

Abstract
Despite the absence of conventional lymphatics, there is efficient drainage of both cerebrospinal fluid (CSF) and interstitial fluid (ISF) from the CNS to regional lymph nodes. CSF drains from the subarachnoid space by channels that pass through the cribriform plate of the ethmoid bone to the nasal mucosa and cervical lymph nodes in animals and in humans; antigen presenting cells (APC) migrate along this pathway to lymph nodes. ISF and solutes drain from the brain parenchyma to cervical lymph nodes by a separate route along 100–150 nm wide basement membranes in the walls of cerebral capillaries and arteries. This pathway is too narrow for the migration of APC so it is unlikely that APC traffic directly from brain parenchyma to lymph nodes by this route. We present a model for the pivotal involvement of regional lymph nodes in immunological reactions of the CNS. The role of regional lymph nodes in immune reactions of the CNS in virus infections, the remote influence of the gut microbiota, multiple sclerosis and stroke are discussed. Evidence is presented for the role of cervical lymph nodes in the induction of tolerance and its influence on neuroimmunological reactions. We look to the future by examining how nanoparticle technology will enhance our understanding of CNS-lymph node connections and by reviewing the implications of lymphatic drainage of the brain for diagnosis and therapy of diseases of the CNS ranging from neuroimmunological disorders to dementias. Finally, we review the challenges and opportunities for progress in CNS-lymph node interactions and their involvement in disease processes.
Keywords: Antigen presenting cells, CSF, Interstitial fluid, Nasal lymphatics, Cervical lymph nodes, Dendritic cells, Perivascular drainage, Immune privilege, Multiple sclerosis, Regulatory T and B cells, Tolerance, Stroke
Introduction
In most organs of the body, apart from the eye and central nervous system (CNS), antigens, lymphocytes and antigen presenting cells (APC) drain from the tissues to regional lymph nodes along defined lymphatic channels lined by specialised endothelial cells. There are no similar defined lymphatic channels associated with the eye, brain or spinal cord. Instead, there is a distinct and well-documented lymphatic drainage pathway for the CSF via the nasal mucosa (Weller et al. 2009; Johanson et al. 2008) and a different lymphatic drainage pathway for interstitial fluid and solutes from the brain parenchyma along the walls of capillaries and arteries to regional lymph nodes in the neck (Szentistvanyi et al. 1984; Weller et al. 2009). In contrast to other organs, however, the lymphatic drainage pathway from the brain parenchyma does not allow the traffic of antigen presenting cells to regional lymph nodes (Carare et al. 2008). Tracer studies in animals have shown that the lymphatic drainage pathway from brain parenchyma is along the basement membranes in the walls of capillaries and arteries to cervical lymph nodes (Carare et al. 2008). Although no tracer studies have been performed in humans, amyloid-β (Aβ) and other amyloidogenic proteins are deposited as insoluble fibrils in the basement membranes of capillaries and arteries in the ageing brain and in Alzheimer’s disease as cerebral amyloid angiopathy (Preston et al. 2003), which is, in effect, a protein elimination-failure angiopathy (PEFA) (Weller et al. 2008; Carare et al. 2013). The failure of elimination of certain proteins along the ageing perivascular lymphatic drainage pathways from the brain appears to be a major factor in the accumulation of insoluble amyloid deposits in the brain in a variety of neurodegenerative diseases including Alzheimer’s disease and other dementias (Carare et al. 2013).
The CNS is considered to have immune privilege whereby allografts implanted into the CNS have an extended survival time when compared with other tissues. Allografts are rapidly rejected from the CNS if the same allograft is implanted in the skin or similar peripheral site (Galea et al. 2007; Weller et al. 2010). These experiments suggest that an immune response to an allograft in the brain is a combination of (a) drainage of soluble antigen from the allograft to the cervical lymph nodes (b) a peripheral immune response and (c) the immunoregulatory characteristics of the CNS-resident cells and their microenvironment. These elements appear to be required to finally trigger rejection of the allograft. As the brain is protected from the external environment, no microorganisms or foreign material enter the brain unless they pass through peripheral tissues. In a natural environment, organisms entering the brain would have passed through respiratory or gut epithelium or skin before entering the brain thus inducing a peripheral immune response. In this way, an immune response in the CNS may be triggered by the drainage of antigens along the perivascular lymphatic drainage pathways to lymph nodes that are already sensitised to those antigens (Weller et al. 2010). It is not that the central nervous system lacks lymphatic drainage it is just different from other organs in the body.
The main significance of the lymphatic drainage pathways from the CNS for human disease falls into two main areas: a) Infections and neuroimmunological diseases in which the perivascular lymphatic drainage pathways appear to be the only route for drainage of antigens from the CNS parenchyma to regional lymph nodes; b) Neurodegenerative diseases and dementias. As arteries age and become stiffer, Aβ and a variety of other proteins become entrapped in the perivascular lymphatic drainage pathways and this may impair further drainage of proteins and metabolites inducing loss of homoeostasis of the extracellular environment in the brain. The full implications of protein elimination-failure, angiopathy (PEFA) have yet to be assessed and the therapeutic implications for resolving such failure have yet to be fully appreciated (Carare et al. 2013).
In this review we examine the pathways by which cells and antigens drain from peripheral organs to regional lymph nodes and then we examine in detail the structure and functional aspects of pathways for the drainage of antigens and the trafficking of cells from CNS to regional lymph nodes. The functional aspects of these pathways in relation to virus infections and autoimmune disorders of the CNS are reviewed and we comment upon recent research on the pathophysiology of immune reactions in the brain and spinal cord.
Why is the issue of cell and antigen exit from the CNS to the periphery of importance to a large audience of immunologists, neurologists, pathologists, as well as clinician-investigators from other disciplines?
First, on a very general level, emphasizing the egress of cells and antigens from the brain helps to further challenge the widely held notion that the relative immune privilege of the CNS is based upon a lack of access to secondary lymphoid organs (Engelhardt and Coisne 2011). Indeed, there are no classical lymph vessels in the CNS, but several routes out of the brain for cells and antigens are now well-established (Weller et al. 2010; Laman and Weller 2012); the least well known route is the drainage of interstitial fluid (ISF) and solutes along 100–150 nm wide basement membranes in the walls of capillaries and arteries (Carare et al. 2008) (Fig. 1).
Fig. 1.
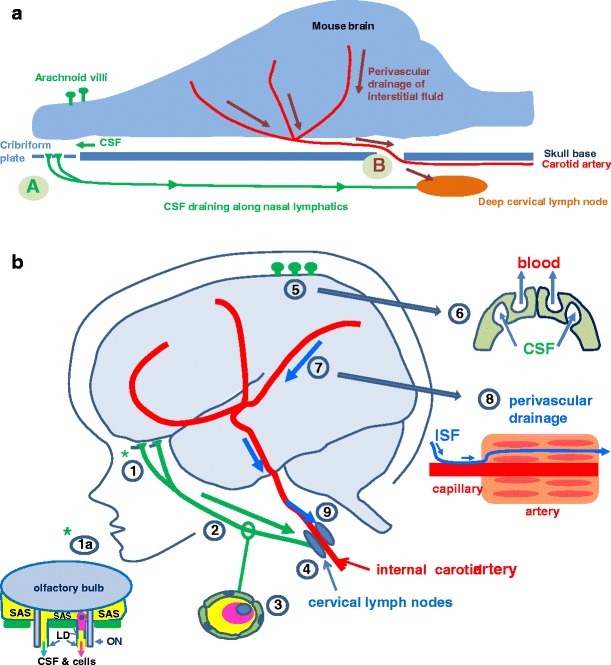
Pathways for the drainage of fluid, solutes and cells from the brain to cervical lymph nodes in rodents and humans. Two of the major fluids associated with the CNS are: (i) CSF in the ventricles and subarachnoid space, and (ii) interstitial fluid (ISF) in the brain parenchyma. CSF and ISF have separate drainage pathways. a Lymphatic drainage of the rodent brain. In rodents, a large proportion of CSF drains via the cribriform plate and nasal mucosa (A) to deep cervical lymph nodes. Arachnoid villi are very small. ISF and solutes drain from brain parenchyma along 100–150 nm-thick basement membranes in the walls of capillaries and arteries to cervical lymph nodes (B). b Lymphatic drainage of the human brain. Lymphatic drainage of CSF. CSF drains from the subarachnoid space through the cribriform plate into nasal lymphatics (1). An enlarged view of the cribriform plate (1a) shows how direct channels for the drainage of CSF from the subarachnoid space (SAS) pass alongside branches of olfactory nerves (ON) into the nasal mucosa. This pathway allows drainage of CSF and antigen presenting cells (see pink cell with blue nucleus in the drainage pathway). (2) Conventional lymphatic vessels carry fluid and cells (3) to cervical lymph nodes (4). CSF also drains into the blood through arachnoid villi and granulations (5) by macromolecular transport (6) through venous endothelial cells. Lymphatic drainage of Interstitial Fluid (ISF). ISF drains from the brain parenchyma along perivascular pathways (7); ISF and solutes diffuse through extracellular spaces of the brain and (8) enter 100–150 nm thick basement membranes in the walls of capillaries and the tunica media of arteries to drain from the brain, depicted as a blue line in (8). Fluid and solutes draining along the walls of cerebral arteries (7) pass into the wall of the internal carotid artery in the neck from which they drain to cervical lymph nodes (9). Modified from (Laman and Weller 2012)
Second, a seminal study by Kraal and coworkers (Wolvers et al. 1999) has demonstrated that cervical lymph nodes (CLN) draining the brain and the oral and nasal mucosa have special properties for driving tolerance. Conversely, surgical removal of CLN reduces disease load and/or lesion load in cryolesion-enhanced EAE (experimental autoimmune encephalomyelitis) in rats (Phillips et al. 1997), in spontaneous EAE in a transgenic model (Furtado et al. 2008), and in relapsing-remitting EAE (van Zwam et al. 2009a). Apparently immune tolerance and pathogenic autoreactivity are differentially regulated in the CLN, by mechanisms that are only partly understood. Cells trafficking from the brain to the CLN may contribute to such mechanisms.
Third, CNS drainage is likely to be critical for neurotropic viral infections, development of multiple sclerosis, and for the immunological consequences of stroke. In addition, therapeutic approaches to CNS diseases such as multiple sclerosis (MS) can potentially target the CLN (de Vos et al. 2002; van Zwam et al. 2009b) and the lumbar lymph nodes draining the spinal cord. Hence, they present an attractive alternative to targeting cell populations hidden behind tissue barriers between the periphery, perivascular spaces, and the brain parenchyma that are difficult to surmount.
Excellent recent reviews related to this article are available, dealing with CNS leukocyte traffic, antigen presenting cell subsets of the CNS, and CNS-lymphoid tissue interactions (Clarkson et al. 2012; Mohammad et al. 2012; Ousman and Kubes 2012; Ransohoff 2012; Prinz et al. 2011; Ransohoff and Engelhardt 2012).
Communications between the CNS and secondary lymphoid organs
There are two major fluids associated with the CNS that are potential media for the transport of antigens, antigen presenting cells and lymphocytes from the CNS to regional lymph nodes. The largest in volume is the 280 ml of ISF (Bergsneider 2001) in the human brain that drains almost exclusively to regional lymph nodes along the walls of capillaries and arteries (Weller et al. 2009). The most studied of the fluids is cerebrospinal fluid (CSF) with a total volume of 140 ml in the human brain; 30 ml of CSF is in the cerebral ventricles and 110 ml in the cerebral and spinal subarachnoid spaces (Bergsneider 2001). CSF drains partly into the blood and partly to regional lymph nodes in the cervical and lumbar regions (Weller et al. 2009). Due to the very narrow and restricted pathways for the drainage of ISF from CNS parenchyma, it is unlikely that antigen presenting cells and lymphocytes traffic from CNS to lymph nodes with ISF. Conversely, the relatively wide channels that lead from the subarachnoid space and carry CSF into nasal (Kida et al. 1993a) or lumbar lymphatics have adequate capacity for the transport of antigen presenting cells and lymphocytes to regional lymph nodes. Here we examine in detail the pathways for the drainage of CSF and ISF to determine their capacity for the transfer of antigens and immune cells from the CNS to regional lymph nodes.
CSF: production and drainage
The majority of CSF is produced by the choroid plexuses in the cerebral ventricles at the rate of 350 μl/min (Davson et al. 1987), although a proportion of CSF may be derived from ISF (Johanson et al. 2008). CSF passes through the ventricular system into the subarachnoid space on the surface of the brain and spinal cord and drains directly into the blood via arachnoid villi and granulations (Fig. 1a and b). Although arachnoid villi and granulations are prominent in the major venous sinuses associated with the brain and spinal cord in humans, arachnoid villi are very small in rodents and sheep (Johanson et al. 2008). In rats, small arachnoid villi are associated with veins on the dorsal surface of the olfactory bulbs (Fig. 1a) (Kida et al. 1993a). CSF passes from the subarachnoid space into a series of channels in the cores of arachnoid granulations in humans to drain through channels in the cellular arachnoid cap of the granulation to reach the endothelium lining venous sinuses (Fig. 1b) (Weller 1998, 2005). Experiments in the monkeys suggest there is bulk flow of CSF from arachnoid villi into the venous sinuses by macrovacuolar transfer across the venous endothelium (Fig. 1b) (Tripathi and Tripathi 1974). However, it is not known whether the restricted trans-cellular route would allow the transfer of antigen presenting cells or lymphocytes from CSF into the blood.
Lymphatic drainage of the CSF was recorded in rodents more than 100 years ago (Schwalbe 1869) and in humans at the beginning of the 20th century (Zwillinger 1912). More recent qualitative and quantitative data suggest that at least 50 % of CSF drains to lymph nodes in rodents (Cserr and Knopf 1992), and that the lymphatic drainage pathways are well developed in large mammals and in humans (Johanson et al. 2008). In rodents, the subarachnoid space containing CSF over the surface of the brain is restricted to channels alongside the major cerebral arteries and to the capacious basal cisterns (Zhang et al. 1992; Kida et al. 1993a). Injection studies have shown that CSF passes through the subarachnoid space to the inferior aspect of the olfactory bulbs and then drains into nasal lymphatics by passing through the cribriform plate of the ethmoid bone along defined channels adjacent to olfactory nerves (Kida et al. 1993a; Johanson et al. 2008) (Fig. 1a). Similar drainage pathways have been demonstrated in humans (Weller et al. 2009); tracers injected into the CSF, drain from the subarachnoid space into sheaths of lymphatics in the nasal mucosa (Johanson et al. 2008) (Fig. 1b). Few studies have investigated lymphatic drainage of CSF from the spinal canal, but it has been shown that tracers in the spinal CSF drain to lumbar lymph nodes (Kida et al. 1993a).
Interstitial fluid (ISF): production and drainage
Interstitial fluid is derived from the blood at the rate of 0.1–0.3 μlmin−1g−1 in rodents (Abbott 2004). Fluid and soluble nutrients pass through the endothelium of capillaries (the blood–brain barrier) (Fig. 2a) and are distributed along capillary basement membranes and through the narrow extracellular spaces to cells of the CNS (Abbott 2004). Clearance of ISF and soluble metabolites from the CNS is first by diffusion through the extracellular spaces, and then by drainage out of the brain along basement membranes in the walls of capillaries (Figs. 1 and 2a) and basement membranes between smooth muscle cells in the tunica media of arteries (Figs. 1 and 2b). Veins and venules do not appear to be involved in the perivascular lymphatic drainage pathway for ISF from brain parenchyma (Carare et al. 2008) (Fig. 1). This is possibly because there is no direct anatomical route from the brain parenchyma to cervical lymph nodes along perivenous spaces, whereas there is a continuous route for lymphatic drainage along the walls of capillaries and arteries to the carotid and vertebral arteries in the neck that are in proximity to cervical lymph nodes (Fig. 1) (Weller et al. 2009).
Fig. 2.
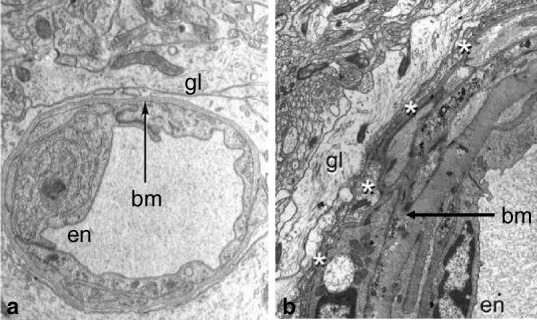
Basement membranes in the walls of a human capillary and artery. (a) Capillary: the lumen is surrounded by endothelium (en) and the basement membrane (bm) encompasses the abluminal surface of the endothelium, separating it from the perivascular glia (gl). X 14000 (b) Artery wall: the endothelium (en) surrounds the lumen and basement membrane (bm) is interposed between the smooth muscle cells in the tunica media. A thin-layer of leptomeningeal cells (white asterisks) separates the perivascular glia (gl) from the artery wall. Basement membranes in the walls of capillaries and arteries are the conduits for perivascular lymphatic drainage of fluid and solutes from the CNS parenchyma as shown by tracer studies and by the distribution of Aβ in cerebral amyloid angiopathy in humans (Carare et al. 2013). X 8400 Reproduced with permission from (a) (Preston et al. 2003) (b) (Zhang et al. 1990)
The perivascular pathway serves as the route for lymphatic drainage of fluid and solutes from the human brain (Weller et al. 2009). This is well demonstrated by the entrapment of Aβ as insoluble deposits in basement membranes of cerebral capillaries and arteries as cerebral amyloid angiopathy in the ageing brain and in Alzheimer’s disease (Carare et al. 2013). Evidence that basement membranes in the walls of cerebral capillaries and arteries are the pathway for drainage of fluid and solutes comes for experimental studies in which tracers injected into the brain parenchyma co-localise with basement membrane proteins (Carare et al. 2008; Carare et al. 2013). In humans, Aβ is deposited in capillary and arterial basement membranes in CAA (Carare et al. 2013; Preston et al. 2003) which suggests that the perivascular drainage pathways are important routes for the elimination of soluble metabolites from the human brain (Weller et al. 2008). The motive force for such drainage appears to be the contrary (reflection) waves that follow the pulse wave in artery walls (Schley et al. 2006). Expansion and recoil in the walls of arteries with the passage of the pulse wave may alter the conformation of basement membranes to produce a valve-like action that prevents reflux of interstitial fluid during the passage of the pulse wave. The motive force for drainage of ISF and solutes appears to fail as cerebral arteries stiffen with age with the consequent deposition of Aβ in vessel walls (Carare et al. 2013).
Quantitative studies suggest that at least 85 % of ISF drains along the walls of cerebral and leptomeningeal arteries through the base of the skull to cervical lymph nodes (Szentistvanyi et al. 1984). Furthermore, there is a certain degree of directionality of the lymphatic drainage as when radioiodinated serum albumin is injected into one side of the brain it drains to lymph nodes on the same side of the neck as the injection. The speed of lymphatic drainage from the brain is equivalent to the speed of lymphatic drainage from other organs (Szentistvanyi et al. 1984). The capacity of perivascular lymphatic drainage from the brain with regard to molecular size has been tested by showing that solutes ranging from 30–150 kDa drain along basement membranes in the walls of cerebral arteries (Carare et al. 2008; Hawkes et al. 2011; Barua et al. 2012). Larger particles, such as Indian ink and 0.2 μm diameter fluorospheres do not drain along basement membranes in artery walls but instead they accumulate in expanded periarterial spaces between the vessel walls and the brain parenchyma (Zhang et al. 1992; Carare et al. 2008). These are not true drainage pathways as the injected particles remain in the perivascular spaces for many months or years (Zhang et al. 1992).
Relationship between CSF and ISF
The relationships between CSF and ISF are different in white and grey matter. For example, in acute hydrocephalus, when drainage of CSF from the ventricles is impeded, CSF passes into the periventricular white matter, often through ruptured ependyma resulting in interstitial oedema. However, the periventricular grey matter remains largely unaffected (Weller 1998). When tracers such as horseradish peroxidase and fluorescent Aβ are injected into the CSF, they pass into the brain alongside arteries as they penetrate the surface of the brain (Rennels et al. 1985; Iliff et al. 2012). Tracers then spread through the interstitial spaces of CNS tissue to paravenous spaces. These experiments suggest that the relationships between CSF and ISF are complex and may vary.
Scanning electron microscope studies have shown that the pia mater in humans is reflected from the surface of the brain and spinal cord to cover arteries and veins in the subarachnoid space, thus separating the CSF from the surface of the brain and from perivascular spaces (Hutchings and Weller 1986; Nicholas and Weller 1988; Weller 2005). Although red blood cells and other particulate matter in the CSF do not penetrate the pia mater, macrophages, fluid and solutes appear to pass readily through the thin layer of pia mater (Hutchings and Weller 1986). The main barrier to the passage of inflammatory cells from the CSF into the surface layers of the brain appears to be the compacted astrocyte processes of the glia limitans (Weller 1999).
Drainage of soluble antigen from CNS to regional lymph nodes
Detailed studies by Helen Cserr and her group have shown that radioiodinated serum albumin injected into both CSF and brain parenchyma drain to cervical lymph nodes (Szentistvanyi et al. 1984). Furthermore, antigens injected into the brain and into CSF result in the formation of antibodies in the cervical lymph nodes and spleen (Cserr et al. 1992). This was demonstrated by the injection of albumin or myelin basic protein into the CSF of rats; intracerebral injection of such antigens is more immunogenic than injection of antigen into the CSF (Cserr et al. 1992).
Traffic of antigen presenting cells from CNS to regional lymph nodes
The question of migration of antigen presenting cells from the CNS parenchyma to regional lymph nodes is still not completely resolved.
Resident antigen presenting cells in the CNS consist of microglia and perivascular macrophages. Dendritic cells, derived from the bone marrow, are also found in association with the healthy non-inflamed brain; they are located in the dura, leptomeninges and choroid plexus in the rat and in small numbers in periventricular regions the brain (Chinnery et al. 2010; Bulloch et al. 2008). Dendritic cells have also been identified in the human choroid plexus (Serot et al. 1997). In the inflamed brain, there is activation of microglia and perivascular macrophages, with increased expression of MHC class II antigen (Pachter et al. 2003; Kida et al. 1993b); dendritic cells, derived from the bone marrow are also increased in number in the inflamed brain (Hatterer et al. 2008).
Microglia do not appear to migrate from the brain parenchyma. Perivascular macrophages are a population of antigen presenting cells that are distinct from pericytes and microglia (Kida et al. 1993b); they act as scavengers in the perivascular spaces taking up solutes draining along perivascular pathways (Carare et al. 2008; Hawkes and McLaurin 2009; Hawkes et al. 2012). Perivascular cells also ingest particles in the perivascular spaces, and when laden with particulate matter they do not seem to migrate from the central nervous system (Kida et al. 1993b).
Much attention has been focused upon the migration of dendritic cells from CNS to cervical lymph nodes. Dendritic cells in most organs of the body migrate to regional lymph nodes and act as efficient stimulators of T lymphocytes. In healthy animals dendritic cells injected into the CSF, migrate to regional lymph nodes but home to the B-cell areas rather than the T-cell areas of cervical lymph nodes (Hatterer et al. 2006). However, when they are injected into the parenchyma of the brain, dendritic cells move only a short distance from the injection site and do not reach cervical lymph nodes (Hatterer et al. 2006). In EAE, dendritic cells appear to migrate from the CSF into brain tissue and also migrate to cervical lymph nodes leading to an accentuation of clinical signs (Hatterer et al. 2008). The most probable route for traffic of dendritic cells and macrophages from the CSF to cervical lymph nodes is through the drainage channels that pass from the subarachnoid space inferior to the olfactory bulbs, through the cribriform plate into nasal lymphatics (Fig. 1) (Kaminski et al. 2012). T-cells follow the same route (Goldmann et al. 2006). Upon injection into rat striatum, microglia and bone marrow-derived dendritic cells generated in vitro leave the CNS via the blood stream, to reach deep cervical lymph nodes, mesenteric lymph nodes, and spleen (Hochmeister et al. 2008).
Given the restricted nature of ISF drainage pathways along 100–150 nm wide basement membranes in the walls of capillaries and arteries, it is unlikely that antigen presenting cells traffic from the brain to cervical lymph nodes by this route (Carare et al. 2008; Weller et al. 2010). When particulate matter, much smaller in size than migrating antigen presenting cells, is injected into the brain parenchyma, it accumulates within perivascular spaces but does not seem to drain out of the brain (Carare et al. 2008; Zhang et al. 1992). It seems unlikely therefore that antigen presenting cells migrate along blood vessel walls directly to lymph nodes. However, macrophages in perivascular spaces near the surface of the brain or spinal cord may well migrate into the CSF and traffic to lymph nodes by this route.
Role of regional lymph nodes in immune reactions in the brain
One of the methods of inducing immunological reactions in the brain is experimental autoimmune encephalomyelitis (EAE) induced by the injection of myelin antigens and Freund’s adjuvant into the footpad or another peripheral site. Classically, acute active EAE is characterised by invasion of lymphocytes and microglia activation mainly in the spinal cord 12–15 days post inoculation (dpi) of antigen. Few lesions occur in the cerebral hemispheres in acute EAE until 20 dpi (Sun et al. 2000). However, a cryolesion on the surface of the brain, 8 days dpi results in a 6-fold enhancement of cerebral EAE at 15 dpi. Removal of cervical lymph nodes at the time of the cryolesion reduces the level of cerebral EAE by 40 % (Phillips et al. 1997). Clinical signs related to spinal cord disease in these animals and no clinical signs attributable to cerebral inflammation were detected. Adoptive transfer of lymphocytes from animals with cryolesion-EAE induced EAE in the recipient’s brain in preference to the spinal cord, suggesting that lymphocytes derived from cryolesion-EAE animals specifically target the brain (Lake et al. 1999). The results of these studies suggest that immune responses in cervical lymph nodes play a significant role in the induction of EAE, possibly associated with lymphatic drainage of antigens from the brain or by neural stimulation (Sun et al. 2000). Excision of cervical and lumbar lymph nodes draining brain and spinal cord delayed relapse and EAE pathology within the spinal cord in chronic relapsing EAE suggesting again that antigen-specific immune responses in the CNS are initiated within the CNS-draining lymph nodes (van Zwam et al. 2009a).
Mechanisms for involvement of cervical lymph nodes in immune reactions in the brain
The central nervous system normally has no direct physical contact with the external environment. Therefore, microorganisms or other antigens only enter the brain following passage through peripheral tissues such as the nasal mucosa, other parts of the respiratory system, skin, or gastrointestinal tract. Inevitably, therefore, such antigens are presented in lymph nodes draining those peripheral organs before the organisms such as viruses enter the brain. The only exception to this is when antigens enter the CNS through direct trauma or surgical implantation. In some ways, therefore, some of the experiments demonstrating immunological privilege are not within the normal range of antigen presentation to the CNS. It has been shown that allografts of skin implanted in the brain survive for longer periods than if they are implanted into other organs of the body. However, if the same allograft is implanted in the skin, there is rapid rejection of the allograft in the brain (Galea et al. 2007). Such experiments strongly suggest that T-lymphocyte dependent immunological reactions in the brain depend upon previous exposure of the antigen to lymph nodes in non-CNS sites. Immunological privilege may depend upon the lack of effective traffic of antigen presenting cells from the brain parenchyma to regional lymph nodes. Figure 3 summarises a possible mechanism for immunological reactions in the brain (Weller et al. 2010). Following entry of antigen, such as a virus, into the body, B-cell and T-cell responses are induced in regional lymph nodes draining tissues such as lung and gut. Lymphocytes, sensitised to these antigens, migrate via the blood to lymph nodes throughout the body, including the cervical and lumbar lymph nodes that drain the CNS. Following arrival of the foreign antigen in the CNS or the release of auto-antigen from the brain, antigens drain to cervical or lumbar lymph nodes in which they encounter T lymphocytes, sensitised to those same antigens. The last step may be the addressing of antigen-sensitised lymphocytes to target the CNS, possibly by the induction of specific surface integrins (Engelhardt 2008). In this way, it is possible that the drainage of soluble antigens along the perivascular pathways from CNS parenchyma to cervical or lumbar lymph nodes could fulfil all the conditions required for the induction of immunological reactions in the CNS without the absolute requirement of trafficking of antigen presenting cells from CNS parenchyma to regional lymph nodes. Alternatively, induction of immunological reactions in the CNS could rely upon a combination of antigen drainage along perivascular pathways and migration of antigen presenting cells along CSF lymphatic drainage pathways.
Fig. 3.
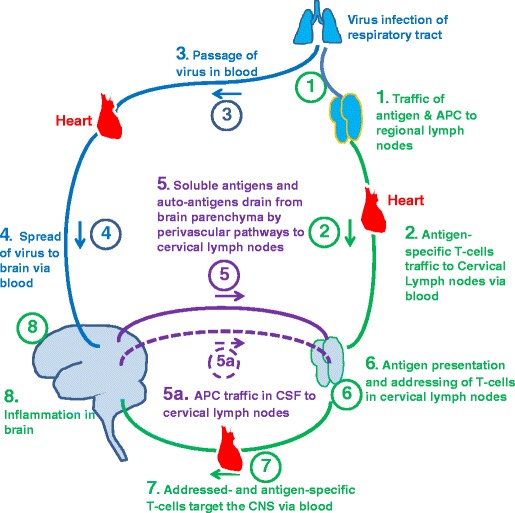
Generation of immune and autoimmune reactions in the brain. Foreign organisms enter the body via exposed surfaces such as skin, respiratory tract and gut. 1) Antigen and antigen presenting cells from those organs traffic to regional lymph nodes and stimulate the production of antigen specific T lymphocytes that, 2) traffic to cervical lymph nodes and other groups of lymph nodes via the blood. 3 &4) organisms such as viruses reach the brain via blood. 5) Soluble viral antigens and autoantigens drain along perivascular pathways to cervical lymph nodes and 5a) antigen presenting cells (APC) in the CSF also traffic to cervical lymph nodes via CSF drainage pathways. 6) Soluble antigens and autoantigens are presented to antigen-specific T lymphocytes in the cervical lymph nodes and T cells are addressed through the induction of integrins to target the brain. 7) Integrin specific, antigen specific T lymphocytes enter the blood and traffic to the brain and induce inflammation (8)
Another aspect to consider is the difference between the immunological environment of the brain parenchyma and the CSF. Although the brain parenchyma exhibits a high level of immune control, the subarachnoid space containing CSF does not exhibit the same mechanisms and intensity of such control. Inflammatory cells readily enter the CSF via veins and venules in the subarachnoid space but the glia limitans presents a firm barrier to the spread of inflammatory cells into the CNS (Weller 1999). Furthermore, cytokines injected into the brain parenchyma may induce an inflammatory reaction in the leptomeninges rather than in the brain parenchyma itself (Andersson et al. 1992). Although dendritic cells and macrophages traffic from the CSF to the cervical lymph nodes, what relationship does this bear to the brain parenchyma? The induction of tolerance by injection of antigen into the CSF suggests that the relationship between brain, CSF and lymphocyte reactions in cervical lymph nodes is complex and depends upon drainage of antigen from brain and CSF and upon trafficking of antigen presenting cells from CSF to lymph nodes. Understanding this process will be key to therapeutic intervention for immunological disease of the CNS, as further detailed below.
The role of lymphatic drainage pathways from the CNS in immunological disease
Infections
Relations between infections, CNS drainage, and cervical lymph nodes as well as lumbar lymph nodes have not been explored in great depth, and there are few functional studies targeting these sentinel nodes. Nevertheless, these lymph nodes appear to be critical for immune responses to CNS infection by neurotropic viruses, such as rubella, varicella zoster, herpes simplex, Epstein-Barr and HIV. Deleidi and Isacson (2012) argue that, based upon recent developments, relations between viral infection, brain inflammation and acute manifestations of neurodegenerative disease are not linear. Instead they propose “causal relationships between viral infection-associated inflammation and cytokine-induced sequelae that together prime relatively specific groups of vulnerable neurons to degenerate, in response to cellular insults”. Hence, there are numerous questions to be pursued on CNS-lymph connections for neurodegenerative disease and infections as well as commensal microbiota.
Selected studies of interest, mostly using Theilers murine encephalomyelitis picornavirus (TMEV) and the mouse hepatitis coronavirus (JHMV), are briefly discussed below.
A debated topic is whether naïve T-cells can be primed in the CNS target organ, or whether secondary lymphoid organs are required. McMahon et al. (2005) provided evidence using T-cell receptor transgenic mice that in both TMEV and in EAE, epitope spreading is initiated in the CNS, and not in the spleen or cervical lymph nodes. Local antigen presenting cells for naïve transgenic T-cells had a phenotype compatible with dendritic cells (F4/80-CD11c+CD45hi). In a complementary approach, mice lacking antigen presenting cells of the lymphoreticular system and of the CNS parenchyma still suffered from CNS invasion upon adoptive transfer of activated autoreactive CD4+ T-cells (Greter et al. 2005). CNS myeloid dendritic cells presenting endogenous myelin peptides biased Th17 differentiation in accordance with expression of TGF-beta, IL-6 and IL-23. Plasmacytoid dendritic cells and CD8alpha+ dendritic cells were inferior, but still more efficient than macrophages (Bailey et al. 2007). In mice transgenic for PLP139-151, CD4+ T cells were hyperactivated in the cervical lymph nodes but not spleen, suggesting initial peripheral activation occurs in that site (Zhang et al. 2008).
Navarrete-Talloni et al. (2010) used expression microarrays to assess compartmentalization in deep cervical lymph nodes versus spleen of the immune response in TMEV. In the early phase (day 14) coinciding with intense disease, the B cell immune response, antigen presenting cell activation, and complement expression were the most upregulated pathways. An increase in CD68- and lysozyme-positive cells, presumably macrophages, was seen. During the chronic phase (days 42 and 98), transcription was reduced in the deep cervical lymph nodes. In the early phase, DAP12 was highly upregulated; DAP12 is involved in activation of several leukocyte populations, and is a mediator of dendritic cell migration from peripheral organs to lymph nodes. Similarly, CD68 has a role in dendritic cell maturation and migration of antigen presenting cells from the CNS. Since TMEV itself is not detectable in the deep cervical lymph node with different methods, it appears that the immune response is initiated by antigen presenting cells migrating from the CNS.
Adoptive transfer of Treg limits T-cell proliferation and demyelination in JHMV infection within the cervical lymph nodes, and not in the CNS (Trandem et al. 2010). CD8+ T cells against this mouse hepatitis virus are first detected in the cervical lymph nodes. Expansion in the spleen lags behind, but still precedes CD8+ T-cell accumulation in the CNS (Marten et al. 2003). Cellular immunity against this virus is controlled by CD8+ T cells, antibody responses regulate persistence in the CNS. After virus clearance, antibody secreting cells declined in cervical lymph nodes and spleen, but were retained in the CNS, demonstrating that this environment supports antibody formation even while viral infection has been cleared and chronic inflammation is no longer detectable (Tschen et al. 2006).
Using a gliatropic mouse hepatitis virus strain and conditional depletion of Treg, it was shown that Treg help to support immune privilege during viral infection by acting on the action of autoreactive T cells in the cervical lymph nodes (Cervantes-Barragan et al. 2012).
Transgenic expression of CCL21 by astrocytes is sufficient to support homeostatic CD4+ T-cell proliferation in the cervical lymph nodes even when endogenous CCL19/CCL2 is lacking. Upon Toxoplasma gondii infection, CCL21 facilitates CD4+ T-cell migration into the CNS parenchyma (Ploix et al. 2011).
Goverman’s group demonstrated with elegant transgenic systems that CNS autoimmunity can be driven by CD8+ T-cells expressing dual T-cell receptors recognizing both an MBP epitope and viral antigen (Ji et al. 2010). Although this is to be further substantiated in humans, these data suggest the fascinating mechanism that ubiquitous viral infection in only a subgroup of infected individuals triggers autoimmune disease. This fundamental study helps to at least partly explain the poorly understood chance effect that drives MS risk in concert with genetics and environmental influences (notably EBV and vitamin D) (Ransohoff 2010).
In conclusion, there is a clear need for additional analysis of the role of cervical lymph nodes in relation to infection, for instance to extend the evidence that naïve T cells can be activated in the CNS under certain conditions, and how immunity versus autoreactivity are controlled. For this purpose, excision of cervical and lumbar lymph nodes could be employed in mouse models of neurotropic infection, as done in distinct EAE models.
Gut microbiota
Intense interest in immune-driving activity of the microbiota under homeostasis and during disease has led to several important findings on cervical lymph nodes. Antibiotic treatment limits development of EAE, and transfer of IL-10+CD25+Foxp3+CD4+ T-cells from cervical lymph nodes suggested a role of Treg (Ochoa-Reparaz et al. 2009). Bacteroides fragilis with polysaccharide A (PSA) protects against EAE, with increased numbers of Treg in the cervical lymph nodes (Ochoa-Reparaz et al. 2010a). PSA treatment enhances CD103+ dendritic cells that accumulate in the lymph nodes, and IL-10 is critical for EAE protection (Ochoa-Reparaz et al. 2010b). CD103 is the alphaE integrin that binds to E-cadherin.
In the spontaneous EAE model driven by transgenic expression of the CD4+ T-cell receptor for MOG92-106 on the SJL/J background, the commensal microbiota is critical for disease development. There was good evidence for an ongoing MOG-specific germinal centre reaction in the cervical lymph nodes, critically dependent on MOG expression (Berer et al. 2011). In active rat EAE, memory T-MBP-GPF cells are located in many secondary lymphoid organs in comparable frequencies, including the spleen, inguinal, mesenteric, mediastinal, para-aortic and cervical lymph nodes. Five days after intratracheal immunization, the lung and the mediastinal lymph nodes have the highest frequencies of memory cells, and not the cervical lymph nodes (Odoardi et al. 2012) (supplemental fig. 4).
Upcoming studies likely will soon reveal more about the anatomical and mechanistic links between gut microbiota and cervical lymph node action.
Multiple sclerosis and EAE
MS and its different variations of EAE models obviously are major focus points of questions on CNS-lymphoid drainage, and protective versus autoreactive immunity. A selection of studies is discussed.
Intracerebral injection with OVA protein leads to recruitment of CD205+CD11c+ dendritic cells which ingest this antigen. Subsequently, CD8+ OVA-specific T cells accumulate in the cervical lymph nodes and thereafter in the CNS (Ling et al. 2003). Upon intracerebral injection, mature OVA-loaded DC migrate from the CNS to cervical lymph nodes to prime OVA-specific T-cell responses (Karman et al. 2004) reviewed by (Clarkson et al. 2012).
We demonstrated that different myelin compounds are present in antigen presenting cells within cervical lymph nodes of MS patients, rhesus monkeys with acute EAE, and marmoset monkeys with chronic EAE (de Vos et al. 2002). Our claims to the identity of these antigen presenting cells were purposely cautious, as much more detailed typing of functional subsets would be required. We used more or less generic markers such as MHC-II, acid phosphatase (relatively characteristic for macrophages), CD40, CD80, CD86, CD83, DC-SIGN, and CCR7. For instance, co-expression of CD83 and lysosomal acid phosphatase suggested intermediate or transitional cell types between macrophage and dendritic cell.
In a subsequent study, we compared a series of animal models of CNS insults including inflammation, cuprizone toxicity, and trauma by enthorinal cortex lesion and facial nerve axotomy, for burden of neuronal antigens in the cervical lymph nodes (van Zwam et al. 2009b), in view of axonal damage. Differential loading of antigen presenting cells with neuronal antigens in the cervical lymph nodes was found, generally correlating with the extent of CNS damage. Some evidence was found for pro- versus anti-inflammatory phenotype of antigen presenting cells loaded with myelin versus axonal antigen (further discussed below).
The central issue of soluble antigen drainage versus transport by cells remains unresolved, and is in need of innovative techniques (see Box 1). An informative analysis demonstrated abundant extracellular myelin particles in the perivascular spaces and meninges of MS patients, and not in several reference donor groups. It is not clear why meningeal macrophages do not ingest this debris, other than in stroke patients (Kooi et al. 2009). This study provides firm support for the existence of myelin antigen debris during MS.
Box 1. Challenges and opportunities for progress on CNS drainage and immunity
| - Relative dearth of thoroughly trained (neuro)pathologists, anatomists, and neurologists with an interest in the topic. Many (respected) medical schools have modified their curricula at the expense of basic science training, including anatomy and physiology. In addition, oftentimes a considerable conceptual gap is evident between immunologists and those morphologists. |
| - Anatomic complexity: Subtle and major differences between humans and individual animal model species. Different and overlapping routes for cells, CSF and ISF. Different relationships between ISF and CSF in white versus grey matter. Lack of sensitive in vivo imaging techniques for fluid flow and cell migration. |
| - Highly variable understanding and use of the blood–brain barrier concept, originally defined for exclusion of dyes by Paul Ehrlich, and now also employed (sometimes confusingly for the novice) for leukocyte migration (Bechmann et al. 2007). Similarly, an overly binary interpretation instead of a more balanced use of the immune privilege concept, often taking decades to overcome widespread dogmas such as ‘the CNS has no connection with the lymphoid system’ (Galea et al. 2007). |
| - The tendency-necessity to reduce large studies to a brief summary statement that sometimes becomes overly schematic in subsequent use. For instance, our study on CNS antigens in cervical lymph nodes in MS patients and two monkey species (de Vos et al. 2002) is sometimes cited as claiming it is only dendritic cells transporting antigens, or only macrophages, or both, or intermediate cell types. |
| - Subset complexity: The origins, locations, dynamics and functions of CNS macrophages, microglia and dendritic cells have become much better defined recently (Prinz et al. 2011). The diversity in location and function in health and during disease development is much higher than previously anticipated. In addition, to the occasional understandable confusion and bewilderment of the non-immunologist, functional ‘subsetology’ of leukocytes is taking an unprecedented flight. Functional Th-cell subsets display considerable plasticity, despite the initial concept of true terminal differentiation. Innate lymphoid cells (ILC) form a completely new system functionally comparable to Th-cell subsets. B-cell subsets as well as CD8 T-cell subsets can have regulatory functions, and CD4 Th-subsets can display killer action. Dendritic cell subsets with differential functions are being (re)defined continuously, and phenotypically similar DC can have opposing functions in cervical lymph nodes versus lung e.g. Dunne et al. (2009) (Dunne et al. 2009). The prime role of DC in EAE induction has recently been fundamentally challenged, and their role in induction of Treg further emphasized (Yogev et al. 2012). |
| - The animal models to mimic a single disease such as MS (Gold et al. 2006; Ransohoff 2012), can be very different, even when the same mouse strain, self peptide and adjuvant are used. Seemingly minor differences in the induction protocol can have major consequences. Another critical factor is the microbial environment and notably the gut microbiota (Goverman et al. 1993; Berer et al. 2011), which can differ considerably even for the same strain when bred in house versus individual commercial vendors. |
| - Technical complexities of visualizing cell migration from CNS cross-sectionally or in real time. CNS injections are disruptive by definition, and even volumes of 10 μl can provoke undue tissue damage. If volumes of fluid greater than 0.5 μl are injected into the brain parenchyma, there is always the possibility that the inoculum will leak into the CSF. Postmortem detection of CNS compounds in lymphoid organs does not prove migration. Labeling CNS compounds in vivo (e.g. transgenically, chemically, by PET-tracers) is difficult, and detection should include the migration pathways, not solely the lymphoid organs. Valuable but taxing parabiosis experiments are not universally allowed. Bone marrow chimera approaches are useful but limited by CNS integrity irradiation artefacts. Genomic DNA amplification of transgenes to identify migrating cells may be affected by proliferation outside the CNS. In vivo real time imaging is complicated by limited penetration depth of 2-photon technology, and the long migration times, presumably days, perhaps with the exception of experimental stroke where extensive CNS damage is associated with abundant CNS antigen load already at 24 h (van Zwam et al. 2009b). |
Importantly, Ji et al. (2013) have shown that in CD4+ driven EAE, MHC-I restricted myelin epitopes can be cross-presented by Tip-dendritic cells, characterized by phenotypic similarity to inflammatory monocytes, TNF and iNOS production and not IL-12, and the lack of the transcription factor Zbtb46/zDC expressed by classical dendritic cells (Meredith et al. 2012). Tip-dendritic cells activate both naïve and effector CD8+ T cells ex vivo, and naïve MBP-specific killer cells were activated directly in the CNS. In line with this, the Tip-dendritic cells lack CCR7. This shows that, at least in this system, determinant spreading can occur in the target organ itself and promote killer responses against oligodendrocytes. Intriguingly, these killer cells may also have beneficial action by lysing the Tip-dendritic cells.
An issue very little pursued thus far is whether regulatory B cells are generated in the cervical lymph nodes, in the spleen, and/or in the CNS. With respect to classic B cells, von Budingen et al. (2012) employed next generation sequencing to provide evidence that B-cell autoimmune diversification in MS is initiated and supported both in the periphery (as measured in peripheral blood), and in the cerebrospinal fluid.
Clearly there is a need for additional work on cell transport versus drainage of extracellular antigen, different types of functional antigen presenting cell subsets in the cervical lymph nodes, and regulatory B cell function.
Stroke
We (van Zwam et al. 2009b) and several other groups have considered how tissue damage in (experimental) stroke could lead to brain antigen localization in the cervical lymph nodes and beyond, potentially contributing to autoimmunity and/or regulation, mostly addressed for Treg. In spontaneously hypersensitive rats, E-selectin specific Treg promote efficacy of neurogenesis after ischemia and promote repair after brain injury. Increased numbers of Treg are present in the cervical lymph node and ischemic brain (Ishibashi et al. 2009). The immunity of stroke has been comprehensively reviewed very recently (Chamorro et al. 2012).
Indeed brain-derived antigens can be found in palatine tonsils and cervical lymph nodes after stroke (Planas et al. 2012). This includes neuronal compounds (MAP-2 and NR-2A) as well as MBP and MOG. Antigen presenting cells containing these antigens were MHC-II+, had macrophage features, and were located in T-cell areas. Although based on a limited case series, interesting correlations were found between increased reactivity to neuronal antigens with smaller infarcts and better long-term outcome, versus stroke severity on admission, larger infarcts, and worse outcome associated with high load of myelin antigens. Intriguingly, we previously found that in human MS CLN, neuronal antigens were present in pro-inflammatory antigen-presenting cells, whereas the majority of myelin-containing cells had anti-inflammatory features. This could reflect a different anatomical origin of the cells or different drainage kinetics and mechanisms. Indeed, the neuronal antigen-containing cells in human cervical lymph nodes did not express the lymph node homing receptor CCR7, whereas myelin antigen-containing cells in situ and in vitro did. At present the implications of these parallel findings by Planas et al. (2012) and van Zwam et al. (2009b) are not clear and are in need of corroboration and followup.
Inducible deletion of oligodendrocytes in mice led to myelin antigen localization in deep cervical lymph nodes. However, not even using the most biased approaches, did immune activation or relaxation of tolerance induce adaptive immunity and CNS inflammation (Locatelli et al. 2012).
Tolerance
The specialized role of cervical lymph nodes in induction of mucosal tolerance is firmly established. This includes nasal antigen instillation and application locally in the oral cavity, for instance sublingual. A non-exhaustive listing of mechanisms includes: Induction of regulatory T cells producing IL-10, TGF-beta and/or IL-4 e.g. (Burkhart et al. 1999; Rynda et al. 2008; Zhong et al. 2012); Preferential expression of IgG2b in the microenvironment of the cervical lymph node, differentially activating dendritic cells through inhibitory Fc-gamma IIIB-receptor signalling (van Helvoort et al. 2004; Samsom et al. 2005); and regulation of the threshold for mucosal tolerance by secretory leukoprotease inhibitor (SLIPI) (Samsom et al. 2007). A less classic means of regulation is the induction of Foxp3+ invariant natural killer T cells (iNKT) by TFG-beta in cervical lymph nodes upon administration of their T-cell receptor agonist alpha-galactosylceramide (Monteiro et al. 2010) that limits EAE.
The general impression is that the tolerance inducing propensity of the cervical lymph node is readily overcome when some sort of adjuvant action is introduced (Samsom et al. 2004). In accordance, using three mouse EAE model variations, we could clearly demonstrate that excision of lumbar and cervical lymph nodes never led to aggravated disease, but instead limited relapsing disease (van Zwam et al. 2009a). Hence, in these three distinct EAE models there was no discernible disease limiting-tolerizing action of the cervical lymph nodes. Adjuvant action is employed in sublingual tolerance induction against allergens, and TLR ligands evoke mixed Th1-Treg CD4 T-cell responses. Compounds such as dexamethasone plus vitamin D3, or probiotics are potential Treg adjuvants by means of IL-10 induction (Moingeon and Mascarell 2012).
Proteins applied in the sublingual mucosa are captured by several subsets of antigen presenting cells in 15 to 30 min, which migrate to draining cervical submaxillary lymph nodes. Local dendritic cell types include Langerhans cells (mucosa), myeloid dendritic cells (lamina propria), and plasmacytoid DC (submucosa) (Moingeon and Mascarell 2012). Interestingly, oral macrophage-like cells (CD11b+CD11c-CD206+RALDH2+) (Mascarell et al. 2011) can mediate sublingual tolerance induction by migration to the cervical lymph nodes and stimulation of IFN-gamma/IL-10 producing Foxp3 CD4+ Treg. This emphasizes that not only dendritic cells can have tolerogenic action in this location.
Compared to sublingual soluble antigen transport, Wraith and colleagues demonstrated that intranasally applied MBP (myelin basic protein) peptide accessed lymphoid tissues and blood peaking around 2.5 to 4 h. High concentrations developed in the draining cervical lymph nodes, but significant and physiologically relevant levels were also found in the spleen, and in non-draining inguinal lymph nodes. These findings emphasize at least some systemic distribution of soluble antigens downstream of the cervical lymph node, an important fact in the interpretation in studies employing nasal instillation.
After entorhinal cortex lesion in mice, EAE induction by PLP (proteolipid protein) was reduced, coinciding with the presence of antigen presenting cells laden with CNS antigens (Mutlu et al. 2007). This would suggest that—at least under certain conditions—self antigens released from the CNS may promote tolerance.
In a very recent study, Yamazaki and colleagues (2012) demonstrate that cervical lymph nodes have high frequencies of Foxp3+ Treg as well as of ROR-gammat+CD4+ T-cells compared to other lymph nodes. Mice lacking Myd88 and TICAM-1/TRIF had no reduction in high frequencies of these Treg. Conversely, these TLR adaptor molecules did drive the high frequency of ROR-gammat+ CD4+-cells in the cervical lymph node. Dendritic cells from the oral cavity drive Foxp3 T-reg induction. However, other than in the intestine, this does not appear to be driven by CD103+ dendritic cells, and there is no RALDH2 expression. Instead, CD8- dendritic cells may be responsible (Yamazaki et al. 2012). The group of Turka (Lieberman et al. 2012) showed that upon transfer of polyclonal Treg from cervical lymph nodes of Foxp3-GFP reporter mice, these do not home directly to cervical lymph nodes. Instead, they accumulate over weeks, and CD69 as a marker of recently activated Treg cells is associated with more rapid and pronounced enrichment. The experiments suggest that antigen specificity may control this accrual, likely resulting from a combination of local proliferation, survival and retention.
In conclusion, features of the cervical lymph nodes in relation to tolerizing action are being more clearly defined.
Insights from (nano)particle technology
Innovative use of new and classic particle technologies enhances understanding of CNS-lymph connections, and improves manipulation of tolerogenic and immunogenic responses in the cervical lymph node. Some selected examples are discussed. Use of superparamagnetic iron oxide particles (SPIO) has been comprehensively reviewed recently (Weinstein et al. 2010), including a welcome definition of nanotechnology: ‘nano-sized structures that contain at least one dimension between 1 to 100 nm in size … and possess new or enhanced properties that are unattainable at both smaller (quantum) and larger (macromolecular) levels’ (Hartman et al. 2008). Virus-sized SPIO (Combidex) were applied by different routes in rats. Intraventricular and intracerebral injection provided MRI signals in the deep cervical lymph nodes, and lesser signals in the superficial cervical nodes. Iron could be detected already within 2 h (Muldoon et al. 2004). Upon intravenous injection of ultrasmall SPIO into Lewis rats with acute EAE resulted in MRI brain signals within 1 h. USPIO accumulated in the cervical lymph nodes from 24 h onward, and stabilising at 72 h. They were present in infiltrated macrophages both in medulla and subcapsular sinus (Oude Engberink et al. 2010).
By densely coating nanoparticles with poly-ethylene glycol (PEG), Nance and colleagues (2012) (Nance et al. 2012) showed that sizes as large as 114 nm diameter diffused within human and rat brain. This is in contrast to the previously established upper limit of 64 nm. There are extracellular spaces larger than 200 nm, and more than one quarter of all pores are larger than 100 nm. Uncoated nanoparticles are immobile, even the 40 nm particles.
Nanoparticles have been used to manipulate nasal tolerance versus humoral immunity: poly-lactic-co-glycolic acid (PLGA), PLGA with N-trimethyl chitosan (TMC) and TMC-tri-polyphosphate (TMC-TPP). From these, only PLGA nanoparticles induced immunoregulation by enhanced Foxp3 in cervical lymph nodes (Keijzer et al. 2011). Wang et al. (2012) used a nanoemulsion soybean oil adjuvant to effectively induce tolerance in an experimental autoimmune thyroiditis, characterized by increased percentages of Tregs in the cervical lymph nodes, and enhanced Foxp3 and TGF-beta. Frenkel et al. (2008) used a nasal proteasome adjuvant to activate microglia and reduce amyloid deposition in a transgenic Alzheimer model. The adjuvant reached the cervical lymph nodes, and scavenger receptor-A positive cells were found.
We would like to propose that differential migration, uptake and retention properties of selected particle technologies can be experimentally utilized to assess immunization versus tolerance. For instance, coupling model antigens or myelin antigens to SPIO and USPIO, also by means of linkers digestible by proteases, in theory would allow analysis of T-cell and antibody responses first in the CNS and then in the cervical lymph nodes. A parallel approach has been used to analyse immune responses against OVA-Sepharose beads trapped in the lung bed (Willart et al. 2009).
Implications for diagnosis, manipulation and therapy
Whereas diagnostic assessment of cervical lymph nodes is central to cancer of the head-neck region, not much is available with respect to inflammatory disease of the CNS. There is little evidence of palpable increases in size and/or numbers, for instance during MS. In vivo sampling by means of ultrasound guided fine needle aspiration of MS patients and healthy volunteers is possible but cell yield is limited (Fabriek et al. 2005). A very recent study assessed cervical lymph nodes in thyroiditis by sonography, demonstrating increased numbers of benign hyperplastic lymph nodes (Brancato et al. 2013). These and other innovative imaging techniques will hopefully be further developed for imaging of secondary lymphoid organs in CNS disease as well.
As partly previously argued by us (Weller et al. 2010; de Vos et al. 2002) and others e.g. (Mohammad et al. 2012), in addition to improved insight into immunopathology, detailed mechanistic understanding of drainage of cells and solutes from the CNS potentially has considerable implications for therapy. Secondary lymphoid organs including the cervical and lumbar lymph nodes, as well as the spleen are directly connected to the systemic and lymphatic circulation. Hence, systemically administered (immuno)therapeutics much more easily attain pharmacologically useful titers in those organs than in the MS lesions in the CNS parenchyma hidden behind multiple tissue barriers. When indeed both detrimental (including epitope spreading) and regulatory T- and B-cell responses are for the most part initiated in the secondary lymphoid organs, local blockade or stimulation may prove effective. Therapeutic relevance of CNS drainage is underscored by a very recent intriguing study that in Th1- and Th17-driven EAE models, disease can be ameliorated by administration of Aβ (Grant et al. 2012).
Further identification of differential migration mechanisms of functional leukocyte subsets out of the CNS might translate into approaches limiting or stimulating exit dependent on the distinct function to be manipulated. For instance in stroke, differential kinetics of antigen drainage (early) versus Treg induction (later) could be employed. As an example, migration of cornea (partly immune privileged) dendritic cells to cervical lymph nodes is driven by VEGFR-3 (Hamrah and Dana 2007). The recent findings on accumulation versus transport of Aβ from the CNS (Mawuenyega et al. 2010; Cramer et al. 2012) suggest appealing new options—albeit highly speculative at this point—to promote transport by manipulating CSF and ISF flow for instance by improving vessel flexibility and movement, and perhaps by stimulating cell migration. The dogma that microglia cannot migrate out of the CNS may be attacked with novel immunophenotypic marker profiles currently becoming available.
Concluding remarks and future directions
Taking the work of Schwalbe (1869) as an arbitrary initial founding observation on the lymph-CNS connection, it could be argued that progress in almost 150 years on this topic has been slow to develop. Conversely, when taking the groundbreaking work of Cserr and Knopf as the benchmark, the insights have improved considerably in the past three decades. In Box 1, we attempt to (perhaps somewhat presumptuously) identify major challenges which when addressed could expedite progress in this field considerably, mostly for the benefit of investigators new to the field. We very recently outlined a series of major outstanding questions (see Table 1 in Laman and Weller (2012)). In brief, we argue here that major advances would be: the development of in vivo imaging techniques for (labeled) myelin compounds transported by CSF or ISF flow; imaging in real time of antigen presenting cells migrating with sequestered self antigen; careful identification of phenotype and functionality of subsets of antigen presenting cell subsets both in the CNS and lymphoid organs, because these might well be a spectrum with shared characteristics of macrophages and dendritic cells, and with activating versus regulatory functions; a further emphasis on drainage of antigen and cells from the spinal cord to lumbar lymph nodes, since this has been explored only to a very limited extent; exploration of differential action of current and novel immunotherapies (notably those interfering with traffic such as natalizumab and fingolimod) for MS on antigen presenting cells containing self antigens; and finally whether antigen and cell drainage might provide insight into very serious side effects of immunotherapy, such as the life-threatening and highly disabling progressive multifocal leukoencephalopathy (PML) occurring in 1 out of 500 MS patients treated with natalizumab (anti-VLA4) due to recrudescence of the JC polyoma virus.
Acknowledgments
Work at Erasmus MC Rotterdam is supported by individual (MS-01-471) and program grants (MS-10-490c) of the Dutch MS Research Society. Work in Southampton is supported by Alzheimer Research UK. We apologize to authors of relevant studies we could not cite here due to space constraints, and most of those studies are retrieved from the reviews we do cite. The authors acknowledge insightful discussions with Dr. Roxana Carare (Southampton, United Kingdom) and Prof. Ingo Bechmann (Leipzig, Germany), and their critical reading of the manuscript.
Abbreviations
- APC
Antigen presenting cell
- CLN
Cervical lymph nodes
- CNS
Central nervous system
- CSF
Cerebrospinal fluid
- DC
Dendritic cells
- EAE
Experimental autoimmune/allergic encephalomyelitis
- GFP
Green fluorescent protein
- ISF
Interstitial fluid
- MBP
Myelin basic protein
- MOG
Myelin oligodendrocyte glycoprotein
- MRI
Magnetic resonance imaging
- MS
Multiple sclerosis
- OVA
Ovalbumin
- PLP
Proteolipid protein
- Treg
Regulatory T cells
Contributor Information
Jon D. Laman, Phone: +31-10-704-3796, FAX: +31-10-7044731, Email: j.laman@erasmusmc.nl
Roy O. Weller, Email: row@soton.ac.uk
References
- Abbott NJ. Evidence for bulk flow of brain interstitial fluid: significance for physiology and pathology. Neurochem Int. 2004;45:545–552. doi: 10.1016/j.neuint.2003.11.006. [DOI] [PubMed] [Google Scholar]
- Andersson PB, Perry VH, Gordon S. Intracerebral injection of proinflammatory cytokines or leukocyte chemotaxins induces minimal myelomonocytic cell recruitment to the parenchyma of the central nervous system. J Exp Med. 1992;176:255–259. doi: 10.1084/jem.176.1.255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey SL, Schreiner B, McMahon EJ, Miller SD. CNS myeloid DCs presenting endogenous myelin peptides ‘preferentially’ polarize CD4+ T(H)-17 cells in relapsing EAE. Nat Immunol. 2007;8(2):172–180. doi: 10.1038/ni1430. [DOI] [PubMed] [Google Scholar]
- Barua NU, Bienemann AS, Hesketh S, Wyatt MJ, Castrique E, Love S, Gill SS. Intrastriatal convection-enhanced delivery results in widespread perivascular distribution in a pre-clinical model. Fluids Barriers CNS. 2012;9:2. doi: 10.1186/2045-8118-9-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bechmann I, Galea I, Perry VH. What is the blood–brain barrier (not)? Trends Immunol. 2007;28:5–11. doi: 10.1016/j.it.2006.11.007. [DOI] [PubMed] [Google Scholar]
- Berer K, Mues M, Koutrolos M, Rasbi ZA, Boziki M, Johner C, Wekerle H, Krishnamoorthy G. Commensal microbiota and myelin autoantigen cooperate to trigger autoimmune demyelination. Nature. 2011;479(7374):538–541. doi: 10.1038/nature10554. [DOI] [PubMed] [Google Scholar]
- Bergsneider M. Evolving concepts of cerebrospinal fluid. Neurosurg Clin N Am. 2001;36:631–638. [PubMed] [Google Scholar]
- Brancato D, Citarrella R, Richiusa P, Amato MC, Vetro C, Galluzzo CG. Neck lymph nodes in chronic autoimmune thyroiditis: the sonographic pattern. Thyroid Off J Am Thyroid Assoc. 2013;23(2):173–177. doi: 10.1089/thy.2012.0375. [DOI] [PubMed] [Google Scholar]
- Bulloch K, Miller MM, Gal-Toth J, Milner TA, Gottfried-Blackmore A, Waters EM, Kaunzner UW, Liu K, Lindquist R, Nussenzweig MC, Steinman RM, McEwen BS. CD11c/EYFP transgene illuminates a discrete network of dendritic cells within the embryonic, neonatal, adult, and injured mouse brain. J Comp Neurol. 2008;508:687–710. doi: 10.1002/cne.21668. [DOI] [PubMed] [Google Scholar]
- Burkhart C, Liu GY, Anderton SM, Metzler B, Wraith DC. Peptide-induced T cell regulation of experimental autoimmune encephalomyelitis: a role for IL-10. Int Immunol. 1999;11(10):1625–1634. doi: 10.1093/intimm/11.10.1625. [DOI] [PubMed] [Google Scholar]
- Carare RO, Bernardes-Silva M, Newman TA, Page AM, Nicoll JAR, Perry VH, Weller RO. Solutes, but not cells, drain from the brain parenchyma along basement membranes of capillaries and arteries. Significance for cerebral amyloid angiopathy and neuroimmunology. Neuropathol Appl Neurobiol. 2008;34:131–144. doi: 10.1111/j.1365-2990.2007.00926.x. [DOI] [PubMed] [Google Scholar]
- Carare RO, Hawkes CA, Jeffrey M, Kalaria RN, Weller RO. Cerebral amyloid angiopathy, prion angiopathy, CADASIL and the spectrum of protein elimination-failure angiopathies (PEFA) in neurodegenerative disease with a focus on therapy. Neuropathol Appl Neurobiol. 2013 doi: 10.1111/nan.12042. [DOI] [PubMed] [Google Scholar]
- Cervantes-Barragan L, Firner S, Bechmann I, Waisman A, Lahl K, Sparwasser T, Thiel V, Ludewig B. Regulatory T cells selectively preserve immune privilege of self-antigens during viral central nervous system infection. J Immunol. 2012;188(8):3678–3685. doi: 10.4049/jimmunol.1102422. [DOI] [PubMed] [Google Scholar]
- Chamorro A, Meisel A, Planas AM, Urra X, van de Beek D, Veltkamp R. The immunology of acute stroke. Nat Rev Neurol. 2012;8(7):401–410. doi: 10.1038/nrneurol.2012.98. [DOI] [PubMed] [Google Scholar]
- Chinnery HR, Ruitenberg MJ, McMenamin PG. Novel characterization of monocyte-derived cell populations in the meninges and choroid plexus and their rates of replenishment in bone marrow chimeric mice. J Neuropathol Exp Neurol. 2010;69:896–909. doi: 10.1097/NEN.0b013e3181edbc1a. [DOI] [PubMed] [Google Scholar]
- Clarkson BD, Heninger E, Harris MG, Lee J, Sandor M, Fabry Z. Innate-adaptive crosstalk: how dendritic cells shape immune responses in the CNS. Adv Exp Med Biol. 2012;946:309–333. doi: 10.1007/978-1-4614-0106-3_18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cramer PE, Cirrito JR, Wesson DW, Lee CY, Karlo JC, Zinn AE, Casali BT, Restivo JL, Goebel WD, James MJ, Brunden KR, Wilson DA, Landreth GE. ApoE-directed therapeutics rapidly clear beta-amyloid and reverse deficits in AD mouse models. Science. 2012;335(6075):1503–1506. doi: 10.1126/science.1217697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cserr HF, Knopf PM. Cervical lymphatics, the blood–brain barrier and the immunoreactivity of the brain: a new view. Immunol Today. 1992;13(12):507–512. doi: 10.1016/0167-5699(92)90027-5. [DOI] [PubMed] [Google Scholar]
- Cserr HF, Harling-Berg CJ, Knopf PM. Drainage of brain extracellular fluid into blood and deep cervical lymph and its immunological significance. Brain Pathol. 1992;2:269–276. doi: 10.1111/j.1750-3639.1992.tb00703.x. [DOI] [PubMed] [Google Scholar]
- Davson H, Welch K, Segal MB. Physiology and pathophysiology of the cerebrospinal fluid. Edinburgh: Churchill Livingstone; 1987. [Google Scholar]
- de Vos AF, van Meurs M, Brok HP, Boven LA, Hintzen RQ, van der Valk P, Ravid R, Rensing S, Boon L, t Hart BA, Laman JD. Transfer of central nervous system autoantigens and presentation in secondary lymphoid organs. J Immunol. 2002;169(10):5415–5423. doi: 10.4049/jimmunol.169.10.5415. [DOI] [PubMed] [Google Scholar]
- Deleidi M, Isacson O. Viral and inflammatory triggers of neurodegenerative diseases. Sci Transl Med. 2012;4(121):121ps123. doi: 10.1126/scitranslmed.3003492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunne PJ, Moran B, Cummins RC, Mills KH. CD11c+CD8alpha+ dendritic cells promote protective immunity to respiratory infection with Bordetella pertussis. J Immunol. 2009;183(1):400–410. doi: 10.4049/jimmunol.0900169. [DOI] [PubMed] [Google Scholar]
- Engelhardt B. Immune cell entry into the central nervous system: involvement of adhesion molecules and chemokines. J Neurol Sci. 2008;274:23–26. doi: 10.1016/j.jns.2008.05.019. [DOI] [PubMed] [Google Scholar]
- Engelhardt B, Coisne C. Fluids and barriers of the CNS establish immune privilege by confining immune surveillance to a two-walled castle moat surrounding the CNS castle. Fluids Barriers CNS. 2011;8:4–9. doi: 10.1186/2045-8118-8-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fabriek BO, Zwemmer JN, Teunissen CE, Dijkstra CD, Polman CH, Laman JD, Castelijns JA. In vivo detection of myelin proteins in cervical lymph nodes of MS patients using ultrasound-guided fine-needle aspiration cytology. J Neuroimmunol. 2005;161:190–194. doi: 10.1016/j.jneuroim.2004.12.018. [DOI] [PubMed] [Google Scholar]
- Frenkel D, Puckett L, Petrovic S, Xia W, Chen G, Vega J, Dembinsky-Vaknin A, Shen J, Plante M, Burt DS, Weiner HL. A nasal proteosome adjuvant activates microglia and prevents amyloid deposition. Ann Neurol. 2008;63(5):591–601. doi: 10.1002/ana.21340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furtado GC, Marcondes MC, Latkowski JA, Tsai J, Wensky A, Lafaille JJ. Swift entry of myelin-specific T lymphocytes into the central nervous system in spontaneous autoimmune encephalomyelitis. J Immunol. 2008;181:4648–4655. doi: 10.4049/jimmunol.181.7.4648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galea I, Bechmann I, Perry VH. What is immune privilege (not)? Trends Immunol. 2007;28:12–18. doi: 10.1016/j.it.2006.11.004. [DOI] [PubMed] [Google Scholar]
- Gold R, Linington C, Lassmann H. Understanding pathogenesis and therapy of multiple sclerosis via animal models: 70 years of merits and culprits in experimental autoimmune encephalomyelitis research. Brain. 2006;129(Pt 8):1953–1971. doi: 10.1093/brain/awl075. [DOI] [PubMed] [Google Scholar]
- Goldmann J, Kwidzinski E, Brandt C, Mahlo J, Richter D, Bechmann I. T cells traffic from brain to cervical lymph nodes via the cribroid plate and the nasal mucosa. J Leukoc Biol. 2006;80:797–801. doi: 10.1189/jlb.0306176. [DOI] [PubMed] [Google Scholar]
- Goverman J, Woods A, Larson L, Weiner LP, Hood L, Zaller DM. Transgenic mice that express a myelin basic protein-specific T cell receptor develop spontaneous autoimmunity. Cell. 1993;72(4):551–560. doi: 10.1016/0092-8674(93)90074-Z. [DOI] [PubMed] [Google Scholar]
- Grant JL, Ghosn EE, Axtell RC, Herges K, Kuipers HF, Woodling NS, Andreasson K, Herzenberg LA, Steinman L. Reversal of paralysis and reduced inflammation from peripheral administration of beta-amyloid in TH1 and TH17 versions of experimental autoimmune encephalomyelitis. Sci Transl Med. 2012;4(145):145ra105. doi: 10.1126/scitranslmed.3004145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greter M, Heppner FL, Lemos MP, Odermatt BM, Goebels N, Laufer T, Noelle RJ, Becher B. Dendritic cells permit immune invasion of the CNS in an animal model of multiple sclerosis. Nat Med. 2005;11:328–334. doi: 10.1038/nm1197. [DOI] [PubMed] [Google Scholar]
- Hamrah P, Dana MR. Corneal antigen-presenting cells. Chem Immunol Allergy. 2007;92:58–70. doi: 10.1159/000099254. [DOI] [PubMed] [Google Scholar]
- Hartman KB, Wilson LJ, Rosenblum MG. Detecting and treating cancer with nanotechnology. Mol Diagn Ther. 2008;12(1):1–14. doi: 10.1007/BF03256264. [DOI] [PubMed] [Google Scholar]
- Hatterer E, Davoust N, Didier-Bazes M, Vuaillat C, Malcus C, Belin MF, Nataf S. How to drain without lymphatics? dendritic cells migrate from the cerebrospinal fluid to the B-cell follicles of cervical lymph nodes. Blood. 2006;107:806–812. doi: 10.1182/blood-2005-01-0154. [DOI] [PubMed] [Google Scholar]
- Hatterer E, Touret M, Belin MF, Honnorat J, Nataf S. Cerebrospinal fluid dendritic cells infiltrate the brain parenchyma and target the cervical lymph nodes under neuroinflammatory conditions. PLoS One. 2008;3:e3321. doi: 10.1371/journal.pone.0003321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawkes CA, McLaurin J. Selective targeting of perivascular macrophages for clearance of beta-amyloid in cerebral amyloid angiopathy. Proc Natl Acad Sci U S A. 2009;106:1261–1266. doi: 10.1073/pnas.0805453106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawkes CA, Hartig W, Kacza J, Schliebs R, Weller RO, Nicoll JA, Carare RO. Perivascular drainage of solutes is impaired in the ageing mouse brain and in the presence of cerebral amyloid angiopathy. Acta Neuropathol. 2011;121:431–443. doi: 10.1007/s00401-011-0801-7. [DOI] [PubMed] [Google Scholar]
- Hawkes CA, Sullivan PM, Hands S, Weller RO, Nicoll JA, Carare RO. Disruption of arterial perivascular drainage of amyloid-beta from the brains of mice expressing the human APOE epsilon4 allele. PLoS One. 2012;7:e41636. doi: 10.1371/journal.pone.0041636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hochmeister S, Zeitelhofer M, Bauer J, Nicolussi EM, Fischer MT, Heinke B, Selzer E, Lassmann H, Bradl M. After injection into the striatum, in vitro-differentiated microglia- and bone marrow-derived dendritic cells can leave the central nervous system via the blood stream. Am J Pathol. 2008;173:1669–1681. doi: 10.2353/ajpath.2008.080234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchings M, Weller RO. Anatomical relationships of the pia mater to cerebral blood vessels in man. J Neurosurg. 1986;65:316–325. doi: 10.3171/jns.1986.65.3.0316. [DOI] [PubMed] [Google Scholar]
- Iliff JJ, Wang M, Liao Y, Plogg BA, Peng W, Gundersen GA, Benveniste H, Vates GE, Deane R, Goldman SA, Nagelhus EA, Nedergaard M. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci Transl Med. 2012;15:147ra111. doi: 10.1126/scitranslmed.3003748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishibashi S, Maric D, Mou Y, Ohtani R, Ruetzler C, Hallenbeck JM. Mucosal tolerance to E-selectin promotes the survival of newly generated neuroblasts via regulatory T-cell induction after stroke in spontaneously hypertensive rats. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2009;29(3):606–620. doi: 10.1038/jcbfm.2008.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ji Q, Perchellet A, Goverman JM. Viral infection triggers central nervous system autoimmunity via activation of CD8+ T cells expressing dual TCRs. Nat Immunol. 2010;11(7):628–634. doi: 10.1038/ni.1888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ji Q, Castelli L, Goverman JM. MHC class I-restricted myelin epitopes are cross-presented by Tip-DCs that promote determinant spreading to CD8(+) T cells. Nat Immunol. 2013;14(3):254–261. doi: 10.1038/ni.2513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johanson CE, Duncan JA, Klinge PM, Brinker T, Stopa EG, Silverberg GD. Multiplicity of cerebrospinal fluid functions: new challenges in health and disease. Cerebrospinal Fluid Res. 2008;5:10. doi: 10.1186/1743-8454-5-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaminski M, Bechmann I, Pohland M, Kiwit J, Nitsch R, Glumm J. Migration of monocytes after intracerebral injection at entorhinal cortex lesion site. J Leukoc Biol. 2012;92:31–39. doi: 10.1189/jlb.0511241. [DOI] [PubMed] [Google Scholar]
- Karman J, Ling C, Sandor M, Fabry Z. Initiation of immune responses in brain is promoted by local dendritic cells. J Immunol. 2004;173:2353–2361. doi: 10.4049/jimmunol.173.4.2353. [DOI] [PubMed] [Google Scholar]
- Keijzer C, Slutter B, van der Zee R, Jiskoot W, van Eden W, Broere F. PLGA, PLGA-TMC and TMC-TPP nanoparticles differentially modulate the outcome of nasal vaccination by inducing tolerance or enhancing humoral immunity. PLoS One. 2011;6(11):e26684. doi: 10.1371/journal.pone.0026684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kida S, Pantazis A, Weller RO. CSF drains directly from the subarachnoid space into nasal lymphatics in the rat. Anatomy, histology and immunological significance. Neuropathol Appl Neurobiol. 1993;19:480–488. doi: 10.1111/j.1365-2990.1993.tb00476.x. [DOI] [PubMed] [Google Scholar]
- Kida S, Steart PV, Zhang ET, Weller RO. Perivascular cells act as scavengers in the cerebral perivascular spaces and remain distinct from pericytes, microglia and macrophages. Acta Neuropathol. 1993;85:646–652. doi: 10.1007/BF00334675. [DOI] [PubMed] [Google Scholar]
- Kooi EJ, van Horssen J, Witte ME, Amor S, Bo L, Dijkstra CD, van der Valk P, Geurts JJ. Abundant extracellular myelin in the meninges of patients with multiple sclerosis. Neuropathol Appl Neurobiol. 2009;35(3):283–295. doi: 10.1111/j.1365-2990.2008.00986.x. [DOI] [PubMed] [Google Scholar]
- Lake J, Weller RO, Phillips MJ, Needham M. Lymphocyte targeting of the brain in adoptive transfer cryolesion-EAE. J Pathol. 1999;187(2):259–265. doi: 10.1002/(SICI)1096-9896(199901)187:2<259::AID-PATH212>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- Laman JD, Weller RO. Editorial: route by which monocytes leave the brain is revealed. J Leukoc Biol. 2012;92:6–9. doi: 10.1189/jlb.0212110. [DOI] [PubMed] [Google Scholar]
- Lieberman SM, Kim JS, Corbo-Rodgers E, Kambayashi T, Maltzman JS, Behrens EM, Turka LA. Site-specific accumulation of recently activated CD4+ Foxp3+ regulatory T cells following adoptive transfer. Eur J Immunol. 2012;42(6):1429–1435. doi: 10.1002/eji.201142286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ling C, Sandor M, Fabry Z. In situ processing and distribution of intracerebrally injected OVA in the CNS. J Neuroimmunol. 2003;141:90–98. doi: 10.1016/S0165-5728(03)00249-2. [DOI] [PubMed] [Google Scholar]
- Locatelli G, Wortge S, Buch T, Ingold B, Frommer F, Sobottka B, Kruger M, Karram K, Buhlmann C, Bechmann I, Heppner FL, Waisman A, Becher B. Primary oligodendrocyte death does not elicit anti-CNS immunity. Nat Neurosci. 2012;15:543–550. doi: 10.1038/nn.3062. [DOI] [PubMed] [Google Scholar]
- Marten NW, Stohlman SA, Zhou J, Bergmann CC. Kinetics of virus-specific CD8+ -T-cell expansion and trafficking following central nervous system infection. J Virol. 2003;77(4):2775–2778. doi: 10.1128/JVI.77.4.2775-2778.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mascarell L, Saint-Lu N, Moussu H, Zimmer A, Louise A, Lone Y, Ladant D, Leclerc C, Tourdot S, Van Overtvelt L, Moingeon P. Oral macrophage-like cells play a key role in tolerance induction following sublingual immunotherapy of asthmatic mice. Mucosal Immunol. 2011;4(6):638–647. doi: 10.1038/mi.2011.28. [DOI] [PubMed] [Google Scholar]
- Mawuenyega KG, Sigurdson W, Ovod V, Munsell L, Kasten T, Morris JC, Yarasheski KE, Bateman RJ. Decreased clearance of CNS beta-amyloid in Alzheimer’s disease. Science. 2010;330:1774. doi: 10.1126/science.1197623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McMahon EJ, Bailey SL, Castenada CV, Waldner H, Miller SD. Epitope spreading initiates in the CNS in two mouse models of multiple sclerosis. Nat Med. 2005;11(3):335–339. doi: 10.1038/nm1202. [DOI] [PubMed] [Google Scholar]
- Meredith MM, Liu K, Kamphorst AO, Idoyaga J, Yamane A, Guermonprez P, Rihn S, Yao KH, Silva IT, Oliveira TY, Skokos D, Casellas R, Nussenzweig MC. Zinc finger transcription factor zDC is a negative regulator required to prevent activation of classical dendritic cells in the steady state. J Exp Med. 2012;209(9):1583–1593. doi: 10.1084/jem.20121003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohammad MG, Hassanpour M, Tsai VW, Li H, Ruitenberg MJ, Booth DW, Serrats J, Hart PH, Symonds GP, Sawchenko PE, Breit SN, Brown DA. Dendritic cells and multiple sclerosis: disease, tolerance and therapy. Int J Mol Sci. 2012;14(1):547–562. doi: 10.3390/ijms14010547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moingeon P, Mascarell L. Induction of tolerance via the sublingual route: mechanisms and applications. Clin Dev Immunol. 2012;2012:623474. doi: 10.1155/2012/623474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monteiro M, Almeida CF, Caridade M, Ribot JC, Duarte J, Agua-Doce A, Wollenberg I, Silva-Santos B, Graca L. Identification of regulatory Foxp3+ invariant NKT cells induced by TGF-beta. J Immunol. 2010;185(4):2157–2163. doi: 10.4049/jimmunol.1000359. [DOI] [PubMed] [Google Scholar]
- Muldoon LL, Varallyay P, Kraemer DF, Kiwic G, Pinkston K, Walker-Rosenfeld SL, Neuwelt EA. Trafficking of superparamagnetic iron oxide particles (Combidex) from brain to lymph nodes in the rat. Neuropathol Appl Neurobiol. 2004;30(1):70–79. doi: 10.1046/j.0305-1846.2003.00512.x. [DOI] [PubMed] [Google Scholar]
- Mutlu L, Brandt C, Kwidzinski E, Sawitzki B, Gimsa U, Mahlo J, Aktas O, Nitsch R, van Zwam M, Laman JD, Bechmann I. Tolerogenic effect of fiber tract injury: reduced EAE severity following entorhinal cortex lesion. Exp Brain Res. 2007;178:542–553. doi: 10.1007/s00221-006-0758-2. [DOI] [PubMed] [Google Scholar]
- Nance EA, Woodworth GF, Sailor KA, Shih TY, Xu Q, Swaminathan G, Xiang D, Eberhart C, Hanes J. A dense poly(ethylene glycol) coating improves penetration of large polymeric nanoparticles within brain tissue. Sci Transl Med. 2012;4(149):149ra119. doi: 10.1126/scitranslmed.3003594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navarrete-Talloni MJ, Kalkuhl A, Deschl U, Ulrich R, Kummerfeld M, Rohn K, Baumgartner W, Beineke A. Transient peripheral immune response and central nervous system leaky compartmentalization in a viral model for multiple sclerosis. Brain Pathol. 2010;20(5):890–901. doi: 10.1111/j.1750-3639.2010.00383.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicholas DS, Weller RO. The fine anatomy of the human spinal meninges. A light and scanning electron microscopy study. J Neurosurg. 1988;69:276–282. doi: 10.3171/jns.1988.69.2.0276. [DOI] [PubMed] [Google Scholar]
- Ochoa-Reparaz J, Mielcarz DW, Ditrio LE, Burroughs AR, Foureau DM, Haque-Begum S, Kasper LH. Role of gut commensal microflora in the development of experimental autoimmune encephalomyelitis. J Immunol. 2009;183(10):6041–6050. doi: 10.4049/jimmunol.0900747. [DOI] [PubMed] [Google Scholar]
- Ochoa-Reparaz J, Mielcarz DW, Ditrio LE, Burroughs AR, Begum-Haque S, Dasgupta S, Kasper DL, Kasper LH. Central nervous system demyelinating disease protection by the human commensal Bacteroides fragilis depends on polysaccharide A expression. J Immunol. 2010;185(7):4101–4108. doi: 10.4049/jimmunol.1001443. [DOI] [PubMed] [Google Scholar]
- Ochoa-Reparaz J, Mielcarz DW, Wang Y, Begum-Haque S, Dasgupta S, Kasper DL, Kasper LH. A polysaccharide from the human commensal Bacteroides fragilis protects against CNS demyelinating disease. Mucosal Immunol. 2010;3(5):487–495. doi: 10.1038/mi.2010.29. [DOI] [PubMed] [Google Scholar]
- Odoardi F, Sie C, Streyl K, Ulaganathan VK, Schlager C, Lodygin D, Heckelsmiller K, Nietfeld W, Ellwart J, Klinkert WE, Lottaz C, Nosov M, Brinkmann V, Spang R, Lehrach H, Vingron M, Wekerle H, Flugel-Koch C, Flugel A. T cells become licensed in the lung to enter the central nervous system. Nature. 2012;488(7413):675–679. doi: 10.1038/nature11337. [DOI] [PubMed] [Google Scholar]
- Oude Engberink RD, Blezer EL, Dijkstra CD, van der Pol SM, van der Toorn A, de Vries HE. Dynamics and fate of USPIO in the central nervous system in experimental autoimmune encephalomyelitis. NMR Biomed. 2010;23(9):1087–1096. doi: 10.1002/nbm.1536. [DOI] [PubMed] [Google Scholar]
- Ousman SS, Kubes P. Immune surveillance in the central nervous system. Nat Neurosci. 2012;15(8):1096–1101. doi: 10.1038/nn.3161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pachter JS, de Vries HE, Fabry Z. The blood–brain barrier and its role in immune privilege in the central nervous system. J Neuropathol Exp Neurol. 2003;62:593–604. doi: 10.1093/jnen/62.6.593. [DOI] [PubMed] [Google Scholar]
- Phillips MJ, Needham M, Weller RO. Role of cervical lymph nodes in autoimmune encephalomyelitis in the Lewis rat. J Pathol. 1997;182:457–464. doi: 10.1002/(SICI)1096-9896(199708)182:4<457::AID-PATH870>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- Planas AM, Gomez-Choco M, Urra X, Gorina R, Caballero M, Chamorro A. Brain-derived antigens in lymphoid tissue of patients with acute stroke. J Immunol. 2012;188(5):2156–2163. doi: 10.4049/jimmunol.1102289. [DOI] [PubMed] [Google Scholar]
- Ploix CC, Noor S, Crane J, Masek K, Carter W, Lo DD, Wilson EH, Carson MJ. CNS-derived CCL21 is both sufficient to drive homeostatic CD4+ T cell proliferation and necessary for efficient CD4+ T cell migration into the CNS parenchyma following Toxoplasma gondii infection. Brain Behav Immun. 2011;25(5):883–896. doi: 10.1016/j.bbi.2010.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preston SD, Steart PV, Wilkinson A, Nicoll JAR, Weller RO. Capillary and arterial amyloid angiopathy in Alzheimer’s disease: defining the perivascular route for the elimination of amyloid beta from the human brain. Neuropathol Appl Neurobiol. 2003;29:106–117. doi: 10.1046/j.1365-2990.2003.00424.x. [DOI] [PubMed] [Google Scholar]
- Prinz M, Priller J, Sisodia SS, Ransohoff RM. Heterogeneity of CNS myeloid cells and their roles in neurodegeneration. Nat Neurosci. 2011;14(10):1227–1235. doi: 10.1038/nn.2923. [DOI] [PubMed] [Google Scholar]
- Ransohoff RM. Turning over the chance card on MS susceptibility. Nat Immunol. 2010;11(7):570–572. doi: 10.1038/ni0710-570. [DOI] [PubMed] [Google Scholar]
- Ransohoff RM. Animal models of multiple sclerosis: the good, the bad and the bottom line. Nat Neurosci. 2012;15(8):1074–1077. doi: 10.1038/nn.3168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ransohoff RM, Engelhardt B. The anatomical and cellular basis of immune surveillance in the central nervous system. Nat Rev Immunol. 2012;12(9):623–635. doi: 10.1038/nri3265. [DOI] [PubMed] [Google Scholar]
- Rennels ML, Gregory TF, Blaumanis OR, Fujimoto K, Grady PA. Evidence for a ‘paravascular’ fluid circulation in the mammalian central nervous system, provided by the rapid distribution of tracer protein throughout the brain from the subarachnoid space. Brain Res. 1985;326:47–63. doi: 10.1016/0006-8993(85)91383-6. [DOI] [PubMed] [Google Scholar]
- Rynda A, Maddaloni M, Mierzejewska D, Ochoa-Reparaz J, Maslanka T, Crist K, Riccardi C, Barszczewska B, Fujihashi K, McGhee JR, Pascual DW. Low-dose tolerance is mediated by the microfold cell ligand, reovirus protein sigma1. J Immunol. 2008;180(8):5187–5200. doi: 10.4049/jimmunol.180.8.5187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samsom JN, Hauet-Broere F, Unger WW, Vanb LA, Kraal G. Early events in antigen-specific regulatory T cell induction via nasal and oral mucosa. Ann N Y Acad Sci. 2004;1029:385–389. doi: 10.1196/annals.1309.045. [DOI] [PubMed] [Google Scholar]
- Samsom JN, van Berkel LA, van Helvoort JM, Unger WW, Jansen W, Thepen T, Mebius RE, Verbeek SS, Kraal G. Fc gamma RIIB regulates nasal and oral tolerance: a role for dendritic cells. J Immunol. 2005;174(9):5279–5287. doi: 10.4049/jimmunol.174.9.5279. [DOI] [PubMed] [Google Scholar]
- Samsom JN, van der Marel AP, van Berkel LA, van Helvoort JM, Simons-Oosterhuis Y, Jansen W, Greuter M, Nelissen RL, Meeuwisse CM, Nieuwenhuis EE, Mebius RE, Kraal G. Secretory leukoprotease inhibitor in mucosal lymph node dendritic cells regulates the threshold for mucosal tolerance. J Immunol. 2007;179(10):6588–6595. doi: 10.4049/jimmunol.179.10.6588. [DOI] [PubMed] [Google Scholar]
- Schley D, Carare-Nnadi R, Please CP, Perry VH, Weller RO. Mechanisms to explain the reverse perivascular transport of solutes out of the brain. J Theor Biol. 2006;238:962–974. doi: 10.1016/j.jtbi.2005.07.005. [DOI] [PubMed] [Google Scholar]
- Schwalbe G. Der Arachnoidalraum, ein Lymphraum und sein Zusammenhang mit dem Perichoroidalraum. Zentralb Med Wiss. 1869;7:465–467. [Google Scholar]
- Serot JM, Foliguet B, Bene MC, Faure GC. Ultrastructural and immunohistological evidence for dendritic-like cells within human choroid plexus epithelium. Neuroreport. 1997;8:1995–1998. doi: 10.1097/00001756-199705260-00039. [DOI] [PubMed] [Google Scholar]
- Sun D, Tani M, Newman TA, Krivacic K, Phillips M, Chernosky A, Gill P, Wei T, Griswold KJ, Ransohoff RM, Weller RO. Role of chemokines, neuronal projections, and the blood–brain barrier in the enhancement of cerebral EAE following focal brain damage. J Neuropathol Exp Neurol. 2000;59:1031–1043. doi: 10.1093/jnen/59.12.1031. [DOI] [PubMed] [Google Scholar]
- Szentistvanyi I, Patlak CS, Ellis RA, Cserr HF. Drainage of interstitial fluid from different regions of rat brain. Am J Physiol. 1984;246:F835–F844. doi: 10.1152/ajprenal.1984.246.6.F835. [DOI] [PubMed] [Google Scholar]
- Trandem K, Anghelina D, Zhao J, Perlman S. Regulatory T cells inhibit T cell proliferation and decrease demyelination in mice chronically infected with a coronavirus. J Immunol. 2010;184(8):4391–4400. doi: 10.4049/jimmunol.0903918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tripathi BJ, Tripathi RC. Vacuolar transcellular channels as a drainage pathway for cerebrospinal fluid. J Physiol. 1974;239:195–206. doi: 10.1113/jphysiol.1974.sp010563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tschen SI, Stohlman SA, Ramakrishna C, Hinton DR, Atkinson RD, Bergmann CC. CNS viral infection diverts homing of antibody-secreting cells from lymphoid organs to the CNS. Eur J Immunol. 2006;36(3):603–612. doi: 10.1002/eji.200535123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Helvoort JM, Samsom JN, Chantry D, Jansen W, Schadee-Eestermans I, Thepen T, Mebius RE, Kraal G. Preferential expression of IgG2b in nose draining cervical lymph nodes and its putative role in mucosal tolerance induction. Allergy. 2004;59(11):1211–1218. doi: 10.1111/j.1398-9995.2004.00510.x. [DOI] [PubMed] [Google Scholar]
- van Zwam M, Huizinga R, Heijmans N, van Meurs M, Wierenga-Wolf AF, Melief M-J, Hintzen RQ, ’t Hart BA, Amor S, Boven LA, Laman JD. Surgical excision of CNS-draining lymph nodes reduces relapse severity in chronic-relapsing EAE. J Pathol. 2009a;217:543–551. doi: 10.1002/path.2476. [DOI] [PubMed] [Google Scholar]
- van Zwam M, Huizinga R, Melief MJ, Wierenga-Wolf AF, van Meurs M, Voerman JS, Biber KP, Boddeke HW, Höpken UE, Meisel C, Meisel A, Bechmann I, Hintzen RQ, ’t Hart BA, Amor S, Laman JD, Boven LA. Brain antigens in functionally distinct antigen-presenting cell populations in cervical lymph nodes in MS and EAE. J Mol Med. 2009b;87:273–286. doi: 10.1007/s00109-008-0421-4. [DOI] [PubMed] [Google Scholar]
- von Budingen HC, Kuo TC, Sirota M, van Belle CJ, Apeltsin L, Glanville J, Cree BA, Gourraud PA, Schwartzburg A, Huerta G, Telman D, Sundar PD, Casey T, Cox DR, Hauser SL. B cell exchange across the blood–brain barrier in multiple sclerosis. J Clin Investig. 2012;122(12):4533–4543. doi: 10.1172/JCI63842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang SH, Fan Y, Makidon PE, Cao Z, Baker JR. Induction of immune tolerance in mice with a novel mucosal nanoemulsion adjuvant and self-antigen. Nanomedicine (London) 2012;7(6):867–876. doi: 10.2217/nnm.11.187. [DOI] [PubMed] [Google Scholar]
- Weinstein JS, Varallyay CG, Dosa E, Gahramanov S, Hamilton B, Rooney WD, Muldoon LL, Neuwelt EA. Superparamagnetic iron oxide nanoparticles: diagnostic magnetic resonance imaging and potential therapeutic applications in neurooncology and central nervous system inflammatory pathologies, a review. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2010;30(1):15–35. doi: 10.1038/jcbfm.2009.192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weller RO. Pathology of cerebrospinal fluid and interstitial fluid of the CNS: significance for Alzheimer disease, prion disorders and multiple sclerosis. J Neuropathol Exp Neurol. 1998;57:885–894. doi: 10.1097/00005072-199810000-00001. [DOI] [PubMed] [Google Scholar]
- Weller RO. Reaction of intrathecal and epidural spaces to infection and inflammation. In: Yaksh TL, editor. Spinal drug delivery. Amsterdam: Elsevier; 1999. pp. 297–315. [Google Scholar]
- Weller RO. Microscopic morphology and histology of the human meninges. Morphologie. 2005;89:22–34. doi: 10.1016/S1286-0115(05)83235-7. [DOI] [PubMed] [Google Scholar]
- Weller RO, Subash M, Preston SD, Mazanti I, Carare RO. Perivascular drainage of amyloid-beta peptides from the brain and its failure in cerebral amyloid angiopathy and Alzheimer’s disease. Brain Pathol. 2008;18:253–266. doi: 10.1111/j.1750-3639.2008.00133.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weller RO, Djuanda E, Yow HY, Carare RO. Lymphatic drainage of the brain and the pathophysiology of neurological disease. Acta Neuropathol. 2009;117:1–14. doi: 10.1007/s00401-008-0457-0. [DOI] [PubMed] [Google Scholar]
- Weller RO, Galea I, Carare RO, Minagar A. Pathophysiology of the lymphatic drainage of the central nervous system: implications for pathogenesis and therapy of multiple sclerosis. Pathophysiol Off J Int Soc Pathophysiol/ISP. 2010;17:295–306. doi: 10.1016/j.pathophys.2009.10.007. [DOI] [PubMed] [Google Scholar]
- Willart MA, Jan de Heer H, Hammad H, Soullie T, Deswarte K, Clausen BE, Boon L, Hoogsteden HC, Lambrecht BN. The lung vascular filter as a site of immune induction for T cell responses to large embolic antigen. J Exp Med. 2009;206(12):2823–2835. doi: 10.1084/jem.20082401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolvers DA, Coenen-de Roo CJ, Mebius RE, van der Cammen MJ, Tirion F, Miltenburg AM, Kraal G. Intranasally induced immunological tolerance is determined by characteristics of the draining lymph nodes: studies with OVA and human cartilage gp-39. J Immunol. 1999;162:1994–1998. [PubMed] [Google Scholar]
- Yamazaki S, Maruyama A, Okada K, Matsumoto M, Morita A, Seya T. Dendritic cells from oral cavity induce Foxp3(+) regulatory T cells upon antigen stimulation. PLoS One. 2012;7(12):e51665. doi: 10.1371/journal.pone.0051665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yogev N, Frommer F, Lukas D, Kautz-Neu K, Karram K, Ielo D, von Stebut E, Probst HC, van den Broek M, Riethmacher D, Birnberg T, Blank T, Reizis B, Korn T, Wiendl H, Jung S, Prinz M, Kurschus FC, Waisman A. Dendritic cells ameliorate autoimmunity in the CNS by controlling the homeostasis of PD-1 receptor(+) regulatory T cells. Immunity. 2012;37(2):264–275. doi: 10.1016/j.immuni.2012.05.025. [DOI] [PubMed] [Google Scholar]
- Zhang ET, Inman CB, Weller RO. Interrelationships of the pia mater and the perivascular (Virchow-Robin) spaces in the human cerebrum. J Anat. 1990;170:111–123. [PMC free article] [PubMed] [Google Scholar]
- Zhang ET, Richards HK, Kida S, Weller RO. Directional and compartmentalised drainage of interstitial fluid and cerebrospinal fluid from the rat brain. Acta Neuropathol. 1992;83:233–239. doi: 10.1007/BF00296784. [DOI] [PubMed] [Google Scholar]
- Zhang H, Podojil JR, Luo X, Miller SD. Intrinsic and induced regulation of the age-associated onset of spontaneous experimental autoimmune encephalomyelitis. J Immunol. 2008;181(7):4638–4647. doi: 10.4049/jimmunol.181.7.4638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong Y, Wang X, Ji Q, Mao X, Tang H, Yi G, Meng K, Yang X, Zeng Q. CD4+LAP + and CD4 +CD25 +Foxp3 + regulatory T cells induced by nasal oxidized low-density lipoprotein suppress effector T cells response and attenuate atherosclerosis in ApoE-/- mice. J Clin Immunol. 2012;32(5):1104–1117. doi: 10.1007/s10875-012-9699-7. [DOI] [PubMed] [Google Scholar]
- Zwillinger H. Die Lymphbahnen des oberen Nasalschnittes und deren Beziehungen zu den perimeningealen Lymphraumen. Arch Laryngol und Rhinol. 1912;26:66–78. [Google Scholar]