferred in the colostrum [5]. This transfer of PEDV immunity is affected by the length of time when the piglets are suckling and the antibody titer in the sow colostrum [5]. Inactivated vaccines are relatively expensive and are inefficient at inducing a mucosal immune response. Given the prevalence of PEDV infection, novel preventive measures are urgently needed [6–9].
PEDV is a member of the genus Alphacoronavirus, family Coronaviridae, order Nidovirales [10]. It is a pleomorphic enveloped virus containing a positive-stranded RNA genome, which is approximately 28 kb in length and has a 5′ cap and a 3′ polyadenylated tail. The virus contains four structural proteins: spike (S), integral membrane (M), envelope (E), and nucleocapsid (N) [11]. The S protein, a type I membrane glycoprotein, is required for virus entry into host cells and binding to the viral receptor porcine amino peptidase. There are two domains in the S protein, S1 (amino acids [aa] 1-735) and S2 (from 736 to the last aa). Previous work has shown that neutralizing epitopes are mainly located in S1 [11–14]. The antigenic variation in the S protein, especially in the S1 domain, makes prevention of PEDV infection more difficult [15–17]. However, based on the surface expression and the function of the S protein, S1 is an attractive target for genetically engineering a PEDV vaccine [18].
Poxvirus has been used as a platform for vaccine development in humans and animals [19–21]. Swinepox virus (SPV) is a naturally mild virus that can potentially function as a vaccination vehicle for the following reasons: a) SPV would not be interfered with by maternal antibodies and b) SPV is host-specific and not transmissible to other species [22]. SPV can express foreign genes effectively and can be modified to include large amounts of recombinant DNA. Moreover, it has low delivery costs and can be a useful platform for live vaccines [23]. Those advantages of SPV suggest that it could be a viable platform for developing a safe live vaccine for PEDV [24, 25].
The aim of this study was to construct a recombinant SPV vaccine expressing a truncated S protein of PEDV, with a focus on its protective efficacy in pigs.
Materials and methods
Cells and viruses
Vero cells (ATCC:E6) and porcine kidney cells (PK-15, ATCC:CCL-33) were purchased from the American Type Culture Collection. Wild-type (wt) SPV NT1501, which has a titer of 1 × 108 plaque-forming units (PFU)/mL as determined in PK-15 cells, was isolated in our laboratory. Two strains of PEDV, the classical CV777 strain, and the novel SQ2014 strain (GenBank: KP728470.1), were also isolated and characterized in our laboratory; the titers of both isolates were determined by plaque assay in Vero cells.
Construction of SPV expressing a truncated S protein (St)
A portion of the S gene of PEDV SQ2014 encoding aa 386-815 of the N-terminus of the S protein, was inserted into the pUSG11 vector [26]. Briefly, the PEDV St gene (1257 bp, KP728470) was amplified from the virus genome using the following two primers: St1 (5’-GCGTCGACATGGTTTTACCTCCTACCGTCA-3’) and St2 (5’-CGGGATCCTTACGCACAATCAACACTAACA-3’). The resulting PCR product was inserted into the pUSG11 plasmid to construct pUSG11-St (Fig. 1a).
Fig. 1.

Characterization of rSPV-St. (A) A schematic diagram for vector construction. (B) Schematic representation of the four antigenic sites in the S protein of PEDV. Shown are antigenic sites 1, 2, 3, 4 (from left to right) of the PEDV S protein. (C) Fluorescence observed for a) rSPV-St and b) wtSPV. (D) Expression of St protein in PK-15 cells infected with a) rSPV-St or b) wtSPV as indicated by IFA. (E) Representative image of a western blot showing St protein expression in cells infected with rSPV-St but not wtSPV. β-actin was used as an internal control. (F) Growth kinetics of rSPV-St and wtSPV in PK-15 cells
The method used to generate the recombinant SPV expressing St (rSPV-St) and green fluorescent protein (GFP) has been described previously [26, 27]. Briefly, a sub-confluent culture of PK-15 cells was infected with wtSPV at a multiplicity of infection (MOI) of 0.02 for 2 h before the cells were transfected with 10 µg of the pUSG11-St plasmid using the Exfect™ Transfection Reagent (Vazyme Biotech Co., Ltd). The cells were harvested at 72 h and lysed by five rounds of freezing and thawing. The lysate was then used to infect PK-15 cells in a 12-well plate for plaque purification of the recombinant virus by fluorescence microscopy (Zeiss, Germany). Briefly, 1.5 mL of medium containing 1% LMP agarose (DingGuo, Beijing, China) was added to each well of the 12-well plate, and the infected cells were incubated for 5 days until plaques became visible under a fluorescence microscope. The plaques were picked and resuspended in 0.3 mL of medium containing 2% FBS. Plaque purification was repeated for 5-6 rounds until all of the plaques in a given well showed green fluorescence. The insertion of the St gene and expression of the St protein were confirm by PCR and indirect immunofluorescence assay (IFA), respectively.
PCR analysis of rSPV-St
DNA was isolated from PK-15 cells infected with rSPV-St and extracted using the SDS-protease K-phenol method. Genomic DNA from PK-15 cells infected with wtSPV was used as a negative control. Amplifications were performed with DNA polymerase (Vazyme Biotech, Jiangsu, China) using the following primers: St1 (5’-GCGTCGACATGGTTTTACCTCCTACCGTCA-3’) and St2 (5’-CGGGATCCTTACGCACAATCAACACTAACA-3’). The expected PCR product was approximately 1257 bp long.
Growth characteristics of rSPV-St in vitro
To analyze the growth characteristics of rSPV-St virus in vitro, PK-15 cells growing in 24-well plates were infected with wtSPV and rSPV-St at an MOI of 0.1. Supernatants of the infected cells were harvested at 6, 12, 24, 36, 48, 60, 72 and 84 h postinfection. Viral titers in the supernatant samples were determined by plaque assay.
Immunofluorescene assay (IFA)
IFA was performed as described previously [28]. PK-15 cells were grown in 24-well plates and infected with the wtSPV and rSPV-St at 1 × 108 plaque-forming units (PFU) in 0.5 mL per well. PBS-treated cells were used as a negative control. At 72 h postinfection, the cells were washed three times with PBS + 0.05% Tween-20 (PBST) and fixed with cold methanol for 10 min at -20 °C. Cells were then washed three times with PBST and blocked with PBST + 10% bovine serum albumin (BSA), followed by incubation for 1 h at 37 °C with antiserum raised against PEDV strain SQ2014 (1:1000 in PBST + 1% BSA). After three washes with PBST, the cells were incubated with rhodamine-conjugated protein A (Boshide, Wuhan, China) for 30 min at 37 °C. After a final wash with PBS, the cells were examined wbyith fluorescence microscopy (Zeiss, Germany).
Western blot analysis
PK-15 cell monolayers were infected with wtSPV and rSPV-St at an MOI of 5 and incubated for 72 h at 37 °C. A lysate of approximately 105 cells was run through a 12% SDS-polyacrylamide gel and then transferred onto a PVDF membrane. The membrane was blocked with 5% skimmed milk in PBST overnight at 4 °C incubated with swine convalescent serum containing antibodies against PEDV strain SQ2014 (1:1000 dilution) for 2 h at 37 °C, and washed three times with PBST. The membrane was washed and then incubated with horseradish peroxidase (HRP)-conjugated anti-swine antibody. After washing with PBST, the protein bands were developed in ECL Western blotting substrate (Bio-Rad, USA) and imaged using an Alpha Innotech Fluor Chem FC2 imager (R&D Systems, Minneapolis, MN).
Animal experiments
All experimental protocols involving pigs were approved by the Laboratory Animal Monitoring Committee of Jiangsu Province, and all experimental animals were negative for SPV and PEDV antibodies by ELISA. Fifteen one-month-old pigs (Large White) were randomly divided into five groups (3 per group) and were immunized twice on days 0 and 21 with (1) rSPV-St (1 × 108 PFU in 2 ml of PBS, protein concentration 4.9 mg/mL), (2) inactivated PEDV strain SQ2014 (1 × 106 PFU in 2 mL of PBS, protein concentration 4.4 mg/mL), (3) inactivated PEDV strain CV777 (1 × 106 PFU in 2 mL of PBS, protein concentration 4.7 mg/mL), (4) wtSPV (1 × 108 PFU in 2 mL of PBS, protein concentration 4.6 mg/mL), (5) PBS alone (negative control). The protein concentration was determined using a NanoDrop 2000 spectrophotometer. The live vaccine and the negative control were administered subcutaneously and orally (1:1 volume ratio), while the inactivated vaccines were mixed with the Freund’s incomplete adjuvant and administered intramuscularly. Sera were collected from the animals on days 0, 7, 14, 21, 28, 35, and 42 post-primary immunization for antibody titration or passive transfer study. To test the protection efficacy of the rSPV-St vaccine, sera collected from each group at day 42 post-immunization were pooled and used to feed one-day-old piglets. Briefly, 50 one-day-old piglets, verified as PEDV negative, were randomly divided into five groups (10 per group) and were fed with sera from each of the vaccination and control groups. The sera were added to milk at a ratio of 5 ml of serum per liter of milk. Each piglet was fed with 50 ml of the serum-milk mixture five times per day [29, 30]. On the second day of serum feeding, five piglets randomly selected from each group were infected with PEDV SQ2014 (1 × 106 PFU in 1 mL of PBS, orally via a gastric tube), while the remaining five piglets in each group were infected with PEDV CV777 (1 × 106 PFU in 1 mL of PBS, orally via a gastric tube). Animals were monitored for 10 days for clinical signs. Small-intestine tissue samples were collected from animals that died from virus challenge for histology. On day 10 post-challenge, the surviving animals were euthanized by intramuscular injection of a combination of ketamine (20 mg/kg) and xylazine (2 mg/kg), followed by intravenous injection of a pentobarbital sodium overdose (60 mg/kg), and the intestinal tissues were sampled. The collected small intestine samples were processed for histological analysis.
Fecal samples were collected from pigs daily. The feces were used homogenized in PBS (10% w/v), frozen and thawed three times, vortexed, and centrifuged at 6000 rpm for 15 min at 4 °C. The supernatant was removed and centrifuged again at 12,000 rpm for 15 min at 4 °C. The final supernatant samples were used for virus titration. PEDV infection was confirmed by a PEDV-M-gene-based reverse transcription polymerase chain reaction (RT-PCR) [31, 32].
Antibody titer measurement
Antibody titers in sera from immunized pigs were measured by ELISA. Briefly, purified PEDV SQ2014 was added to the plates and incubated overnight at 4 °C, and the plates were washed three times with PBST and blocked with 5% skim milk in PBST at 37 °C for 2 h. Serum samples were serially diluted and incubated at 37 °C for 1 h. After the incubation, the plates were washed three times with PBST, and HRP-conjugated goat anti-SPA IgG or goat anti-porcine IgA diluted in PBST was then added to each well. The plates were incubated at room temperature in the dark for 30 min and then washed three times with PBST. The TMB microwell peroxidase substrate system (TIANGEN) was added for color development. After incubation for 20 min, the reaction was stopped with 2 M sulfuric acid. The assays were run in duplicate wells. A microplate reader (Bio-Rad) was used to measure the absorbance at 450 nm [33].
Cytokine assay
The assay was carried out as described previously [34]. The cellular immune response was assessed indirectly by measuring the levels of IFN-γand IL-4 in peripheral blood lymphocytes. The blood was collected at day 30 after the primary immunization with wtSPV, rSPV-St, inactivated-CV777, inactivated-SQ2014 or PBS. Blood lymphocytes were isolated, counted, and diluted to a density of 2 × 107 cells/100 μl. The cells were aliquoted into 96-well plates (100 μl/well), and 100 μl of DMEM containing 200 TCID50 of PEDV SQ2014 was added to each well. After incubation for 60 h, the supernatants were tested using an ELISA kit (ExCell Bio, China) according to the manufacturer’s instructions. Standard curves were generated using control IFN- and IL-4 that were serially diluted in PBS and coated onto ELISA plates overnight at 37 °C. The levels of serum IFN-γ and IL-4 were calculated according to standard curves.
Microneutralization assay
The microneutralization assay was performed as described previously [24, 25]. Sera were collected at 0, 7, 21, 35, and 42 days after primary immunization from pigs immunized with inactivated SQ2014, inactivated CV777, wtSPV, rSPV-St, or PBS. One hundred µl of twofold serially diluted serum samples was mixed with an equal volume of 70 PFU of PEDV SQ2014 at 37 °C. After a 1.5-h incubation, the antibody-virus mixtures were used to infect Vero cells in 96-well plates, and the cells were overlaid with agar. The cells were incubated for three days at 37 °C in a 5% CO2 atmosphere. The microneutralization titer was expressed as the reciprocal of the highest serum dilution that inhibited the cytopathic effect (CPE).
Statistical analysis
All data were analyzed using one-way ANOVA. The threshold for significance was established at P < 0.05.
Results
Engineering of rSPV-St
To engineer an effective live PDEV vaccine, we used SPV as the vector to express the St of the novel PEDV strain SQ2014. The resultant vaccine was referred as rSPV-St. The insertion of the St gene was confirmed by PCR, which amplified a 1257-bp fragment of the S gene (Fig. 1B). The infectious titers of rSPV-St and wtSPV were both approximately 1 × 108 PFU/mL. PK-15 cells infected with rSPV-St showed green fluorescence when observed by fluorescence microscopy, as shown in Fig. 1C, and the expression of the St protein was confirmed by IFA (Fig. 1D). In addition, when probed with convalescent serum from a PEDV-infected pig, a 46-kDa band, consistent with the predicted size of the St protein, was detected by western blot in cells infected with rSPV-St, but not wtSPV (Fig. 1E). Taken together, these observations indicated that the St protein was efficiently expressed by rSPV-St. In addition, rSPV-St exhibited growth characteristic similar to that of wtSPV (Fig. 1F).
Safety of rSPV-St in pigs
The safety of the rSPV-St vaccine was assessed in 30-day-old pigs for up to 2 weeks post-immunization. Five days post-immunization, pigs that received wtSPV and rSPV-St had pox marks approximately 0.2 cm in diameter around the sites of injection. By 10 days post-immunization, the pox marks were substantially reduced, and all of the pigs recovered after 2 weeks. There were no indications of abnormal body temperature or appetite in any of the vaccinated animals. These results demonstrated that both rSPV-St and wtSPV were well tolerated in pigs.
Humoral and cellular immune response to rSPV-St in pigs
The humoral immune response to vaccination with rSPV-St was assessed by indirect ELISA. Pigs received two vaccinations, on days 0 and 21, and an antibody response was detected as early as day 14. The S-specific immune response increased gradually over time in animals immunized with rSPV-St (Fig. 2A). The IgG titers in animals vaccinated with rSPV-St were significantly higher than those that received wtSPV or PBS but were lower than those in the inactivated SQ2014- and CV777- vaccinated groups (P < 0.01; n = 5). The IgA titers in the rSPV-St group were significantly higher than in animals that received inactivated vaccines or the PBS control (Fig. 2B). In addition, neutralizing antibodies were detected in the rSPV-St group with a mean titer of 24 at 42 days post-vaccination. The neutralizing antibody titers of rSPV-St group were lower than those in the inactivated SQ2014 group but were higher than those in the group vaccinated with inactivated-CV777 (Fig. 3B; P < 0.05; 5 animals per time point).
Fig. 2.

Humoral immune response to rSPV-St in pigs. (A) IgG antibodies (OD values) against PEDV were detected by indirect ELISA. (B) IgA antibodies (OD values) were detected by indirect ELISA. Data are shown as the mean ± S.D
Fig. 3.
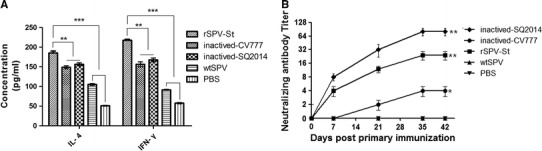
Cellular immune response and neutralizing antibody titers induced by rSPV-St. Pigs (1 month old) were immunized with rSPV-St, and the cellular immune response and neutralizing antibody titers were measured. (A) The IL-4 and IFN-γ levels post-vaccination. (B) The neutralizing antibody titers were determined using twofold serial dilutions. The titers of neutralizing antibodies were expressed as the reciprocal of the highest serum dilution that prevented CPE. Data are shown as the mean ± S.D
We examined the cellular immune response in pigs vaccinated with inactivated SQ2014, inactivated CV777, rSPV-St, wtSPV and PBS on day 30 post-immunization (Fig. 3A). The concentrations of IL-4 and IFN-γ in rSPV-St-vaccinated pigs were the highest among the five experimental groups, while the inactivated vaccines resulted in higher IL-4 and IFN-γ concentrations than did wtSPV and PBS.
Efficacy of passive immunity from rSPV-St vaccination
One of the most common practices for protecting piglets from PEDV infection is passive transfer of maternal antibodies from vaccinated sows. To examine the protective efficacy of antibodies induced by the rSPV-St vaccine, piglets were fed sera from animals vaccinated with rSPV-St, inactivated SQ2014, inactivated CV777, wtSPV, or PBS. On the following day, animals were challenged with either SQ2014 or CV777. The piglets fed sera from pigs vaccinated with rSPV-St and inactivated vaccines (inactivated SQ2014, inactivated CV777) were fully protected against the homologous virus challenge, while all animals succumbed to the challenge with the heterologous virus (Fig. 4A and B, Tables 1 and 2). As expected, animals fed sera from pigs vaccinated with wtSPV or PBS exhibited clinical signs including diarrhea, loss of appetite, reduced weight gain, and increased mental instability, and all animals died between 8 and 10 days post-challenge. Histological examination revealed no lesions in the intestinal tissues from piglets in the homologous challenge groups. However, evident histological changes were observed in tissues from animals that died from the heterologous virus challenge, as well as those that received sera from wtSPV or PBS vaccination but died from either virus challenge. These changes included degeneration and necrosis of intestinal microvilli and infiltration of inflammatory cells (Fig. 5, Table 3). Taken together, these results show that the sera from the rSPV-St and inactivated-SQ2014-immunized animals provided complete protection against infection with the PEDV SQ2014 strain upon passive transfer.
Fig. 4.
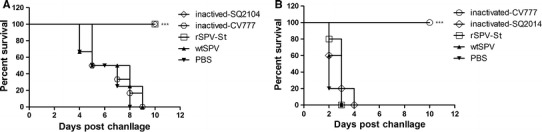
Efficacy of passively transferred PEDV immunity in piglets. Piglets were fed immune sera from pigs vaccinated with inactivated SQ2014, inactivated CV777, rSPV-Sa, wtSPV or PBS. Survival was followed for 12 days after challenge with either (A) SQ2014 or (B) CV777
Table 1.
Severity of disease in pigs passively transferred with sera and challenged with PEDV SQ2014
| Sow group | Days after challenge with PEDV strain SQ2014 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| rSPV-St | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d |
| Inactivated-SQ2014 | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d |
| Inactivated-CV777 | +a/–b/-c/+d | ++a/+b/+c/+d | +++a/++b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | ND | ND |
| wtSPV | +a/–b/-c/+d | ++a/+b/+c/+d | +++a/++b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | ND | ND | ND |
| PBS | +a/–b/-c/+d | ++a/+b/+c/+d | +++a/++b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | ND | ND | ND | ND |
ND: not determined due to the death of this piglet
a Severity of diarrhea: – no diarrhea; + mild diarrhea; ++ moderate diarrhea; +++ severe diarrhea
b Degree of appetite loss: – normal appetite; + slight loss of appetite; ++ moderate loss of appetite; +++ no appetite
c Loss of mental stability: - mental stability; + mentally unstable
d PEDV detection in feces by RT-PCR: - negative; + positive
Table 2.
Severity of disease after passive transfer of sera and challenge with PEDV CV777
| Sow group | Days after challenge with PEDV strain CV777 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| rSPV-St | ++a/+b/+c/+d | +++a/+++b/+c/+d | ND | ND | ND | ND | ND | ND | ND | ND |
| Inactivated-CV777 | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d | –a/–b/-c/-d |
| Inactivated-SQ2014 | ++a/+b/+c/+d | +++a/+++b/+c/+d | +++a/+++b/+c/+d | ND | ND | ND | ND | ND | ND | ND |
| wtSPV | ++a/+b/+c/+d | +++a/+++b/+c/+d | ND | ND | ND | ND | ND | ND | ND | ND |
| PBS | ++a/+b/+c/+d | +++a/++b/+c/+d | ND | ND | ND | ND | ND | ND | ND | ND |
ND: not determined due to the death of this piglet
a Severity of diarrhea: – no diarrhea; + mild diarrhea; ++ moderate diarrhea; +++ severe diarrhea
b Degree of appetite loss: – normal appetite; + slight loss of appetite; ++ moderate loss of appetite; +++ no appetite
c Loss of mental stability: - mental stability; + mentally unstable
d PEDV detection in feces by RT-PCR: - negative; + positive
Fig. 5.
Representative images of histological changes in the small intestine from piglets fed sera from pigs immunized with (a) rSPV-St, (b) inactivated SQ2014, (c) wtSPV, or (d) PBS, followed by challenge with PEDV SQ2014. The samples were obtained 3 days post-challenge. The intestinal tissue from a normal healthy pig (e) was included for comparison. The tissues were stained with hematoxylin and eosin (HE). Magnification, 100×
Table 3.
Evaluation of intestinal lesions in pigs from different treatment groups
| Group | n | Gross scorea | Microscopic scoreb |
|---|---|---|---|
| rSPV-St | 5 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Inactivated-SQ2014 | 5 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| wtSPV | 5 | 6.55 ± 0.35* | 7.05 ± 0.55* |
| PBS | 5 | 5.95 ± 0.45* | 7.85 ± 0.65* |
| Blank control group | 5 | 0.00 ± 0.00 | 0.00 ± 0.00 |
* Indicates significantly (P < 0.05) higher lesion scores of the wtSPV- and PBS-treated groups compared to groups immunized with rSPV-St and inactivated-SQ2014 vaccines
a The gross score was obtained by summing I and III according to the following description and lesion scores from 0 (normal) to 10 (severe lesion). Shown are mean ± S.D
I. Watery stool in small intestine: 0 (none) to 4 (severe diarrhea)
II. Small intestine bleeding: 0 (no bleeding) to 3 (severe bleeding)
III. Small intestine flatulence: 0 (no flatulence) to 3 (severe flatulence)
b The microscopic score was obtained by summing I to IV according to the following description and lesion scores from 0 (normal) to 10 (severe lesion). Shown are mean ± S.D
I. Infiltrations of inflammatory cells: 0, none; 1 (mild); 2 (moderate); 3 (severe)
II. Small intestine villi degeneration and necrosis: 0 (no lesion); 1 (mild); 2 (moderate); 3 (severe)
III. Inflammatory infiltration involvement of different layers of intestine: 0 (none) to 4 (severe)
Sequence differences in the S proteins of SQ2014 and CV777
The amino acid sequences in the S proteins of the two viruses were compared. Sequence alignment indicated that the amino acid sequence identity between the two strains was 96%. However, the S protein of SQ2014 had an amino acid deletion at position 152 from the N-terminus, within a neutralizing epitope [11–14], and there were three consecutive amino acid differences at positions 151-153 [11–14]. There were also seven amino acid mutations in three previously predicted antigenic domains, 499-638, 748-755 and 764-771 (Fig. 6). The difference between the sequences of neutralizing epitope(s) may have contributed to the observed differences in the clinical response to vaccination with rSPV-St.
Fig. 6.

Alignment of the St aa sequences from the PEDV strains SQ2014 and CV777. The neutralizing epitopes are boxed. The GenBank accession numbers of the viruses are listed in the text
Discussion
We have engineered a live SPV vaccine that expresses the truncated S protein (rSPV-St) from the novel PEDV strain SQ2014. The vaccine induced a robust immune response to the S antigen of PEDV in pigs and provided full protection against homologous challenge with wt SQ2014. Our results suggest that rSPV-St is a promising live vaccine candidate against PEDV. It has been reported that some epitopes are conserved among various PEDV strains [35–37]. However, it is unclear whether these different strains of PEDV can induce a cross-immune response in pigs. Most commercial PED vaccines in China are inactivated vaccines, the protective efficacy of which is weakened due to the different virus genotypes used for vaccine generation [37]. Furthermore, inactivated vaccines do not induce an efficient mucosal immune response, and this is a potential reason why there are many reports about the lack of efficacy of commercial PEDV vaccines from 2010.
The S protein is a key component of coronaviruses and contains neutralizing epitopes, but it is genetically variable, which increases the difficulty of generating a vaccine against PEDV. Our results indicated that vaccination with inactivated SQ2014 and inactivated CV777 resulted in effective protection against a homologous virus challenge, but not a heterologous virus challenge. rSPV-St, expressing the truncated S protein from the SQ2014 strain, did not provide any protection against the classical CV777 strain, indicating a lack of cross-protection. These results may also provide insight into why commercial vaccines developed from classical PEDV strains such as CV777 are no longer effective in China due to the emergence of novel PEDV strains.
Although SPV caused mild lesions in our study, the reaction around the injection site subsided quickly without leaving a scar in the skin. The pig immune reactions in our experiments were interesting. Immunization with rSPV-St resulted in the highest IgA antibody titer and cytokine response among the five experimental groups, but the IgG titers were lower than those elicited by inactivated SQ2014. However, rSPV-St vaccine provided efficient protection in newborn pigs that was as effective as that provided by the inactivated SQ2014 vaccine. These findings indicated that the neutralizing antibody titer is not the only criterion for whether a vaccine is effective; the IgA antibody titer and cytokines also contribution to protection. These results show that the efficacy of a vaccine cannot be evaluated by only measuring the neutralizing antibody titer in vitro.
In recent years, poxvirus vector vaccine research has attracted widespread attention. Vaccinia, fowlpox and canarypox viruses have numerous potential applications. A FPV vector has been used in a commercial poultry vaccine against avian influenza virus [23]. The use of poxvirus as a vaccine delivery vector has been extensively explored in mammalian and avian species [38, 39]. SPV has also aroused the interest of researchers. Our data suggest that the SPV live vaccine could induce a more sustainable humoral immune response than the inactive vaccine. Furthermore, the live SPV-based vaccines induced a stronger mucosal immune response, while the inactivated vaccines failed to induce a strong mucosal immune response and elicited lower IgA antibody titers.
Compared to the traditional attenuated or inactivated vaccines, the novel SPV vector vaccine has an advantage in terms of the route of immunization. This novel SPV vector vaccine can trigger the immune response simply by oral administration or subcutaneous piercing without any immunoadjuvant. It not only saves effort during vaccination but also results in less stress to the animals. Thus, this novel SPV vaccine shows a promising future for application in the swine industry.
Given that SPV is in itself a pathogen that elicits an immune response, endemic SPV infection may represents a concern for vaccination using a recombinant SPV. To address this problem, we collected 2000 random serum samples from reserve and feeder pigs to determine the rate of SPV positivity by agarose gel diffusion assay (data not shown). The SPV positive rate was approximately 1%, and there have been no reports of an SPV outbreak in recent years. Therefore, endemic SPV is unlikely to interfere with vaccination programs using an rSPV vector.
In summary, we describe the immunogenicity and protective efficacy of a recombinant SPV-based PEDV vaccine expressing a truncated S protein. The results indicate that both inactivated SQ2014 and rSPV-St vaccine induced complete protection in a passive-transfer piglet model upon homologues virus challenge but failed to protect against heterologous virus challenge. It is expected that a PEDV multivalent vaccine constructed using SPV will be able to prevent both homologous and heterologous viral infections.
Acknowledgements
This study was supported by the National Transgenic Major Program (2014ZX0800946B), the Special Fund for Agro-Scientific Research in the Public Interest (201403054), the Jiangsu Agricultural Science and Technology Innovation Fund (CX(16)1028), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Author contributions statement
X.Y., H.L. and H.F. designed the study. X.Y., H.L. and H.F. performed and collected data from experiment and analyzed data. B.L and K.H participated in animal experiments. X.Y. wrote the manuscript.
Compliance with ethical standards
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies with human participants by any of the authors. All experiments were conducted at the College of Veterinary Medicine, Nanjing Agricultural University. Animal experiments were approved by the Laboratory Animal Monitoring Committee of Jiangsu Province. The study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals. Protocols for the animal studies were approved by the Committee on the Ethics of Animal Experiments of the College of Veterinary Medicine, Nanjing Agricultural University.
Funding
The authors of this paper have no financial or personal relationship with other people or organizations that could inappropriately influence or bias the content of the paper.
Footnotes
Xiaomin Yuan and Huixing Lin contributed equally to this work.
References
- 1.Debouck P, Pensaert M. Experimental infection of pigs with a new porcine enteric coronavirus, CV 777. Am J Vet Res. 1980;41(2):219–223. [PubMed] [Google Scholar]
- 2.Pijpers A, van Nieuwstadt AP, Terpstra C, Verheijden JH. Porcine epidemic diarrhoea virus as a cause of persistent diarrhoea in a herd of breeding and finishing pigs. Vet Rec. 1993;132(6):129–131. doi: 10.1136/vr.132.6.129. [DOI] [PubMed] [Google Scholar]
- 3.Pensaert MB, de Bouck P. A new coronavirus-like particle associated with diarrhea in swine. Arch Virol. 1978;58(3):243–247. doi: 10.1007/BF01317606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sun RQ, Cai RJ, Chen YQ, Liang PS, Chen DK, Song CX. Outbreak of porcine epidemic diarrhea in suckling piglets, China. Emerg Infect Dis. 2012;18(1):161–163. doi: 10.3201/eid1801.111259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Crawford K, Lager KM, Kulshreshtha V, Miller LC, Faaberg KS. Status of vaccines for porcine epidemic diarrhea virus in the United States and Canada. Virus Res. 2016;226:108–116. doi: 10.1016/j.virusres.2016.08.005. [DOI] [PubMed] [Google Scholar]
- 6.Jinghui F, Yijing L. Cloning and sequence analysis of the M gene of porcine epidemic diarrhea virus LJB/03. Virus Genes. 2005;30(1):69–73. doi: 10.1007/s11262-004-4583-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kusanagi K, Kuwahara H, Katoh T, Nunoya T, Ishikawa Y, Samejima T, Tajima M. Isolation and serial propagation of porcine epidemic diarrhea virus in cell cultures and partial characterization of the isolate. J Vet Med Sci. 1992;54(2):313–318. doi: 10.1292/jvms.54.313. [DOI] [PubMed] [Google Scholar]
- 8.Song D, Park B. Porcine epidemic diarrhoea virus: a comprehensive review of molecular epidemiology, diagnosis, and vaccines. Virus Genes. 2012;44(2):167–175. doi: 10.1007/s11262-012-0713-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Takahashi K, Okada K, Ohshima K. An outbreak of swine diarrhea of a new-type associated with coronavirus-like particles in Japan. Nihon Juigaku Zasshi. 1983;45(6):829–832. doi: 10.1292/jvms1939.45.829. [DOI] [PubMed] [Google Scholar]
- 10.Kocherhans R, Bridgen A, Ackermann M, Tobler K. Completion of the porcine epidemic diarrhoea coronavirus (PEDV) genome sequence. Virus Genes. 2001;23(2):137–144. doi: 10.1023/A:1011831902219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Duarte M, Tobler K, Bridgen A, Rasschaert D, Ackermann M, Laude H. Sequence analysis of the porcine epidemic diarrhea virus genome between the nucleocapsid and spike protein genes reveals a polymorphic ORF. Virology. 1994;198(2):466–476. doi: 10.1006/viro.1994.1058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jackwood MW, Hilt DA, Callison SA, Lee CW, Plaza H, Wade E. Spike glycoprotein cleavage recognition site analysis of infectious bronchitis virus. Avian Dis. 2001;45(2):366–372. doi: 10.2307/1592976. [DOI] [PubMed] [Google Scholar]
- 13.Lee DK, Park CK, Kim SH, Lee C. Heterogeneity in spike protein genes of porcine epidemic diarrhea viruses isolated in Korea. Virus Res. 2010;149(2):175–182. doi: 10.1016/j.virusres.2010.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sun DB, Feng L, Shi HY, Chen JF, Liu SW, Chen HY, Wang YF. Spike protein region (aa 636789) of porcine epidemic diarrhea virus is essential for induction of neutralizing antibodies. Acta Virol. 2007;51(3):149–156. [PubMed] [Google Scholar]
- 15.Chen Q, Li G, Stasko J, Thomas JT, Stensland WR, Pillatzki AE, Gauger PC, Schwartz KJ, Madson D, Yoon KJ, Stevenson GW, Burrough ER, Harmon KM, Main RG, Zhang J. Isolation and characterization of porcine epidemic diarrhea viruses associated with the 2013 disease outbreak among swine in the United States. J Clin Microbiol. 2014;52(1):234–243. doi: 10.1128/JCM.02820-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Park SJ, Moon HJ, Yang JS, Lee CS, Song DS, Kang BK, Park BK. Sequence analysis of the partial spike glycoprotein gene of porcine epidemic diarrhea viruses isolated in Korea. Virus Genes. 2007;35(2):321–332. doi: 10.1007/s11262-007-0096-x. [DOI] [PubMed] [Google Scholar]
- 17.Lee S. Lee C (2014) Outbreak-related porcine epidemic diarrhea virus strains similar to US strains, South Korea. Emerg Infect Dis. 2013;20(7):1223–1226. doi: 10.3201/eid2007.140294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chang SH, Bae JL, Kang TJ, Kim J, Chung GH, Lim CW, Laude H, Yang MS, Jang YS. Identification of the epitope region capable of inducing neutralizing antibodies against the porcine epidemic diarrhea virus. Mol Cells. 2002;14(2):295–299. [PubMed] [Google Scholar]
- 19.Vinicor F, Jack L., Jr 25 years and counting: Centers for Disease Control and Prevention identifies opportunities and challenges for diabetes prevention and control. Ann Intern Med. 2004;140(11):943–944. doi: 10.7326/0003-4819-140-11-200406010-00034. [DOI] [PubMed] [Google Scholar]
- 20.Pastoret PP, Brochier B. Development of a recombinant vaccinia-rabies vaccine for oral vaccination of foxes against rabies. Dev Biol Stand. 1992;79:105–111. [PubMed] [Google Scholar]
- 21.Taylor J, Weinberg R, Languet B, Desmettre P, Paoletti E. Recombinant fowlpox virus inducing protective immunity in non-avian species. Vaccine. 1988;6(6):497–503. doi: 10.1016/0264-410X(88)90100-4. [DOI] [PubMed] [Google Scholar]
- 22.Barcena J, Blasco R. Recombinant swinepox virus expressing beta-galactosidase: investigation of viral host range and gene expression levels in cell culture. Virology. 1998;243(2):396–405. doi: 10.1006/viro.1998.9053. [DOI] [PubMed] [Google Scholar]
- 23.Hahn J, Park SH, Song JY, An SH, Ahn BY. Construction of recombinant swinepox viruses and expression of the classical swine fever virus E2 protein. J Virol Methods. 2001;93(1–2):49–56. doi: 10.1016/S0166-0934(00)00296-2. [DOI] [PubMed] [Google Scholar]
- 24.Huang D, Zhu H, Lin H, Xu J, Lu C. First insights into the protective effects of a recombinant swinepox virus expressing truncated MRP of Streptococcus suis type 2 in mice. Berl Munch Tierarztl Wochenschr. 2012;125(3–4):144–152. [PubMed] [Google Scholar]
- 25.Xu J, Huang D, Liu S, Lin H, Zhu H, Liu B, Lu C. Immune responses and protection efficacy of a recombinant swinepox virus expressing HA1 against swine H3N2 influenza virus in mice and pigs. Virus Res. 2012;167(2):188–195. doi: 10.1016/j.virusres.2012.04.014. [DOI] [PubMed] [Google Scholar]
- 26.Lin HX, Ma Z, Yang XQ, Fan HJ, Lu CP. A novel vaccine against Porcine circovirus type 2 (PCV2) and Streptococcus equi ssp. zooepidemicus (SEZ) co-infection. Vet Microbiol. 2014;171(1–2):198–205. doi: 10.1016/j.vetmic.2014.03.018. [DOI] [PubMed] [Google Scholar]
- 27.Lin HX, Huang DY, Wang Y, Lu CP, Fan HJ. A novel vaccine against Streptococcus equi ssp. zooepidemicus infections: the recombinant swinepox virus expressing M-like protein. Vaccine. 2011;29(40):7027–7034. doi: 10.1016/j.vaccine.2011.07.074. [DOI] [PubMed] [Google Scholar]
- 28.Soprana E, Panigada M, Knauf M, Radaelli A, Vigevani L, Palini A, Villa C, Malnati M, Cassina G, Kurth R, Norley S, Siccardi AG. Joint production of prime/boost pairs of Fowlpox Virus and Modified Vaccinia Ankara recombinants carrying the same transgene. J Virol Methods. 2011;174(1–2):22–28. doi: 10.1016/j.jviromet.2011.03.013. [DOI] [PubMed] [Google Scholar]
- 29.Torres JM, Sanchez C, Sune C, Smerdou C, Prevec L, Graham F, Enjuanes L. Induction of antibodies protecting against transmissible gastroenteritis coronavirus (TGEV) by recombinant adenovirus expressing TGEV spike protein. Virology. 1995;213(2):503–516. doi: 10.1006/viro.1995.0023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yuan X, Lin H, Fan H. Efficacy and immunogenicity of recombinant swinepox virus expressing the A epitope of the TGEV S protein. Vaccine. 2015;33(32):3900–3906. doi: 10.1016/j.vaccine.2015.06.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ishikawa K, Sekiguchi H, Ogino T, Suzuki S. Direct and rapid detection of porcine epidemic diarrhea virus by RT-PCR. J Virol Methods. 1997;69(1–2):191–195. doi: 10.1016/S0166-0934(97)00157-2. [DOI] [PubMed] [Google Scholar]
- 32.Sun M, Ma J, Wang Y, Wang M, Song W, Zhang W, Lu C, Yao H. Genomic and epidemiological characteristics provide new insights into the phylogeographical and spatiotemporal spread of porcine epidemic diarrhea virus in Asia. J Clin Microbiol. 2015;53(5):1484–1492. doi: 10.1128/JCM.02898-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Tang M, Harp JA, Wesley RD. Recombinant adenovirus encoding the HA gene from swine H3N2 influenza virus partially protects mice from challenge with heterologous virus: A/HK/1/68 (H3N2) Arch Virol. 2002;147(11):2125–2141. doi: 10.1007/s00705-002-0870-y. [DOI] [PubMed] [Google Scholar]
- 34.Du Y, Li Y, He H, Qi J, Jiang W, Wang X, Tang B, Cao J, Jiang P. Enhanced immunogenicity of multiple-epitopes of foot-and-mouth disease virus fused with porcine interferon alpha in mice and protective efficacy in guinea pigs and swine. J Virol Methods. 2008;149(1):144–152. doi: 10.1016/j.jviromet.2007.12.018. [DOI] [PubMed] [Google Scholar]
- 35.Chen F, Zhu Y, Wu M, Ku X, Ye S, Li Z, Guo X, He Q. Comparative genomic analysis of classical and variant virulent parental/attenuated strains of porcine epidemic diarrhea virus. Viruses. 2015;7(10):5525–5538. doi: 10.3390/v7102891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Islam MT, Kubota T, Ujike M, Yahara Y, Taguchi F. Phylogenetic and antigenic characterization of newly isolated porcine epidemic diarrhea viruses in Japan. Virus Res. 2016;222:113–119. doi: 10.1016/j.virusres.2016.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lin CM, Gao X, Oka T, Vlasova AN, Esseili MA, Wang Q, Saif LJ. Antigenic relationships among porcine epidemic diarrhea virus and transmissible gastroenteritis virus strains. J Virol. 2015;89(6):3332–3342. doi: 10.1128/JVI.03196-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chen H, Huang Q, Cui B, Li X, Guan Q. Construction of recombinant fowlpox virus coexpressing HA from subtype H5 of avian influenza virus and chicken interleukin-18. Wei Sheng Wu Xue Bao. 2008;48(8):1025–1030. [PubMed] [Google Scholar]
- 39.Kyriakis CS, De Vleeschauwer A, Barbe F, Bublot M, Van Reeth K. Safety, immunogenicity and efficacy of poxvirus-based vector vaccines expressing the haemagglutinin gene of a highly pathogenic H5N1 avian influenza virus in pigs. Vaccine. 2009;27(16):2258–2264. doi: 10.1016/j.vaccine.2009.02.006. [DOI] [PubMed] [Google Scholar]