

Abstract
Leucine-rich alpha-2-glycoprotein-1 (LRG) is a serum glycoprotein of unknown function that has shown promise based on qualitative assessments as a biomarker for certain diseases including microbial infections and cancer. However, the lack of a quantitative assay for LRG has limited its application. Here an indirect enzyme-linked immunosorbent assay (ELISA) for quantifying LRG in human serum is described in which cytochrome c is employed as the capturing ligand and a monoclonal antibody specific for LRG is used to detect the captured glycoprotein. Application of this assay in quantifying LRG in various patients' sera is demonstrated. The concentration of LRG in sera of control subjects as determined by this assay is approximately 50 μg/ml. Consistent with expectations from published reports, LRG was found to be significantly elevated in the sera of some patients with a bacterial infection (toxic shock syndrome, TSS). LRG was only slightly elevated in patients infected with the human immunodeficiency virus as compared to uninfected control subjects, while normal levels of LRG were observed in patients with non-infectious diseases (inflammatory arthritis and neurological disorders, primarily Parkinson's disease). Although LRG and C-reactive protein (CRP) are both produced by the liver and are classified as acute-phase proteins, there was no significant correlation between the levels of LRG and CRP in the sera of the patients. Thus, LRG and CRP measurements are non-redundant and indicate different physiological contexts. The ELISA described in this report should be useful to further assess serum LRG as a biomarker for clinical diagnostics.
Abbreviations: CRP, C-reactive protein; Cyt c, cytochrome c; HIV, human immunodeficiency virus; LRG, leucine-rich alpha-2-glycoprotein-1; mAb, monoclonal antibody; MALDI-TOF MS, matrix-assisted laser desorption ionization time-of-flight mass spectrometry; NCBI GDS, National Center for Biotechnology Information Gene Expression Omnibus data set; PD, Parkinson's disease; SARS, severe acute respiratory syndrome; TSS, toxic shock syndrome
Keywords: Biomarker, Cytochrome c, Sandwich ELISA, Leucine-rich alpha-2-glycoprotein-1, Toxic shock syndrome
1. Introduction
Evaluation of serum components is one of the least invasive approaches to clinical diagnosis and prognosis. There is considerable potential to exploit this approach even further than at present. Indeed, the impact of a number of serum proteins in disease diagnosis and prognosis is not fully known, limited in large part by the availability of rapid, quantitative assays. A case in point is the serum glycoprotein leucine-rich-alpha-2-glycoprotein-1 (LRG). LRG was first purified from human serum in 1977 (Haupt and Baudner, 1977) and its amino acid sequence was determined 8 years later (Takahashi et al., 1985). The glycoprotein has a molecular weight of approximately 50 kDa and contains 23% carbohydrate by weight (Haupt and Baudner, 1977). It is comprised of repetitive sequences with a leucine-rich motif and leucine residues make up 17% of the amino acid composition (Takahashi et al., 1985). Based on mRNA analysis, LRG is predicted to be expressed by liver cells (NCBI GDS 182, GDS 423, and GDS 2068) and by neutrophils (O'Donnell et al., 2002) and is up-regulated in a mouse model of inflammation (Norkina et al., 2004). The function of LRG is not known, although it has been shown to bind cytochrome c (Cyt c; Cummings et al., 2006) and has been hypothesized to play a role in cell survival (Thompson et al., 2007).
Increased serum levels of the glycoprotein have been qualitatively demonstrated in several groups of patients by proteomics approaches involving two-dimensional gel electrophoresis. LRG was consistently elevated by qualitative assessment in the sera of patients with bacterial infections including Haemophilis influenzae type b, Salmonella, and Streptococcus pyogenes and was often, but not always, increased during viral infections including Varicella-Zoster, measles, and mumps (Bini et al., 1996). LRG was also overexpressed along with other acute-phase proteins in patients with severe acute respiratory syndrome (SARS; Chen et al., 2004). In addition, increased serum LRG was observed in patients with several types of cancer including pancreatic (Kakisaka et al., 2007), liver (Kawakami et al., 2005), and lung cancers (Okano et al., 2006) by two-dimensional gel electrophoresis in conjunction with mass spectrometry and validation by Western blotting.
Evaluation of LRG as a serum biomarker would be enhanced by the development of a quantitative assay to measure the glycoprotein. To that end we prepared a mouse monoclonal antibody (mAb) reactive with human LRG and have used it in conjunction with Cyt c as the capturing ligand to develop an indirect enzyme-linked immunosorbent assay (ELISA) for quantifying LRG. Employing this assay we examined serum levels of LRG in patients with a variety of disorders including inflammatory arthritis, neurological disorders including Parkinson's disease (PD), toxic shock syndrome (TSS) resulting from infection with Staphylococcal aureus bacteria, and infection with human immunodeficiency virus (HIV). C-reactive protein (CRP), a prominent acute-phase protein, was also quantified and compared to LRG to determine if the two biomarkers are redundant or provide different clinical information.
2. Materials and methods
2.1. Human LRG
LRG from pooled human sera (obtained from several volunteer donors as approved by the University of Minnesota Institutional Review Board) was purified as previously described with slight modifications (Cummings et al., 2006). For adsorption on DEAE-Sephacel (40 ml; Sigma-Aldrich Chem. Co., St. Louis, MO) the supernate from 50% ammonium sulfate precipitation of serum was dialyzed against 10 mM sodium phosphate, pH 7.0. Stepwise two-fold increases in sodium chloride concentration beginning at 40 mM resulted in the elution of human LRG at 160 mM sodium chloride. The eluate containing LRG was adsorbed to 1 ml horse Cyt c-Affi-Gel 10 (Bio-Rad Laboratories, Richmond, CA), washed in phosphate-buffered saline, pH 7.4 (PBS, 3 × 10 ml), and eluted stepwise with 4 ml 0.05 M acetic acid followed by 0.5 M acetic acid. The 0.05 M acetic acid eluate which contained approximately 80% of the LRG was lyophilized, dissolved in 200 μl 10 mM sodium phosphate pH 7.0, dialyzed against 1 mM sodium phosphate pH 7.0, and passed through 200 μl of hydroxylapatite (Bio-Rad) to remove any acidic proteins that may have bound to the Cyt c-Affi-Gel 10 column due to electrostatic interactions (Haupt and Baudner, 1977). Protein purity was assessed by Gel Code Blue staining of SDS-PAGE gels (4 to 20% acrylamide gradient; Pierce Biotechnology, Rockford, IL). The concentration of LRG was determined from the absorbance at 280 nm assuming an extinction coefficient (1 mM) of 32.8 as calculated from the amino acid composition (Gill and von Hippel, 1989).
2.2. Patients' sera
Individual blood samples were obtained from patients and control subjects following protocols that were approved by the University of Minnesota Institutional Review Board. After clotting at room temperature, the blood samples were refrigerated for 1 h then centrifuged at 5000 rpm to separate sera from the clots. Sera were frozen in aliquots at − 20 °C until assayed for LRG and CRP.
2.3. Antibodies
To enhance its immunogenicity, human LRG was covalently coupled in an equimolar amount to hemocyanin using glutaraldehyde (Jemmerson, 1987). BALB/c mice were injected i.p. with 25 μg protein emulsified 1:1 in complete Freund's adjuvant and challenged 3 weeks later with the conjugated protein (25 μg) emulsified in incomplete Freund's adjuvant. The protocol for injection and surgical procedure for removing spleens from immunized mice were approved by the University of Minnesota Institutional Animal Care and Use Committee. A monoclonal antibody specific for human LRG (mAb 2F5.A2, determined to express the γ 1 and κ heavy and light chain isotypes, respectively, by indirect ELISA) was obtained by fusing splenocytes of an immunized mouse, 3 weeks after secondary challenge, with P3X63 myeloma cells using polyethylene glycol (Kearney et al., 1979). Antibody-secreting hybridomas were selected by ELISA based on the ability of the antibodies produced to bind to human LRG that was captured from serum by Cyt c on the assay plate. Selected hybridomas were subcloned in limiting dilution using normal mouse splenocytes as feeder cells.
2.4. Immunoaffinity chromatography
mAb 2F5.A2 was purified and coupled to beads that were used in immunoaffinity chromatography to adsorb LRG complexed to Cyt c. The mAb was purified from ascites fluid in two steps: precipitation in 50% saturated ammonium and flow through DEAE-Sephacel beads in 40 mM sodium phosphate, pH 8.0. The mAb (3 mg) was coupled to 300 mg cyanogen-bromide activated Sepharose 4B (Sigma-Aldrich Chem. Co.) according to the manufacturer's instructions. Cyt c and LRG were incubated in molar ratios of 4:1 and 20:1 for 2 h with continual mixing and then the complexes were adsorbed on the immunoaffinity column. From the absorbances at 280 nm and 410 nm of the proteins eluted in 0.5 M acetic acid following washing of the immunoaffinity column with 20 ml PBS, the molar ratio of Cyt c bound to LRG was determined. To detect other possible proteins bound to LRG in serum, human serum was diluted to 10% in PBS and LRG was adsorbed onto the mAb column. A similar protocol was employed to adsorb serum proteins that bind to Cyt c employing horse Cyt c coupled to Affi-Gel 10.
2.5. Mass spectrometry
MS determination of proteins in gel slices was performed in the University of Minnesota Center for Mass Spectrometry and Proteomics as previously described (Cummings et al., 2006) employing a QSTAR XL Quadrupole MALDI-TOF (Applied Biosystems, Inc., Foster City, CA.); 4800 MALDI-TOF/TOF, MALDI-TOF-TOF Analyzer (Applied Biosystems, Inc.); or Biflex III, MALDI-TOF Delayed Extraction (Bruker Daltonics, Inc., Billerica MA.). Statistical analysis was performed with Protein Pilot (v. 2.01) software (Applied Biosystems, Inc.).
2.6. Western blotting
After SDS-PAGE, protein bands were elecrophoretically transferred to a Hybond-ECL nitrocellulose membrane (Amersham Biosciences, Piscataway, NJ), blocked with 5% dry milk in PBS containing 0.1% Triton X-100, and probed for Cyt c using mAb 7H8.2C12 (Liu et al., 1996) followed by HRP-coupled goat anti-mouse immunoglobulin (Pierce Biotechnology). The blot was developed using SuperSignal West Femto substrate (Pierce Biotechnology) with a 5 s exposure to Hyperfilm ECL (Amersham Biosciences).
2.7. Indirect ELISA to quantify LRG
Binding of LRG to Cyt c was assessed employing indirect ELISA (Engvall, 1980). Since the binding of LRG to Cyt c is evolutionarily conserved (Thompson et al., 2007), horse Cyt c was substituted for human Cyt c and adsorbed to microtiter plates to capture LRG. The protein was dissolved at a concentration of 0.5 μM in PBS, pH 7.4 and 50 μl were adsorbed for 1.5 h to 96 well MaxiSorp plates (Nalge Nunc International, Rochester, N.Y.). The plates were washed twice in PBS containing 0.1% Triton X-100, then dilutions of purified human LRG as standards or human sera diluted 500-fold or greater were added to the wells for 1.5 h After two washes, mouse anti-human LRG mAb (1 μg/ml in PBS containing 0.1% Triton X-100), was added to each well and incubated for 1.5 h The plates were washed twice and incubated with 50 μl of a 1/2000 dilution of goat anti-mouse IgG (whole molecule) conjugated to horseradish peroxidase (Sigma-Aldrich Chem. Co.). After 1.5 h, the plates were washed three times and citrate-phosphate buffer, pH 5.0 containing substrate (hydrogen peroxide) and color indicator (o-phenylenediamine) was added to each well to 100 μl. The reaction was allowed to continue for 15 min and stopped by the addition of 50 μl 4 N sulfuric acid. The absorbance at 492 nm was determined employing a Titertek Multiskan plate reader (Flow Laboratories, McClean, VA.). Serum samples were assayed in triplicate. LRG concentrations were determined from averaged absorbance readings and a standard curve plotted as a sigmoid dose response equivalent to the 4 parameter logistic equation employing the Prism 4.03 program (GraphPad Software Inc., San Diego, CA). Statistical significance was also analyzed employing the Prism 4.03 program.
2.8. Quantification of CRP
A commercial kit was employed to quantify CRP (AlerCHEK, Portland, ME). Sera were diluted 10,000-fold to 500,000-fold for testing depending upon the quantity of CRP.
3. Results
3.1. ELISA for LRG
An indirect sandwich ELISA to detect and quantify human LRG was prepared employing the ligand Cyt c to capture the glycoprotein and a mAb specific for human LRG to detect it. Since human LRG binds Cyt c from many species including a variety of mammals, birds, and fishes (Cummings et al., 2006, Thompson et al., 2007), horse Cyt c was employed to capture human LRG in the assay described. A typical curve for purified LRG used as a standard in the assay to quantify LRG is shown in Fig. 1 . Following its purification from pooled human sera, LRG migrated in SDS-PAGE with the expected molecular weight of approximately 50 kDa (Fig. 1A). A typical standard curve for the ELISA is shown in Fig. 1B. Since we calculated the concentration of LRG in control serum to be approximately 50 μg/ml (see below), sera must be diluted at least 500-fold for effective quantification in this assay.
Fig. 1.
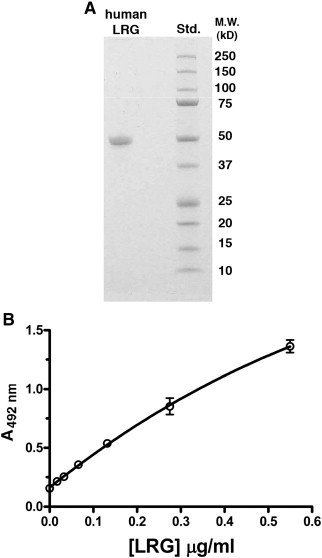
Standard curve for purified human LRG in ELISA. The purity of LRG was assessed by SDS-PAGE (A). The range of LRG concentrations quantifiable by a standard curve for LRG (B) requires that normal human sera be diluted approximately 500-fold.
3.2. Endogenous Cyt c does not interfere with the assay
Although Cyt c has been detected in normal serum, its reported concentration of 2 to 25 ng/ml (Barczyk et al., 2005, Pullerits et al., 2005) is too low, theoretically, to inhibit the Cyt c-mediated capture of LRG which is more than 1000-fold more concentrated than Cyt c in serum. Nevertheless, to assess whether Cyt c or some other component in serum might bind LRG and interfere with its quantification in the ELISA, serum (diluted to 10% in PBS) was passed through a 1 ml column of Sepharose beads to which the LRG-specific mAb was covalently bound and adsorbed proteins were eluted in 0.5 M acetic acid, then electrophoresed in SDS-PAGE. The complex of purified LRG and Cyt c, formed in vitro using purified LRG and Cyt c in a molar ratio of 4:1, was effectively adsorbed by the mAb-coupled beads (Fig. 2 A). In this experiment the molar ratio of Cyt c to LRG in the bound complex was calculated to be 0.7 based on the extinction coefficients of the two proteins at 410 nm and 280 nm. When the concentration of Cyt c relative to LRG in the incubation solution was increased to 20 fold, the molar ratio of the two components in the complex approached 1.0 (results not shown).
Fig. 2.
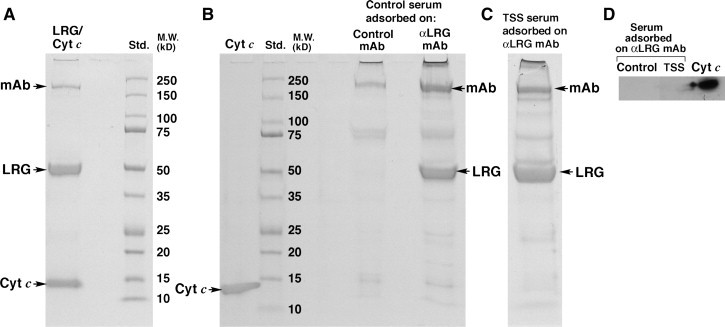
Serum LRG is not complexed to Cyt c or any other protein to any significant extent as assessed by SDS-PAGE of proteins adsorbed by and eluted from Sepharose beads to which anti-LRG mAb molecules were covalently attached. The complex of purified Cyt c and purified LRG is readily adsorbed by the mAb column (A) demonstrating the effectiveness of this approach in defining molecules that might be complexed to LRG in serum. The only molecules eluted from the column in substantial amounts following adsorption of 4.7 ml control serum (B) and 3.0 ml serum pooled from TSS patients (C) were LRG and the mAb. Cyt c was not observed. In a Western blot probed for Cyt c (2.5 ng standard) the protein was not detected when either control sera or sera pooled from TSS patients were adsorbed and eluted from the anti-LRG mAb column (D).
When proteins in normal human serum were allowed to adsorb to the same mAb-coated beads, LRG and the mAb were the dominant proteins eluted from the column as determined by Coomassie blue staining of the gel (Fig. 2B). No significant bands beyond what also eluted from control γ 1 κ mAb-coupled Sepharose beads were observed. In particular, Cyt c (migrating at 12.5 kDa) was not detected in the eluates. Adsorption of pooled sera from TSS patients with elevated LRG (see below) gave the same results as normal sera (Fig. 2C). In another experiment, in which bands from a gel similar to that in 2B and 2C were transferred to nitrocellulose and probed with a Cyt c-specific mAb using a sensitive chemiluminescent reaction for detection, we also did not observe Cyt c adsorbed from the sera of either control subjects or TSS patients, although purified Cyt c was detected (Fig. 2D, lane 3). From these experiments we conclude that LRG is not bound to Cyt c to any significant extent in the sera of the patients that were examined.
In a competitive ELISA in which increasing amounts of soluble, purified Cyt c were added to the wells prior to the addition of serum containing LRG (diluted 100-fold), inhibition was not observed below a concentration of 10− 8 M soluble Cyt c (Fig. 3 A). This represents 5-fold more than the upper limit of Cyt c estimated to be present in normal human serum (Pullerits et al., 2005). Complete inhibition of LRG detection was achieved at approximately 3 × 10− 7 M or 150-fold the upper limit of Cyt c in normal serum. This indicates that caution will be necessary when employing the assay described herein to quantify LRG in the sera of patients with exceptionally high levels of Cyt c e.g., patients with some forms of liver disease or patients undergoing a course of cancer chemotherapy (Ben-Ari et al., 2003, Barczyk et al., 2005).
Fig. 3.
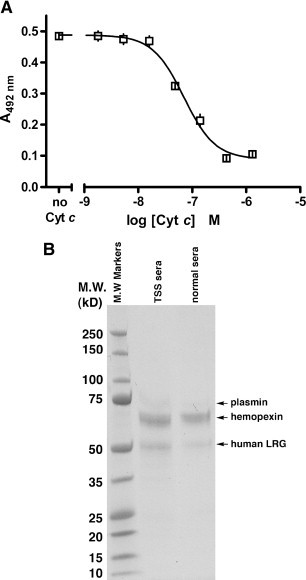
Cyt c at concentrations reported to be found in normal serum does not inhibit LRG detection in the ELISA as shown in a competitive experiment in which increasing concentrations of soluble Cyt c inhibit the binding of LRG (present in 1% serum) to plate-bound Cyt c (A). LRG is not the only serum protein adsorbed on Cyt c-Affi-gel 10 (B), but is the only Cyt c-binding serum protein that binds specifically to native Cyt c and is recognized by mAb 2F5.A2 (see text). Proteins in sera pooled from control subjects and from TSS patients (1 ml total) were adsorbed on horse Cyt c-Affi-gel and eluted, visualized by SDS-PAGE, and identified by mass spectrometry (see the text). Hemopexin binds the exposed heme in denatured Cyt c, while plasmin likely adsorbed to the affinity column due to its affinity for lysine residues.
In addition to showing that Cyt c is not co-immunoprecipitated with LRG from serum, the data in Fig. 2B and C indicate that there is no prominent serum protein that can be co-immunoprecipitated with LRG using mAb 2F5.A2. To confirm this and to identify other serum proteins that might be adsorbed by Cyt c in the ELISA for LRG, we passed 1 ml normal human serum (diluted 10-fold in or sera pooled from TSS patients through Cyt c-Affi-gel and visualized, by Gel Code Blue staining, protein bands in the eluates (100 μl serum equivalent). Proteins were identified by mass spectrometry. As shown in Fig. 3B, in addition to the band at 50 kDa expected for human LRG, a prominent band at 63 kDa was observed along with a lesser intense band with a molecular weight of approximately 70 kDa.
The 50 kDa protein was confirmed as LRG by MALDI-TOF MS (p < 0.05) and MS/MS on two individual peptides yielding Mascot Scores of 33 and 45. The 63 kDa protein was identified as hemopexin employing both MALDI-TOF delayed extraction (p < 0.0001) and MALDI-TOF/TOF in which 16 unique peptides of hemopexin were observed with 99% confidence. The 70 kDa protein was identified as plasmin employing MALDI-TOF/TOF. Twelve unique peptides with complete b and y ions were defined with 99% confidence including a peptide just two amino acids removed from the carboxyl terminus (residues 799–808) and the most amino terminus-proximal peptide, residues 127–36. Although the 12 peptides observed are also present in plasminogen, based on the size of the protein and the particular peptides that were observed extending from residues 127 through 808 we conclude that this protein is plasmin and not plasminogen.
Because plasmin has affinity for lysine (Robbins et al., 1981) and horse Cyt c contains 16% lysine residues, some plasmin molecules in the serum likely adsorbed to Cyt c-Affi-Gel 10 on this basis. We obtained human plasmin from a commercial supplier (Sigma-Aldrich Chem. Co.) and found that when employed in the ELISA described herein, binding of mAb 2F5.A2 to Cyt c-coated plates incubated with plasmin was not observed (data not shown).
Although hemopexin does not bind native Cyt c, it does bind the heme moiety exposed in chemically-cleaved fragments of the molecule (Jemmerson, 1978). Denatured molecules of Cyt c with exposed heme likely arose during pre-washing of the Cyt c-Affi-gel column with 1 M acetic acid and/or during chemical coupling of the protein to the matrix allowing for hemopexin to be adsorbed from the serum. However, when the 1–65 cyanogen-bromide-cleaved fragment of horse Cyt c, known to bind hemopexin (Jemmerson, 1978), was substituted for the native protein in the ELISA, binding of mAb 2F5.A2 was not observed. This indicates that any hemopexin that may have bound to (denatured) Cyt c in this assay was not detected using this mAb (results not shown). Furthermore, hemopexin was not immunoprecipitated from serum with mAb 2F5.A2 (Fig. 2B and C).
3.3. Quantification of LRG in sera of control subjects and several patient groups
Sera from several groups of patients (Table 1 ) were assessed for LRG employing the indirect ELISA described above. Microbial infections were represented by two groups of patients, TSS (bacterium infected) and HIV (virus infected). Two other groups of patients not previously assessed for serum LRG were patients with inflammatory arthritis and neurological disorders, including PD. Control subjects spanned the ages of most individuals in the patient groups. Both males and females were included in all groups examined.
Table 1.
Characterization of human subjects participating in this study
| Group | Gender | Age range | Disorder |
|---|---|---|---|
| Controlsa | 6 males, 11 females, 3 unknown | 24–93 median 55 | Normal |
| Arthritis patients | 10 males, 12 females | 25–77 median 45 | Rheumatoid arthritis (13)b; psoriatic arthritis (6); undifferentiated spondyloarthropathy (3) |
| Neurology patients | 13 males, 6 females, 3 unknown | 23–87 median 58 | PD or PD tremor (13); brain stem stroke (3); other (6) |
| TSS patients | 6 males, 7 females, 4 unknown | < 1–76 median age unknown | Mild TSS (1) to severe TSS (most others) |
| HIV patients | 10 males, 2 females | 26–59 median 41–42 | Early/presymptomatic (6); acute (2); AIDS (4) |
The control group for the HIV patients not listed here included 4 males, ages 28–43, confirmed as uninfected.
Numbers of subjects.
The average level of LRG in TSS patients (160 μg/ml) was significantly greater than the level in control subjects' sera (39 μg/ml) tested in the same assay (p = 0.04 by the 2-tailed t test and p < 0.001 by the Bonferonni post test after 2-way ANOVA) as shown in Fig. 4 . This result is consistent with the published assessment of serum LRG by qualitative means in several groups of patients with various bacterial infections (Bini et al., 1996). In another ELISA, serum LRG in HIV patients (38 μg/ml) was slightly higher than the level in uninfected control subjects (24 μg/ml; p = 0.01 by the 2-tailed t test), but not significantly different from the control group (p > 0.05) by the Bonferonni post test after 2-way ANOVA). Serum LRG in patients with inflammatory arthritis (Fig. 4) averaged 82 μg/ml compared to 69 μg/ml for the control subjects tested in the same assay with no statistical distinction (p = 0.13 by the 2-tailed t test and p > 0.05 by the Bonferonni post test after 2-way ANOVA). Serum LRG levels in neurology patients averaged 69 μg/ml compared to 62 μg/ml in control subjects tested in the same assay (p = 0.21 by the 2-tailed t test and p > 0.05 by the Bonferonni post test after 2-way ANOVA; Fig. 4). One PD patient had a high LRG level and was not included in the data set because it was not known whether this outlier may have resulted from a microbial infection.
Fig. 4.
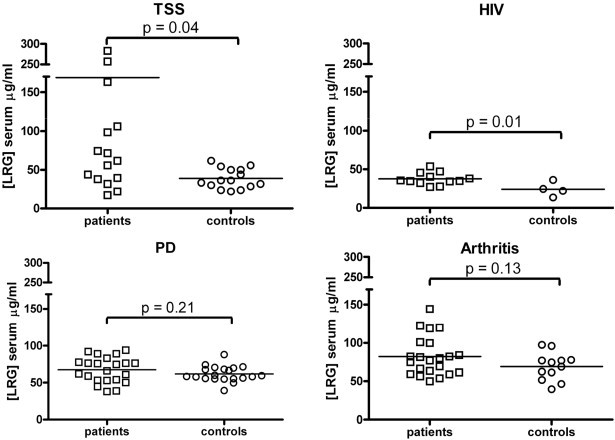
LRG levels in sera of control subjects versus several patient groups quantified by indirect ELISA. Average values are indicated by horizontal lines through the data points. P values shown are from the 2-tailed t test. For p values from Bonferonni post tests after 2-way ANOVA see the text.
The concentration of LRG in control subjects averaged across all of the tests was 54 μg/ml with a standard error of 2.7 and standard deviation of 20. The median concentration of LRG was 55 μg/ml with the 25th percentile at 36 μg/ml and the 75th percentile at 68 μg/ml. Based on the yield of LRG during its purification from pooled human serum, the concentration of LRG was previously estimated to be 21 μg/ml (Takahashi et al., 1985). The lower amount quantified in the earlier study is likely due to loss of protein during the purification process. The slight variation in the average LRG level within the same pool of control subjects among the experiments in the present study may be due to differences in the preparations of LRG employed as standards in each of the experiments.
3.4. Comparison of CRP and LRG concentrations in patients' sera
LRG and CRP are both produced by the liver in response to inflammation as acute-phase proteins so it was possible that measurement of both biomarkers could be redundant. Therefore, we also quantified CRP in several of the patient groups employing a commercial sandwich ELISA for CRP to determine if CRP and LRG provided independent measurements. As shown in Fig. 5 , two TSS patients with the highest levels of serum LRG had low to moderate levels of CRP. Note that the median level of CRP in healthy individuals is 1.5 mg/ml (Ridker, 2003). Two TSS patients with extremely high CRP levels had only slightly elevated levels of LRG and two other TSS patients with extremely high LRG levels had normal levels of CRP. While there were a few patients with above average levels of both LRG and CRP, for TSS patients the two biomarkers did not correlate (r 2 = 0.023, p = 0.56). Inflammatory arthritis patients, while not statistically different from control subjects on the basis of serum LRG, often had above median levels of CRP. Again there was no correlation between quantities of LRG and CRP in sera (r 2 = 0.023, p = 0.50). With the HIV-infected group of patients there was also no correlation (r 2 = 0.097, p = 0.42). Among all the patients tested for CRP, there was no correlation between serum LRG and CRP (r 2 = 0.002, p = 0.78).
Fig. 5.
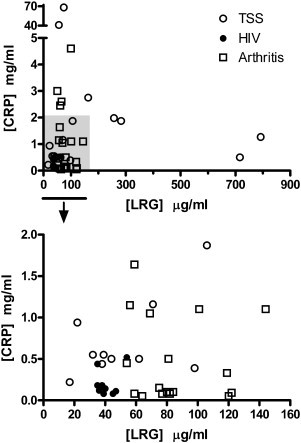
Comparison of LRG and CRP levels in sera of TSS, HIV, and arthritis patients. The shaded area in the upper graph is expanded in the lower graph. There was no correlation between the two biomarkers (r2 = 0.002, p = 0.78).
4. Discussion
In this report an indirect ELISA for human LRG was described that employs the LRG ligand, Cyt c, as the capturing agent. The usefulness of the assay in quantification of LRG in human sera was also assessed. The assay is sensitive to at least 20 ng/ml requiring that normal serum, which contains approximately 50 μg/ml LRG, be diluted 500-fold for testing. Sera containing higher concentrations of LRG may require further dilution.
Due to the extremely low concentrations of Cyt c that may be present in serum (2 to 25 ng/ml) relative to the amount of LRG (50 μg/ml) quantification of LRG in this assay is not inhibited by endogenous Cyt c. Employing an LRG-specific mAb to immunoprecipitate LRG along with bound Cyt c, we could not detect Cyt c associated with LRG in human sera using a Western blot sensitive to picogram quantities (Fig. 2). In a competitive ELISA, soluble Cyt c-mediated inhibition of LRG capture only began to be observed at a concentration 5-fold higher than the upper limit of Cyt c estimated to be present in normal serum. However, complete inhibition was observed at 150-fold higher concentration indicating that caution in interpreting the results of the ELISA described herein will be required in patients with high serum Cyt c levels. Because LRG has been shown to inhibit Cyt c detection in at least one sandwich ELISA format (Cummings et al., 2006), both LRG and Cyt c levels should be measured for optimal clinical assessment of these two biomarkers.
LRG was found to be elevated in certain patient groups and not in others. Patients with inflammatory arthritis and neurological conditions including PD have normal levels of LRG. In contrast, at least half of the patients with TSS have elevated levels of LRG. In another study to be reported elsewhere, this assay identified elevated serum LRG levels in a number of patients with ovarian cancer (Dr. Amy Skubitz, University of Minnesota, personal communication). Thus, our findings support those of several other groups who employed qualitative approaches such as two-dimensional gel electrophoresis and Western blotting to assess LRG levels in the sera of microbially-infected individuals and patients with several types of cancer (Bini et al., 1996, Chen et al., 2004, Jiang et al., 2005, Kakisaka et al., 2007, Kawakami et al., 2005, Okano et al., 2006).
Distinctions between LRG and CRP levels in several patient groups suggest that the two biomarkers serve as distinct indicators of disease even though both proteins are secreted by the liver. While CRP and LRG are both produced in response to inflammation and are classified as acute-phase proteins, low levels of LRG were observed in inflammatory arthritis patients who frequently had high levels of CRP. Two TSS patients with the highest serum levels of LRG had below average to moderate levels of CRP, while two patients with extremely high CRP levels had low serum LRG levels. There was no statistical correlation between serum LRG and CRP in the patient groups that we examined. Our findings are consistent with Kakisaka et al. who observed above normal levels of serum LRG but low levels of CRP in pancreatic cancer patients.
The relevance of elevated LRG in certain patient groups is not understood as the function of the protein is unknown. Our previous finding that LRG binds Cyt c (Cummings et al., 2006) and that LRG competes with Apaf-1 for binding Cyt c (Thompson et al., 2007) suggest that LRG may be an anti-apoptotic factor allowing survival of cells in response to stress signals. In support of this hypothesis, the LRG gene is up-regulated in lung and muscle cells within 24 to 48 h following exposure to poisons (NCBI GDS 234, GDS 1244) and then it is downregulated by 72 h LRG and Apaf-1 mRNAs are also inversely expressed in many tissues in the mouse including the liver, suggesting opposing functions (NCBI GDS 2068). In regard to cancer, LRG mRNA expression was found to be elevated in 5 of 10 ovarian cancer cell lines examined, consistent with an increase in serum LRG in at least half of ovarian cancer patients (NCBI GDS 1523). It remains to be determined whether LRG in sera of cancer patients is produced by the cancer cells or some other tissues, such as the liver or neutrophils. In their study of pancreatic cancer, Kakisaka et al. reported that cultured pancreatic cancer cells did not secrete LRG. LRG was also not observed in cells infected with the coronavirus causing SARS (Jiang et al., 2005). The elevated LRG in serum could result from the body's defense against cancer or could be a reaction of the liver to serum factors increased in response to cancer.
While the potential importance of LRG as a unique biomarker is highlighted in this study, further clinical analysis is required to determine whether quantification of serum LRG has prognostic or diagnostic value. If elevated serum LRG results from the body's defense against cancer, then high LRG levels may bode well for patients. However, if circulating LRG levels are associated with cancer cell survival, then a poor prognosis may be indicated. The sandwich ELISA described herein should be a useful tool to assess these and other questions in future clinical trials.
Acknowledgements
This study was partially funded by grants from the Minnesota Medical Foundation and the University of Minnesota Graduate School to R.J. The authors thank Drs. Ameeta Kelekar and Amy Skubitz for helpful discussions and Dr. LeeAnn Higgins for MS analysis.
References
- Barczyk K., Kreuter M., Pryjma J., Booy E.P., Maddika S., Ghavami S., Berdel W.E., Roth J., Los M. Serum cytochrome c indicates in vivo apoptosis and can serve as a prognostic marker during cancer therapy. Int. J. Cancer. 2005;116:167. doi: 10.1002/ijc.21037. [DOI] [PubMed] [Google Scholar]
- Ben-Ari Z., Schmilovotz-Weiss H., Belinki A., Pappo O., Sulkes J., Neuman M.G., Kaganovsky E., Kfir B., Tur-Kaspa R., Klein T. Circulating soluble cytochrome c in liver disease as a marker of apoptosis. J. Intern. Med. 2003;254:168. doi: 10.1046/j.1365-2796.2003.01171.x. [DOI] [PubMed] [Google Scholar]
- Bini L., Magi B., Marzocchi B., Cellesi C., Berti B., Raggiaschi R., Rossolini A., Pallini V. Two-dimensional electrophoretic patterns of acute-phase human serum proteins in the course of bacterial and viral diseases. Electrophoresis. 1996;17:612. doi: 10.1002/elps.1150170333. [DOI] [PubMed] [Google Scholar]
- Chen J.-H., Chang Y.-W., Yao C.-W., Chiueh T.-S., Huang S.-C., Chien K.-Y., Chen A., Chang F.-Y., Wong C.-H., Chen Y.-J. Plasma proteome of severe acute respiratory syndrome analyzed by two-dimensional gel electrophoresis and mass spectrometry. Proc. Natl. Acad. Sci. U. S. A. 2004;101:17039. doi: 10.1073/pnas.0407992101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cummings C., Walder J., Treeful A., Jemmerson R. Serum leucine-rich alpha-2-glycoprotein-1 binds cytochrome c and inhibits antibody detection of this apoptotic marker in enzyme-linked immunosorbent assay. Apoptosis. 2006;11:1121. doi: 10.1007/s10495-006-8159-3. [DOI] [PubMed] [Google Scholar]
- Engvall E. Enzyme immunoassay ELISA and EMIT. Methods Enzymol. 1980;70:419. doi: 10.1016/s0076-6879(80)70067-8. [DOI] [PubMed] [Google Scholar]
- Gill S.C., von Hippel P.H. Calculation of protein extinction coefficients from amino acid sequence data. Anal. Biochem. 1989;182:319. doi: 10.1016/0003-2697(89)90602-7. [DOI] [PubMed] [Google Scholar]
- Haupt H., Baudner S. Isolierung und charakterisierung eines bisher unbekannten leucinreichen 3.1 S-α2-glykoproteins aus h-umanserum. Hoppe-Seyler Z. Physiol. Chem. 1977;358:639. [PubMed] [Google Scholar]
- Jemmerson, R., 1978. The specificity of rabbit antibodies directed against horse and rabbit cytochromes c. Doctoral dissertation, Northwestern University, Evanston, IL.
- Jemmerson R. Antigenicity and native structure of globular proteins: low frequency of peptide reactive antibodies. Proc. Natl. Acad. Sci. U. S. A. 1987;84:9180. doi: 10.1073/pnas.84.24.9180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang X.-S., Tang L.-Y., Dai J., Zhou H., Li S.-J., Xia Q.-C., Wu J.-R., Zeng R. Quantitative analysis of severe acute respiratory syndrome (SARS)-associated coronavirus-infected cells using proteomic approaches: implications for cellular response to virus infection. Mol. Cell Proteomics. 2005;4:902. doi: 10.1074/mcp.M400112-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kakisaka T., Kondo T., Okano T., Fujii K., Honda K., Endo M., Tsuchida A., Aoki T., Itoi T., Moriyasu F., Yamada T., Kato H., Nishimura T., Todo S., Hirohashi S. Plasma proteomics of pancreatic cancer patients by multi-dimensional liquid chromatography and two-dimensional difference gel electrophoresis (2D-DIGE): up-regulation of leucine-rich alpha-2-glycoprotein in pancreatic cancer. J. Chromatogr., B, Biomed. Sci. Appl. 2007;852:257. doi: 10.1016/j.jchromb.2007.01.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawakami T., Hoshida Y., Kanai F., Tanaka Y., Tateishi K., Ikenoue T., Obi S., Sato S., Teratani T., Shiina S., Kawabe T., Suzuki T., Hatano N., Taniguchi H., Omata M. Proteomic analysis of sera from hepatocellular carcinoma patients after radiofrequency ablation treatment. Proteomics. 2005;5:4287. doi: 10.1002/pmic.200401287. [DOI] [PubMed] [Google Scholar]
- Kearney J.F., Radbruch A., Liesegang B., Rajewsky K. A new mouse myeloma cell line that has lost immunoglobulin expression but permits the construction of antibody-secreting hybrid cell lines. J. Immunol. 1979;123:1548. [PubMed] [Google Scholar]
- Liu X., Kim C.N., Yang J., Jemmerson R., Wang X. Induction of apoptotic program in cell-free extracts: requirement for dATP and cytochrome c. Cell. 1996;86:147. doi: 10.1016/s0092-8674(00)80085-9. [DOI] [PubMed] [Google Scholar]
- Norkina O., Kaur S., Ziemer D., De Lisle R.C. Inflammation of the cystic fibrosis mouse small intestine. Am. J. Physiol.: Gasterointest. Liver Physiol. 2004;286:G1032. doi: 10.1152/ajpgi.00473.2003. [DOI] [PubMed] [Google Scholar]
- O'Donnell L.C., Druhan L.J., Avalos B.R. Molecular characterization and expression analysis of leucine-rich α2-glycoprotein, a novel marker of granulocytic differentiation. J. Leukoc. Biol. 2002;72:478. [PubMed] [Google Scholar]
- Okano T., Kondo T., Kakisaka T., Fujii K., Yamada M., Kato H., Nishimura T., Gemma A., Kudoh S., Hirohashi S. Plasma proteomics of lung cancer by a linkage of multi-dimensional liquid chromatography and two-dimensional difference gel electrophoresis. Proteomics. 2006;6:3938. doi: 10.1002/pmic.200500883. [DOI] [PubMed] [Google Scholar]
- Pullerits R., Bokarewa M., Jonsson I.-M., Verdrengh M., Tarkowski A. Extracellular cytochrome c, a mitochondrial apoptosis-related protein, induces arthritis. Rheumatology. 2005;44:32. doi: 10.1093/rheumatology/keh406. [DOI] [PubMed] [Google Scholar]
- Ridker P.M. Clinical application of C-reactive protein for cardiovascular disease detection and prevention. Circulation. 2003;107:363. doi: 10.1161/01.cir.0000053730.47739.3c. [DOI] [PubMed] [Google Scholar]
- Robbins K.C., Summaria L., Wohl R.C. Human plasmin. Methods Enzymol. 1981;80:379. doi: 10.1016/s0076-6879(81)80032-8. [DOI] [PubMed] [Google Scholar]
- Takahashi N., Takahashi Y., Putnam F.W. Periodicity of leucine and tandem repetition of a 24-amino acid segment in the primary structure of leucine-rich α2-glycoprotein of human serum. Proc. Natl. Acad. Sci. U. S. A. 1985;82:1906. doi: 10.1073/pnas.82.7.1906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson C., Kelekar A., Jemmerson R. Keystone Symposium on Apoptosis, Abstract No. 217. 2007. Is the cytochrome c ligand, leucine-rich α2 glycoprotein-1, a survival protein? [Google Scholar]