

Abstract
Accumulating evidence has indicated that natural killer cells (NK cells) play an important role in immune responses generated in the liver. However, the underlying molecular basis for local immune regulation is poorly understood. Mice were intraperitoneally injected with polyinosinic-polycytidylic acid (PolyI:C) at a dose of 20 mg/kg body wt. The percentage and absolute number of NK cells in the liver were analysed with flow cytometry. LSECtin knockout mice and LSECtin cDNA plasmids were used for analyze the role of LSECtin in hepatic NK cell regulation in vivo. Here, we show that the C-type lectin LSECtin, a member of the DC-SIGN family, is a novel liver regulator for NK cells. LSECtin could bind to NK cells in a carbohydrate-dependent manner and could regulate the number of hepatic NK cells. In the NK cell-mediated acute liver injury model induced with PolyI:C, the exogenous expression of LSECtin accelerated NK cell-induced liver injury, whereas the absence of LSECtin ameliorated this condition. Our results reveal that LSECtin is a novel, liver-specific NK cell regulator that may be a target for the treatment of inflammatory diseases in the liver.
Keywords: Liver, The natural killer cells, Hepatic immunology
Abbreviations: LSECtin, liver node sinusoidal endothelial cell lectin; NK cells, natural killer cells; PolyI:C, polyinosinic-polycytidylic acid; LSECs, liver node sinusoidal endothelial cells; DC-SIGN, dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin; ALT, alanine transaminase; AST, aspartate aminotransferase; CRD, carbohydrate recognition domain
Highlights
-
•
Liver-specific C-type lectin, LSECtin, binds to NK cells.
-
•
LSECtin expression in vivo accelerates inflammatory liver injury induced by polyI:C.
-
•
Absence of LSECtin in vivo decreased the member of hepatic NK cells.
-
•
LSECtin is not only an activated T cells but also a NK cells modulator in the liver.
1. Introduction
Natural killer cells (NK cells) are large granular lymphocytes that display cytotoxic activity against most pathogens [1]. NK cells are present in the blood and the spleen, and they are enriched in the population of resident liver lymphocytes [2], [3]. Mouse liver lymphocytes contain approximately 10% NK cells, whereas rat and human liver lymphocytes contain approximately 30%–50% NK cells [4]. The signals transmitted by stimulatory and inhibitory receptors control the activity of NK cells [5], [6]. However, the mechanisms of NK cell trafficking to the liver and recruitment within the liver during inflammation are still poorly understood.
Recently, we reported a novel c-type lectin, LSECtin (liver node sinusoidal endothelial cell lectin), that is specifically expressed on LSECs and Kupffer cells. LSECtin is a member of a family comprising CD23, DC-SIGN and DC-SIGNR, possesses a typical carbohydrate recognition domain (CRD) and binds to mannose, N-acetylglucosamine (GlcNAc) and fucose in a Ca2+-dependent manner [7]. We found that LSECtin recognises activated T cells and negatively regulates T-cell receptor-mediated signalling in vitro and the hepatic T cell immune response in vivo [8], [9]. Then, we sought to determine whether LSECtin can regulate other immune cells and if it contributes to hepatic immune regulation.
Here, we show that LSECtin can recognize to NK cells in a carbohydrate-dependent manner. We observed decreased accumulation of NK cells in the liver in the absence of LSECtin. Upon treatment with the double-stranded RNA polyinosinic-polycytidylic acid (polyI:C), LSECtin expression accelerates inflammatory liver injury in vivo, with slight elevation of serum alanine transaminase (ALT)/aspartate aminotransferase (AST) levels. In contrast, the absence of LSECtin ameliorated this condition. Thus, our results reveal that LSECtin plays an important role in the local recruitment of hepatic NK cells.
2. Materials and methods
2.1. mAbs, reagents and plasmids
The mAb against human CD3 (clone 515), CD56 (clone B159) were from BD Pharmingen; the mAb against mouse CD3 (145-2C11), pan-DX5 were (clone DX5) from e-bioscience; Recombine LSECtin protein were from R&D systems (LSECtin extracellular region Ser54-Cys293). The mAb against LSECtin were prepared as described [8]. Polyinosinic–polycytidylic acid (polyI:C) sodium were from Sigma. The cDNA coding full-length mLSECtin were sub-cloned into the expression vector pcDNA3.1a (Invitrogen).
2.2. Mice treatment
Male C57 mice, 6–8 week-old, were purchased from Vitonglihua Co. (Beijing, China). LSECtin knock-out mice was backcrossed to C57BL/6 strains as describe [8]. All mice were maintained under controlled conditions (22 °C, 55% humidity, and 12 h day/night rhythm) in compliance with the regulations of animal care of China. PolyI:C was dissolved in the pyrogen-free saline at the concentration of 1 mg/ml. For in vivo stimulation of NK cells, mice were intraperitoneally injected with polyI:C at dose of 20 mg/kg body. Serum ALT/AST levels were estimated using a detection kit. The mLSECtin cDNA were delivered in vivo using a modified ‘hydrodynamic transfection method [9]. The mice were injected with full-length mLSECtin cDNA plasmid or control plasmid (1 mg/kg body weight i. v. in 2 ml of saline).
2.3. Cell preparations
Murine livers were removed and passed through a 200-gauge stainless steel mesh and then suspended in RPMI 1640 medium containing 2% FBS. After washing, the cells were re-suspended in 30% Percoll solution and were centrifuged at 500 g for 30 min at room temperature. The pellet was re-suspended in a red cell lysis (BD pharmagen). After incubation on the ice for 7 min, cells were harvested by centrifugation and washed twice in HBSS containing 5% FCS before use. Primary hepatocyte isolation was performed as previously described [10]. The hepatic NK cells were isolated from liver MNCs by using FACSAria III BD flow cytometry.
2.4. Flow cytometry
PBMCs from healthy donors were isolated using Ficoll-Hypaque following the manufacturer’s protocol. For analysis of LSECtin binding, peripheral blood mononuclear cells were incubated for 1 h on ice with 10 μg/ml LSECtin a binding buffer, washed with the same buffer, and incubated with mAb against LSECtin (10 μg/ml) for 40 min on ice, washed and incubated with FITC-conjugated goat anti-mouse IgG, and analysed on a FACScalibur. In inhibitory experiments, LSECtin was pre-incubated with the indicated concentrations of EGTA, mannose or GlcNAc.
The phenotype of the liver MNCs was analysed using mAb in conjunction with two-color immunofluorescence. To prevent nonspecific binding, respective isotype antibodies were used as controls.
2.5. Cytotoxicity assay
NK cells cytotoxicity was analysed using a standard lactate dehydrogenase release assay. Target cells used were YAC-1 cells or primary hepatocytes. The hepatic NK cells were used as effector cells. Each assay was performed in triplicate. After 4 h incubation at 37 °C, the released LDH was measured using the lactate dehydrogenase Cytotoxic assay (Sigma). Percentage of lysis was calculated as follows: 100 × (corrected experimental release-spontaneous release)/(maximum release-spontaneous release).
2.6. Statistical analysis
The Student’s t-test was used for statistical analysis. Results with a P value of less than 0.05 were considered statistically significant.
3. Results
3.1. LSECtin binds to NK cells
The liver is considered an immune organ; it has a reticuloendothelial system and contains a large member of lymphocytes [3]. LSECtin is specifically expressed on LSECs and Kupffer cells. Therefore, LSECtin might act as a cell adhesion molecule in the reticuloendothelial system. The recombine LSECtin protein was used in screening human PBMCs to identify the LSECtin binding cells by flow cytometry. Here, we found that LSECtin recognises CD3(−)CD56(+) cells. As a C-type lectin, LSECtin exhibits calcium-dependent binding of saccharides. LSECtin binds mannose and GlcNAc as well as fucose in a Ca2+-dependent manner. Therefore, we sought to determine whether LSECtin binds to NK cells through protein-glycan interaction. Actually, we found that EGTA, mannose and N-acetylglucosamine (GlcNAc) could inhibit this adhesion, which suggests that LSECtin binds NK cells though protein-glycan recognition (Fig. 1 B, C). These results indicate that the C-type lectin LSECtin can bind to NK cells.
Fig. 1.
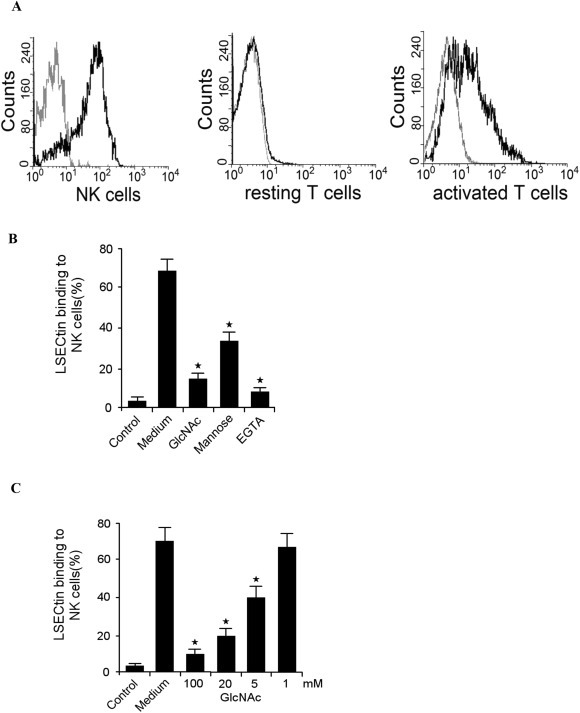
LSECtin binds to NK cells (A) Adhesion of LSECtin to NK cells. Human PBMCs were isolated and subsequently analysed for CD3 and CD56 expression using specific mAbs. LSECtin binding to activated T cells as positive control. (B) Inhibition of LSECtin binding to NK cells by D-mannose, GlcNAc and EGTA. (C) Inhibition of LSECtin binding to NK cells by GlcNAc at different concentrations. Results are the mean ± S.D. from three independent experiments. ★, p < 0.05.
3.2. LSECtin expression in vivo accelerates inflammatory liver injury induced by polyI:C
Given that LSECtin recognises NK cells, we investigated whether LSECtin regulates NK cells in the liver. PolyI:C is an artificial mimic of viral RNA, which may trigger the immune response resembling viral infection via toll-like receptors. Following PolyI:C treatment, NK cells accumulation and activation occurred in the mouse liver and results in liver damage [10], [11]. To demonstrate the effect of exogenous LSECtin expression on NK cell-induced liver injury, a gene therapy protocol was used in which full-length LSECtin or a control plasmid was injected into mice. The mice that were administered the LSECtin plasmid before treatment with polyI:C showed a higher disease severity, with increased serum ALT/AST levels (Fig. 2 A), and more serious liver histological damage was observed (Fig. 2B). Then, the phenotype of the mouse liver MNCs was analysed. The LSECtin plasmid-treated mice displayed increased numbers of NK (CD3-DX5+) cells in the liver (Fig. 2C). Furthermore, the cytotoxicity of liver NK cells against the NK cell-sensitive cell line YAC-1 or primary hepatocytes was also significantly enhanced in LSECtin plasmid-treated mice (Fig. 2D). Moreover, LSECtin plasmid treatment increased the numbers of IFN-γ-producing NK cells in liver (Fig. 2E). These results indicate that LSECtin can regulate hepatic NK cells after polyI:C-induced liver injury.
Fig. 2.
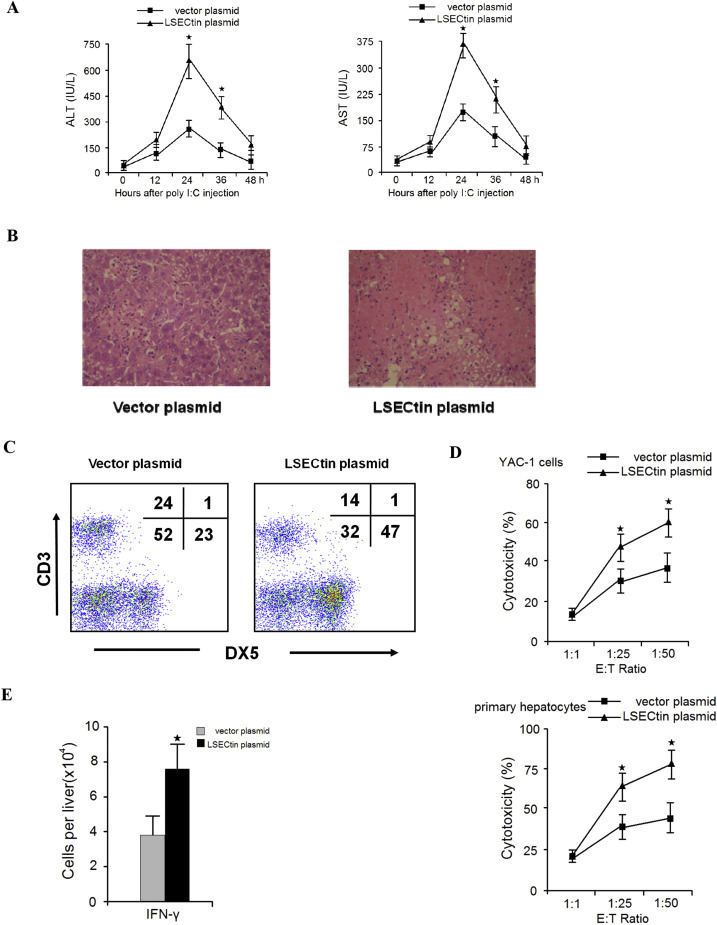
LSECtin expression accelerates inflammatory liver injury induced by polyI:C Mice were injected with control or LSECtin cDNA plasmids 24 h before polyI:C injection. (A) The plasma ALT/AST levels were measured. (B) Liver samples were prepared and stained with haematoxylin-eosin. (C) The liver MNCs were collected, and lymphocyte populations were analysed using anti-CD3 and DX-5 antibodies. (D) The cytotoxicity of the hepatic NK cells was tested against YAC-1 cells or primary hepatocytes at the indicated effector-to-target (E/T) ratios. (E) Total cell numbers of IFN-γ-producing NK cells were counted by intracellular cytokine staining. Results are the mean ± S.D. from three independent experiments. ★, p < 0.05.
3.3. An absence of LSECtin in vivo decreased the inflammatory liver injury induced by polyI:C
The expression of adhesion molecules and chemokines is crucial to the control of lymphocyte adhesion. The mechanisms that regulate the adhesion and migration of NK cells to liver in vivo were poorly understood. To determine the in vivo function of LSECtin on NK cells recruitment in the liver, the LSECtin-deficient mice were used in this study. Our results showed that LSECtin KO mice displayed normal numbers of CD3−DX5+ cells in the spleen compared with the WT mice. In contrast, a decrease in CD3−DX5+ cells (9% in wt mice vs. 5% in KO mice) was found in the liver (Fig. 3 A, B). Next, we observed that polyI:C administration caused a significant increase in liver damage in wild-type mice compared with LSECtin KO mice, with increased serum ALT/AST and more serious histological damage to the liver in the wild-type mice (Fig. 4 A, B). The number of NK cells in the liver was decreased in LSECtin KO mice compared with WT mice (Fig. 4C). The cytotoxicity of hepatic NK cells against YAC-1 or primary hepatocytes was lower in the polyI:C-treated LSECtin KO mice (Fig. 4D). We also investigated the in vivo effect of LSECtin in recruitment of NK cells by delivering LSECtin-encoding or control plasmid into the LSECtin KO mouse liver by hydrodynamic tail vein injection. In concordance with data from the normal mice, the LSECtin plasmid treated in KO mice exhibited higher number of hepatic NK cells compared to control plasmid treated mice (Fig. 4E). Combining the results from in vitro and in vivo studies, it is reasonable to conclude that LSECtin is involved in regulating NK cell recruitment in the liver.
Fig. 3.
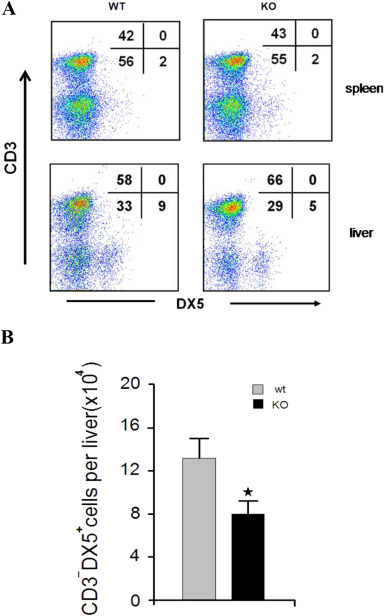
Absence of LSECtin in vivo decreased the member of hepatic NK cells (A) Lymphocytes were isolated from the spleen and liver of wt and KO mice (8–12 weeks old) and subsequently analysed for CD3 and DX5 expression using specific mAbs. (B) Total cell numbers of CD3−DX5+ cells were counted. The liver MNCs were collected, and lymphocyte populations were analysed using anti-CD3 and DX-5 antibodies. Results are the mean ± S.D. from three independent experiments. ★, p < 0.05.
Fig. 4.
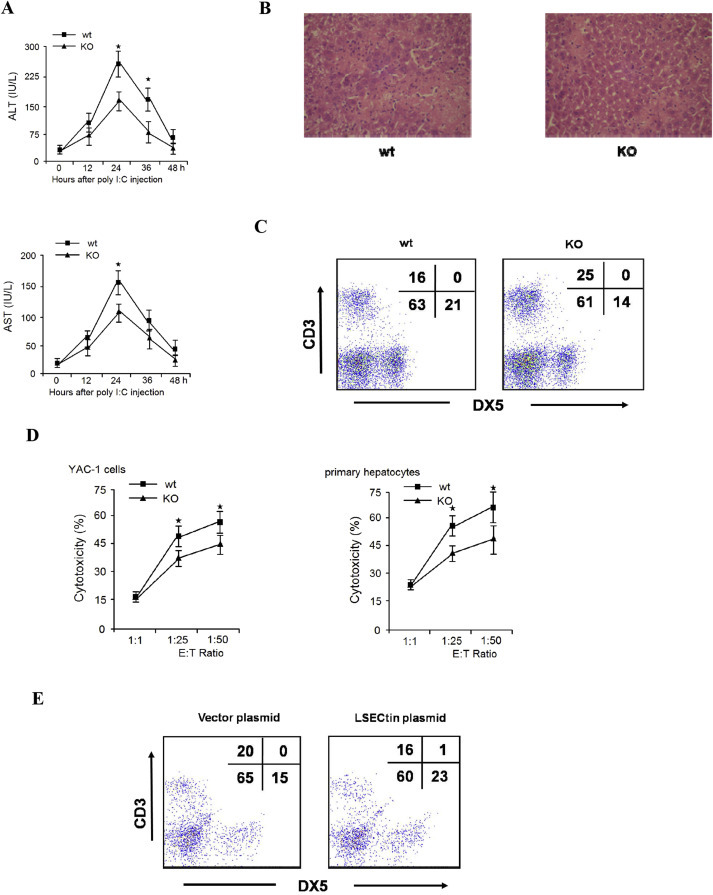
Absence of LSECtin in vivo decreased inflammatory liver injury induced by polyI:C (A) Wt or KO mice were treated with polyI:C. Plasma ALT/AST levels were measured. (B) The liver samples were collected for haematoxylin-eosin staining. (C) The liver MNCs were collected, and lymphocyte populations were analysed using anti-CD3 and DX-5 antibodies. (D) The cytotoxicity of the hepatic NK cells were tested against YAC-1 cells or primary hepatocytes at the indicated effector-to-target (E/T) ratios. (E) KO mice were treated with control or LSECtin plasmid before treated with polyI:C. The liver MNCs were collected, and lymphocyte populations were analysed using anti-CD3 and DX-5 antibodies. Results are the mean ± S.D. from three independent experiments. ★, p < 0.05.
4. Discussion
The liver is considered an immune organ, which is justified by the role of the “lymphoid liver” as a constituent of the body’s immune system [3], [12]. NK cells represent a larger percentage of the lymphoid population in liver than other organs, suggesting that NK cells express some unique function in liver diseases. NK cells play a vital role in limiting viral replication. Recently study showed that the particular populations of the NK cells are associated with the early control and clearance of HCV infection [13]. The specific defect in CD56 (dim) NK cell activation and the reduced capacity to produce anti-viral and Th1-skewing cytokines may play a role in HBV persistence [14]. NK cells are activated during acute HCV regardless of infection outcome [15]. NK cells recruitment and activation is a critical step for NK cell functioning during infection or tumor metastasis. The expression of adhesion molecules and chemokines is crucial to the control of lymphocyte adhesion. LSECs and Kupffer cells play significant roles in the regulation of hepatic NK cells [16]. LSECtin was specifically expressed in LSECs and Kupffer cells in tissue arrays with 44 normal human tissues. Its specific localization and roles in regulation of NK cell give rise to its important function in hepatic immune regulation [8]. Thus, LSECtin might be involved in the mechanisms underlying the interaction between LSECs or Kupffer cells and NK cells.
Our results show that LSECtin can bind NK cells via protein-glycan recognition. We have indicated that LSECtin recognized CD44 on activated T cells [9]. However, LSECtin binding to NK cells could not be inhibited by blocking antibody against CD44 (Figure S1). NK cells express a wide array of adhesion molecules, including some integrins, VCAM-1, ICAM-1, PSGL-1, and L-selectin [17], [18]. Recirculation of lymphocyte is regulated by differential expression of adhesion molecules under different conditions. Our results show that LSECtin is involved in hepatic NK cell-mediated immunity. However, LSECtin is not the sole element contributing to hepatic NK cell recruitment since DX5+ cells can still be found in the liver in the absence of LSECtin. It is highly likely that hepatic NK cells regulation is multifactorial. So, future studies should be aimed at this issue: identifying the receptor on NK cells and its involvement in LSECtin-mediated immune regulation.
The liver possesses a strong innate immune system [19], [20]. Hepatic NK cells are critical effectors of the liver innate immune system and mediate a rapid first line of defence against most pathogens and cancer cells [21], [22]. In particular, interaction of LSECtin with the surface glycoproteins of severe acute respiratory syndrome (SARS) coronavirus and Ebola virus has been described, and LSECtin-mediated infection of cells by Ebola virus has been demonstrated [23], [24], [25]. Under disease conditions the molecules involved are relatively regulation and result in selective recruitment of NK cells. Our recently study showed that the expression of LSECtin in liver tissue from patients with various liver diseases was different. The expression of LSECtin in hepatocellular carcinoma tissue is lower. The expression of LSECtin in chronic active hepatitis was higher. Some research proved that the NK cells promote chronic infection by limiting CD8+ T-cell immunity during virus infection [26]. In addition, LSECtin is a novel regulator of T cells and expose a crucial mechanism for hepatic T-cell immune suppression. In this study, we demonstrate that LSECtin is an important regulator for modulating hepatic NK cells and involves the innate immune responses. So LSECtin is not only an activated T cells but also a NK cells modulator in the liver. This hepatic immune regulation mechanism may provide a target for the treatment of chronic inflammatory diseases or the therapeutic manipulation of tumours in the liver.
Conflict of interest
The authors have declared that no competing interests exist.
Acknowledgments
The study was supported partially by the Chinese National Natural Science Foundation Projects (30730050, 31000405); National Program on Key Basic Research Project of China (2006CB910800, 2009CB522506, 2010CB911902, 2015CB910700); National High-tech R&D Program of China (2012AA020201); National Science and Technology Major Project of the Ministry of Science and Technology of China (2009ZX09103-631, 2013ZX10002009-001, 2013YQ140405).
Footnotes
Transparency document related to this article can be found online at http://dx.doi.org/10.1016/j.bbrc.2016.05.072.
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.bbrc.2016.05.072.
Transparency document
Appendix A. Supplementary data
The following is the supplementary data related to this article:
Figure S1. The binding of LSECtin to NK cells could not inhibit by mAbs against CD44. Results are the mean ± S.D. from three independent experiments.
References
- 1.Moretta L., Bottino C., Pende D., Mingari M.C., Biassoni R., Moretta A. Human natural killer cells: their origin, receptors and function. Eur. J. Immunol. 2002;32:1205–1211. doi: 10.1002/1521-4141(200205)32:5<1205::AID-IMMU1205>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 2.Vivier E., Tomasello E., Baratin M., Walzer T., Ugolini S. Functions of natural killer cells. Nat. Immunol. 2008;9:503–510. doi: 10.1038/ni1582. [DOI] [PubMed] [Google Scholar]
- 3.Racanelli V., Rehermann B. The liver as an immunological organ. Hepatology. 2006;431:S54–S62. doi: 10.1002/hep.21060. [DOI] [PubMed] [Google Scholar]
- 4.Gao B., Jeong W., Tian Z. Liver: an organ with predominant innate immunity. Hepatology. 2008;47:729–736. doi: 10.1002/hep.22034. [DOI] [PubMed] [Google Scholar]
- 5.Held W. NK cell education: licensing, arming, disarming, tuning? Which model? Tolerance and reactivity of NK cells: two sides of the same coin? Eur. J. Immunol. 2008;38:2930–2933. doi: 10.1002/eji.200838755. [DOI] [PubMed] [Google Scholar]
- 6.Lanier L.L. Up on the tightrope: natural killer cell activation and inhibition. Nat. Immunol. 2008;9:495–502. doi: 10.1038/ni1581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Liu W.L., Tang L., Zhang G., Wei H.D., Cui Y.F., Guo L.H., Gou Z.K., Chen X.X., Jiang D.F., Zhu Y.P., Kang G.F., He F.C. Characterization of a novel C-type lectin-like gene, LSECtin - demonstration of carbohydrate node. J. Biol. Chem. 2004;279:18748–18758. doi: 10.1074/jbc.M311227200. [DOI] [PubMed] [Google Scholar]
- 8.Tang L., Yang J., Liu W., Tang X., Chen J., Zhao D., Wang M., Xu F., Lu Y., Liu B., Sun Q., Zhang L., He F. Liver sinusoidal endothelial cell lectin, LSECtin, negatively regulates hepatic t-cell immune response. Gastroenterology. 2009;137:1498–1508. doi: 10.1053/j.gastro.2009.07.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tang L., Yang J., Tang X., Ying W., Qian X., He F. The DC-SIGN family member LSECtin is a novel ligand of CD44 on activated T cells. Eur. J. Immunol. 2010;40:1185–1191. doi: 10.1002/eji.200939936. [DOI] [PubMed] [Google Scholar]
- 10.Dong Z.J., Wei H.M., Sun R., Hu Z.Q., Gao B., Tian Z.G. Involvement of natural killer cells in PolyI : C-induced liver injury. J. Hepatol. 2004;41:966–973. doi: 10.1016/j.jhep.2004.08.021. [DOI] [PubMed] [Google Scholar]
- 11.Underhill D.M. Toll-like receptors: networking for success. Eur. J. Immunol. 2003;33:1767–1775. doi: 10.1002/eji.200324037. [DOI] [PubMed] [Google Scholar]
- 12.Crispe I.N. The liver as a lymphoid organ. Annu. Rev. Immunol. 2009:147–163. doi: 10.1146/annurev.immunol.021908.132629. [DOI] [PubMed] [Google Scholar]
- 13.Alter G., Jost S., Rihn S., Reyor L.L., Nolan B.E., Ghebremichael M., Bosch R., Altfeld M., Lauer G.M. Reduced frequencies of NKp30+NKp46+, CD161+, and NKG2D+NK cells in acute HCV infection may predict viral clearance. J. Hepatol. 2011;55:278–288. doi: 10.1016/j.jhep.2010.11.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tjwa E.T.T.L., van Oord G.W., Hegmans J.P., Janssen H.L.A., Woltman A.M. Viral load reduction improves activation and function of natural killer cells in patients with chronic hepatitis B. J. Hepatol. 2011;54:209–218. doi: 10.1016/j.jhep.2010.07.009. [DOI] [PubMed] [Google Scholar]
- 15.Pelletier S., Drouin C., Bedard N., Khakoo S.I., Bruneau J., Shoukry N.H. Increased degranulation of natural killer cells during acute HCV correlates with the magnitude of virus-specific T cell responses. J. Hepatol. 2010;53:805–816. doi: 10.1016/j.jhep.2010.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tu Z., Bozorgzadeh A., Pierce R.H., Kurtis J., Crispe I.N., Orloff M.S. TLR-dependent cross talk between human Kupffer cells and NK cells. J. Exp. Med. 2008;205:233–244. doi: 10.1084/jem.20072195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Morris M.A., Ley K. Trafficking of natural killer cells. Curr. Mol. Med. 2004;4:431–438. doi: 10.2174/1566524043360609. [DOI] [PubMed] [Google Scholar]
- 18.Robertson M.J. Role of chemokines in the biology of natural killer cells. J. Leukoc. Biol. 2002;71:173–183. [PubMed] [Google Scholar]
- 19.Gao B., Radaeva S., Park O. Liver natural killer and natural killer T cells: immunobiology and emerging roles in liver diseases. J. Leukoc. Biol. 2009;86:513–528. doi: 10.1189/jlb.0309135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schwabe R.F., Seki E., Brenner D.A. Toll-like receptor signaling in the liver. Gastroenterology. 2006;130:1886–1900. [Google Scholar]
- 21.Mondelli M.U., Varchetta S., Oliviero B. Natural killer cells in viral hepatitis: facts and controversies. Eur. J. Clin. Investig. 2010;40:851–863. doi: 10.1111/j.1365-2362.2010.02332.x. [DOI] [PubMed] [Google Scholar]
- 22.Newman K.C., Riley E.M. Whatever turns you on: accessory-cell-dependent activation of NK cells by pathogens. Nat. Rev. Immunol. 2007;7:279–291. doi: 10.1038/nri2057. [DOI] [PubMed] [Google Scholar]
- 23.Pipirou Z., Powlesland A.S., Steffen I., Poehlmann S., Taylor M.E., Drickamer K. Mouse LSECtin as a model for a human Ebola virus receptor. Glycobiology. 2011;21:806–812. doi: 10.1093/glycob/cwr008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Powlesland A.S., Fisch T., Taylor M.E., Smith D.F., Tissot B., Dell A., Poehlmann S., Drickamer K. A novel mechanism for LSECtin binding to Ebola virus surface glycoprotein through truncated glycans. J. Biol. Chem. 2008;283:593–602. doi: 10.1074/jbc.M706292200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gramberg T., Hofmann H., Moller P., Lalor P.F., Marzi A., Geier M., Krumbiegel M., Winkler T., Kirchhoff F., Adams D.H., Becker S., Munch J., Pohlmann S. LSECtin interacts with filovirus glycoproteins and the spike protein of SARS coronavirus. Virology. 2005;340:224–236. doi: 10.1016/j.virol.2005.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lang P.A., Lang K.S., Xu H.C., Grusdat M., Parish I.A., Recher M., Elford A.R., Dhanji S., Shaabani N., Tran C.W., Dissanayake D., Rahbar R., Ghazarian M., Bruestle A., Fine J., Chen P., Weaver C.T., Klose C., Diefenbach A., Haeussinger D., Carlyle J.R., Kaech S.M., Mak T.W., Ohashi P.S. Natural killer cell activation enhances immune pathology and promotes chronic infection by limiting CD8(+) T-cell immunity. P Natl. Acad. Sci. U. S. A. 2012;109:1210–1215. doi: 10.1073/pnas.1118834109. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. The binding of LSECtin to NK cells could not inhibit by mAbs against CD44. Results are the mean ± S.D. from three independent experiments.