

Abstract
Background
Human papillomavirus (HPV) infection is one of the most common sexually transmitted infections. However, only a small percentage of high-risk (HR) HPV infections progress to cervical precancer and cancer. In this study, we investigated the role of the cervicovaginal microbiome (CVM) in the natural history of HR-HPV.
Methods
This study was nested within the placebo arm of the Costa Rica HPV Vaccine Trial that included women aged 18–25 years of age. Cervical samples from two visits of women with an incident HR-HPV infection (n = 273 women) were used to evaluate the prospective role of the CVM on the natural history of HR-HPV. We focus specifically on infection clearance, persistence, and progression to cervical intraepithelial neoplasia grade 2 and 3 (CIN2+). The CVM was characterized by amplification and sequencing the bacterial 16S V4 rRNA gene region and the fungal ITS1 region using an Illumina MiSeq platform. OTU clustering was performed using QIIME2. Functional groups were imputed using PICRUSt and statistical analyses were performed using R.
Results
At Visit 1 (V1) abundance of Lactobacillus iners was associated with clearance of incident HR-HPV infections (Linear Discriminant Analysis (LDA)>4.0), whereas V1 Gardnerella was the dominant biomarker for HR-HPV progression (LDA>4.0). At visit 2 (V2), increased microbial Shannon diversity was significantly associated with progression to CIN2+ (p = 0.027). Multivariate mediation analysis revealed that the positive association of V1 Gardnerella with CIN2+ progression was due to the increased cervicovaginal diversity at V2 (p = 0.040). A full multivariate model of key components of the CVM showed significant protective effects via V1 genus Lactobacillus, OR = 0.41 (0.22–0.79), V1 fungal diversity, OR = 0.90 (0.82–1.00) and V1 functional Cell Motility pathway, OR = 0.75 (0.62–0.92), whereas V2 bacterial diversity, OR = 1.19 (1.03–1.38) was shown to be predictive of progression to CIN2+.
Conclusion
This study demonstrates that features of the cervicovaginal microbiome are associated with HR-HPV progression in a prospective longitudinal cohort. The analyses indicated that the association of Gardnerella and progression to CIN2+ may actually be mediated by subsequent elevation of microbial diversity. Identified features of the microbiome associated with HR-HPV progression may be targets for therapeutic manipulation to prevent CIN2+.
Trial registration
ClinicalTrials.gov NCT00128661.
Author summary
Despite being the most common sexually transmitted infection and the causal agent of cervical cancer, it is still not clear why only a small proportion of high-risk HPV (HR-HPV) infections progress to cervical cancer. Our study utilizes longitudinal cervicovaginal samples from a prospective cohort, along with advanced epidemiological modeling and mediation analysis, to investigate the association between the cervicovaginal microbiome (CVM) and progression of an incident HR-HPV infection to cervical precancer. The results of our study suggest a novel association between the effect of Gardnerella and disruption of CVM homeostasis that can influence the pathway of HR-HPV infection progression to cervical precancer. We further show the interplay between several key components of the cervicovaginal microbiome and demonstrate that within the context of HR-HPV natural history the effect of Gardnerella is mediated by increased cervicovaginal bacterial diversity directly preceding the progression of a persistent infection to precancer.
Introduction
Persistent cervical infections by high-risk (HR) human papillomavirus (HPV) cause virtually all cervical cancers and their immediate precursor lesions [1]. Most sexually active women have been infected with HPV at some point in their lives and in the vast majority the infection is cleared within a few months [2]. However, a subset of women develop a persistent HPV infection that places them at high risk for cervical precancer and cancer [2–5]. Fig 1 illustrates this paradigm canonically referred to as HPV natural history.
Fig 1. HPV natural history.

The natural history of HR-HPV is depicted. Briefly, an incident HR-HPV infection can occur by entering the basal layer through an epithelial abrasion. Most incidence infections are cleared, however some remain persistent for years and decades. Persistence of a HR-HPV infection combined with known risk factors (e.g., smoking) may allow the persistent HR-HPV infection to progress to precancer (cervical intraepithelial neoplasia, CIN). If the lesion does not regress and the HR-HPV is able to successfully integrate into the host-cell genome, clonal expansion may occur and result in an invasive cancer.
Non-viral factors (HPV co-factors) associated with the outcomes of HR-HPV infections have not been fully elucidated. While smoking [6–9], hormonal contraceptive use [10, 11], and parity [12] are associated with developing precancer and cancer; systemic and local immune responses are thought to be important for clearance and control of infection (persistence vs. clearance) [13, 14]. In addition, specific host immune regulatory alleles (e.g., human leukocyte antigen) are associated with risk of cervical cancer [15, 16].
The local, cervical microenvironment, including the microbiome, may also influence the natural history of HPV infection [17]. Other studies have recently implicated the microbiome’s role in the natural history of other viral infections [18] and a variety of cancers [19–21]. The cervicovaginal microbiome (CVM) is of particular interested because it has been well characterized and specific features have been associated with gynecologic disease and reproductive health [22–24]. The CVM has been categorized into community state types (CSTs) generally defined by a dominance of a specific Lactobacillus species (Lactobacillus crispatus, Lactobacillus iners, Lactobacillus gasseri or Lactobacillus jensenii), or a state of polymicrobialism [25, 26]. Transitions from Lactobacillus dominated CSTs have been linked to detrimental health outcomes including elevated risks for sexually transmitted infections [27], as well as higher incidences of preterm births [28].
An association between increased CVM diversity and prevalence of HR-HPV infection and/or cervical abnormalities (vs. HPV negative) has been reported in several studies [29–33]. Higher abundance of L. crispatus has been shown to be associated with lower HPV prevalence [34] and increased detection of normal cytology [35]. Long-term use of vaginal probiotics with Lactobacillus spp. has been associated with increase clearance of HPV compared to short-term use [36]. However, evidence is conflicting on the association of CVM diversity and the severity of cervical neoplasia [37–39]. Additionally, most studies looking at the natural history of HPV and the microbiome are cross-sectional and therefore lack the ability to draw potential causal links.
For the current study, we leveraged longitudinal data and specimens from the placebo arm of a large randomized HPV vaccine trial [40] to examine the impact of the CVM on the natural history of incident HR-HPV infections to study: 1) progression to cervical precancer, 2) viral persistence, and 3) viral clearance.
Results
Subject characteristics and cervicovaginal microbiome features
A total of 273 women with an incident HR-HPV infection were included in the analyses, of whom 266 had a second sample with an average sampling interval of 1.5±0.9 years. Table 1 presents the sample demographic information and summarizes the bacterial and fungal sequencing results after taxonomic assignment for each infection outcome at baseline. There were no significant differences between groups in age (p = 0.13), 16S rRNA gene OTU clustered read counts (p = 0.33), or ITS1 OTU clustered read counts (p = 0.53).
Table 1. CVT cohort characteristics.
| Variable | Clearance | Persistence | Progression | p-val |
|---|---|---|---|---|
| Baseline Sample Count | 70 | 170 | 33 | |
| Age (years) | 23.4 ± 2.8 | 22.6 ± 2.3 | 22.70 ± 2.9 | 0.13 |
| Current/former Smoker | 15.7% (11/70) | 27.1% (46/170) | 26.7% (8/30) | 0.16 |
| Total number of sexual partners as of visit | 3 ± 3 | 3 ± 2 | 3 ± 2 | 0.63 |
| Contraceptive use* | 97.1% (68/70) | 97.6% (166/170) | 96.7% (29/30) | 0.86 |
| Condom use prior to visit | 44.1% (15/34) | 39.3% (33/84) | 58.3% (7/12) | 0.46 |
| Contraceptive pill prior to visit | 68.8% (33/48) | 68.5% (76/111) | 52.6% (10/19) | 0.40 |
| Injectable contraceptive prior to visit | 14.8% (8/54) | 21.7% (26/120) | 23.8% (5/21) | 0.53 |
| Other contraceptive prior to visit** | 4.6% (3/65) | 0% (0/150) | 0% (0/26) | 0.029 |
| Chlamydia/Gonorrhea Positive | 46.2% (6/13) | 30.4% (14/46) | 0% (0/3) | 0.32 |
| HPV16 Positive | 37.1% (26/70) | 34.7% (59/170) | 45.5% (15/33) | 0.49 |
| 16SV4 Reads | 10,642 ± 4,910 | 11,482 ± 4,985 | 11,041 ± 4,809 | 0.33 |
| ITS1 Reads | 1,942 ± 3,433 | 3,299 ± 7,055 | 4,660 ± 11,500 | 0.53 |
Continuous data is presented using median ± standard deviation, significance assessed using the Kruskal-Wallis test.
Count data is presented using percent with proportions shown in parenthesis, significance is assessed using Fisher's exact test.
Proportion of count data is based on only the samples which had data for a given count variable.
*Counts use of any of the following prior to the visit: condom, birth control pills, diaphragm, injectable, iud, spermicide, sponge and or other** types of contraceptives.
Fig 2 summarizes the bacterial Shannon diversity measures of the three ordered categorical HR-HPV outcomes (see Fig 1) within the CVT cohort at V1 and V2. Alpha diversity analysis did not reveal any significant differences at V1 in terms of bacterial Shannon alpha diversity (trend p = 0.52). At V2 there was a significant trend of rising diversity based on the Shannon diversity index (trend p = 0.024).
Fig 2. Bacterial Shannon diversity by visit.

HR-HPV category specific microbial Shannon diversity is shown for V1 and V2. Horizontal strip labels at the top of the figure indicate visit number. V1 has an elevated diversity in the progression group, but the overall trend did not achieve statistical significance (p = 0.52). At V2 the observed trend of a rising Shannon alpha diversity from clearance to persistence to progression was statistically significant, p = 0.024.
To evaluate the overall structure of the cervicovaginal microbiome, we performed hierarchical clustering on all available samples (n = 539) (Fig 3A). This analysis revealed four distinct bacterial community state types (CSTs). Two CSTs were dominated by species of the genus Lactobacillus (Lactobacillus iners, 143/539, 26.5% and Lactobacillus crispatus, 83/539, 15.4%), one CST by Gardnerella vaginalis (94/539, 17.4%), and the other CST did not contain a major group but had a highly diverse microbiome (219/539, 40.6%).
Fig 3. Bacterial and fungal communities within the study cohort.

A. The abundance plot represents the bacterial community structure of the study subjects. The operational taxonomic units (OTUs) were collapsed at the species level and the top 10 species are presented. Figure boxes labeled: L. iners, L. crispatus, Gardnerella and Diversity represent the vaginal community state types (CSTs) identified using hierarchical clustering. B. Heatmap showing the top 20 fungal species identified within the study subjects. C. albicans has the highest mean abundance. There were three vaginal fungal community states identified using hierarchical clustering as indicated by the separate boxes.
Fig 3B shows results of hierarchical clustering based on the detected fungal species. Candida albicans was the dominant fungal taxa. In terms of fungal clusters, there appears to be a single clade dominated by C. albicans (43/208, 20.7%), one dominated by an unidentified fungal species (12/208, 5.8%) and one that is composed of a diverse fungal community (153/208, 73.6%).
Analysis of bacterial taxonomic categories of the microbiome associated with persistence/progression vs. clearance revealed a total of 24 taxa that were significant (LDA>2.0) at V1. G. vaginalis was the bacterial species with the highest positive correlation to progression (Fig 4A), while L. iners was the most positively associated taxon with clearance based on relative abundance. Amongst V2 samples there were a total of 13 significant taxa identified (three taxa associated with clearance and 10 with progression) (Fig 4B). At V2, bacteria that are commonly associated with bacterial vaginosis, such as Prevotella amnii and Anaerococcus prevotii, were significantly correlated with progression. We used a generalized linear model (GLM) to validate LEfSe biomarkers with adjustments for key covariates (age, smoking status, HPV16 and visit CST) (S1 Table).
Fig 4. Bacteria associated with progression to CIN2+ identified using LEfSe.

HR-HPV bacterial biomarkers for visit 1 (panel A) and visit 2 (panel B), comparing clearance vs. progression, were identified using the LEfSe tool. Only the significant bacterial taxa (LDA>2.0) are shown for both visits.
Fungal biomarker discovery revealed five fungal taxa associated with HPV progression (S1A Fig). The average of the five combined fungal taxa were detected at rates of 0.59%, 3.55% and 3.76% for the clearance, persistence and progression outcomes, respectively (p = 0.0080, S1B Fig). However in the adjusted GLM analysis, none of the fungal biomarkers were determined to be significant at either visit (S2 Table).
To evaluate whether some common function of bacteria might be associated with HR-HPV outcomes, we used a functional analysis of gene groups imputed from the microbiome data as described in the methods. This analysis revealed 8 functional pathways at KEGG Level 2 significantly associated with progression to CIN2+ (S3 Table). Of the 8 identified pathways, only 2 had a mean read coverage of >1% and were considered for further analysis. The two identified pathways were “Xenobiotics Biodegradation and Metabolism” pathway, which was positively associated with progression (p = 0.0020) and the Cell Motility pathway, which was negatively associated with progression (p = 0.019). These two pathways were significantly correlated (Pearson correlation = -0.80, S2 Fig), and we chose to use Cell Motility in multivariate modeling since it produced a more stable GLM estimate (S3 Table).
Microbiome and HR-HPV natural history: GLM modeling
To investigate the contribution of components of the microbiome over time, we used a GLM in order to adjust for known covariates of CIN2+ progression that may influence the relationship of the CVM and progression to CIN2+ (e.g. age, smoking, HPV16 and CST). We utilized a GLM with a binary outcome (clearance/progression) and the significant microbial features identified in preceding sections as predictors. Specifically, we used the abundance of Gardnerella at V1, the abundance of Lactobacillus at V1, the Observed fungal species diversity at V1, the imputed Cell Motility pathway at V1 and the microbial diversity at V2. The model was adjusted for age, CST, smoking and HPV16 infection status. Fig 5 shows the model estimates of the resulting GLM analysis. The multivariate analysis revealed a significant protective effect of V1 Lactobacillus (genus) abundance, OR = 0.41 (0.22–0.79), V1 fungal species diversity, OR = 0.90 (0.82–1.00) and imputed V1 Cell Motility pathway OR = 0.75 (0.62–0.92). In addition, the model showed that the V2 microbial diversity was a significant risk factor for CIN2+ progression, OR = 1.17 (1.02–1.29).
Fig 5. Generalized Linear Model (GLM) results showing the odds ratios of key microbial components in association with progression to CIN2+.

The forest plot shows the results of variables evaluated in the univariate analysis that were then entered into a GLM. The model shows ORs (small circle) and 95% confidence interval (line extending on either side of the circle) of the microbial features associated with clearance/progression at either Visit 1 (V1) and/or Visit 2 (V2). The main variables included V1 Gardnerella, V1 Lactobacillus, V1 Fungal Observed OTUs and V1 Cell Motility and V2 Shannon diversity. The model was adjusted for age, bacterial CSTs, smoking and HPV16 infection status. 95% CIs that did not cross the Odds Ratio of 1.0 (dotted vertical line) are considered statistically significant.
Following the multivariate analysis, we wanted to explore the reason for V1 Gardnerella being insignificant despite being the top microbial risk factor in differential abundance in all previous analyses (Fig 4, S3 Fig and S1 Table). Thus, we performed a mediation analysis to determine if V1 Gardnerella was acting by inducing the elevated diversity at V2 (Fig 6A and 6B). This analysis showed that after adjustment for V1 Gardnerella there was a significant association of V2 Shannon diversity with progression to CIN2+, p = 0.04. The Average Direct Effect (ADE) showed that V1 Gardnerella wasn’t significant after adjustment for the V2 Shannon diversity, p = 0.23 supporting our mediation hypothesis (Fig 6A).
Fig 6. Diversity model for HPV progression with mediation analysis.
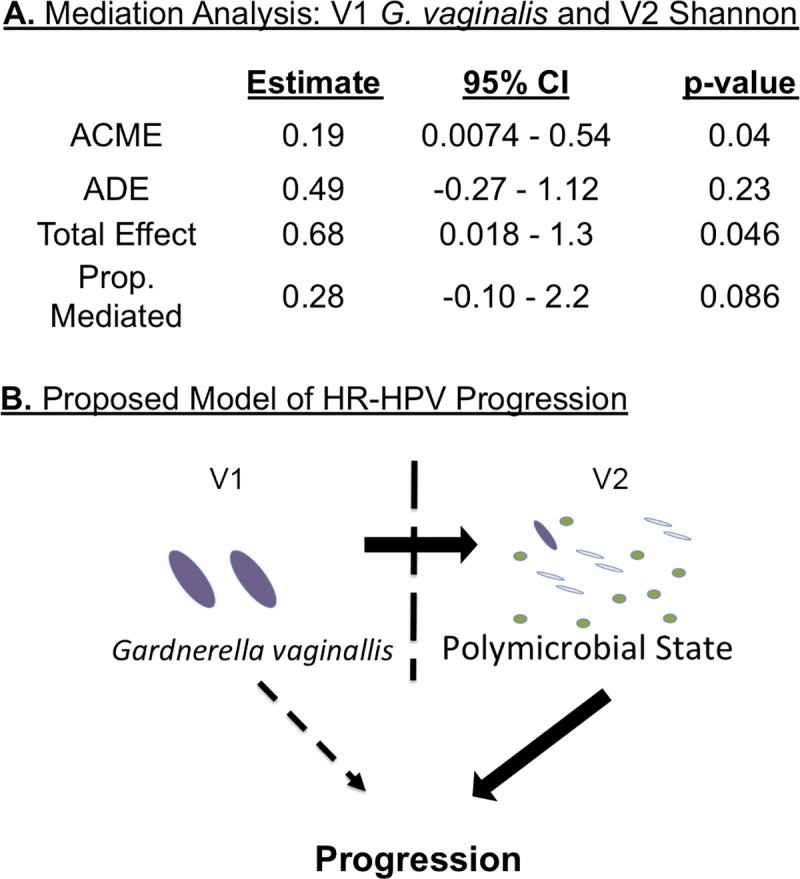
Panel A shows the results of the mediation analysis that focus on V1 Gardnerella and V2 Shannon diversity. Top row shows Average Causal Mediation Effect (ACME) which is the full mediation effect of V2 Shannon diversity after adjusting for the direct effect of V1 Gardnerella on case status. The second row shows Average Direct Effect (ADE) which is the direct effect of Gardnerella on the clearance/progression outcome after accounting for the mediation effect of V2 Shannon diversity. The third row shows the Total Effect which is the direct, unadjusted effect, of Gardnerella on case outcome. The last row shows the Proportion (Prop.) Mediated, which is the proportion of the model that is mediated by V2 Shannon diversity. Based on GLM modeling, we propose the above model (Panel B) in which V1 Gardnerella causes an expansion of bacterial diversity at V2, which acts as a risk factor for the progression of a HR_HPV infection into a CIN2+ lesion.
Power for detection of the effects of each microbiome component was performed using the lmSupport package. Specifically each of the microbiome components (i.e., V1 Gardnerella, V1 Lactobacillus, V1 Fungal Observed OTUs, V1 Cell Motility, and V2 Shannon diversity) was tested separately to assess power with adjustment for age, CST, HPV16 status and smoking. S4 Table shows the results of the power calculation. Given large effect sizes for V1 Lactobacillus, V1 Fungal Observed OTUs, V1 Cell Motility, and V2 Shannon diversity we calculated that these could be detected with a power >98%, while the V1 Gardnerella had a power of 82% at the 0.05 alpha level.
Discussion
Previous cross-sectional studies analyzing the association between the cervicovaginal microbiome and HPV infection outcomes have consistently identified Gardnerella as a key biomarker associated with CIN2+. This has been reported in studies that utilized both next-generation sequencing [39, 41] and other methods of microbiome analyses [52, 43]. We present data that Gardnerella is in fact associated with CIN2+ lesions, but rather than directly causing the CIN2+ lesion, appears to induce a higher diversity CVM over time as measured at V2, which in turn mediates the observed effect of Gardnerella in HR-HPV disease progression. Although it is not clear how a state of polymicrobialism in the presence of a persistent HR-HPV infection leads to the development of epithelial dysplasia, recent studies on the microbiome’s role in other cancers suggests that the answer lies in the establishment of a microbial microenvironment, perhaps a biofilm. For instance, it has been shown that certain cancers (e.g., colorectal cancer and prostate cancer) have distinct microbial communities at the tumor site that are associated with tumor development [44, 45]. In fact, other data from our lab using cervical biopsy tissue samples indicates that there are distinct microbial differences as cervical cancer progresses to advanced FIGO stages (manuscript in preparation). This idea is further supported by data indicating that cervical precancerous lesions that regress, compared to those that progress to cancer, harbor a different immune microenvironment [46]. The local interplay between the microbiome and the local host immune system may be important to understanding the progression of HR-HPV infection to cervical cancer.
The protective microbial biomarkers identified at V1 also suggest an association of the microbiome and host innate and acquired immunity in progression to CIN2+. Specifically, the protective effect of bacterial Cell Motility may be due to the known phenomena of bacterial flagella activating host immunity [47–49]. This local activation may facilitate the innate immune system’s ability to clear an active HPV infection. Such stimulation may be critical in HPV control since cervical lesions have been shown to be associated with local immunosuppression through the reduction of factors such as IL-17 [50]. Despite the presence of studies to support this conjecture it should be noted that this type of immune activation needs to be confirmed with rigorous experimental precision.
Gardnerella, as discussed above, continuously emerges as a risk factor for CIN2+ development and progression. Based on our findings and published data, the association may be tied to the ability of Gardnerella to be immunosuppressive in the cervicovaginal region [51]. Whereas, it seems that the presence of commensal bacteria (e.g. Lactobacillus) with the ability to stimulate a local immune response may be contributing factors to the clearance of incident HR-HPV infections. Moreover, the presence of bacteria with immunosuppressive attributes, such as Gardnerella, may promote viral persistence and progression.
Alternatively, there may be other explanations for the observed associations between the cervicovaginal microbiome and HPV’s natural history. One possible explanation is a host developed or inherited immune deficiency that is a common cofactor for both cervical cancer progression and microbial diversity. For example, elevation of a particular inflammatory cytokine may be both necessary for successful tumor growth and be a causal factor in increasing vaginal microbial diversity. Such a factor may also explain the consistent identification of Gardnerella, which is commonly identified as a biomarker for increased diversity in the CVM [52] and a risk factor for CIN2+.
We have identified distinct microbial biomarkers that either protect, or promote the progression of a HR-HPV infection to CIN2+ lesions. In the context of what is known about the cervicovaginal microbiome, it may be that these factors act to suppress (in the case of progression) or activate (in the case of clearance) a localized immune response, which in turn influences the natural history of a HR-HPV infection. However, additional prospective studies are needed to establish a causal link between the cervicovaginal microbiome, the immune system and the natural history of HPV. Nevertheless, our results suggest a marker for identifying women with persistent HR-HPV infection at risk for progression by monitoring the presence of Gardnerella and subsequent elevation in microbial diversity. If future studies support a causal role of the cervicovaginal microbiome and disease progression, then it might be possible to manipulate the CVM in a manner to activate a local immune response. It is possible that HPV vaccination might influence the CVM and future research will be needed to evaluate such potential changes.
The strength of this study includes the prospective design and availability of a longitudinal cohort. In addition we used advanced epidemiological methods in a novel way to investigate potential causative factors in cervical intraepithelial neoplasia. Potential weaknesses in this study include the relatively small sample size, homogeneity of the population and the use of only two time points.
In summary, through the use of longitudinal samples from the CVT cohort we investigated and identified key features of the cervicovaginal microbiome potentially associated with progression of HR-HPV infection [28, 53–56] (e.g., Gardnerella and subsequent increase vaginal microbial diversity). Additional studies are required to validate the model proposed in this report.
Materials and methods
Clinical trial information
The study of cervicovaginal microbiome and HR-HPV natural history was a nested analysis within the previously reported CVT [57] (clinical trials registration NCT00128661). Written informed consent was obtained from all participants in CVT. The trial protocol can be obtained from the original trial publication [57].
Ethics statement
All CVT participants were adult women between the ages of 18–25 years. All participants were shown a video describing the study design and were then required to provide written consent to continue participating in the trial. Institutional review board approval was obtained for the informed consent forms at both the NCI and in Costa Rica. Registered with Clinicaltrials.gov NCT00128661
Study population and case definitions
Subjects for this nested study were selected from the placebo arm of a community-based clinical trial of the HPV 16/18 vaccine in Costa Rica that had enrolled women 18 to 25 years of age in 2004–2005 [57]. Women with an incident HR-HPV infection (HPV16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, or 59) were selected for analysis. Incident infections were classified based on outcomes from the well-established model of HR-HPV natural history including outcomes of clearance, persistence and progression [58]. Specifically, outcomes related to the incident HR-HPV infection included women who developed a CIN2 or CIN3 (CIN2+) lesion (progression), women with an infection for 2 or more years with the same type in the absence of a CIN2+ diagnosis (persistent), or women who cleared their incident HR-HPV infections within 1 year (clearance). This analysis included 273 women of whom all had available samples at V1 (first visit positive for the studied HPV type) and 266 who had a second sample at V2 (for persistent, visit that was positive for the same type and at least 305 days after V1; progression, closest visit before diagnosis of CIN2+; clearance, following visit that was negative for that type); all had clinical follow-up data. Seven women, one with clearance, and six with persistence either did not have an available V2 sample or the sample failed in lab testing.
Cervical microbiome characterization
DNA samples [59] were shipped to the Burk Lab on dry ice where the microbiome analysis was performed. DNA had been extracted from cervical brush samples by DDL Diagnostic Laboratory (Voorberg, The Netherlands) where they had been tested for HPV as previously described [60]. Laboratory procedures for the microbiome analyses were performed within a hood (AirClean Systems, Creedmoor, NC) in an isolated room to minimize environmental contamination and water-blank negative controls were used throughout the testing.
Bacterial DNA was amplified using barcoded-primers 16SV4_515F (GTGYCAGCMGCCGCGGTA) and 16SV4_806R (GGACTACHVGGGTWTCTAAT) that amplify the V4 variable region of the 16S rRNA gene [61]. This region has been demonstrated to accurately amplify and resolve vaginal bacteria [62]. PCR reactions were performed with 17.75 μl of nuclease-free PCR-grade water (Lonza, Rockland, ME), 2.5 μl of 10X Buffer w/ MgCl2 (Affymetrix, Santa Clara, CA), 1 μl of MgCl2 (25 mM, Affymetrix, Santa Clara, California, USA), 0.5 μl of dNTPs (10 mM, Roche, Basel, Switzerland), 0.25 μl of AmpliTaq Gold DNA Polymerase (5 U/μl, Applied Biosystems, Foster City, California), 0.5 μl of HotStart-IT FideliTaq (2.5 U/μl, Affymetrix, Santa Clara, CA), 1 μl of each primer (5 μM), and 0.5 μl of sample DNA. Thermal cycling conditions consisted of initial denaturation at 95°C for 5 min, followed by 15 cycles at 95°C for 1 min, 55°C for 1 min, and 68°C for 1 min, followed by 15 cycles at 95°C for 1 min, 60°C for 1 min, and 68°C for 1 min, and a final extension for 10 min at 68°C on a GeneAmp PCR System 9700 (Applied Biosystems, Foster City, CA).
The fungal DNA ITS1 region was amplified using barcoded-primers ITS1_48F (ACACACCGCCCGTCGCTACT) and ITS1_217R (TTTCGCTGCGTTCTTCATCG) as previously described [63]. PCR reactions were performed with 8.25 μl of nuclease-free PCR-grade water (Lonza), 2.5 μl of 10X Buffer w/ MgCl2 (Affymetrix), 1 μl of MgCl2 (25 mM, Affymetrix), 0.5 μl of dNTPs (10 mM, Roche), 0.25 μl of AmpliTaq Gold DNA Polymerase (5 U/μl, Applied Biosystems), 0.5 μl of HotStart-IT FideliTaq (2.5 U/μl, Affymetrix), 1μl of each primer (5 μM), and 10 μl of sample DNA. Thermal cycling conditions consisted of initial denaturation of 95°C for 3 min, followed by 35 cycles at 95°C for 30 sec, 55°C for 30 sec, and 68°C for 2 min, followed by a final extension for 10 min at 68°C on a GeneAmp PCR System 9700 (Applied Biosystems).
For both amplicon experiments, 20 negative controls were randomly mixed amongst samples. Negative controls were created using nuclease-free PCR-grade water (Lonza) as described above instead of extracted DNA.
Barcoded-PCR products were combined for each amplicon type and the DNA fragments (~356 bp for 16S V4 and 400–600 for ITS1) were isolated by gel purification using a QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany). Purified PCR products were quantified using a Qubit 2.0 Fluorometic High Sensitivity dsDNA Assay (Life Technologies, Carlsbad, CA) prior to library construction using a KAPA LTP Library Preparation Kit (Kapa Biosystems, Wilmington, MA). Size integrity of the amplicons was validated with a 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA). High-throughput amplicon sequencing of 2x300 paired-end reads was conducted on an Illumina MiSeq (Illumina, San Diego, CA).
Bioinformatics
Illumina reads were trimmed to remove bases that had a PHRED score of <25 using prinseq-lite V0.0.4 [64]. Quality trimmed reads were then demultiplexed using Novobarcode [65]. Paired-end reads were joined using PANDAseq with default settings [66]. The merged reads were processed through the VSEARCH quality-filtering pipeline [67] to dereplicate the sequences, reduce noise and remove chimeric reads.
For bacterial 16S V4 rRNA gene reads, closed-reference OTU picking was performed using VSEARCH [67] with a custom database that contained sequences from the GreenGenes database [68] the Human Oral Microbiome Database (HOMD) [69] and cervicovaginal microbiome 16S reference sequences retrieved from NCBI [70]. Representative sequences were aligned using PyNAST [71] and taxonomy was assigned using VSEARCH [67].
PICRUSt was used to impute microbial functional gene content and to collapse identified genes into functional pathways [72]. Pathways that were associated with HR-HPV clearance were identified through the use of a generalized linear model (GLM) based on statistical significance (p<0.05) and relative abundance (1% or higher across all reads).
For fungal ITS1 reads, open-reference OTU picking was performed using VSEARCH [67] and the UNITE database [73]. Taxonomy of representative fungal sequences was assigned using BLAST [74].
Phyloseq [75] was used to import BIOM data for 16S and ITS assays into R separately, followed by the determination of Shannon and Chao1 alpha diversity. For all analyses, bacterial data was subsampled for 2,500 reads. For fungal analyses subsampling was performed at 500 reads. Biomarker discovery analysis was performed using the LEfSe tool [76]. Linear discriminant analysis (LDA) scores greater than 2.0 are considered to be significant [76]. Microbial community state types (CSTs) were assigned on the basis of hierarchical clustering of the 20 most abundant OTUs. Prior to clustering, OTUs were agglomerated at the species level or the lowest identified taxonomic level. Clustering was performed using the wardD2 algorithm using Euclidian distances.
Statistical analysis
R v3.4.2 [77] was used for statistical analyses. The Kruskal-Wallis test was used to assess significance of continuous data. Linear regression was used to assess the significance of variables associated with the ordered outcome states of a HR-HPV infection (1). Logistic regression was performed using the GLM function and a binomial family generalized linear model in R. For categorical data, dummy variables were created and each individual factor level was tested in a univariate GLM analysis. Models were adjusted for age, smoking, HPV16 and CSTs. Smoking status was determined through a questionnaire and incorporated into the model as ordered categories: never smoked, former smoker and current smoker [40]. Power of GLM results was computed using the lmSupport package [78].
We performed a statistical mediation analysis to test whether V1 Gardnerella (an independent variable) could be acting by inducing a subsequent elevated microbiome diversity at V2 (mediator variable) that influences the outcome of HR-HPV progression using the package mediation [79]. The outcome model we used was binary clearance/progression. Models were adjusted for age, CST, smoking status and HPV16 infection status. In the results we present the mediation effect (average causal mediation effects (ACME)), which is the total effect of V2 Shannon diversity and V1 Gardnerella minus the direct effect of V1 Gardnerella. Additionally, we estimate the direct of effect (presented using the average direct effect (ADE)) of V1 Gardnerella on the binary outcome clearance/progression, minus the effect of the V2 Shannon diversity mediator; the total effect, which is the sum between the indirect effect of the V2 Shannon diversity and the direct effect of the V1 Gardnerella; and the proportion mediated which is the ratio of the ACME and total effect estimates.
Investigators in the International Agency for Research on Cancer/World Health Organization
Where authors are identified as personnel of the International Agency for Research on Cancer/ World Health Organization, the authors alone are responsible for the views expressed in this article and they do not necessarily represent the decisions, policy or views of the International Agency for Research on Cancer / World Health Organization.
Investigators in the Costa Rica HPV Vaccine Trial (CVT) group
Proyecto Epidemiológico Guanacaste, Fundación INCIENSA, San José, Costa Rica—Bernal Cortés (specimen and repository manager), Paula González (LTFU: co-principal investigator), Rolando Herrero (CVT: co-principal investigator), Silvia E. Jiménez (trial coordinator), Carolina Porras (co-investigator), Ana Cecilia Rodríguez (co-investigator).
United States National Cancer Institute, Bethesda, MD, USA—Allan Hildesheim (co-principal investigator & NCI co-project officer), Aimée R. Kreimer (LTFU: co-principal investigator & NCI co-project officer), Douglas R. Lowy (HPV virologist), Mark Schiffman (CVT: medical monitor & NCI co-project officer), John T. Schiller (HPV virologist), Mark Sherman (CVT: QC pathologist), Sholom Wacholder (statistician).
Leidos Biomedical Research, Inc., Frederick National Laboratory for Cancer Research, Frederick, MD, USA (HPV Immunology Laboratory)—Ligia A. Pinto, Troy J. Kemp
Georgetown University, Washington, DC, USA—Mary K. Sidawy (CVT: histopathologist)
DDL Diagnostic Laboratory, Netherlands (HPV DNA Testing)—Wim Quint, Leen-Jan van Doorn, Linda Struijk.
University of California, San Francisco, CA, USA—Joel M. Palefsky (expert on anal HPV infection and disease diagnosis and management), Teresa M. Darragh (pathologist and clinical management)
University of Virginia, Charlottesville, VA, USA—Mark H. Stoler (QC pathologist)
Supporting information
(CSV)
(CSV)
(CSV)
(XLSX)
Panel A shows specific fungal taxa, identified as being significant with the three HR-HPV outcomes. Values that are higher than LDA score of 2.0 are considered to be significant. Panel B shows the main fungal taxa identified in panel A with their relative abundances. The box represents the median value (with the 25–75% confidence interval as the box and the 95% confidence interval with the whiskers) for the taxa in each outcome (shown in the three separate panels and labeled at the top of the panel). There is a statistically significant increase in the sum of the five progression associated taxa when going from clearance to persistence to progression, p = 0.0080. The y-axis is the log of the relative abundance.
(PDF)
The two pathways identified after PICRUSt that were significantly different between clearance and progression were analyzed to determine their correlation to each other. Plot shows a significant negative correlation (Pearson correlation = -0.80) between the two pathways and thus they are highly correlated.
(PDF)
Panel A shows the 16S bacterial Shannon diversity index in each cytology group. Panel B shows the relative abundance of Gardnerella across the cytology groups. Significance is shown above bar plots as indicated in the figure at the top right.
(PDF)
Acknowledgments
The NCI and Costa Rica investigators are responsible for the design and conduct of the study; collection, management, analysis, and interpretation of the data; and preparation of the manuscript. We extend a special thanks to the women of Guanacaste and Puntarenas, Costa Rica, who gave of themselves in participating in this effort. In Costa Rica, we acknowledge the tremendous effort and dedication of the staff involved in this project; we would like to specifically acknowledge the meaningful contributions by Carlos Avila, Loretto Carvajal, Rebeca Ocampo, Cristian Montero, Diego Guillen, Jorge Morales and Mario Alfaro. In the United States, we extend our appreciation to the team from Information Management Services (IMS) responsible for the development and maintenance of the data system used in the trial and who serve as the data management center for this effort, especially Jean Cyr, Julie Buckland, John Schussler, and Brian Befano. We thank Dr. Diane Solomon (CVT: medical monitor & QC pathologist) for her invaluable contributions during the randomized blinded phase of the trial and the design of the LTFU and Nora Macklin (CVT) and Kate Torres (LTFU) for the expertise in coordinating the study. We thank the members of the Data and Safety Monitoring Board charged with protecting the safety and interest of participants during the randomized, blinded phase of our study (Steve Self, Chair, Adriana Benavides, Luis Diego Calzada, Ruth Karron, Ritu Nayar, and Nancy Roach) and members of the external Scientific HPV Working Group who have contributed to the success of our efforts over the years (Joanna Cain and Elizabeth Fontham, Co-Chairs, Diane Davey, Anne Gershon, Elizabeth Holly, Silvia Lara, Henriette Raventós, Wasima Rida, Richard Roden, Maria del Rocío Sáenz Madrigal, Gypsyamber D’Souza, and Margaret Stanley).
Data Availability
Data cannot be shared publicly because of the use of confidential patient data. Data are available from the National Cancer Institute Institutional Data Access / Ethics Committee (contact via BefanoB@imsweb.com) for researchers who meet the criteria for access to confidential data.
Funding Statement
The Costa Rica HPV Vaccine Trial is a long-standing collaboration between investigators in Costa Rica and the NCI. The trial is sponsored and funded by the NCI (contract N01-CP-11005), with funding support from the National Institutes of Health Office of Research on Women's Health. GlaxoSmithKline Biologicals (GSK) provided vaccine and support for aspects of the trial associated with regulatory submission needs of the company under a Clinical Trials Agreement (FDA BB-IND 7920) during the four-year, randomized blinded phase of our study. MU, RDB, CPZ and AG were funded by the National Cancer Institute (NCI) (U01 CA78527). The original Costa Rica vaccine trial was sponsored and funded by the NCI (contract N01-CP-11005). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Schiffman M, Doorbar J, Wentzensen N, de Sanjose S, Fakhry C, Monk BJ, et al. Carcinogenic human papillomavirus infection. Nature reviews Disease primers. 2016;2:16086 Epub 2016/12/03. 10.1038/nrdp.2016.86 . [DOI] [PubMed] [Google Scholar]
- 2.Ho GY, Burk RD, Klein S, Kadish AS, Chang CJ, Palan P, et al. Persistent genital human papillomavirus infection as a risk factor for persistent cervical dysplasia. J Natl Cancer Inst. 1995;87(18):1365–71. 10.1093/jnci/87.18.1365 . [DOI] [PubMed] [Google Scholar]
- 3.Castle PE, Rodriguez AC, Burk RD, Herrero R, Wacholder S, Alfaro M, et al. Short term persistence of human papillomavirus and risk of cervical precancer and cancer: population based cohort study. BMJ. 2009;339:b2569 Epub 2009/07/30. 10.1136/bmj.b2569 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kjaer SK, Frederiksen K, Munk C, Iftner T. Long-term absolute risk of cervical intraepithelial neoplasia grade 3 or worse following human papillomavirus infection: role of persistence. J Natl Cancer Inst. 2010;102(19):1478–88. Epub 2010/09/16. 10.1093/jnci/djq356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ho G. Y., Burk R. D., Klein S., Kadish A. S., Chang C. J., Palan P., … & Romney S. (1995). Persistent genital human papillomavirus infection as a risk factor for persistent cervical dysplasia. JNCI: Journal of the National Cancer Institute, 87(18), 1365–1371. [DOI] [PubMed] [Google Scholar]
- 6.Castle PE, Wacholder S, Lorincz AT, Scott DR, Sherman ME, Glass AG, et al. A prospective study of high-grade cervical neoplasia risk among human papillomavirus-infected women. J Natl Cancer Inst. 2002;94(18):1406–14. Epub 2002/09/19. 10.1093/jnci/94.18.1406 . [DOI] [PubMed] [Google Scholar]
- 7.Roura E, Castellsague X, Pawlita M, Travier N, Waterboer T, Margall N, et al. Smoking as a major risk factor for cervical cancer and pre-cancer: results from the EPIC cohort. Int J Cancer. 2014;135(2):453–66. 10.1002/ijc.28666 . [DOI] [PubMed] [Google Scholar]
- 8.Plummer M, Herrero R, Franceschi S, Meijer CJ, Snijders P, Bosch FX, et al. Smoking and cervical cancer: pooled analysis of the IARC multi-centric case—control study. Cancer Causes Control. 2003;14(9):805–14. 10.1023/b:caco.0000003811.98261.3e . [DOI] [PubMed] [Google Scholar]
- 9.Eldridge RC, Pawlita M, Wilson L, Castle PE, Waterboer T, Gravitt PE, et al. Smoking and subsequent human papillomavirus infection: a mediation analysis. Ann Epidemiol. 2017. Epub 2017/11/07. 10.1016/j.annepidem.2017.10.004 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Smith JS, Green J, Berrington de Gonzalez A, Appleby P, Peto J, Plummer M, et al. Cervical cancer and use of hormonal contraceptives: a systematic review. Lancet. 2003;361(9364):1159–67. Epub 2003/04/11. 10.1016/s0140-6736(03)12949-2 . [DOI] [PubMed] [Google Scholar]
- 11.Moreno V, Bosch FX, Munoz N, Meijer CJ, Shah KV, Walboomers JM, et al. Effect of oral contraceptives on risk of cervical cancer in women with human papillomavirus infection: the IARC multicentric case-control study. Lancet. 2002;359(9312):1085–92. 10.1016/S0140-6736(02)08150-3 . [DOI] [PubMed] [Google Scholar]
- 12.Munoz N, Franceschi S, Bosetti C, Moreno V, Herrero R, Smith JS, et al. Role of parity and human papillomavirus in cervical cancer: the IARC multicentric case-control study. Lancet. 2002;359(9312):1093–101. 10.1016/S0140-6736(02)08151-5 . [DOI] [PubMed] [Google Scholar]
- 13.Stanley MA, Sterling JC. Host Responses to Infection with Human Papillomavirus. Curr Probl Dermatol. 2014;45:58–74. Epub 2014/03/20. 10.1159/000355964 . [DOI] [PubMed] [Google Scholar]
- 14.Doorbar J. Host control of human papillomavirus infection and disease. Best practice & research Clinical obstetrics & gynaecology. 2018;47:27–41. Epub 2017/09/19. 10.1016/j.bpobgyn.2017.08.001 . [DOI] [PubMed] [Google Scholar]
- 15.Leo PJ, Madeleine MM, Wang S, Schwartz SM, Newell F, Pettersson-Kymmer U, et al. Defining the genetic susceptibility to cervical neoplasia-A genome-wide association study. PLoS genetics. 2017;13(8):e1006866 10.1371/journal.pgen.1006866 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Habbous S, Pang V, Xu W, Amir E, Liu G. Human papillomavirus and host genetic polymorphisms in carcinogenesis: A systematic review and meta-analysis. J Clin Virol. 2014. 10.1016/j.jcv.2014.07.019 . [DOI] [PubMed] [Google Scholar]
- 17.Castle PE, Giuliano AR. Chapter 4: Genital tract infections, cervical inflammation, and antioxidant nutrients—assessing their roles as human papillomavirus cofactors. J Natl Cancer Inst Monogr. 2003;(31):29–34. Epub 2003/06/17. 10.1093/oxfordjournals.jncimonographs.a003478 . [DOI] [PubMed] [Google Scholar]
- 18.Pfeiffer JK. Host response: Microbiota prime antiviral response. Nature microbiology. 2016;1(2):15029. [DOI] [PubMed] [Google Scholar]
- 19.Vallianou NG, Tzortzatou-Stathopoulou F. Microbiota and cancer: an update. J Chemother. 2019;31(2):59–63. 10.1080/1120009X.2018.1541046 [DOI] [PubMed] [Google Scholar]
- 20.Rajagopala SV, Vashee S, Oldfield LM, Suzuki Y, Venter JC, Telenti A, et al. The human microbiome and cancer. Cancer Prevention Research. 2017;10(4):226–34. 10.1158/1940-6207.CAPR-16-0249 [DOI] [PubMed] [Google Scholar]
- 21.Vogtmann E, Hua X, Zeller G, Sunagawa S, Voigt AY, Hercog R, et al. Colorectal cancer and the human gut microbiome: reproducibility with whole-genome shotgun sequencing. PloS one. 2016;11(5):e0155362 10.1371/journal.pone.0155362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Champer M, Wong AM, Champer J, Brito IL, Messer PW, Hou JY, et al. The role of the vaginal microbiome in gynecological cancer: a review. BJOG: an international journal of obstetrics and gynaecology. 2017. Epub 2017/03/10. 10.1111/1471-0528.14631 . [DOI] [PubMed] [Google Scholar]
- 23.Kroon SJ, Ravel J, Huston WM. Cervicovaginal microbiota, women's health, and reproductive outcomes. Fertil Steril. 2018;110(3):327–36. 10.1016/j.fertnstert.2018.06.036 [DOI] [PubMed] [Google Scholar]
- 24.Kyrgiou M, Mitra A, Moscicki A-B. Does the vaginal microbiota play a role in the development of cervical cancer? Translational Research. 2017;179:168–82. 10.1016/j.trsl.2016.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ravel J, Gajer P, Abdo Z, Schneider GM, Koenig SS, McCulle SL, et al. Vaginal microbiome of reproductive-age women. Proc Natl Acad Sci U S A. 2011;108 Suppl 1:4680–7. Epub 2010/06/11. 10.1073/pnas.1002611107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Smith BC, Zolnik CP, Usyk M, Chen Z, Kaiser K, Nucci-Sack A, et al. Focus: Microbiome: Distinct Ecological Niche of Anal, Oral, and Cervical Mucosal Microbiomes in Adolescent Women. The Yale journal of biology and medicine. 2016;89(3):277 [PMC free article] [PubMed] [Google Scholar]
- 27.Bik EM, Bird SW, Bustamante JP, Leon LE, Nieto PA, Addae K, et al. A novel sequencing-based vaginal health assay combining self-sampling, HPV detection and genotyping, STI detection, and vaginal microbiome analysis. PloS one. 2019;14(5):e0215945 10.1371/journal.pone.0215945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Freitas AC, Bocking A, Hill JE, Money DM. Increased richness and diversity of the vaginal microbiota and spontaneous preterm birth. Microbiome. 2018;6(1):117 10.1186/s40168-018-0502-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Di C, Jin F. Value of combined detection of claudin 4 and high-risk human papilloma virus in high-grade squamous intraepithelial lesion and cervix squamous cell carcinoma. Zhejiang da xue xue bao Yi xue ban = Journal of Zhejiang University Medical sciences. 2018;47(4):344–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Audirac-Chalifour A, Torres-Poveda K, Bahena-Román M, Téllez-Sosa J, Martínez-Barnetche J, Cortina-Ceballos B, et al. Cervical microbiome and cytokine profile at various stages of cervical cancer: a pilot study. PloS one. 2016;11(4):e0153274 10.1371/journal.pone.0153274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dareng E, Ma B, Famooto A, Akarolo-Anthony S, Offiong R, Olaniyan O, et al. Prevalent high-risk HPV infection and vaginal microbiota in Nigerian women. Epidemiol Infect. 2016;144(1):123–37. 10.1017/S0950268815000965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gao W, Weng J, Gao Y, Chen X. Comparison of the vaginal microbiota diversity of women with and without human papillomavirus infection: a cross-sectional study. BMC infectious diseases. 2013;13(1):271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lee JE, Lee S, Lee H, Song Y-M, Lee K, Han MJ, et al. Association of the vaginal microbiota with human papillomavirus infection in a Korean twin cohort. PloS one. 2013;8(5):e63514 10.1371/journal.pone.0063514 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Reimers LL, Mehta SD, Massad LS, Burk RD, Xie X, Ravel J, et al. The cervicovaginal microbiota and its associations with human papillomavirus detection in HIV-infected and HIV-uninfected women. The Journal of infectious diseases. 2016;214(9):1361–9. 10.1093/infdis/jiw374 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Audirac-Chalifour A, Torres-Poveda K, Bahena-Roman M, Tellez-Sosa J, Martinez-Barnetche J, Cortina-Ceballos B, et al. Cervical Microbiome and Cytokine Profile at Various Stages of Cervical Cancer: A Pilot Study. PLoS One. 2016;11(4):e0153274 10.1371/journal.pone.0153274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Palma E, Recine N, Domenici L, Giorgini M, Pierangeli A, Panici PB. Long-term Lactobacillus rhamnosus BMX 54 application to restore a balanced vaginal ecosystem: a promising solution against HPV-infection. BMC infectious diseases. 2018;18(1):13 10.1186/s12879-017-2938-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Piyathilake CJ, Ollberding NJ, Kumar R, Macaluso M, Alvarez RD, Morrow CD. Cervical Microbiota Associated with Risk of Higher Grade Cervical Intraepithelial Neoplasia in Women Infected with High-Risk Human Papillomaviruses. Cancer Prev Res (Phila). 2016. Epub 2016/03/05. 10.1158/1940-6207.capr-15-0350 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mitra A, MacIntyre DA, Lee YS, Smith A, Marchesi JR, Lehne B, et al. Cervical intraepithelial neoplasia disease progression is associated with increased vaginal microbiome diversity. Scientific reports. 2015;5:16865 10.1038/srep16865 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Seo SS, Oh HY, Lee JK, Kong JS, Lee DO, Kim MK. Combined effect of diet and cervical microbiome on the risk of cervical intraepithelial neoplasia. Clin Nutr. 2016. 10.1016/j.clnu.2016.03.019 . [DOI] [PubMed] [Google Scholar]
- 40.Hildesheim A, Herrero R, Wacholder S, Rodriguez AC, Solomon D, Bratti MC, et al. Effect of human papillomavirus 16/18 L1 viruslike particle vaccine among young women with preexisting infection: a randomized trial. JAMA. 2007;298(7):743–53. Epub 2007/08/21. 10.1001/jama.298.7.743 . [DOI] [PubMed] [Google Scholar]
- 41.Zhang C, Liu Y, Gao W, Pan Y, Gao Y, Shen J, et al. The direct and indirect association of cervical microbiota with the risk of cervical intraepithelial neoplasia. Cancer medicine. 2018. Epub 2018/04/03. 10.1002/cam4.1471 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Takač I, Marin J, Gorišek B. Human papillomavirus 16 and 18 infection of the uterine cervix in women with different grades of cervical intraepithelial neoplasia (CIN). International Journal of Gynecology & Obstetrics. 1998;61(3):269–73. [DOI] [PubMed] [Google Scholar]
- 43.Di Paola M, Sani C, Clemente AM, Iossa A, Perissi E, Castronovo G, et al. Characterization of cervico-vaginal microbiota in women developing persistent high-risk Human Papillomavirus infection. Scientific reports. 2017;7(1):10200 10.1038/s41598-017-09842-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Louis P, Hold GL, Flint HJ. The gut microbiota, bacterial metabolites and colorectal cancer. Nature Reviews Microbiology. 2014;12(10):661 10.1038/nrmicro3344 [DOI] [PubMed] [Google Scholar]
- 45.Sfanos KS, Yegnasubramanian S, Nelson WG, De Marzo AM. The inflammatory microenvironment and microbiome in prostate cancer development. Nature Reviews Urology. 2018;15(1):11 10.1038/nrurol.2017.167 [DOI] [PubMed] [Google Scholar]
- 46.Øvestad IT, Gudlaugsson E, Skaland I, Malpica A, Kruse A-J, Janssen EA, et al. Local immune response in the microenvironment of CIN2–3 with and without spontaneous regression. Mod Pathol. 2010;23(9):1231 10.1038/modpathol.2010.109 [DOI] [PubMed] [Google Scholar]
- 47.Ramos HC, Rumbo M, Sirard J-C. Bacterial flagellins: mediators of pathogenicity and host immune responses in mucosa. Trends Microbiol. 2004;12(11):509–17. 10.1016/j.tim.2004.09.002 [DOI] [PubMed] [Google Scholar]
- 48.Vijay-Kumar M, Gewirtz A. Flagellin: key target of mucosal innate immunity. Mucosal immunology. 2009;2(3):197 10.1038/mi.2009.9 [DOI] [PubMed] [Google Scholar]
- 49.Cullender TC, Chassaing B, Janzon A, Kumar K, Muller CE, Werner JJ, et al. Innate and adaptive immunity interact to quench microbiome flagellar motility in the gut. Cell host & microbe. 2013;14(5):571–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Gosmann C, Mattarollo SR, Bridge JA, Frazer IH, Blumenthal A. IL-17 Suppresses Immune Effector Functions in Human Papillomavirus-Associated Epithelial Hyperplasia. J Immunol. 2014. 10.4049/jimmunol.1400216 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Murphy K, Mitchell CM. The interplay of host immunity, environment and the risk of bacterial vaginosis and associated reproductive health outcomes. The Journal of infectious diseases. 2016;214(suppl_1):S29–S35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Schellenberg JJ, Patterson MH, Hill JE. Gardnerella vaginalis diversity and ecology in relation to vaginal symptoms. Res Microbiol. 2017;168(9–10):837–44. 10.1016/j.resmic.2017.02.011 [DOI] [PubMed] [Google Scholar]
- 53.Arokiyaraj S, Seo SS, Kwon M, Lee JK, Kim MK. Association of cervical microbial community with persistence, clearance and negativity of Human Papillomavirus in Korean women: a longitudinal study. Scientific reports. 2018;8(1):15479 Epub 2018/10/21. 10.1038/s41598-018-33750-y . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Godoy-Vitorino F, Romaguera J, Zhao C, Vargas-Robles D, Ortiz-Morales G, Vazquez-Sanchez F, et al. Cervicovaginal Fungi and Bacteria Associated With Cervical Intraepithelial Neoplasia and High-Risk Human Papillomavirus Infections in a Hispanic Population. Front Microbiol. 2018;9:2533 Epub 2018/11/09. 10.3389/fmicb.2018.02533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kwasniewski W, Wolun-Cholewa M, Kotarski J, Warchol W, Kuzma D, Kwasniewska A, et al. Microbiota dysbiosis is associated with HPV-induced cervical carcinogenesis. Oncology letters. 2018;16(6):7035–47. Epub 2018/12/14. 10.3892/ol.2018.9509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Suehiro TT, Malaguti N, Damke E, Uchimura NS, Gimenes F, Souza RP, et al. Association of human papillomavirus and bacterial vaginosis with increased risk of high-grade squamous intraepithelial cervical lesions. International journal of gynecological cancer: official journal of the International Gynecological Cancer Society. 2019. Epub 2019/01/12. 10.1136/ijgc-2018-000076 . [DOI] [PubMed] [Google Scholar]
- 57.Herrero R, Hildesheim A, Rodríguez AC, Wacholder S, Bratti C, Solomon D, et al. Rationale and design of a community-based double-blind randomized clinical trial of an HPV 16 and 18 vaccine in Guanacaste, Costa Rica. Vaccine. 2008;26(37):4795–808. 10.1016/j.vaccine.2008.07.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Schiffman M, Wentzensen N. Human papillomavirus infection and the multistage carcinogenesis of cervical cancer. Cancer Epidemiol Biomarkers Prev. 2013;22(4):553–60. 10.1158/1055-9965.EPI-12-1406 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hildesheim A, Wacholder S, Catteau G, Struyf F, Dubin G, Herrero R, et al. Efficacy of the HPV-16/18 vaccine: final according to protocol results from the blinded phase of the randomized Costa Rica HPV-16/18 vaccine trial. Vaccine. 2014;32(39):5087–97. 10.1016/j.vaccine.2014.06.038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Herrero R, Wacholder S, Rodríguez AC, Solomon D, González P, Kreimer AR, et al. Prevention of persistent human papillomavirus infection by an HPV16/18 vaccine: a community-based randomized clinical trial in Guanacaste, Costa Rica. Cancer discovery. 2011;1(5):408–19. 10.1158/2159-8290.CD-11-0131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Huntley J, Fierer N, et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. The ISME journal. 2012;6(8):1621 10.1038/ismej.2012.8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Van Der Pol WJ, Kumar R, Morrow CD, Blanchard EE, Taylor CM, Martin DH, et al. In silico and experimental evaluation of primer sets for species-level resolution of the vaginal microbiota using 16s ribosomal rna gene sequencing. The Journal of infectious diseases. 2018;219(2):305–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Usyk M, Zolnik CP, Patel H, Levi MH, Burk RD. Novel ITS1 fungal primers for characterization of the mycobiome. mSphere. 2017;2(6):e00488–17. 10.1128/mSphere.00488-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Schmieder R, Edwards R. Quality control and preprocessing of metagenomic datasets. Bioinformatics. 2011;27(6):863–4. Epub 2011/02/01. 10.1093/bioinformatics/btr026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Hercus C. Novocraft short read alignment package. Website http://www.novocraft.com. 2009. [Google Scholar]
- 66.Masella AP, Bartram AK, Truszkowski JM, Brown DG, Neufeld JD. PANDAseq: paired-end assembler for illumina sequences. BMC Bioinformatics. 2012;13:31 Epub 2012/02/16. 10.1186/1471-2105-13-31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Rognes T, Flouri T, Nichols B, Quince C, Mahé F. VSEARCH: a versatile open source tool for metagenomics. PeerJ. 2016;4:e2584 10.7717/peerj.2584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.DeSantis TZ, Hugenholtz P, Larsen N, Rojas M, Brodie EL, Keller K, et al. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl Environ Microbiol. 2006;72(7):5069–72. 10.1128/AEM.03006-05 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chen HS, Bromberg-White J, Conway MJ, Alam S, Meyers C. Study of infectious virus production from HPV18/16 capsid chimeras. Virology. 2010;405(2):289–99. 10.1016/j.virol.2010.05.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Coordinators NR. Database resources of the national center for biotechnology information. Nucleic acids research. 2017;45(Database issue):D12 10.1093/nar/gkw1071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. QIIME allows analysis of high-throughput community sequencing data. Nature methods. 2010;7(5):335–6. 10.1038/nmeth.f.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Langille MG, Zaneveld J, Caporaso JG, McDonald D, Knights D, Reyes JA, et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat Biotechnol. 2013;31(9):814 10.1038/nbt.2676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kõljalg U, Larsson KH, Abarenkov K, Nilsson RH, Alexander IJ, Eberhardt U, et al. UNITE: a database providing web‐based methods for the molecular identification of ectomycorrhizal fungi. New Phytologist. 2005;166(3):1063–8. 10.1111/j.1469-8137.2005.01376.x [DOI] [PubMed] [Google Scholar]
- 74.Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. Journal of molecular biology. 1990;215(3):403–10. 10.1016/S0022-2836(05)80360-2 [DOI] [PubMed] [Google Scholar]
- 75.McMurdie PJ, Holmes S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PloS one. 2013;8(4):e61217 10.1371/journal.pone.0061217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, et al. Metagenomic biomarker discovery and explanation. Genome biology. 2011;12(6):R60 10.1186/gb-2011-12-6-r60 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Team RC. R: A language and environment for statistical computing. 2013. [Google Scholar]
- 78.Curtin J. lmSupport: support for linear models. R package version. 2015;2(2). [Google Scholar]
- 79.Tingley D, Yamamoto T, Hirose K, Keele L, Imai K. Mediation: R package for causal mediation analysis. 2014. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(CSV)
(CSV)
(CSV)
(XLSX)
Panel A shows specific fungal taxa, identified as being significant with the three HR-HPV outcomes. Values that are higher than LDA score of 2.0 are considered to be significant. Panel B shows the main fungal taxa identified in panel A with their relative abundances. The box represents the median value (with the 25–75% confidence interval as the box and the 95% confidence interval with the whiskers) for the taxa in each outcome (shown in the three separate panels and labeled at the top of the panel). There is a statistically significant increase in the sum of the five progression associated taxa when going from clearance to persistence to progression, p = 0.0080. The y-axis is the log of the relative abundance.
(PDF)
The two pathways identified after PICRUSt that were significantly different between clearance and progression were analyzed to determine their correlation to each other. Plot shows a significant negative correlation (Pearson correlation = -0.80) between the two pathways and thus they are highly correlated.
(PDF)
Panel A shows the 16S bacterial Shannon diversity index in each cytology group. Panel B shows the relative abundance of Gardnerella across the cytology groups. Significance is shown above bar plots as indicated in the figure at the top right.
(PDF)
Data Availability Statement
Data cannot be shared publicly because of the use of confidential patient data. Data are available from the National Cancer Institute Institutional Data Access / Ethics Committee (contact via BefanoB@imsweb.com) for researchers who meet the criteria for access to confidential data.