

Abstract
Background
Influenza is an acute viral respiratory illness that causes high morbidity and mortality globally. Therapeutic actions are limited to vaccines and a few anti-viral drugs. Polygala (P.) japonica herba is rich in Polygalasaponin F (PSF, C53H86O23), used for acute bronchitis, pharyngitis, pneumonia, amygdalitis, and respiratory tract infections treatment in China. Hypercytokinemia is often correlated with severe pneumonia caused by several influenza viruses. PSF was reported to have anti-inflammatory effects and its mechanism is associated with the nuclear factor (NF)-κB signaling pathway. The action of PSF to alleviate pulmonary inflammation caused by influenza A virus (IAV) infection requires careful assessment. In the present study, we evaluated the effect and mechanism of PSF on mice with pneumonia caused by influenza H1N1 (A/FM/1/47).
Methods
Mice were infected intranasally with fifteen 50% mouse lethal challenge doses (MLD50) of influenza virus. BALB/c mice were treated with PSF or oseltamivir (oral administration) for 2 h post-infection and received concomitant treatment for 5 days after infection. On day 6 post-infection, 10 mice per group were killed to collect related samples, measure body weight and lung wet weight, and detect the viral load, cytokine, prostaglandins, pathological changes, and cell pathway protein expression in the lungs. In addition, the survival experiments were carried out to investigate the survival of mice. The expression profile of cell pathway proteins was detected and analyzed using a broad pathway antibody array and confirmed the findings from the array by western blotting.
Results
Polygalasaponin F and oseltamivir can protect against influenza viral infection in mice. PSF and oseltamivir significantly relieved the signs and symptoms, reduced body weight loss, and improved the survival rate of H1N1-infected mice. Moreover, PSF efficiently decreased the level of interleukin (IL)-1β, tumor necrosis factor (TNF)-α, IL-4, interferon (IFN)-γ, thromboxane A2 (TXA2), and prostaglandin E2 (PGE2) in lung tissues of mice infected with influenza virus (p < 0.05–0.01). Oseltamivir had a similar effect to lung cytokine of PSF, but did not decrease the levels of TXA2 and PGE2. There was a twofold or greater increase in four cell pathway protein, namely NF-κB p65 (2.68-fold), I-kappa-B-alpha (IκBα) (2.56-fold), and MAPK/ERK kinase 1 (MEK1) (7.15-fold) assessed in the array induced by influenza virus. Western blotting showed that the expression of these proteins was significantly decreased in lung after influenza virus challenge in PSF and oseltamivir-treated mice (p < 0.05–0.01).
Conclusion
Polygalasaponin F appears to be able to augment protection against IAV infection in mice via attenuation of pulmonary inflammatory responses. Its effect on IAV-induced pulmonary inflammation was associated with suppression of Raf/MEK/ERK and NF-κB expressions.
Keywords: Polygalasaponin F, Pneumonia, Influenza virus, Signaling pathway
Introduction
Influenza A virus (IAV) causes respiratory tract infection in humans and animals with strong infectivity, rapid epidemic, and variation (Cheung and Poon 2007). Anti-influenza drugs currently used in the clinic include influenza virus matrix protein M2 ion channel inhibitors (amantadine), neuraminidase (NA) inhibitors (oseltamivir) (Pawestri et al. 2018; Pires de Mello et al. 2018), and RNA polymerase inhibitors (ribavirin) (De Clercq 2006). However, the toxicity and side effects of long-term and large-scale use of anti-influenza drugs and their resistance to viruses have attracted much attention (Choi et al. 2009). NA inhibitor, oseltamivir, has been approved by the US Food and Drug Administration for the treatment of influenza and oseltamivir-resistant IAV emerge frequently (Kiso et al. 2010; Liao et al. 2017).
When the respiratory tract is infected with IAV, monocytes and macrophages gather around the lungs, producing and releasing a large number of proinflammatory cytokines and chemokines, which would cause inflammatory damages to the lung tissue. This process is called “cytokine storm” (Perrone et al. 2008), which is considered as a predictor of morbidity and mortalities during influenza infection (Ding et al. 2017). Viral pneumonia accounts for 40–66% of hospitalized patients with influenza, while 22.9–42% of pneumonia patients can develop acute respiratory distress syndrome (ARDS) (Denholm et al. 2010), which is the main cause of mortality in patients with severe influenza (Chien et al. 2010).
Polygala (P.) japonica herba, a traditional Chinese herb, has been used as an expectorant, antitussive, antibacterial, and ataractic agent (Zhang et al. 1996). It is widely used in China to treat acute bronchitis, pharyngitis, pneumonia, amygdalitis, and respiratory tract infections in children (Zhou et al. 2015). The compounds isolated from P. japonica so far mainly include saponins, flavonoids, xanthones, polysaccharides, etc. Saponins are one of the key components of P. japonica (Fu et al., 2008). These activities may be due to the presence of various saponins in P. japonica, since studies have showed that the saponins found in P. japonica have antipsychotic, anti-inflammatory, and expectorant effects (Chung et al. 2002; Wang et al. 2008). Polygalasaponin F (PSF, C53H86O23, Fig. 1) is a triterpenoid saponin and is also one of the major active constituents of P. japonica used for the quality control of the medicine (Zhou et al. 2015). Previous research has reported that PSF has anti-inflammatory effects and its mechanism is associated with the NF-κB signaling pathway (Wei et al. 2014; Yan et al. 2015; Wang et al. 2008). Inflammation is associated with influenza. Importantly, hypercytokinemia is often correlated with severe pneumonia caused by several influenza viruses. When infected by influenza virus, the cytokines will be duplicated with the support of various cell-signaling pathways such as the changes in Raf/MEK/ERK, nuclear factor (NF)-κB, and phosphatidylinositol 3-kinase (PI3K) induced by IAV (Ehrhardt et al. 2006). Therefore, a reduction in cytokine expression and viral replications by interfering the signaling events is considered as a beneficial method to treat IAV infections. PSF has an important role in anti-inflammatory activity and inhibition of (NF)-κB (Yan et al. 2015; Wei et al. 2014). PSF may also have the potential to treat pneumonia induced by IAV. This study was designed to investigate whether PSF could provide protection against pneumonia caused by influenza virus. The mechanisms of PSF anti-influenza virus using high-throughput proteome array and western blotting were also discussed.
Fig. 1.
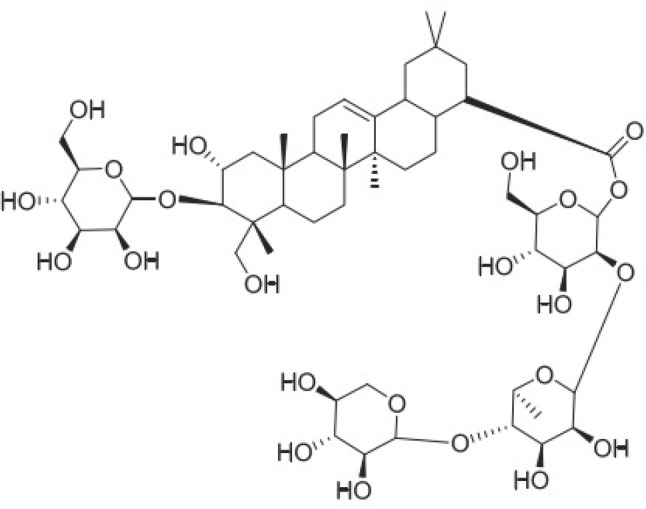
Structure of polygalasaponin F
Methods
Animals and Viruses and cells
Female BALB/C mice, provided by Medical Experiment Center of Guangdong province (Guangzhou, China), aged 6–8 weeks, specific-pathogen-free (SPF) were used. The study has been approved by the Animal Care and Use Committee of Guangzhou University of Chinese Medicine (Guangzhou, China, 2018, No.151). Humane endpoints were used through euthanasia and anaesthesia for the experimental mice (e.g. survival study).
A strain of mouse-adapted influenza virus, A/Font Monmouth/47(H1N1, FM1), was obtained from Chinese Center for Disease Control and Prevention (CDC). The virus was then plaque purified in Madin–Darby canine kidney (MDCK) cells and replicated in 9-day-old chicken embryos. Before the study, the virus pool was pretitrated in mice, so that a suitable challenge dose can be determined first.
Compounds
Polygalasaponin F was obtained from Shanghai Yuanye Bio-Technology Co., Ltd. (purity ≥ 98%; Lot: Z13D5B1; Shanghai, China) and diluted to the desired concentrations with sterile saline (suspended in 0.5% Tween 80). Oseltamivir was acquired from Hoffmann-La Roche, Ltd. (Basel, Switzerland; Lot: M1301).
Murine model of influenza virus infection
BALB/c mice aged 8 weeks were divided into six groups, with ten mice in each, including negative control (NC), IAV control (IAV-C), PSF groups (PSF 50 mg/kg, 100 mg/kg, and 200 mg/kg), and the positive control oseltamivir (10 mg/kg). The dose of infection of influenza virus was used as described previously (Li et al. 2017). The dose of PSF and oseltamivir were referred to pharmacological experiments in mice as described previously (Chung et al. 2002; Langlois et al. 2010). Intranasal infection of mice (except NC group) were carried out with 50 μL fifteen 50% mouse lethal challenge doses (MLD50) of influenza virus in sterile PBS (pre-cooled) with mild anaesthesia. Mice were orally administered with PSF or oseltamivir and received concomitant treatment for 5 days post-infection. On the 6th day after infection, 10 mice in each group were executed to measure body weight and lung wet weight, and to detect viral load, pathological changes, prostaglandins, cytokines, and expression of cellular pathway proteins in the lungs.
Survival experiments
BALB/c mice were intranasally infected by five MLD50 influenza viruses in pre-cooled sterile PBS. Mice were treated for 2 h after infection with PSF or oseltamivir (oral gavage). After infection, the mice were treated for 5 days and observed for 2 weeks every day. The weight loss, survival status, clinical symptoms, and signs of illness of mice were observed every day and the survival rate was evaluated.
Lung index
Lung index is used to reflect the severity of inflammation and pulmonary edema. After measuring the body weight, the lungs of mice were removed, washed, and measured. Lung index was calculated as follows: lung weight/body weight × 100%.
Lung pathology
The lungs were treated with 10% PBS buffer, formaldehyde, dehydrated with graded ethanol, and imbedded in low-melting paraffin. The lungs were then cut into 5-µm pieces and then the pieces were sectioned for further examination. The sections were stained with HE and observed under microscope by double-blinded method.
The histopathology scores were examined to determine the extent of pneumonia in a blinded fashion as previously described (Chen et al. 2015): 0 represented no pneumonia; 1 mild interstitial pneumonia (< 25% of the lung); 2 moderate interstitial pneumonia (25–50% of the lung); 3 severe interstitial pneumonia (50–75% of the lung), and 4 very severe interstitial pneumonia (> 75% of the lung).
Viral load analysis by quantitative polymerase chain reaction (qPCR)
The viral load of lungs are determined as previously described (Li et al. 2017). Briefly, the lung was homogenized and centrifuged at 4000g for 15 min at 4 °C. Total RNA was extracted by RNeasy kit (TAKARA Biotechnology Co., Ltd, Dalian, China). Revert Aid First Strand cDNA Synthesis Kit (MBI Fermentas, Vilnius, Lithuania) was used to reverse transcribe it into cDNA as per the instruction of the manufacturer. The primer sequences of IAV M gene were: forward 5′-AATGGTGCAGGCGATGAGAG-3′ and reverse 5′-TACTTGCGGCAACAACGAGAG-3′. GAPDH primers used for internal controls of cellular RNAs were: forward 5′-CCTCGTCCCGTAGACAAAATG-3′ and reverse 5′-TGAGGTCAATGAAGGGGTCG-3′. Quantitative reverse transcriptase PCR (qRT-PCR) was performed with Platinum SYBR Green qPCR SuperMix-UDG kit (Invitrogen) in an ABI Applied Biosystems ViiA 7™ Real-Time PCR System (Applied Biosystems, Foster city, CA, USA) based on the following parameters: 95 °C for 2 min, 40 cycles of 95 °C for 15 s, 62.5 °C for 30 s, and 72 °C for 30 s. The analysis of each sample is carried out in triplicate. IAV quantities of PCR products were expressed as per mode for normalized expression (2−ΔΔct).
Cytokine in lung homogenate in mice infected with IAV
The lungs were homogenized and centrifuged at 4000g for 15 min at 4 °C. Supernatants of the lung homogenates are collected and stored at − 80 °C to analyze the cytokine levels. Then the interleukin (IL)-4, tumor necrosis factor (TNF)-α, IL-1β, and interferon (IFN)-γ concentrations from the homogenates were measured using Mouse IL-4, TNF-α, IL-1β, and IFN-γ ELISA Kit (Shanghai Enzyme-linked Biotechnology Co., Ltd., Shanghai, China) based on the instructions of the manufacturer.
The levels of thromboxane A2 (TXA2) and prostaglandin E2 (PGE2) in lung homogenate in mice infected with IAV
The lungs were homogenized and centrifuged at 4000g for 15 min at 4 °C. Supernatants of the lung homogenates were collected and stored at − 80 °C to analyze the cytokine levels. Then the levels of TXA2 and PGE2 in lung using Mouse TXA2 and PGE2 ELISA Kit (Shanghai Enzyme-linked Biotechnology Co., Ltd., Shanghai, China) were assayed according to the instructions of the manufacturer.
CSP100 broad pathway antibody array
The expression profiles of cell pathway proteins were detected and analyzed by broad pathway antibody array (CSP100, Fullmoon Biosystems, Sunnyvale, CA, USA). Mice were divided into two groups: IAV control group (IAV + sterile saline, suspended in 0.5% Tween 80) and PSF group (IAV + PSF 100 mg/kg). Mice were fed with PSF for 2 days before viral administration and treated for 5 days after infection. On day 6 after infection, mice were killed from each group to collect the lungs for the study. Lungs were lysed with extraction buffer as instructed by the manufacturer. The biotin reagent dissolved in N, N-dimethylformamide (DMF) is used to biotinylate the protein samples. 10 µL protein sample was blend with 40 µL labeling buffer and 1:7 biotin/DMF reagent was added to the mixture. The protein samples labeled with biotin were conjugated to pathway antibody array. The slides of antibody array were incubated on a rotating shaker at ambient temperature using blocking solution, then rinsed three times with water and dried naturally. Next, the protein coupling mix was added to the array slide and incubated at 4 °C overnight. The slides were washed twice, each time for 10 min using 1 × washing solution. The Cy3-streptavidin solution was added and shaked (55 rpm) for 20 min at ambient temperature. The slides were scanned using GenePix 4000B Array Scanner. The fold changes in intensities of protein between IAV control group and PSF group were calculated.
Immunoblotting
The lungs were lysed with radio-immunoprecipitation assay (RIPA) buffer (Exon Biotechnology Inc., Guangzhou, China) and quantitated by the previously described method. The lysates were resolved by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto a polyvinylidene fluoride (PVDF) membrane (Bio-Rad Laboratories, Inc, CA, USA), which was blocked in 5% milk and blotted with various primary antibodies in 5% milk dissolved in TBS and Tween 20 (TBST) solution. The immunoblotting was tested with anti-NF-κB p65 (1:500), anti-IκBα (1:500), and anti-MEK1 (1:500) antibodies (Cell Signaling Technology, Inc., Danvers, MA, 01923, USA). Gel analysis software (GelPro5.0, Media Cybernetics, Bethesda, MD, USA) was used to quantify the density of the band by calculating the average optical density of each field.
Statistical analysis
PASW for Windows, version 18.0 (SPSS, Chicago, IL, USA) was used for statistical analysis. The data were represented by means ± SD in the analysis. The differences between the groups were compared by one-way analysis of variance (ANOVA) and the least significant difference test. The ANOVA and Tukey’s multiple comparison test were also adopted. p< 0.05 was significant statistically and Kruskal–Wallis test was conducted for the scoring of lung damage. A log-rank (Mantel–Cox) test was also performed for survival studies.
Results
Polygalasaponin F treatment protected mice from influenza challenge
This study assessed the effect of PSF on protecting IAV-infected mice. The clinical symptoms of mice appeared on the 3rd day after inoculation with IAV. More serious clinical symptoms, such as inactivity, ruffled fur, rapid shallow breathing, poor appetite, and weight loss, were detected by day 6.
Results show that the clinical symptoms of mice can be relieved by PSF or oseltamivir. The infected mice began to die on the 4th day after inoculation and reached the peak level on the 6th day (Fig. 2a, b). A significant decrease in the body weight of mice on days 3–9 after infection was compared with the NC group (Fig. 2a). However, the mice treated with oseltamivir and PSF (50, 100 and 200 mg/kg) had higher survival rate and body weight than the IAV-C group. All the infected mice of IAV-C group died within 15 days with the average survival time of only 5 days. In contrast, infected mice had longer survival time when treated with PSF (50, 100 and 200 mg/kg), as shown in Fig. 2b, c (p < 0.05–0.01). Therefore, PSF and oseltamivir can help to inhibit the mortality rate, reduce body weight loss, and prolong the survival time of mice. The mice of the NC group showed no clinical symptoms or died and their body weight increased during the study period. This indicated that PSF could efficiently protect mice from influenza virus.
Fig. 2.

Polygalasaponin F treatment protected mice from influenza challenge. BALB/c mice(n = 10) were treated for 2 h after viral challenge and 6 days after infection and monitored daily for signs and symptoms, body weight loss, and survival for 15 consecutive days. a Body weight; b survival rate; c survival time. Values are mean ± SD. Asterisks denote the significance levels: *p < 0.05; **p < 0.01; *** p < 0.001, compared with IAV-C group
PSF attenuated pulmonary inflammation in mice
To check the pathological changes in mice, we studied the HE staining and lung injury score. On day 6 after inoculation, the infected mice presented lesions in the lungs such as inflammatory cell infiltration, perivascular interstitial edema, and capillary bronchitis with edema (Fig. 3a—IAV-C). However, the pathological changes in the lung were improved when treated with PSF. No histological changes were found in the mice of the NC group. Infected mice treated with PSF (50–200 mg/kg) had lower lung histopathology score than the IAV-C group (Fig. 3b). On day 6 after infection, the lung index of IAV-C group increased significantly compared with that of the NC group (p < 0.001). However, after treated with PSF (50–200 mg/kg), the lung index dropped significantly (Fig. 3c) (p < 0.05–0.01). Similar to PSF, oseltamivir could also decrease the lung index (p < 0.01). Compared with the IAV-C group, the mRNA expression of influenza virus M gene was significantly inhibited when PSF (50–200 mg/kg) was used (Fig. 3d, p < 0.01), which indicated that PSF could reduce the inflammation induced by influenza virus.
Fig. 3.

Polygalasaponin F alleviated the severity of IAV-induced lung injuries at day 6 post-infection (n = 6–10). a Pathological changes of lung tissues (HE, × 100); b pathological scores. c Lung index. d Relative quantitation of influenza A virus in lung. Data were presented as mean ± SD. A black arrow shows representative damages, including interstitial congestion, edema, infiltration of inflammatory cells, pulmonary hemorrhage, and alveolar edema. Asterisks denote the significance levels: *p < 0.05; **p < 0.01; ***p < 0.001, compared with IAV-C group
PSF inhibited cytokine levels in lung of mice infected with IAV
As hypercytokinemia is usually related to IAV-induced pneumonia, we conducted the following experiments to determine whether PSF could affect the release of viral-induced proinflammatory cytokines.
After the infection by influenza virus, we extracted lung tissue from mice and analyzed the cytokines (IL-4, TNF-α, IL-1β and IFN-γ). The analysis showed that the IL-1β, TNF-α and IFN-γ levels in lung homogenate had a significant rise after 5 days of infection compared with the NC group (Fig. 4). But treatment with PSF (50, 100, and 200 mg/kg) had inhibited the increase of levels of IL-1β, TNF-α, IL-4, and IFN-γ in lung homogenates (Fig. 4a, c, d, p < 0.05–0.01). Oseltamivir also had a similar effect to that of PSF. So the experiment showed that PSF could significantly decrease the level of pulmonary cytokines induced by influenza virus.
Fig. 4.

Effects of polygalasaponin F on cytokine in lung homogenate in mice infected with IAV (n = 10). Proinflammatory cytokine levels were analyzed in lung homogenates of mice on day 6 after infection by ELISA. Data were presented as mean ± SD. Asterisks denote the significance levels: *p < 0.05; **p < 0.01; ***p < 0.001, compared with IAV-C group. a The level of IL-1beta in lung homogenate; b the level of IL-4 in lung homogenate; c the level of IFN-gamma in lung homogenate; d the level of TNF-alpha in lung homogenate
PSF inhibited the levels of TXA2 and PGE2 in lung of mice infected with IAV
Lung inflammation induced by influenza virus is accompanied by increased lung production of prostaglandins, leukotrienes, and lipid mediators (McCarthy and Weinberg 2012).
On day 6 post-infection, the levels of TXA2 and PGE2 of IAV-C group increased significantly compared with that of the NC group (p < 0.001). However, after treated with PSF (100–200 mg/kg), the levels of TXA2 and PGE2 declined significantly (Fig. 5a, b) (p < 0.05–0.001). Oseltamivir did not decrease the levels of TXA2 and PGE2 (p > 0.05).
Fig. 5.
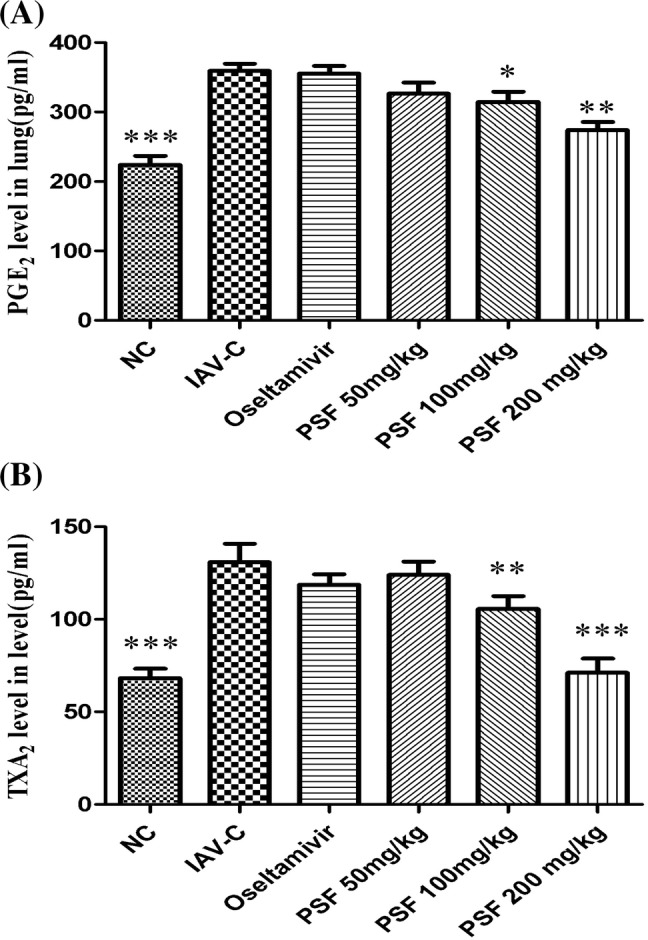
Effects of polygalasaponin F on lung TXA2 and PGE2 in mice infected with IAV (n = 10). The levels of TXA2 and PGE2 were analyzed in lung homogenates of mice on day 6 after infection by ELISA. Data were presented as mean ± SD. Asterisks denote the significance levels: *p < 0.05; **p < 0.01; ***p < 0.001, compared with IAV-C group. a The level of PGE2 in lung homogenate; b the level of TXA2 in lung homogenate
Polygalasaponin F decreased signaling pathway proteins involved in mice infected by IAV.
A broader antibody microarray (CSP100) was used to study whether PSF could alter the signaling pathway (Table 1). This antibody array contains 93 signaling pathway proteins, each with six replicates. Using a cutoff ratio of 0.88, we identified four cell pathway proteins, namely NF-κB p65, IκBα, and MEK1, were decreased by PSF in lung of mice infected with IAV (Table 1). There was a 2-fold or greater increase in 6 of the 93 proteins assessed in the array induced by influenza virus and the expression of MEK1 (Ab-217) increased the most (7.15-fold) (Table 1).
Table 1.
Effects of polygalasaponin F on signaling pathway proteins involved in mice infected by IAV
| Cell pathway protein | Fold change in intensities from protein between IAV-C group and PSF (100 mg/kg)group |
|---|---|
| IkB-alpha (Ab-42) | 5.19 |
| IkB-alpha (Phospho-Tyr42) | 2.56 |
| MEK1 (Ab-217) | 7.15 |
| MEK1 (Phospho-Ser221) | 2.39 |
| NFkB-p65 (Ab-529) | 2.68 |
| NFkB-p65 (Phospho-Thr254) | 1.46 |
Polygalasaponin F inhibited influenza virus-induced cell-signaling pathway protein expression in lungs
The expression of NF-κB p65, IκBα, and MEK1 in the lung of infected mice was assessed using western blot analysis to testify the results of antibody array (Fig. 6a). Results show that PSF could significantly inhibit the increase of the expression of NF-κB p65, IκBα, and MEK1 (Fig. 6b–d), which were also proved to be the targets of PSF.
Fig. 6.

Polygalasaponin F inhibited signaling pathway proteins expression in lung of mice infected with IAV by western blot analysis (n = 6). GAPDH protein was used as a loading control. a Representative PAGE of cell-signaling pathway proteins expression by western blotting. b Semi-quantitative analysis of IκBα protein. c Semi-quantitative analysis of NF-κB p65 protein. d Semi-quantitative analysis of MEK1 protein. Values are mean ± SD. Asterisks denote the significance levels: *p < 0.05; **p < 0.01; ***p < 0.001, compared with IAV-C group
Discussion
Influenza virus can infect upper and lower respiratory tracts and cause serious pneumonia with acute respiratory distress syndrome or even lead to death (Clemans et al. 2000). IAV infection would cause tissue damages or trigger unbalanced production of antiviral cytokines, also known as “cytokine burst” or “hypercytokinemia”, which had a certain connection with severe pneumonia caused by IAV strains (Cheung et al. 2002; de Jong et al. 2006). This study assessed the effect of PSF on the mice infected by IAV. The results showed that PSF could significantly relieve clinical symptoms, reduce the loss of body weight, improve the survival time, and alleviate lung lesions and life-threatening complications of infected mice. These results indicated that PSF could effectively protect mice from IAV infection.
Influenza viruses induce inflammatory cells to infiltrate lung tissue and release proinflammatory cytokines (such as IL-1beta and TNF-alpha), causing severe secondary pneumonia that contributed to the mortality, due to influenza infection (Matikainen et al. 2006; Bellani et al. 2010). In this study, PSF-treated mice had lower histopathology scores in the lung, showing that PSF might reduce the occurrence rate of pulmonary edema and relieve lung inflammation induced by influenza virus. In addition, PSF could inhibit the production of proinflammatory cytokines (e.g. TNF-α), and thus suppress the inflammatory responses in the tissues and cells (Yan et al. 2015; Wang et al. 2008). Our data show that influenza virus would significantly raise the IL-1β, TNF-α, IL-4, and IFN-γ levels in the lung tissue and the use of PSF could reduce the up-regulated proinflammatory cytokines. Besides, PSF could inhibit inflammation in the lung tissues and suppress excessive innate inflammatory responses, so that to protect mice infected with IAV from further damages.
This study found that influenza virus induce viral pneumonia in mice characterized by interstitial congestion, edema, infiltration of inflammatory cells, pulmonary hemorrhage, and alveolar edema. The lung index and pulmonary vascular permeability appeared to progressively increase, suggesting that the increase of pulmonary vascular permeability is one of the early lesions. In the IAV-C group and the PSF treatment group, lung TXA2 and PGE2 levels showed a synchronous change trend with pulmonary vascular permeability, lung index, and lung pathological changes, suggesting that lung damage caused by IAV was closely related to the production of TXA2 and PGE2. Prostaglandins [PGs, including prostaglandin E2 (PGE2), PGD2, PGF2α, PGI2, TXA2] are bioactive lipid mediators generated from arachidonic acid via the enzymatic activity of cyclooxygenases (COXs) and participate in the pathogenesis of inflammatory disease (Park et al. 2006; Davies et al. 1984). TXA2 is by far the strongest platelet aggregator and vasoconstrictor of vascular and bronchial smooth muscle (van Ooijen et al. 1990). TXA2 constricts pulmonary vascular and bronchial smooth muscle, promotes platelet aggregation and secretion, and aggravates lung injury. In addition, TXA2 can mediate the adhesion and aggregation of neutrophils, release harmful factors such as oxygen free radicals, leukotrienes, and platelet activating factors, thus aggravating lung damage (Westphal et al. 2005). PGE2 is the most abundant prostanoid in the mammalian body and plays a role in regulation of immune responses, blood pressure, gastrointestinal integrity, and inflammation (Coulombe and Divangahi 2014). PGE2 acts as a mediator of pain and fever in inflammatory response and participates in the pathophysiological process of inflammation. All the classic signs of inflammation (swelling, redness, heat, and pain) can be attributed to PGE2 (Funk 2001). IAV hyper induces the COX-2 and PGE2 production (Liu et al. 2012). In this study, PSF can inhibit the release of TXA2 and PGE2 to a certain extent, thereby effectively alleviate the inflammatory response, which may be one of the mechanisms of anti-influenza effect of PSF. Our study showed that even if the viral replication have been suppressed in the mice treated with oseltamivir, levels of TXA2 and PGE2 were still similar to the IAV-C mice, which were significantly higher than those in the mice receiving PSF therapy. This finding suggests that once the viral infection has triggered the cytokine storm, inflammatory cytokines will continue to drive the immunopathologic progression (Zheng et al. 2008) and antiviral therapy cannot completely inhibit this process.
Influenza virus infection would activate intracellular signaling pathways to support efficient replications and Raf/MEK/ERK-cascade and NF-κB would be activated by the influenza virus during the replications (Haasbach et al. 2013). As the main regulator of influenza cytokines and chemokines production (e.g. TNF-α, TNF-β, IL-1 β, IL-4, IL-6, IL-10 and IFN-γ), the activation of NF-κB is a hallmark of host cells responding to influenza virus infection (Barnes and Karin 1997). Overexpression of IAV-induced cytokines and chemokines is subjected to the NF-κB signaling pathway (Barnes and Karin 1997). The activation and inactivation of NF-κB could directly regulate the transcription of inflammatory cytokines and adhesion factors such as interferon (IFN)-β and tumor necrosis factor-α (TNF-α) (Chu et al. 1999). These cytokines and adhesion factors are considered to have connection with acute lung injury (ALI) (Lee et al. 2005). As previously studied, the nuclear translocation of IκBα and NF-κB -dependent transcriptional activity can be inhibited by treating with PSF (Yan et al., 2015; Wei et al., 2014). The expression of NF-κB p65 (major component of NF-κB) and MEK1 had a significant decrease in the lung of IAV-infected mice after treated with PSF. PSF could inhibit signaling by Raf/MEK/ERK and NF-KappaB, which would interfere with the events and suppress virus production and inflammatory responses.
Polygala japonica Houtt (Gua-zi-jin in Chinese) is used in traditional medicine for the treatment of various inflammatory disorders such as acute tonsillitis, pharyngitis and pulmonitis (Wang et al., 2008). Previous studies have reported triterpenoid saponins from P. japonica exhibited the anti-inflammatory activities on carageenan-induced mouse paw edema and nitric oxide (NO) production in LPS-stimulated RAW264.7 macrophages (Wang et al. 2008; Kou et al. 2006; Kim et al. 2009). Triterpenoid saponins from the roots of P. japonica Houtt. have anti-inflammatory activity through inhibition of nitrite and PGE2 production, iNOS and, COX-2 protein expression and TNF-α, IL-1β, IL-6, and IL-12 mRNA expression in LPS-stimulated BV2 cells (Kim et al. 2009; Ahn et al. 2005). Previous research has reported that PSF inhibit the secretion of inflammatory cytokines induced by LPS and its mechanism is associated with the NF-κB signaling pathway (Wei et al. 2014; Yan et al. 2015), but the effect and mechanism vary in different conditions. This study demonstrated that PSF can block the production of cytokines and prostaglandins and inhibit viral pneumonia in mice infected with IAV, and its mechanism is associated with the suppression of Raf/MEK/ERK and NF-κB expressions, which may support the utility of P. japonica in respiratory infection, especially IAV-induced pneumonia.
There is an urgent need to find an effective treatment strategy against IAV infection in humans, because of the substantial mortality associated with these viruses. Although oseltamivir is, currently, most widely used for the treatment of influenza, it still exhibited high fatality, which can be attributed to delayed initiation of therapy (Zheng et al. 2008). Influenza virus infection triggers the cytokine storm and includes following hallmarks: excessive proinflammatory cytokines, chemokines, and inflammatory cells recruitment into lungs (La Gruta et al. 2007). Drugs with anti-inflammatory actions have controlled excessive inflammation from severe influenza in animal models such as the COX-2 inhibitor, naproxen (Lejal et al. 2013). This finding suggests that viral replication is suppressed by oseltamivir, but the immunopathologic progression is not completely suppressed. The antiviral activity of oseltamivir involves NA inhibition, which differs from the anti-inflammatory mechanism of PSF.
In sum, our study supports the new evidence that PSF blocks cytokine production and inhibits viral pneumonia in mice infected with influenza A virus. However, the mechanism of PSF in other subtypes and strains of influenza virus in vitro and the effect of PSF on influenza viral pneumonia patients need further study.
Acknowledgements
We thank International Science Editing (Co. Clare, Ireland) for the language editorial assistance in the preparation of the manuscript.
Compliance with ethical standards
Conflict of interest
We wish to confirm that there are no known conflicts of interest associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Ahn KS, Noh EJ, Zhao HL, Jung SH, Kang SS, Kim YS. Inhibition of inducible nitric oxide synthase and cyclooxygenase II by Platycodon grandiflorum saponins via suppression of nuclear factor-kappaB activation in RAW 264.7 cells. Life Sci. 2005;76(20):2315–2328. doi: 10.1016/j.lfs.2004.10.042. [DOI] [PubMed] [Google Scholar]
- Barnes PJ, Karin M. Nuclear factor-kappaB: a pivotal transcription factor in chronic inflammatory diseases. N Engl J Med. 1997;336(15):1066–1071. doi: 10.1056/NEJM199704103361506. [DOI] [PubMed] [Google Scholar]
- Bellani G, Guerra L, Pesenti A, Messa C. Imaging of lung inflammation during severe influenza A: H1N1. Intensive Care Med. 2010;36(4):717–718. doi: 10.1007/s00134-010-1756-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen L, Yan X, Yan Q, Fan J, Huang H, Shi X, Han L, Han T, Zhu H. The modified JiuWei QiangHuo decoction alleviated severe lung injury induced by H1N1 influenza virus by regulating the NF-kappa B pathway in mice. Evid Based Compl Alternat Med. 2015;2015:790739. doi: 10.1155/2015/790739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheung TK, Poon LL. Biology of influenza a virus. Ann NY Acad Sci. 2007;1102:1–25. doi: 10.1196/annals.1408.001. [DOI] [PubMed] [Google Scholar]
- Cheung CY, Poon LL, Lau AS, Luk W, Lau YL, Shortridge KF, Gordon S, Guan Y, Peiris JS. Induction of proinflammatory cytokines in human macrophages by influenza A (H5N1) viruses: a mechanism for the unusual severity of human disease? Lancet. 2002;360(9348):1831–1837. doi: 10.1016/s0140-6736(02)11772-7. [DOI] [PubMed] [Google Scholar]
- Chien YS, Su CP, Tsai HT, Huang AS, Lien CE, Hung MN, Chuang JH, Kuo HS, Chang SC. Predictors and outcomes of respiratory failure among hospitalized pneumonia patients with 2009 H1N1 influenza in Taiwan. J Infect. 2010;60(2):168–174. doi: 10.1016/j.jinf.2009.12.012. [DOI] [PubMed] [Google Scholar]
- Choi WY, Kim S, Lee N, Kwon M, Yang I, Kim MJ, Cheong SG, Kwon D, Lee JY, Oh HB, Kang C. Amantadine-resistant influenza A viruses isolated in South Korea from 2003 to 2009. Antiviral Res. 2009;84(2):199–202. doi: 10.1016/j.antiviral.2009.08.006. [DOI] [PubMed] [Google Scholar]
- Chu WM, Ostertag D, Li ZW, Chang L, Chen Y, Hu Y, Williams B, Perrault J, Karin M. JNK2 and IKKbeta are required for activating the innate response to viral infection. Immunity. 1999;11(6):721–731. doi: 10.1016/s1074-7613(00)80146-6. [DOI] [PubMed] [Google Scholar]
- Chung IW, Moore NA, Oh WK, O'Neill MF, Ahn JS, Park JB, Kang UG, Kim YS. Behavioural pharmacology of polygalasaponins indicates potential antipsychotic efficacy. Pharmacol Biochem Behav. 2002;71(1–2):191–195. doi: 10.1016/s0091-3057(01)00648-7. [DOI] [PubMed] [Google Scholar]
- Clemans DL, Bauer RJ, Hanson JA, Hobbs MV, 3rd St Geme JW, Marrs CF, Gilsdorf JR. Induction of proinflammatory cytokines from human respiratory epithelial cells after stimulation by nontypeable Haemophilus influenzae. Infect Immun. 2000;68(8):4430–4440. doi: 10.1128/iai.68.8.4430-4440.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coulombe F, Divangahi M. Targeting eicosanoid pathways in the development of novel anti-influenza drugs. Expert Rev Anti Infect Ther. 2014;12(11):1337–1343. doi: 10.1586/14787210.2014.966082. [DOI] [PubMed] [Google Scholar]
- Davies P, Bailey PJ, Goldenberg MM, Ford-Hutchinson AW. The role of arachidonic acid oxygenation products in pain and inflammation. Annu Rev Immunol. 1984;2:335–357. doi: 10.1146/annurev.iy.02.040184.002003. [DOI] [PubMed] [Google Scholar]
- De Clercq E. Antiviral agents active against influenza A viruses. Nat Rev Drug Discov. 2006;5(12):1015–1025. doi: 10.1038/nrd2175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Jong MD, Simmons CP, Thanh TT, Hien VM, Smith GJ, Chau TN, Hoang DM, Chau NV, Khanh TH, Dong VC, Qui PT, Cam BV, do Ha Q, Guan Y, Peiris JS, Chinh NT, Hien TT, Farrar J. Fatal outcome of human influenza A (H5N1) is associated with high viral load and hypercytokinemia. Nat Med. 2006;12(10):1203–1207. doi: 10.1038/nm1477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denholm JT, Gordon CL, Johnson PD, Hewagama SS, Stuart RL, Aboltins C, Jeremiah C, Knox J, Lane GP, Tramontana AR, Slavin MA, Schulz TR, Richards M, Birch CJ, Cheng AC. Hospitalised adult patients with pandemic (H1N1) 2009 influenza in Melbourne, Australia. Med J Aust. 2010;192(2):84–86. doi: 10.5694/j.1326-5377.2010.tb03424.x. [DOI] [PubMed] [Google Scholar]
- Ding Y, Cao Z, Cao L, Ding G, Wang Z, Xiao W. Antiviral activity of chlorogenic acid against influenza A (H1N1/H3N2) virus and its inhibition of neuraminidase. Sci Rep. 2017;7:45723. doi: 10.1038/srep45723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrhardt C, Marjuki H, Wolff T, Nurnberg B, Planz O, Pleschka S, Ludwig S. Bivalent role of the phosphatidylinositol-3-kinase (PI3K) during influenza virus infection and host cell defence. Cell Microbiol. 2006;8(8):1336–1348. doi: 10.1111/j.1462-5822.2006.00713.x. [DOI] [PubMed] [Google Scholar]
- Fu J, Zuo L, Yang J, Chen R, Zhang D. Oligosaccharide polyester and triterpenoid saponins from the roots of Polygala japonica. Phytochemistry. 2008;69(7):1617–1624. doi: 10.1016/j.phytochem.2008.01.010. [DOI] [PubMed] [Google Scholar]
- Funk CD. Prostaglandins and leukotrienes: advances in eicosanoid biology. Science. 2001;294(5548):1871–1875. doi: 10.1126/science.294.5548.1871. [DOI] [PubMed] [Google Scholar]
- Haasbach E, Reiling SJ, Ehrhardt C, Droebner K, Ruckle A, Hrincius ER, Leban J, Strobl S, Vitt D, Ludwig S, Planz O. The NF-kappaB inhibitor SC75741 protects mice against highly pathogenic avian influenza A virus. Antiviral Res. 2013;99(3):336–344. doi: 10.1016/j.antiviral.2013.06.008. [DOI] [PubMed] [Google Scholar]
- Kim SH, Jang SD, Lee KY, Sung SH, Kim YC. Chemical constituents isolated from Polygala japonica leaves and their inhibitory effect on nitric oxide production in vitro. J Enzyme Inhib Med Chem. 2009;24(1):230–233. doi: 10.1080/14756360802051362. [DOI] [PubMed] [Google Scholar]
- Kiso M, Shinya K, Shimojima M, Takano R, Takahashi K, Katsura H, Kakugawa S, Le MT, Yamashita M, Furuta Y, Ozawa M, Kawaoka Y. Characterization of oseltamivir-resistant 2009 H1N1 pandemic influenza A viruses. PLoS Pathog. 2010;6(8):e1001079. doi: 10.1371/journal.ppat.1001079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kou J, Si M, Dai G, Lin Y, Zhu D. Antiinflammatory activity of Polygala japonica extract. Fitoterapia. 2006;77(6):411–415. doi: 10.1016/j.fitote.2006.04.011. [DOI] [PubMed] [Google Scholar]
- La Gruta NL, Kedzierska K, Stambas J, Doherty PC. A question of self-preservation: immunopathology in influenza virus infection. Immunol Cell Biol. 2007;85(2):85–92. doi: 10.1038/sj.icb.7100026. [DOI] [PubMed] [Google Scholar]
- Langlois RA, Meyerholz DK, Coleman RA, Cook RT, Waldschmidt TJ, Legge KL. Oseltamivir treatment prevents the increased influenza virus disease severity and lethality occurring in chronic ethanol consuming mice. Alcohol Clin Exp Res. 2010;34(8):1425–1431. doi: 10.1111/j.1530-0277.2010.01226.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee CJ, Liao CL, Lin YL. Flavivirus activates phosphatidylinositol 3-kinase signaling to block caspase-dependent apoptotic cell death at the early stage of virus infection. J Virol. 2005;79(13):8388–8399. doi: 10.1128/JVI.79.13.8388-8399.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lejal N, Tarus B, Bouguyon E, Chenavas S, Bertho N, Delmas B, Ruigrok RW, Di Primo C, Slama-Schwok A. Structure-based discovery of the novel antiviral properties of naproxen against the nucleoprotein of influenza A virus. Antimicrob Agents Chemother. 2013;57(5):2231–2242. doi: 10.1128/AAC.02335-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y, Xu YL, Lai YN, Liao SH, Liu N, Xu PP. Intranasal co-administration of 1,8-cineole with influenza vaccine provide cross-protection against influenza virus infection. Phytomedicine. 2017;34:127–135. doi: 10.1016/j.phymed.2017.08.014. [DOI] [PubMed] [Google Scholar]
- Liao SH, Li Y, Lai YN, Liu N, Zhang FX, Xu PP. Ribavirin attenuates the respiratory immune responses to influenza viral infection in mice. Arch Virol. 2017;162(6):1661–1669. doi: 10.1007/s00705-017-3291-7. [DOI] [PubMed] [Google Scholar]
- Liu L, Cao Z, Chen J, Li R, Cao Y, Zhu C, Wu K, Wu J, Liu F, Zhu Y. Influenza A virus induces interleukin-27 through cyclooxygenase-2 and protein kinase A signaling. J Biol Chem. 2012;287(15):11899–11910. doi: 10.1074/jbc.M111.308064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matikainen S, Siren J, Tissari J, Veckman V, Pirhonen J, Severa M, Sun Q, Lin R, Meri S, Uze G, Hiscott J, Julkunen I. Tumor necrosis factor alpha enhances influenza A virus-induced expression of antiviral cytokines by activating RIG-I gene expression. J Virol. 2006;80(7):3515–3522. doi: 10.1128/JVI.80.7.3515-3522.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCarthy MK, Weinberg JB. Eicosanoids and respiratory viral infection: coordinators of inflammation and potential therapeutic targets. Mediators Inflamm. 2012;2012:236345. doi: 10.1155/2012/236345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park JY, Pillinger MH, Abramson SB. Prostaglandin E2 synthesis and secretion: the role of PGE2 synthases. Clin Immunol. 2006;119(3):229–240. doi: 10.1016/j.clim.2006.01.016. [DOI] [PubMed] [Google Scholar]
- Pawestri HA, Nugraha AA, Hariastuti NI, Setiawaty V. Detection of neuraminidase inhibitor-resistant influenza A (H1N1)pdm09 viruses obtained from influenza surveillance in Indonesia. SAGE Open Med. 2018;6:2050312118818293. doi: 10.1177/2050312118818293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrone LA, Plowden JK, Garcia-Sastre A, Katz JM, Tumpey TM. H5N1 and 1918 pandemic influenza virus infection results in early and excessive infiltration of macrophages and neutrophils in the lungs of mice. PLoS Pathog. 2008;4(8):e1000115. doi: 10.1371/journal.ppat.1000115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pires de Mello CP, Drusano GL, Adams JR, Shudt M, Kulawy R, Brown AN. Oseltamivir-zanamivir combination therapy suppresses drug-resistant H1N1 influenza A viruses in the hollow fiber infection model (HFIM) system. Eur J Pharm Sci. 2018;111:443–449. doi: 10.1016/j.ejps.2017.10.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Ooijen B, Kort WJ, Tinga CJ, Wilson JH. Significance of thromboxane A2 and prostaglandin I2 in acute necrotizing pancreatitis in rats. Dig Dis Sci. 1990;35(9):1078–1084. doi: 10.1007/BF01537578. [DOI] [PubMed] [Google Scholar]
- Wang H, Gao J, Kou J, Zhu D, Yu B. Anti-inflammatory activities of triterpenoid saponins from Polygala japonica. Phytomedicine. 2008;15(5):321–326. doi: 10.1016/j.phymed.2007.09.014. [DOI] [PubMed] [Google Scholar]
- Wei W, Yuan YH, Gao YN, Yan WF, Li CJ, Zhang DM, Chen NH. Polygalasaponin F inhibits secretion of inflammatory cytokines via NF-kappaB pathway regulation. J Asian Nat Prod Res. 2014;16(8):865–875. doi: 10.1080/10286020.2014.918962. [DOI] [PubMed] [Google Scholar]
- Westphal M, Noshima S, Isago T, Fujioka K, Maybauer MO, Maybauer DM, Traber LD, Flynn JT, Westphal-Varghese BB, Traber DL. Selective thromboxane A2 synthase inhibition by OKY-046 prevents cardiopulmonary dysfunction after ovine smoke inhalation injury. Anesthesiology. 2005;102(5):954–961. doi: 10.1097/00000542-200505000-00014. [DOI] [PubMed] [Google Scholar]
- Yan WF, Shao QH, Zhang DM, Yuan YH, Chen NH. The molecular mechanism of polygalasaponin F-mediated decreases in TNFalpha: emphasizing the role of the TLR4-PI3K/AKT-NF-kappaB pathway. J Asian Nat Prod Res. 2015;17(6):662–670. doi: 10.1080/10286020.2015.1056166. [DOI] [PubMed] [Google Scholar]
- Zhang D, Miyase T, Kuroyanagi M, Umehara K, Ueno A. Five new triterpene saponins, polygalasaponins XXVIII-XXXII from the root of Polygala japonica Houtt. Chem Pharm Bull (Tokyo) 1996;44(4):810–815. doi: 10.1248/cpb.44.810. [DOI] [PubMed] [Google Scholar]
- Zheng BJ, Chan KW, Lin YP, Zhao GY, Chan C, Zhang HJ, Chen HL, Wong SS, Lau SK, Woo PC, Chan KH, Jin DY, Yuen KY. Delayed antiviral plus immunomodulator treatment still reduces mortality in mice infected by high inoculum of influenza A/H5N1 virus. Proc Natl Acad Sci USA. 2008;105(23):8091–8096. doi: 10.1073/pnas.0711942105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou W, Zhang J, Xu W, Sun J. Quantification of polygalasaponin F in rat plasma using liquid chromatography-tandem mass spectrometry and its pharmacokinetics application. Biomed Chromatogr. 2015;29(9):1388–1392. doi: 10.1002/bmc.3435. [DOI] [PubMed] [Google Scholar]