

Abstract
Since the discovery that certain small viral membrane proteins, collectively termed as viroporins, can permeabilize host cellular membranes and also behave as ion channels, attempts have been made to link this feature to specific biological roles. In parallel, most viroporins identified so far are virulence factors, and interest has focused toward the discovery of channel inhibitors that would have a therapeutic effect, or be used as research tools to understand the biological roles of viroporin ion channel activity. However, this paradigm is being shifted by the difficulties inherent to small viral membrane proteins, and by the realization that protein–protein interactions and other diverse roles in the virus life cycle may represent an equal, if not, more important target. Therefore, although targeting the channel activity of viroporins can probably be therapeutically useful in some cases, the focus may shift to their other functions in following years. Small-molecule inhibitors have been mostly developed against the influenza A M2 (IAV M2 or AM2). This is not surprising since AM2 is the best characterized viroporin to date, with a well-established biological role in viral pathogenesis combined the most extensive structural investigations conducted, and has emerged as a validated drug target. For other viroporins, these studies are still mostly in their infancy, and together with those for AM2, are the subject of the present review.
Keywords: Viroporins, Viruses, Channel activity, Drug discovery, Inhibitors, Protein–protein interactions
1. Introduction
The field of viroporins was born about 20 years ago from the realization that viruses inflict injuries in the membranes of cells during infection, which results in increased membrane permeability (Carrasco, 1995). This early observation heralded the current interest in viroporins, along with the therapeutic opportunities afforded by the modulation of their channel activity using small molecules (Fischer et al., 2012, Scott and Griffin, 2015), and the deeper understanding of the roles of viroporin ion channel activity on viral replication and pathogenesis (Nieto-Torres, Verdia-Baguena, Castano-Rodriguez, Aguilella, & Enjuanes, 2015).
This interest will likely increase with the discovery of new viroporins along with their ever-growing roles in the virus life cycle. Indeed, the number of medically important viroporins known is expected to rise as new human viruses continuously emerge from animal hosts. It is also possible that new viroporins may have novel features and structural motifs which will contribute in updating their classifications in terms of topology and number of transmembrane α-helical domains (Nieva, Madan, & Carrasco, 2012). In vertebrates alone, the number of viruses was estimated to be around one million (Morse, 1993). More recent estimates quantify the viral diversity in mammals, which are the reservoir hosts of the majority of emerging zoonoses, as 320,000 (Anthony et al., 2013). This strongly contrasts with the few thousand viruses identified to date, which means that more than 99% of viruses—and their viroporins, if any—are still unknown.
Most viroporins have been identified as virulence factors that lead to viral attenuation when deleted. This attenuation is attributed only in part to their channel activity, but nevertheless small-molecule channel inhibitors have been sought after. The vast majority of these channel inhibitors have been developed against the influenza A virus M2 (IAV M2 or AM2) protein (reviewed recently in Du et al., 2012, Gu et al., 2013), which is the first viroporin discovered. This is not surprising since AM2 is the best characterized viroporin to date, with a well-established biological role in viral pathogenesis combined the most extensive structural investigations conducted, and has emerged as a validated drug target. For other viroporins, these studies are still mostly in their infancy, although a high-resolution structure of the hepatitis C virus (HCV) p7 protein has been recently described (Ouyang et al., 2013) that may be useful for the rational design of p7 channel inhibitors in the future.
However, attempts to rationally design small-molecule inhibitors against viroporins will encounter several challenges. One is the potential for sequence variability in the viroporin. Indeed, it has been argued that since the number of genes susceptible to deleterious mutations increase with genome size, mutation rates are higher for small genomes (Drake, 1969). A related problem is that viral proteins, when compared to prokaryotic and eukaryotic proteins, have a higher degree of structural flexibility that may represent adaptation mechanisms to increase resistance to random mutations, especially in the case of RNA viruses, which mutate and evolve faster than DNA viruses. A less dense packing, and a concomitant smaller number of network interactions, translates not only to a smaller difference in energy between folded and unfolded states, but an also smaller percentual contribution of each mutation to stability (Tokuriki, Oldfield, Uversky, Berezovsky, & Tawfik, 2009). Additionally, this enhanced flexibility may have an impact on the ability of a single protein to perform a variety of tasks, consistent with the economical use of resources in viruses. Adding to this intrinsic flexibility, another problem in structure-based drug discovery concerning viroporins is the strong dependence of the protein structure on environment. Indeed, solution NMR can be applied to study small-membrane proteins in membrane-mimicking environments that allow rapid reorientation, but these artificial systems may also alter the intrinsic properties of the proteins (Cross et al., 2012, Cross et al., 2011). Last, some viroporins seem to form a protein–lipid complex rather than a purely proteinaceous pore, as shown for the VP4 protein in the triatoma virus (Sanchez-Eugenia, Goikolea, Gil-Carton, Sanchez-Magraner, & Guerin, 2015), and similar observations have also been reported for coronavirus envelope proteins (Verdia-Baguena et al., 2012).
In the quest to discover small molecules targeting viroporins, amantadine (Amt) can be considered to be the first viroporin channel inhibitor, and one of the first antivirals licensed for use in humans. Although it was known that this adamantane was active against IAV (Davies et al., 1964, Wingfield et al., 1969), the mechanism of inhibition was described only in 1992 (Duff and Ashley, 1992, Pinto et al., 1992). The target itself, AM2, was not known until 1985 (Hay, Wolstenholme, Skehel, & Smith, 1985). Even today, Amt, together with its methylated derivative rimantadine (Rim) which was licensed in the 1980s, are the only licensed antiviral drugs that target viroporins. This contrasts with the availability of hundreds of compounds that exert their effects with subnanomolar affinities on nonviral ion channels. Although this phenomenon could be explained in principle by a lack of structural data for viroporins, this is not the case for AM2, where precise structural data has been obtained, even for some of its Amt-resistant mutants (Pielak and Chou, 2011, Wang, Qiu, et al., 2011). Unfortunately, an effective alternative to adamantanes has not yet been licensed.
2. The Influenza A Virus Matrix Protein 2 (IAV M2 or AM2)
With our current knowledge, the AM2 channel provides yet another paradigm valid for other viroporins. It has a very simple structure compared to ion channels from higher organisms (such as K+ and Ca2 + channels), but the mechanism of proton conduction and drugs binding and inhibition are not yet fully understood. Indeed, the structural and functional properties of the AM2 protein vary widely in changing experimental conditions, such as peptide length, drug binding, lipid composition, lipid thickness, or pH (Acharya et al., 2010, Cady et al., 2011, Cross et al., 2012, Kovacs et al., 2000, Thomaston et al., 2013, Zhou and Cross, 2013). An important factor in this sensitivity is the direct contact of its TM domains with the lipid molecules, in contrast to higher organism ion channels (eg, K+ channels) (Cuello, Jogini, Cortes, & Perozo, 2010), where pore-forming α-helices are surrounded and protected by additional transmembrane helices. However, while the AM2 channel exists in a variety of conformational states in the apo-form, inhibitor-binding significantly reduces the conformational flexibility of the channel. The structural dynamics of the channel represents another drawback in the use of high-resolution structures for rationally designed drug discovery.
Amantadine is just one of the four antivirals against IAV available: two neuraminidase inhibitors (oseltamivir and zanamivir) and two AM2 channel blockers (Amt and Rim) (Vanderlinden & Naesens, 2014). The seasonal flu caused by IAV is associated with high medical burden (Molinari et al., 2007) and sudden pandemics with high mortality rates (Hay et al., 2001, Neumann et al., 2009). Currently, most circulating strains are Amt resistant (Bright et al., 2006, Bright et al., 2005, Deyde et al., 2007, Hayden and De Jong, 2011) and therefore the use of Amt and Rim has been discontinued for the human population (Fiore et al., 2011).
The AM2 channel is a homotetramer (Sakaguchi, Tu, Pinto, & Lamb, 1997), where each monomer has 97 amino acids containing one α-helical TM domain (residues 25–46). The extramembrane C-terminal domain (residues 47–97) is exposed to the cytoplasmic side (Lamb, Zebedee, & Richardson, 1985) and contains an amphipathic helix (residues 51–59). Four of these amphipathic helices stabilize the tetrameric channel, forming a base on the membrane surface almost perpendicular to the TM bundle. The end of this C-terminal tail is disordered and interacts with the viral matrix protein 1 (M1). The M2 protein performs several critical roles for virus replication (Pinto & Lamb, 2006). For instance, M2-mediated acidification of the interior of the virus is required for the uncoating of genetic material inside the cell (Pinto et al., 1992). M2 is also required to regulate the intralumenal pH of the Golgi apparatus preventing the premature conformational change of hemagglutinin (Takeuchi & Lamb, 1994).
Influenza B, the closest relative of the influenza A virus, accounts for about 50% of all influenza disease in recent years (according to the US Centers for Disease Control and Prevention website, www.cdc.gov) and has a similarly essential pH-activated proton channel M2 viroporin, the BM2 (Hatta et al., 2004, Pinto and Lamb, 2006). Although AM2 and BM2 share almost no sequence identity, both have a single TM domain and form homotetramers (Paterson et al., 2003, Wang, Pielak, et al., 2009). In comparison, BM2 is completely insensitive to Amt or Rim (Mould et al., 2003, Pinto and Lamb, 2006) and no BM2 inhibitors have been identified yet.
The location of the binding site for the adamantanes Amt and Rim in AM2 was initially controversial. A crystallographic structure suggested a pore-binding model in the amino-terminal part of the TM (22–46), ie, the P-binding site (Stouffer et al., 2008) (Fig. 1A–C). In the same and in the following year, a solution NMR-based structure of a longer construct of AM2 (18–60) (Pielak et al., 2009, Schnell and Chou, 2008) proposed a surface-binding site (the S-binding site) for Rim, on the carboxy-terminal, lipid-facing surface of the helices (Fig. 1D–G), suggesting an alternative allosteric mechanism of inhibition. In this allosteric binding site, the adamantane group interacted favourably with the hydrophobic side chains of Leu-40, Leu-43, and Ile-42, whereas the positively charged ammonium group interacted with the polar patch formed by Asp-44 and Arg-45 from the adjacent subunit (Fig. 1G). A dilemmatic feature of allosteric sites is that viral mutations at these sites are easily accommodated by viruses without dramatic loss of fitness. In HCV, for example, protease inhibitor monotherapy can rapidly select resistant viral populations within a few days (Schmidt et al., 2014).
Figure 1.

P- and S-binding sites for Amt and Rim to IAV M2. (A–C) AM2 TM domain crystallized in detergent, with the most critical residues identified by site-directed mutagenesis lining the AM2 pore (A); omit map showing electron density in the Amt binding region (B); structure of amantadine (nitrogen in cyan (light gray in the print version) and carbon in white) inside the binding site showing the surface associated with residues Val-27 (red (light gray in the print version) surface), Ala-30 (green (dark gray in the print version)), Ser-31 (blue (light gray in the print version)), and Gly-34 (orange (light gray in the print version)) (C); (D and E), an ensemble of 15 low-energy structures derived from NMR restraints, with TM α-helices (residues 25–46) and amphipathic helices (residues 51–59) superimposed separately (D); ribbon representation, showing the drug Rim (colored in red (light gray in the print version)) (E); (F and G), surface representation of the Rim-binding pocket, showing the Asp-44, the indole amine of Trp-41, and Arg-45, which form the polar patch, as well as the hydrophobic wall composed of Leu-40, Ile-42, and Leu-43; (H–J) solution NMR structures of the (AM2–BM2)TM channel in the absence and presence of Rim; ensembles of 15 low-energy structures of drug-bound chimera channels. Rim is highlighted in red (light gray in the print version) (H); ribbon representation of drug-free (AM2–BM2)TM tetramer (left), and overlay of its AM2 and BM2 regions (green (dark gray in the print version)) with the corresponding regions (yellow (light gray in the print version)) of the AM2 (PDB code: 2RLF) and BM2 (PDB code: 2KIX) structures (right) (I); overlay of the drug-free (white) and the drug-bound (cyan (light gray in the print version)) chimera structures, showing substantial differences in helical packing (J).
Adapted by permission from Macmillan Publishers Ltd.: Stouffer, A. L., Acharya, R., Salom, D., Levine, A. S., Di Costanzo, L., Soto, C. S., et al. (2008). Structural basis for the function and inhibition of an influenza virus proton channel. Nature, 451(7178), 596–599. Copyright (2008); Schnell, J. R., & Chou, J. J. (2008). Structure and mechanism of the M2 proton channel of influenza A virus. Nature, 451(7178), 591–595. doi: 10.1038/nature06531. Copyright (2008). Adapted by permission from Elsevier: Pielak, R. M., Oxenoid, K., & Chou, J. J. (2011). Structural investigation of rimantadine inhibition of the AM2-BM2 chimera channel of influenza viruses. Structure, 19(11), 1655–1663. doi: 10.1016/j.str.2011.09.003. Copyright (2011).
However, clear support for the P-binding site was obtained from solid-state NMR in lipid bilayers (PDB ID: 2KQT) (Cady et al., 2010); the inhibitor was found in a hydrophilic pocket formed by Ala-30, Ser-31, and Gly-34, ie, occluding the AM2 channel. In this case, hydrophobic interactions were detected between the adamantane group and Val-27 side chains, whereas the ammonium group was hydrogen-bonded to pore-facing residues and water molecules. The latter work revealed that the S-binding site was in fact just a low-affinity site, and only observed at high concentrations of the drug in the membrane. Importantly, Amt was found to undergo significant motion in the N-terminal lumen, suggesting that its structure can be improved for a better fit to the AM2 channel. This pore-location (P-binding) was also further validated by Chou et al. by solution NMR (Pielak, Oxenoid, & Chou, 2011) using an M2 chimeric sample, where the N-terminal part of the M2 TM domain corresponded to that of the AM2, whereas the C-terminal part was from the Amt-insensitive BM2 (Fig. 1H–J). In that model, methyl groups of Val-27 and Ala-30 from four subunits form a hydrophobic pocket around the adamantane, with the amino group in polar contact with the backbone oxygen of Ala-30. This ended the controversy, with the acceptance of the initially proposed P-binding site.
There are 12 structures of the wild-type and drug-resistant mutant M2 channels available in the Protein Data Bank solved by different techniques and conditions (see summary in Gu et al., 2013). Among these, structure 2RLF solved in micelle (Schnell & Chou, 2008) and structure 2L0J in the bilayer environment contain, in addition to the TM domain, short C-terminal intracellular amphipathic helices. In 2RLF, the C-terminal helices were connected to the transmembrane helices via a short loop (residue 47–51) and were exposed to the solution. However, in 2L0J, the C-terminal helices were connected to the TM helices through a rigid turn (residue 47) and were positioned on the bilayer surface (Sharma et al., 2010), and this conformation was stabilized by extensive hydrophobic interactions among these helices, and polar residues Asp-44 and Arg-45 were buried by hydrophobic residues of the intracellular amphipathic helices, making the S-binding site more hydrophobic than that in micelles. Therefore, the S-binding site was absent in 2L0J (Sharma et al., 2010).
Molecular dynamics simulations of the M2 proton channel with inhibitors binding at different sites have shown that the P-binding site is more stable for drug binding, but a higher energy barrier needs to be overcome for binding to occur (Gu et al., 2011). The S-binding site was less stable for drug binding but it was nearly barrier less and was easily accessed. Therefore, the P-binding site represents the thermodynamic binding site where the drug molecule binds slowly and stably and dissociates even more slowly, whereas the S-binding site is a kinetic binding site where the drug molecule binds readily but less stably and dissociates easily.
Over the last four decades, systematic studies of amantadine analogues and library screening have elucidated structure–activity relationships (SARs) and helped to identify potent wild-type (WT) AM2 channel blockers (De Clercq, 2006, Lagoja and De Clercq, 2008). Several other molecules were not based on adamantanes. For example, the spirene guanidine analogue, 2-[3-azaspiro(5,5)undecanol]-2-imidazoline (BL-1743) (Fig. 2A) was discovered during a high-throughput screen based on the ability of inhibitors to reverse the toxicity associated with M2 channels expressed in the yeast Saccharomyces cerevisiae (Kurtz et al., 1995). Based on this non-adamantane, a SAR study using a combination of viral plaque assays and two-electrode voltage clamp (TEVC) (Wang, Cady, et al., 2009) generated 3-azaspiro[5,5]undecane hydrochloride (Fig. 2B), which lacks the imidazoline group of BL-1743. This compound showed an IC50 of 0.92 ± 0.11 μM against AM2, ie, more than an order of magnitude more potent than amantadine (IC50 = 16 μM) and more than 45-fold increase in potency relative to the parental BL-1743.
Figure 2.
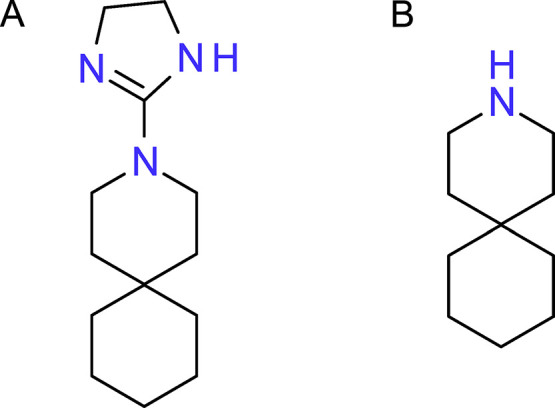
Example of IAV M2 inhibitors not based on the adamantane framework. (A) BL-1743; (B) spiro-piperidine developed from (A) (Wang, Cady, et al., 2009).
Compared to Amt, this spiro-piperidine induced a more homogeneous conformation of the AM2 peptide, reducing dynamic disorder near the water-filled central cavity of the helical bundle, suggesting a more extensive binding to the AM2 channel, thus leading to stronger inhibitory potency. However, this compound was only effective against the wild-type AM2. This is crucial because of the current prevalence of Amt-resistant variants of AM2, eg, L26F, V27A, and S31N, which comprise more than 99% of the reported resistance mutants (Furuse et al., 2009, Suzuki et al., 2003). Of these three mutations, V27A is the only one known to originate from drug selection pressure (Furuse et al., 2009), and this mutant is the only one among the three that is completely resistant to Amt and Rim (Balannik et al., 2009). On the other hand, S31N is found in more than 98% of the mutated M2 channels in the IAV subtype H3N2 viruses (Bright et al., 2005), and causes a 10–20 times decrease in the IC50, from low micromolar to 200 μM (Amt) and more than 2 mM (Rim) (Wang, Ma, et al., 2013).
The first non-adamantane inhibitor of the V27A mutant was a polycyclic pyrrolidine, a dual inhibitor of WT (IC50 of 3.4 μM) and V27A (IC50 of 0.29 μM) (Rey-Carrizo et al., 2013). Later, a compound with triple inhibitor efficacy was reported by the same authors, active against the WT, V27A, and L26F mutants, with IC50 of 18, 0.7, and 9 μM, respectively (Rey-Carrizo et al., 2014). The inhibitory activity of the compounds was tested on AM2 channels expressed in Xenopus laevis oocytes using the two-electrode voltage clamp (TEVC) technique, but most of the compounds were cytotoxic (more than Amt and Rim), as tested in cell-based antiviral plaque reduction assays. Therefore, despite the encouraging results obtained in vitro, the toxic properties of these compounds still hinder their therapeutic potential.
The spiro-piperidine inspired the generation of another compound (Fig. 3C) that could inhibit not just the WT AM2 (IC50 = 18 μM) but also mutants L26F (IC50 = 6 μM) and V27A (IC50 = 0.3 μM) (Wang, Ma, Fiorin, et al., 2011). Direct interaction of this compound with the V27A mutant was assessed by solid-state NMR, and inhibition was further demonstrated in plaque reduction assays. This compound, however, was inactive against the S31N mutant. Such observation was rationalized by the fact that the S31N mutant is more dynamic and hydrated in the pore than the corresponding WT protein (Pielak et al., 2009), as shown by NMR studies. This lead to a disruption in the size and polarity of the pore in precisely the region that accommodates the adamantane group, thereby hampering the discovery of inhibitors of S31N. This led to attempts to increase the polarity or the dimensions of the adamantane core in order to capture more interactions with the backbone of the channel.
Figure 3.

Inhibitors for IAV M2. (A) Amantadine (Amt); (B) rimantadine (Rim); (C) compound 9 in Wang, Ma, Fiorin, et al. (2011); (D) benzenediol (Wang, Ma, et al., 2013); (E and F) compounds with IC50 of ~ 16 μM (Wang, Wu, et al., 2013); (G) compound found in Wu et al. (2014).
A variety of scaffolds have been used to substitute for the hydrophobic adamantane group (Hu et al., 2010, Wang, Ma, et al., 2011a). These compounds showed excellent activity against the WT AM2, and some were highly active against V27A and L26F (Wang, Ma, Fiorin, et al., 2011). However, none was better than Amt against S31N. Attempts to introduce additional groups to the amine to enhance affinity for both S31N and WT via additional electrostatic interactions led to the discovery of benzyl-substituted amantadine derivatives (Fig. 3D) as dual WT and S31N inhibitors (Wang, Ma, et al., 2013) with IC50 of 35 and 59 μM for S31N and WT, respectively.
The DeGrado group later discovered small-molecule drugs, eg, M2WJ332, that locked the dynamic S31N mutant into a well-defined conformation, enabling high-resolution structure determination by solution NMR that showed the drug bound in the homotetrameric channel, threaded between the side chains of Asn-31. These compounds were formed by conjugation of a-CH2-heteroaryl group to the amine of Amt (Fig. 3E and F, IC50 of 16 μM) and inhibited S31N with potencies greater than Amt against AM2 (Wang, Wu, et al., 2013). The charged ammonium binds as a hydrate to one of three sites aligned along the central cavity that might stabilize hydronium-like species formed as protons diffuse through the outer channel to the proton-shuttling residue His-37 near the cytoplasmic end of the channel.
Based on structural and molecular dynamics data, the DeGrado group realized that inhibitors that are active against both WT and S31N channels bind in opposite orientations: the aromatic headgroup faces toward the C-terminus in WT, and toward the N-terminus in S31N. Investigation of substitutions led to the identification of halide-substituted thiophene compounds. The most potent of which, N-[(5-bromothiophen-2-yl)methyl]adamantan-1-amine (Fig. 3G), inhibited both WT and S31N mutant with comparable affinities as Amt against WT (Wu et al., 2014).
It would be desirable to develop the same paradigm found in IAV AM2 and their inhibitors for other viroporins, so that their channel inhibitors can be validated. For example, many amantadine-resistant influenza viruses can be selected in cell culture (Grambas et al., 1992, Grambas and Hay, 1992, Hay et al., 1985), and some of these are also found in infected patients undergoing treatment with amantadine (Shiraishi et al., 2003). In turn, engineered viruses harboring these pore-lining mutations, while competent to replicate in mouse (Abed, Goyette, & Boivin, 2005), give rise to attenuated viruses that tend to revert to the WT in the absence of drug pressure (Grambas and Hay, 1992, Suzuki et al., 2003).
3. The Hepatitis C Virus p7 Protein (HCV-p7)
Another very important viroporin is p7, found in the Hepatitis C virus (HCV), a member of the Flaviviridae family that has six genotypes (GTs). HCV is a positive-strand RNA virus that causes severe liver disease and is the major cause of hepatocellular carcinomas in the developed world. HCV has chronically infected 170 million people worldwide and is responsible for more than 300,000 yearly deaths, while no prophylactic or therapeutic vaccine is available.
HCV infection can be cured with drugs targeting viral enzymes and proteins, as well as host cellular proteins. One host-targeted antiviral agent (HTAs) is the type I IFN-α (Hoofnagle et al., 1986), which exerts antiviral activities against both RNA and DNA viruses through IFN-stimulated gene (ISG) products. Pegylated IFN has been used in combination with ribavirin (RBV), a synthetic guanosine analogue (Herrmann, Lee, Marinos, Modi, & Zeuzem, 2003), but this antiviral therapy can only last up to 1 year, associated with side effects, and is not always effective (Manns et al., 2006, Pawlotsky et al., 2007). Other drugs target HCV proteins with high potency, such as those against the HCV NS3/4A protease, the NS5A protein or the NS5B RNA-dependent RNA polymerase (RdRP) (Sulkowski et al., 2014) [see Pawlotsky, 2014, Schmidt et al., 2014, for recent reviews]. However, drug resistance creates a burden for patients, worsened by the rapid turnover of HCV replication, which produces up to 1012 virions daily (Neumann et al., 1998). This is compounded by the error-prone activity of the HCV RNA polymerase, which leads to the high genetic diversity of HCV and the formation of “quasi-species.” In this complicated context, the clinical niche of p7 inhibitors is unclear.
The viroporin HCV p7 is encoded within a single polyprotein precursor (Simmonds, 2013) processed by host and viral proteases (Moradpour & Penin, 2013). It separates the structural proteins (core and envelope glycoproteins E1 and E2) from the nonstructural proteins NS2, NS3, NS4A, NS4B, NS5A, and NS5B (Lindenbach & Rice, 2005). p7 results from cleavage of E2-p7-NS2 and E2-p7 by a signal peptidase at the ER (Lin et al., 1994, Mizushima et al., 1994). The p7 protein is 63 residues long with two TM domains, found mainly at the ER membrane. Although p7 is not necessary for RNA replication (Lohmann et al., 1999), it is essential for productive HCV propagation in vivo (Sakai et al., 2003). However, the precise role of the p7 in virus production was only determined when it was possible to replicate a genotype 2a isolate from a Japanese patient suffering from fulminant hepatitis (designated JFH-1) (Kato et al., 2003, Wakita et al., 2005). This isolate efficiently replicated in human hepatoma cell lines and produced infectious virus particles. This was used to demonstrate that p7 is essential for virus particle assembly and release (Jones et al., 2007, Steinmann et al., 2007). The bovine viral diarrhea virus (BVDV) (Harada, Tautz, & Thiel, 2000) and the hepacivirus GB virus B (GBV-B) (Takikawa et al., 2006), HCV's closest relatives, also have a p7 protein crucial for virus replication.
The channel activity of p7 was examined more than 10 years ago in experiments involving artificial lipid bilayers. It was suggested that p7 has preference for cations vs anions, with permeability ratios in the range 7–11 (Griffin et al., 2003, Montserret et al., 2010, Premkumar et al., 2004, Pavlovic et al., 2003) and more recently in Xenopus oocytes (Ouyang et al., 2013). This selectivity is rather low compared to other channels, eg, for cystic fibrosis transmembrane conductance regulator (CFTR) Cl− channels, selectivity for anions over cations is 10–30 (Anderson et al., 1991, Bear et al., 1992). For potassium channels, which conduct K+ ions near the diffusion limit, the selectivity for K+ over Na+ is about 10,000–1 (Doyle et al., 1998), and in aquaporins water to ion flux ratio is about 109 (Pohl, 2004). However, the biological importance of p7 cation channel activity may be relative, in the light of its recently found role to permeabilize membranes to protons, and acting to prevent acidification of intracellular vesicles (Wozniak et al., 2010). This proton channel activity was found to be crucial for the production of infectious viruses, and was observed in intracellular vesicles harboring p7. Recently, this proton channel activity has been directly confirmed in vitro using purified nontagged p7 protein in a liposome-based assay (Gan, Surya, Vararattanavech, & Torres, 2014), where the lipid composition used was the mixture PAESC (PA/PE/PS/PC 5:2:2:1 w/w) (Gervais et al., 2011).
Several initial papers also reported the inhibition of channel activity by some compounds. For example, His-tagged p7 from genotype 1b was inhibited by 1 μM amantadine (Griffin et al., 2003), whereas synthetic p7 from genotype 1a (strain H77) was inhibited by 100 μM hexamethyleneamiloride (HMA, Fig. 4A), although this peptide was largely unpurified (Premkumar et al., 2004). Another synthetic p7, also unpurified, was inhibited by 50–100 μM of the long-alkyl-chain imino sugar derivatives NN-DGJ, NN-DNJ (Fig. 4B) and N-7-oxanonyl-6-deoxy-DGJ (Pavlovic et al., 2003). However, the channel activity of purified synthetic p7 HCV-J genotype 1b in artificial lipid bilayers was inhibited by just 50% in the presence of 200 μM HMA, whereas Amt did not show any effect even at 1 mM concentration (Montserret et al., 2010). In fact, the efficacy of Amt in HCV culture systems is low (Steinmann et al., 2007), and no clinical benefit of Amt-based combination therapy was found in comparison to standard of care (peg-IFNα plus ribavirin) (Castelain et al., 2007, Mihm et al., 2006). Some of these compounds have been studied in infected cells, where they reduced virus production up to 10-fold in a genotype-dependent manner (Foster et al., 2014, Gottwein et al., 2011, Steinmann et al., 2007), reducing secretion of HCV particles by inhibition of the acidification of virus-containing compartments (Wozniak et al., 2010), without impairment of the infectivity of intracellular virions (Foster et al., 2011, Griffin et al., 2008).
Figure 4.

Some other inhibitors for viroporins reported in the literature. (A) HMA, (B) NN-DNJ, (C) BIT225, (D) pyronin-B.
To discover more p7 channel inhibitors, a liposome-based dye (carboxyfluorescein, CF) release assay has been proposed that involves the addition of p7 protein to preloaded liposomes (StGelais et al., 2007). The method has been claimed to be sensitive to inhibitors such as Amt and Rim, and proposed as a high-throughput functional assay to test drug sensitivity of p7 mutants, or to discover new p7 channel activity inhibitors (Foster et al., 2011, Gervais et al., 2011). These dye release assays have been previously performed with tagged p7 protein, either with FLAG (Li et al., 2012, StGelais et al., 2009) or flu-antigen (Foster et al., 2011). However, this method has been put into question since just the C-terminal half of p7, p7(27–63) was as efficient as full-length p7 in releasing CF, while a properly reconstituted α-helical sample of p7 was not able to elicit CF release (Gan et al., 2014).
Nevertheless, p7 inhibitors may be able to inhibit both the proton channel activity and dye release if the mechanisms are similar, eg, via an allosteric effect that increases the rigidity of the C-terminal half (TM2) of p7. This is supported by the fact that TM2 of p7 interacts with Amt and is the site of inhibition by adamantanes. Indeed, Amt interacts with leucines 51–57, and these leucines have been shown unequivocally to be part of a membrane inserted α-helix (Cook and Opella, 2009, Ouyang et al., 2013). Binding of Amt to p7 in dihexanoylphosphatidylcholine (DHPC) micelles led to resonance shifts in several residues, some of which are the leucines 51–57 (but not 53), ie, located in TM2 (Cook & Opella, 2009). Consistent with this, Leu to Ala mutations at this region produced an Amt-resistant p7 mutant revealed using the CF release assay (StGelais et al., 2009).
The imino sugar NN-DNJ and Rim inhibited secreted infectivity in Amt-resistant JFH-1 and CON-1/JFH-1c3 viruses, whereas mutation L20F conferred resistance to rimantadine (Foster et al., 2011), consistent with Rim resistance was found in GT1b patients not responding to the triple therapy IFN/Rib/Amt (Mihm et al., 2006). Using an oligomerization assay in DHPC gels, the GT1b J4 p7 oligomerization was abolished in presence of imino-sugars, whereas Rim did not affect oligomerization, suggesting two different mechanisms of action for these two compounds (Foster et al., 2011). The p7 of GT3a, but not GT1b, was found to be resistant to NN-DNJ both in vitro and in culture, and this was attributed to a F25A mutation (Foster et al., 2011, Griffin et al., 2008). However, direct interaction between HMA or imino sugar derivatives with p7 reconstituted in a detergent or membrane has not been reported. Addition of Rim to GT1b J4 p7 monomer in methanol (Foster et al., 2014) showed disruption at several residues, but not especially at Leu-20. Thus, this observation is either inconsistent with the proposed role of L20F in acquired resistance, or the methanol environment is not a good system to test this interaction.
Structural data for p7 was obtained initially in solvents and micelles by solution NMR. For example, solution NMR data was obtained for synthetic p7 (C27A mutant) in 50% of the helix inducer trifluoroethanol (TFE) (Montserret et al., 2010), and for recombinant p7 (C27S mutant) in DHPC micelles (Cook & Opella, 2010). The prevalent model for the p7 monomer according to these studies was that of an α-helical hairpin with two α-helical TMs kinked in the middle (Cook et al., 2010, Montserret et al., 2010), where the N-terminal TM helix (TM1) would face the lumen of a channel (Carrere-Kremer et al., 2002, Chew et al., 2009, Patargias et al., 2006) formed by either six or seven monomers (Clarke et al., 2006, Griffin et al., 2003, Luik et al., 2009, Montserret et al., 2010).
In a more recently published structural model of p7 (Ouyang et al., 2013), the oligomeric size is still ambiguous since this data could not be obtained in the same environment used in electron microscopy. From that model, the “old” TM1 and TM2 domains would correspond to three helical segments H1–H3, ie, H1 and first half of H2 (TM1) and second half of H2 and H3 (TM2). There are concerns, however, that this model can be present in a physiological context since the folding implied is difficult to explain by our current understanding of protein folding in the cell (Madan & Bartenschlager, 2015). In the Ouyang et al. paper, the authors systematically tested p7 amino acid sequences from various HCV genotypes and found that the sequence from GT5a (EUH1480 strain) generated samples adequate for structure determination. Although the oligomeric size of p7 in dodecyl phosphocholine (DPC) micelles was not determined, this was combined with data from negative stain electron microscopy (EM), which showed hexameric, flower-shaped particles. The latter were similar to those p7 hexamers (of GT2aJFH-1) observed in EM in the presence of DHPC micelles used earlier for single-particle reconstruction (Luik et al., 2009). The NMR-based model showed that the C-terminal helix, p7(27–63), is not the pore-lining sequence but instead forms a “lipid facing” part of the molecule, whereas the N-terminal half would orient lining the lumen (Ouyang et al., 2013). This is in agreement with previous models based on the partial inhibition of p7 channel activity by Cu2 +, but not by Mg2 +, which suggested exposure to a His residue located in the N-terminal half to the lumen of the channel (Chew et al., 2009). Although this interpretation may be correct, it is unfortunate that synthetic peptides corresponding to the N-terminal half “TM1,” eg, p7(1–34) or the C-terminal half “TM2,” eg, p7(35–63) aggregate easily, such that individual channel activity of these peptides cannot be properly measured (Montserret et al., 2010). In fact, neither of these helices separately forms α-helical structure in micelles or membranes, and peptide p7(1–26) was totally insoluble in TFE or in detergent (Gan et al., 2014), a difficulty also encountered for the N-terminal fragment of p7(1–34) (Montserret et al., 2010). In contrast, TM domains of other viroporins are completely α-helical, eg, the IAV M2 (Torres, Kukol, & Arkin, 2000), severe acute respiratory syndrome coronavirus (SARS-CoV) E (Torres et al., 2006), or respiratory syncytial virus (RSV) SH (Gan et al., 2012) produce sharp amide I bands in the infrared spectrum consistent with ~ 100% α-helix.
Nevertheless, the Amt binding site in the p7 channel was determined by identifying Nuclear Overhauser Enhancements (NOEs) between the protein backbone amide protons and drug protons (Ouyang et al., 2013) (Fig. 5 ). Although the full-scale structure determination of the complex could not be achieved, Amt (10 mM) showed NOE cross-peaks between the adamantane protons and the amide protons of Val-26, Leu-55, Leu-56, and Arg-57, ie, between the pore-forming and peripheral helices, suggesting a pocket formed by Leu-52, Val-53, and Leu-56 from H3, and Phe-20, Val-25, and Val-26 from H2, with the polar amino group of Amt pointing to the channel lumen, with similar results also obtained for Rim. Amt and Rim have also been reported previously to interact with some residues in the N-terminal half (Cook and Opella, 2009, StGelais et al., 2009). Isothermal titration calorimetry and NMR chemical shift perturbation analyses of p7(GT5a)–Rim interaction produced a binding constant (Kd) from 50 to 100 μM at 3 mM detergent concentration. The authors proposed that large differences in drug efficacies observed between different HCV genotypes are probably due to variations in the hydrophobicity of the binding pocket among p7 variants, and that the adamantane derivatives inhibit channel activity by restricting the structural rearrangement of the channel, ie, an allosteric mechanism.
Figure 5.

Interaction between HCV subtype 5a p7 and Amt. Left, Amt docked into the p7(5a) hexamer using restraints from NOEs contacts with backbone amide protons; right, a close-up view of Amt in the binding pocket.
Adapted by permission from Macmillan Publishers Ltd.: Ouyang, B., Xie, S., Berardi, M. J., Zhao, X., Dev, J., Yu, W., et al. (2013). Unusual architecture of the p7 channel from hepatitis C virus. Nature, 498(7455), 521–525. doi: 10.1038/nature12283. Copyright (2013).
Perhaps the most efficient effect for a p7 inhibitor has been reported for BIT225 (N-[5-(1-methyl-1H-pyrazol-4-yl)-napthalene-2-carbonyl]-guanidine) (Fig. 4C). This amiloride was identified in a bacterial assay that involved expression of p7 in E. coli; only in presence of inhibitors the normal ionic gradients are maintained and cells are able to grow despite the presence of p7. This compound had antiviral activity against the HCV model pestivirus bovine viral diarrhea virus (BVDV) with an IC50 of 314 nM. The addition of 100 μM of BIT225 blocked ion channel activity of synthetic HCV p7 protein of GT1a H77, although no data was reported in the context of HCV infection (Luscombe et al., 2010). While some docking efforts have been performed (Bichmann, Wang, & Fischer, 2014), direct interaction of BIT225 with p7 has not been proven using biophysical or structural assays, and therefore rational optimization of the compound is still not possible.
In addition to HCV p7, the human immunodeficiency virus (HIV) viroporin Vpu is also targeted; BIT225 was tested against the Vpu TM domain using a black lipid membrane (BLM) system, where the target was exposed to 40 μM of the drug (Khoury, Ewart, Luscombe, Miller, & Wilkinson, 2010). Although direct interaction with Vpu was not observed, this was assumed since the drug was not active against HIV-2, which has no Vpu gene.
One early clinical trial (phase Ib/IIa) involving the treatment of 24 naïve GT1 HCV patients indicated a significant reduction of viral load after 12 weeks treatment with BIT225 in combination with IFNα/ribavirin (www.biotron.com.au: biotron 2011). Currently, a phase 2a/2 clinical trial is in progress and the first results indicate activity against several HCV genotypes, including the difficult-to-treat GT3a and GT1a (www.biotron.com.au; May 29, 2015). Since it is also active against the HIV-1 Vpu, BIT225 might be suitable for treatment of HCV/HIV coinfected patients. In a Jul. 2015 update of a 3-month dosing trial with BIT225, there was a recommendation that future trials would focus on patients infected with HCV GT3 and to further study BIT225 in combination with other direct-acting antiviral drugs. In these trials, patients received 400 mg of BIT225 twice daily for 3 months. A phase 2a trial on HCV demonstrated that 100% of the patients infected with HCV GT1 who received BIT225 (400 mg) in combination with current standard of care therapies, IFN/RBV, had undetectable virus after 48 weeks. A phase 2 trial in HIV/HCV coinfected patients showed that all HCV GT3 patients completing 28 days of treatment with BIT225 in combination with IFN/RBV have undetectable HCV load for 12 weeks (sustained viral response, SVR12) after completing all therapy. BIT225 is also in development for treatment of HIV, and is the pioneer in a new class of antiviral drugs that may provide a new approach to the eradication of this virus. It has shown clinical efficacy against HIV in reservoir cells, and has the potential to be combined with new or existing antiretroviral drugs to eradicate long-lived pools of virus that are not successfully eliminated with current treatments. Despite these promising results, however, it remains to be demonstrated that BIT225 targets p7 or Vpu in infected cells.
4. The Coronavirus Envelope Protein (CoV E)
The most studied viroporin in coronaviruses is the envelope (E) protein. Coronaviruses (CoV) typically affect the respiratory tract and gut of mammals and birds. CoVs belong to the subfamily Coronavirinae in the family Coronaviridae, and are organized into four genera (Enjuanes et al., 2000): α, β, γ, and δ. Approximately 30% of common colds are caused by two human coronaviruses—OC43 and 229E. Of particular medical interest is the virus responsible for the severe acute respiratory syndrome (SARS), which produced a near pandemic in 2003 (Holmes, 2003), and the recently emerged Middle East respiratory syndrome coronavirus (MERS-CoV), which after 3 years has caused hundreds of deaths (Raj, Osterhaus, Fouchier, & Haagmans, 2014). Other coronavirus viroporins, eg, the SARS-CoV3a (Lu et al., 2006), a 274-amino acid protein with three putative TM domains, are also attracting increasing interest (Chien et al., 2013, Hsu and Fischer, 2012), but no inhibitors or structural data is available so far.
Currently, no effective licensed treatment exist against coronavirus infection (Kilianski and Baker, 2014, Kilianski et al., 2013, Lou et al., 2014), although live vaccines consisting of attenuated viruses is a promising strategy (Enjuanes et al., 2011, Graham et al., 2012), along with fusion inhibitors (reviewed in Heald-Sargent & Gallagher, 2012). Even so, the possibility of reappearance of virulent phenotypes, drug side effects, and resistance calls for continued antiviral development.
The envelope (E) proteins are 76–109 residues long with one TM domain (Li, Surya, et al., 2014, Parthasarathy et al., 2008, Parthasarathy et al., 2012, Pervushin et al., 2009, Torres et al., 2006, Torres et al., 2005), and most of them have a cytoplasmic C-terminal domain and a lumenal N-terminus (Corse and Machamer, 2000, Nieto-Torres et al., 2011, Raamsman et al., 2000, Ruch and Machamer, 2012).
Most CoV E proteins are present at low concentrations in virions (Corse and Machamer, 2000, Godet et al., 1992, Liao et al., 2006, Yu et al., 1994), but found abundantly in internal membranes of infected hosts (Corse and Machamer, 2003, Liao et al., 2006, Raamsman et al., 2000, Tung et al., 1992), eg, the ER–Golgi intermediate compartment (ERGIC), where virions assemble (Lopez et al., 2008, Nieto-Torres et al., 2011).
CoV E proteins have been found to be critical for viral pathogenesis, while having a protective antiapoptotic effect on infected cells (DeDiego et al., 2011) which may help the virus evade premature death of host cells, thereby allowing viral replication. Interestingly, deletion of E protein reduced pathogenicity and mortality in animal models through a reduced inflammation (DeDiego et al., 2014)—as discussed later—and this has led to the development of live attenuated vaccines based on E-deleted or E-truncated virions (Almazán et al., 2013, Lamirande et al., 2008, Netland et al., 2010).
The only structural data available for a CoV E protein is for SARS-CoV E, where the TM domain (E-TM) has been characterized in some detail in lipid membranes (Torres et al., 2006). Later, solution NMR of the selectively labelled synthetic E-TM (residues 8–38) in DPC micelles produced a similar pentameric left-handed parallel bundle (Pervushin et al., 2009). In these models, Asn-15 is facing the lumen of the channel (Torres et al., 2006) whereas Val-25 is involved in helix–helix interactions with other subunits (Pervushin et al., 2009). Mutations at these residues, N15A and V25F, abolished channel activity in vitro (Torres et al., 2007), and introduction of these mutations in a recombinant SARS-CoV resulted in in vivo attenuation in a mouse model. The integrity of the TM domain, and preservation of channel activity, were shown to be important for inflammasome activation and elevated production of the proinflammatory cytokine IL-1β (Nieto-Torres et al., 2014). In the same study, revertant mutants of the channel-inactive E that regained fitness and pathogenicity also regained channel activity, as measured in black lipid membranes. Both E-TM and the full-length E protein form homopentameric channels with poor ion selectivity that can be partially inhibited by the drug HMA with micromolar affinity (Verdia-Baguena et al., 2012, Wilson, Gage and Ewart, 2006).
The NOEs between HMA and E-TM suggested the presence of two binding sites at both ends of the TM domain. At the N-terminal domain, Leu-19 exhibited the largest chemical shift, and the relative intensities of the HMA proton cross-peaks indicated an HMA:E-TM stoichiometry of 1:5 at the N-terminal binding site near Asn-15, suggesting one HMA molecule per E-TM pentamer. This is consistent with a pore-blocking inhibition mode similar to Amt in AM2, where in this case the HMA molecule may be stabilized by a hydrogen bonding network to the Asn-15 side chains of the SARS-CoV E, with the cyclohexamethylene ring pointing away from the center of the channel (Fig. 6A). At the C-terminal of the TM domain, near Arg-38, the HMA:E-TM stoichiometry was 1:2, suggesting for this site a rapid exchange, in the chemical shift timescale, between E-TM-bound and micelle-bound forms of HMA. At this location, the amiloride group of HMA is likely to be involved in interactions with the guanidinium groups of Arg-38 (Fig. 6B). These results were confirmed using the extended peptide E (8–65) in mixed SDS/DPC (1:4 molar ratio) micelles (Li, Surya, et al., 2014), where the addition of HMA perturbed residues clustered near the N-terminal end of the TM domain, eg, Glu-8, Gly-10, Thr-11, Val-14, Asn-15, and Ser-16. At the C-terminal end of the TM, Leu-37 was the most affected, suggesting that the interaction of HMA at the Hɛ of Arg-38 reported previously may have been an artifact due to the use of an E-TM (8–38) peptide. In any case, these results suggest a similar behaviour of HMA-to-E protein and Amt-to-AM2 protein, ie, the presence of two binding sites, one pore-binding and one surface-binding in the TM domain. This structure provided a possible rationale for inhibition and a platform for future structure-based drug design of this potential pharmacological target.
Figure 6.

Binding of HMA to the SARS-CoV E-TM pentameric channel (Pervushin et al., 2009). (A) Side view of the binding of HMA to the vicinity of Asn-15. The side chains of amino acids interacting with HMA are shown using a stick representation; (B) binding of HMA to the C-terminal binding site of the channel, in the vicinity of Thr-35 and Arg-38. For clarity, one of the E-TM monomers has been removed.
5. The Respiratory Syncytial Virus Small Hydrophobic Protein (RSV SH)
The small hydrophobic (SH) protein is found in the human respiratory syncytial virus (hRSV), an enveloped pneumovirus in the Paramyxoviridae family. hRSV is the leading cause of bronchiolitis and pneumonia in infants and elderly (Dowell et al., 1996), and the most frequent cause of hospitalization of infants and young children in industrialized countries. In developing countries, hRSV is a significant cause of death, with global estimates of more than 70,000 deaths in young children. hRSV is the third most important cause of deadly childhood pneumonia after Streptococcus pneumoniae and Haemophilus influenzae (Nair et al., 2010).
Several compounds target the RSV fusion (F) protein, which facilitates viral entry through the host cell membrane (Zhao, Singh, Malashkevich, & Kim, 2000) through formation of a 6-helix bundle (Douglas et al., 2005, Lambert et al., 1996, Pastey et al., 2000, Razinkov et al., 2001, Roymans et al., 2010, Shepherd et al., 2006). Other approaches have involved gene transfer to expose viral proteins to cells (Kumar et al., 2002), or siRNA against specific viral proteins (Zhang et al., 2005). Vaccines have been recently designed based on a stabilized RSV-F form which preserves a highly antigenic site in its prefusion state, yielding RSV-specific neutralizing antibodies in mice and macaques (McLellan, Chen, et al., 2013a, McLellan, Chen, et al., 2013b). Recently, a vaccine candidate based on the extracellular domain (C-terminal) of the RSV viroporin, the small hydrophobic (SH) protein, has been reported (Schepens et al., 2014). In addition, the prevention of nasopulmonary infection in mice caused by RSV has been reported using stapled peptides targeting the fusogenic F-protein 6-helix bundle (Bird et al., 2014). However, despite all these efforts, new FDA-approved drugs have yet to emerge. The only licensed drug for use in infected individuals is ribavirin, a nucleoside analogue, but its efficacy is very limited (Hall et al., 1986).
The hRSV genome transcribes 11 proteins (Collins & Melero, 2011). One of these is the viroporin SH protein, which is 64–65 residues long (depending on subgroup, A or B) and has a single α-helical TM domain with N- and C-terminal extramembrane domains oriented cytoplasmically and lumenally/extracellularly, respectively (Collins and Mottet, 1993, Gan et al., 2012). Most SH protein accumulates at the membranes of the Golgi complex in infected cells, but it has also been detected in the ER and plasma membranes (Rixon et al., 2004).
RSV that lacks SH (RSVΔSH) is still viable, and still forms syncytia (Bukreyev et al., 1997, Karron et al., 1997, Techaarpornkul et al., 2001), but is attenuated in vivo. In mouse, RSV ΔSH replicated 10-fold less efficiently in the upper respiratory tract (Bukreyev et al., 1997), whereas chimpanzees developed significantly less rhinorrhea than those infected with wild-type RSV (Whitehead et al., 1999). Other reports have shown that the lack of SH protein leads to an attenuated phenotype in children and in rats (Jin et al., 2000, Karron et al., 1997). Overall, these results indicate involvement of hRSV SH protein in replication and pathogenesis.
In common to the SARS-CoV E protein, SH protein blocks or delays apoptosis in infected cells (Fuentes, Tran, Luthra, Teng, & He, 2007), and a similar antiapoptotic effect of SH protein has been observed for other members of the Paramyxoviridae family, eg, J Paramyxovirus (JPV) (Jack et al., 2005, Li et al., 2011), mumps virus (MuV), and the parainfluenza virus 5 (PIV5), formerly known as simian virus 5 (SV5). In all these systems, the SH protein seems to block apoptosis during infection—at least in part—through inhibition of the TNF-α pathway (Fuentes et al., 2007, Li et al., 2011, Lin et al., 2003, Wilson, Fuentes, et al., 2006).
The mutual orientation of the TM α-helices that form the ion channel was determined in lipid bilayers using site-specific infrared dichroism (Gan, Ng, Xin, Gong, & Torres, 2008). A description of the full-length RSV SH protein monomer has been obtained by solution NMR in DPC micelles (Gan et al., 2012) and later in bicelles, only for the extramembrane domains (Li, To, et al., 2014). Like SARS-CoV E, the RSV SH protein forms homopentameric channels (Gan et al., 2008, Gan et al., 2012) that have low ion selectivity (Li, To, et al., 2014). The TM domain of SH protein has a funnel-like architecture (Gan et al., 2012), as observed in other viroporins, eg, the IAV M2 (Schnell & Chou, 2008), SARS-CoV E (Pervushin et al., 2009) and HCV p7 (Ouyang et al., 2013). A narrower region (Gan et al., 2012) in the TM domain is lined with hydrophobic side chains (Ile-32, Ile-36, Ile-40, and Leu-44) whereas the more open N-terminal region is lined by polar residues, ie, His-22, Thr-25, and Ser-29. The TM α-helix extends up to His-51 in the C-terminal region, followed by a loop, whereas the N-terminal cytoplasmic extramembrane domain forms a short α-helix (residues 5–14) present both in micelles and in bicelles (Li, To, et al., 2014).
Only one compound, pyronin-B, has been reported to inhibit the channel activity of the SH protein. This compound was obtained from liposome-based fluorescence assay, and was tested against purified SH protein reconstituted in planar lipid bilayers (BLM). A concentration of 10 μM led to a ~ 60% inhibition of channel activity, with a Kd of ~ 7 μM. The effect of pyronin-B was tested against RSV replication in Vero cells, where the tissue culture infectivity (50% infective dose, TCID50) was zero at a drug concentration of 0.25 μM.
In the NMR studies, the binding of pyronin-B to the SH protein revealed backbone resonance perturbations at both ends of the TM domain. At the N-terminal cytoplasmic end, residues more affected were Ile-6 and Ile-21. At the lumenal C-terminal end, the residue most affected was Ala-39, and a group of nearby residues, Ile-38, Ile-40, Leu-41, and Lys-43 (Fig. 7A). Interestingly, most of these juxtamembrane residues (residues 38–43) form a conserved motif in the SH protein “A39ILNKL43,” which suggests that this is a critical region for SH protein with a high barrier to resistance. Determination of intermolecular nuclear Overhauser effects (NOEs) for the drug–protein complex was not possible, probably due to weak interaction.
Figure 7.

Mapping of pyronin-B binding to the RSV SH protein. (A) Chemical shift perturbation (CSP) values in presence of pyronin-B, mapped onto the structure of SH protein; larger (CSP ≥ 0.07 ppm) and smaller (CSP ≥ 0.04 ppm) shifts in red (dark gray in the print version) and yellow (light gray in the print version), respectively; (B and C) two predicted pyronin-B binding sites on the SH pentamer, at the N-terminal (B) and C-terminal (C) ends of the TM domain (dotted circles). Only two monomers of SH protein pentamer (i and i + 1) are shown for simplicity; (D) druggable pockets (green (dark gray in the print version) mesh) predicted by DoGSiteScorer (Volkamer, Kuhn, Rippmann, & Rarey, 2012). The main residues that showed largest NMR chemical shift changes are shown in red (dark gray in the print version). Two monomers have been removed from the pentamer for clarity.
Docking studies of pyronin-B to the SH pentameric model obtained in DPC micelles (Gan et al., 2012) produced two predicted binding sites at both ends of the TM domain. Indeed, a majority of structures showed the drug located near residues with largest chemical shift perturbation, ie, Ile-6, Ile-21, and Ala-39 (Fig. 7). The binding site near the N-terminus is formed by Phe-14, Ile-21 in one monomer (i + 1), and Ile-6 of the previous (i) monomer. The one near the C-terminus is formed by residues Ile-40, Leu-41, and Lys-43 in one monomer (i + 1) and Ile-38 and Ala-39 of the previous (i) monomer (Fig. 7A–C). Similar “druggable pockets” on the SH pentamer surface were identified using DoGSiteScorer (Volkamer et al., 2012) (Fig. 7D). However, the C-terminal end of the TM domain (residues 38–41) is sufficient for inhibition, since a conservative mutation A39S almost completely prevented inhibition (~ 10% inhibition).
6. The Human Immunodeficiency Virus Viral Protein U (HIV-1-Vpu)
Vpu is a 81-residue small-membrane protein encoded by the single-stranded positive-sense RNA human immunodeficiency virus 1 (HIV-1), and some simian immunodeficiency virus (SIV) isolates. Vpu is one of four HIV-1 accessory proteins involved in the release of new virions from host cell membranes (Cohen et al., 1988, Strebel et al., 1988, Terwilliger et al., 1989). Without Vpu, HIV particles fail to detach from the plasma membrane and accumulate in large numbers in the cell surface (Klimkait, Strebel, Hoggan, Martin, & Orenstein, 1990).
The initial structural studies of Vpu were performed more than 20 years ago by solution NMR in TFE/water using synthetic peptides of various overlapping lengths, which led to a model of an N-terminal hydrophobic transmembrane helix (the TM domain) and a C-terminal cytoplasmic domain with a helix–loop–helix arrangement [reviewed recently in Opella, 2015]. Structural similarity of Vpu with AM2 suggested that Vpu could form pores (Maldarelli, Chen, Willey, & Strebel, 1993) and was later shown to have cation-selective channel activity (Ewart et al., 1996, Schubert et al., 1996), but a unique structural model for the Vpu oligomer cannot be defined [reviewed in Radoicic, Lu, & Opella, 2014]. An analysis of the Vpu TM (residues 1–40) by solid-state NMR in membranes, combined with analytical centrifugation measurements in detergent, produced data compatible with a variety of oligomers coexisting in micelles and phospholipid bilayers (Lu, Sharpe, Ghirlando, Yau, & Tycko, 2010).
This led to the proposal that regulation of virus release by Vpu might involve an alteration of the plasma membrane potential, and this was supported by (i) the reduced rate of HIV-1 virus release after membrane depolarization (Hsu, Han, Shinlapawittayatorn, Deschenes, & Marbán, 2010), (ii) the observed reduction in viral load and viral pathogenicity when Vpu TM domain is scrambled in the context of a simian–human immunodeficiency virus (SHIV) (Hout et al., 2005), and (iii) the substitution of Ala-18 by a histidine (A18H) rendered HIV-1 infections susceptible to Rim (Hout et al., 2006). However, recent studies challenge this model and argue against a critical involvement of Vpu ion channel activity in Vpu-enhanced virus release. Indeed, mutations in the Vpu TM domain that affect channel activity do not impair the ability to enhance virus release, whereas mutations that affect virus release retain ion channel activity (Bolduan et al., 2011). Thus, the functional significance of Vpu ion channel activity for HIV replication, and the role as a target for channel inhibitors, remains unclear.
7. 6K, Agnoprotein, and 2B
There are several other viroporins of which only very limited structural data is available, and no channel inhibitors have been reported. The following section will briefly describe three of such viroporins: the alphavirus 6K protein, polyomavirus agnoprotein, and the 2B protein found in picornaviruses.
7.1. The Alphavirus 6K Protein
The 6K protein is 58–61 amino acids long and has one predicted α-helical TM domain. This viroporin is found in several members of the alphavirus genus in the positive-sense RNA family Togaviridae. This includes, for example, equine encephalitis viruses, Chikungunya virus, Sindbis virus, Semliki Forest viruses (SFV), Ross River virus (RRV), and Barmah Forest virus (BFV). In these viruses, structural proteins are translated in the ER from a subgenomic mRNA as a single polyprotein later cleaved by proteases, in the order capsid-PE2-6K-E1 (Liljestrom and Garoff, 1991, Strauss and Strauss, 1994).
The 6K protein was first described as early as 1980 (Welch & Sefton, 1980). Its expression in E. coli caused an increase in membrane permeability, whereas in eukaryotic cells it promoted viral budding (Sanz, Perez, & Carrasco, 1994). Further, 6K proteins of RRV and BFV formed cation-selective ion channels in planar lipid bilayers (Melton et al., 2002). This suggested a role for viroporins in alphavirus infection, since an increased permeability of cells to monovalent cations is followed by budding of virions.
Whatever the mechanism, 6K proteins have an important role in virus assembly and budding. For example, in Sindbis virus, cysteine-less mutants dramatically reduced the release of virus particles, leading to aberrant enveloped particles containing multiple nucleocapsids (Gaedigk-Nitschko, Ding, Levy, & Schlesinger, 1990). Other results are consistent with an indirect role in the packing of the virus spike glycoproteins; mutation C39S resulted in defective budding and a revertant contained two additional mutations at the ectodomain of the virus glycoprotein (Ivanova, Lustig, & Schlesinger, 1995). Further, deletion of 6K in a full-length cDNA clone of SFV resulted in a dramatic reduction in virus release (Liljestrom, Lusa, Huylebroeck, & Garoff, 1991), and BHK cells expressing a Δ6K strain produced thermolabile particles, suggesting a disrupted glycoprotein packing (Loewy, Smyth, von Bonsdorff, Liljestrom, & Schlesinger, 1995). Despite these investigations, the specific role of 6K channel activity in these processes is still uncertain.
7.2. The Polyomavirus Agnoprotein
Agnoprotein is a 71-amino acids long membrane protein with one predicted α-helical TM domain required for the productive replication of the human polyomavirus JCV (John Cunningham virus). JCV causes progressive multifocal leukoencephalopathy, a fatal demyelinating disease resulting from lytic infection of oligodendrocytes. Agnoproteinis also found in other polyomaviruses, including BK virus, and simian virus 40 (Sariyer, Saribas, White, & Safak, 2011). Agnoprotein was shown to play important regulatory roles in the JCV replication cycle (Ellis et al., 2012, Ellis et al., 2013, Saribas et al., 2013), and possesses properties commonly associated with viroporins (Suzuki et al., 2013, Suzuki, Orba, et al., 2010a, Suzuki, Orba, et al., 2010b). For example, a deletion mutant is defective in virion release and viral propagation. Also, agnoprotein localizes to the ER during early stages of infection, but is also found at the plasma membrane late in infection. Finally, agnoprotein, an integral membrane protein, forms homo-oligomers that enhance permeability of cells to hygromycin B and induce the influx of extracellular Ca2 +, leading to membrane dysfunction and enhancement of virus release. Only a solution NMR structure of a synthetic peptide corresponding to the TM domain (Thr-17 to Glu-55) is available, but with an uncertain oligomeric size in lipid bilayers (Coric et al., 2014). As with the 6K protein, characterization of this viroporin is still in its infancy.
7.3. The Picornavirus 2B Protein
Protein 2B is a ~ 100 residues long nonstructural protein found in enteroviruses [see Ao, Sun, & Guo, 2014 for a recent review], eg, poliovirus (Agirre, Barco, Carrasco, & Nieva, 2002). Enteroviruses belong to the single-stranded positive-sense RNA Picornaviridae family, and include poliovirus, Coxsackie virus and human enterovirus 71 (EV71). The single open reading frame encodes a peptide chain divided into three regions, P1 to P3. The 2B protein is one of the products from the polypeptides in the P2 region, and has two TM domains, one more hydrophobic than the other (de Jong et al., 2003, Nieva et al., 2003). The C- and N-termini of the poliovirus 2B protein are exposed to the ER and Golgi lumen when expressed in cells (de Jong et al., 2003), but the 2B protein in Coxsackie virus adopts an opposite orientation (Abed et al., 2005, van Kuppeveld et al., 1996). Poliovirus 2B protein was found to increase the concentrations of free calcium in the cytosol, which has been proposed to promote viral replication and repression of the antiviral immune response of host cells (Aldabe, Irurzun, & Carrasco, 1997). However, precise structural data is not yet available for this protein or its oligomers.
8. Host–Cell Responses Triggered by Viroporin Activity
One common, specific effect of viroporin channel activity may be associated with the observed ability of viruses to activate the host inflammasome, which activates caspase-1 to produce proinflammatory cytokines, eg, IL-1β. These effects have been shown in many cases to be caused by the disruption of cellular ion homeostasis by allowing ion leakage from intracellular organelles into the cytosol, through the expression of viroporins [reviewed in Guo, Jin, Zhi, Yan, & Sun, 2015]. For example, the IAV activates the NLRP3 inflammasome as a result of H+ or ion flux from Golgi mediated by its M2 ion channel (Ichinohe, Pang, & Iwasaki, 2010). The 2B protein from several picornaviruses, including the encephalomyocarditis virus (EMCV), poliovirus, enterovirus 71, and human rhinovirus (Triantafilou, Kar, van Kuppeveld, & Triantafilou, 2013) were shown to induce NLRP3 cytoplasmic relocalization and inflammasome activation in an intracellular Ca2 +-mediated manner (Ito, Yanagi, & Ichinohe, 2012). The channel activity of the SARS-CoV E protein is required for the processing of IL-1β (Nieto-Torres et al., 2014), which requires caspase-1 activation. Channel activity of the SH protein may also result directly or indirectly in activation of the NLRP3 inflammasome (Segovia et al., 2012, Triantafilou, Kar, et al., 2013a). In these cases, therefore, development of channel inhibitors should improve the pathological effects of host inflammation caused by these viruses.
9. Modulation of Protein–Protein Interactions Involving Viroporins
Apart from the effects of their channel activity, viroporins are involved in many protein–protein interactions (PPIs) that are susceptible for therapeutic intervention, and may be increasingly important once these interactions are properly characterized [see recent reviews Fischer et al., 2014, Fischer et al., 2012, Nieva and Carrasco, 2015].
For example, the cytoplasmic regions of AM2 and BM2 are important for proper virus assembly and interaction with M1 (Chen et al., 2008, Imai et al., 2008, Imai et al., 2004, McCown and Pekosz, 2006), which can be proposed as an attractive drug target.
Also, the HCV p7 interacts with other viral structural and nonstructural proteins that are important to promote virus assembly and release (Gentzsch et al., 2013, Haqshenas et al., 2007, Pietschmann et al., 2006, Sakai et al., 2003, Yi et al., 2007), and functions related to capsid assembly and localization of several viral proteins are not affected by either channel-inactivating mutations or treatment with Rim (Tedbury et al., 2011, Wozniak et al., 2010), eg, the transfer of HCV core protein from lipid droplets to the ER depends on the interaction between p7 and NS2 (Boson, Granio, Bartenschlager, & Cosset, 2011). Therefore, the disruption of such interactions would require the action of PPI modulators rather than channel inhibitors.
The interaction partners of E and SH proteins have been reviewed recently (Torres, Surya, Li, & Liu, 2015). In the case of CoV E proteins, the interaction with M protein has long been reported to contribute to M localization and virion formation (Corse and Machamer, 2003, Hogue and Machamer, 2008, Ilk et al., 2011, Lim and Liu, 2001, Siu et al., 2008, Vennema et al., 1996). Recent reports have highlighted interactions with Protein Associated with Lin Seven 1 (PALS1) (Teoh et al., 2010) and syntenin through PDZ domains of E proteins, which are consistent with alterations of lung epithelia integrity and inflammation observed during CoV infection. Disruption of these pathways may have clear therapeutic implications, since in SARS-CoV-infected patients it is an exacerbated inflammatory response that leads to epithelial and endothelial damage, edema, and acute respiratory distress syndrome (Smits et al., 2010). Noticeably, several other viruses, eg, human papillomavirus and influenza A, have been found to enhance pathogenesis through proteins containing PDZ binding motifs (reviewed in Javier & Rice, 2011), which suggests that this is a particular case of a widely used viral strategy.
Vpu is known for its ability to degrade the HIV-receptor CD4 (Bour & Strebel, 2003), which requires interaction between Vpu and CD4 cytoplasmic domains (McNatt et al., 2013, Singh et al., 2012) and probably TM domains (Do et al., 2013, Magadán and Bonifacino, 2012) of the two proteins. This may not even require formation of oligomers, and even less channel activity. Vpu also affects virus release via interaction with interferon-induced protein tetherin (BST-2) (Miyagi et al., 2009, Neil et al., 2008, Skasko et al., 2012, Van Damme et al., 2008), an interferon-induced host protein that inhibits the release of many enveloped viruses by tethering virions to the cell surface [reviewed in Strebel, 2014].
In another case, the interaction between RSV SH and G proteins has been reported previously in infected cells (Low et al., 2008, Rixon et al., 2005), although the significance is not clear yet. The B-cell receptor-associated protein 31 (BAP31) has been recently reported to interact with the SH protein (Li et al., 2015). Although this interaction has not been reported in infected cells, it could prevent the cleavage of BAP31 and the formation of proapoptotic fragment p20, thus delaying apoptosis of infected cells. Interaction with BAP31 was also observed in the E5 viroporin from the high-risk human papillomavirus HPV-16 and HPV-31, where it is similarly thought to regulate apoptosis (Regan & Laimins, 2008).
JCV agnoprotein has been shown to interact with a number of cellular and viral proteins (Gerits and Moens, 2012, Gerits et al., 2015). However, despite all these intense research, the exact regulatory function of agnoprotein in viral replication is not fully understood. JCV agnoprotein specifically interacts with the adaptor protein complex 3 through its delta subunit. This interaction interrupts adaptor protein complex 3-mediated vesicular trafficking. The protein is then not targeted to the lysosomal degradation pathway, while allowing transport of agnoprotein to the plasma membrane. The findings suggest that the viroporins of other viruses may also be highly regulated by specific interactions with host cell proteins (Suzuki et al., 2013).
These pathologically relevant interactions between viroporins and their associated viral and host proteins present an attractive therapeutic target for disruption by small molecules, as well as peptides stabilized by internal covalent linkage (stapled peptides) (Walensky & Bird, 2014). In principle, both the transmembrane and extramembrane domains of viroporins can be potential templates for peptide-based modulators. For example, one could design a stapled peptide mimicking the α-helical cytoplasmic domain of the SH protein to disrupt the interactions between SH and a variant of the death effector domain (vDED) of BAP31, which would lead to enhanced apoptosis and cell death of infected cells. One could also mimic the transmembrane domain to disrupt monomer–monomer interactions, as shown recently for Halobacterium salinarum where efflux by a small multidrug resistance protein was inhibited with peptides targeting its TM domain (Bellmann-Sickert, Stone, Poulsen, & Deber, 2014). In the case of viroporins, the disruption of the oligomeric status would abolish channel activity, a classical strategy revisited with a different approach. However, more detailed mechanistic information should be available before these conceptual strategies can be widely used.
Acknowledgments
This work has been funded by Singapore MOE Tier 1 grant RG 51/13 (J.T.).
References
- Abed Y., Goyette N., Boivin G. Generation and characterization of recombinant influenza A (H1N1) viruses harboring amantadine resistance mutations. Antimicrobial Agents and Chemotherapy. 2005;49(2):556–559. doi: 10.1128/AAC.49.2.556-559.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Acharya R., Carnevale V., Fiorin G., Levine B.G., Polishchuk A.L., Balannik V. Structure and mechanism of proton transport through the transmembrane tetrameric M2 protein bundle of the influenza A virus. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(34):15075–15080. doi: 10.1073/pnas.1007071107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agirre A., Barco A., Carrasco L., Nieva J.L. Viroporin-mediated membrane permeabilization. Pore formation by nonstructural poliovirus 2B protein. Journal of Biological Chemistry. 2002;277(43):40434–40441. doi: 10.1074/jbc.M205393200. [DOI] [PubMed] [Google Scholar]
- Aldabe R., Irurzun A., Carrasco L. Poliovirus protein 2BC increases cytosolic free calcium concentrations. Journal of Virology. 1997;71(8):6214–6217. doi: 10.1128/jvi.71.8.6214-6217.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almazán F., DeDiego M.L., Sola I., Zuñiga S., Nieto-Torres J.L., Marquez-Jurado S. Engineering a replication-competent, propagation-defective middle east respiratory syndrome coronavirus as a vaccine candidate. mBio. 2013;4(5):1–11. doi: 10.1128/mBio.00650-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson M.P., Gregory R.J., Thompson S., Souza D.W., Paul S., Mulligan R.C. Demonstration that cftr is a chloride channel by alteration of its anion selectivity. Science. 1991;253(5016):202–205. doi: 10.1126/science.1712984. [DOI] [PubMed] [Google Scholar]
- Anthony S.J., Epstein J.H., Murray K.A., Navarrete-Macias I., Zambrana-Torrelio C.M., Solovyov A. A strategy to estimate unknown viral diversity in mammals. mBio. 2013;4(5) doi: 10.1128/mBio.00598-13. e00598-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ao D., Sun S.Q., Guo H.C. Topology and biological function of enterovirus non-structural protein 2B as a member of the viroporin family. Veterinary Research. 2014;45:87. doi: 10.1186/s13567-014-0087-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balannik V., Wang J., Ohigashi Y., Jing X.H., Magavern E., Lamb R.A. Design and pharmacological characterization of inhibitors of amantadine-resistant mutants of the M2 Ion channel of influenza A virus. Biochemistry. 2009;48(50):11872–11882. doi: 10.1021/bi9014488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bear C.E., Li C.H., Kartner N., Bridges R.J., Jensen T.J., Ramjeesingh M., Riordan J.R. Purification and functional reconstitution of the cystic-fibrosis transmembrane conductance regulator (cftr) Cell. 1992;68(4):809–818. doi: 10.1016/0092-8674(92)90155-6. [DOI] [PubMed] [Google Scholar]
- Bellmann-Sickert K., Stone T.A., Poulsen B.E., Deber C.M. Efflux by small multidrug resistance proteins is inhibited by membrane-interactive helix-stapled peptides. The Journal of Biological Chemistry. 2014;290(3):1752–1759. doi: 10.1074/jbc.M114.616185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bichmann L., Wang Y.T., Fischer W.B. Docking assay of small molecule antivirals to p7 of HCV. Computational Biology and Chemistry. 2014;53(B):308–317. doi: 10.1016/j.compbiolchem.2014.11.001. [DOI] [PubMed] [Google Scholar]
- Bird G.H., Boyapalle S., Wong T., Opoku-Nsiah K., Bedi R., Crannell W.C. Mucosal delivery of a double-stapled RSV peptide prevents nasopulmonary infection. Journal of Clinical Investigation. 2014;124(5):2113–2124. doi: 10.1172/Jci71856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolduan S., Votteler J., Lodermeyer V., Greiner T., Koppensteiner H., Schindler M. Ion channel activity of HIV-1 Vpu is dispensable for counteraction of CD317. Virology. 2011;416(1–2):75–85. doi: 10.1016/j.virol.2011.04.009. [DOI] [PubMed] [Google Scholar]
- Boson B., Granio O., Bartenschlager R., Cosset F.L. A concerted action of hepatitis C virus P7 and nonstructural protein 2 regulates core localization at the endoplasmic reticulum and virus assembly. PLoS Pathogens. 2011;7(7) doi: 10.1371/journal.ppat.1002144. e1002144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bour S., Strebel K. The HIV-1 Vpu protein: A multifunctional enhancer of viral particle release. Microbes and Infection. 2003;5(11):1029–1039. doi: 10.1016/S1286-4579(03)00191-6. [DOI] [PubMed] [Google Scholar]
- Bright R.A., Medina M.J., Xu X., Perez-Oronoz G., Wallis T.R., Davis X.M. Incidence of adamantane resistance among influenza A (H3N2) viruses isolated worldwide from 1994 to 2005: A cause for concern. Lancet. 2005;366(9492):1175–1181. doi: 10.1016/S0140-6736(05)67338-2. [DOI] [PubMed] [Google Scholar]
- Bright R.A., Shay D.K., Shu B., Cox N.J., Klimov A.I. Adamantane resistance among influenza A viruses isolated early during the 2005–2006 influenza season in the United States. Journal of the American Medical Association. 2006;295(8):891–894. doi: 10.1001/jama.295.8.joc60020. [DOI] [PubMed] [Google Scholar]
- Bukreyev A., Whitehead S.S., Murphy B.R., Collins P.L. Recombinant respiratory syncytial virus from which the entire SH gene has been deleted grows efficiently in cell culture and exhibits site-specific attenuation in the respiratory tract of the mouse. Journal of Virology. 1997;71(12):8973–8982. doi: 10.1128/jvi.71.12.8973-8982.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cady S.D., Schmidt-Rohr K., Wang J., Soto C.S., Degrado W.F., Hong M. Structure of the amantadine binding site of influenza M2 proton channels in lipid bilayers. Nature. 2010;463(7281):689–692. doi: 10.1038/nature08722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cady S., Wang T., Hong M. Membrane-dependent effects of a cytoplasmic helix on the structure and drug binding of the influenza virus M2 protein. Journal of the American Chemical Society. 2011;133(30):11572–11579. doi: 10.1021/ja202051n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco L. Modification of membrane permeability by animal viruses. Advances in Virus Research. 1995;45:61–112. doi: 10.1016/S0065-3527(08)60058-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrere-Kremer S., Montpellier-Pala C., Cocquerel L., Wychowski C., Penin F., Dubuisson J. Subcellular localization and topology of the p7 polypeptide of hepatitis C virus. Journal of Virology. 2002;76(8):3720–3730. doi: 10.1128/JVI.76.8.3720-3730.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castelain S., Bonte D., Penin F., François C., Capron D., Dedeurwaerder S. Hepatitis C virus p7 membrane protein quasispecies variability in chronically infected patients treated with interferon and ribavirin, with or without amantadine. Journal of Medical Virology. 2007;79(2):144–154. doi: 10.1002/jmv.20772. [DOI] [PubMed] [Google Scholar]
- Chen B.J., Leser G.P., Jackson D., Lamb R.A. The influenza virus M2 protein cytoplasmic tail interacts with the M1 protein and influences virus assembly at the site of virus budding. Journal of Virology. 2008;82(20):10059–10070. doi: 10.1128/JVI.01184-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chew C.F., Vijayan R., Chang J., Zitzmann N., Biggin P.C. Determination of pore-lining residues in the hepatitis C virus p7 protein. Biophysical Journal. 2009;96(2):L10–L12. doi: 10.1016/j.bpj.2008.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chien T.H., Chiang Y.L., Chen C.P., Henklein P., Hanel K., Hwang I.S. Assembling an ion channel: ORF 3a from SARS-CoV. Biopolymers. 2013;99(9):628–635. doi: 10.1002/bip.22230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke D., Griffin S., Beales L., Gelais C.S., Burgess S., Harris M., Rowlands D. Evidence for the formation of a heptameric ion channel complex by the hepatitis C virus P7 protein in vitro. Journal of Biological Chemistry. 2006;281(48):37057–37068. doi: 10.1074/jbc.M602434200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen E.A., Terwilliger E.F., Sodroski J.G., Haseltine W.A. Identification of a protein encoded by the vpu gene of HIV-1. Nature. 1988;334(6182):532–534. doi: 10.1038/334532a0. [DOI] [PubMed] [Google Scholar]
- Collins P.L., Melero J.A. Progress in understanding and controlling respiratory syncytial virus: Still crazy after all these years. Virus Research. 2011;162(1–2):80–99. doi: 10.1016/j.virusres.2011.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins P.L., Mottet G. Membrane orientation and oligomerization of the small hydrophobic protein of human respiratory syncytial virus. Journal of General Virology. 1993;74:1445–1450. doi: 10.1099/0022-1317-74-7-1445. [DOI] [PubMed] [Google Scholar]
- Cook G.A., Opella S.J. NMR studies of p7 protein from hepatitis C virus. European Biophysics Journal. 2009;39(7) doi: 10.1007/s00249-009-0533-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cook G.A., Opella S.J. Secondary structure, dynamics, and architecture of the p7 membrane protein from hepatitis C virus by NMR spectroscopy. Biochimica et Biophysica Acta. 2010;1808:1448–1453. doi: 10.1016/j.bbamem.2010.08.010. S0005-2736(10)00291-9 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cook G.A., Zhang H., Park S.H., Wang Y., Opella S.J. Comparative NMR studies demonstrate profound differences between two viroporins: p7 of HCV and Vpu of HIV-1. Biochimica et Biophysica Acta. 2010;1808(2):554–560. doi: 10.1016/j.bbamem.2010.08.005. S0005-2736(10)00286-5 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coric P., Saribas A.S., Abou-Gharbia M., Childers W., White M.K., Bouaziz S., Safak M. Nuclear magnetic resonance structure revealed that the human polyomavirus JC virus agnoprotein contains an alpha-helix encompassing the Leu/Ile/Phe-rich domain. Journal of Virology. 2014;88(12):6556–6575. doi: 10.1128/Jvi.00146-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corse E., Machamer C.E. Infectious bronchitis virus E protein is targeted to the Golgi complex and directs release of virus-like particles. Journal of Virology. 2000;74(9):4319–4326. doi: 10.1128/jvi.74.9.4319-4326.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corse E., Machamer C.E. The cytoplasmic tails of infectious bronchitis virus E and M proteins mediate their interaction. Virology. 2003;312(1):25–34. doi: 10.1016/S0042-6822(03)00175-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cross T.A., Dong H., Sharma M., Busath D.D., Zhou H.X. M2 protein from influenza A: From multiple structures to biophysical and functional insights. Current Opinion in Virology. 2012;2(2):128–133. doi: 10.1016/j.coviro.2012.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cross T.A., Sharma M., Yi M., Zhou H.X. Influence of solubilizing environments on membrane protein structures. Trends in Biochemical Sciences. 2011;36(2):117–125. doi: 10.1016/j.tibs.2010.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuello L.G., Jogini V., Cortes D.M., Perozo E. Structural mechanism of C-type inactivation in K + channels. Nature. 2010;466(7303) doi: 10.1038/Nature09153. 203-U273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies W.L., Grunert R.R., Haff R.F., McGahen J.W., Neumayer E.M., Paulshock M. Antiviral activity of 1-adamantanamine (amantadine) Science. 1964;144(3620):862–863. doi: 10.1126/science.144.3620.862. [DOI] [PubMed] [Google Scholar]
- De Clercq E. Antiviral agents active against influenza A viruses. Nature Reviews. Drug Discovery. 2006;5(12):1015–1025. doi: 10.1038/nrd2175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Jong A.S., Wessels E., Dijkman H.B.P.M., Galama J.M.D., Melchers W.J.G., Willems P.H.G.M., van Kuppeveld F.J.M. Determinants for membrane association and permeabilization of the coxsackievirus 2B protein and the identification of the Golgi complex as the target organelle. Journal of Biological Chemistry. 2003;278(2):1012–1021. doi: 10.1074/jbc.M207745200. [DOI] [PubMed] [Google Scholar]
- DeDiego M.L., Nieto-Torres J.L., Jimenez-Guardeno J.M., Regla-Nava J.A., Alvarez E., Oliveros J.C. Severe acute respiratory syndrome coronavirus envelope protein regulates cell stress response and apoptosis. PLoS Pathogens. 2011;7(10) doi: 10.1371/journal.ppat.1002315. e1002315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeDiego M.L., Nieto-Torres J.L., Regla-Nava J.A., Jimenez-Guardeño J.M., Fernandez-Delgado R., Fett C. Inhibition of NF-κB-mediated inflammation in severe acute respiratory syndrome coronavirus-infected mice increases survival. Journal of Virology. 2014;88(2):913–924. doi: 10.1128/JVI.02576-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deyde V.M., Xu X., Bright R.A., Shaw M., Smith C.B., Zhang Y. Surveillance of resistance to adamantanes among influenza A(H3N2) and A(H1N1) viruses isolated worldwide. Journal of Infectious Diseases. 2007;196(2):249–257. doi: 10.1086/518936. [DOI] [PubMed] [Google Scholar]
- Do H.Q., Wittlich M., Glück J.M., Möckel L., Willbold D., Koenig B.W., Heise H. Full-length Vpu and human CD4(372–433) in phospholipid bilayers as seen by magic angle spinning NMR. Biological Chemistry. 2013;394(11):1453–1463. doi: 10.1515/hsz-2013-0194. [DOI] [PubMed] [Google Scholar]
- Douglas J.L., Panis M.L., Ho E., Lin K.Y., Krawczyk S.H., Grant D.M. Small molecules VP-14637 and JNJ-2408068 inhibit respiratory syncytial virus fusion by similar mechanisms. Antimicrobial Agents and Chemotherapy. 2005;49(6):2460–2466. doi: 10.1128/AAC.49.6.2460-2466.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dowell S.F., Anderson L.J., Gary H.E., Jr., Erdman D.D., Plouffe J.F., File T.M., Jr. Respiratory syncytial virus is an important cause of community-acquired lower respiratory infection among hospitalized adults. Journal of Infectious Diseases. 1996;174(3):456–462. doi: 10.1093/infdis/174.3.456. [DOI] [PubMed] [Google Scholar]
- Doyle D.A., Cabral J.M., Pfuetzner R.A., Kuo A., Gulbis J.M., Cohen S.L. The structure of the potassium channel: Molecular basis of K + conduction and selectivity. Science. 1998;280(5360):69–77. doi: 10.1126/science.280.5360.69. [DOI] [PubMed] [Google Scholar]
- Drake J.W. Comparative rates of spontaneous mutation. Nature. 1969;221(5186):1132. doi: 10.1038/2211132a0. [DOI] [PubMed] [Google Scholar]
- Du J., Cross T.A., Zhou H.X. Recent progress in structure-based anti-influenza drug design. Drug Discovery Today. 2012;17(19–20):1111–1120. doi: 10.1016/j.drudis.2012.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duff K.C., Ashley R.H. The transmembrane domain of influenza A M2 protein forms amantadine-sensitive proton channels in planar lipid bilayers. Virology. 1992;190(1):485–489. doi: 10.1016/0042-6822(92)91239-q. [DOI] [PubMed] [Google Scholar]
- Ellis L., Dang X., Norton E., Koralnik I. Role of agnogene deletion and archetype-like regulatory region in a JCV strain isolated from the brain of a patient with JCV Encephalopathy (JCVE) Journal of Neurovirology. 2012;18:33–34. [Google Scholar]
- Ellis L.C., Norton E., Dang X., Koralnik I.J. Agnogene deletion in a novel pathogenic JC virus isolate impairs VP1 expression and virion production. PLoS One. 2013;8(11) doi: 10.1371/journal.pone.0080840. e80840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enjuanes L., Brian D., Cavanagh D., Holmes K., Lai M.M.C., Laude H. Coronaviridae. In: van Regenmortel M.H.V., Fauquet C.M., Bishop D.H.L., Carsten E.B., Estes M.K., Lemon S.M., McGeoch D.J., Maniloff J., Mayo M.A., Pringle C.R., Wickner R.B., editors. Virus taxonomy. Classification and nomenclature of viruses. Academic Press; San Diego: 2000. pp. 835–849. [Google Scholar]
- Enjuanes L., Nieto-Torres J.L., Jimenez-Guardeno J.M., DeDiego M.L. Recombinant live vaccines to protect against the severe acute respiratory syndrome coronavirus. In: Dormitzer P.R., editor. Replicating vaccines. Springer; Basel: 2011. pp. 73–97. (Birkhauser advances in infectious diseases). [Google Scholar]
- Ewart G.D., Sutherland T., Gage P.W., Cox G.B. The Vpu protein of human immunodeficiency virus type 1 forms cation-selective ion channels. Journal of Virology. 1996;70(10):7108–7115. doi: 10.1128/jvi.70.10.7108-7115.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiore A.E., Fry A., Shay D., Gubareva L., Bresee J.S., Uyeki T.M. Antiviral agents for the treatment and chemoprophylaxis of influenza—Recommendations of the Advisory Committee on Immunization Practices (ACIP) MMWR Surveillance Summaries. 2011;60(1):1–24. [PubMed] [Google Scholar]
- Fischer W.B., Li L.H., Mahato D.R., Wang Y.T., Chen C.P. Viral channel proteins in intracellular protein-protein communication: Vpu of HIV-1, E5 of HPV16 and p7 of HCV. Biochimica et Biophysica Acta. 2014;1838(4):1113–1121. doi: 10.1016/j.bbamem.2013.08.017. [DOI] [PubMed] [Google Scholar]
- Fischer W.B., Wang Y.T., Schindler C., Chen C.P. Mechanism of function of viral channel proteins and implications for drug development. International Review of Cell and Molecular Biology. 2012;294:259–321. doi: 10.1016/B978-0-12-394305-7.00006-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foster T.L., Thompson G.S., Kalverda A.P., Kankanala J., Bentham M., Wetherill L.F. Structure-guided design affirms inhibitors of hepatitis C virus p7 as a viable class of antivirals targeting virion release. Hepatology. 2014;59(2):408–422. doi: 10.1002/hep.26685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foster T.L., Verow M., Wozniak A.L., Bentham M.J., Thompson J., Atkins E. Resistance mutations define specific antiviral effects for inhibitors of the hepatitis C virus p7 ion channel. Hepatology. 2011;54(1):79–90. doi: 10.1002/hep.24371. [DOI] [PubMed] [Google Scholar]
- Fuentes S., Tran K.C., Luthra P., Teng M.N., He B. Function of the respiratory syncytial virus small hydrophobic protein. Journal of Virology. 2007;81(15):8361–8366. doi: 10.1128/JVI.02717-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furuse Y., Suzuki A., Oshitani H. Large-scale sequence analysis of M gene of influenza A viruses from different species: Mechanisms for emergence and spread of amantadine resistance. Antimicrobial Agents and Chemotherapy. 2009;53(10):4457–4463. doi: 10.1128/aac.00650-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaedigk-Nitschko K., Ding M., Levy M.A., Schlesinger M.J. Site-directed mutations in the Sindbis virus 6K protein reveal sites for fatty acylation and the underacylated protein affects virus release and virion structure. Virology. 1990;175(1):282–291. doi: 10.1016/0042-6822(90)90210-i. [DOI] [PubMed] [Google Scholar]
- Gan S.W., Ng L., Xin L., Gong X., Torres J. Structure and ion channel activity of the human respiratory syncytial virus (hRSV) small hydrophobic protein transmembrane domain. Protein Science. 2008;17:813–820. doi: 10.1110/ps.073366208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gan S.W., Surya W., Vararattanavech A., Torres J. Two different conformations in hepatitis C virus p7 protein account for proton transport and dye release. PLoS One. 2014;9(1) doi: 10.1371/journal.pone.0078494. e78494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gan S.W., Tan E., Lin X., Yu D., Wang J., Tan G.M.-Y. The small hydrophobic protein of the human respiratory syncytial virus forms pentameric ion channels. Journal of Biological Chemistry. 2012;287(29):24671–24689. doi: 10.1074/jbc.M111.332791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gentzsch J., Brohm C., Steinmann E., Friesland M., Menzel N., Vieyres G. Hepatitis C virus p7 is critical for capsid assembly and envelopment. PLoS Pathogens. 2013;9(5):1–24. doi: 10.1371/journal.ppat.1003355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerits N., Johannessen M., Tummler C., Walquist M., Kostenko S., Snapkov I. Agnoprotein of polyomavirus BK interacts with proliferating cell nuclear antigen and inhibits DNA replication. Virology Journal. 2015;12:7. doi: 10.1186/S12985-014-0220-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerits N., Moens U. Agnoprotein of mammalian polyomaviruses. Virology. 2012;432(2):316–326. doi: 10.1016/j.virol.2012.05.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gervais C., Do F., Cantin A., Kukolj G., White P.W., Gauthier A., Vaillancourt F.H. Development and validation of a high-throughput screening assay for the hepatitis C virus p7 viroporin. Journal of Biomolecular Screening. 2011;16(3):363–369. doi: 10.1177/1087057110396215. [DOI] [PubMed] [Google Scholar]
- Godet M., L'Haridon R., Vautherot J.-F., Laude H. TGEV corona virus ORF4 encodes a membrane protein that is incorporated into virions. Virology. 1992;188(2):666–675. doi: 10.1016/0042-6822(92)90521-P. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottwein J.M., Jensen T.B., Mathiesen C.K., Meuleman P., Serre S.B.N., Lademann J.B. Development and application of hepatitis C reporter viruses with genotype 1 to 7 core-nonstructural protein 2 (NS2) expressing fluorescent proteins or luciferase in modified JFH1 NS5A. Journal of Virology. 2011;85(17):8913–8928. doi: 10.1128/jvi.00049-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham R.L., Becker M.M., Eckerle L.D., Bolles M., Denison M.R., Baric R.S. A live, impaired-fidelity coronavirus vaccine protects in an aged, immunocompromised mouse model of lethal disease. Nature Medicine. 2012;18(12):1820–1826. doi: 10.1038/Nm.2972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grambas S., Bennett M.S., Hay A.J. Influence of amantadine resistance mutations on the pH regulatory function of the M2 protein of influenza A viruses. Virology. 1992;191(2):541–549. doi: 10.1016/0042-6822(92)90229-i. [DOI] [PubMed] [Google Scholar]
- Grambas S., Hay A.J. Maturation of influenza A virus hemagglutinin—Estimates of the pH encountered during transport and its regulation by the M2 protein. Virology. 1992;190(1):11–18. doi: 10.1016/0042-6822(92)91187-y. [DOI] [PubMed] [Google Scholar]
- Griffin S.D., Beales L.P., Clarke D.S., Worsfold O., Evans S.D., Jaeger J. The p7 protein of hepatitis C virus forms an ion channel that is blocked by the antiviral drug, Amantadine. FEBS Letters. 2003;535(1–3):34–38. doi: 10.1016/s0014-5793(02)03851-6. [DOI] [PubMed] [Google Scholar]
- Griffin S., Stgelais C., Owsianka A.M., Patel A.H., Rowlands D., Harris M. Genotype-dependent sensitivity of hepatitis C virus to inhibitors of the p7 ion channel. Hepatology. 2008;48(6):1779–1790. doi: 10.1002/hep.22555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu R.X., Liu L.A., Wei D.Q. Structural and energetic analysis of drug inhibition of the influenza A M2 proton channel. Trends in Pharmacological Sciences. 2013;34(10):571–580. doi: 10.1016/j.tips.2013.08.003. [DOI] [PubMed] [Google Scholar]
- Gu R.X., Liu L.A., Wei D.Q., Du J.G., Liu L., Liu H. Free energy calculations on the two drug binding sites in the M2 proton channel. Journal of the American Chemical Society. 2011;133(28):10817–10825. doi: 10.1021/ja1114198. [DOI] [PubMed] [Google Scholar]
- Guo H.C., Jin Y., Zhi X.Y., Yan D., Sun S.Q. NLRP3 inflammasome activation by viroporins of animal viruses. Viruses. 2015;7(7):3380–3391. doi: 10.3390/v7072777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall C.B., Powell K.R., Macdonald N.E., Gala C.L., Menegus M.E., Suffin S.C., Cohen H.J. Respiratory syncytial viral-infection in children with compromised immune function. The New England Journal of Medicine. 1986;315(2):77–81. doi: 10.1056/Nejm198607103150201. [DOI] [PubMed] [Google Scholar]
- Haqshenas G., Dong X., Ewart G., Bowden S., Gowans E.J. A 2a/1b full-length p7 inter-genotypic chimeric genome of hepatitis C virus is infectious in vitro. Virology. 2007;360(1):17–26. doi: 10.1016/j.virol.2006.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harada T., Tautz N., Thiel H.J. E2-p7 region of the bovine viral diarrhea virus polyprotein: Processing and functional studies. Journal of Virology. 2000;74(20):9498–9506. doi: 10.1128/jvi.74.20.9498-9506.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatta M., Goto H., Kawaoka Y. Influenza B virus requires BM2 protein for replication. Journal of Virology. 2004;78(11):5576–5583. doi: 10.1128/JVI.78.11.5576-5583.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hay A.J., Gregory V., Douglas A.R., Yi P.L. The evolution of human influenza viruses. Philosophical Transactions of the Royal Society, B: Biological Sciences. 2001;356(1416):1861–1870. doi: 10.1098/rstb.2001.0999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hay A.J., Wolstenholme A.J., Skehel J.J., Smith M.H. The molecular basis of the specific anti-influenza action of amantadine. EMBO Journal. 1985;4(11):3021–3024. doi: 10.1002/j.1460-2075.1985.tb04038.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayden F.G., De Jong M.D. Emerging influenza antiviral resistance threats. Journal of Infectious Diseases. 2011;203(1):6–10. doi: 10.1093/infdis/jiq012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heald-Sargent T., Gallagher T. Ready, set, fuse! The coronavirus spike protein and acquisition of fusion competence. Viruses. 2012;4(4):557–580. doi: 10.3390/v4040557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrmann E., Lee J.H., Marinos G., Modi M., Zeuzem S. Effect of ribavirin on hepatitis C viral kinetics in patients treated with pegylated interferon. Hepatology. 2003;37(6):1351–1358. doi: 10.1053/jhep.2003.50218. [DOI] [PubMed] [Google Scholar]
- Hogue B.G., Machamer C.E. Coronavirus structural proteins and virus assembly. In: Perlman S., Gallagher T., Snijder E.J., editors. Nidoviruses. ASM Press; Washington DC, USA: 2008. pp. 179–200. [Google Scholar]
- Holmes K.V. SARS coronavirus: A new challenge for prevention and therapy. The Journal of Clinical Investigation. 2003;111(11):1605–1609. doi: 10.1172/JCI18819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoofnagle J.H., Mullen K.D., Jones D.B., Rustgi V.K., Dibisceglie A.M., Peters M.G. Treatment of chronic non-a non-B hepatitis with recombinant human alpha-interferon. Hepatology. 1986;6(5):1220. doi: 10.1056/NEJM198612183152503. [DOI] [PubMed] [Google Scholar]
- Hout D.R., Gomez M.L., Pacyniak E., Gomez L.M., Inbody S.H., Mulcahy E.R. Scrambling of the amino acids within the transmembrane domain of Vpu results in a simian-human immunodeficiency virus (SHIVTM) that is less pathogenic for pig-tailed macaques. Virology. 2005;339(1):56–69. doi: 10.1016/j.virol.2005.04.038. [DOI] [PubMed] [Google Scholar]
- Hout D.R., Gomez L.M., Pacyniak E., Miller J.M., Hill M.S., Stephens E.B. A single amino acid substitution within the transmembrane domain of the human immunodeficiency virus type 1 Vpu protein renders simian-human immunodeficiency virus (SHIVKU-1bMC33) susceptible to rimantadine. Virology. 2006;348(2):449–461. doi: 10.1016/j.virol.2005.12.025. [DOI] [PubMed] [Google Scholar]
- Hsu H.J., Fischer W.B. In silico investigations of possible routes of assembly of ORF 3a from SARS-CoV. Journal of Molecular Modeling. 2012;18(2):501–514. doi: 10.1007/s00894-011-1092-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu K., Han J., Shinlapawittayatorn K., Deschenes I., Marbán E. Membrane potential depolarization as a triggering mechanism for Vpu-mediated HIV-1 release. Biophysical Journal. 2010;99(6):1718–1725. doi: 10.1016/j.bpj.2010.07.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu W., Zeng S., Li C., Jie Y., Li Z., Chen L. Identification of hits as matrix-2 protein inhibitors through the focused screening of a small primary amine library. Journal of Medicinal Chemistry. 2010;53(9):3831–3834. doi: 10.1021/jm901664a. [DOI] [PubMed] [Google Scholar]
- Ichinohe T., Pang I.K., Iwasaki A. Influenza virus activates inflammasomes via its intracellular M2 ion channel. Nature Immunology. 2010;11(5):404–410. doi: 10.1038/ni.1861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ilk N., Egelseer E.M., Sleytr U.B. S-layer fusion proteins-construction principles and applications. Current Opinion in Biotechnology. 2011;22(6):824–831. doi: 10.1016/j.copbio.2011.05.510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imai M., Kawasaki K., Odagiri T. Cytoplasmic domain of influenza B virus BM2 protein plays critical roles in production of infectious virus. Journal of Virology. 2008;82(2):728–739. doi: 10.1128/JVI.01752-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imai M., Watanabe S., Ninomiya A., Obuchi M., Odagiri T. Influenza B virus BM2 protein is a crucial component for incorporation of viral ribonucleoprotein complex into virions during virus assembly. Journal of Virology. 2004;78(20):11007–11015. doi: 10.1128/JVI.78.20.11007-11015.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito M., Yanagi Y., Ichinohe T. Encephalomyocarditis virus viroporin 2B activates NLRP3 inflammasome. PLoS Pathogens. 2012;8(8) doi: 10.1371/journal.ppat.1002857. e1002857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivanova L., Lustig S., Schlesinger M.J. A pseudo-revertant of a Sindbis virus 6K protein mutant, which corrects for aberrant particle formation, contains two new mutations that map to the ectodomain of the E2 glycoprotein. Virology. 1995;206(2):1027–1034. doi: 10.1006/viro.1995.1025. [DOI] [PubMed] [Google Scholar]
- Jack P.J.M., Boyle D.B., Eaton B.T., Wang L.F. The complete genome sequence of J virus reveals a unique genome structure in the family Paramyxoviridae. Journal of Virology. 2005;79(16):10690–10700. doi: 10.1128/Jvi.79.16.10690-10700.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Javier R.T., Rice A.P. Emerging theme: Cellular PDZ proteins as common targets of pathogenic viruses. Journal of Virology. 2011;85(22):11544–11556. doi: 10.1128/JVI.05410-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin H., Zhou H., Cheng X., Tang R., Munoz M., Nguyen N. Recombinant respiratory syncytial viruses with deletions in the NS1, NS2, SH, and M2-2 genes are attenuated in vitro and in vivo. Virology. 2000;273(1):210–218. doi: 10.1006/viro.2000.0393. [DOI] [PubMed] [Google Scholar]
- Jones C.T., Murray C.L., Eastman D.K., Tassello J., Rice C.M. Hepatitis C virus p7 and NS2 proteins are essential for production of infectious virus. Journal of Virology. 2007;81(16):8374–8383. doi: 10.1128/JVI.00690-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karron R.A., Buonagurio D.A., Georgiu A.F., Whitehead S.S., Adamus J.E., Clements-Mann M.L. Respiratory syncytial virus (RSV) SH and G proteins are not essential for viral replication in vitro: Clinical evaluation and molecular characterization of a cold-passaged, attenuated RSV subgroup B mutant. Proceedings of the National Academy of Sciences of the United States of America. 1997;94(25):13961–13966. doi: 10.1073/pnas.94.25.13961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kato T., Date T., Miyamoto M., Furusaka A., Tokushige K., Mizokami M., Wakita T. Efficient replication of the genotype 2a hepatitis C virus subgenomic replicon. Gastroenterology. 2003;125(6):1808–1817. doi: 10.1053/j.gastro.2003.09.023. [DOI] [PubMed] [Google Scholar]
- Khoury G., Ewart G., Luscombe C., Miller M., Wilkinson J. Antiviral efficacy of the novel compound BIT225 against HIV-1 release from human macrophages. Antimicrobial Agents and Chemotherapy. 2010;54(2):835–845. doi: 10.1128/aac.01308-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilianski A., Baker S.C. Cell-based antiviral screening against coronaviruses: Developing virus-specific and broad-spectrum inhibitors. Antiviral Research. 2014;101:105–112. doi: 10.1016/j.antiviral.2013.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilianski A., Mielech A.M., Deng X., Baker S.C. Assessing activity and inhibition of Middle East respiratory syndrome coronavirus papain-like and 3C-like proteases using luciferase-based biosensors. Journal of Virology. 2013;87(21):11955–11962. doi: 10.1128/JVI.02105-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klimkait T., Strebel K., Hoggan M.D., Martin M.A., Orenstein J.M. The human immunodeficiency virus type 1-specific protein vpu is required for efficient virus maturation and release. Journal of Virology. 1990;64(2):621–629. doi: 10.1128/jvi.64.2.621-629.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kovacs F.A., Denny J.K., Song Z., Quine J.R., Cross T.A. Helix tilt of the M2 transmembrane peptide from influenza A virus: An intrinsic property. Journal of Molecular Biology. 2000;295(1):117–125. doi: 10.1006/jmbi.1999.3322. [DOI] [PubMed] [Google Scholar]
- Kumar M., Behera A.K., Lockey R.F., Zhang J., Bhullar G., De La Cruz C.P. Intranasal gene transfer by chitosan-DNA nanospheres protects BALB/c mice against acute respiratory syncytial virus infection. Human Gene Therapy. 2002;13(12):1415–1425. doi: 10.1089/10430340260185058. [DOI] [PubMed] [Google Scholar]
- Kurtz S., Luo G.X., Hahnenberger K.M., Brooks C., Gecha O., Ingalls K. Growth impairment resulting from expression of influenza-virus M2 protein in Saccharomyces cerevisiae: Identification of a novel inhibitor of influenza-virus. Antimicrobial Agents and Chemotherapy. 1995;39(10):2204–2209. doi: 10.1128/aac.39.10.2204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagoja I.M., De Clercq E. Anti-influenza virus agents: Synthesis and mode of action. Medicinal Research Reviews. 2008;28(1):1–38. doi: 10.1002/med.20096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamb R.A., Zebedee S.L., Richardson C.D. Influenza virus M2 protein is an integral membrane protein expressed on the infected-cell surface. Cell. 1985;40(3):627–633. doi: 10.1016/0092-8674(85)90211-9. [DOI] [PubMed] [Google Scholar]
- Lambert D.M., Barney S., Lambert A.L., Guthrie K., Medinas R., Davis D.E. Peptides from conserved regions of paramyxovirus fusion (F) proteins are potent inhibitors of viral fusion. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(5):2186–2191. doi: 10.1073/pnas.93.5.2186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamirande E.W., DeDiego M.L., Roberts A., Jackson J.P., Alvarez E., Sheahan T. A live attenuated severe acute respiratory syndrome coronavirus is immunogenic and efficacious in golden Syrian hamsters. Journal of Virology. 2008;82(15):7721–7724. doi: 10.1128/JVI.00304-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H., Atkins E., Bruckner J., McArdle S., Qiu W.C., Thomassen L.V. Genetic and functional heterogeneity of the hepatitis C virus p7 ion channel during natural chronic infection. Virology. 2012;423(1):30–37. doi: 10.1016/j.virol.2011.11.011. [DOI] [PubMed] [Google Scholar]
- Li Y., Jain N., Limpanawat S., To J., Quistgaard E.M., Nordlund P. Interaction between human BAP31 and respiratory syncytial virus small hydrophobic (SH) protein. Virology. 2015;482:105–110. doi: 10.1016/j.virol.2015.03.034. [DOI] [PubMed] [Google Scholar]
- Li Y., Surya W., Claudine S., Torres J. Structure of a conserved Golgi complex-targeting signal in coronavirus envelope proteins. Journal of Biological Chemistry. 2014;289(18):12535–12549. doi: 10.1074/jbc.M114.560094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y., To J., Verdià-Baguena C., Dossena S., Surya W., Huang M. Inhibition of the human respiratory syncytial virus small hydrophobic protein and structural variations in a bicelle environment. Journal of Virology. 2014;88(20):11899–11914. doi: 10.1128/JVI.00839-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Z., Xu J., Patel J., Fuentes S., Lin Y.A., Anderson D. Function of the small hydrophobic protein of J paramyxovirus. Journal of Virology. 2011;85(1):32–42. doi: 10.1128/Jvi.01673-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao Y., Yuan Q., Torres J., Tam J.P., Liu D.X. Biochemical and functional characterization of the membrane association and membrane permeabilizing activity of the severe acute respiratory syndrome coronavirus envelope protein. Virology. 2006;349(2):264–275. doi: 10.1016/j.virol.2006.01.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liljestrom P., Garoff H. Internally located cleavable signal sequences direct the formation of Semliki Forest virus membrane proteins from a polyprotein precursor. Journal of Virology. 1991;65(1):147–154. doi: 10.1128/jvi.65.1.147-154.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liljestrom P., Lusa S., Huylebroeck D., Garoff H. In vitro mutagenesis of a full-length cDNA clone of Semliki Forest virus: The small 6,000-molecular-weight membrane protein modulates virus release. Journal of Virology. 1991;65(8):4107–4113. doi: 10.1128/jvi.65.8.4107-4113.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim K.P., Liu D.X. The missing link in coronavirus assembly. Retention of the avian coronavirus infectious bronchitis virus envelope protein in the pre-Golgi compartments and physical interaction between the envelope and membrane proteins. Journal of Biological Chemistry. 2001;276(20):17515–17523. doi: 10.1074/jbc.M009731200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Y., Bright A.C., Rothermel T.A., He B. Induction of apoptosis by paramyxovirus simian virus 5 lacking a small hydrophobic gene. Journal of Virology. 2003;77(6):3371–3383. doi: 10.1128/JVI.77.6.3371-3383.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin C., Lindenbach B.D., Prágai B.M., McCourt D.W., Rice C.M. Processing in the hepatitis C virus E2-NS2 region: Identification of p7 and two distinct E2-specific products with different C termini. Journal of Virology. 1994;68(8):5063–5073. doi: 10.1128/jvi.68.8.5063-5073.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindenbach B.D., Rice C.M. Unravelling hepatitis C virus replication from genome to function. Nature. 2005;436(7053):933–938. doi: 10.1038/nature04077. [DOI] [PubMed] [Google Scholar]
- Loewy A., Smyth J., von Bonsdorff C.H., Liljestrom P., Schlesinger M.J. The 6-kilodalton membrane protein of Semliki Forest virus is involved in the budding process. Journal of Virology. 1995;69(1):469–475. doi: 10.1128/jvi.69.1.469-475.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lohmann V., Korner F., Koch J., Herian U., Theilmann L., Bartenschlager R. Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science. 1999;285(5424):110–113. doi: 10.1126/science.285.5424.110. [DOI] [PubMed] [Google Scholar]
- Lopez L.A., Riffle A.J., Pike S.L., Gardner D., Hogue B.G. Importance of conserved cysteine residues in the coronavirus envelope protein. Journal of Virology. 2008;82(6):3000–3010. doi: 10.1128/JVI.01914-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lou Z., Sun Y., Rao Z. Current progress in antiviral strategies. Trends in Pharmacological Sciences. 2014;35(2):86–102. doi: 10.1016/j.tips.2013.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Low K.W., Tan T., Ng K., Tan B.H., Sugrue R.J. The RSV F and G glycoproteins interact to form a complex on the surface of infected cells. Biochemical and Biophysical Research Communications. 2008;366(2):308–313. doi: 10.1016/j.bbrc.2007.11.042. [DOI] [PubMed] [Google Scholar]
- Lu J.X., Sharpe S., Ghirlando R., Yau W.M., Tycko R. Oligomerization state and supramolecular structure of the HIV-1 Vpu protein transmembrane segment in phospholipid bilayers. Protein Science. 2010;19(10):1877–1896. doi: 10.1002/pro.474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu W., Zheng B.J., Xu K., Schwarz W., Du L.Y., Wong C.K.L. Severe acute respiratory syndrome-associated coronavirus 3a protein forms an ion channel and modulates virus release. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(33):12540–12545. doi: 10.1073/pnas.0605402103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luik P., Chew C., Aittoniemi J., Chang J., Wentworth P., Jr., Dwek R.A. The 3-dimensional structure of a hepatitis C virus p7 ion channel by electron microscopy. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(31):12712–12716. doi: 10.1073/pnas.0905966106. 0905966106 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luscombe C.A., Huang Z., Murray M.G., Miller M., Wilkinson J., Ewart G.D. A novel Hepatitis C virus p7 ion channel inhibitor, BIT225, inhibits bovine viral diarrhea virus in vitro and shows synergism with recombinant interferon-α-2b and nucleoside analogues. Antiviral Research. 2010;86(2):144–153. doi: 10.1016/j.antiviral.2010.02.312. [DOI] [PubMed] [Google Scholar]
- Madan V., Bartenschlager R. Structural and functional properties of the hepatitis C virus p7 viroporin. Viruses. 2015;7(8):4461–4481. doi: 10.3390/v7082826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magadán J.G., Bonifacino J.S. Transmembrane domain determinants of CD4 downregulation by HIV-1 Vpu. Journal of Virology. 2012;86(2):757–772. doi: 10.1128/jvi.05933-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maldarelli F., Chen M.Y., Willey R.L., Strebel K. Human immunodeficiency virus type 1 Vpu protein is an oligomeric type I integral membrane protein. Journal of Virology. 1993;67(8):5056–5061. doi: 10.1128/jvi.67.8.5056-5061.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manns M.P., Wedemeyer H., Cornberg M. Treating viral hepatitis C: Efficacy, side effects, and complications. Gut. 2006;55(9):1350–1359. doi: 10.1136/gut.2005.076646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCown M.F., Pekosz A. Distinct domains of the influenza a virus M2 protein cytoplasmic tail mediate binding to the M1 protein and facilitate infectious virus production. Journal of Virology. 2006;80(16):8178–8189. doi: 10.1128/JVI.00627-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLellan J.S., Chen M., Joyce M.G., Sastry M., Stewart-Jones G.B.E., Yang Y.P. Structure-based design of a fusion glycoprotein vaccine for respiratory syncytial virus. Science. 2013;342(6158):592–598. doi: 10.1126/science.1243283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLellan J.S., Chen M., Leung S., Graepel K.W., Du X.L., Yang Y.P. Structure of RSV fusion glycoprotein trimer bound to a prefusion-specific neutralizing antibody. Science. 2013;340(6136):1113–1117. doi: 10.1126/science.1234914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNatt M.W., Zang T., Bieniasz P.D. Vpu binds directly to tetherin and displaces it from nascent virions. PLoS Pathogens. 2013;9(4) doi: 10.1371/journal.ppat.1003299. e1003299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melton J.V., Ewart G.D., Weir R.C., Board P.G., Lee E., Gage P.W. Alphavirus 6K proteins form ion channels. Journal of Biological Chemistry. 2002;277(49):46923–46931. doi: 10.1074/jbc.M207847200. [DOI] [PubMed] [Google Scholar]
- Mihm U., Grigorian N., Welsch C., Herrmann E., Kronenberger B., Teuber G. Amino acid variations in hepatitis C virus p7 and sensitivity to antiviral combination therapy with amantadine in chronic hepatitis C. Antiviral Therapy. 2006;11(4):507–519. [PubMed] [Google Scholar]
- Miyagi E., Andrew A.J., Kao S., Strebe K. Vpu enhances HIV-1 virus release in the absence of Bst-2 cell surface down-modulation and intracellular depletion. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(8):2868–2873. doi: 10.1073/pnas.0813223106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mizushima H., Hijikata M., Asabe S.I., Hirota M., Kimura K., Shimotohno K. Two hepatitis C virus glycoprotein E2 products with different C termini. Journal of Virology. 1994;68(10):6215–6222. doi: 10.1128/jvi.68.10.6215-6222.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molinari N.A.M., Ortega-Sanchez I.R., Messonnier M.L., Thompson W.W., Wortley P.M., Weintraub E., Bridges C.B. The annual impact of seasonal influenza in the US: Measuring disease burden and costs. Vaccine. 2007;25(27):5086–5096. doi: 10.1016/j.vaccine.2007.03.046. [DOI] [PubMed] [Google Scholar]
- Montserret R., Saint N., Vanbelle C., Salvay A.G., Simorre J.P., Ebel C. NMR structure and ion channel activity of the p7 protein from hepatitis C virus. Journal of Biological Chemistry. 2010;285(41):31446–31461. doi: 10.1074/jbc.M110.122895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moradpour D., Penin F. Hepatitis C virus proteins: From structure to function. 2013. pp. 113–142.http://dx.doi.org/10.1007/978-3-642-27340-7_5 (369 Hepatitis C virus: From molecular virology to antiviral therapy). [DOI] [PubMed] [Google Scholar]
- Morse S.S. Oxford University Press; New York, Oxford: 1993. Emerging viruses. [Google Scholar]
- Mould J.A., Paterson R.G., Takeda M., Ohigashi Y., Venkataraman P., Lamb R.A., Pinto L.H. Influenza B virus BM2 protein has ion channel activity that conducts protons across membranes. Developmental Cell. 2003;5(1):175–184. doi: 10.1016/s1534-5807(03)00190-4. [DOI] [PubMed] [Google Scholar]
- Nair H., Nokes D.J., Gessner B.D., Dherani M., Madhi S.A., Singleton R.J. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: A systematic review and meta-analysis. Lancet. 2010;375(9725):1545–1555. doi: 10.1016/S0140-6736(10)60206-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neil S.J.D., Zang T., Bieniasz P.D. Tetherin inhibits retrovirus release and is antagonized by HIV-1 Vpu. Nature. 2008;451(7177):425–430. doi: 10.1038/nature06553. [DOI] [PubMed] [Google Scholar]
- Netland J., DeDiego M.L., Zhao J., Fett C., Alvarez E., Nieto-Torres J.L. Immunization with an attenuated severe acute respiratory syndrome coronavirus deleted in E protein protects against lethal respiratory disease. Virology. 2010;399(1):120–128. doi: 10.1016/j.virol.2010.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neumann A.U., Lam N.P., Dahari H., Gretch D.R., Wiley T.E., Layden T.J., Perelson A.S. Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-alpha therapy. Science. 1998;282(5386):103–107. doi: 10.1126/science.282.5386.103. [DOI] [PubMed] [Google Scholar]
- Neumann G., Noda T., Kawaoka Y. Emergence and pandemic potential of swine-origin H1N1 influenza virus. Nature. 2009;459(7249):931–939. doi: 10.1038/nature08157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieto-Torres J.L., DeDiego M.L., Alvarez E., Jimenez-Guardeno J.M., Regla-Nava J.A., Llorente M. Subcellular location and topology of severe acute respiratory syndrome coronavirus envelope protein. Virology. 2011;415(2):69–82. doi: 10.1016/j.virol.2011.03.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieto-Torres J.L., Dediego M.L., Verdia-Baguena C., Jimenez-Guardeno J.M., Regla-Nava J.A., Fernandez-Delgado R. Severe acute respiratory syndrome coronavirus envelope protein ion channel activity promotes virus fitness and pathogenesis. PLoS Pathogens. 2014;10(5) doi: 10.1371/journal.ppat.1004077. e1004077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieto-Torres J.L., Verdia-Baguena C., Castano-Rodriguez C., Aguilella V.M., Enjuanes L. Relevance of viroporin ion channel activity on viral replication and pathogenesis. Viruses. 2015;7(7):3552–3573. doi: 10.3390/v7072786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieva J.L., Agirre A., Nir S., Carrasco L. Mechanisms of membrane permeabilization by picornavirus 2B viroporin. FEBS Letters. 2003;552(1):68–73. doi: 10.1016/S0014-5793(03)00852-4. [DOI] [PubMed] [Google Scholar]
- Nieva J.L., Carrasco L. Viroporins: Structures and functions beyond cell membrane permeabilization. Viruses. 2015;7(10):5169–5171. doi: 10.3390/v7102866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieva J.L., Madan V., Carrasco L. Viroporins: Structure and biological functions. Nature Reviews. Microbiology. 2012;10(8):563–574. doi: 10.1038/Nrmicro2820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Opella S.J. Relating structure and function of viral membrane-spanning miniproteins. Current Opinion in Virology. 2015;12:121–125. doi: 10.1016/j.coviro.2015.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ouyang B., Xie S., Berardi M.J., Zhao X., Dev J., Yu W. Unusual architecture of the p7 channel from hepatitis C virus. Nature. 2013;498(7455):521–525. doi: 10.1038/nature12283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parthasarathy K., Lu H., Surya W., Vararattanavech A., Pervushin K., Torres A. Expression and purification of coronavirus envelope proteins using a modified beta-barrel construct. Protein Expression and Purification. 2012;85(1):133–141. doi: 10.1016/j.pep.2012.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parthasarathy K., Ng L., Lin X., Liu D.X., Pervushin K., Gong X., Torres J. Structural flexibility of the pentameric SARS coronavirus envelope protein ion channel. Biophysical Journal. 2008;95(6):L39–L41. doi: 10.1529/biophysj.108.133041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pastey M.K., Gower T.L., Spearman P.W., Crowe J.E., Graham B.S. A RhoA-derived peptide inhibits syncytium formation induced by respiratory syncytial virus and parainfluenza virus type 3. Nature Medicine. 2000;6(1):35–40. doi: 10.1038/71503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patargias G., Zitzmann N., Dwek R., Fischer W.B. Protein-protein interactions: Modeling the hepatitis C virus ion channel p7. Journal of Medicinal Chemistry. 2006;49(2):648–655. doi: 10.1021/jm050721e. [DOI] [PubMed] [Google Scholar]
- Paterson R.G., Takeda M., Ohigashi Y., Pinto L.H., Lamb R.A. Influenza B virus BM2 protein is an oligomeric integral membrane protein expressed at the cell surface. Virology. 2003;306(1):7–17. doi: 10.1016/s0042-6822(02)00083-1. [DOI] [PubMed] [Google Scholar]
- Pavlovic D., Neville D.C., Argaud O., Blumberg B., Dwek R.A., Fischer W.B., Zitzmann N. The hepatitis C virus p7 protein forms an ion channel that is inhibited by long-alkyl-chain iminosugar derivatives. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(10):6104–6108. doi: 10.1073/pnas.1031527100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pawlotsky J.M. New hepatitis C therapies: The toolbox, strategies, and challenges. Gastroenterology. 2014;146(5):1176–1192. doi: 10.1053/j.gastro.2014.03.003. [DOI] [PubMed] [Google Scholar]
- Pawlotsky J., Chevaliez S., McHutchison J.G. The hepatitis C virus life cycle as a target for new antiviral therapies. Gastroenterology. 2007;132(5):1979–1998. doi: 10.1053/j.gastro.2007.03.116. [DOI] [PubMed] [Google Scholar]
- Pervushin K., Tan E., Parthasarathy K., Lin X., Jiang F.L., Yu D. Structure and inhibition of the SARS coronavirus envelope protein ion channel. PLoS Pathogens. 2009;5(7) doi: 10.1371/journal.ppat.1000511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pielak R.M., Chou J.J. Influenza M2 proton channels. Biochimica et Biophysica Acta—Biomembranes. 2011;1808(2):522–529. doi: 10.1016/j.bbamem.2010.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pielak R.M., Oxenoid K., Chou J.J. Structural investigation of rimantadine inhibition of the AM2-BM2 chimera channel of influenza viruses. Structure. 2011;19(11):1655–1663. doi: 10.1016/j.str.2011.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pielak R.M., Schnell J.R., Chou J.J. Mechanism of drug inhibition and drug resistance of influenza A M2 channel. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(18):7379–7384. doi: 10.1073/pnas.0902548106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pietschmann T., Kaul A., Koutsoudakis G., Shavinskaya A., Kallis S., Steinmann E. Construction and characterization of infectious intragenotypic and intergenotypic hepatitis C virus chimeras. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(19):7408–7413. doi: 10.1073/pnas.0504877103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinto L.H., Holsinger L.J., Lamb R.A. Influenza virus M2 protein has ion channel activity. Cell. 1992;69(3):517–528. doi: 10.1016/0092-8674(92)90452-i. [DOI] [PubMed] [Google Scholar]
- Pinto L.H., Lamb R.A. The M2 proton channels of influenza A and B viruses. Journal of Biological Chemistry. 2006;281(14):8997–9000. doi: 10.1074/jbc.R500020200. [DOI] [PubMed] [Google Scholar]
- Pohl P. Combined transport of water and ions through membrane channels. Biological Chemistry. 2004;385(10):921–926. doi: 10.1515/Bc.2004.120. [DOI] [PubMed] [Google Scholar]
- Premkumar A., Wilson L., Ewart G.D., Gage P.W. Cation-selective ion channels formed by p7 of hepatitis C virus are blocked by hexamethylene amiloride. FEBS Letters. 2004;557(1–3):99–103. doi: 10.1016/s0014-5793(03)01453-4. [DOI] [PubMed] [Google Scholar]
- Raamsman M.J.B., Krijnse Locker J., De Hooge A., De Vries A.A.F., Griffiths G., Vennema H., Rottier P.J.M. Characterization of the coronavirus mouse hepatitis virus strain A59 small membrane protein E. Journal of Virology. 2000;74(5):2333–2342. doi: 10.1128/jvi.74.5.2333-2342.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radoicic J., Lu G.J., Opella S.J. NMR structures of membrane proteins in phospholipid bilayers. Quarterly Reviews of Biophysics. 2014;47(3):249–283. doi: 10.1017/S0033583514000080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raj V.S., Osterhaus A.D.M.E., Fouchier R.A.M., Haagmans B.L. MERS: Emergence of a novel human coronavirus. Current Opinion in Virology. 2014;5:58–62. doi: 10.1016/j.coviro.2014.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Razinkov V., Gazumyan A., Nikitenko A., Ellestad G., Krishnamurthy G. RFI-641 inhibits entry of respiratory syncytial virus via interactions with fusion protein. Chemistry and Biology. 2001;8(7):645–659. doi: 10.1016/s1074-5521(01)00042-4. [DOI] [PubMed] [Google Scholar]
- Regan J.A., Laimins L.A. Bap31 is a novel target of the human papillomavirus E5 protein. Journal of Virology. 2008;82(20):10042–10051. doi: 10.1128/JVI.01240-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rey-Carrizo M., Barniol-Xicota M., Ma C., Frigolé-Vivas M., Torres E., Naesens L. Easily accessible polycyclic amines that inhibit the wild-type and amantadine-resistant mutants of the M2 channel of influenza A virus. Journal of Medicinal Chemistry. 2014;57(13):5738–5747. doi: 10.1021/jm5005804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rey-Carrizo M., Torres E., Ma C., Barniol-Xicota M., Wang J., Wu Y. 3-azatetracyclo[5.2.1.15,8.01,5]undecane derivatives: From wild-type inhibitors of the M2 ion channel of influenza A virus to derivatives with potent activity against the V27A mutant. Journal of Medicinal Chemistry. 2013;56(22):9265–9274. doi: 10.1021/jm401340p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rixon H.W., Brown G., Aitken J., McDonald T., Graham S., Sugrue R.J. The small hydrophobic (SH) protein accumulates within lipid-raft structures of the Golgi complex during respiratory syncytial virus infection. Journal of General Virology. 2004;85(Pt. 5):1153–1165. doi: 10.1099/vir.0.19769-0. [DOI] [PubMed] [Google Scholar]
- Rixon H.W., Brown G., Murray J.T., Sugrue R.J. The respiratory syncytial virus small hydrophobic protein is phosphorylated via a mitogen-activated protein kinase p38-dependent tyrosine kinase activity during virus infection. Journal of General Virology. 2005;86(Pt. 2):375–384. doi: 10.1099/vir.0.80563-0. [DOI] [PubMed] [Google Scholar]
- Roymans D., De Bondt H.L., Arnoult E., Geluykens P., Gevers T., Van Ginderen M. Binding of a potent small-molecule inhibitor of six-helix bundle formation requires interactions with both heptad-repeats of the RSV fusion protein. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(1):308–313. doi: 10.1073/pnas.0910108106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruch T.R., Machamer C.E. A single polar residue and distinct membrane topologies impact the function of the infectious bronchitis coronavirus E protein. PLoS Pathogens. 2012;8(5) doi: 10.1371/journal.ppat.1002674. e1002674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakaguchi T., Tu Q.A., Pinto L.H., Lamb R.A. The active oligomeric state of the minimalistic influenza virus M-2 ion channel is a tetramer. Proceedings of the National Academy of Sciences of the United States of America. 1997;94(10):5000–5005. doi: 10.1073/pnas.94.10.5000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakai A., Claire M.S., Faulk K., Govindarajan S., Emerson S.U., Purcell R.H., Bukh J. The p7 polypeptide of hepatitis C virus is critical for infectivity and contains functionally important genotype-specific sequences. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(20):11646–11651. doi: 10.1073/pnas.1834545100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez-Eugenia R., Goikolea J., Gil-Carton D., Sanchez-Magraner L., Guerin D.M. Triatoma virus recombinant VP4 protein induces membrane permeability through dynamic pores. Journal of Virology. 2015;89(8):4645–4654. doi: 10.1128/JVI.00011-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanz M.A., Perez L., Carrasco L. Semliki Forest virus 6K protein modifies membrane permeability after inducible expression in Escherichia coli cells. The Journal of Biological Chemistry. 1994;269(16):12106–12110. [PubMed] [Google Scholar]
- Saribas A.S., Abou-Gharbia M., Childers W., Sariyer I.K., White M.K., Safak M. Essential roles of Leu/Ile/Phe-rich domain of JC virus agnoprotein in dimer/oligomer formation, protein stability and splicing of viral transcripts. Virology. 2013;443(1):161–176. doi: 10.1016/j.virol.2013.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sariyer I.K., Saribas A.S., White M.K., Safak M. Infection by agnoprotein-negative mutants of polyomavirus JC and SV40 results in the release of virions that are mostly deficient in DNA content. Virology Journal. 2011;8:255. doi: 10.1186/1743-422x-8-255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schepens B., Sedeyn K., Vande Ginste L., De Baets S., Schotsaert M., Roose K. Protection and mechanism of action of a novel human respiratory syncytial virus vaccine candidate based on the extracellular domain of small hydrophobic protein. EMBO Molecular Medicine. 2014;6:1436–1454. doi: 10.15252/emmm.201404005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt W.N., Nelson D.R., Pawlotsky J.M., Sherman K.E., Thomas D.L., Chung R.T. Direct-acting antiviral agents and the path to interferon independence. Clinical Gastroenterology and Hepatology. 2014;12(5):728–737. doi: 10.1016/j.cgh.2013.06.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schnell J.R., Chou J.J. Structure and mechanism of the M2 proton channel of influenza A virus. Nature. 2008;451(7178):591–595. doi: 10.1038/nature06531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schubert U., Ferrer-Montiel A.V., Oblatt-Montal M., Henklein P., Strebel K., Montal M. Identification of an ion channel activity of the Vpu transmembrane domain and its involvement in the regulation of virus release from HIV-1-infected cells. FEBS Letters. 1996;398(1):12–18. doi: 10.1016/s0014-5793(96)01146-5. [DOI] [PubMed] [Google Scholar]
- Scott C., Griffin S. Viroporins: Structure, function and potential as antiviral targets. Journal of General Virology. 2015;96(8):2000–2027. doi: 10.1099/vir.0.000201. [DOI] [PubMed] [Google Scholar]
- Segovia J., Sabbah A., Mgbemena V., Tsai S.Y., Chang T.H., Berton M.T. TLR2/MyD88/NF-kappa B pathway, reactive oxygen species, potassium efflux activates NLRP3/ASC inflammasome during respiratory syncytial virus infection. PLoS One. 2012;7(1) doi: 10.1371/journal.pone.0029695. e29695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma M., Yi M., Dong H., Qin H., Peterson E., Busath D.D.…Cross T.A. Insight into the mechanism of the influenza A proton channel from a structure in a lipid bilayer. Science. 2010;330(6003):509–512. doi: 10.1126/science.1191750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shepherd N.E., Hoang H.N., Desai V.S., Letouze E., Young P.R., Fairlie D.P. Modular alpha-helical mimetics with antiviral activity against respiratory syncitial virus. Journal of the American Chemical Society. 2006;128(40):13284–13289. doi: 10.1021/Ja064058a. [DOI] [PubMed] [Google Scholar]
- Shiraishi K., Mitamura K., Sakai-Tagawa Y., Goto H., Sugaya N., Kawaoka Y. High frequency of resistant viruses harboring different mutations in amantadine-treated children with influenza. The Journal of Infectious Diseases. 2003;188(1):57–61. doi: 10.1086/375799. [DOI] [PubMed] [Google Scholar]
- Simmonds P. The origin of Hepatitis C virus. 2013. pp. 1–15.http://dx.doi.org/10.1007/978-3-642-27340-7_1 (369 Hepatitis C virus: From molecular virology to antiviral therapy). [Google Scholar]
- Singh S.K., Möckel L., Thiagarajan-Rosenkranz P., Wittlich M., Willbold D., Koenig B.W. Mapping the interaction between the cytoplasmic domains of HIV-1 viral protein U and human CD4 with NMR spectroscopy. FEBS Journal. 2012;279(19):3705–3714. doi: 10.1111/j.1742-4658.2012.08732.x. [DOI] [PubMed] [Google Scholar]
- Siu Y.L., Teoh K.T., Lo J., Chan C.M., Kien F., Escriou N. The M, E, and N structural proteins of the severe acute respiratory syndrome coronavirus are required for efficient assembly, trafficking, and release of virus-like particles. Journal of Virology. 2008;82(22):11318–11330. doi: 10.1128/JVI.01052-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skasko M., Wang Y., Tian Y., Tokarev A., Munguia J., Ruiz A. HIV-1 Vpu protein antagonizes innate restriction factor BST-2 via lipid-embedded helix-helix interactions. Journal of Biological Chemistry. 2012;287(1):58–67. doi: 10.1074/jbc.M111.296772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smits S.L., de Lang A., van den Brand J.M.A., Leijten L.M., van IJcken W.F., Eijkemans M.J.C. Exacerbated innate host response to SARS-CoV in aged non-human primates. PLoS Pathogens. 2010;6(2) doi: 10.1371/journal.ppat.1000756. e1000756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinmann E., Penin F., Kallis S., Patel A.H., Bartenschlager R., Pietschmann T. Hepatitis C virus p7 protein is crucial for assembly and release of infectious virions. PLoS Pathogens. 2007;3(7):962–971. doi: 10.1371/journal.ppat.0030103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- StGelais C., Foster T.L., Verow M., Atkins E., Fishwick C.W.G., Rowlands D. Determinants of hepatitis C virus p7 ion channel function and drug sensitivity identified in vitro. Journal of Virology. 2009;83(16):7970–7981. doi: 10.1128/JVI.00521-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- StGelais C., Tuthill T.J., Clarke D.S., Rowlands D.J., Harris M., Griffin S. Inhibition of hepatitis C virus p7 membrane channels in a liposome-based assay system. Antiviral Research. 2007;76(1):48–58. doi: 10.1016/j.antiviral.2007.05.001. S0166-3542(07)00334-8 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stouffer A.L., Acharya R., Salom D., Levine A.S., Di Costanzo L., Soto C.S. Structural basis for the function and inhibition of an influenza virus proton channel. Nature. 2008;451(7178):596–599. doi: 10.1038/nature06528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strauss J.H., Strauss E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiological Reviews. 1994;58(3):491–562. doi: 10.1128/mr.58.3.491-562.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strebel K. HIV-1 Vpu—An ion channel in search of a job. Biochimica et Biophysica Acta—Biomembranes. 2014;1838(4):1074–1081. doi: 10.1016/j.bbamem.2013.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strebel K., Klimkait T., Martin M.A. A novel gene of HIV-1, vpu, and its 16-kilodalton product. Science. 1988;241(4870):1221–1223. doi: 10.1126/science.3261888. [DOI] [PubMed] [Google Scholar]
- Sulkowski M.S., Gardiner D.F., Rodriguez-Torres M., Reddy K.R., Hassanein T., Jacobson I. Daclatasvir plus sofosbuvir for previously treated or untreated chronic HCV infection. The New England Journal of Medicine. 2014;370(3):211–221. doi: 10.1056/NEJMoa1306218. [DOI] [PubMed] [Google Scholar]
- Suzuki T., Orba Y., Makino Y., Okada Y., Sunden Y., Hasegawa H. Viroporin activity of the JC polyomavirus is regulated by interactions with the adaptor protein complex 3. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(46):18668–18673. doi: 10.1073/pnas.1311457110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki T., Orba Y., Malcino Y., Okada Y., Sunden Y., Kimura T. Disruption of intracellular vesicular trafficking by agnoprotein is essential for viroporin activity and JC virus replication. Journal of Neurovirology. 2010;16:84. [Google Scholar]
- Suzuki T., Orba Y., Okada Y., Sunden Y., Kimura T., Tanaka S. The human polyoma JC virus agnoprotein acts as a viroporin. PLoS Pathogens. 2010;6(3) doi: 10.1371/journal.ppat.1000801. e1000801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki H., Saito R., Masuda H., Oshitani H., Sato M., Sato I. Emergence of amantadine-resistant influenza A viruses: Epidemiological study. Journal of Infection and Chemotherapy. 2003;9(3):195–200. doi: 10.1007/s10156-003-0262-6. [DOI] [PubMed] [Google Scholar]
- Takeuchi K., Lamb R.A. Influenza virus M2 protein ion channel activity stabilizes the native form of fowl plague virus hemagglutinin during intracellular transport. Journal of Virology. 1994;68(2):911–919. doi: 10.1128/jvi.68.2.911-919.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takikawa S., Engle R.E., Emerson S.U., Purcell R.H., St. Claire M., Bukh J. Functional analyses of GB virus B p13 protein: Development of a recombinant GB virus B hepatitis virus with a p7 protein. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(9):3345–3350. doi: 10.1073/pnas.0511297103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Techaarpornkul S., Barretto N., Peeples M.E. Functional analysis of recombinant respiratory syncytial virus deletion mutants lacking the small hydrophobic and/or attachment glycoprotein gene. Journal of Virology. 2001;75(15):6825–6834. doi: 10.1128/JVI.75.15.6825-6834.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tedbury P., Welbourn S., Pause A., King B., Griffin S., Harris M. The subcellular localization of the hepatitis C virus non-structural protein NS2 is regulated by an ion channel-independent function of the p7 protein. Journal of General Virology. 2011;92:819–830. doi: 10.1099/vir.0.027441-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teoh K.T., Siu Y.L., Chan W.L., Schluter M.A., Liu C.J., Peiris J.S. The SARS coronavirus E protein interacts with PALS1 and alters tight junction formation and epithelial morphogenesis. Molecular Biology of the Cell. 2010;21(22):3838–3852. doi: 10.1091/mbc.E10-04-0338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terwilliger E.F., Cohen E.A., Lu Y., Sodroski J.G., Haseltine W.A. Functional role of human immunodeficiency virus type 1 vpu. Proceedings of the National Academy of Sciences of the United States of America. 1989;86(13):5163–5167. doi: 10.1073/pnas.86.13.5163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomaston J.L., Nguyen P.A., Brown E.C., Upshur M.A., Wang J., DeGrado W.F., Howard K.P. Detection of drug-induced conformational change of a transmembrane protein in lipid bilayers using site-directed spin labeling. Protein Science. 2013;22(1):65–73. doi: 10.1002/Pro.2186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tokuriki N., Oldfield C.J., Uversky V.N., Berezovsky I.N., Tawfik D.S. Do viral proteins possess unique biophysical features? Trends in Biochemical Sciences. 2009;34(2):53–59. doi: 10.1016/j.tibs.2008.10.009. S0968-0004(08)00253-3 [pii] [DOI] [PubMed] [Google Scholar]
- Torres J., Kukol A., Arkin I.T. Use of a single glycine residue to determine the tilt and orientation of a transmembrane helix. A new structural label for infrared spectroscopy. Biophysical Journal. 2000;79(6):3139–3143. doi: 10.1016/S0006-3495(00)76547-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres J., Maheswari U., Parthasarathy K., Ng L.F., Liu D.X., Gong X.D. Conductance and amantadine binding of a pore formed by a lysine-flanked transmembrane domain of SARS coronavirus envelope protein. Protein Science. 2007;16(9):2065–2071. doi: 10.1110/ps.062730007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres J., Parthasarathy K., Lin X., Saravanan R., Kukol A., Ding X.L. Model of a putative pore: The pentameric α-helical bundle of SARS coronavirus E protein in lipid bilayers. Biophysical Journal. 2006;91(3):938–947. doi: 10.1529/biophysj.105.080119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres J., Surya W., Li Y., Liu D.X. Protein-protein interactions of viroporins in coronaviruses and paramyxoviruses: New targets for antivirals? Viruses. 2015;7(6):2858–2883. doi: 10.3390/v7062750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres J., Wang J., Parthasarathy K., Liu D.X. The transmembrane oligomers of coronavirus protein E. Biophysical Journal. 2005;88(2):1283–1290. doi: 10.1529/biophysj.104.051730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Triantafilou K., Kar S., Vakakis E., Kotecha S., Triantafilou M. Human respiratory syncytial virus viroporin SH: A viral recognition pathway used by the host to signal inflammasome activation. Immunology. 2013;140:87–88. doi: 10.1136/thoraxjnl-2012-202182. [DOI] [PubMed] [Google Scholar]
- Triantafilou K., Kar S., van Kuppeveld F.J., Triantafilou M. Rhinovirus-induced calcium flux triggers NLRP3 and NLRC5 activation in bronchial cells. American Journal of Respiratory Cell and Molecular Biology. 2013;49(6):923–934. doi: 10.1165/rcmb.2013-0032OC. [DOI] [PubMed] [Google Scholar]
- Tung F.Y.T., Abraham S., Sethna M., Hung S.-L., Sethna P., Hogue B.G., Brian D.A. The 9-kDa hydrophobic protein encoded at the 3′ end of the porcine transmissible gastroenteritis coronavirus genome is membrane-associated. Virology. 1992;186(2):676–683. doi: 10.1016/0042-6822(92)90034-M. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Damme N., Goff D., Katsura C., Jorgenson R.L., Mitchell R., Johnson M.C. The interferon-induced protein BST-2 restricts HIV-1 release and is downregulated from the cell surface by the viral Vpu protein. Cell Host & Microbe. 2008;3(4):245–252. doi: 10.1016/j.chom.2008.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Kuppeveld F.J.M., Galama J.M.D., Zoll J., van den Hurk P.J.J.C., Melchers W.J.G. Coxsackie B3 virus protein 2B contains a cationic amphipathic helix that is required for viral RNA replication. Journal of Virology. 1996;70(6):3876–3886. doi: 10.1128/jvi.70.6.3876-3886.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanderlinden E., Naesens L. Emerging antiviral strategies to interfere with influenza virus entry. Medicinal Research Reviews. 2014;34(2):301–339. doi: 10.1002/med.21289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vennema H., Godeke G.J., Rossen J.W., Voorhout W.F., Horzinek M.C., Opstelten D.J., Rottier P.J. Nucleocapsid-independent assembly of coronavirus-like particles by co-expression of viral envelope protein genes. EMBO Journal. 1996;15(8):2020–2028. doi: 10.1002/j.1460-2075.1996.tb00553.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verdia-Baguena C., Nieto-Torres J.L., Alcaraz A., DeDiego M.L., Torres J., Aguilella V.M., Enjuanes L. Coronavirus E protein forms ion channels with functionally and structurally-involved membrane lipids. Virology. 2012;432(2):485–494. doi: 10.1016/j.virol.2012.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkamer A., Kuhn D., Rippmann F., Rarey M. DoGSiteScorer: A web server for automatic binding site prediction, analysis and druggability assessment. Bioinformatics. 2012;28(15):2074–2075. doi: 10.1093/bioinformatics/bts310. [DOI] [PubMed] [Google Scholar]
- Wakita T., Pietschmann T., Kato T., Date T., Miyamoto M., Zhao Z. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nature Medicine. 2005;11(7):791–796. doi: 10.1038/nm1268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walensky L.D., Bird G.H. Hydrocarbon-stapled peptides: Principles, practice, and progress. Journal of Medicinal Chemistry. 2014;57(15):6275–6288. doi: 10.1021/Jm4011675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Cady S.D., Balannik V., Pinto L.H., DeGrado W.F., Hong M. Discovery of spiro-piperidine inhibitors and their modulation of the dynamics of the M2 proton channel from influenza a virus. Journal of the American Chemical Society. 2009;131(23):8066–8076. doi: 10.1021/ja900063s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Ma C., Balannik V., Pinto L.H., Lamb R.A., Degrado W.F. Exploring the requirements for the hydrophobic scaffold and polar amine in inhibitors of M2 from influenza A virus. ACS Medicinal Chemistry Letters. 2011;2(4):307–312. doi: 10.1021/ml100297w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Ma C., Fiorin G., Carnevale V., Wang T., Hu F. Molecular dynamics simulation directed rational design of inhibitors targeting drug-resistant mutants of influenza A virus M2. Journal of the American Chemical Society. 2011;133(32):12834–12841. doi: 10.1021/ja204969m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J.Z., Ma C.L., Wang J., Jo H., Canturk B., Fiorin G. Discovery of novel dual inhibitors of the wild-type and the most prevalent drug-resistant mutant, S31N, of the M2 proton channel from influenza a virus. Journal of Medicinal Chemistry. 2013;56(7):2804–2812. doi: 10.1021/jm301538e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Pielak R.M., McClintock M.A., Chou J.J. Solution structure and functional analysis of the influenza B proton channel. Nature Structural & Molecular Biology. 2009;16(12):1267–1271. doi: 10.1038/nsmb.1707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Qiu J.X., Soto C., Degrado W.F. Structural and dynamic mechanisms for the function and inhibition of the M2 proton channel from influenza A virus. Current Opinion in Structural Biology. 2011;21(1):68–80. doi: 10.1016/j.sbi.2010.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Wu Y.B., Ma C.L., Fiorin G., Wang J.Z., Pinto L.H. Structure and inhibition of the drug-resistant S31N mutant of the M2 ion channel of influenza A virus. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(4):1315–1320. doi: 10.1073/pnas.1216526110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welch W.J., Sefton B.M. Characterization of a small, nonstructural viral polypeptide present late during infection of BHK cells by Semliki Forest virus. Journal of Virology. 1980;33(1):230–237. doi: 10.1128/jvi.33.1.230-237.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitehead S.S., Bukreyev A., Teng M.N., Firestone C.Y., St Claire M., Elkins W.R. Recombinant respiratory syncytial virus bearing a deletion of either the NS2 or SH gene is attenuated in chimpanzees. Journal of Virology. 1999;73(4):3438–3442. doi: 10.1128/jvi.73.4.3438-3442.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson R.L., Fuentes S.M., Wang P., Taddeo E.C., Klatt A., Henderson A.J., He B. Function of small hydrophobic proteins of paramyxovirus. Journal of Virology. 2006;80(4):1700–1709. doi: 10.1128/JVI.80.4.1700-1709.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson L., Gage P., Ewart G. Hexamethylene amiloride blocks E protein ion channels and inhibits coronavirus replication. Virology. 2006;353(2):294–306. doi: 10.1016/j.virol.2006.05.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wingfield W.L., Pollack D., Grunert R.R. Therapeutic efficacy of amantadine HCl and rimantadine HCl in naturally occurring influenza A2 respiratory illness in man. The New England Journal of Medicine. 1969;281(11):579–584. doi: 10.1056/NEJM196909112811102. [DOI] [PubMed] [Google Scholar]
- Wozniak A.L., Griffin S., Rowlands D., Harris M., Yi M., Lemon S.M., Weinman S.A. Intracellular proton conductance of the hepatitis C virus p7 protein and its contribution to infectious virus production. PLoS Pathogens. 2010;6(9) doi: 10.1371/journal.ppat.1001087. e1001087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu Y., Canturk B., Jo H., Ma C., Gianti E., Klein M.L. Flipping in the pore: Discovery of dual inhibitors that bind in different orientations to the wild-type versus the amantadine-resistant s31n mutant of the influenza a virus m2 proton channel. Journal of the American Chemical Society. 2014;136(52):17987–17995. doi: 10.1021/ja508461m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yi M., Ma Y., Yates J., Lemon S.M. Compensatory mutations in E1, p7, NS2, and NS3 enhance yields of cell culture-infectious intergenotypic chirneric hepatitis C virus. Journal of Virology. 2007;81(2):629–638. doi: 10.1128/JVI.01890-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu X., Bi W., Weiss S.R., Leibowitz J.L. Mouse hepatitis virus gene 5b protein is a new virion envelope protein. Virology. 1994;202(2):1018–1023. doi: 10.1006/viro.1994.1430. [DOI] [PubMed] [Google Scholar]
- Zhang W., Yang H., Kong X., Mohapatra S., San Juan-Vergara H., Hellermann G. Inhibition of respiratory syncytial virus infection with intranasal siRNA nanoparticles targeting the viral NS1 gene (vol 11, pg 56, 2005) Nature Medicine. 2005;11(2):233. doi: 10.1038/Nm0205-233b. [DOI] [PubMed] [Google Scholar]
- Zhao X., Singh M., Malashkevich V.N., Kim P.S. Structural characterization of the human respiratory syncytial virus fusion protein core 2. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(26):14172–14177. doi: 10.1073/pnas.260499197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou H.X., Cross T.A. Modeling the membrane environment has implications for membrane protein structure and function: Influenza A M2 protein. Protein Science. 2013;22(4):381–394. doi: 10.1002/Pro.2232. [DOI] [PMC free article] [PubMed] [Google Scholar]