

Abstract
Influenza is a major public health concern, infecting 5–15% of the global population annually. Influenza virus belongs to family Orthomyxoviridae, and has three types A, B and C. Infection by influenza virus A is most common and severe, generally found in humans. It spreads rapidly and affects human population across large geographical region within short period of time with varying degree of pathology from mild to severe. Wild aquatic birds and other animal species like birds, pigs, ferret, horses, seals, whales, mink, giant anteaters, cats and dogs are the reservoir for the influenza A virus. Influenza B and C viruses have very limited host range and appear predominantly in humans. Influenza virus gains pandemic potential through genetic reassortment called “genetic shift” with complete renewal of surface antigen and a small but gradual genetic change by mutations which make it to adapt efficiently in human population called “genetic drift”. Although, the epidemiology related to influenza infection has been studied from several years but some facts associated to disease transmission has poorly understood. This article reviews the important aspects of virological, epidemiological and clinical features related to influenza virus for better understanding of disease transmission and its pathogenesis.
Keywords: Influenza virus, H1N1, Pathogenesis, Epidemiology, Pandemic, Swine flu
Introduction
Influenza virus infection is a highly contagious airborne disease that causes acute febrile respiratory illness and results in variable degrees of systemic symptoms, ranging from mild fatigue to respiratory failure and death. Generally, it occurs during winter season of both Northern and Southern hemispheres. It leads to high morbidity rates for people of all ages and high mortality rate particularly for children and adults over 60 years old, patients with chronic illnesses and pregnant women [1–4]. A report by World Health Organization shows that there are 3–5 million severe influenza cases and 250,000–500,000 mortality every year.
Influenza virus belongs to family Orthomyxoviridae, and has three types A, B and C. Infection by influenza virus A is most common and severe, generally found in humans [5, 6]. Wild aquatic birds are the primary reservoir for the influenza A virus, in which infection is largely intestinal, waterborne, and asymptomatic. It also naturally infect a variety of other animal species like birds, pigs, ferret, horses, seals, whales, mink, giant anteaters, cats and dogs. Influenza B and C viruses have very limited host range and appear predominantly in humans. Type B influenza is usually associated with a mild form of the disease and is more common in the pediatric population. Infection with type C is infrequently seen (Table 1).
Table 1.
Properties of the subtypes of influenza virus
| Properties | A | B | C |
|---|---|---|---|
| Occurrence | Most frequent | Very low | Rare |
| Severity | Virulent | Mild | Not virulent |
| Outbreaks | Pandemic/epidemic [6] | Epidemic [6] | Sporadic [6] |
| Mutation & recombination rate | High (antigenic shift and antigenic drift) | Low (only antigenic drift) | No recombination |
| Reservoir | Wild aquatic bird | No | No |
| Host range | Wide (Human, Birds, Horse, Pig, Ferret, mice) | Limited to human | Only in humans |
| Subtypes | Many with different combination of 16 HA & 9 NA | No subtypes | No subtypes |
| RNA genome | Consist of eight segments | Eight segments | Seven segments |
Recent pandemic of swine flu infection (H1N1) in 2009 has greatly affected human population worldwide which was the first devastating pandemic of twenty-first century, caused high morbidity and mortality in humans. According to WHO, approximately 526,060 cases of pandemic H1N1/09 infection and at least 6770 deaths were reported all over the world by 15 November 2009 [7]. This has suddenly arisen in North America and spread rapidly in Europe, Asia and South Africa via human to human transmission within a short period of time. Influenza virus gains pandemic potential through genetic reassortment called “genetic shift” with complete renewal of surface antigen. A small but gradual genetic change also occurs by mutations which make it to adapt efficiently in human population called “genetic drift”. Pig acts as a reservoir for influenza virus and doesn’t show any significant respiratory illness against H1N1 infection while it is fatal in other mammalian species (human, monkeys, ferrets, macaques and mice). This review will focus the basic virology related to current 2009 pandemic virus, epidemiology, clinical features, diagnosis, treatment and vaccines which would help in understanding the pathogenesis associated to influenza virus. It would also highlight the molecular determinants of H1N1 virus that causes immunopathogenesis in human during infection and host’s immune response against swine flu virus (H1N1) that enhances pathogenesis/severity of infection (leading to the death of host).
Influenza Virus
All three subtypes of influenza virus (A, B, C) are similar in structure but vary antigenically. Influenza viruses are about 80–120 nm in size, roughly spherical/ovoid in shape. All contain single-stranded, negative-sense, segmented RNA genome (having eight segments of variable length). These encodes 11 proteins including nine structural [Polymerase proteins—polymerase basic protein 1 (PB1), PB1-F2, PB2 and polymerase acidic protein (PA), surface glycoproteins—haemagglutinin (HA) and neuraminidase (NA), Nucleocapsid protein—nucleoproteins (NP), Matrix proteins—M1, Ion pore protein—M2] and two nonstructural proteins (NS1 and NS2) respectively [8].
Structurally, virus is enclosed by outermost envelope made up of lipid bilayer from host membrane that consists of two surface glycoproteins, HA and NA which act as antigenic determinants and induce humoral immune response in host (Fig. 1). HA is mainly responsible for virus attachment to target cells and entry of viral genome into the host cell while NA participates in release of progeny virus particles from infected host cells by cleaving terminal sialic acid residue on host cell surface. Influenza A virus is classified into subtypes on the basis of these surface glycoproteins. To date, 16 serologically distinct HA and 9 NA have been identified and are sequentially named as H1, H2, H3 and so on and N1, N2……etc. Currently, H1, H2 and H3 in combination with N1 and N2 are most frequently circulating types in human. Remaining subtypes remain to be zoonotic, causing disease mainly in fowl and nonhuman primates [5, 6]. Viral proteins encoded by segmented RNA genome have various functions biological as well as in pathogenesis during infection. The three largest RNA segments encode proteins PA, PB1 and PB2, jointly act as a RNA dependent RNA polymerase (RdRP). PB1 gene segment also encode small 87 residue nonstructural protein PB1-F2 having apoptotic function [9]. Surface proteins HA, NA and NP are encoded by intermediate sized RNA segments. Remaining RNA segments encode M1 matrix protein and M2 ion pore protein. Two nonstructural proteins NS1A and NS2/NEP are encoded by very small RNA segments, functions of all these proteins are shown in Table 2. Influenza virus make entry into host by attachment to surface glycoproteins containing sialic acid residue on host respiratory epithelium cells with help of viral surface HA protein as shown in Fig. 2. To date, human influenza viruses isolated from 1957 and 1968 pandemics and recent pandemic swine origin influenza virus A (SO-IVA) H1N1 virus preferentially bind to cell-surface glycoproteins or glycolipids containing terminal 2-6 linked sialyl-galactosyl moieties [Neu5Ac(α2-6)Gal] while Avian influenza virus prefer α2-3 linked glycan sialic acid [Neu5Ac(@2-3)Gal] [10]. Recent studies depict that pandemic 2009 H1N1 (SO-IVA) virus is well adapted to mammalian hosts and binds to both α2,6-linked and α2,3-linked cellular receptors, which are present in conjunctiva, distal airways, and alveolar pneumocytes [11]. The viral HA molecules found in the form of trimers play another role in release of viral ribo-nucleoprotein particles (vRNP) into the cytoplasm [12]. The HA precursor (HA0) contains two domain, one sialic acid receptor binding domain (HA1) and remaining ectodomain (HA2) also called as fusion peptide, both linked by disulfide linkage. Virus is internalized within endosome through the mechanism of receptor mediated endocytosis. After internalization, M2 viral protein (Ion channel protein) play role in acidification of endosome by enhancing proton transport from cytoplasm. Hence, acidification of the endosomes facilitate irreversible conformational changes in HA leads to dissociation of HA1 from the endosomal membrane and enable HA2, a fusion peptide to attach to the endosomal membrane. All this process initiates the fusion of the viral and endosomal membranes, resulting in the release of the RNP into the cytoplasm [13, 14].
Fig. 1.
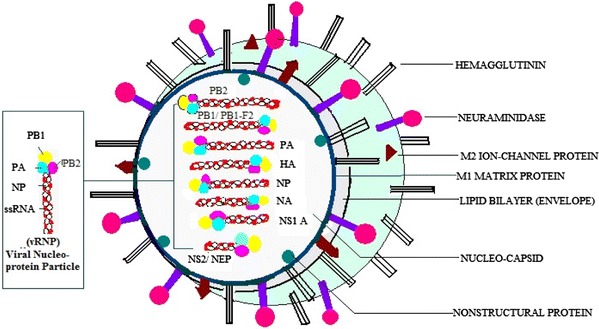
Structure of influenza virus
Table 2.
Viral proteins & its functions in pathogenesis
| S. no. | Viral proteins | Function |
|---|---|---|
| 1 | HA (haemagglutinin) | Viral entry into target cell |
| 2 | NA (neuraminidase) | Release of viral particles from target cells and help in dissemination of virion particles throughout respiratory tract |
| 3 | PA (acidic polymerase protein) | Endonuclease activity and help in cap snatching mechanism |
| 4 | PB1 (basic polymerase protein-1) | Viral mRNA transcription by 5′ cap snatching mechanism |
| 5 | PB2 (basic polymerase protein-2) | Unprimed replication of viral mRNA |
| 6 | PB1-F2 | Apoptosis of host cell |
| 7 | M1 (matrix protein) | Role in attachment of vRNP to cell membrane & provide stability |
| 8 | M2 (ion-channel protein) | Help in releasing of vRNP from endosome to cytoplasm |
| 9 | NP (nucleoprotein particle) | Participate in the nuclear import and export of vRNP & viral replication |
| 10 | NS1 (nonstructural protein-1) | Suppress IFN-β and host protein production |
| 11 | NS2 (nonstructural protein-2) | Help in nuclear import of vRNP |
Fig. 2.
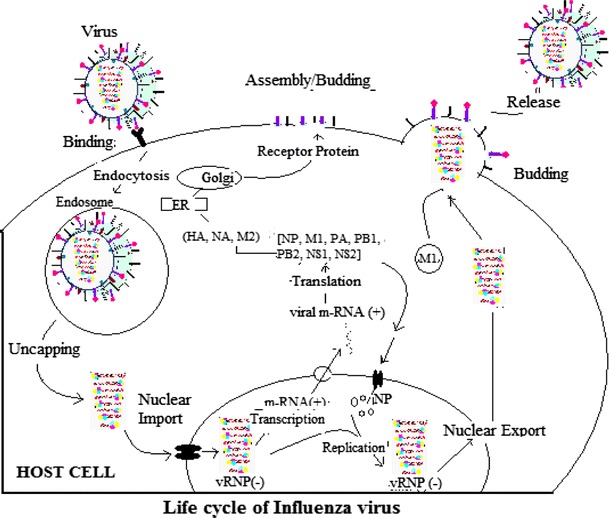
Life cycle of influenza virus A
A number of X-ray crystallographic and NMR studies revealed that M2 protein is 97 residues long and has a transmembrane (TM) domain and a C-terminal cytoplasmic amphipilic helix. The tetramers of TM helices are arranged in a specific way to form a proton channel [15]. According to A. Okada et al., the highly conserved residues His37 and Trp41 of TM helices located in the proton channel are critical in the proton transport process. His37 is protonated at low pH which triggers proton flow [16] and at high pH Trp41 residues positioned adjacent to His37, are clustered to form a channel gate that blocks the proton channel. At low pH, this gate opens up in association with the rearrangement of TM helices [10]. In future, new therapeutic drugs may be designed that target these two conserved amino acid residues which interfere in the arrangement of TM helices and in turn block proton transport in endosome and indirectly block viral replication in host cell. The vRNP attached to the lipid bilayer membrane via M1 matrix protein are released into cytoplasm through M2 channels due to influx of protons from endosome to virus. The released vRNP are transported into nucleus by recognition of the NLSs sequences (nuclear localization sequences) on NP [17]. Multiple copies of NP molecules are complexed with viral RNA genome and polymerase complex and also participate in export of vRNPs and viral replication and interact with host proteins [18]. In nucleus, viral polymerase initiates viral transcription via “cap snatching mechanism” in which PB2 of viral polymerase complex binds to 5′ cap (m7GTP) of a host pre-mRNA by its cap binding domain (PB2cap) and PA polymerase subunit (that has endonuclease and protease activities) [19], cleave a phosphodiester bond 10–13 nucleotides downstream of the cap and initiates viral m-RNA transcription at the cleaved 3′ end of the capped segment) [20–22]. These nascent viral m-RNAs are transported to the cytoplasm for translation into viral proteins (HA, M2 and NA) that are processed in the endoplasmic reticulum (ER), glycosylated in the Golgi apparatus and transported to the cell membrane. The P complex also replicates viral RNAs by different mechanism called unprimed replication in two steps [(−) vRNA → (+) cRNA → (−) vRNA] which require NP molecules that is deposited on the cRNA to form vRNP. This newly synthesized vRNP exported to the cytoplasm through M2–NS2 complex. The exported vRNAs reach the cell membrane and to be incorporated into new viruses that are budded out and then disseminated by the activity of NA [11].
Epidemiology
Influenza infection spreads around the world as seasonal epidemic every year. While, pandemic occurs rarely every 10–50 years, through which a new version of the influenza virus emerges in human population causing serious outbreak of disease. Pandemics lead to widespread illness and death even in otherwise healthy people who do not have any prior immunity against newly emerged antigenically and genetically novel influenza virus, responsible for social disruption and economic loss. The first pandemic was documented in sixteenth century [23]. In the last 400 years, at least 31 pandemics have been recorded [24]. Three major pandemics have been documented during the twentieth century, mortality impact of which ranged from devastating to moderate or mild—the “Spanish” flu in 1918–1919 (caused by a H1N1 subtype of influenza virus A), the “Asian” flu in 1957–1958 (H2N2 subtype), and the “Hong Kong” flu in 1968–1969 (H3N2 subtype) [25] (Table 3). The 1918 influenza A pandemic H1N1 was particularly severe with high death rate among young adults than Asian and Hong Kong flu. The overall attack rate and the age distribution of cases were not dramatically different from those observed during other pandemics of sixteenth century [26]. The 1957 pandemic was caused by H3N2 influenza virus and originated in China with highest attack rate (>50%) in young children aged 5–19. Genetic analysis studies revealed that it was an assortment virus possessing five genes similar to 1918 H1N1 virus and remaining three acquired from avian influenza virus (H2, N2 and PB1). This reassortment event was followed by replacement of H2 and PB1 gene with avian H3 and PB1 genes to create the 1968 Hong Kong influenza (H3N2) virus [27]. Recently, the incident of 2009 pandemic of twenty-first century occurred worldwide by H1N1 influenza virus as a result of multiple reassortment events including avian, human and swine influenza A virus strains [28].
Table 3.
Pandemics of influenza virus infection
| Pandemics during nineteenth century | “Asiatic (Russian Flu)” 1889–1890 | 1 million death, strain H3N8 |
| Influenza pandemics during twentieth century | “Spanish” flu (1918–1919) (devastating pandemic) | Most deadliest pandemic caused by H1N1, >500,000 people died in the United State and up to 50 million people worldwide [70] |
| “Asian” flu (1957–1958) (moderate) | Caused by new H2N2 strain and killed two million people [23] | |
| “Hong Kong” flu (1968–1969) (mild) | Caused by H3N2 strain and killed one million people [25] | |
| Influenza pandemics during twenty-first century | “Swine flu” (2009) | Strain (H1N1), 6770 deaths reported till 16 Nov [7] |
Epidemiology of 2009 Pandemic H1N1
A novel influenza A (HINI) virus of swine origin (hereafter named as SO-IVA) emerged in Mexico and USA in early April 2009. It spread rapidly across 206 countries by 15 November 2009. Approximately 526,060 cases of pandemic H1N1/09 infection and at least 6770 deaths were reported [7]. The World Health Organization (WHO) declared its pandemic alert to the phase 6, highest level on June 11, 2009 indicating widespread community transmission on at least two continents [19]. In India as of 20 January 2010, the Ministry of Health and Family Welfare (MoHFW) reported over 28,000 confirmed cases and 1152 deaths giving a case fatality rate of 4%. The first SO-IAV death was reported on 3rd August in Pune. Studies show that Pune, Delhi and Mumbai have been the worst affected cities in India. The disease pattern in severe cases of this pandemic was quite different from the epidemic of seasonal influenza infection (H1N1) in that it greatly affected healthy young people and had higher transmissibility than seasonal influenza but lower clinical severity than the 1918 influenza pandemic [29, 30]. This novel SO-IVA emerged with completely new genome by reassortment of gene segments of avian, human and swine influenza viruses, when all three viruses simultaneously infected the same cell within intermediate host, i.e., Pig (mechanism called “antigenic shift”). Pig that acts as a reservoir for influenza virus continually harbors influenza viruses from different sources without any significant illness. Accumulation of gradual and small point mutations due to the polymerase activity of error prone RdRP in viral genome makes it able to cross the interspecies barrier and cause of epidemic (antigenic drift).
Genetic characterization studies revealed that the current pandemic H1N1 virus is antigenically similar to 1918 Spanish flu virus but it was quite different from seasonal H1N1 virus (that was causing epidemic outbreaks since 1970 and was maintained in human population). Many Laboratory and epidemiologic studies demonstrated that these antigenic similarities between the 1918 virus and the 2009 H1N1 resulted in long-term immunity against infection with 2009 H1N1 for many persons born before 1947 showed decreased risk of 2009 H1N1 infection among older adults compared to younger age groups [31, 32]. It contained completely new antigen by reassortment of gene segments from different sources within intermediate host pig. Pig act as a reservoir of avian as well as human H1N1 virus. It exhibits the combination of gene segments that previously has not been reported in swine and human influenza viruses in the United States or elsewhere. The NA and M segments of this virus have been considered from Eurasian swine genetic lineages of avian origin (1979), the HA, NS and NP gene segments are of classical swine lineages which also came from avian virus during 1918. The PB1, PB2 and PA genetic lineages were identified to triple reassortant swine viruses, of which PB1 came into pig indirectly from birds that was first seeded to human around 1968 and then from human to pig population in North America in 1998. The PA and PB2 also came into swine population of North America from birds around 1998 [33]. The outcome of all these studies represents that the new virus is made from pieces of flu viruses that infect pigs from North America, pigs from Eurasia, and pieces from human and bird flu viruses that were all mixed together and acquire the ability of pandemic potential to efficiently infect human population (Fig. 3).
Fig. 3.
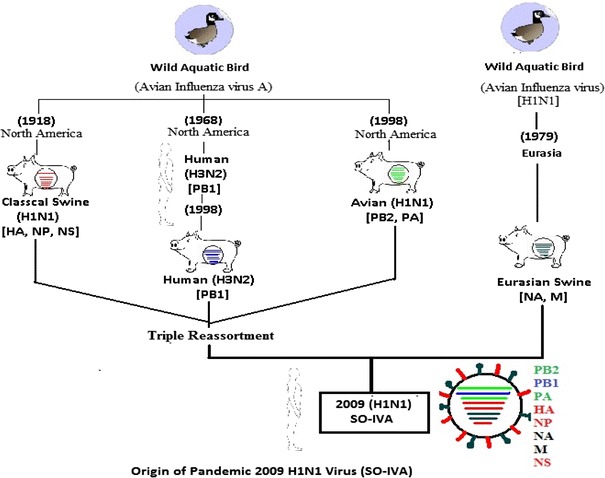
Origin of pandemic 2009 H1N1 virus (SO-IVA) Available sequence data indicate that currently circulating pandemic 2009 influenza virus A (SO-IVA) are related to two distinct lineages of swine influenza viruses, one from North American and another from European pig populations. The six segments of SO-IVA derive from swine viruses of North America and that the NA and M genes from Europe and Asia
Clinical Features
Influenza strains circulate throughout the year and cause seasonal epidemics with one or two peaks during rainy season in tropical countries. It can seriously affect all age groups, but the highest risk of complications occurs among children younger than age two, adults age 65 or older and people of any age with certain medical conditions such as chronic heart, lung, kidney, liver, blood or metabolic diseases (such as diabetes), or weakened immune systems. Generally, seasonal influenza shows mild to moderate respiratory infection with high attack rate and low mortality while pandemic has high mortality due to highly virulent strain [34, 35]. Clinical features concerned to seasonal influenza are the sudden onset of high fever, cough (usually dry), headache, muscle and joint pain, severe malaise (feeling unwell), sore throat and runny nose. It is self limiting, so most of the people recover from fever and other symptoms within a week without requiring medical attention. But influenza can cause severe illness or death in people at high risk. The time from infection to illness, known as the incubation period, is about two days (range, 1–5 days). Accurate diagnosis of influenza A or B infection based solely on clinical criteria is difficult because of the overlapping symptoms caused by the various viruses associated with upper respiratory tract infection (URTI) that present influenza like illness (ILI). Common symptoms of the flu such as fever, headache, and fatigue are the result of the huge amounts of proinflammatory cytokines and chemokines (such as interferon or tumor necrosis factor) produced from influenza-infected cells. In addition, several serious viruses, including adenoviruses, enteroviruses, and paramyxoviruses may initially cause influenza like symptoms. The early presentation of mild or moderate cases of flavivirus infections (e.g. Dengue) may initially mimic influenza. ILI includes a person with sudden onset of fever >38°C, cough or sore throat and/or running nose in the absence of other diagnoses. In severe cases of influenza, adults are accompanied by severe myalgias and dry cough with or without hemoptysis and children below 5 years of age present cough or difficult breathing or fast breathing with any of the following signs, i.e., chest in drawing or stridor in a calm child. The myalgias of seasonal influenza infection are peculiarly severe and are localized to the neck and back. Retro-orbital pain is common, and conjunctival suffusion may be present in severe seasonal human influenza A but conjunctival suffusion may be present in avian influenza (H5N1) but is uncommon with swine influenza (H1N1) [36, 37].
Pandemic influenza A affects primarily young healthy adults rather than the very young, elderly, and immunocompromised. Most fatal cases have occurred in young healthy adults. Clinical manifestation associated with pandemic H1N1 (2009) virus infection with mild symptoms in affected individuals shows symptoms like seasonal influenza infection but in severe cases it deeply affect lower respiratory tract leading to Pneumonia that begins abruptly [38]. A proportion of affected patients also presented gastrointestinal symptoms such as diarrhea and vomiting. Critically ill patients show acute respiratory distress syndrome (ARDS) with refractory hypoxaemia resulting in respiratory failure and in severe condition secondary bacterial infections, septic shock and multi-organ dysfunction. This is evidenced by one study on hospitalized patients with 2009 H1N1 influenza in the United States during April–June 2009. This study reported that hospitalized patients typically present with fever (98%) cough (88%), shortness of breath (60%), fatigue (43%), runny nose (38%), sore throat (31%), headache (34%), and myalgia (36%) [38]. These patients usually present symptoms within a week of exposure and are infectious for about 8 days after onset of symptoms [1]. Most of the human infections with pandemic 2009 H1N1 results in self limiting uncomplicated disease with clinical symptoms similar to seasonal influenza. Clinical signs atypical for seasonal influenza including vomiting and diarrhea have been reported, in a relatively large proportion of cases. Some patients have required hospitalization because of severe pneumonia and respiratory failure, with a fatal outcome. In contrast to seasonal influenza, a substantial proportion of the cases of severe illness and death have occurred among previously healthy adults aged 18–50 years, as well as among adults with underlying disease and pregnant women cases [39, 40].
Host Immune Response and Immunopathogenesis
The most consistent histopathological findings suggest that main pathological changes associated with fatal cases of H1N1 infection are localized to the lungs with varying degrees of diffused alveolar damage with hyaline membranes and septal edema tracheitis and necrotizing bronchiolitis [41, 42]. Cytopathic effect has also been found in the bronchial and alveolar epithelial cells, epithelial hyperplasia, and squamous metaplasia of the large airways with pulmonary vascular congestion and alveolar hemorrhage. It not only infects cells of upper respiratory tracheobrochial epithelial cells and mucosal glands but also infects alveolar epithelia cells (type-I and II pneumocytes). Influenza viral proteins were also detected in alveolar epithelial cells, macrophages as well as in epithelial cells of upper respiratory tract.
Although, the pattern of Innate and adaptive immune response in patients affected with pandemic 2009 H1N1 (SO-IVA) are not fully characterized but the data related to lung pathology of influenza infected patients represents that influenza virus infection induces a dysregulated and complex network of host immune responses. Host response involves early innate immune response as well as humoral and cell-mediated responses. Innate immune response show first line of defense against infection and mediated by the production of Type I Interferon (IFNγ) that are produced by infected respiratory epithelial cells and alveolar macrophages (AMs). Respiratory epithelial and AMs are primary targets for influenza virus H1N1 and H5N1. Infection causes necrosis and apoptotic death of epithelia and AMs that triggers innate immune response by producing Pro-inflammatory cytokines [Tumor necrosis factor (TNF-α), interleukin [IL] 2R, interleukin (IL-1) and chemokines [Monocyte chemoattractant protein (MCP-1), regulated on activation, normal T-cell expressed and secreted (RANTES), MIP-1α/β (macrophase inflammatory protein-1α/β), interferon induced protein (IP-1) and IL-8)] [43–46]. The release of extreme Pro-inflammatory cytokines and chemokines generate exaggerated inflammatory response resulting in recruitment of monocytes/macrophages, neutrophils, Dendritic cells (DCs) and lymphocytes into alveolar spaces, thereby further amplifying these pro-inflammatory cascades. There is evidence that this early inflammatory pathology contributes severe disease, especially with the highly lethal H5N1 viruses. Comparative studies with highly pathogenic pandemic (2009) H1N1 [SO-IFA], avian influenza virus and seasonal influenza virus H1N1 in primary human cells in vitro demonstrate that H5N1 induces proinflammatory cytokines more strongly as compared to pandemic (2009) H1N1 virus [SO-IVA] and seasonal H1N1 virus and that seasonal and pandemic (2009) H1N1 viruses induce similar proinflammatory mediated immune response[47]. In vivo studies revealed that patients who died or who had ARDS had higher plasma level of interleukin-6, interleukin-10 and interleukin-15 throughout the illness and of granuolocyte colony-stimulating factor, interleukin-1α, interleukin-8, and interferon—inducible protein 10, and tumor necrosis factor during the late phase of illness [48].
Few in vitro studies of H5N1 infection on primary human monocyte derived macrophages reports that H5N1 infection induce higher level of cytokines and chemokines such as TNF-α, IFN-β, IL-6, IL-1β, CXCL10/IFN-γ-inducible protein 10 (IP-10), CCL5/RANTES, CCL2/MCP-1, CCL3/MIP-1α, and CCL4/MIP-1β, when compared to seasonal H1N1 and S-OIV [49–51]. Similarly, H5N1 virus genotypes differentially up-regulate these cytokines from alveolar epithelial cells in vitro. There are two proteins p38 kinase and IRF3 (IFN regulatory factor 3) which play important roles in H5N1-mediated up-regulation of IFN-β, TNF-α and IFN-λ1 transcription by different pathways. TNF-α, IFN-β and IFN-λ1 are three cytokines act as primary mediators directly induced by H5N1 virus and others being the result of complex autocrine/paracrine cascade. In this differentially activated cytokine response induced by H5N1 virus it is not fully understood that which of these mediators are primarily induced by the virus and which ones result from autocrine or paracrine feedback. It has been reported that NS1 protein of human influenza viruses help virus to evade the antiviral defenses of the host by inhibiting IRF3 (IFN regulatory factor 3) activation which is a transcription factor of IFN-β [4, 52]. So, there may be the possibility that seasonal H1N1 and S-OIV produce little cytokine and chemokines responses. It has been recently reported that mediators from H5N1 virus-infected macrophages amplify and broaden the cytokine cascade through their interaction with alveolar epithelial cells. This study reveals that cyclooxygenase 2 (COX2) plays a regulatory role in this cytokine cascade and that COX2 inhibitors modulate this response [53]. In mice infected with H5N1 virus, others have shown that COX2 inhibitors enhanced the beneficial effect of antiviral drugs [54].
These findings reflect that viral genetic factors solely not responsible for varying degree of pathogenesis induced by influenza virus. But differential host immune responses especially innate immune responses have need to be explored.
Diagnosis
For diagnosis of influenza virus infection four tests are available that have been performed in the clinical laboratories. These include rapid antigen test (RIDT—rapid influenza diagnostic test), direct immunofluorescence assay (DFA) test, viral culture and molecular diagnosis by reverse transcriptase polymerase chain reaction (RT-PCR). RIDT lack sensitivity as compared to virus culture method and molecular methods and require technical expertise. It does not distinguish between various types of influenza such as seasonal H1N1, H3N2 and 2009 H1N1. DFA and virus culture have ability to distinguish between other respiratory viruses such as adenovirus, parainfluenza, respiratory syncytial virus and metapneumomvirus that commonly circulate with influenza virus during influenza season but unable to distinguish subtypes of influenza virus, time taking and require expertise in microscopy. The most sensitive and reliable method is performing molecular techniques such as RT-PCR that distinguishes all influenza viruses targeting on conserved matrix M1 gene, followed by RT-PCR assay specific for different influenza A haemagglutinin genes, which would discriminate between pandemic H1, seasonal H1 and seasonal H3 viruses. Thus, Nucleic acid-based approaches are superior to conventional nonmolecular tests (such as RIDT, DFA and culture methods) because of their rapidity, excellent sensitivity, specificity and ability to distinguish between seasonal and pandemic influenza A, often in a single step. The negative results from RIDT and DFA test do not confirm the absence of pandemic H1N1 (2009) so confirmation of cases is only made by real-time RT-PCR and viral culture according to CDC guidelines [55, 56].
Treatment
Two classes of antiviral compounds are widely accepted for the treatment of influenza viruses that is licensed by the US FDA. Adamantanes grouped drugs (amantadine and rimantadine) block the activity of the viral M2 proton channel, thus preventing virus uncoating and inhibiting release of the viral genome into host cells [57]. The second group of drugs is NA inhibitors (Oseltamivir/Tamiflu and zanamivir/Relenza) that bind and block enzymatic activity of the viral NA protein [57]. This prevents efficient release of mature virions from the surface of infected cells and thereby limits viral spread. A current study on treatment implies that drug Oseltamivir resistant strains of seasonal H1N1 and H5N1 have been emerged, although most H1N1 viruses remain sensitive to zanamivir [58, 59]. The researches have been made to develop novel antivirals due to continuous emergence of Oseltamivir resistant strains among highly pathogenic H5N1 viruses which may cause another devastating pandemic in future [60]. Although early isolates of the novel swine-origin pandemic H1N1 influenza A virus were demonstrated to be susceptible to the commonly used antivirals, some Oseltamivir-resistant mutants of this virus have recently been reported [61, 62]. The global influenza surveillance network (GISN), supported by WHO Collaborating Centres had identified 35 Oseltamivir-resistant pandemic H1N1 influenza viruses worldwide by October 16, 2009, all exhibited H275Y mutation which causes resistance to Oseltamivir, but not to Zanamivir [63]. Thus, it is evident that new antivirals are continuously needed. Although a number of studies have proposed multiple excellent strategies for the future development of novel anti-influenza drugs [64, 65] an influenza epidemic emerges during each winter’s flu season.
Vaccine
Each year there are two flu seasons due to the occurrence of influenza at different times in the Northern and Southern Hemispheres. Hence, WHO recommended two vaccines per year for seasonal epidemics of influenza, one for Northern and another for Southern Hemisphere. The most common prevailing influenza A subtypes that infect humans are H1N1 and H3N2. Each year, the trivalent vaccine used worldwide contains influenza A strains from H1N1 and H3N2, along with an influenza B strain. Flu vaccines are available either as TIV [injection of trivalent (three strains; usually A/H1N1, A/H3N2, and B) inactivated vaccine] or LAIV (nasal spray of live attenuated influenza vaccine). Every year these vaccines are reformulated with identifying circulating strains of influenza virus A and B in Northern and Southern geographical region that have been confirmed by WHO. The composition of virus vaccines for use in the 2009–2010 Northern Hemisphere influenza season recommended by the World Health Organization on February 12, 2009 [66] was an A/Brisbane/59/2007 (H1N1)-like virus; an A/Brisbane/10/2007 (H3N2)-like virus; a B/Brisbane/60/2008-like virus [67]. Since the A/Brisbane/59/2007 (H1N1)-like virus used in the vaccine is an unrelated seasonal strain of influenza, it probably cannot create immunity to the new, non-seasonal strain of influenza A virus subtype H1N1 responsible for the 2009 swine flu outbreak [68]. Annual vaccine formulated during 2010 Southern Hemisphere winter season consist of an A/California/7/2009 (H1N1)-like virus; an A/Perth/16/2009 (H3N2)-like virus; a B/Brisbane/60/2008-like virus [69]. The H1N1 strain, i.e., A/California/7/2009 (H1N1)-like virus used in this composition is the same strain used in the 2009 flu pandemic vaccine [69].
Conclusion
Influenza virus infection is a zoonotic disease that spreads rapidly and greatly affects human population across the large geographical regions within a short period of time. The segmented nature of viral genome augments the diversity of virus through genetic reassortment, in addition to normal mutation. High mutational rate in surface glycoproteins such as HA and NA enhances capacity of virus to adapt efficiently in human population and makes it responsible for varying degree of pathology from mild to severe. Although, the epidemiology related to influenza infection has been studied from several years but some facts associated to disease transmission has been poorly understood.
To overcome the disease researches have been done in field of vaccine formulation to develop a universal vaccine that will be effective against any type of influenza strain. Chemotherapy against influenza infection includes few drugs such as Oseltamivir and Zanamivir available in market. Excessive uses of these drugs in treatment of seasonal and pandemic influenza in post-prophylaxis cases are also challenged by emergence of drug-resistant mutants. A lot of research has been done all over the world but knowledge is still required for better understanding the molecular mechanism behind several unrevealed metabolic pathways triggered by virus during pathogenesis which will further help in designing new drugs and vaccines.
References
- 1.Dawood F, Jain S, Finelli L, et al. Emergence of a novel swine-origin influenza A (H1N1) virus in humans. N Engl J Med. 2009;360:2605–2615. doi: 10.1056/NEJMoa0903810. [DOI] [PubMed] [Google Scholar]
- 2.Reed C, Angulo FJ, Swerdlow DL, et al. Estimates of the prevalence of pandemic (H1N1) 2009, United States, April–July 2009. Emerg Infect Dis. 2009;15:7–2004. doi: 10.3201/eid1512.091413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Donaldson LJ, Rutter PD, Ellis BM, et al. Mortality from pandemic A/H1N12009 influenza in England: public health surveillance study. Br Med J. 2009;339:b5213. doi: 10.1136/bmj.b5213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Li KS, Guan Y, Wang J, et al. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature. 2004;430:209–213. doi: 10.1038/nature02746. [DOI] [PubMed] [Google Scholar]
- 5.Cox NJ, Subbarao K. Global epidemiology of influenza: past and present. Annu Rev Med. 2000;51:407–421. doi: 10.1146/annurev.med.51.1.407. [DOI] [PubMed] [Google Scholar]
- 6.Treanor JJ. Influenza virus. In: Mandell GL, Bennett JE, Dolin R, editors. Principles and practice of infectious diseases. Philadelphia: Elsevier; 2005. pp. 2060–2085. [Google Scholar]
- 7.World Health Organization (2009) Pandemic influenza (H1N1) 2009 Update 75. http://www.who.int/csr/don/2009. Accessed 8 July 2011
- 8.Fields BN, Knipe DM, Howley PM. Fields virology. Philadelphia: Wolters Kluwer Health/Lippincott Williams & Wilkins; 2007. [Google Scholar]
- 9.Chen W, et al. A novel influenza A virus mitochondrial protein that induces cell death. Nat Med. 2007;7:1306–1312. doi: 10.1038/nm1201-1306. [DOI] [PubMed] [Google Scholar]
- 10.Matrosovich MN, Matrosovich TY, Gray T, et al. Human and avian influenza viruses target different cell types in cultures of human airway epithelium. Proc Natl Acad Sci USA. 2004;101:4620–4624. doi: 10.1073/pnas.0308001101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Nayak DP, Hui EK, Barman S. Assembly and budding of influenza virus. Virus Res. 2004;106:147–165. doi: 10.1016/j.virusres.2004.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wilson IA, Skehel JJ, Wiley DC. Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3 Å resolutions. Nature. 1981;289:366–373. doi: 10.1038/289366a0. [DOI] [PubMed] [Google Scholar]
- 13.Skehel JJ, Wiley DC. Receptor binding and membrane fusion in virus entry: the influenza hemagglutinin. Annu Rev Biochem. 2000;69:531–569. doi: 10.1146/annurev.biochem.69.1.531. [DOI] [PubMed] [Google Scholar]
- 14.Harrison SC. Viral membrane fusion. Nat Struct Mol Biol. 2008;15:690–698. doi: 10.1038/nsmb.1456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pinto LH, Lamb RA. The M2 proton channels of influenza A and B viruses. J Biol Chem. 2006;281:8997–9000. doi: 10.1074/jbc.R500020200. [DOI] [PubMed] [Google Scholar]
- 16.Wang C, Lamb RA, Pinto LH. Activation of the M2 ion channel of influenza virus: a role for the transmembrane domain histidine residue. Biophys J. 1995;69:1363–1371. doi: 10.1016/S0006-3495(95)80003-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wu WWW, Pante N. The directionality of the nuclear transport of the influenza A genome is driven by selective exposure of nuclear localization sequences on nucleoprotein. Virol J. 2009;6:68. doi: 10.1186/1743-422X-6-68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Portela A, Digard P. The influenza virus nucleoprotein: a multifunctional RNA binding protein pivotal to virus replication. J Gen Virol. 2002;83:723–734. doi: 10.1099/0022-1317-83-4-723. [DOI] [PubMed] [Google Scholar]
- 19.Sanz-Ezquerro JJ, Zurcher T, de la Luna S, Ortin J, Nieto A. The amino-terminal one-third of the influenza virus PA protein is responsible for the induction of proteolysis. J Virol. 1996;70:1905–1911. doi: 10.1128/jvi.70.3.1905-1911.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ulmanen I, Broni BA, Krug RM. The role of two of the influenza virus core P proteins in recognizing cap 1 structures (m 7 GpppNm) on RNAs and in initiating viral RNA transcription. Proc Natl Acad Sci USA. 1981;78:7355–7359. doi: 10.1073/pnas.78.12.7355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Plotch SJ, Bouloy M, Ulmanen I, Krug RM. A unique cap (m 7 GpppXm) dependent influenza virion endonuclease cleaves capped RNAs to generate the primers that initiate viral RNA transcription. Cell. 1981;23:847–858. doi: 10.1016/0092-8674(81)90449-9. [DOI] [PubMed] [Google Scholar]
- 22.Hagen M, Chung TDDY, Butcher A, Krystal M. Recombinant influenza virus polymerase: requirement of both 5 and 3 viral ends for endonuclease activity. J Virol. 1991;68:1509–1515. doi: 10.1128/jvi.68.3.1509-1515.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.WHO . Avian influenza: assessing the pandemic threat. Geneva: WHO; 2005. [Google Scholar]
- 24.Lazzari S, Stohr K. Avian influenza and influenza pandemics. Bull World Health Organ. 2004;82:242. [PMC free article] [PubMed] [Google Scholar]
- 25.Simonsen L, Clarke MJ, Schonberger LB, Arden NH, Cox NJ, Fukuda K. Pandemic versus epidemic influenza mortality: a pattern of changing age distribution. J Infect Dis. 1998;178:53–60. doi: 10.1086/515616. [DOI] [PubMed] [Google Scholar]
- 26.Glezen WP. Emerging infections: pandemic influenza. Epidemiol Rev. 1996;18:64–76. doi: 10.1093/oxfordjournals.epirev.a017917. [DOI] [PubMed] [Google Scholar]
- 27.Morens DM, Taubenberger JK, Fauci AS. The persistent legacy of the 1918 influenza virus. N Engl J Med. 2009;361:225–229. doi: 10.1056/NEJMp0904819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brockwell-Staats C, Webster RG, Webby RJ. Diversity of influenza viruses in swine and the emergence of a novel human pandemic influenza A (H1N1) Influenza Other Respi Viruses. 2009;3:207–213. doi: 10.1111/j.1750-2659.2009.00096.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.World Health Organization (WHO) (2009) World now at the start of 2009 influenza pandemic. http://www.who.int/mediacentre/news/statements/2009/h1n1. Accessed 8 July 2011
- 30.Fraser C, Donnelly CA, Cauchemez S, et al. Pandemic potential of a strain of influenza A (H1N1): early findings. Science. 2009;324:1557. doi: 10.1126/science.1176062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hancock K, Veguilla V, Lu X, et al. Cross-reactive antibody responses to the 2009 pandemic H1N1 influenza virus. N Engl J Med. 2009;361:1–8. doi: 10.1056/NEJMoa0906453. [DOI] [PubMed] [Google Scholar]
- 32.Gras S, Kedzierski L, Valkenburg S, et al. Cross-reactive CD81 T-cell immunity between the pandemic H1N1–2009 and H1N1–1918 influenza A viruses. Proc Natl Acad Sci USA. 2010;107:12599–12604. doi: 10.1073/pnas.1007270107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Garten RJ, Davis CT, Russell CA, Shu B, Lindstrom S, Balish A, et al. Antigenic and genetic characteristics of swine-origin 2009 A (H1N1) influenza viruses circulating in humans. Science. 2009;325:197–201. doi: 10.1126/science.1176225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sym D, Patel PM, Chaar GM. Seasonal, avian and novel H1N1 influenza: prevention and treatment modalities. Ann Pharmacother. 2009;43:2001–2011. doi: 10.1345/aph.1M557. [DOI] [PubMed] [Google Scholar]
- 35.Debre R, Couvreur J. Influenza: clinical features. In: Debre R, Celers J, editors. Clinical virology: the evaluation and management of human viral infections. Philadelphia: WB Saunders; 1970. pp. 507–515. [Google Scholar]
- 36.Harper SA, Bradley JS, Englund JA, et al. Seasonal influenza in adults and children: diagnosis, treatment, chemoprophylaxis and institutional outbreak management: clinical practice guidelines of the Infectious Diseases Society of America. Clin Infect Dis. 2009;48:1003–1032. doi: 10.1086/598513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cunha BA. Pneumonia essentials. 3. Sudbury: Jones & Bartlett; 2010. [Google Scholar]
- 38.Jain S, Kamimoto L, Bramley AM, et al. Hospitalized patients with 2009 H1N1 influenza in the United States, April–June 2009. N Engl J Med. 2009;361:1935–1944. doi: 10.1056/NEJMoa0906695. [DOI] [PubMed] [Google Scholar]
- 39.Munster VJ, de Wit E, van den Brand JM, et al. Pathogenesis and transmission of swine origin 2009 A(H1N1) influenza virus in ferrets. Science. 2009;325:481–483. doi: 10.1126/science.1177127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Shieh WJ, Blau DM, Denison AM, et al. Pandemic influenza A (H1N1): pathology and pathogenesis of 100 fatal cases in the United States. Am J Pathol. 2010;177:166–175. doi: 10.2353/ajpath.2010.100115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Gill JR, Sheng Z, Ely SF, et al. Pulmonary pathological findings of fatal 2009 pandemic influenza A/H1N1 viral infections. Arch Pathol Lab Med. 2010;134:225–243. doi: 10.5858/134.2.235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bender BS, Small PA., Jr Influenza: pathogenesis and host defense. Semin Respir Infect. 1992;7:38–45. [PubMed] [Google Scholar]
- 43.Fesq H, Bacher M, Nain M, Gemsa D. Programmed cell death (apoptosis) in human monocytes infected by influenza A virus. Immunobiology. 1994;190:175–182. doi: 10.1016/S0171-2985(11)80292-5. [DOI] [PubMed] [Google Scholar]
- 44.Julkunen I, Melen K, Nyqvist M, Pirhonen J, Sareneva T, Matikainen S. Inflammatory responses in influenza A virus infection. Vaccine. 2000;19(Suppl 1):S32–S37. doi: 10.1016/S0264-410X(00)00275-9. [DOI] [PubMed] [Google Scholar]
- 45.Julkunen I, Sareneva T, Pirhonen J, Ronni T, Melen K, Matikainen S. Molecular pathogenesis of influenza A virus infection and virus-induced regulation of cytokine gene expression. Cytokine Growth Factor Rev. 2001;12:171–180. doi: 10.1016/S1359-6101(00)00026-5. [DOI] [PubMed] [Google Scholar]
- 46.Woo PC, Tung ET, Chan KH, Lau CC, Lau SK, Yuen KY, et al. Cytokine profiles induced by the novel swine-origin influenza A/H1N1 virus: implications for treatment strategies. J Infect Dis. 2010;201:346–353. doi: 10.1086/649785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hung KK, Li IF, et al. Delayed clearance of viral load and marked cytokine activation in severe cases of pandemic H1N1 2009 influenza virus infection. Clin Infect Dis. 2010;50:850–859. doi: 10.1086/650581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cheung CY, Poon LL, Lau AS, Luk W, Lau YL, Shortridge KF, Gordon S, Guan Y, Peiris JS. Induction of proinflammatory cytokines in human macrophages by influenza A (H5N1) viruses: a mechanism for the unusual severity of human disease? Lancet. 2002;360:1831–1837. doi: 10.1016/S0140-6736(02)11772-7. [DOI] [PubMed] [Google Scholar]
- 49.Perrone LA, Plowden JK, Garcia-Sastre A, Katz JM, Tumpey TM. H5N1 and 1918 pandemic influenza virus infection results in early and excessive infiltration of macrophages and neutrophils in the lungs of mice. PLoS Pathog. 2008;4:E1000115. doi: 10.1371/journal.ppat.1000115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Guan Y, Poon LL, Cheung CY, Ellis TM, Lim W, Lipatov AS, Chan KH, Sturm-Ramirez KM, Cheung CL, Leung YH, et al. H5N1 influenza: a protean pandemic threat. Proc Natl Acad Sci USA. 2004;101:8156–8161. doi: 10.1073/pnas.0402443101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kochs G, Garcia-Sastre A, Martinez-Sobrido L. Multiple anti-interferon actions of the influenza A virus NS1 protein. J Virol. 2007;81:7011–7021. doi: 10.1128/JVI.02581-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lee SM, Cheung CY, Nicholls JM, Hui KP, Leung CY, Uiprasertkul M, Tipoe GL, Lau YL, Poon LL, Ip NY, et al. Hyperinduction of cyclooxygenase-2-mediated proinflammatory cascade: a mechanism for the pathogenesis of avian influenza H5N1 infection. J Infect Dis. 2008;198:525–535. doi: 10.1086/590499. [DOI] [PubMed] [Google Scholar]
- 53.Hui KPY, Lee SMY, Cheung C, Ng IHY, Poon LLM, et al. Induction of proinflammatory cytokines in primary human macrophages by influenza A virus (H5N1) Is selectively regulated by IFN regulatory factor 3 and p38 MAPK. J Immunol. 2009;182:1088–1098. doi: 10.4049/jimmunol.182.2.1088. [DOI] [PubMed] [Google Scholar]
- 54.Centers for Disease Control and Prevention (CDC) Interim guidance for the detection of novel influenza A virus using rapid influenza diagnostic tests. Atlanta: CDC; 2009. [Google Scholar]
- 55.Centers for Disease Control and Prevention (CDC) (2009) Interim recommendations for clinical use of influenza diagnostic tests during 2009–10 influenza season. www.cdc.gov/h1n1flu/guidance/diagnostic_tests.htm. Accessed 8 July 2011
- 56.Palese P, Shaw ML. Orthomyxoviridae: the viruses and their replication. In: Knipe DM, Howley PM, editors. Fields virology. Philadelphia: Lippincott Williams & Wilkins; 2007. pp. 1647–1689. [Google Scholar]
- 57.CDC Update: influenza activity—United States, September 28, 2008–April 4, 2009, and composition of the 2009–2010 influenza vaccine. Morb Mortal Wkly Rep. 2009;58:369–374. [PubMed] [Google Scholar]
- 58.Le MT, Wertheim HF, Nguyen HD, et al. Influenza A H5N1 clade 2.3.4 virus with a different antiviral susceptibility profile replaced clade 1 virus in humans in northern Vietnam. PLoS One. 2008;3:e3339. doi: 10.1371/journal.pone.0003339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Le QM, Kiso M, Someya K, et al. Avian flu: isolation of drug-resistant H5N1 virus. Nature. 2005;437:1108. doi: 10.1038/4371108a. [DOI] [PubMed] [Google Scholar]
- 60.Itoh Y, Shinya K, Kiso M, et al. In vitro and in vivo characterization of new swine-origin H1N1 influenza viruses. Nature. 2009;460:1021–1025. doi: 10.1038/nature08260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.CDC Update: drug susceptibility of swine-origin influenza A (H1N1) viruses, April 2009. Morb Mortal Wkly Rep. 2009;58:433–435. [PubMed] [Google Scholar]
- 62.World Health Organization (WHO) (2009) Pandemic (H1N1) 2009—update 70 virological surveillance data. http://www.who.int/csr/disease/swineflu/laboratory16_10_2009/en/index.html. Accessed 8 July 2011
- 63.Hoffmann HH, Palese P, Shaw ML. Modulation of influenza virus replication by alteration of sodium ion transport and protein kinase C activity. Antivir Res. 2008;80:124–134. doi: 10.1016/j.antiviral.2008.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ludwig S. Targeting cell signalling pathways to fight the flu: towards a paradigm change in anti-influenza therapy. J Antimicrob Chemother. 2009;64:1–4. doi: 10.1093/jac/dkp161. [DOI] [PubMed] [Google Scholar]
- 65.WHO information meeting on influenza vaccine composition for the Northern Hemisphere 2009–2010 and round-table discussion on matters relating to influenza vaccine development and production. WHO headquarters, Executive Board Room, Geneva
- 66.Barr IG, McCauley J, Cox N, Daniels R, et al. Epidemiological, antigenic and genetic characteristics of seasonal influenza A(H1N1), A(H3N2) and B influenza viruses: basis for the WHO recommendation on the composition of influenza vaccines for use in the 2009–2010 Northern Hemisphere season. Vaccine. 2010;28:1156–1167. doi: 10.1016/j.vaccine.2009.11.043. [DOI] [PubMed] [Google Scholar]
- 67.CDC article (2009) 2008–2009 influenza season week 21 ending May 30, 2009. http://www.cdc.gov/flu/weekly/weeklyarchives2008-2009/weekly21.htm. Accessed 5 June 2009
- 68.WHO. Recommended composition of influenza virus vaccines for use in the 2010 southern hemisphere influenza season
- 69.Centres for Disease Control and Prevention (2009) Update: Influenza A (H1N1) 2009 monovalent vaccines. Morb Mortal Wkly Rep 58:1100–1101 [PubMed]
- 70.Johnson NP, Mueller J. Updating the accounts: global mortality of the 1918–1920 “Spanish” influenza pandemic. Bull Hist Med. 2002;76:105–115. doi: 10.1353/bhm.2002.0022. [DOI] [PubMed] [Google Scholar]