

Abstract
The collection of microbes that live in and on the human body - the human microbiome - can impact on cancer initiation, progression, and response to therapy, including cancer immunotherapy. The mechanisms by which microbiomes impact on cancers can yield new diagnostics and treatments, but much remains unknown. The interactions between microbes, diet, host factors, drugs, and cell-cell interactions within the cancer itself likely involve intricate feedbacks, and no single component can explain all the behavior of the system. Understanding the role of host-associated microbial communities in cancer systems will require a multidisciplinary approach combining microbial ecology, immunology, cancer cell biology, and computational biology - a systems biology approach.
The Roles of Microbiomes in Cancer Are Unclear and Require More Investigation
A statistic often cited says that infectious agents cause 20% of cancers worldwide [1]. That fact perhaps explains why cancer biology tends to view microbes such as viruses (Box 1) and bacteria (Box 2) as carcinogens to be eliminated. The microbial communities that make up the human microbiome (see Glossary) impact on many aspects of host biology: nutrient uptake, drug metabolism, inflammation [2], and even behavior [3]. They most likely also influence cancer progress-sion and treatment, and in ways that can range from harmful to beneficial. Understanding how microbiomes impact on cancers - directly and indirectly, harmfully and beneficially - could bring new opportunities for cancer prevention, treatment, and management [4].
Box 1. Oncogenic Viruses and the Microbiome.
The discovery by Rous in 1911 that a virus caused sarcomas in chickens first linked a microbe to cancer [66]. The virus was present in a filtrate that Rous extracted from cancerous chicken tissue and caused cancer when injected into healthy chickens. Later, Varmus, Bishop, and colleagues expanded on this discovery to detail a genetic mechanism [67]. The first finding of a genetic origin of cancer was a viral discovery, and it set off an era of cancer molecular biology.
Viruses have had a key role in our understanding of cancer as a genetic disease, but those viruses which have already been discovered to cause cancer in humans exemplify how the interface between microbes and cancer can aid cancer prevention. Vaccines against hepatitis B virus and human papillomavirus (HPV) are remarkably successful to prevent the cancers that these viruses cause: hepatocellular carcinoma and cervical cancer, respectively [68]. Given the link between viruses and cancer, it is unfortunate that most microbiome studies have a bias for bacteria, and largely ignore viruses. Surprisingly, other microbial constituents of microbiomes also likely influence the progression of the virus-caused cancers. This is important because most people infected with oncoviruses will never develop cancer, and the factors that determine the fate of a viral infection remain unclear. In a study of Puerto Rican women with high-risk HPV infections and high cervical cancer rates, significant bacterial and fungal profile changes were associated with cervical squamous intraepithelial lesions and HPV infections (Figure I). The changes in the cervicovaginal microbiota, which resides at an interface between the host and the environment, may thus affect susceptibility to disease [69].
Viral microbiome research has challenges that warrant immediate attention: metagenomics and bioinformatics pipelines such as ViromeScan [70] and virMine [71] can find known viruses, but discovering new viruses that are not yet listed in databases will require new approaches [72]. New experimental methods may selectively detect viruses - including RNA viruses - from microbiome samples [73]. Solving such technical challenges could help us to understand better the role of bacteriophages in microbiome ecology. This is therapeutically important: bacteriophage transfer during FMT and abiotic FMT was associated with a positive outcome in patients with re-current C. difficile infection [74–76], highlighting the complex interkingdom interactions within the microbiota community.
Figure I. Cervical Cancer Comes with a Shift in the Composition of the Cervicovaginal Microbiome.
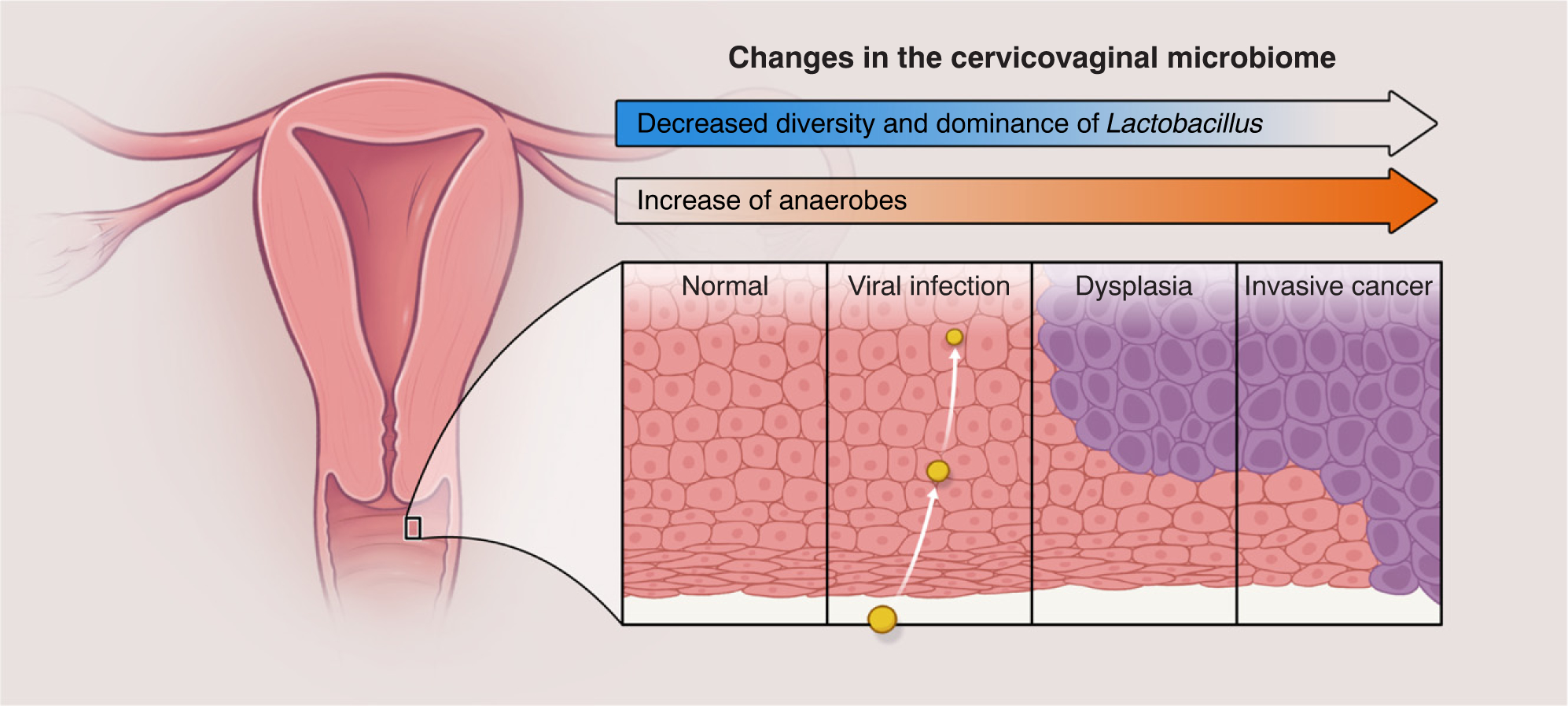
The microbiome changes from a Lactobacillus-dominated community to a community enriched in strict anaerobes such as Sneathia sanguinigens and Gardnerella vaginallis. Bacterial vaginosis - a dysbiotic phenotype - is associated with higher risk of HIV and human papillomavirus (HPV) acquisition and transmission, preterm births, and pelvic inflammatory disease and cancer.
Box 2. The Duplicitous Role of Microbes in Cancer.
The best-known bacterial cause of cancer is Helicobacter pylori, which colonizes 50% of people worldwide [77] and is responsible for ~90% of gastric cancers [1]. Stomach cancer led the list of deadly cancers for the first half of the 20th century. Its incidence has decreased in Western countries, possibly thanks to hygiene and antibiotic use, because some antibiotics taken to treat other infections kill H. pylori unintentionally [78]. However, H. pylori exemplifies a paradox: this bacterium may have coevolved with humans because it can be transmitted from person to person and could be considered as a normal component of the stomach microbiome [79]. H. pylori colonization may even benefit human health because decolonization increases the risk of severe gastroesophageal reflux disease and its sequelae, Barrett’s esophagus, and adenocarcinoma of the esophagus [80]. It may also protect against asthma, multiple sclerosis, and IBD [81–83], although it is a risk factor for type 2 diabetes and some other diseases [84,85]. Therefore, the same microbe could both harm (by causing gastric cancer and some diseases) and benefit human health (by preventing other diseases and even cancers in other organs).
This duplicity applies to other microbes [86]. Perhaps the difference between harm and benefit lies in the type of inflammation caused by the microbe. Human immunity is a subtle balance, and a measured immune response is essential for homeostasis: a weak immune response can fail to protect against infection, but an excessive response can destroy tissues or cause autoimmune disorders. IBD, a form of chronic gut inflammation, alters the composition of the luminal microbiota and enriches for Gram-negative bacteria. Patients with IBD have a higher risk of colorectal cancer, which may be due to bacteria enriched in the luminal microbiota such as pks+ E. coli [32]. However, inflammation caused by microbes can be leveraged against cancer. Bacillus Calmette-Guérin (BCG) - a vaccine prepared from an attenuated strain of Mycobacterium bovis - is one of the most successful biotherapies that can treat bladder cancer [87]. Moreover, microbiomes may affect cancers beyond the presence or absence of a specific microbe, and the simpler cases where the effect is attributed to a single organism may be exceptions more than a general rule. Understanding microbiomes in cancer may require knowledge of networks to distinguish a homeostatic network from a carcinogenic network, to understand whether those networks are static or plastic, and to devise methods to convert a carcinogenic network back into a homeostatic network.
Many knowledge gaps remain: microbiomes are complex ecosystems with spatiotemporal dynamics that emerge from interactions between microbes and with the cells of their host. Each cancer is itself an ecosystem in which cancer cells interact with each other and with stromal cells in intricate, dynamic ways. Common factors such as circulating metabolites, systemic immunity, etc. can impact on - and be affected by - the two types of ecosystems simultaneously. The relevant links between microbiomes and cancers may be impossible to reduce to a single component. Exploring the role of microbiomes in cancer will likely require a systems biology approach.
The International Cancer Microbiome Consortiumi made a frank statement in 2019: ‘there is currently no direct evidence that the human commensal microbiome is a key determinant in the aetiopathogenesis of cancer’ [5]. It will be crucial to distinguish direct microbiome effects on cancer from indirect effects. Direct effects could occur when microbes contact the cancerous tissue directly and impact on their behaviors. Indirect effects could occur when microbiomes impact on tumors at distant sites, such as for example, when the gut microbiome influences host physiology or systemic inflammation, which then impacts on cancer progression elsewhere (Figure 1). Although there are few examples so far of microbiomes that impact on cancers directly, the past decade of microbiome research has showed us that microbiomes influence host physiology in many ways. It is likely that some of those indirect effects determine the etiopathogenesis of cancer.
Figure 1. The Human Microbiome May Interact with a Cancer through Intricate Feedback Loops, and Investigating the Interaction Networks May Require a Systems Biology Approach.
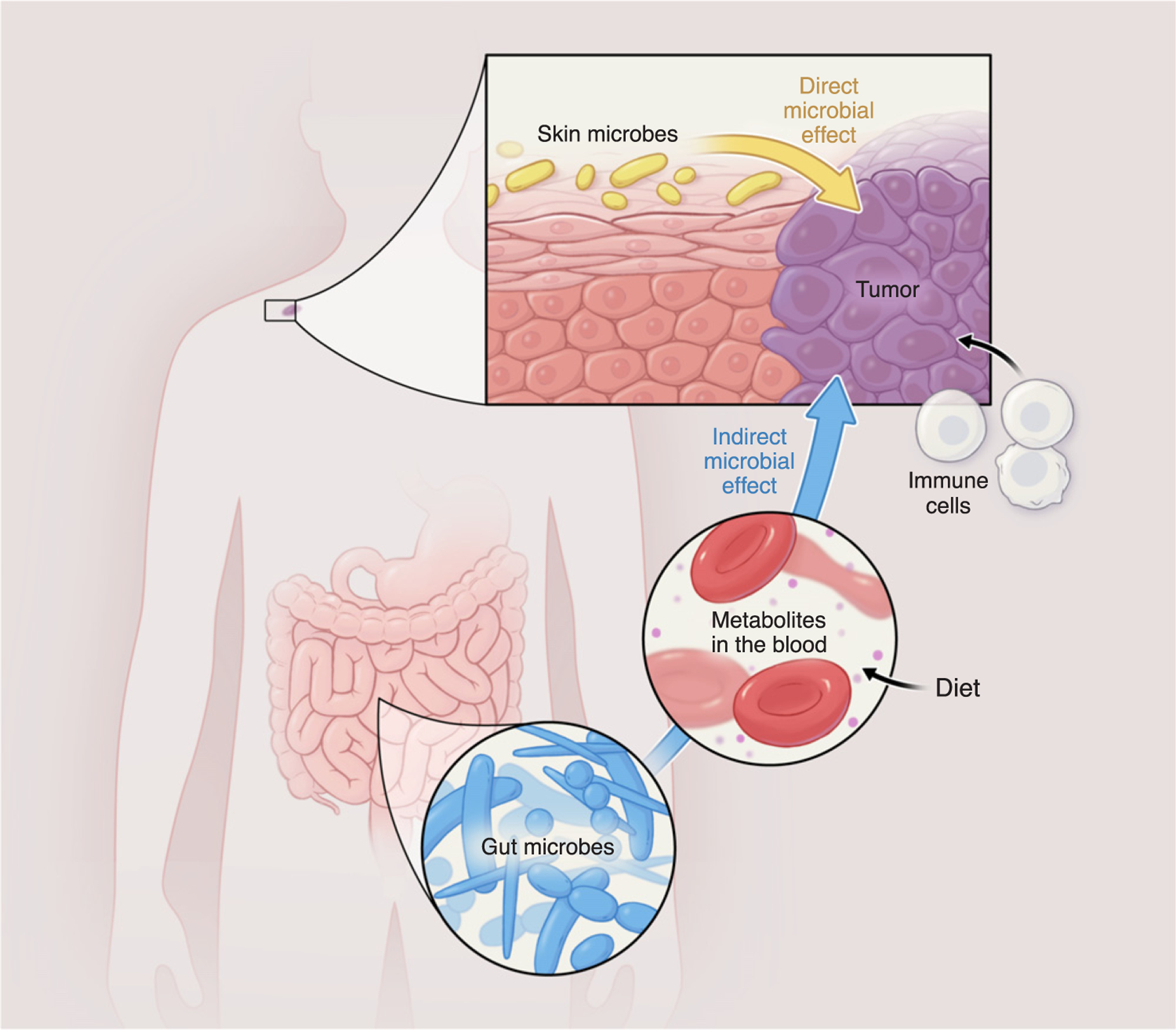
The effect of a microbiome on a cancer may be direct or indirect, and this is an important distinction. Direct interaction could take place between microbiomes residing in tissues where the cancer emerges, such as the hypothetical roles illustrated here of the skin microbiome interacting directly with melanoma; indirect interactions could occur between a microbiome and a cancer residing in a different tissue. For example, the gut microbiome alters circulating metabolites which in turn affect general host physiology and could have an indirect effect on the progression of skin cancer or its response to therapy. Diet will also play a role because it affects the circulating levels of metabolites and the microbiome composition.
Indirect Effects of Microbiomes on Cancer through Metabolism
One aspect of host physiology where the impact of the gut microbiome is well established is the systemic balance of circulating levels of nutrients and metabolites [6]. A seminal study in the microbiome literature showed that, when the gut bacteria of obese mice were transferred into lean germ-free mice, the recipient mice became obese [2]. Importantly, the recipient mice from obese donors gained more fat than control recipient mice from lean donors, even though both groups consumed the same number of calories. The bacteria residing in the gut microbiome catalyze many bioreactions that convert metabolites that reach the intestine - either through the diet or released by the host. The host can absorb or reabsorb some of those microbially converted metabolites which then enter the circulation [7], and from circulation the microbial products can reach distant tissues and impact on cancer progression there. In liver cancer, for example, gut commensals can metabolize bile acids from primary to secondary structures which can recirculate and modulate recruitment of natural killer T cells to cancerous liver cells [8]. Other bacteria in the gut metabolize estrogen, potentially altering the risk of postmenopausal estrogen receptor-positive breast cancer, even though the breast is far from the gut [9,10]. In men, microbes can generate 11-oxyandrogens from host glucocorticoids in the gut and urinary tract, which may contribute to prostate cancer [11–13].
Antibiotics can damage the gut microbiome and impact on metabolite levels in the plasma [14], which can affect host cells that interact with cancers in distant tissues. Two metabolites affected indirectly by antibiotic use - trimethylamine N-oxide and betaine - impact on peritoneal macrophages in mice, changing their phenotype to display markers associated with atherosclerosis [15]. In that example, the antibiotic treatment impacted on the microbiome, which then affected the levels of circulating metabolites in a way that harmed host health. However, the demonstration that microbiota manipulations can alter macrophage markers could inspire strategies against cancer. Macrophages are phenotypically very versatile, and some of their phenotypes can actually promote tumor growth [16]. Engineering a microbiome that reduces the systemic levels of metabolites, such as lactate that polarizes tumor-associated macrophages [17,18], could potentially turn off the tumor-promoting phenotypes of those macrophages.
The gut microbiome may also influence the central nervous system, mood, and behavior, and potentially impact on cancer. The link between microbiomes and animal behaviors is a hot topic [19], but this link is probably ancient: animals have always coexisted with microbes and have many symbiotic relationships with their commensals which affect behavior [3]. The reward system of the brain, a key circuit in emotional processes, can potentiate an antitumor immune response. In a recent study also in mice, activating the reward system with chemogenetics shrank lung tumors [20]. The mechanism involved a cascade of effects: the reward system attenuated noradrenergic input to a major immunological site, the bone marrow. The myeloid-derived suppressor cells became less immunosuppressive as a result and caused an antitumor response.
The microbiota can indirectly impact on the host reward systems [21] and even the host response to drugs that affect those systems [22]. Therefore, the microbiome could in principle enhance antitumor immunity and decrease cancer progression indirectly by altering the psychological state of the host. A cancer therapy that works on this principle may seem to be science fiction today, but the use of probiotics to reduce pain - either directly or indirectly by potentiating analgesics such as opioids - is already under study (Figure 2) [23]. The interaction between microbiomes and the central nervous system could at least be leveraged to control cancer symptoms when cures are unavailable.
Figure 2. Microbiome Engineering Could Reduce Pain in Cancer Patients.
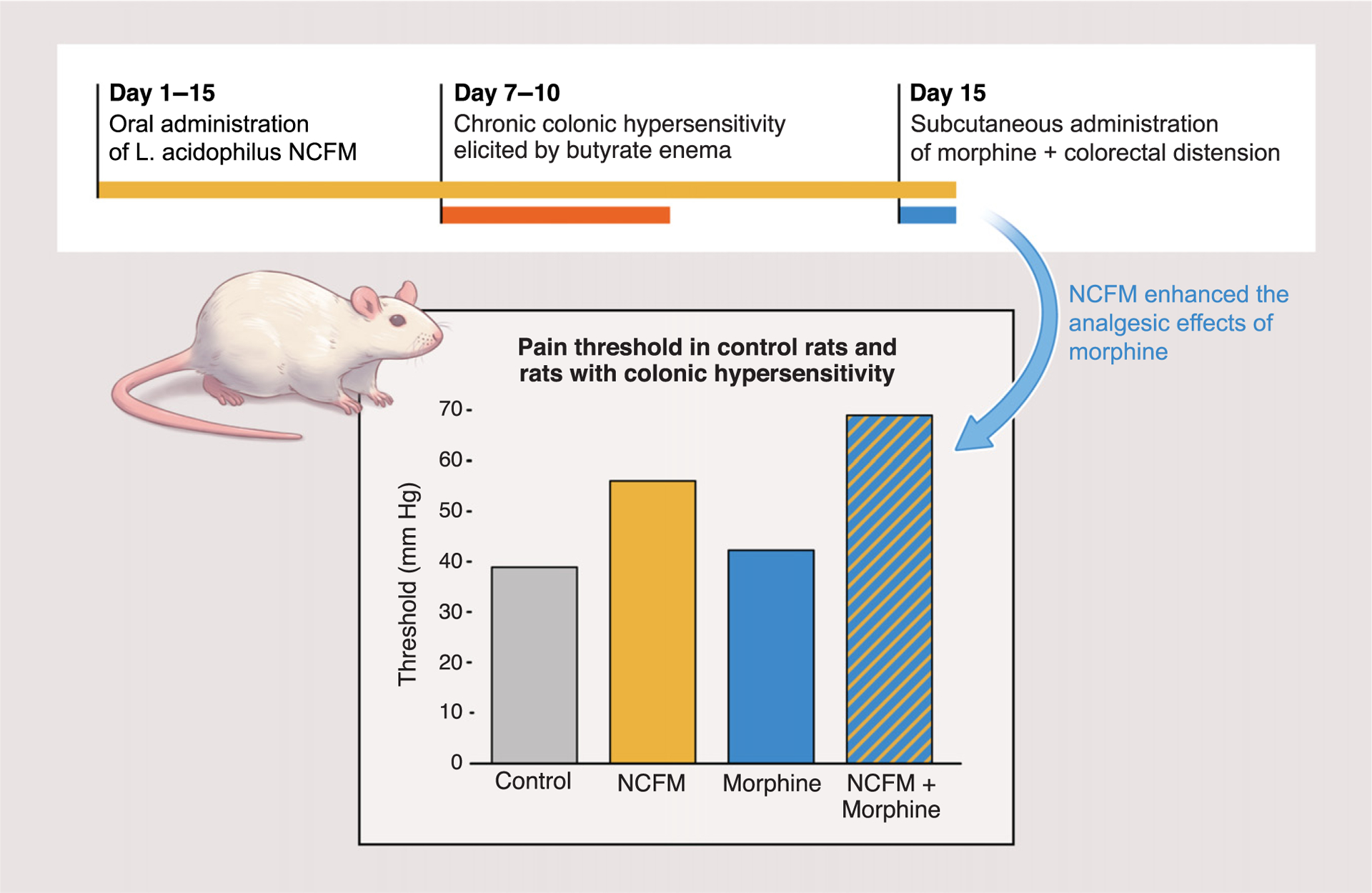
In a mouse model, the probiotic Lactobacillus acidophilus NCFM strain decreased pain relative to controls and potentiated the effect of opioid analgesics [23].
Taking full advantage of the potential of the microbiome against cancer requires a better understanding of how commensals function. Fortunately, the microbiome field is rapidly expanding from taxonomic studies to functional studies (Box 3). Some beneficial functions may be pinned down to specific microbes, such as when a specific commensal belonging to the Bifidobacterium genus promotes antitumor immunity in mice [24]. However, other functions, such as metabolic capabilities, may spread across broad phylogenetic clades. In these cases, several microbes could be used to provide the same function, assuming conservation of function within clades [25]. In addition, studying the composition of microbiomes in large populations can reveal new links between commensal members of the microbiome and cancer incidence (Box 4).
Box 3. Emerging Technologies to Study the Role of Microbiomes in Cancer.
Studying the role of microbiomes in cancer will require distinguishing direct effects from indirect effects because they warrant different approaches and even different technologies. Direct effects concern microbes that belong to the ‘tumor microbiome’, where they can contact cancer cells directly and affect their behaviors. Spatial concerns and tumor heterogeneity play a strong role. These studies may require sophisticated imaging in situ which leaves spatial structure intact. We could leverage multiplexed hybridization techniques that label multiple players at the same time: bacteria, mucus, and even levels of gene expression in the intestinal cells [88,89]. Promising hybridization protocols include CLASI-FISH, which uses a combination of bacterial probes to show the spatial organization of the microbes [90], and MERFISH, which reveals gene expression in single cells [91]. Multiplexed hybridization protocols destroy the sample, which means that the technique cannot be used to follow the same sample across time. The high-resolution cross-sectional data that these methods do provide are, however, uniquely suited to study spatial organization: the data can lead to new agent-based computational models that describe how cancer and microbial cells organize themselves in tumors. Agent-based packages such as PhysiCell, which combines simulation of diffusional gradients and mechanical processes [92], could be expanded to include bacterial cells and then applied to study inflammation feedback mechanisms that mediate interactions between cancer cells and microbiomes. Models should be developed with close collaboration between modelers and experiment-talists to foster feedback between experiment and simulation.
Indirect effects of microbiomes will require analytical methods to study systemic microbiome function; these methods should integrate microbial composition and function with other relevant data for systems modeling such as habitat, host information, other omic data, etc. Some bioinformatics tools are already available to study microbiome function, such as BugBase [93] and PICRUSt2 [94] which seek to quantify the functional composition of commensal microbes using broad phenotypic categories such as ‘anaerobe’, ‘glucose utilizing’, and ‘denitrifier’. Other tools, such as QIIME 2 [95], strive to provide a platform for microbiome multi-omics, or integration of microbiome datatypes, such as taxonomic profiles, metagenomes, and metabolomes, to support simultaneous investigation of which microbes are present, their functional potential, and their metabolic activities, to move beyond studies of microbiome associations to studies of microbiome mechanisms. Functional studies are already common in other fields of applied microbiology, such as in the biological treatment of wastewater [96], and could be an important tool to link microbiome composition and function.
Box 4. The Impact of the Environment on Microbiome Composition and its Role in Cancer Incidence.
Microbiome studies in large populations can help us to understand changes in the incidence of cancer and lead to new mechanisms linking microbes and cancer. The microbiota of industrialized populations differs from populations that retain a preindustrial lifestyle, such as the Hadza people who are hunter-gatherers in Tanzania [97,98]. Industrialized microbiota are less diverse, and the lower diversity may explain high incidences of autoimmune and inflammatory disorders in parts of the world where our symbiotic relationship with the microbiota has been most affected [99]. Antibiotics, the amount of fiber in diet [100], food additives [101,102], and even excessive hygiene [103] can also change the microbiome composition. All these changes can potentially impact on immune responses, cause autoimmune problems, and compromise our ability to fight pathogens [99], all of which are risk factors for cancer.
A recent study on the microbiomes of US immigrant populations (Figure I) showed that the gut microbiome of migrants changed when they came to the USA: strains typical of US populations and metagenomic functions displaced native strains and functions in the guts of migrants immediately after migration [104]. The longer the migrants stayed in the USA, the more their microbiomes changed. The change was even greater in the next generation and - importantly - correlated with obesity. The changes in microbiome composition in migrants could be simply due to changes in diet. However, it is unknown whether the different gut microbes could have come from other humans, raw fruits or vegetables, animals, or even other environmental sources. The loss of microbial diversity could be due to the selective pressure imposed by a different diet, but it could also be due to exposure to a microbiome in the US environment which might be less diverse than that of the original country. Nevertheless, the suggestion that immigration impacts on microbiome diversity and correlates with health problems that can lead to cancer, such as obesity, justifies efforts to establish microbiome banks that could be used for microbiome reconstitution. The nonprofit Microbiota Vaultii has a reservoir of ‘lost bugs’ to rescue the diversity depleted in the modern era [105]. Another nonprofit venture, the OpenBiome projectiii, has a large sample bank for FMT motivated by the successes of FMT against Clostridioides difficile colitis [76,106].
Figure I. Migration to the USA Can Lower the Diversity of the Gut Microbiota.
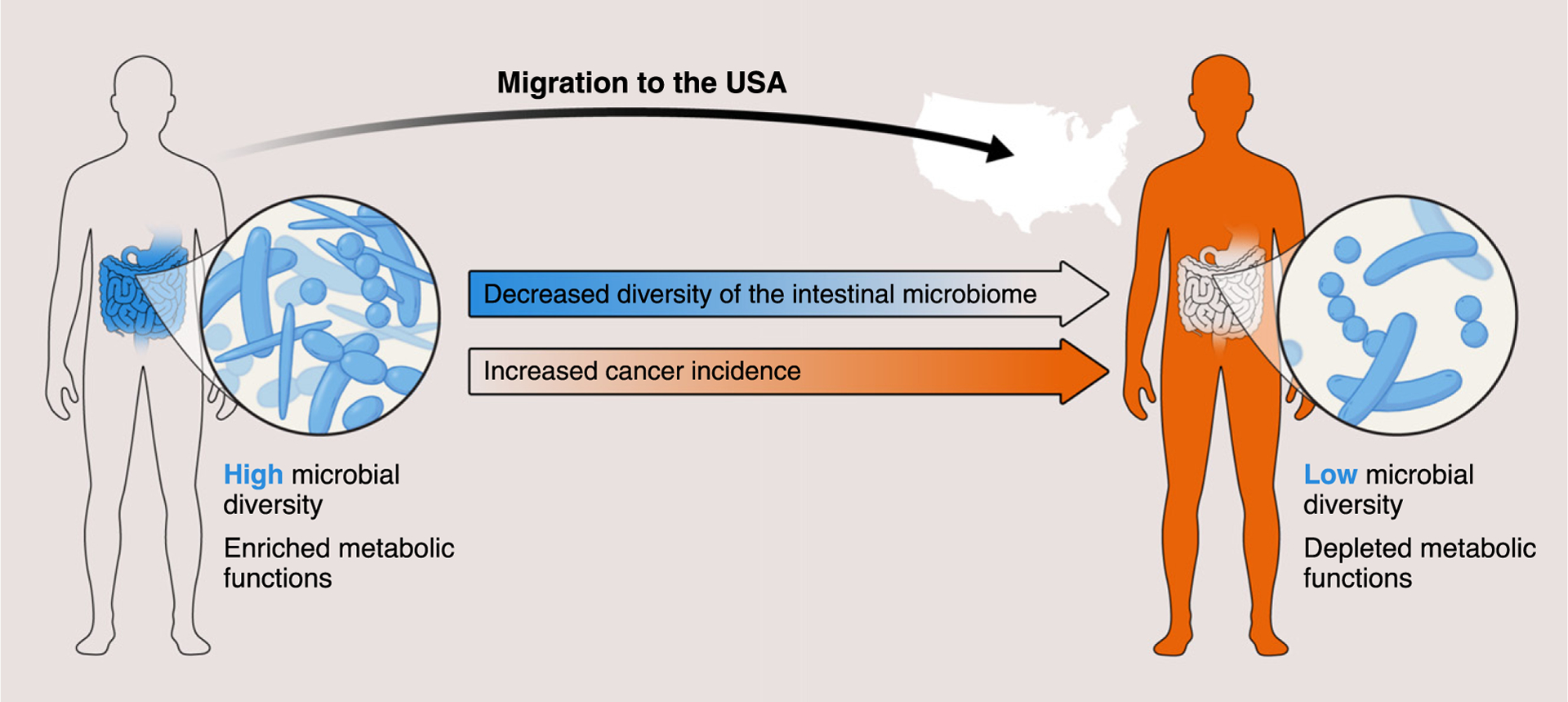
This could explain why the incidence of particular diseases - including obesity and some cancers - increases in some populations of US immigrants. Studying this intriguing phenomenon could provide new clues to the role of microbiomes in cancer.
Direct Effects of Microbiomes on Cancer
Most studies of direct effects come, perhaps expectedly, from tumors in tissues naturally colonized by commensals: the gut microbiome on colorectal cancer, the lung microbiome on lung cancer, the skin microbiome on melanoma, the vaginal microbiome on cervical and endometrial cancers, etc. However, microbes in unexpected tissues can impact on cancer progression and response to therapy. Pancreatic cancers can have bacteria accidentally introduced by medical intervention [26–28], and colorectal cancers can bring along gut bacteria when they metastasize to other tissues such as the liver [29]. Regardless of whether a tissue is already colonized, the direct interaction between microbes and cancer cells remains poorly understood.
This area of research would benefit greatly from a system that allows modular experiments in vivo, where each component can be changed independently such that the mechanisms of interaction between cancer cells and microbes can be untangled. Colorectal cancer is the second largest cause of cancer death in the USA [30]. The key risk factor is genetic predisposition, but the remaining factors - diet, lifestyle, and microbiota composition - account for the majority of cases [31]. Untangling these risks could benefit from a systems approach. Colorectal cancer already has animal models such as Il10−/− mice, which could provide a modular experimental system. Depending on the genetic background, up to 100% of Il10−/− mice develop colitis; with the addition of the colon-specific carcinogen azoxymethane, 60–80% of these mice develop colon tumors [32]. Colorectal cancer is itself a modular disease that progresses in stages [33]. The tissue is directly colonized by a microbial community and its composition in bacteria, virus, and fungi changes as the disease proceeds through its stages [34–36]. The combination of a well-established model and a disease with a pathway may enable cancer microbiome studies that discern the roles and mechanisms of microbes in distinct disease stages [37].
Western diets have many nutrients that can increase intestinal tumors [38]. These diets abound in animal protein and fat, and can increase bile acid secretion into the gastrointestinal tract. Gut commensals can conjugate these bile acids before they are reabsorbed into circulation. One species, Bilophila wadsworthia, converts taurine to acetate and ammonia, and releases hydrogen sulfide which is carcinogenic [39]. However, even when specific microbes such as B. wadsworthia contribute to cancer progression, other microbes may also impact on the process, which means that the mechanisms involved must be approached systematically. Spatial structure may also play a key role. For example, microbial biofilm organization has been associated with the location of development of colorectal cancer [40] and was also shown to promote carcinogenesis in preclinical models [41].
The transition from a localized cancer to a disseminated disease - metastasis - is a key turning point when many cancers evolve from a curable to an incurable stage. A recent study showed that microbes can travel from primary tumor sites to distal sites together with metastatic cells. The study focused on Fusobacterium nucleatum, one of the most prevalent bacteria in colorectal cancers. Patient biopsies suggested that Fusobacterium and other bacteria from the gut microbiome travel from the primary tumor to distant metastases (Figure 3). Mouse xenografts from primary colorectal adenocarcinomas show that cancer cells can indeed carry viable Fusobacterium and transport them to metastatic sites. The ability to carry bacteria was maintained after several pas-sages and was important for malignancy: reducing the bacterial load by treating the mice with antibiotics reduced tumor growth. These results suggested that antimicrobial interventions could help in treating colorectal cancers associated with Fusobacterium [42].
Figure 3. Metastases Can Carry Bacteria from the Microbiome of Primary Tumors to Distal Tissues.
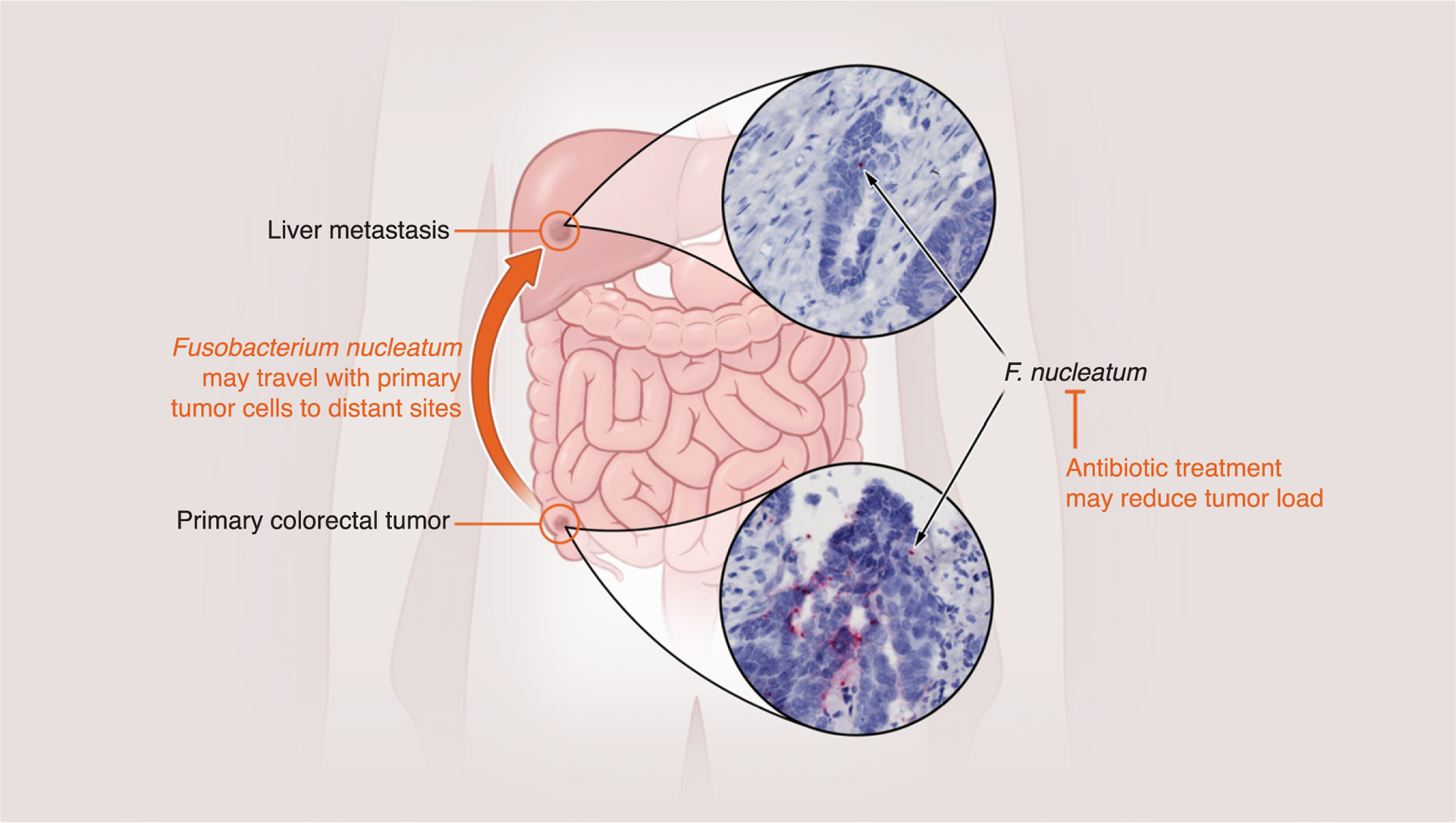
A recent study [42] investigated patient biopsies from colorectal cancer and liver metastases from the same patients. Bacteria found in primary tumors, including Fusobacterium nucleatum, were also found in metastases, suggesting they had traveled with the metastatic cells. Experiments with mice validated these findings and showed that antibiotic treatment could reduce tumor load.
However, bacteria that interact directly with cancer cells could actually help to treat cancers by causing localized inflammation [43]. This serves as additional evidence of the duplicitous role of bacteria (discussed in Box 2). The interaction becomes even more complex when we consider that bacteria can drive drug resistance: some bacteria can metabolize the chemotherapeutic cancer drug gemcitabine (2’,2’-difluorodeoxycytidine) into its inactive form, 2’,2’-difluorodeoxyuridine. The inactivation of the cancer drug requires that bacteria express a long isoform of cytidine deam-inase, an enzyme that is predominantly present in gammaproteobacteria. Experiments in a mouse model with subcutaneous syngeneic tumors showed that gammaproteobacteria living inside the tumor can increase tumor resistance to gemcitabine. A combination therapy of gemcitabine and ciprofloxacin - a broad-spectrum antibiotic that kills gammaproteobacteria - halted resistance to the cancer drug [26], serving as another example of how antibiotics could be used in cancer treatment.
The Microbiome and Cancer Immunotherapy
Understanding the intricate feedback between microbes, immunity, and cancer - and their impact on immunotherapy - will likely require multidisciplinary approaches and a systems perspective. Immunotherapy can treat advanced melanoma, and its successes may apply to other cancers. Up to 60% of patients with metastatic melanoma can now achieve durable remissions thanks to the ability to unleash the immune system with checkpoint inhibitors [44]. Three papers published recently suggest that the gut microbiome of a patient can influence their response to immunotherapy [45–47]. These studies found differences in the gut microbiota composition of patients who responded to checkpoint inhibitors and patients who did not respond. Intriguingly, the three studies did not agree on which specific microbes mattered for the success of immunotherapy [48]. In addition to a lack of agreement about which commensal microbes matter the most, the mechanisms by which the gut microbiome influences immunotherapy remain unclear [49]. The microbiomes of responders can induce the production of interferon (IFN)-γ and gran- zyme B by CD4 and CD8 T cells, which would help to recruit antitumor macrophages. Checkpoint inhibitors are less successful against models of melanoma in germ-free and antibiotic-treated mice compared with mice with intact gut microbiota [24]. Some studies even suggest that specific bacteria enriched in responders, such as Bacteroidetes thetaiotaomicron and Faecalibacterium prausnitzii [50], may be used in precision probiotic treatments to augment the efficacy of checkpoint inhibitors.
A recent study showed that short-chain fatty acids (SCFAs) produced by gut bacteria promote the memory potential of antigen-activated CD8+ T cells [51]. This important finding addresses the healthy skepticism about the disparity in taxa discovered by different groups. F. prausnitzii, which several groups had identified in patients who responded favorably to checkpoint blockade, is a key clostridial producer of SCFAs including butyrate. The literature on IBD and gut graft-versus-host disease suggests that these cluster XIVa and IV clostridia produce SCFAs that then induce a more tolerant and anti-inflammatory environment by inducing regulatory T cells and interleukin (IL)-10. It had seemed contradictory that gut microbes such as F. prausnitzii would be beneficial in the context of cancer immunotherapy, but this recent study showed that these SCFAs may induce different phenotypes depending on various environmental and host-specific factors [51]. The conditional mechanism explaining why SCFA-producing clostridia impact on immune checkpoint blockade efficacy echoes the point raised earlier that several gut microbiota members may have a common functional output (e.g., SCFA, butyrate production), and the metabolites that they produce are functionally important.
There are other reasons to investigate the gut microbiome of patients receiving immunotherapy: the therapy has side effects and can itself impact on the gut microbiome, causing more complications. Checkpoint inhibition can cause immune-mediated colitis [52]. A prospective study of patients with metastatic melanoma undergoing ipilimumab treatment identified gut bacteria in the precolitis microbiota of patients that later developed colitis [53]. The patients had fewer bacteria from the Bacteroidetes phylum and also lower expression of genes involved in polyamine transport and B vitamin biosynthesis. If these biomarkers indeed explain why some patients developed colitis, then this finding could lead to interventions that ameliorate the microbiota and reduce the risk of inflammatory complications from cancer immunotherapy. More broadly, the prospective study illustrates that immunotherapy can cause gut inflammation that then alters the composition of the microbiome, causing a feedback loop that can lead to more complications. Remarkably, in a case report serial fecal microbiota transplantation (FMT) alleviated programmed cell death protein 1 (PD-1)-induced refractory colitis in patients [54].
Restoring Microbiomes after Therapy
Cancer treatments such as radiation and chemotherapy can damage the microbiome and lead to complications. Patients with prostate, gynecological, or gastrointestinal cancers, for example, pelvic tumors, often receive pelvic radiotherapy which can be extremely effective (e.g., 90% effective for prostate cancer). However, pelvic radiotherapy causes chronic diarrhea for which there is no effective treatment [55–57]. The composition of the gut microbiome at the time a patient receives the radiation may be responsible for this complication. Specifically, low levels of bacterial richness in the lower gut before radiation is linked to a higher risk of diarrhea after radiation treatment [58]. Understanding how microbiome composition at the time of the treatment may influence post-treatment complications requires more systematic analysis.
Cancer patients receiving bone marrow transplantations experience drastic changes in their gut microbiota as a result of the large doses of antibiotics given to prevent or treat infections. Antibiotics are essential because any infection becomes life-threatening if it occurs before the bone marrow transplant has had time to engraft and the patient is neutropenic. However, the antibiotics cause collateral damage to the gut commensals: the microbiota loses diversity and may become domi-nated by bacteria such as vancomycin-resistant Enterococcus faecium [59], which increases the risk of bloodstream infections and patient mortality [60]. A recent trial conducted with transplant patients showed that autologous fecal microbiota transplantation (auto-FMT) can reconstitute the microbiota composition in patients after they receive bone marrow transplantation (Figure 4) [61]. The study concluded that auto-FMT restored microbiota composition for the average patient, but the success varied significantly from patient to patient. In the best case, a patient recovered practically 100% of their original microbiota composition but, in the worst case, a patient recovered only 50%. Why successes varied so widely is not known, but could reflect factors such as the actual composition of the microbiota transplanted, the state of the microbiota before the auto-FMT, or even personal factors including patient genetics or underlying disease. Understanding the variable success of FMT is an important direction for future research.
Figure 4. Many Cancer Therapies such as Chemotherapy and Pelvic Irradiation Can Damage Healthy Microbiomes.
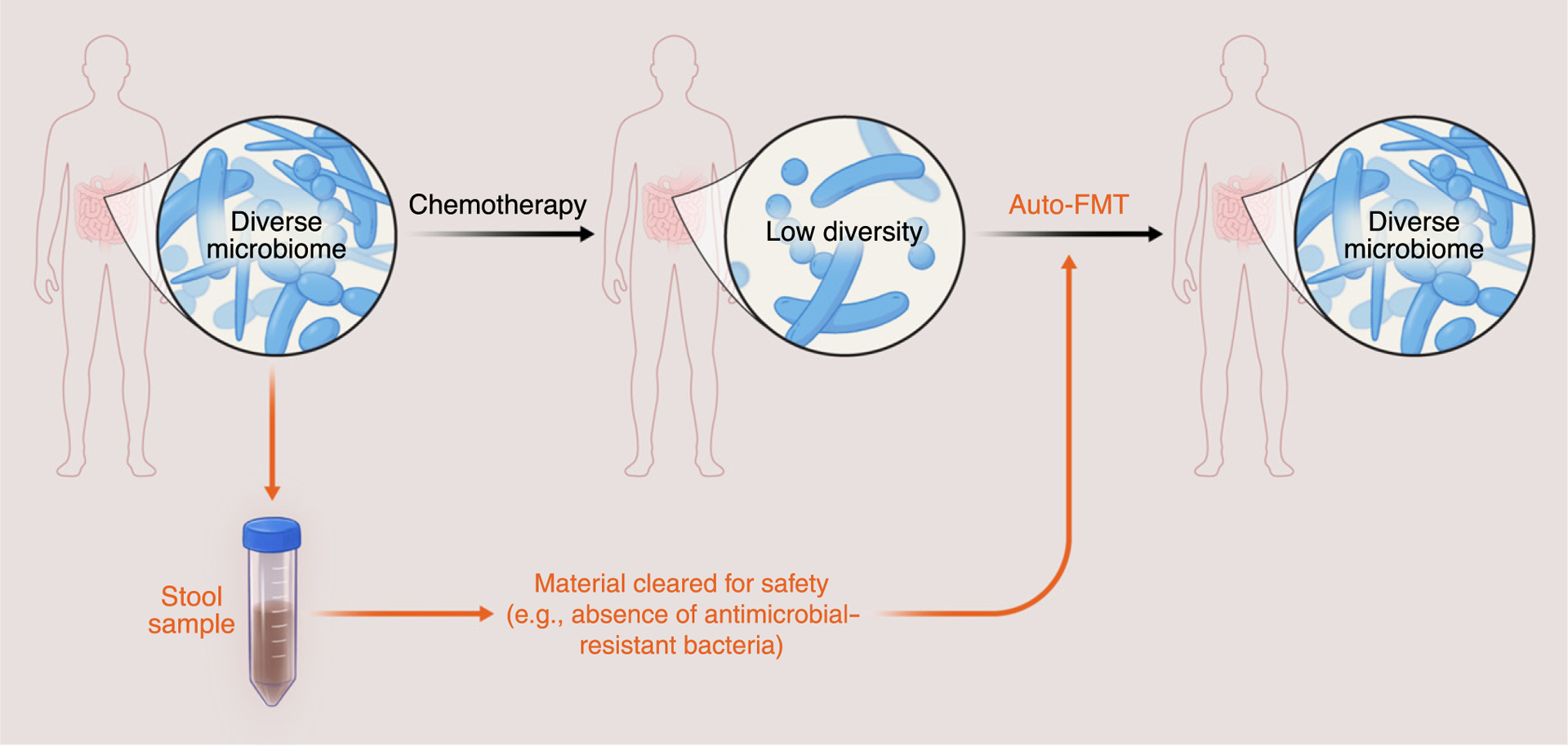
Transplants of intestinal microbiota such as autologous fecal microbiota transplant (auto-FMT), where a stool sample is collected, cleared for safety, and stored to then be given as a treatment after microbiota damage, could be used to safely remediate the microbiome of a patient. Auto-FMT has been applied to cancer patients receiving hematopoietic cell transplantation. Applications to other cancer patients should proceed with caution because we do not fully understand the link between the gut microbiome and cancer; auto-FMT raises a concern that restoring the pretreatment microbiome composition could increase the odds of cancer recurrence.
Probiotic therapies also vary markedly depending on the resident microbiota of the host [62], but also depending on differences in genes expressed by the host. This indicates complex interplay between microbiota ecology and host factors. Stable colonization of the gut microbiome by a probiotic strain of Bifidobacterium longum depends on the resident microbiome through unclear mechanisms [63]. In addition, it is important to consider that success of auto-FMT can be at least partially compromised by probiotics [64].
Despite the initial successes of auto-FMT to restore the microbiota of cancer patients receiving hematopoietic cell transplantations, the same idea may not work as well for other cancers. If the microbiota has a crucial role in cancer initiation and progression, performing auto-FMT in those patients could increase the odds of recurrence. Given the current state of our knowledge, auto-FMT cannot be deemed a risk-free approach for every cancer without further understanding the role of the microbiome in a particular disease. We should try to understand better how the microbes residing in the gut interact with each other and with the host to improve the success of probiotics and FMT, and – especially – future personalized cancer therapies based on microbiota cocktails [65].
Concluding Remarks
The many microbes living in and on the human body impact on many aspects of human physiology. It seems logical, then, that human microbiomes can impact on cancer prevention, development, treatment, and management, although the specific mechanisms remain unclear. The departure from physiological homeostasis caused by cancer and by cancer therapies can damage microbiomes, which may then increase the risk of more complications. Cancer, cancer treatment, tumor-associated microbes, and the human microbiome may all be linked through intricate feedback networks. Untangling these networks and their plasticity requires a systems biology approach.
The examples discussed here could be taken as invitations to investigate a scientific frontier that remains largely unexplored (see Outstanding Questions). The field should benefit from recent advances in microbiome research, as a field evolving from cross-sectional studies that first sought to catalogue the human microbiome to longitudinal studies that seek to understand function. The evolution of the microbiome field brings us closer to understanding the dynamic role of microbiomes in carcinogenesis and cancer progression. The human microbiome is an integrated part of a large cancer-host-microbiome system. Although each single component may appear to be insignificant compared with the magnitude of the disease, recent research already shows that microbiome manipulations can nudge cancer systems towards desired states.
Outstanding Questions.
What other members of the human microbiome beyond bacteria - archaea, fungi, and viruses (including bacteriophages) - are important for cancer?
How can we integrate microbial composition and function with other relevant data such as habitat, host information, and other variables quantified by omic approaches?
Does the human microbiome play a role in the link between migration and cancer incidence?
Why does the success of microbiota transplants vary from patient to patient?
Can the ability of gut microbes to influence the central nervous system be leveraged to manage pain in cancer patients or even cancer development?
Can we engineer microbiomes to turn immunotherapy nonresponders into responders?
Without better ways to prevent, detect, and treat cancer at an early stage, the number of new people diagnosed with cancer will continue to outpace the treatments available - even as new treatments are discovered. That means many of us may need to learn to live with cancer in our lifetime. It is likely that the solution for prolonging quality and fulfilling lives, even when a cancer cure is unavailable, resides in our future ability to leverage the human microbiome to help to provide sustainable, long-term control of the disease. This could help make incurable cancer a chronic but manageable disease. Once better understood, the homeostasis between microbes and cancer systems may ultimately contribute to healthier and longer lives.
Highlights.
Microbiomes impact on human health in many ways and likely impact on cancer progression and response to therapy.
The effects of microbiomes on cancer may be direct or indirect.
The interactions between microbiomes and cancers - two complex systems on their own - require a systems biology approach.
Treatments that alter microbiome composition of cancer patients are already under investigation in trials.
Acknowledgments
This article emerged from the conversations of participants of an innovation laboratory entitled ‘Systems Biology for the Cancer Microbiome’. The innovation laboratory and this work were supported by the National Institutes of Health (grant U54 CA209975 to J.B.X.).
Glossary
- Autologous fecal microbiota transplant (auto-FMT)
the microbiota of a patient are stored before a treatment that will damage their microbiota, and are then used to restore the damaged microbiota
- Commensals
microbes that are considered to be nonpathogenic
- Human microbiome
the collection of microbes that live in and on the human body
- Microbiota cocktails
mixtures of microbes concocted for a specific therapy
- Probiotics
live microorganisms used in treatments to provide health benefits
Footnotes
References
- 1.de Martel C et al. (2012) Global burden of cancers attributable to infections in 2008: a review and synthetic analysis. Lancet Oncol. 13, 607–615 [DOI] [PubMed] [Google Scholar]
- 2.Turnbaugh PJ et al. (2006) An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444, 1027–1031 [DOI] [PubMed] [Google Scholar]
- 3.Ezenwa VO et al. (2012) Animal behavior and the microbiome. Science 338, 198–199 [DOI] [PubMed] [Google Scholar]
- 4.Jobin C (2018) Precision medicine using microbiota. Science 359, 32–34 [DOI] [PubMed] [Google Scholar]
- 5.Scott AJ et al. (2019) International Cancer Microbiome Consortium consensus statement on the role of the human microbiome in carcinogenesis. Gut 68, 1624–1632 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Arends J (2010) Metabolism in cancer patients. Anticancer Res. 30, 1863–1868 [PubMed] [Google Scholar]
- 7.Fujisaka S et al. (2018) Diet, genetics, and the gut microbiome drive dynamic changes in plasma metabolites. Cell Rep. 22, 3072–3086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ma C et al. (2018) Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science 360, eaan5931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kwa M et al. (2016) The intestinal microbiome and estrogen receptor-positive female breast cancer. J. Natl. Cancer Inst 108, djw029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Baker JM et al. (2017) Estrogen-gut microbiome axis: physiological and clinical implications. Maturitas 103, 45–53 [DOI] [PubMed] [Google Scholar]
- 11.Devendran S et al. (2017) Identification and characterization of a 20beta-HSDH from the anaerobic gut bacterium Butyricicoccus desmolans ATCC 43058. J. Lipid Res 58, 916–925 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Devendran S et al. (2018) The desA and desB genes from Clostridium scindens ATCC 35704 encode steroid-17,20-desmolase. J. Lipid Res 59, 1005–1014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zimmermann M et al. (2019) Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 570, 462–467 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Schulfer AF et al. (2019) The impact of early-life sub-therapeutic antibiotic treatment (STAT) on excessive weight is robust despite transfer of intestinal microbes. ISMEJ. 13,1280–1292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang Z et al. (2011) Gut flora metabolism of phosphatidyl-choline promotes cardiovascular disease. Nature 472, 57–63 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Carmona-Fontaine C et al. (2013) Emergence of spatial structure in the tumor microenvironment due to the Warburg effect. Proc. Natl. Acad. Sci 110, 19402–19407 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Carmona-Fontaine C et al. (2017) Metabolic origins of spatia organization in the tumor microenvironment. Proc. Natl. Acad. Sci. U. S. A 114, 2934–2939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Colegio OR et al. (2014) Functional polarization of tumour-associated macrophages by tumour-derived lactic acid. Nature 513, 559–563 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sharon G et al. (2016) The central nervous system and the gut microbiome. Cell 167, 915–932 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ben-Shaanan TL et al. (2018) Modulation of anti-tumor immunity by the brain’s reward system. Nat. Commun 9, 2723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Alcock J et al. (2014) Is eating behavior manipulated by the gastrointestinal microbiota? Evolutionary pressures and potential mechanisms. Bioessays 36, 940–949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kiraly DD et al. (2016) Alterations of the host microbiome affect behavioral responses to cocaine. Sci. Rep 6, 35455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rousseaux C et al. (2007) Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat. Med 13, 35–37 [DOI] [PubMed] [Google Scholar]
- 24.Sivan A et al. (2015) Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 350,1084–1089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhou YJ et al. (2017) Cancer killers in the human gut microbiota: diverse phylogeny and broad spectra. Oncotarget 8, 49574–49591 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Geller LT et al. (2017) Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science 357,1156–1160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pushalkar S et al. (2018) The pancreatic cancer microbiome promotes oncogenesis by induction of innate and adaptive immune suppression. Cancer Discov. 8, 403–416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Thomas RM et al. (2018) Intestinal microbiota enhances pancreatic carcinogenesis in preclinical models. Carcinogenesis 39, 1068–1078 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kostic AD et al. (2012) Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 22, 292–298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Mojica CM et al. (2018) Interventions promoting colorectal cancer acreening among Latino men: a systematic review. Prev. Chronic Dis 15, E31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nistal E et al. (2015) Factors determining colorectal cancer: the role of the intestinal microbiota. Front. Oncol 5, 220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Arthur JC et al. (2012) Intestinal inflammation targets cancer-inducing activity of the microbiota. Science 338, 120–123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Markowitz SD and Bertagnolli MM (2009) Molecular origins of cancer: molecular basis of colorectal cancer. N. Engl. J. Med 361, 2449–2460 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Coker OO et al. (2019) Enteric fungal microbiota dysbiosis and ecological alterations in colorectal cancer. Gut 68, 654–662 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Nakatsu G et al. (2015) Gut mucosal microbiome across stages of colorectal carcinogenesis. Nat. Commun 6, 8727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Nakatsu G et al. (2018) Alterations in enteric virome are associated with colorectal cancer and survival outcomes. Gastroenterology 155, 529–541 [DOI] [PubMed] [Google Scholar]
- 37.Yachida S et al. (2019) Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. Nat. Med 25, 968–976 [DOI] [PubMed] [Google Scholar]
- 38.Goncalves MD et al. (2019) High-fructose corn syrup enhances intestinal tumor growth in mice. Science 363, 1345–1349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ridlon JM et al. (2016) Taurocholic acid metabolism by gut microbes and colon cancer. Gut Microbes 7, 201–215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Dejea CM et al. (2014) Microbiota organization is a distinct feature of proximal colorectal cancers. Proc. Natl. Acad. Sci. U. S. A 111, 18321–18326 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tomkovich S et al. (2019) Human colon mucosal biofilms from healthy or colon cancer hosts are carcinogenic. J. Clin. Invest 130, 1699–1712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bullman S et al. (2017) Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 358, 1443–1448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Song S et al. (2018) The role of bacteria in cancer therapy - enemies in the past, but allies at present. Infect. Agents Cancer 13, 9–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Larkin J et al. (2015) Combined nivolumab and ipilimumab or monotherapy in untreated melanoma. N. Engl. J. Med 373, 23–34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gopalakrishnan V et al. (2018) Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science 359, 97–103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Matson V et al. (2018) The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science 359,104–108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Routy B et al. (2018) Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 359, 91–97 [DOI] [PubMed] [Google Scholar]
- 48.Gharaibeh RZ and Jobin C (2019) Microbiota and cancer immunotherapy: in search of microbial signals. Gut 68, 385–288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Spor A et al. (2011) Unravelling the effects of the environment and host genotype on the gut microbiome. Nat. Rev. Microbiol 9, 279–290 [DOI] [PubMed] [Google Scholar]
- 50.Frankel AE et al. (2017) Metagenomic shotgun sequencing and unbiased metabolomic profiling identify specific human gut microbiota and metabolites associated with immune checkpoint therapy efficacy in melanoma patients. Neoplasia 19, 848–855 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bachem A et al. (2019) Microbiota-derived short-chain fatty acids promote the memory potential of antigen-activated CD8+ T cells. Immunity 20, 285–297 [DOI] [PubMed] [Google Scholar]
- 52.Weber JS et al. (2013) Patterns of onset and resolution of immune-related adverse events of special interest with ipilimumab: detailed safety analysis from a phase 3 trial in patients with advanced melanoma. Cancer 119, 1675–1682 [DOI] [PubMed] [Google Scholar]
- 53.Dubin K et al. (2016) Intestinal microbiome analyses identify melanoma patients at risk for checkpoint-blockade-induced colitis. Nat. Commun 7, 10391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Wang Y et al. (2018) Fecal microbiota transplantation for refractory immune checkpoint inhibitor-associated colitis. Nat. Med 24, 1804–1808 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Benson AB 3rd et al. (2004) Recommended guidelines for the treatment of cancer treatment-induced diarrhea. J. Clin. Oncol 22, 2918–2026 [DOI] [PubMed] [Google Scholar]
- 56.Morris KA and Haboubi NY (2015) Pelvic radiation therapy: between delight and disaster. World J. Gastrointest. Surg 7,279–288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Theis VS et al. (2010) Chronic radiation enteritis. Clin. Oncol. (R. Coll. Radiol.) 22, 70–83 [DOI] [PubMed] [Google Scholar]
- 58.Wang A et al. (2015) Gut microbial dysbiosis may predict diarrhea and fatigue in patients undergoing pelvic cancer radiotherapy: a pilot study. PLoS One 10, e0126312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Taur Y et al. (2012) Intestinal domination and the risk of bacteremia in patients undergoing allogeneic hematopoietic stem cell transplantation. Clln. Infect. Dis 55, 905–914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Taur Y et al. (2014) The effects of intestinal tract bacterial diversity on mortality following allogeneic hematopoietic stem cell transplantation. Blood 124, 1174–1182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Taur Y et al. (2018) Reconstitution of the gut microbiota of antibiotic-treated patients by autologous fecal microbiota transplant. Sci. Transl. Med 10, eaap9489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Zmora N et al. (2018) Personalized gut mucosal colonization resistance to empiric probiotics is associated with unique host and microbiome features. Cell 174, 1388–1405 [DOI] [PubMed] [Google Scholar]
- 63.Maldonado-Gomez MX et al. (2016) Stable engraftment of Biffdobacterium longum AH1206 in the human gut depends on individualized features of the resident microbiome. Cell Host Microbe 20, 515–526 [DOI] [PubMed] [Google Scholar]
- 64.Suez J et al. (2018) Post-antibiotic gut mucosal microbiome reconstitution is impaired by probiotics and improved by autologous FMT. Cell 174, 1406–1423 [DOI] [PubMed] [Google Scholar]
- 65.Baxter NT et al. (2019) Dynamics of human gut microbiota and short-chain fatty acids in response to dietary interventions with three fermentable fibers. MBio 10, e02566–18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Rous P (1911) A sarcoma of the fowl transmissible by an agent separable from the tumor cells. J. Exp. Med 13, 397–411 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Stehelin D et al. (1976) DNA related to the transforming gene (s) of avian sarcoma viruses is present in normal avian DNA. Nature 260, 170–173 [DOI] [PubMed] [Google Scholar]
- 68.Lollini PL et al. (2006) Vaccines for tumour prevention. Nat. Rev. Cancer 6, 204–216 [DOI] [PubMed] [Google Scholar]
- 69.Godoy-Vitorino F et al. (2018) Cervicovaginal fungi and bacteria associated with cervical intraepithelial neoplasia and high-risk human papillomavirus infections in a Hispanic population. Front. Microbiol 9, 2533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Rampelli S et al. (2016) ViromeScan: a new tool for metagenomic viral community profiling. BMC Genomics 17,165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Garretto A et al. (2019) virMine: automated detection of viral sequences from complex metagenomic samples. PeerJ 7, e6695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Barrientos-Somarribas M et al. (2018) Discovering viral genomes in human metagenomic data by predicting unknown protein families. Sci. Rep 8, 28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Hannigan GD et al. (2018) Diagnostic potential and interactive dynamics of the colorectal cancer virome. MBio 9, e02248–18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ott SJ et al. (2017) Efficacy of sterile fecal filtrate transfer for treating patents with Clostridium difficile infection. Gastroenterology 152, 799–811 [DOI] [PubMed] [Google Scholar]
- 75.Zuo T et al. (2018) Bacteriophage transfer during faecal microbiota transplantation in Clostridium difficile infection is associated with treatment outcome. Gut 67, 634–643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hourigan SK et al. (2019) Fecal transplant in children with Clostridioides difficile gives sustained reduction in antimicrobial resistance and potential pathogen burden. Open Forum Infect. Dis 6, ofz379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Amieva M and Peek RM Jr. (2016) Pathobiology of Helicobacter pylor’-Induced Gastric Cancer. Gastroenterology 150, 64–78 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Amiri M et al. (2011) The decline in stomach cancer mortality: exploration of future trends in seven European countries. Eur J. Epidemiol 26, 23–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Linz B et al. (2007) An African origin for the intimate association between humans and Helicobacter pylori. Nature 445, 915–918 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Blaser MJ and Atherton JC (2004) Hellcobacter pylori persistence: biology and disease. J. Clin. Invest 113,321–333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Chen C et al. (2017) Accumulated evidence on Helicobacter pylori infection and the risk of asthma: a meta-analysis. Ann. Allergy Asthma Immunol 119, 137–145 [DOI] [PubMed] [Google Scholar]
- 82.Kira JI and Isobe N (2019) Helicobacter pylori infection and demyelinating disease of the central nervous system. J. Neuroimmunol 329, 14–19 [DOI] [PubMed] [Google Scholar]
- 83.Piovani D et al. (2019) Environmental risk factors for inflammatory bowel diseases: an umbrella review of meta-analyses. Gastroenterology 157, 647–659 [DOI] [PubMed] [Google Scholar]
- 84.Gravina AG et al. (2018) Helicobacter pylori and extragastric diseases: a review. World J. Gastroenterol 24, 3204–3221 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Hosseininasab Nodoushan SA and Nabavi A (2019) The interaction of Helicobacter pylori infection and type 2 diabetes mellitus. Adv. Biomed. Res 8, 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Dzutsev A et al. (2017) Microbes and cancer. Annu. Rev. Immunol 35, 199–228 [DOI] [PubMed] [Google Scholar]
- 87.Redelman-Sidi G et al. (2014) The mechanism of action of BCG therapy for bladder cancer - a current perspective. Nat. Rev. Urol 11, 153–162 [DOI] [PubMed] [Google Scholar]
- 88.Earle KA et al. (2015) Quantitative Imaging of gut microbiota spatial organization. Cell Host Microbe 18, 478–488 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Tropini C et al. (2017) The gut microbiome: connecting spatia organization to function. Cell Host Microbe 21, 433–442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Mark Welch JL et al. (2017) Spatial organization of a model 15-member human gut microbiota established in gnotobiotic mice. Proc. Natl. Acad. Sci. U. S. A 114, E9105–E9114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Moffitt JR et al. (2016) High-performance multiplexed fluorescence in situ hybridization in culture and tissue with matrix imprinting and clearing. Proc. Natl. Acad. Sci. U. S. A 113, 14456–14461 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Ghaffarizadeh A et al. (2018) PhysiCell: an open source physics-based cell simulator for 3-D multicellular systems. PLoS Comput. Biol 14, e1005991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Ward T et al. (2017) BugBase predicts organism-level microbiome phenotypes. bioRxiv. Published online May 7, 2017 10.1101/133462 [DOI] [Google Scholar]
- 94.Douglas GM et al. (2019) PICRUSt2: an improved and extensible approach for metagenome inference. bioRxiv. Published online June 15, 2019. 10.1101/672295 [DOI] [Google Scholar]
- 95.Bolyen E et al. (2019) Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol 37, 852–857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Xavier JB et al. (2007) Multi-scale individual-based model of microbial and bioconversion dynamics in aerobic granular sludge. Environ. Sci. Technol 41, 6410–6417 [DOI] [PubMed] [Google Scholar]
- 97.Pasolli E et al. (2019) Extensive unexplored human microbiome diversity revealed by over 150,000 genomes from metagenomes spanning age, geography, and lifestyle. Cell 176, 649–662.e20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Smits SA et al. (2017) Seasonal cycling in the gut microbiome of the Hadza hunter-gatherers of Tanzania. Science 357, 802–806 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Belkaid Y and Hand TW (2014) Role of the microbiota in immunity and inflammation. Cell 157,121–141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.David LA et al. (2014) Diet rapidly and reproducibly alters the human gut microbiome. Nature 505, 559–563 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Chassaing B et al. (2015) Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syn-drome. Nature 519, 92–96 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Viennois E et al. (2017) Dietary emulsifier-induced low-grade inflammation promotes colon carcinogenesis. Cancer Res. 77, 27–40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Scudellari M (2017) Cleaning up the hygiene hypothesis. Proc. Natl. Acad. Sci. U. S. A 114, 1433–1436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Vangay P et al. (2018) US immigration westernizes the human gut microbiome. Cell 175, 962–972 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Bello MGD et al. (2018) Preserving microbial diversity. Science 362, 33–34 [DOI] [PubMed] [Google Scholar]
- 106.Rohlke F and Stollman N (2012) Fecal microbiota transplantation in relapsing Clostridium difficile infection. Ther. Adv. Gastroenterol 5, 403–420 [DOI] [PMC free article] [PubMed] [Google Scholar]