

Abstract
Paramyxovirus might adopt a molecular mechanism of membrane fusion similar to that of other class I viruses in which the heptad repeat (HR) regions of fusion protein (F) HR1 and HR2 form a six-helix bundle structure inducing membrane fusion. In this study, we examined the structure and function of HR1 and HR2 from the avian paramyxovirus-2 (APMV-2) F protein. The study showed that APMV-2 HR1 and HR2 formed a stable six-helix bundle. Only a soluble APMV-2 HR2 peptide showed potent and specific virus-cell fusion inhibition activity. Cross-inhibiting activity with APMV-1 (Newcastle disease virus, NDV) was not found. A possible mechanism of membrane fusion inhibition by the paramyxovirus HR2 peptide is discussed.
Keywords: Paramyxovirus, Heptad repeat, Six-helix bundle, Cross-inhibiting activity
Membrane fusion between the virus envelope and host cells is the first step of the enveloped virus’s entry into the host cells. This process involves the interaction of viral envelope glycoproteins and their cellular receptors (proteins or sialic aids), which leads to the conformational changes of the envelope glycoproteins [1]. The viral envelope contains two important glycoproteins, including attachment glycoprotein and fusion glycoprotein, which have two highly conserved heptad repeat (HR) 1 HR1 and HR2 regions in class I viral envelope proteins. When membrane fusion occurs, HR1 and HR2 can form a six-helix bundle structure. Synthetic peptides corresponding to HR1 or HR2 regions exhibit the ability to inhibit virus infection [2]. These are shown in many enveloped viruses, including human immunodeficiency virus (HIV) and Avian sarcoma and leucosis virus (ASLV) of Retroviridae [3], [4], [5], possibly human cytomegalovirus (CMV) and Bovine herpesvirus-1 (BoHv-1) of Herpesviridae [6], [7], severe acute respiratory syndrome cononavirus (SARS) of Coronaviridae [8], [9], Ebola virus of Filoviridae [10], and paramyxoviruses.
Paramyxoviridae family viruses are important human and animal respiratory tract pathogens [11]. Paramyxovirus contains two envelope glycoproteins, hemagglutinin-neuraminidase (HN) and fusion protein (F). HN is involved in the virus attachment to the sialic acid-containing host-cell surface receptor. The current favored mechanism [12], [13], [14] involves binding sialates to HN. This alters its binding conformation so that it can activate the F protein to undergo three conformation states including native state, pre-hairpin intermediate state, and post-fusion hairpin state. In the process, the F protein HR1 and HR2 tend to interact with each other to form a stable six-helix bundle, and then to pull the cellular and viral membranes closer to mediate the membrane fusion. The added synthetic HR1 or HR2, or both, of paramyxoviruses would prevent the formation of a six-helix bundle by binding to the respective partner in the F protein intermediate conformational state. Consensus fusion mechanism has been proposed for some paramyxoviruses, such as Avian paramyxovirus-1 (APMV-1) (popular nomenclature is Newcastle disease virus, NDV) [15], Sendai virus (SeV) [16], [17], Mumps virus (MuV) [18], Simian virus 5 (SV5) [19], Measles virus (MeV) [20], and Respiratory syncytial virus (RSV) [21].
Avian paramyxovirus-2 (APMV-2) infects a wide variety of avian species, thus posing a potential threat to the animal industry. APMV-2 contains HN and F glycoproteins, the inactive precursor of APMV-2, F0 (55.75 kDa), which is cleaved by the enzyme isolating F1 and F2 subunits [22], [23]. APMV-1 (NDV) is a model virus for the study of viral fusion mechanism, which together with APMV-2 is a member of the genus Rubulavirus in the family Paramyxoviridae. No correlative report exists, however, regarding APMV-2 membrane fusion. In this paper, the HR1 and HR2 regions of APMV-2 F glycoproteins were predicted by the computer programs LearnCoil-VMF and ExPASy-Coils, in which HR1 and HR2 were synthesized and studied to determine a series of structure and functions. To examine the ability of the HR1 and HR2 peptides to form a stable six-helix bundle, we performed circular dichroism (CD) and gel-filtration experiments. A cell fusion inhibition assay and a cross-inhibition assay were designed to examine the specificity and susceptibility of APMV-1 and APMV-2 Env-mediated fusion to inhibition by homologous or heterologous HR peptides. Certain conflicts exist concerning the inhibition mechanism of viral–cell membrane fusion, and this paper discusses the novel possible mechanism based on the structure and function of paramyxovirus HR peptides.
Materials and methods
Gene construction
In this study, fusion protein (F) from the APMV-2 strain Yucaipa (GenBank Accession No. D13977) was used. We chose the LearnCoil-VMF(http://nightingale.lcs.mit.edu) and ExPASy-Coils (http://www.ch.embnet.org/software/COILS) programs, as they were designed specifically to identify viral heptad repeat coiled coils and have been used successfully for a number of virus fusion proteins. As shown in Fig. 1 , the programs indeed predicted the F1 protein HR1 and HR2 regions, which were quite similar to each other, but sequences predicted by LearnCoil-VMF were longer than those predicted by the ExPASy-Coils programs. The actual HR1 and HR2 sequences chosen in this paper were somewhat longer than those predicted by LearnCoil-VMF. For HR1 and HR2 gene construction, the HR1 region used was derived from amino acids 124 to 170, which gave rise to seven additional amino acids. The HR2 region used was derived from amino acids 443 to 474, which gave rise to one additional amino acid in comparison with the LearnCoil-VMF prediction. The HR1 and HR2 genes, respectively, were constructed by PCR and cloned into the GST fusion expression vector pGEX-6P-1BamHI–XhoI restriction sites, in which there is a rhinovirus 3C protease cleavage site for the fusion protein (the same as the commercial PreScission protease cleavage site). The positive plasmids were verified by direct DNA sequencing.
Fig. 1.
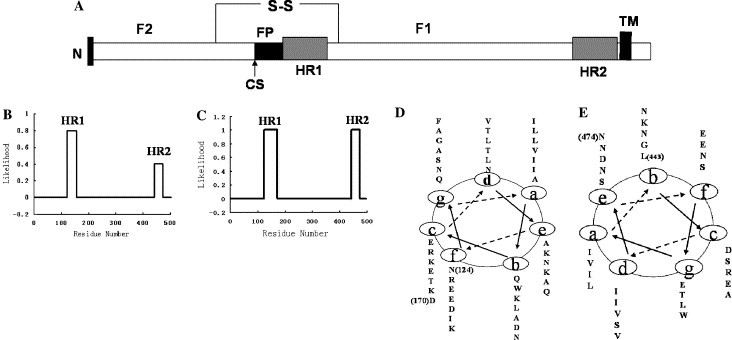
Heptad repeat regions of the APMV-2 F protein. (A) Schematic diagram of the APMV-2 F protein with the location of structurally significant domains. “S-S” represents the disulfide bond linking the F1 and F2; CS, cleavage site; FP, fusion peptide; HR, heptad repeat; and TM, transmembrane region. (B) HR1 and HR2 sequences were predicted by the ExPASy-Coils program. (C) HR1 and HR2 sequences were predicted by the LearnCoil-VMF program. (D) Helical wheel of the HR1 (amino acids 124–170) is depicted. (E) Helical wheel of the HR2 (amino acids 443–474) is depicted.
Protein expression and purification
Escherichia coli strain BL21 (DE3) transformed with the recombinant pGEX-6p-1 plasmid was grown at 37 °C in 2× YTA to an optical density of 0.8–1.0 (OD at 590 nm) before induction with 1 mM IPTG for 4 h. Bacterial cells were harvested and lysed by sonication in phosphate-buffered saline (PBS, 10 mM sodium phosphate, pH 7.3; 150 mM NaCl). Triton X-100 was then added to a final concentration of 1%, and the lysate was incubated for 30 min at 0 °C and subsequently clarified by centrifugation at 12,000g for 30 min at 4 °C. The clarified supernatants were passed over glutathione–Sepharose 4B column (equalized by PBS). The GST fusion protein-bound column was washed with PBS over 10 column volumes and eluted with reduced glutathione (10 mM) for three column volumes. The GST fusion proteins were then cleaved by GST fusion rhinovirus 3C protease at 5 °C for 16 h in the cleavage buffer (50 mM Tris–HCl, pH 7.0; 150 mM NaCl; 1 mM DTT; and 1 mM EDTA, pH 8.0). The cleaved HR1 and HR2 proteins were purified by ultrafiltration and affinity filtration. The resultant protein was dialyzed against PBS and reduced to a proper concentration by ultrafiltration and stored at −70 °C for further analysis. GST fusion proteins and cleaved HR1 and HR2 proteins were analyzed on 15% SDS–PAGE.
The HR1 peptide (amino acid 137–198) and HR2 peptide (amino acid 462–502) of APMV-1 (NDV) were expressed and purified by the above-mentioned methods. HR sequences were consistent with predictions and published reports (data not shown).
CD spectroscopy
CD spectra were performed on a Jasco J-715 spectrophotometer with HR1, HR2, and mixtures of APMV-2 HR1 and HR2 proteins in PBS (10 mM sodium phosphate, pH 7.3; 150 mM NaCl). Wavelength spectra were recorded at 20 °C using a 0.1-cm path-length cuvette. Thermodynamic stability was measured at 222 nm by recording the CD signal in the temperature range of 20–85 °C with a scan rate of 1 °C/min.
Gel-filtration analysis
GST-removed APMV-2 HR1 and HR2 proteins were mixed and incubated at room temperature for 1 h. The HR1 and HR2 mixtures were then loaded onto the Superdex G100 column (Pharmacia 2*100 cm). The peak molecular weight was estimated by comparison with the protein standards running on the same column. The fractions of the peak were collected and run on Tris–tricine SDS–PAGE.
Cell fusion assay
For the fusion inhibition assay, monolayers of HeLa T4 cells were infected with APMV-1 (NDV) (strain F48E9) and APMV-2 (strain Yucaipa) at proper-diluted (100*TCID50) concentration (TCID50 of APMV-2 was 105, TCID50 of NDV was 107) to form syncytia, respectively. In the meantime, a series of dilutions of homologous HR peptides were added to the cells for 1.5 h (APMV-2) or 1 h (APMV-1) at 37 °C. For the fusion cross-inhibition assay, monolayers of HeLa T4 cells were inoculated with virus in the presence of heterologous HR peptides. The inoculum was removed and DMEM with 1% FCS was added to the substrate. The cells were scored for fusion at 48 h (APMV-2) or 24 h (APMV-1) after incubation at 37 °C in a 5% CO2 incubator. After staining with Gimsa, cell fusion was measured by polykaryons formation and recorded as the percentage of nuclei numbers in polykaryons to numbers of total nuclei.
At least five random different fields under a light microscope were counted, and the IC50 values were calculated according to the Reed–Muench method. The fusion assays offer highly sensitive direct methods for obtaining quantitative information on the fusion process. GST-HR2 and cleaved HR2 have a proximal inhibition effect as shown in APMV-1 [24], therefore GST fusion proteins were used in this paper.
Results and discussion
GST fusion protein expressed as a highly soluble protein
In this study, we predicted APMV-2 HR1 and HR2 regions by using LearnCoil-VMF and ExPASy-Coils biosoftware, which identified the highly approximate condition (Fig. 1). The GST fusion proteins were expressed as a soluble protein and the GST-removed proteins were also highly soluble under PBS buffer at concentrations of about 6 mg/ml. SDS–PAGE of the GST fusion proteins and cleaved proteins is shown in Fig. 2 ; the molecular weight (MW) matches the theoretical value. After GST-3C cleavage, APMV-1 HR1 MW was about 6.1 kDa; APMV-1 HR2 MV was about 4.8 kDa; APMV-2 HR1 MW was about 6.4 kDa; and APMV-2 HR2 MW was about 4.1 kDa, respectively (Fig. 2). MW of the shorter HR2 peptide was about 4.1 kDa, which was close to the mass spectrometer result (4.084 kDa, data not shown).
Fig. 2.
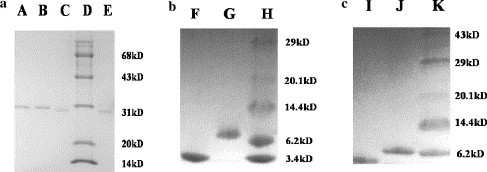
SDS–PAGE analysis of the GST fusion proteins and GST-removed proteins. (a) Lane A, Purified APMV-2 GST-HR1 protein; lane B, purified APMV-1 GST-HR1 protein; lane C, purified APMV-1 GST-HR2 protein; lane D, protein molecular weight markers (14, 20, 31, 43, and 68 kDa); lane E, purified APMV-2 GST-HR2 protein. (b) Lane F, APMV-2 HR2 after GST-3C cleavage; lane G, APMV-2 HR1 after GST-3C cleavage; and lane H, protein molecular weight markers (3.4, 6.2, 14.4, 20.1, and 29 kDa). (c) Lane I, APMV-1 HR2 after GST-3C cleavage; lane J, APMV-1 HR1 after GST-3C cleavage; lane K, protein molecular weight markers (6.2, 14.4, 20.1, 29, and 43 kDa).
Expressed GST-HR fusion proteins and cleaved HR proteins by 3C protease were passed over glutathione–Sepharose 4B column to purify. We achieved the highly purified GST-HR and HR proteins by the aforementioned method.
Cleaved APMV-2 HR1 and HR2 form a stable heterotrimeric α-helical bundle
To test the secondary structure of the APMV-2 purified peptides, CD spectrometry was carried out as described in Materials and methods. The results showed that the HR2 protein was unfolded and that the tendency of the HR1 and HR2 protein mixture towards an α-helix secondary structure was more obvious than with HR1. Thermostability results indicated that the melting temperature of the mixture of the two proteins was over 90 °C, and HR1 melted at 44 °C, which indicated that the mixture of the two proteins was extremely stable (shown in Fig. 3 ).
Fig. 3.
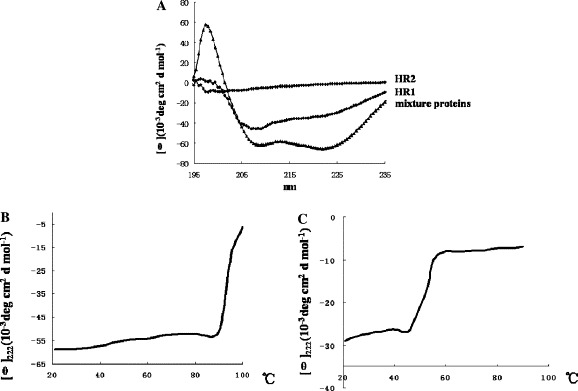
CD spectra analysis of the APMV-2HR1 and HR2 proteins. CD spectra at 25 °C and thermal denaturing curves were recorded at 222 nm with a scan rate of 1 °C/min of proteins in PBS. (A) The HR2 protein was unfolded; HR1 protein and the HR1 and HR2 protein mixtures gave a typical α-helix structure, with double minima at 208 and 222 nm. (B) The HR1 and HR2 protein mixtures showed extreme stability and a thermal denaturing temperature of over 90 °C. (C) The thermal denaturing temperature of HR1 was 44 °C.
The purified mixture of the two proteins was loaded onto Superdex G100 column (gel-filtration). A single symmetrical peak was observed on the gel-filtration and the MW was estimated to be 31 kDa. The two proteins of equivalent concentrations whose MW matched the HR1 and HR2 monomer, respectively, were shown on SDS–PAGE, which showed the formation of the heterotrimeric structure (approximately 31 kDa, about the sum of three molecular HR1 and three molecular HR2) (Fig. 4 ). The results of gel-filtration and CD showed that the APMV-2 HR1 and HR2 formed a stable six-helix bundle structure, which shared the common features of APMV-1 (NDV) [24] HR peptides and other paramyxovirus [14].
Fig. 4.
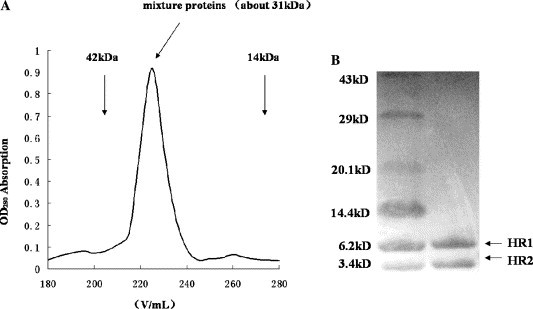
Gel-filtration analysis of the APMV-2HR1 and HR2 protein (complex) mixtures. (A) On the Superdex G100 column a clear peak was eluted between the eluted volumes corresponding to 42 and 14 kDa standards, about 31 kDa. (B) Inset picture indicates SDS–PAGE analysis of the peak, the two proteins of equivalent concentrations whose molecular weights matched the HR1 and HR2 monomer, respectively, indicating the formation of the heterotrimer structure (31 kDa is the approximate sum of three molecular HR1 and three molecular HR2).
Homologous APMV-1 and APMV-2 HR2 peptide a potent inhibitor
A fusion inhibition assay result showed that APMV-2 synthetic HR2 had APMV-2 fusion inhibition activity, and a complete syncytia reduction concentration was 16 μM; IC50 was 4.2 μM. A fusion inhibition assay result also indicated that APMV-1 (NDV) synthetic HR2 had APMV-1 fusion inhibition activity, and a complete syncytia reduction concentration was 14 μM; IC50 was 2.1 μM. These results indicated that homologous HR2 was a strong inhibitor preventing paramyxovirus-cell membrane fusion (Fig. 5 ). Indeed, clinical trials involving a number of promising agents have commenced, including the fusion inhibitor T-20 (brand name: Fuzeon). Fuzeon was recently approved by the US FDA as the first HIV entry inhibitor to be used for the treatment of AIDS patients who failed to respond to the current antiretroviral drugs, e.g., the reverse transcriptase inhibitors and protease inhibitors [25], [26]. Meanwhile, currently in medical development [20] is the mimicking of a carboxy-terminal region (HR2) paramyxovirus MeV and RSV inhibitor. The search for a membrane fusion inhibitor is an area of avid interest to a growing number of laboratories.
Fig. 5.
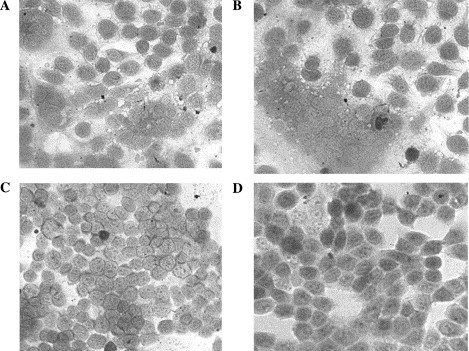
Membrane fusion and fusion inhibition tests (syncytia forming reduction). (A) Syncytia formation cells infected by APMV-2. (B) Syncytia formation cells infected by APMV-1 (NDV). (C) The homologous APMV-2 HR2 protein completely inhibited formation of syncytia cells when the HR2 concentration was added to 16 μM. (D) The homologous APMV-1 HR2 completely inhibited formation of syncytia cells when the HR2 concentration was added to 14 μM.
Homologous APMV-2 and APMV-1 HR2 a specific inhibitor
This fusion cross-inhibition assay result showed that only APMV-2 synthetic HR2 protein was a fusion inhibitor of APMV-2, and neither APMV-2 HR1 protein, APMV-1 HR1 protein, nor APMV-1 HR2 protein showed any cross-inhibition activity for APMV-2. The result also showed that only APMV-1 synthetic HR2 protein was a fusion inhibitor of APMV-1, and neither APMV-1 HR1 protein, APMV-2 HR1 protein, nor APMV-2 HR2 protein showed any cross-inhibition activity for APMV-1 (Fig. 6 ). It is becoming increasingly clear that among different members of the Paramyxoviridae, HN glycoprotein is specific [12]. There have been further reports about the antiviral activity of homologous HR, but few papers have reported examining HR specificity. For Retroviridae, HR2 peptides from human immunodeficiency virus-1 (HIV-1) and HIV-2 and Simian immunodeficiency virus (SIV) all inhibited HIV-1 Env-mediated fusion, but three HR2 peptides were poor inhibitors of HIV-2 and SIV fusion [27]. The reason [28] could be that folding into a six-helix bundle is much faster for HIV-1 than for SIV. For Herpesviridae, the HR region of Bovine herpesvirus-1 (BoHV-1) glycoprotein inhibited the Herpes simplex virus (HSV) and the Pseudorabies virus (PRV) [7]. However, the human cytomegalovirus (HCMV) HR region blocked HCMV entry, though neither affected Murine cytomegalovirus (MCMV) nor HSV entry [6]. For Paramyxoviridae, APMV-1 and APMV-2 synthetic HR2 peptides of the genus Rubulavirus had specific virus-cell fusion inhibition activity, and in this study demonstrated no cross-inhibiting activity between APMV-1 and APMV-2. To pursue the possible factors that were correlative with the virus cross-inhibiting activity of the HR peptide, we compared the peptide’s amino acid sequences between viruses. We found the following: the HR peptide amino acid identity between HIV-1 and SIV was about 50%, which peptides had partly virus cross-inhibiting activity; the HR peptide amino acid identity between BoHV-1 and HSV and PRV was over 60%, which peptides had potent virus cross-inhibiting activity; and the HR peptide amino acid identity between HCMV and MCMV and HSV was under 50% (45 and 20%, respectively), which peptides had no virus cross-inhibiting activity. We also found that the HR peptide amino acid identity between APMV-1 and APMV-2 of the genus Rubulavirus was about 40% and no virus cross-inhibitory activity was detected. These results indicate that the correlation of virus cross-inhibition HR peptide activity with sequence homology of the peptide between viruses is remarkable, in which 50% HR peptide sequence homology could be a key factor, though it has still not been determined.
Fig. 6.
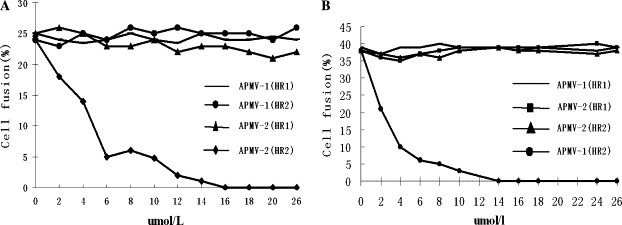
Cross-inhibition curves of cell fusion by HR protein preparations. (A) For APMV-2, only the APMV-2 synthetic HR2 peptide had specific virus-cell fusion inhibition activity, and the APMV-2 HR1 peptide had no fusion inhibition activity even if the concentration of the peptide was added to 26 μM; APMV-1 HR1 and HR2 peptides also displayed no fusion inhibition activity. (B) For APMV-1, only the APMV-1 synthetic HR2 peptide had specific virus-cell fusion inhibition activity, and the APMV-1 HR1 peptide had no fusion inhibition activity even if the concentration of the peptide was added to 26 μM; APMV-2 HR1 and HR2 peptides also displayed no fusion inhibition activity.
An unresolved issue exists regarding the mechanism of HR2 membrane fusion inhibition. The interaction assays of APMV HR1 and HR2 support the notion that the synthetic HR2 could prevent formation of the six-helix bundle structure inhibiting membrane fusion by binding to the homologous partner, which is consistent with the hypothesis introduced above [13], [19], [21], [24]. Meanwhile, studies have found that the F protein HR2 peptide is able to interact with some HN protein domains before a conformation change of the virus (including APMV-1 [29], [30], [31]) F protein. Therefore, some researchers speculate that the synthetic HR2 might block F protein activation by the HN protein, to prevent an F protein conformation state from inhibiting virus entry [30], [31], [32]. This is another possible reason for HR2 inhibition activity, although the interaction mechanism between HN and HR2 is unclear. In our study, the polykaryon formation infected by APMV-1 or APMV-2 was reduced in the presence of homologous HR2 peptides, indicating that the synthetic HR2 might have inhibition activity in the early state of APMV entry. Other reports, however, have indicated that the paramyxovirus HR2 peptide could inhibit for some time [13], [21], [24] virus-cell membrane fusion after virus-cell incubation. In a word, synthetic HR2 might prevent viral membrane fusion in the early state (before an F protein conformation change), and not just inhibit membrane fusion in the later state (by binding to HR1 in the intermediate conformational state). This study speculates that on the basis of two different mechanisms synthetic HR2 might prevent APMV entry. The paper supports and supplements a current favored mechanism and provides a basis for developing antiviral drugs.
Acknowledgment
This work was supported by The National Natural Sciences Foundation of China (NSFC) (Grant No. 30228025).
Footnotes
Abbreviations used: HR, heptad repeat; APMV-2, avian paramyxovirus-2; HIV, human immunodeficiency virus; ASLV, Avian sarcoma and leucosis virus; CMV, human cytomegalovirus; BoHv-1, Bovine herpesvirus-1; SARS, severe acute respiratory syndrome cononavirus; HN, hemagglutinin-neuraminidase; SeV, Sendai virus; MuV, Mumps virus; SV5, Simian virus 5; RSV, Respiratory syncytial virus.
References
- 1.Smith A.E., Helenius A. Science. 2004;304:237–242. doi: 10.1126/science.1094823. [DOI] [PubMed] [Google Scholar]
- 2.Colman P.M., Lawrence M.C. Nat. Rev. Mol. Cell Biol. 2003;4:309–319. doi: 10.1038/nrm1076. [DOI] [PubMed] [Google Scholar]
- 3.Chan D.C., Fass D., Berger J.M. Cell. 1997;89:263–273. doi: 10.1016/s0092-8674(00)80205-6. [DOI] [PubMed] [Google Scholar]
- 4.Egelhofer M., Brandenburg G., Martinius H. J. Virol. 2004;78:568–575. doi: 10.1128/JVI.78.2.568-575.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Markosyan R.M., Bates P., Cohen F.S. Biophys. J. 2004;87:3291–3298. doi: 10.1529/biophysj.104.047696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lopper M., Compton T. J. Virol. 2004;8:8333–8341. doi: 10.1128/JVI.78.15.8333-8341.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Okazaki K., Kida H. J. Gen. Virol. 2004;85:2131–2137. doi: 10.1099/vir.0.80051-0. [DOI] [PubMed] [Google Scholar]
- 8.Liu S., Xiao G., Chen Y. Lancet. 2004;363:938–947. doi: 10.1016/S0140-6736(04)15788-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Xu Y.H., Lou Z.Y., Gao G.F. J. Biol. Chem. 2004;279:49414–49419. doi: 10.1074/jbc.M408782200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Weissenhorn W., Carfi A., Lee K.H. Mol. Cell. 1998;2:605–616. doi: 10.1016/s1097-2765(00)80159-8. [DOI] [PubMed] [Google Scholar]
- 11.R.A. Lamb, P.L. Collins, D. Kolakofsky, Paramyxoviridae, ed., Academic Press, San Diego, 2000
- 12.Eckert D.M., Kim P.S. Annu. Rev. Biochem. 2001;70:777–810. doi: 10.1146/annurev.biochem.70.1.777. [DOI] [PubMed] [Google Scholar]
- 13.Koshiba T., Chan D.C. J. Biol. Chem. 2003;278:7573–7579. doi: 10.1074/jbc.M211154200. [DOI] [PubMed] [Google Scholar]
- 14.Morrison T.G. Acta Biochem. Biophys. 2003;1614:73–84. doi: 10.1016/s0005-2736(03)00164-0. [DOI] [PubMed] [Google Scholar]
- 15.Zhu J.Q., Li P., Gao G.F. Protein Eng. 2003;16:373–379. doi: 10.1093/protein/gzg041. [DOI] [PubMed] [Google Scholar]
- 16.Ghosh J.K., Peisajovich S.G., Shai Y. J. Biol. Chem. 1998;273:27182–27190. doi: 10.1074/jbc.273.42.27182. [DOI] [PubMed] [Google Scholar]
- 17.Wang X.J., Wang M., Gao G.F. Acta Crystallogr. D. 2004;60:1509–1512. [Google Scholar]
- 18.Liu Y., Zhu J., Gao G.F. Arch. Biochem. Biophys. 2004;421:143–148. doi: 10.1016/j.abb.2003.09.037. [DOI] [PubMed] [Google Scholar]
- 19.Russell C.J., Jardetzky T.S., Lamb R.A. EMBO J. 2001;20:4024–4034. doi: 10.1093/emboj/20.15.4024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Plemper R.K., Erlandson K.J., Lakdawala A.S. Proc. Natl. Acad. Sci. USA. 2004;101:5628–5633. doi: 10.1073/pnas.0308520101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wang E., Sun X., Qian Y. Biochem. Biophys. Res. Commun. 2003;302:469–475. doi: 10.1016/s0006-291x(03)00197-9. [DOI] [PubMed] [Google Scholar]
- 22.Mao X., Kusagawa S., Tsrudome M. Avian Dis. 1996;40:150–157. [PubMed] [Google Scholar]
- 23.Dutch R.E., Hagglund R.N., Nagel M.A. Virology. 2000;281:138–150. doi: 10.1006/viro.2000.0817. [DOI] [PubMed] [Google Scholar]
- 24.Yu M., Wang E., Liu Y. J. Gen. Virol. 2002;83:623–629. doi: 10.1099/0022-1317-83-3-623. [DOI] [PubMed] [Google Scholar]
- 25.Cilliers T., Patience T., Pillay C. AIDS Res. Hum. Retroviruses. 2004;20:477–482. doi: 10.1089/088922204323087714. [DOI] [PubMed] [Google Scholar]
- 26.Liu S., Jiang S. Curr. Pharm. Des. 2004;10:1827–1843. doi: 10.2174/1381612043384466. [DOI] [PubMed] [Google Scholar]
- 27.Gallo S.A., Sackett K., Rowat S.S. J. Mol. Biol. 2004;340:9–14. doi: 10.1016/j.jmb.2004.04.027. [DOI] [PubMed] [Google Scholar]
- 28.Marti D.N., Bjelic S., Lu M. J. Mol. Biol. 2004;336:1–8. doi: 10.1016/j.jmb.2003.11.058. [DOI] [PubMed] [Google Scholar]
- 29.Gravel K.A., Morrison T.G. J. Virol. 2003;77:11040–11049. doi: 10.1128/JVI.77.20.11040-11049.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tomasi M., Pasti C., Manfrinato C. FEBS Lett. 2003;536:56–60. doi: 10.1016/s0014-5793(03)00010-3. [DOI] [PubMed] [Google Scholar]
- 31.Yuan W., Craig S., Si Z.H. J. Virol. 2004;78:5448–5457. doi: 10.1128/JVI.78.10.5448-5457.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Li J., Quinlan E., Mirza A. J. Virol. 2004;78:5299–5310. doi: 10.1128/JVI.78.10.5299-5310.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]