

Abstract
Marine sponges and their associated bacteria have been proven to be a rich source of novel secondary metabolites with therapeutic usefulness in cancer, infection, and autoimmunity. In this study, 79 strains belonging to 20 genera of the order Actinomycetales and seven strains belonging to two genera of the order Sphingomonadales were cultivated from 18 different Caribbean sponges and identified by 16S rRNA gene sequencing. Seven of these strains are likely to represent novel species. Crude extracts from selected strains were found to exhibit protease inhibition against cathepsins B and L, rhodesain, and falcipain-2 as well as immunomodulatory activities such as induction of cytokine release by human peripheral blood mononuclear cells. These results highlight the significance of marine sponge-associated bacteria to produce bioactive secondary metabolites with therapeutic potential in the treatment of infectious diseases and disorders of the immune system.
Electronic supplementary material
The online version of this article (doi:10.1007/s10126-010-9349-0) contains supplementary material, which is available to authorized users.
Keywords: Actinomycetes, Sphingomonads, Marine sponge, Anti-protease, Immunomodulatory, Phylogenetic analysis
Introduction
Infection, cancer, and autoimmunity pose serious problems to mankind (WHO 2009), and thus, there is a clear and continued need for new therapeutic agents against human diseases (Caspi 2008). Natural product discovery programs have yielded an exhaustive supply of biologically active secondary metabolites from terrestrial plants and microorganisms (Challis 2008). In the last four decades, drug discovery efforts have shifted to the marine environment where invertebrates (sponges, soft corals, ascidians), and plants have been subject to elaborate screening programs. In recent years, marine microorganisms, either free living or as symbionts, have moved into the forefront of attention (Gulder and Moore 2009) fueled largely by the recognition of untapped actinomycete diversity in the marine environment.
Many marine sponges are associated with dense and phylogenetically diverse microbial consortia that can account for nearly half of the animal’s biomass (Hentschel et al. 2006; Taylor et al. 2007). Of special interest are members of the phylum Actinobacteria which have been identified in sponges both by cultivation and cultivation-independent approaches (Abdelmohsen et al. 2010; Hentschel et al. 2002; Jiang et al. 2008; Kim et al. 2005; Montalvo et al. 2005; Pimentel-Elardo et al. 2010; Webster et al. 2001; Zhang et al. 2008). Actinobacteria and specifically members of the order actinomycetes are in fact quite abundant in the marine environment (Maldonado et al. 2005; Mincer et al. 2002; Stach and Bull 2005). A number of obligately marine actinomycete species and genera have been described which indicates true adaptations to the marine environment rather than them simply being washed into the ocean as spores from terrestrial soils. The actinomycetes are of particular relevance due to their unmatched capacity to produce novel and bioactive secondary metabolites. About 7,000 compounds have been isolated from this bacterial taxon alone (Jensen et al. 2005; Lam 2006). The anticancer compounds salinosporamide and sporolide from the actinomycete Salinispora tropica (Buchanan et al. 2005; Fenical et al. 2009), as well as the antitumor antibiotic marinomycin (Kwon et al. 2006) from the obligately marine genus Marinispora, are just two examples of metabolites from marine actinomycetes.
In this study, we aimed to isolate taxonomically novel actinomycetes and sphingomonads from diverse Caribbean sponges. Sphingomonads were included in this study because of their reported immunomodulatory activities due to specific glycosphingolipids (Long et al. 2007). Organic extracts of the bacterial cultures were then subjected to two screenings: (a) anti-protease assays using human, parasitic, and viral proteases as targets. Proteases are relevant drug targets in cardiovascular, inflammatory, and infectious disease areas with about 32 protease inhibitors currently approved for clinical use (Turk 2006). One prominent example is Ritonavir, an aspartic protease inhibitor of HIV-1 virus which has been in clinical use since 1996 for the treatment of AIDS (Danner et al. 1995), and (b) immunomodulatory activities in terms of cytokine release in cultures of human peripheral blood mononuclear cells (PBMC) and induction of cell proliferation. Immunoadjuvants from bacterial cells and their fractions have been used in vaccination and immune therapy (Azuma and Seya 2001). For example, immunostimulatory CpG oligonucleotides have been found to induce maturation, differentiation, and/or proliferation of multiple cell types (Klinman et al. 2010).
Materials and Methods
Sponge Collection
The first group of sponges (Aplysina fistularis, Plakortis sp., Amphimedon compressa, Aiolochroia crassa, Agelas clathrodes, Agelas cerebrum, Ircinia felix, Scopalina ruetzleri, Erylus formosus, Chondrilla nucula, and Aplysina archeri) was collected by SCUBA diving at depths of 3–20 m in Bahamas in July 2008 (GPS—26°27′3.25″ N, 77°54′14.59″ W). The second group of sponges (A. clathrodes, Aplysina insularis, Agelas tubulata, Biemna cribaria, A. crassa, Discodermia dissoluta, S. ruetzleri, Dragmacidon reticulata, I. felix, Monanchora arbuscula, and Plakinastrella onkodes) was collected at depths of 11–23 m including sediments and seawater in El Morro, Santa Marta Bay, Colombia (GPS—11°14′59.04″ N, 74°13′44.47″ W) in December 2008. Sponges were transferred to plastic bags containing seawater and immediately transported to the laboratory. Sponge specimens were rinsed in sterile seawater, cut into pieces of ca. 1 cm3, and then thoroughly homogenized in a sterile mortar with 10 volumes of sterile seawater. The supernatant was diluted in ten-fold series (10−1, 10−2, 10−3) and subsequently plated out on agar plates. The same protocol was repeated for sediment and seawater samples.
Bacterial Isolation
Six different media were used for the isolation of Actinobacteria, such as M1 (1 L artificial sea water (ASW) containing 10 g starch, 4 g yeast extract, 2 g peptone, 18 g Bacto Agar, and 1 L ASW), M2 (1 L ASW was made up of 6 ml of 100% glycerol, 1 g arginine, 1 g K2HPO4, 0.5 g MgSO4, and 18 g Bacto Agar; Mincer et al. 2002), ISP medium 2 (1 L ASW containing 4 g yeast extract, 10 g malt extract, 4 g dextrose, and 18 g Bacto Agar; Shirling and Gottlieb 1966), M7 (1 L ASW containing 2 g peptone, 0.1 g asparagine, 4 g sodium propionate, K2HPO4, 0.1 g MgSO4, 0.0001 g FeSO4, 5 g glycerol, 20 g NaCl, and 18 g Bacto Agar; Webster et al. 2001), NaST21Cx (solution A 750 ml of artificial seawater containing 1 g K2HPO4 and 10 g Bacto Agar and solution B 250 ml artificial seawater containing 1 g KNO3, 1 g MgSO4, 1 g CaCl.22H2O, 0.2 g FeCl3, and 0.1 g MnSO4·7H2O; Magarvey et al. 2004), and oligotropic medium (1 L ASW containing 0.5 g tryptone, 0.1 g sodium glycerophosphate, 0.05 g yeast extract, and 12 g Bacto Agar; Santavy et al. 1990). Heat shock (95°C for 10 min) and incubation with 1.5% phenol at 30°C for 30 min were applied to the sponge homogenates to select for spore-forming actinomycetes and rare actinomycete genera. All plates were incubated and observed for growth of colonies for 6–8 weeks. All media were supplemented with the following antibiotics: cycloheximide (100 μg/ml), nystatin, and nalidixic acid (25 μg/ml) to inhibit the growth of fungi and Gram-negative bacteria. Sphingomonads were isolated on M1, ISP medium 2, and oligotropic medium. The isolates from the Bahamas are abbreviated as “BA” and those from Colombia as “CO”.
Molecular Identification and Phylogenetic Analysis
The 16S rRNA genes from all Bahamian sponge isolates were amplified, cloned, and sequenced according to Hentschel et al. (2001) using the universal primers 27F and 1492R (Lane 1991). Isolates from the Colombian sponges were first sorted into groups according to their morphological characteristics. Restriction length fragment polymorphism (RFLP) analysis was then performed on all isolates to reduce strain replication. Following 16S rRNA gene amplification using the universal primers 27F and 1492R (Lane 1991) and 341F and 907R (Muyzer et al. 1993), the PCR products were digested with the restriction enzymes HaeIII and MspI for 2 h. One to two isolates from each RFLP group were selected for 16S rRNA gene sequencing.
Chimeric sequences were identified using the Pintail program (Ashelford et al. 2005). Sequence alignment and phylogenetic analysis were performed using the ARB software package (Ludwig et al. 2004). The genus-level affiliation of the isolates was validated using the Ribosomal Database Project Classifier (Wang et al. 2007). Tree construction was conducted using neighbor-joining algorithm (Jukes–Cantor correction) with bootstrap values based on 1,000 replications. The 16S rRNA gene sequences of the putatively novel isolates were deposited in GenBank under the accession numbers indicated in parentheses: BA21 (HM005239), BA53 (HM005240), CO58 (HM005241), CO104 (HM005242), CO105 (HM005243), CO132 (HM005244), and CO155 (HM005245). All other 16S rRNA gene sequences (Suppl. Tables 1 and 2) were deposited in GenBank under the accession numbers: HQ398366–HQ398418.
Organic Extract Preparation
Sixteen strains were selected based on phylogenetic novelty (four actinomycetes, three sphingomonads, Fig. 1a, b) or their affiliation to clades for which secondary metabolites had not been reported (nine strains). The isolates were cultured in 100 ml Erlenmeyer flasks each containing 20 ml liquid M1 medium. The cultures were grown for 4–21 days depending on their growth rate at 30°C while shaking at 150 rpm, and 3-ml culture aliquots were taken at different time points (days 4, 7, 14, and 21); 1.5 ml methanol was added to each culture aliquot for cell lysis, and shaking was continued at 150 rpm for 2 h at room temperature (Shaker SM 30, E. Bühler). The broths were centrifuged in 15-ml Falcon tubes at 5,000 rpm for 15 min at room temperature (Megafuge 1.0R, Heraeus), and the supernatants were dried using a rotary evaporator (Heidolph, Germany). Media control using uninoculated M1 was extracted using the same method above.
Fig. 1.
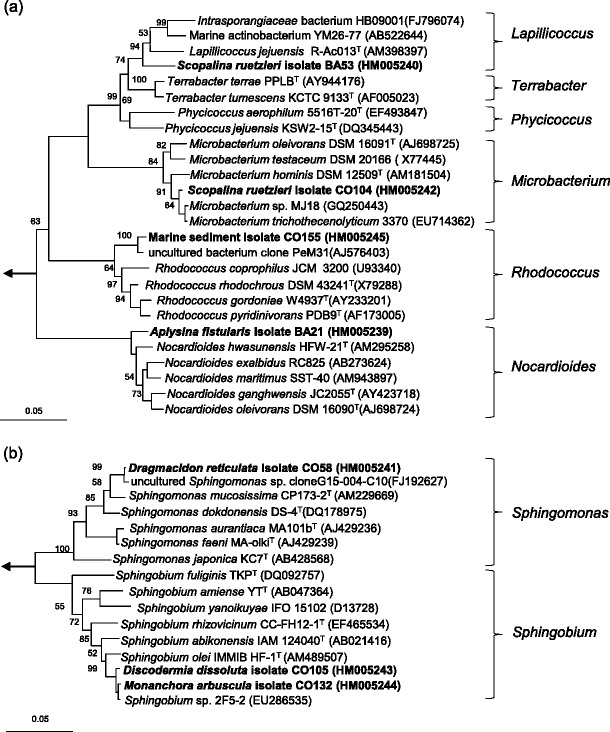
Neighbor-joining trees of isolates and representative species of the order Actinomycetales (a) and Sphingomonadales (b) based on nearly complete 16S rRNA gene sequences. Numbers at the nodes indicate the levels of the bootstrap support based on 1,000 resampled data sets. Only values greater than 50% are shown. The arrow points to the outgroup consisting of four species belonging to Methanosarcinaceae. The scale bar indicates 0.05 substitution per nucleotide position
Eleven strains were grown on M1 agar plates (three square plates per strain with approximately 120 mm diameter each) at 30°C for 7–10 days. Biomass together with the agar was removed from the plates by cutting it into small pieces and macerated overnight with sufficient volume of ethyl acetate to submerge the agar pieces. Extraction with ethyl acetate of the macerated agar was repeated, and the macerations were subsequently filtered by gravity using a Whatman filter paper No. 1. The filtrates were combined and dried by rotary evaporation. Media control using uninoculated M1 agar was extracted using the same method above.
Screening for Anti-protease Activity
Cathepsins B and L and rhodesain protease inhibition assays were performed according to Vicik et al. (2006a, b). Briefly, assays were done at 25°C in a 20-mM Tris–HCl buffer pH 6.0 (cathepsins) or in a 50-mM acetate buffer pH 5.5 (rhodesain) in a total volume of 200 μl. The final substrate concentration was 6.25 and 100.0 mM for cathepsins B and L and 10.0 μM for rhodesain. The final enzyme concentration was 53 ng/ml for cathepsins B and L and 41 nM for rhodesain. The falcipain-2 inhibition assay was performed as described previously by Breuning et al. (2010). Shortly, the enzyme was incubated with crude extracts for 5 min prior to substrate addition in a total volume of 200 μl. The following buffer was used: 50 mM acetate, pH 5.5 supplemented with 5 mM 1,4-dithiothreitol. Substrate (Cbz-Phe-Arg-AMC for all four enzymes) and inhibitor stock solutions were prepared in 10% final concentration dimethyl sulfoxide and were diluted with assay buffer. Crude extracts were tested in duplicates at a final concentration of 20 μg/ml. Protease inhibition assays were carried out on a Cary Eclipse fluorescence spectrophotometer (Varian, Darmstadt, Germany) using a microplate reader (excitation 365 nm, emission 460 nm).
The fluorometric SARS Mpro and Plpro protease inhibition assays were performed according to Kaeppler et al. (2005). Briefly, assays were performed at 25°C in a 20 mM Tris–HCl buffer pH 7.5 in a total volume of 200 μl. The final substrate (H2N-Abz-Ser-Val-Thr-Leu-Gln-Ser-Gly-(NO2)Tyr-Arg-(MTS)-TFA-salt for Mpro and Z-Arg-Leu-Arg-Gly-Gly-AMC-acetate salt for Plpro) concentration for inhibition assays was 50 μM, and the final enzyme concentration was 4.25 μg/ml. Crude extracts were tested in duplicates at a final concentration of 20 μg/ml. Assays were carried out at 325 nm excitation and 425 nm emission.
Screening for Immunomodulatory Activity
PBMC from healthy donors were prepared as a by-product of platelet concentrates obtained with leukoreduction system chambers. The cell concentrate was then diluted in versene (ethylenediaminetetraacetic acid). PBMCs were isolated from this preparation using density gradient centrifugation with lymphocyte separation medium (PAA Laboratories GmbH, Pasching, Austria) and washed with ice-cold buffered salt solution/bovine serum albumin. Cells were counted and cultured in Roswell Park Memorial Institute medium 1640 plus l-glutamine, supplemented with 50 mM mercaptoethanol, MEM non-essential amino acids (100×), 1 mM sodium pyruvate (Gibco BRL, Gaithersburg, MD, USA), 100 U/ml penicillin (Grünenthal GmbH, Aachen, Germany) and 100 U/ml streptomycin sulfate (Riemser Arzeimittel, Greifswald, Germany), 10 mM (2-hydroxyethyl)-1-piperazineethanesulfonic acid (AppliChem GmbH, Darmstadt, Germany), and 10% AB-positive heat-inactivated (30 min at 56°C) human serum (PAA Laboratories GmbH, Pasching, Austria). Cells were stimulated in triplicates using 96-well cell culture plates (Greiner Bio-One, Kremsmünster, Austria; 2 × 105 cells in 200 μl per well) in a humidified incubator at 37°C with 5% CO2. To increase sensitivity of T cells to stimulation, PBMC were precultured for 48 h at 1.5 × 107 cells in 1.5 ml medium in 24-well plates (T.H., unpublished). Supernatants were collected after 24 h and stored at −20°C. A panel of cytokines (TNF, IFN-γ, IL-2, and IL-10) was measured by cytometric bead array (BD Biosciences, San Jose, CA, USA), using an LSR II flow cytometer (BD Biosciences, San Jose, CA, USA) following the manufacturer’s instructions. Results were analyzed using FCAP Array software (Soft Flow, Inc., USA). Forty-eight hours after the stimulation, cell proliferation was measured as radioactivity incorporated into DNA from tritiated thymidine during a 24-h pulse, using a liquid scintillation counter (PerkinElmer, Waltham, MA, USA).
Results and Discussion
Cultivation and Identification of Actinomycetes
A total of 79 actinomycetes were recovered from 18 different Caribbean sponge species, sediment, and seawater, 56 of which were identified based on nearly complete 16S rRNA gene sequence information (Supplementary Table 1). The highest number of actinomycete isolates was recovered from S. ruetzleri (20 isolates), followed by A. insularis (12), D. dissoluta (12), A. fistularis (7), D. reticulata (5), A. clathrodes (5), P. onkodes (4), I. felix (2), Plakortis sp. (3), M. arbuscula (1), A. crassa (1), and E. formosus (1). Actinomycetes were not recovered from the remaining six sponges. Additionally, five isolates were recovered from marine sediments and one isolate from natural seawater. To our knowledge, this is the first report on the isolation of actinomycetes from the sponges A. cerebrum, A. tubulata, A. compressa, A. archeri, B. cribaria, and C. nucula.
In terms of actinomycete diversity, the 79 cultivated strains are represented by 20 different genera namely Microbacterium (21 isolates), Rhodococcus (10), Streptomyces (5), Mycobacterium (6), Micromonospora (5), Knoellia (4), Gordonia (4), Curtobacterium (4), Arthrobacter (4) Salinispora (3), Saccharopolyspora (2), Nocardioides (2), Citromicrobium (2), Sanguibacter (1), Lapillicoccus (1), Kocuria (1), Dietzia (1), Cellulosimicrobium (1) Cellulomonas (1), and Agrococcus (1; Supplementary Table 1).
Several recent publications have shown that Streptomyces, Micromonospora, and Rhodococcus are among the dominant genera commonly isolated from marine sponges (Abdelmohsen et al. 2010; Schneemann et al. 2010; Sun et al. 2010; Zhang et al. 2008). In our study, Microbacterium (27%) was the most dominant strain followed by Rhodococcus (12%) and Streptomyces (9%). Nevertheless, we also present here the isolation of rare Actinobacteria genera namely Cellulosimicrobium, Citromicrobium, Sanguibacter, and Lapillicoccus. Moreover, this is the first report on the cultivation of the genera Citromicrobium, Sanguibacter, and Lapillicoccus from marine sponges and of the genus Lapillicoccus from the marine environment in general.
Remarkably, four actinomycete strains exhibited less than 98.1% 16S rRNA gene sequence similarities to validly described species. These low similarity values suggest that the strains belong to novel taxa (Stackebrandt and Ebers 2006). Phylogenetic analysis (Fig. 1a) revealed that these strains belong to the order Actinomycetales under the following genera: Lapillicoccus, Microbacterium, Rhodococcus, and Nocardioides. Phenotypic and genotypic characterization is further required to confirm the taxonomic affiliation of these strains. Moreover, the isolation of novel actinomycete taxa has previously yielded new chemistry (Feling et al. 2003; Fiedler et al. 2005; Pimentel-Elardo et al. 2008), and thus, this targeted approach offers a more productive route to natural products discovery.
Cultivation and Identification of Sphingomonads
Seven isolates were identified belonging to the order Sphingomonadales and the genera Sphingobium (six isolates) and Sphingomonas (one isolate). Nearly complete 16S rRNA gene information is provided for four isolates (Supplementary Table 2). The low 16S rRNA gene sequence similarities (<98.5%) of two of these strains also suggest the taxonomic novelty at the species level (Fig. 1b). While members of the genus Sphingomonas have previously been isolated from marine organisms such as from a bivalve (Heindl et al. 2010; Romanenko et al. 2007, 2009) and sponges ( Dieckmann et al. 2005; Laroche et al. 2007), we report here on the isolation of members of the genus Sphingobium from a marine source for the first time.
Anti-protease Activity
The crude extracts of 16 isolates cultivated in M1 medium and extracted at four different time points were tested for their anti-protease activities. This involved testing against the human cysteine proteases and cathepsins B and L, which are involved in tumor metastasis (Calkins and Sloane 1995). Extracts were furthermore tested against proteases from the human parasites Plasmodium falciparum (falcipain-2) and Trypanosoma brucei rhodesiense (rhodesain) which are responsible for causing malaria and African trypanosomiasis (sleeping sickness), respectively. Moreover, crude extracts were tested for their capacity to inhibit the papain-like (SARS-CoV PLpro) and the main protease (SARS-CoV Mpro) of the SARS coronavirus. These enzymes are essential for the replication of the severe acute respiratory syndrome (SARS) coronavirus (Anand et al. 2003; Ratia et al. 2006). Cathepsins B and L, rhodesain, and falcipain-2 enzymes belong to the cathepsin L subfamily of cysteine proteases (clan CA, family C1; CAC1). SARS-CoV PLpro also belongs to the clan of cysteine proteases CA but is affiliated to the family C16, which contains polyprotein endopeptidases from coronaviruses. The protease SARS-CoV Mpro belongs to the clan PA (family 30) with a catalytic type of mixed cysteine, serine, and threonine (Rawlings et al. 2010).
The extracts of eight isolates showed a certain specificity to the clan CA (family C1; CAC1) group of proteases, whereas no protease inhibition was observed against viral proteases (Table 1). Extracts were considered active when, at a concentration of 20 μg/ml, an inhibition of at least 40% was observed. The strains Nocardioides sp. BA21, Saccharopolyspora shandongensis strain CO86 and Sphingobium sp. CO132 inhibited rhodesain. The isolates Agrococcus jenensis strain BA22 and Sphingomonas mucosissima strain CO180 inhibited cathepsin B and falcipain-2. Micromonospora coxensis strain CO74 was active only against falcipain-2, and Rhodococcus sp. CO155 was only active against cathepsin L. Extracts of M. coxensis strain CO164 were most active, inhibiting all four proteases tested.
Table 1.
Anti-protease activities of crude extracts from actinomycete and sphingomonad strains grown on M1 media and extracted at different time points (4, 7, 14, and 21 days)
| Strain | % protease inhibition | |||
|---|---|---|---|---|
| Cathepsin B | Cathepsin L | Falcipain-2 | Rhodesain | |
| Actinomycetes | ||||
| Nocardioides sp. BA21a | ND | ND | ND | 40 ± 1e |
| Agrococcus jenensis strain BA22 | 41 ± 1c | ND | 44 ± 2b, 40 ± 4e | ND |
| Micromonospora coxensis strain CO74 | ND | ND | 42 ± 2c | ND |
| Saccharopolyspora shandongensis strain CO86 | ND | ND | ND | 52 ± 1d |
| Rhodococcus sp. CO155a | ND | 44 ± 4d | ND | ND |
| Micromonospora coxensis strain CO164 | 45 ± 3b | 43 ± 2d | 41 ± 2d | 46 ± 3b, 57 ± 5c |
| Sphingomonads | ||||
| Sphingobium sp. CO132a | ND | ND | ND | 53 ± 3d |
| Sphingomonas mucosissima strain CO180 | 49 ± 5b | ND | 45 ± 1d | ND |
Samples were tested in duplicates at a concentration of 20 μg/ml
ND Not detected
aPutatively novel species
b4 days
c21 days
d7 days
e14 days
In fluorometric assays, the enzyme activity is measured by the hydrolysis rate of a fluorogenic or chromogenic substrate (Ludewig et al. 2010). This means that substrate and inhibitor compete for the enzyme’s active site and that depends on the substrate concentration and on the affinity of the substrate to the target enzyme. The inhibition of rhodesain, to which the substrate (Cbz-Phe-Arg-AMC) has a particularly high affinity, by extracts of Nocardioides sp. BA21, S. shandongensis CO86, M. coxensis strain CO164, and Sphingobium sp. CO132, is thus remarkable. Future efforts will be directed at structure elucidation of the anti-protease secondary metabolites from these isolates.
Most of the protease inhibitors reported to date are synthetic molecules developed by structure-based design (Turk 2006). Nevertheless, protease inhibitors have been also found in natural sources. Miraziridine A, a pentapeptide inhibitor of cathepsins B and L, which was isolated from the marine sponge Theonella mirabilis, is one such example (Nakao et al. 2000). A family of aeruginosin inhibitors is active against human serine proteases and was isolated from marine sponges and cyanobacterial waterblooms (Ersmark et al. 2008). Only few protease inhibitors have so far been reported from actinomycetes, leupeptin being a notable exception (Hozumi et al. 1972). In this study, we report on six actinomycete and two sphingomonad isolates as potential sources of novel protease inhibitors.
Stimulation of PBMC
Human PBMC were stimulated with crude extracts prepared from bacterial isolates at four different time points as well as grown on solid media. The CD3-specific monoclonal antibody OKT3, which addresses the T cell antigen receptor complex, was used as a positive control. After 24 h, a panel of cytokines (TNF, IFN-γ, IL-2, and IL-10) was measured in the culture supernatants, and after 48 h, cell proliferation was determined by incorporation of tritiated thymidine.
Human PBMC contain monocytes and lymphocytes, the latter consisting of B cells, T cells (CD4+ T cells, CD8+ T cells, and regulatory T cells), as well as NKT cells. Cytokines are proteins which are secreted by the cells of innate and adaptive immunity and which mediate many cellular functions. In the present study, we chose the following cytokines which are key pro- and anti-inflammatory factors and which are characteristic of particular types of immune responses to pathogens: TNF, the principal mediator of the acute inflammatory response to gram-negative infectious microbes, is responsible for many of the systemic complications of severe infections. IFN-γ is the principal macrophage-activating cytokine and serves critical functions in innate and in adaptive cell-mediated immunity. IL-2 is a cytokine responsible for T cell clonal expansion after antigen recognition. IL-10, a cytokine with anti-inflammatory properties, has a central role in infection by limiting the immune response to pathogens and thereby preventing damage to the host.
When culturing PBMC in the presence of our crude extracts, different patterns of cytokine release were observed. Even though this assay does not distinguish which cells are responsible for cytokine production, we can make some assumptions about their cellular source. This is facilitated by the fact that individual extracts induced quite distinct patterns of cytokine release. For example, A. jenensis strain BA22 was very active in inducing TNF and IL-10, suggesting that this preparation addresses monocytes. In contrast, M. coxensis strain CO164 was a potent inducer of IFN-γ, IL-2, and, to a lesser extent, IL-10. Since the main sources of IFN-γ are TH1 CD4+ T cells and CD8+ T cells, this preparation appears to have T cell activating properties. Furthermore, the induction of IL-2 but not of IFN-γ by S. shandongensis strain CO86 suggests a further specificity for CD4+ T cell activation. Isolates A. jenensis strain BA22, Arthrobacter oxydans strain BA30, and A. oxydans strain BA34 were effective in inducing TNF and IL-10 release (Fig. 2a, d), cytokines induced mainly by monocytes and T cells (TNF: TH1, IL-10: TH2 and Treg). Strains Lapillicoccus sp. BA53 and M. coxensis strain CO164 induced IFN-γ, IL-2, and IL-10 (Fig. 2b–d). IFN-γ and IL-2 can be released by T helper 1 cells and IL-10 by T helper 2 and regulatory T cells as well as monocytes. S. shandongensis strain CO86 effectively induced the release of IL-2 and IL-10 (Fig. 2c, d). IL-2 is mainly released by TH1 cells and IL-10 is mostly produced by macrophages and T helper 2 and regulatory T cells. Sphingobium sp. CO132 induced the release of TNF, IFN-γ, IL-2, and IL-10 (Fig. 2a–d), indicating a stimulatory effect on both T cells and monocytes. The remaining crude extracts from liquid and solid cultures from bacterial isolates did not induce cytokine release in PBMC culture supernatants.
Fig. 2.
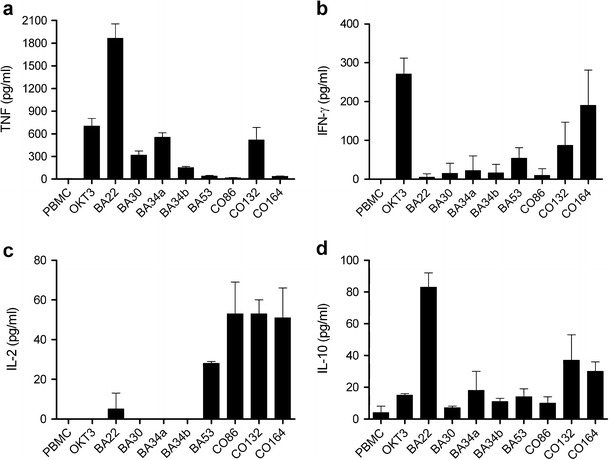
Cytokine responses of precultured human peripheral blood mononuclear cells to crude extracts from bacterial isolates. a TNF, b IFN-γ, c IL-2, and d IL-10. Crude extracts were tested in triplicates at three different concentrations (preparations from liquid cultures—25, 2.5, and 0.25 μg/ml, from solid cultures—10, 1, and 0.1 μg/ml), and the most active one is shown. The isolates A. jenensis strain BA22, A. oxydans strain BA30, A. oxydans strain BA34a, and M. coxensis strain CO164 were grown on M1 agar plates for 7–10 days. Crude extract from isolate A. oxydans strain BA34b was obtained after 4 days of liquid culture, and active extracts from strains Lapillicoccus sp. BA53, S. shandongensis strain CO86 and Sphingobium sp. CO132 were prepared after 7 days of liquid culture
With regard to the induction of proliferation, crude extract prepared from solid culture of strain Sphingobium sp. CO105 exhibited the strongest mitogenic activity, reaching 14% of the positive control (OKT3, activating all T cells; Fig. 3). When strain Sphingobium sp. CO105 was tested for its capacity to induce cytokine release, a weak induction of IL-2 was observed (data not shown), which suggests that T cells are the responsive cell type. The remainder of the samples did not stimulate PBMC proliferation.
Fig. 3.
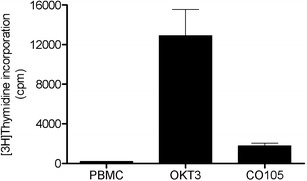
PBMC proliferation, measured as radioactivity incorporated into DNA from tritiated thymidine, in response to stimulation with crude extract from strain Sphingobium sp. CO105. Crude extract was tested in triplicates at a concentration of 65 ng/ml
Sphingomonads are well-known for their production of glycosphingolipids. For example, the glycosphingolipid GSL-1 was found to be produced by a Sphingomonas strain isolated from the sponge Plakortis simplex (Laroche et al. 2007). The immunomodulatory activities of the two sphingomonads presented in this study also do not come as a surprise. Glycosphingolipids are a class of compounds that have been shown to serve as immunomodulatory agents (Long et al. 2007). The substance KRN7000, a synthetic analog of the natural product agelasphin 9b from the sponge Agelas mauritianus, is one such example and is currently in clinical trials for its capacity to stimulate NKT cells (Park et al. 2010). To our knowledge, we present here, for the first time, immunomodulatory activities from actinomycetes.
Conclusions
The cultivation of 79 strains belonging to 20 different Actinomycetales genera as well as seven strains belonging to two Sphingomonadales genera represents considerable diversity of culturable bacteria in 18 Caribbean sponges as shown in this study. The isolation of rare actinomycete genera which have not previously been reported from marine organisms and the identification of seven putatively novel actinomycete and sphingomonad species based on the phylogenetic analysis of their 16S rRNA gene sequence interestingly adds to this caveat. These results further prove that marine sponges still remain a relatively untapped resource for actinomycetes.
Moreover, anti-protease activities against cathepsins B and L, rhodesain, and falcipain-2, as well as immunomodulatory activities specifically the induction of cytokine release by PBMC and induction of cell proliferation, were found to be exhibited by the crude extracts of selected strains. The anti-protease and immunomodulatory activities exhibited by these strains further warrant the need for a bioassay-guided purification and identification of the bioactive secondary metabolites. Furthermore, it will be necessary to identify the responsive immune cell types and to obtain a more detailed mechanistic understanding of the processes underlying their modulation.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
16S rRNA gene based phylogenetic affiliation of actinomycete isolates (DOC 103 kb)
16S rRNA gene based phylogenetic affiliation of sphingomonad isolates (DOC 30.0 kb)
Acknowledgments
We gratefully acknowledge H. Angermeier (U. Würzburg) and J. Pawlik (U. North Carolina, Wilmington) for sponge collection from the Bahamas, C. Heindl, S. Berr, and C. Gernert for technical assistance in the laboratory and P. Römer for the introduction into the immunological techniques (U. Würzburg). We furthermore thank S. Zea (INVEMAR, Santa Marta, Colombia) for help in sponge identification as well as L. Veloza and L.E. Ramírez (Universidad Tecnológica de Pereira, Colombia) for providing the laboratory conditions for the cultivation of the Colombian strains. Financial support was provided by a doctoral fellowship (Graduate College “Immunomodulation GK 520” funded by the DFG) to P.T. and the DFG (SFB 630 TP A5) to U.H.
References
- Abdelmohsen UR, Pimentel-Elardo SM, Hanora A, Radwan M, Abou-El-Ela SH, Ahmed S, Hentschel U. Isolation, phylogenetic analysis and anti-infective activity screening of marine sponge-associated actinomycetes. Mar Drugs. 2010;8:399–412. doi: 10.3390/md8030399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anand K, Ziebuhr J, Wadhwani P, Mesters JR, Hilgenfeld R. Coronavirus main proteinase (3CLpro) structure: basis for design of anti-SARS drugs. Science. 2003;300:1763–1767. doi: 10.1126/science.1085658. [DOI] [PubMed] [Google Scholar]
- Ashelford KE, Chuzhanova NA, Fry JC, Jones AJ, Weightman AJ. At least 1 in 20 16S rRNA sequence records currently held in public repositories is estimated to contain substantial anomalies. Appl Environ Microbiol. 2005;71:7724–7736. doi: 10.1128/AEM.71.12.7724-7736.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azuma I, Seya T. Development of immunoadjuvants for immunotherapy of cancer. Int Immunopharmacol. 2001;1:1249–1259. doi: 10.1016/S1567-5769(01)00055-8. [DOI] [PubMed] [Google Scholar]
- Breuning A, Degel B, Schulz F, Buchold C, Stempka M, Machon U, Heppner S, Gelhaus C, Leippe M, Leyh M, Kisker C, Rath J, Stich A, Gut J, Rosenthal PJ, Schmuck C, Schirmeister T. Michael acceptor based antiplasmodial and antitrypanosomal cysteine protease inhibitors with unusual amino acids. J Med Chem. 2010;53:1951–1963. doi: 10.1021/jm900946n. [DOI] [PubMed] [Google Scholar]
- Buchanan GO, Williams PG, Feling RH, Kauffman CA, Jensen PR, Fenical W. Sporolides A and B: structurally unprecedented halogenated macrolides from the marine actinomycete Salinispora tropica. Org Lett. 2005;7:2731–2734. doi: 10.1021/ol050901i. [DOI] [PubMed] [Google Scholar]
- Calkins CC, Sloane BF. Mammalian cysteine protease inhibitors: biochemical properties and possible roles in tumor progression. Biol Chem Hoppe Seyler. 1995;376:71–80. [PubMed] [Google Scholar]
- Caspi RR. Immunotherapy of autoimmunity and cancer: the penalty for success. Nat Rev Immunol. 2008;8:970–976. doi: 10.1038/nri2438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Challis GL. Mining microbial genomes for new natural products and biosynthetic pathways. Microbiology. 2008;154:1555–1569. doi: 10.1099/mic.0.2008/018523-0. [DOI] [PubMed] [Google Scholar]
- Danner SA, Carr A, Leonard JM, Lehman LM, Gudiol F, Gonzales J, Raventos A, Rubio R, Bouza E, Pintado V, Aguado AG, Delomas JG, Delgado R, Borleffs JCC, Hsu A, Valdes JM, Boucher CAB, Cooper DA, Gimeno C, Clotet B, Tor J, Ferrer E, Martinez PL, Moreno S, Zancada G, Alcami J, Noriega AR, Pulido F, Glassman HN. A short-term study of the safety, pharmacokinetics, and efficacy of Ritonavir, an inhibitor of Hiv-1 protease. New Engl J Med. 1995;333:1528–1533. doi: 10.1056/NEJM199512073332303. [DOI] [PubMed] [Google Scholar]
- Dieckmann R, Graeber I, Kaesler I, Szewzyk U, von Döhren H. Rapid screening and dereplication of bacterial isolates from marine sponges of the Sula Ridge by intact-cell-MALDI-TOF mass spectrometry (ICM-MS) Appl Microbiol Biotechnol. 2005;67:539–548. doi: 10.1007/s00253-004-1812-2. [DOI] [PubMed] [Google Scholar]
- Ersmark K, Del Valle JR, Hanessian S. Chemistry and biology of the aeruginosin family of serine protease inhibitors. Angew Chem Int Edit. 2008;47:1202–1223. doi: 10.1002/anie.200605219. [DOI] [PubMed] [Google Scholar]
- Feling RH, Buchanan GO, Mincer TJ, Kauffman CA, Jensen PR, Fenical W. Salinosporamide A: a highly cytotoxic proteasome inhibitor from a novel microbial source, a marine bacterium of the new genus Salinospora. Angew Chem Int Edit. 2003;42:355–357. doi: 10.1002/anie.200390115. [DOI] [PubMed] [Google Scholar]
- Fenical W, Jensen PR, Palladino MA, Lam KS, Lloyd GK, Potts BC. Discovery and development of the anticancer agent salinosporamide A (NPI-0052) Bioorg Med Chem. 2009;17:2175–2180. doi: 10.1016/j.bmc.2008.10.075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiedler HP, Bruntner C, Bull AT, Ward AC, Goodfellow M, Potterat O, Puder C, Mihm G. Marine actinomycetes as a source of novel secondary metabolites. Antonie van Leeuwenhoek. 2005;87:37–42. doi: 10.1007/s10482-004-6538-8. [DOI] [PubMed] [Google Scholar]
- Gulder TA, Moore BS. Chasing the treasures of the sea—bacterial marine natural products. Curr Opin Microbiol. 2009;12:252–260. doi: 10.1016/j.mib.2009.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heindl H, Wiese J, Thiel V, Imhoff JF. Phylogenetic diversity and antimicrobial activities of bryozoan-associated bacteria isolated from Mediterranean and Baltic Sea habitats. Syst Appl Microbiol. 2010;33:94–104. doi: 10.1016/j.syapm.2009.12.002. [DOI] [PubMed] [Google Scholar]
- Hentschel U, Schmid M, Wagner M, Fieseler L, Gernert C, Hacker J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol Ecol. 2001;35:305–312. doi: 10.1111/j.1574-6941.2001.tb00816.x. [DOI] [PubMed] [Google Scholar]
- Hentschel U, Hopke J, Horn M, Friedrich AB, Wagner M, Hacker J, Moore BS. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl Environ Microbiol. 2002;68:4431–4440. doi: 10.1128/AEM.68.9.4431-4440.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hentschel U, Usher KM, Taylor MW. Marine sponges as microbial fermenters. FEMS Microbiol Ecol. 2006;55:167–177. doi: 10.1111/j.1574-6941.2005.00046.x. [DOI] [PubMed] [Google Scholar]
- Hozumi M, Ogawa M, Sugimura T, Takeuchi T, Umezawa H. Inhibition of tumorigenesis in mouse skin by leupeptin, a protease inhibitor from actinomycetes. Cancer Res. 1972;32:1725–1728. [PubMed] [Google Scholar]
- Jensen PR, Mincer TJ, Williams PG, Fenical W. Marine actinomycete diversity and natural product discovery. Antonie van Leeuwenhoek. 2005;87:43–48. doi: 10.1007/s10482-004-6540-1. [DOI] [PubMed] [Google Scholar]
- Jiang S, Li X, Zhang L, Sun W, Dai S, Xie L, Liu Y, Lee K. Culturable actinobacteria isolated from marine Sponge Iotrochota sp. Mar Biol. 2008;153:945–952. doi: 10.1007/s00227-007-0866-y. [DOI] [Google Scholar]
- Kaeppler U, Stiefl N, Schiller M, Vicik R, Breuning A, Schmitz W, Rupprecht D, Schmuck C, Baumann K, Ziebuhr J, Schirmeister T. A new lead for nonpeptidic active-site-directed inhibitors of the severe acute respiratory syndrome coronavirus main protease discovered by a combination of screening and docking methods. J Med Chem. 2005;48:6832–6842. doi: 10.1021/jm0501782. [DOI] [PubMed] [Google Scholar]
- Kim TK, Garson MJ, Fuerst JA. Marine actinomycetes related to the ‘Salinospora’ group from the Great Barrier Reef sponge Pseudoceratina clavata. Environ Microbiol. 2005;7:509–518. doi: 10.1111/j.1462-2920.2005.00716.x. [DOI] [PubMed] [Google Scholar]
- Klinman DM, Klaschik S, Tomaru K, Shirota H, Tross D, Ikeuchi H. Immunostimulatory CpG oligonucleotides: effect on gene expression and utility as vaccine adjuvants. Vaccine. 2010;28:1919–1923. doi: 10.1016/j.vaccine.2009.10.094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon HC, Kauffman CA, Jensen PR, Fenical W. Marinomycins A–D, antitumor-antibiotics of a new structure class from a marine actinomycete of the recently discovered genus “Marinispora”. J Am Chem Soc. 2006;128:1622–1632. doi: 10.1021/ja0558948. [DOI] [PubMed] [Google Scholar]
- Lam KS. Discovery of novel metabolites from marine actinomycetes. Curr Opin Microbiol. 2006;9:245–251. doi: 10.1016/j.mib.2006.03.004. [DOI] [PubMed] [Google Scholar]
- Lane DJ. 16S/23S rRNA sequencing. Chichester: Wiley; 1991. [Google Scholar]
- Laroche M, Imperatore C, Grozdanov L, Costantino V, Mangoni A, Hentschel U, Fattorusso E. Cellular localisation of secondary metabolites isolated from the Caribbean sponge Plakortis simplex. Mar Biol. 2007;151:1365–1373. doi: 10.1007/s00227-006-0572-1. [DOI] [Google Scholar]
- Long XT, Deng S, Mattner JC, Zang Z, Zhou D, Mcnary N, Goff RD, Teyton L, Bendelac A, Savage PB. Synthesis and evaluation of stimulatory properties of Sphingomonadaceae glycolipids. Nat Chem Biol. 2007;3:559–564. doi: 10.1038/nchembio.2007.19. [DOI] [PubMed] [Google Scholar]
- Ludewig S, Kossner M, Schiller M, Baumann K, Schirmeister T. Enzyme kinetics and hit validation in fluorimetric protease assays. Curr Top Med Chem. 2010;10:368–382. doi: 10.2174/156802610790725498. [DOI] [PubMed] [Google Scholar]
- Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar, Buchner A, Lai T, Steppi S, Jobb G, Forster W, Brettske I, Gerber S, Ginhart AW, Gross O, Grumann S, Hermann S, Jost R, Konig A, Liss T, Lussmann R, May M, Nonhoff B, Reichel B, Strehlow R, Stamatakis A, Stuckmann N, Vilbig A, Lenke M, Ludwig T, Bode A, Schleifer KH. ARB: a software environment for sequence data. Nucleic Acids Res. 2004;32:1363–1371. doi: 10.1093/nar/gkh293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magarvey NA, Keller JM, Bernan V, Dworkin M, Sherman DH. Isolation and characterization of novel marine-derived actinomycete taxa rich in bioactive metabolites. Appl Environ Microbiol. 2004;70:7520–7529. doi: 10.1128/AEM.70.12.7520-7529.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maldonado LA, Stach JEM, Pathom-aree W, Ward AC, Bull AT, Goodfellow M. Diversity of cultivable Actinobacteria in geographically widespread marine sediments. Antonie van Leeuwenhoek. 2005;87:11–18. doi: 10.1007/s10482-004-6525-0. [DOI] [PubMed] [Google Scholar]
- Mincer TJ, Jensen PR, Kauffman CA, Fenical W. Widespread and persistent populations of a major new marine actinomycete taxon in ocean sediments. Appl Environ Microbiol. 2002;68:5005–5011. doi: 10.1128/AEM.68.10.5005-5011.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montalvo NF, Mohamed NM, Enticknap JJ, Hill RT. Novel actinobacteria from marine sponges. Antonie van Leeuwenhoek. 2005;87:29–36. doi: 10.1007/s10482-004-6536-x. [DOI] [PubMed] [Google Scholar]
- Muyzer G, Waal ECD, Uitterlinden AG. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol. 1993;59:695–700. doi: 10.1128/aem.59.3.695-700.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakao Y, Fujita M, Warabi K, Matsunaga S, Fusetani N. Bioactive marine metabolites. Part 104. Miraziridine A, a novel cysteine protease inhibitor from the marine sponge Theonella aff. mirabilis. J Am Chem Soc. 2000;122:10462–10463. doi: 10.1021/ja001859j. [DOI] [Google Scholar]
- Park JJ, Lee JH, Seo KC, Bricard G, Venkataswamy MM, Porcelli SA, Chung SK. Syntheses and biological activities of KRN7000 analogues having aromatic residues in the acyl and backbone chains with varying stereochemistry. Bioorg Med Chem Lett. 2010;20:814–818. doi: 10.1016/j.bmcl.2009.12.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pimentel-Elardo S, Gulder TAM, Hentschel U, Bringmann G. Cebulactams A1 and A2, new macrolactams isolated from Saccharopolyspora cebuensis, the first obligate-marine strain of the genus Saccharopolyspora. Tetrahedron Lett. 2008;49:6889–6892. doi: 10.1016/j.tetlet.2008.09.094. [DOI] [Google Scholar]
- Pimentel-Elardo SM, Kozytska S, Bugni TS, Ireland CM, Moll H, Hentschel U. Anti-parasitic compounds from Streptomyces sp. strains isolated from Mediterranean sponges. Mar Drugs. 2010;8:373–380. doi: 10.3390/md8020373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ratia K, Saikatendu KS, Santarsiero BD, Barretto N, Baker SC, Stevens RC, Mesecar AD. Severe acute respiratory syndrome coronavirus papain-like protease: structure of a viral deubiquitinating enzyme. Proc Natl Acad Sci USA. 2006;103:5717–5722. doi: 10.1073/pnas.0510851103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rawlings ND, Barrett AJ, Bateman A. MEROPS: the peptidase database. Nucleic Acids Res. 2010;38:D227–D233. doi: 10.1093/nar/gkp971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romanenko LA, Uchino M, Frolova GM, Tanaka N, Kalinovskaya NI, Latyshev N, Mikhailov VV. Sphingomonas molluscorum sp. nov., a novel marine isolate with antimicrobial activity. Int J Syst Evol Microbiol. 2007;57:358–363. doi: 10.1099/ijs.0.64441-0. [DOI] [PubMed] [Google Scholar]
- Romanenko LA, Tanaka N, Frolova GM, Mikhailov VV (2009) Arenicella xantha gen. nov., sp. nov., a novel Gammaproteobacterium isolated from a marine sandy sediment from the Sea of Japan. Int J Syst Evol Microbiol. doi: 10.1099/ijs.0.017194-0 [DOI] [PubMed]
- Santavy DL, Willenz P, Colwell RR. Phenotypic study of bacteria associated with the Caribbean sclerosponge, Ceratoporella nicholsoni. Appl Environ Microbiol. 1990;56:1750–1762. doi: 10.1128/aem.56.6.1750-1762.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneemann I, Nagel K, Kajahn I, Labes A, Wiese J, Imhoff JF. Comprehensive investigation of marine Actinobacteria associated with the sponge Halichondria panicea. Appl Environ Microbiol. 2010;76:3702–3714. doi: 10.1128/AEM.00780-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirling E, Gottlieb D. Methods for characterization of Streptomyces species. Int J Syst Bacteriol. 1966;16:317–327. doi: 10.1099/00207713-16-3-313. [DOI] [Google Scholar]
- Stach JE, Bull AT. Estimating and comparing the diversity of marine actinobacteria. Antonie van Leeuwenhoek. 2005;87:3–9. doi: 10.1007/s10482-004-6524-1. [DOI] [PubMed] [Google Scholar]
- Stackebrandt E, Ebers J. Taxonomic parameters revisited: tarnished gold standards. Microbiol Today. 2006;33:152–155. [Google Scholar]
- Sun W, Dai S, Jiang S, Wang G, Liu G, Wu H, Li X. Culture-dependent and culture-independent diversity of Actinobacteria associated with the marine sponge Hymeniacidon perleve from the South China Sea. Antonie van Leeuwenhoek. 2010;98:65–75. doi: 10.1007/s10482-010-9430-8. [DOI] [PubMed] [Google Scholar]
- Taylor MW, Radax R, Steger D, Wagner M. Sponge-associated microorganisms: evolution, ecology, and biotechnological potential. Microbiol Mol Biol Rev. 2007;71:295–347. doi: 10.1128/MMBR.00040-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turk B. Targeting proteases: successes, failures and future prospects. Nat Rev Drug Discov. 2006;5:785–799. doi: 10.1038/nrd2092. [DOI] [PubMed] [Google Scholar]
- Vicik R, Busemann M, Gelhaus C, Stiefl N, Scheiber J, Schmitz W, Schulz F, Mladenovic M, Engels B, Leippe M, Baumann K, Schirmeister T. Aziridide-based inhibitors of cathepsin L: synthesis, inhibition activity, and docking studies. ChemMedChem. 2006;1:1126–1141. doi: 10.1002/cmdc.200600106. [DOI] [PubMed] [Google Scholar]
- Vicik R, Hoerr V, Glaser M, Schultheis M, Hansell E, Mckerrow JH, Holzgrabe U, Caffrey CR, Ponte-Sucre A, Moll H, Stich A, Schirmeister T. Aziridine-2, 3-dicarboxylate inhibitors targeting the major cysteine protease of Trypanosoma brucei as lead trypanocidal agents. Bioorg Med Chem Lett. 2006;16:2753–2757. doi: 10.1016/j.bmcl.2006.02.026. [DOI] [PubMed] [Google Scholar]
- Wang Q, Garrity GM, Tiedje JM, Cole JR. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol. 2007;73:5261–5267. doi: 10.1128/AEM.00062-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster NS, Wilson KJ, Blackall LL, Hill RT. Phylogenetic diversity of bacteria associated with the marine sponge Rhopaloeides odorabile. Appl Environ Microbiol. 2001;67:434–444. doi: 10.1128/AEM.67.1.434-444.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO . World malaria report 2009. Geneva: World Health Organization; 2009. [Google Scholar]
- Zhang HT, Zhang W, Jin Y, Jin MF, Yu XJ. A comparative study on the phylogenetic diversity of culturable Actinobacteria isolated from five marine sponge species. Antonie van Leeuwenhoek. 2008;93:241–248. doi: 10.1007/s10482-007-9196-9. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
16S rRNA gene based phylogenetic affiliation of actinomycete isolates (DOC 103 kb)
16S rRNA gene based phylogenetic affiliation of sphingomonad isolates (DOC 30.0 kb)