

Abstract
Canine parvovirus type 2 (CPV-2) can cause acute haemorrhagic enteritis in dogs and myocarditis in puppies. This disease has become one of the most serious infectious diseases of dogs. During 2014 in China, there were many cases of acute infectious diarrhoea in dogs. Some faecal samples were negative for the CPV-2 antigen based on a colloidal gold test strip but were positive based on PCR, and a viral strain was isolated from one such sample. The cytopathic effect on susceptible cells and the results of the immunoperoxidase monolayer assay, PCR, and sequencing indicated that the pathogen was CPV-2. The strain was named CPV-NY-14, and the full-length genome was sequenced and analysed. A maximum likelihood tree was constructed using the full-length genome and all available CPV-2 genomes. New strains have replaced the original strain in Taiwan and Italy, although the CPV-2a strain is still predominant there. However, CPV-2a still causes many cases of acute infectious diarrhoea in dogs in China.
Keywords: CPV-2, Phylogenetic analysis, Viral evolution
Introduction
Canine parvovirus type 2 (CPV-2) belongs to the genus Protoparvovirus and the Parvoviridae family and was first observed by electron microscopy in 1977 [1]. CPV-2 has three antigenic variants: types 2a, 2b, and 2c. Two antigenic variants, CPV-2a and CPV-2b, are now distributed worldwide [2]. A third CPV-2 variant, which was initially believed to be a Glu-426 mutant and subsequently renamed CPV-2c, was detected in Italy in 2000 [3] and is now circulating there together with types 2a and 2b [4–6]. The new type 2c has also been reported in Vietnam by Nakamura et al. [7], who developed monoclonal antibodies specific for that type [7]. Type 2c has also been reported in the United States and South America [8].
The genome of CPV-2 is a single-stranded DNA molecule that contains two open reading frames (ORF1 and ORF2). ORF1 is located in the 5′ region of the genome and encodes non-structural protein 1 (NS1) and 2 (NS2). ORF2 is located in the right part of the genome and encodes the structural proteins VP1 and VP2. The middle region of the genome also contains a 500 bp sequence [9, 10]. The non-structural proteins NS1 and NS2 are associated with viral replication. NS1 functions as a nickase and helicase and can form a covalent bond at the 5′ end of DNA. Structural proteins VP1 and VP2 compose the nucleocapsid of CPV-2 [11].
In 2014 in China, many acute cases of infectious diarrhoea were reported in dogs. To identify the pathogen, we collected and examined faecal samples from symptomatic dogs. We report here that the disease is associated with a strain of parvovirus, and phylogenetic characterization of the causal isolate showed that isolates from China have long branch lengths; this may indicate an extensive process of accumulation of mutations and substitutions in Chinese lineages. This study provides the basis for further exploration of the CPV-2 variation, the selection of vaccines, and the effective prevention and control of CPV-2 infections.
Methods
Clinical samples
Faeces were collected from dogs exhibiting acute haemorrhagic enteritis. The faecal samples (10 g) were immersed in 10 mL of sterile phosphate-buffered saline (PBS) with antibiotics, and the suspension was centrifuged at 10,000×g for 15 min. The supernatant was filtered and frozen at −80 °C.
Identification of CPV-2 with the colloidal gold test strip and PCR
To determine whether CPV-2 was present in the samples, the filtered supernatants were added to the colloidal gold test strips using the BioNote Rapid Test Kit (Korea, BioNote). The samples were mixed with buffer and then tested for the antigen at room temperature according to the manufacturer’s instructions. The negative sample was prepared for PCR as follows.
The PCR assay was performed in a 20-μL reaction volume that included 2.4 μL of template, 1.8 μL each of VP2-F and VP2-R (Table 1), 2.0 μL of dNTPs, 1.0 μL of KOD FX Neo polymerase (1 U/μL) (TOYOBO Biotechnology Company, Shanghai, China), 5.0 μL of 2 × PCR buffer for KOD FX Neo (TOYOBO Biotechnology Company, Shanghai, China), and sufficient ddH2O to increase the volume to 20 μL. Amplification was carried out in a pre-heated thermocycler (Applied Biosystems 2720 Thermal Cycler) as follows: one cycle at 94 °C for 5 min; followed by 35 cycles at 94 °C for 45 s, 50 °C for 60 s, and 72 °C for 60 s; and a final extension at 72 °C for 10 min. Amplicons were detected by electrophoresing 10-μL aliquots in 2 % agarose gels in 1 × TAE [40 mM Tris–acetate (pH 8.0), 1 mM EDTA].
Table 1.
Sequences of primers used to amplify CPV-2
| Primer designation | Primer sequence (5′-3′) | Position | Fragment size in bp |
|---|---|---|---|
| NS1-F1 | ACCGTTACTGACATTCGCTTC | 781–802 | 2111 |
| NS1-R1 | CCTTACCTCTCCTGGCTC | 2875–2891 | |
| VP2-F2 | GCCGGTGCAGGACAAGTA | 3317–3335 | 2011 |
| VP2-R2 | CAACCCACACCATAACAACA | 5038–5327 | |
| CoF13 | TTTATGTTTTATTACAATTTATTTTAAGATTAG | 1–33 | 5783 |
| CoR13 | CAAACCAACCAAACCACCCA | 5765–5783 |
Identification of CPV-2 with cell culture and IPMA
Feline kidney cell line F81 was obtained from the American Type Culture Collection, USA and was used to isolate viruses from clinical samples and to observe cytopathic effects associated with viral replication. The treated samples were inoculated at a concentration of 1 mL per 25 cm2 flask in a cell monolayer. After adsorbing for 1 h at 37 °C, the inoculum was removed, DMEM with 2 % foetal bovine serum was added, and the cells were again incubated at 37 °C. Cell cultures were observed daily for 4–5 days to monitor the appearance of cytopathic effects (CPE). Subsequently, an IPMA was conducted, and the other flask was frozen at −20 °C and submitted to further passages following the same procedure after freezing and thawing for three times, until the eventual appearance of CPE [3, 12].
Cloning the full-length genomic sequence of CPV-2
DNA and RNA were extracted from the homogenized samples (faeces from infected dogs) with the TIANamp Virus DNA/RNA Kit (Beijing TIANGEN Biotech Company, Beijing, China) according to the manufacturer’s protocol. DNA and RNA were analysed by PCR or RT-PCR, and the presence of other potential pathogens, such as canine distemper virus (CDV), canine adenovirus (CAV), canine coronavirus (CCV), and canine rotavirus (CRV), was investigated.
Table 1 summarizes the sequences of primers used to amplify the virus, and the PCR was conducted as previously described [13–15].
DNA fragments were then cloned into the pMD18-T sample vector (TaKaRa Biotechnology Co., Ltd. Japan). The same set of PCR primers was used to sequence the full-length genome of the isolate NY-14. Sequencing reactions were performed by a commercial corporation (Huada, Beijing).
Sequence alignment and phylogenetic analyses
Sequences were aligned using Clustal X software version 2.1 [16]. After the first alignment, the sequences were manually edited to maintain the reading frames using the SE-Al programme version 2.0 (http://evolve.zoo.ox.ac.uk/software/). The following CPV-2 references were used for the phylogenetic analyses: M23255; M24000; DQ340404; EU659117; DQ340411; M24003; EU659120; DQ340419; DQ340430; DQ340426; M24004; M24001; EU659116; DQ340410; EU659118; DQ340408; DQ340429; DQ340405; EU659119; M74849; DQ340416; DQ340417; DQ340432; EU659121; DQ340413; DQ340431; DQ340409; DQ340421; DQ340418; DQ340425; DQ340433; DQ340412; DQ340427; DQ340434; U22896; DQ340422; DQ340428; DQ340423; DQ340415; M74852; DQ340424; DQ340414; DQ340420; DQ340407; DQ340406; D26079; JN033694; EU310373; JN867618; JN867617; JN867616; JN867615; JN867614; JN867613; JN867612; JN867611; JN867610; EF011664; M19296; KF638400; JX660690; JQ268284; JQ268283; EF988660.
Maximum likelihood trees and bootstrap values were obtained using PhyML software [17]. The KHY model and gamma distribution (Γ) was selected according to the likelihood ratio test (LRT) using jModelTest software [18, 19]. The resulting trees were visualized and edited using FigTree (http://tree.bio.ed.ac.uk/software/figtree/).
Recombination analyses
To determine the extent of recombination in the CPV-2 sequences, RDP v.4 software [20] was used for analyses, which utilizes a collection of methods. An excellent and detailed explanation of each method implemented in the RDP programme can be found in the user’s manual (http://darwin.uvigo.es/rdp/rdp.html).
Results
Identification of CPV-2 with the colloidal gold test strip or PCR
The ill dogs showed acute infectious diarrhoea, and the faecal samples were bloody stools (Fig. 1a). The colloidal gold test strip showed positive (Fig. 1b1) and negative (Fig. 1b2) results, while the PCR results were both positive (Fig. 1c).
Fig. 1.
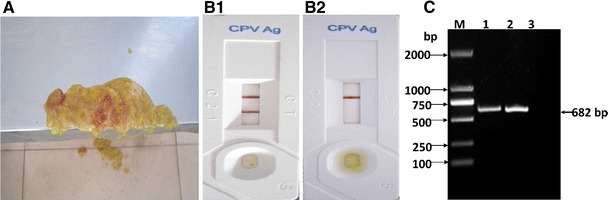
The samples and the colloidal gold test strip and PCR assay. a The haemorrhagic enteritis faecal sample, b Detection of the CPV-2 antigen by colloidal gold test strip. 1 Positive, 2 negative. c Detection of CPV-2 in samples by PCR. M DL2000 DNA marker; 1 sample NY130615; 2 positive control; 3 negative control
Morphological changes in CPV-2-infected cells
One viral strain was isolated from the faeces of dogs that tested negative in the rapid test but positive using PCR. CPE was observed in F81 cells after three passages. CPV-2-infected cell cultures showed vast regions of cell detachment (Fig. 2). The IPMA was conducted with positive sera (Fig. 3).
Fig. 2.
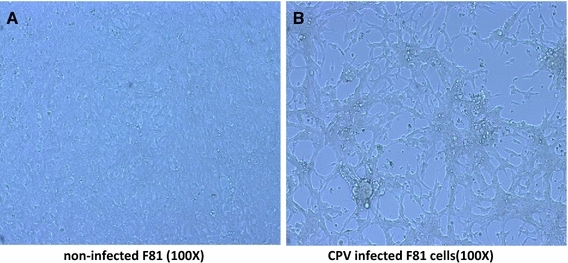
The CPE of CPV-NY-14 cultured in F81 cells. a Non-infected F81 cells (×100); b virus-infected F81 cells (×100)
Fig. 3.
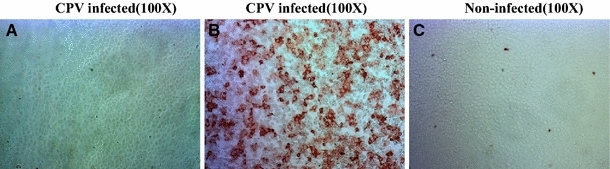
IPMA results detected 24 h after inoculation of CPV-NY-14 in F81 cells (×100) using different sera. a Rabbit negative serum, b rabbit positive serum, c VP2 immune positive serum
Molecular characterization of CPV-2
PCR amplification showed a fragment of 1755 bp corresponding to the VP2 gene of CPV-2. PCR amplification detected the presence of CPV-2 but was negative for other pathogens in the samples analysed. The sequencing results showed that the VP2 gene of CPV-NY-14 is 1755 bp long with no insertions or deletions in the coding region. The results were once again positive for CPV-2. Three experimental animals were inoculated with NY-14 and exhibited clinical symptoms at 14 days post inoculation. CPV-2 was re-isolated from the faeces of the experimental dogs. The whole genome was sequenced, and the CPV-NY-14 complete genome structure was analysed and is shown in Fig. 4.
Fig. 4.
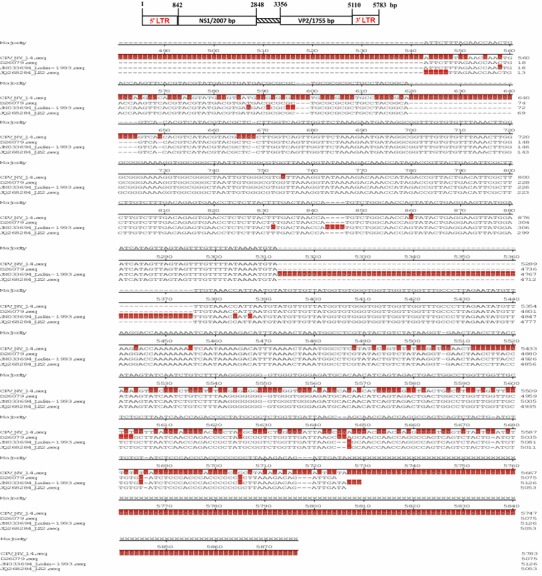
Genomic map of the isolate NY-14
Phylogenetic analysis
A maximum likelihood tree was constructed using the VP2 sequence and full-length genomes of CPV-NY-14 and all available CPV-2 genomes. Figure 4 shows that all the CPV-2 sequences from China clustered in a group, and the CPV-NY-14 isolate is also located in the Chinese clade. Interestingly, located at the base of the Chinese clade is one isolate from Russia (JN033694) sampled in 1993. All sequences used to construct the above-mentioned tree were absent for recombination.
To further examine CPV-2 isolates, we used a larger dataset composed only of VP2 protein sequences. The maximum likelihood tree again showed a monophyletic cluster containing all CPV-2 isolates from China. CPV-NY-14 belongs to the CPV-2a type, and it was inferred that CPV-2a is still circulating in China and is also the main agent of acute haemorrhagic enteritis in dogs.
In addition, both trees showed that isolates from China have long branch lengths; this may indicate an extensive process of accumulation of mutations and substitutions in Chinese lineages.
Discussion
CPV-2 is an acute infectious disease in dogs, which causes acute haemorrhagic enteritis. There are three antigenic variants (2a, 2b, and 2c) of CPV-2 that are distributed worldwide with different frequencies. CPV-2 has two clinical phenotypes: haemorrhagic enteritis and myocarditis. Both are characterized by high mortality and high infectivity. It has become one of the major infections of dogs, causing serious and continuous economic losses.
Since it emerged in 1978, canine parvovirus has spread in the domestic and wild canine population, where it is continuously evolving into new adaptive viral variants. The variability and the intrinsically high mutation rate of the CPV-2 genome allowed the diversity of CPV-2 to rapidly increase as the virus spread through canine populations [21]. Because some viral variants replicate more successfully than others, the virus population changes over time [22]. CPV-2a and CPV-2b are the best examples of variants with a fast spread and replacement capacity since the emergence of CPV-2 [9, 23]. During 2014, there were many cases of acute infectious diarrhoea in dogs. Some samples tested negative for the CPV-2 antigen by a colloidal gold test strip, while they were positive for CPV-2 by PCR. One viral strain was isolated from the faeces of dogs that tested negative in the colloidal gold test strip but positive with PCR. CPV-NY-14, which was described in this study, was found in dogs that exhibited acute haemorrhagic enteritis. Based on the PCR results, the only virus consistently detected in the samples was CPV-2.
Some samples tested with the serology method (colloidal gold test strip) failed to detect the virus, while PCR was able to detect CPV-2, which is a valid and important objective. This may be due to differences in the test sensitivity (PCR is more sensitive than the serological test) or due to a change in the viral genome that leads to a major change in the type of the antigen (viral protein) and the failure of the antibodies to recognize the viral antigen in the serology test. This question needs to be addressed by future experiments.
The isolated virus was then cultivated in F81 cells, and we next inoculated adult dogs with the isolated CPV-2. These animals presented clinical conditions characteristic of CPV-2 infection. PCR was used to demonstrate that CPV-2 was the agent causing the disease in these experimental animals.
CPV-2 is pandemic, and the frequencies of the different antigenic types of CPV-2 vary in different countries. In Uruguay, for example, CPV-2c is the major epidemic strain [24]. In the USA and southern Africa, CPV-2b is the main viral type that causes most outbreaks of CPV-2 infection [23, 25]. In the UK, both CPV-2a and CPV-2b are present, and Germany and Spain have similar frequencies of isolation [26, 27]. In India, the major epidemic strains are CPV-2a and CPV-2c. The new strains have also replaced the original one in Taiwan and Italy, although the CPV-2a strain is still predominant there [28, 29]. However, CPV-2 still causes many cases of acute infectious diarrhoea in dogs in Japan [30].
In this study, molecular phylogenetic analysis of CPV-NY-14 and other CPV-2 isolates in GenBank revealed that CPV-NY-14 is closely related to isolates s5, SC02-11, LZ1, LZ2, and nj01-06, especially isolate LZ2. CPV-2a still causes many cases of acute infectious diarrhoea in dogs in China. The sequence analysis showed that there is little variation, and this is important in choosing a vaccine to prevent this disease. Given the results obtained in the current study, the continuous surveillance of CPV-2 in China is imperative for determining whether CPV-2a will colonize and spread into new territories.
Acknowledgments
This work was partly supported by the Agricultural Science and Technology Innovation Programme of China (ASTIP-IAS15).
Compliance with ethical standards
Conflict of interest
The authors declare that they have no competing interests.
Footnotes
Ling Zhou, Qinghai Tang, Lijun Shi, and Miaomiao Kong have contributed equally to this work.
Contributor Information
Kai Zhao, Phone: +86 531 82767361, Email: zybin395@126.com.
Shangjin Cui, Phone: +86 0 18518437100, Email: cuishangjin@126.com.
References
- 1.Eugster A. Diarrhea in puppies: parvovirus-like particles demonstrated in their feces. Southwest. Vet. 1977;30(1):59–60. [Google Scholar]
- 2.Truyen U. J. Vet. Microbiol. 2006;117(1):9–13. doi: 10.1016/j.vetmic.2006.04.003. [DOI] [PubMed] [Google Scholar]
- 3.Buonavoglia C, Martella V, Pratelli A, Tempesta M, Cavalli A, Buonavoglia D, Bozzo G, Elia G, Decaro N, Carmichael L. J. Gen. Virol. 2001;82(12):3021–3025. doi: 10.1099/0022-1317-82-12-3021. [DOI] [PubMed] [Google Scholar]
- 4.Martella V, Cavalli A, Pratelli A, Bozzo G, Camero M, Buonavoglia D, Narcisi D, Tempesta M, Buonavoglia C. J. Clin. Microbiol. 2004;42(3):1333–1336. doi: 10.1128/JCM.42.3.1333-1336.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Decaro N, Elia G, Campolo M, Desario C, Lucente MS, Bellacicco AL, Buonavoglia C. J. Vet. Med. B. Infect. Dis. Vet. Public Health. 2005;52(7–8):316–319. doi: 10.1111/j.1439-0450.2005.00869.x. [DOI] [PubMed] [Google Scholar]
- 6.Decaro N, Elia G, Martella V, Campolo M, Desario C, Camero M, Cirone F, Lorusso E, Lucente MS, Narcisi D, Scalia P, Buonavoglia C. J. Virol. Methods. 2006;133(1):92–99. doi: 10.1016/j.jviromet.2005.10.026. [DOI] [PubMed] [Google Scholar]
- 7.Nakamura M, Tohya Y, Miyazawa T, Mochizuki M, Phung HT, Nguyen NH, Huynh LM, Nguyen LT, Nguyen PN, Nguyen PV, Nguyen NP, Akashi H. Arch. Virol. 2004;149(11):2261. doi: 10.1007/s00705-004-0367-y. [DOI] [PubMed] [Google Scholar]
- 8.Pérez R, Francia L, Romero V, Maya L, López I, Hernández M. Vet. Microbiol. 2007;124(1–2):147. doi: 10.1016/j.vetmic.2007.04.028. [DOI] [PubMed] [Google Scholar]
- 9.Parrish C, Carmichael L. Virology. 1983;129(2):401–414. doi: 10.1016/0042-6822(83)90179-4. [DOI] [PubMed] [Google Scholar]
- 10.Reed A, Jones E, Miller T. J. Viral. 1988;62(1):266–276. doi: 10.1128/jvi.62.1.266-276.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rhode S, Paradiso P. Parvovirus genome. J. Viral. 1983;45(1):173–184. doi: 10.1128/jvi.45.1.173-184.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Puentes R, Eliopulos N, Pérez R, Franco G, Sosa K, Bianchi P, Furtado A, Hübner SO, Esteves PA. Braz. J. Microbiol. 2012;43(3):1005–1009. doi: 10.1590/S1517-83822012000300022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Calderon MG, Remorini P, Periolo O, Iglesias M, Mattion N, La Torre J. Vet. Microbiol. 2007;125:341–349. doi: 10.1016/j.vetmic.2007.05.020. [DOI] [PubMed] [Google Scholar]
- 14.Chaturvedi U, Tiwari AK, Ratta B, Ravindra PV, Rajawat YS, Palia SK, Rai A. J. Virol. Methods. 2008;149:260–263. doi: 10.1016/j.jviromet.2008.01.024. [DOI] [PubMed] [Google Scholar]
- 15.Pratelli A, Buonavoglia D, Martella V, Tempesta M, Lavazza A, Buonavoglia C. J. Virol. Methods. 2000;84:91–94. doi: 10.1016/S0166-0934(99)00134-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23(21):2947–2948. doi: 10.1093/bioinformatics/btm404. [DOI] [PubMed] [Google Scholar]
- 17.Guindon S, Gascuel O. Syst. Biol. 2003;52(5):696–704. doi: 10.1080/10635150390235520. [DOI] [PubMed] [Google Scholar]
- 18.Posada D. Mol. Biol. Evol. 2008;25(7):1253–1256. doi: 10.1093/molbev/msn083. [DOI] [PubMed] [Google Scholar]
- 19.Posada D, Crandall KA. Bioinformatics. 1998;14(9):817–818. doi: 10.1093/bioinformatics/14.9.817. [DOI] [PubMed] [Google Scholar]
- 20.Martin DP, Williamson C, Posada D. Bioinformatics. 2005;21(2):260–262. doi: 10.1093/bioinformatics/bth490. [DOI] [PubMed] [Google Scholar]
- 21.Shackelton LA, Parrish CR, Truyen U, Holmes EC. Proc. Natl. Acad. Sci. USA. 2005;102:379–384. doi: 10.1073/pnas.0406765102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hoelzer K, Shackelton LA, Parrish CR, Holmes EC. J. Gen. Virol. 2008;89(9):2280–2289. doi: 10.1099/vir.0.2008/002055-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Parrish CR, Aquadro CF, Strassheim ML, Evermann JF, Sgro JY, Mohammed HO. J. Virol. 1991;65(12):6544–6552. doi: 10.1128/jvi.65.12.6544-6552.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Perez R, Calleros L, Marandino A, Sarute N, Iraola G, Grecco S, Blanc H, Vignuzzi M, Isakov O, Shomron N, Carrau L, Hernandez M, Francia L, Sosa K, Tomas G, Panzera Y. PLoS One. 2014;9(11):e111779. doi: 10.1371/journal.pone.0111779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Steinel A, Venter EH, Van Vuuren M, Parrish CR, Truyen U. J. Vet. Res. 1998;65(4):239–242. [PubMed] [Google Scholar]
- 26.Ybáñez RR, Vela C, Cortés E, Simarro I, Casal JI. Vet. Rec. 1995;136(7):174–175. doi: 10.1136/vr.136.7.174. [DOI] [PubMed] [Google Scholar]
- 27.Greenwood NM, Chalmers WS, Baxendale W, Thompson H. Vet. Rec. 1996;138:495–496. doi: 10.1136/vr.138.20.495. [DOI] [PubMed] [Google Scholar]
- 28.Chang W, Chang A, Pan M. Vet. Rec. 1996;138:447. doi: 10.1136/vr.138.18.447. [DOI] [PubMed] [Google Scholar]
- 29.Sagazio P, Tempesta M, Buonavoglia D, Cirone F, Buonavoglia C. J. Virol. Methods. 1998;73(2):197–200. doi: 10.1016/S0166-0934(98)00055-X. [DOI] [PubMed] [Google Scholar]
- 30.Hirasawa T, Yono K, Mikazuki K. J. Vet. Med. Ser. B. 1996;43(9):545–554. doi: 10.1111/j.1439-0450.1996.tb00352.x. [DOI] [PubMed] [Google Scholar]