

ABSTRACT
Introduction: Influenza virus, human respiratory syncytial virus (RSV), and human metapneumovirus (HMPV) are important human respiratory pathogens. Recombinant virus-like particle (VLP) vaccines are suggested to be potential promising platforms to protect against these respiratory viruses. This review updates important progress in the development of VLP vaccines against respiratory viruses.
Areas Covered: This review summarizes progress in developing VLP and nanoparticle-based vaccines against influenza virus, RSV, and HMPV. The PubMed was mainly used to search for important research articles published since 2010 although earlier key articles were also referenced. The research area covered includes VLP and nanoparticle platform vaccines against seasonal, pandemic, and avian influenza viruses as well as RSV and HMPV respiratory viruses. The production methods, immunogenic properties, and vaccine efficacy of respiratory VLP vaccines in preclinical animal models and clinical studies were reviewed in this article.
Expert opinion: Previous and current preclinical and clinical studies suggest that recombinant VLP and nanoparticle vaccines are expected to be developed as promising alternative platforms against respiratory viruses in future. Therefore, continued research efforts are warranted.
KEYWORDS: Influenza virus, Virus-like particles, Vaccines, Protection, Respiratory syncytial virus, Human metapneumovirus virus
1. Introduction
Respiratory viruses cause infections in the upper or lower respiratory tracts. Viral infections in the lower respiratory tracts can cause several respiratory syndromes such as bronchitis, pneumonia, and bronchiolitis [1]. In particular, children and the elderly are greatly affected. There are several important viruses that infect epithelial cells in the human respiratory tracts: respiratory syncytial virus (RSV); influenza A and B viruses; parainfluenza 1, 2, and 3; and adenovirus [2]. Other viruses infecting human respiratory tracts include rhinovirus, human metapneumovirus (HMPV), severe acute respiratory syndrome coronavirus, human coronavirus NL63 and HKU1, parainfluenza 4, and bocavirus [1,3].
Among the respiratory viruses, influenza is the leading cause of respiratory illness. Globally 5–15% of human populations are affected by influenza virus annually. According to the report of World Health Organization, every year 250,000 to 500,000 deaths occur due to the infection of influenza virus [4]. Different platforms for influenza vaccines have been available on the market, such as inactivated split vaccine, live attenuated influenza vaccines (LAIV) and recombinant purified subunit protein vaccines [5,6].
Human respiratory syncytial virus (RSV) causes respiratory tract infections in infants and young children worldwide, leading acute bronchiolitis and viral pneumonia [7]. An extensive epidemic study indicates that approximately 45% of hospital admissions and in-hospital deaths are due to RSV-induced acute lower respiratory infection in children younger than 6 months [8]. Thirty-three million cases worldwide in children under at age 5 are estimated to be infected with RSV-associated acute lower respiratory infections and about 10% of patients were being hospitalized, resulted in about 1-3% of in-hospital deaths [8]. Palivizumab, the licensed monoclonal antibody (Ab), has been used as a prophylactic drug to prevent severe RSV disease in high-risk children [9]. Since palivizumab has safety and efficacy concerns such as anaphylaxis and hypersensitivity reactions, palivizumab is not recommended for the therapeutic-treatment post-RSV infection [10]. There are neither effective therapies nor any licensed vaccines commercially available in the market against RSV so far.
Another important respiratory virus is human metapneumovirus (HMPV) which is also a leading cause of acute lower respiratory tract infection globally. Pediatric population and immunocompromised patients are greatly susceptible to HMPV, causing substantial morbidity and mortality worldwide [11,12]. However, there is currently no HMPV-specific vaccine available.
Virus-like particles (VLPs) are developed and reported, suggesting effective vaccine platforms against respiratory viruses such as influenza virus, RSV, and HMPV infections (Figure 1) [13–16]. VLPs are non-replicating as well as noninfectious because they do not contain viral genetic materials. Viral structural proteins such as envelope viral glycoproteins and capsid core proteins can be expressed and self-assembled into a particulate morphology of VLPs in cell cultures in vitro. VLPs can be produced in bacteria, mammalian cell lines, insect cell lines, yeast, and plant cells [17]. VLPs produced by recombinant baculovirus – derived constructs exhibited higher and broader immune responses compared to those by mammalian – expressed VLP vaccines [18]. It is possible that residual baculoviruses in VLP preparations activates innate immune response at the site of inoculation. VLPs provide an immunogenic platform effectively eliciting T cell and B cell immune responses due to repetitive and high density of viral proteins displayed on the surfaces of VLPs [19,20].
Figure 1.
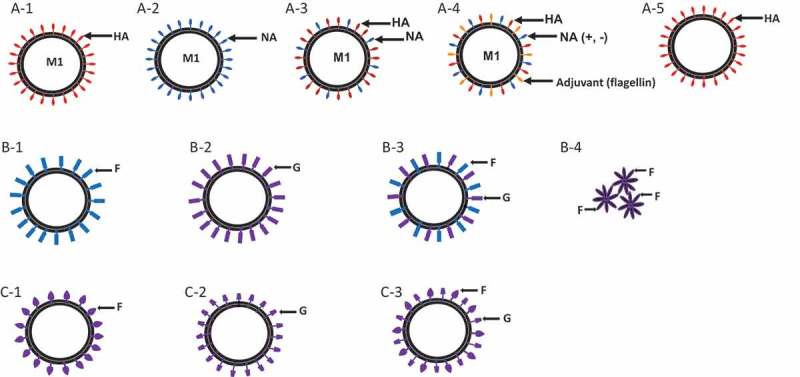
Diagrams for influenza VLPs, RSV VLPs, and HMPV VLPs.
(a) Influenza VLPs expressing hemagglutinin (HA) (A-1), neuraminidase (NA)(A-2), both HA and NA (A-3), HA, NA together with adjuvant flagellin (A-4), or plant-made VLPs containing HA (A-5). Influenza matrix protein (M1) was used as a core protein. Influenza VLPs are generated by insect/rBV system, mammalian cells or plant – derived. Plant-derived VLPs are produced in Nicotiana benthamiana infiltrated in batches with an Agrobacterium inoculum containing influenza antigen expression cassette. (b) RSV VLPs and nanoparticle by insect/rBV system. VLPs expressing RSV fusion protein (F) (B-1), RSV glycoprotein (G) (B-2) or both F and G (B-3) on influenza M1 core protein. RSV F nanoparticle (B-4) extracted and purified from insect cell membranes composed of multiple RSV F oligomers arranged in the form of rosettes. (c) HMPV VLPs: HMPV VLPs derived from retroviral core particles are generated in human embryonic kidney epithelial (293-f) cells by expressing fusion protein (F) (C-1), glycoprotein (G) (C-2) or both F and G (C-3) proteins.
In this review, we attempted to cover the production, immunogenic properties, and vaccine efficacy of VLP vaccines, from preclinical and clinical studies reporting on respiratory viruses of influenza viruses, RSV and HMPV.
2. Influenza VLP vaccines
2.1. Production and immunogenicity of influenza VLP vaccines targeting seasonal influenza viruses
Different approaches have been reported to generate influenza VLP vaccines. Influenza VLPs can be composed of various structural influenza proteins, such as hemagglutinin (HA), neuraminidase (NA), matrix (M1) and M2 proteins Figure 1(a). VLPs containing four structural proteins derived from influenza virus A/Udorn/72 (H3N2) were produced in insect cells via the recombinant baculovirus (rBV) expression system [21]. Three HA, NA, and M1 protein-encoding genes cloned into a single baculovirus construct were expressed for the generation of influenza virus VLPs [22–24] (Table 1). Also, individual rBVs expressing HA and M1 were co-infected together into insect cells to produce influenza VLPs [16,25,26]. There are several advantages of using the baculovirus expression vector system (BEVS). Proteins expressed in BEVS are soluble and functionally active. Other advantages of BEVS include post-translational modifications and higher yields for secreted proteins.
Table 1.
Seasonal influenza VLP vaccines in animal models.
| Source | Component | Protection determined by mouse survival (%) | Host | Reference |
|---|---|---|---|---|
| H1N1 | HA+NA+M1 | Homologous virus (NC/99, H1N1) A/Mexico/4482/09 |
Mice, Ferrets | [53,55] |
| NA+B gag | Not available | NA | [62] | |
| HA | Not available | Mice | [45] | |
| HA+M1 | 100%, A/California/04/2009 80%, A/PR/8/1934 |
Mice | [54] | |
| NA+M1 | 100%, homologous virus (A/PR/8/34); 100%, heterosubtypic virus (A/Phil/82) |
Mice | [52] | |
| HA+M1+ flagelin | (1)100%, homologous virus (A/PR/8/34); 67%, heterosubtypic virus (A/Phil/82) (2)100%, homologous (A/PR/8/34) 100%, heterosubtypic virus (A/Phil/82) |
Mice | [51,56] | |
| M1+ M2e+ Flagellin | A/Philippine/2/82 A/PR/8/34 |
Mice | [77,78] | |
| HA+M1+ CT, alum, CpG, MPL | 100%, homologous virus (A/PR/8/34) | Mice | [89] | |
| HA+NA+M1+ M2/CpG | Homologous virus (1918 influenza A) | Mice | [27] | |
| M2 (A/WSN/33)/inactivated PR8 | 2009 H1N1, H3N2, H5N1 | Mice | [58] | |
| H3N2 | HA+M1 HA+M1/IL-12 |
(1) Not available (2) A/HK (H3N2) (3) NY/04 (H3N2) (4) A/Hong Kong/68 (H3N2) |
(1) Dog (2) Mice, Ferrets |
[25,28,29,53] |
| HA (LAH)+ HBc | Not available | Mice | [85] | |
| HA+NA+M1 | Not available | Mice, Ferrets | [22] | |
| HA+M1/GPI-CCL28 | 20%, A/Philippines/2/1982 60%, A/Aichi/2/1968 |
Mice | [84] | |
| H1, H3 | HA+M1 | 100%, homologous (A/PR/8/34, A/Aichi/2/68), 100%, heterologous (A/WSN/33) |
Mice | [16] |
| H1N1, H3N2, Influenza B | HA+NA+M1 | A/New Caledonia/20/1999 A/New York/55/2004 |
Mice, Ferrets | [53] |
| H1N1 | HA (plant-made) | Stable, interact and activate antigen-presenting cells | Mice | [44,45] |
| H1N1, H3N2, | HA (plant-made) | Proper antigenicity (E+, HI+, VN+, MN+) | Mice, chicken, ferrets | [43] |
E: ELISA; VN: virus neutralization; MN: virus microneutralization; HI: hemagglutination inhibition.
Mammalian cell-expressed VLPs were reported to be produced in human embryonic kidney 293T cells with transfection of DNA plasmids expressing influenza surface proteins and core matrix proteins [30] (Table 2). In brief, mammalian cells were co-transfected with three different cloned recombinant plasmids such as M1, HA, and NA, respectively, M1 from A/Puerto Rico/8/1934 strain, NA from A/Thailand/1(KAN-1)/2004 and codon-optimized HA protein from clade 2 H5N1.
Table 2.
Avian influenza VLP vaccines in animal models.
| H5N1 | HA+NA+M1 | 100%, homologous and heterologous virus (A/Viet Nam/1203/2004, A/Indonesia/05/2005) 100%, homologous virus (A/chicken/Korea/ES/2003, H5N1) |
Mice Ferrets Chicken |
[31–33] |
| HA + NA+ Bgag/M8 HA + NA+ Bgag/Alum HA + NA+ Bgag/Addavax HA + NA+ Bgag/Poly(I:C) |
100%, rgH5N1 (A/Vietnam/1203/2004, A/Thailand/1(KAN-1)/2004,A/Puerto Rico/8/1934) | Mice | [87] | |
| HA+M1 HA |
100%, homologous virus (A/Viet Nam/1203/2004) H5N1 HPAI H5N1 HPAI |
Mice Chicken |
[34,59] | |
| HA+NA+M1+ M2 | 100%, homologous virus (A/Hanoi/30,408/2005, H5N1) | Mice | [35] | |
| COBRA HA+M1 COBRA HA+M1/Alum |
A/Whooper Swan/Mongolia/244/2005 | Mice Ferrets |
[30] | |
| HA+NA+Gag | homologous virus (A/Viet Nam/1203/04) and heterologous virus(A/Indonesia/5/05) | Mice | [36] | |
| HA+NA+M1/CFA | 100%, A/meerkat/Shanghai/SH-1/2012 (clade2.3.2.1) (H5N1) | Mice | [37] | |
| HA+NA+M1/NP | With NP: 100%, heterologous virus A/goose/GD/1996, H5N1, clade 2.3.4 Without NP: 50%, heterologous virus clade 2.3.4 |
Chicken | [38] | |
| H7N9 | HA+NA+M1+ Iscomatrix | NA, A/Shanghai/1/2013 | Human Ferrets |
[24,39,40,60,80] |
| HA+NA+M1+ Iscomatrix | Homologous A/Anhui/1/2013 | |||
| Plant derived-HA-VLP Plant derived-HA+ alum |
100%, A/Anhui/1/2013 | Mice Ferrets |
||
| HA+NA+M1/Novasome rH9/Novasome |
A/Hong Kong/1073/99 (H9N2) | Rat Ferrets Mice |
||
| TM of H3N2 HA+H7-HA+M1 | A/Chicken/Guangdong/53/2014 A/Chicken/Guangdong/MCX/2014 A/Chicken/Guangdong/ZSM/2017 |
Mice | ||
| H7N3, H7N9, H5N1 | HA+ NA+M1+ Iscomatrix | 100%, A/Anhui/1/2013 | Mice | [81] |
| H9N2 | HA+M1/ISA70 HA+NA/Freund’s HA+NA+M1/Novasome HA+NA+M1 |
NA, homologous H9N2 (A/Chicken/Korea/01310/2001) NA NA, homologous H9N2 (A/Hong Kong/1073/99) |
Chicken chickens Mice Ferrets |
[23,41,88] |
| H1, H5 | M2e5x+M1 Split vaccine (prime) + M2e5x VLP (Boost) |
100%, homologous (A/Viet Nam/1203/2004) Not protected 100%, A/Philippine/2/81 (H3N2), rgH5N1 A/Brisbane/59/2007 (BR/59) |
Mice Chicken Mice Ferrets |
[42,64,66,68,69] |
| M2e5x+HA+M1 | 100%, homologous virus (A/Viet Nam/1203/2004) | Mice | [42,78] | |
| M2e5x+ Flagellin | 100%, A/Philippines/2/82 | Mice | ||
| M2e5x+M1 | A/Philippines/2/82 A/California/4/2009 rgH5N1 |
Mice | [63,64,67] | |
| LAIV+M2e5xVLP | rgH5N1 A/PR/8/34 A/Philippines/2/1982 |
Mice | [70] | |
| HA+NA+M1+ M2 | Not available | Mice | [76] | |
| HA+M1/GPI-ISM | 100%, heterologous A/Vietnam/1203/2004 | Mice | [83] | |
| H1N1, H3N8, H5N1, H7N3 | HA+M1 | 100%, H1N1, H2N1, H7N9, H7N1, H10N1, H11N1 90%, H5N1, 88.3%, H10N1 |
Mice | [50] |
| H1N1, H3N2 | HA+M1 | 100%, H1N1 A/PR/8/34, H3N2 A/Aichi/2/68, H1N1 A/WSN/33 | Mice | [47] |
| H5N8 | HA+NA | Not available | Mice | [26] |
| H5/H7/H9/H10 | HA+NA+ gag (immunodeficiency virus gag protein) | Not available | NA | [62] |
| H5, H7 | HA+ DNA plasmid | A/WSN/1933, A/California/07/2009 (CA09), A/Cambodia/P0322095/2005 (CAM05), A/swine/Jiangxi/1/2004, A/chicken/Jiangsu/7/2002, A/Hong Kong/1/1968, A/Guizhou/54/1989, A/Netherlands/219/2003 |
Mice | [61] |
| H5 | HA (plant-made) | Interact and activate antigen-presenting cells | Mice | [44] |
Plant cells are being used for expression and production of recombinant HA influenza VLP vaccines, which are currently under clinical trials to evaluate the immunogenicity, safety, and efficacy [43] (Tables 1 and 2). Various forms of recombinant HA, including monomers, trimers, virus-like particles or chimeric proteins were obtained by plant-based technology. Nicotiana benthamiana plants are vacuum infiltrated in batches with an Agrobacterium inoculum containing an HA expression cassette. VLPs obtained by harvesting plant cells, homogenizing, and purification. Plant-made VLPs showed morphological stable structures over time, eliciting both humoral and cellular responses in mice, ferrets, rabbits, or chickens [43–45] (Tables 1, 2, 4). A plant-derived H1-VLP vaccine was reported to induce significantly higher levels of IgG antibodies, CD4+, and CD8+ T cell responses compared to inactivated virus vaccine in aged mice [45].
Table 4.
Clinical trial studies.
| Source | Component | Protective immunity | Host | Reference |
|---|---|---|---|---|
| H1N1, H5N1 | HA+NA+M1 | Neutralizing antibody induced and 79% sero-protection rate achieved. | 18–64 years old | [125] |
| H5N1 | HA+NA+M1 | 61% seroconversion rate based on HAI and 76% seroconversion rate based on MN. | 18 to 40 years old | [126] |
| H7N9 | HA VLP+ ISCOMATRIX | Neutralizing antibody induced against homologous strain and the heterologous H7-A/Netherlands/219/03 strain | [80] | |
| H1N1 | gH1-Qbeta/alhydrogel | 65.4% seroconversion in adjuvanted group and 65.4% seroconversion innon-adjuvanted group. | 21 and 64 years old | [130] |
| H5N1 | HA | HI+, MN+ | 18-60 years old | [43,45] |
| RSV-A2 | RSV F protein nanoparticle | Neutralizing antibody induced | Older adult | [137] |
| SV-A2 | RSV F protein nanoparticle | Neutralizing antibody induced | Healthy women | [133] |
MN: virus microneutralization; HI: hemagglutination inhibition.
Taken together, influenza VLPs can be composed of HA, NA, or M1 either by containing all of these three proteins or two proteins HA or NA with M1. These VLP vaccines containing structural proteins from seasonal influenza virus H1N1 or H3N2 subtypes were immunogenic inducing both humoral antibodies and cellular immune responses [46].
Studies have reported that H1N1 or H3N2 influenza VLPs could provide protection against homologous and heterologous protection with a different degree of efficacy depending on antigenic closeness [16,17,22,47–52] (Table 1). The rBV expression system produced seasonal trivalent VLP vaccines derived from influenza A virus such as A/New Caledonia/20/1999 H1N1, A/New York/55/2004 H3N2, and influenza B virus B/Shanghai/367/2002 were able to elicit substantial levels of hemagglutination inhibition (HAI) titers against homologous and heterologous virus challenge in mouse and ferret models [53] (Table 1). In this study, mice were immunized with 3 µg, 6 µg, or 12 µg of VLP and ferrets immunized with 15 µg, 3 µg, or 0.6 µg [53]. Trivalent VLP vaccines were found to elicit higher levels of influenza-specific CD8+ T cell responses in mice than those with commercial egg-derived trivalent-inactivated vaccine (TIV). H1N1 or H3N2 influenza VLPs vaccinations provided higher vaccine efficacies against homologous or heterologous virus challenge infections compared to that against heterosubtypic influenza virus, showing less body weight loss (0–12%) in animal models after challenge with homologous or heterologous virus [25,50–52,54–56]. These results indicate that the efficacies of influenza VLP vaccines are encouraging in preclinical studies as an alternative vaccine platform. However, protective efficacies vary with diverse VLP vaccines containing different antigenic components and contents. Most efficacy studies were reported in in-bred mouse models, which might not predict what will happen in humans after vaccination. Influenza VLP vaccines need to be better characterized, tested in out-bred, more relevant animal models, and developed for improved cross-protective immunity for consideration of future vaccines.
2.2. Influenza VLP vaccine against potential pandemic influenza virus
An influenza pandemic is an outbreak of a novel influenza virus that can spread worldwide. During the 2009 influenza pandemic, we observed significant shortages and delays of egg-based vaccines in the global supply [5]. Technology of non-replicating VLPs represents an alternative option since it is immunogenic, safe, and independent of egg supplies. Previous studies reported influenza VLPs containing the HA, NA, and M2 surface proteins from highly pathogenic avian influenza viruses: A/Chicken/FPV/Rostock/1934 (H7N1) and A/Thailand/KAN-1/04 (H5N1), inducing high titers of neutralizing antibodies in mice [57]. Influenza 2009 H1N1 VLPs and H5N1 VLPs generated by using the rBV expression system-induced encouraging protective immunity in mice and ferrets [54,58,59] (Tables 1 and 2). H7 HA VLPs assembled with HA of wild-type (HA-WT) or HA transmembrane (TM) domain replaced by that from H3N2 subtype (HA-TM) were generated [60]. H7 VLPs-TM induced better protection against homologous and heterologous H7N9 viruses challenge, as observed with less weight loss and higher survival rates compared to H7 VLPs-WT [60]. A bivalent heterologous DNA and VLP prime-boost vaccine was reported to provide protection against H1, H5, H9, and H3 and H7 viruses [61]. Mice were primed with DNA plasmid twice at weeks 0 and 3 and boosted with VLP. The immune sera from vaccinated mice were able to neutralize homologous, heterologous (H5, H7) and heterosubtypic H1 virus after passive transfer [61]. Irina Tretyakova and her colleague reported a quadrivalent pseudotype influenza VLP vaccine containing four different types of HA (H5N1, H7N9, H9N2, and H10N8) proteins, NA and bovine immunodeficiency virus gag protein (B gag) as an alternative to M1 matrix protein in insect cells [62] (Table 2). Electron microscopic observation results demonstrated that average sizes of the B gag core protein assembled VLPs are in a range of 150 nm to 200 nm, which was larger than the influenza virus M1 core protein assembled VLPs. These results indicate that vaccine efficacies from avian influenza VLPs are encouraging. B gag core protein assembled VLPs might incorporate more spike antigens than the M1 assembled VLPs.
2.3. Universal influenza VLP vaccines
Influenza virus surface antigenic proteins HA and NA change over time and result in generating novel viruses that are antigenically different to become new HA and/or new HA and NA proteins in influenza viruses. New research efforts to develop universal influenza VLPs containing HA, NA, and M2 proteins are going on for broader immunity.M2e as an extracellular domain of M2 is conserved among influenza A viruses and has been used for cross-protective antigenic target [63]. The cross-protective efficacy was observed after vaccination of mice with M2e5x VLP vaccines containing heterologous tandem M2e repeats (M2e5x) derived from human, swine, and avian influenza viruses [63]. This heterologous tandem M2e5x repeat construct contains multiple M2e conserved epitopes to increase the density of M2e epitopes. A comparative study of vaccine efficacy has been conducted in mice between M2e5x VLP and wild-type M2 expressed VLP. This study demonstrated that M2e5x VLP immunization induced 100% protection against A/California/4/2009 (H1N1), A/PR/8/34 (H1N1), A/Philippines/2/82 (H3N2), and A/Vietnam/1203/2004 (H5N1). However, wild-type M2 VLP-vaccinated mice showed a significant loss in body weight with a substantial delay in recovering body weight [64] (Table 2). M2e5x VLP vaccination could induce long-lasting cross-protection against A/PR/8/34 (H1N1) and reassortant A/Vietnam/1203/2004 (H5N1) virus by passively transferring immune sera from vaccinated mice to the naïve mice [64]. Another experimental data demonstrated that M2e5x VLP vaccine has the ability to show more cross-protective efficacy in BALB/c mice compared to C57BL/6 mice [63,65], suggesting that immunogenicity and efficacy are dependent on host strains. The efficacy of cross-protection by M2e VLP immunity is mediated by weak non-neutralizing immune responses [66–68]. Supplemented HA-based influenza vaccination with M2e5x VLP vaccines could significantly enhance cross-protection in mice, chicken, and ferret [66,69,70]. These results indicate that higher vaccine efficacy or cross-protection induced by M2e VLPs can be induced by supplemented influenza vaccination.
2.4. Chimeric influenza or adjuvanted VLPs vaccines
Different adjuvants were tested in the influenza vaccination with M2e epitopes, which include Freund’s adjuvant, monophosphoryl lipid A, cholera toxin subunits, heat-labile endotoxin, flagellin, and profilin, providing certain levels of broader and enhanced cross-protection [71–78]. ISCOMATRIX is composed of purified fractions of Quillaia saponaria extract, cholesterol, and phospholipid [79] and was tested with influenza VLP vaccine [80]. A/Anhui/1/2013 (H7N9) based HA-NA-M1 VLP vaccine with ISCOMATRIX adjuvant was shown to induce 3-4-fold higher levels of serum HAI and NA antibody titers and to provide better protection in mice against heterosubtypic A/Shanghai/1/2013 (H7H9), A/Chicken/Jalisco/CPA/2012 (H7N3) virus and heterologous A/Indonesia/05/2005 (H5N1) virus, compared to HA-NA-M1 VLP only vaccination [81].
Influenza VLPs were engineered to express molecular adjuvants or immunostimulatory molecules on the surfaces of VLPs. A flagellin molecular adjuvant was engineered to be incorporated into influenza VLP vaccines via a membrane-anchoring domain [51,56] (Figure 1(a) A-4). Flagellin-containing influenza VLPs were produced in insect Sf9 cells by co-infecting with rBVs expressing PR8 HA, M1, and membrane-anchored flagellin. This chimeric flagellin plus HA VLP vaccination-induced heterosubtypic protection against challenge with A/Philippines (H3N2) virus [51]. Also, flagellin VLP was found to play an effective adjuvant role in promoting IgG isotype-switched long-lasting antibody induction and protection of influenza vaccines in a CD4-deficient condition in mice [82].
Using a protein transfer technology, glycosylphosphatidylinositol – anchored immunostimulatory molecules (GPI-ISMs) are incorporated onto influenza VLPs [83]. Mice immunized with VLPs modified by protein transfer with GPI-GM-CSF induced higher levels of IgG antibody responses and protection against a heterologous influenza virus challenge than unmodified VLPs in mice [83]. Intranasal immunization with chimeric VLPs containing influenza HA antigen and GPI-CCL28 was shown to induce long-lasting mucosal immunity against H3N2 virus infection [84]. GPI-anchored CCL-28 chemokine co-expressed on the surface of influenza HA VLPs was shown to be effective in reducing lung viral titers after virus challenge in vaccinated mice. Vaccination of mice with influenza HA VLPs containing GPI-anchored CCL-28 enhanced lung IgA and serum IgG antibody responses, compared to the standard influenza HA VLP vaccine.
Yeast expression system rather than using the bacterial expression system has been used to avoid bacterial endotoxins [85]. In this study, a long alpha helix domain (55 amino acid residues) from the conserved HA stalk region was genetically fused to the hepatitis B virus (HBV) core protein and expressed via the yeast expression system [85]. The hydrophobic stalk domain fragment was presented in immunogenic spikes on the surfaces of self-assembled HBV core protein VLP [85]. This chimeric HA stalk – HBV core VLPs produced in yeast cells were immunogenic, inducing IgG antibody responses to both group 1 and group 2 HA proteins in mice after vaccination in combination with adjuvant CpG (ODN 2395) [85].
M8 as a sequence-optimized 5′-triphosphate-containing RNA (5′pppRNA) RIG-I agonist has been used as adjuvant to enhance influenza VLP efficacy [86]. Immunization with a novel M8 adjuvanted VLPs (M8-VLP) containing H5N1 influenza virus HA and NA induced humoral and cellular mediated protective responses and protection against influenza virus challenge [87]. Intramuscular immunization with M8-VLP induced long-lasting protective antibody responses against influenza virus in mice. Intramuscular immunization with Freund’s adjuvanted H9N2 influenza VLP containing HA and NA induced high levels of HAI titers with significant reduction in virus shedding and substantial homologous protection in chickens [88]. Adjuvants Alum, CpG DNA, monophosphoryl lipid A (MPL), poly IC, gardiquimod, and cholera toxin (CT) have been used to assess immunity in combination with influenza VLPs in mice [89]. Alum, CpG, MPL, or CT adjuvanted VLPs showed higher levels of antibodies in both sera and mucosa. Alum, MPL, or CT adjuvanted VLPs showed higher HAI and virus-neutralizing activities compared to a non – adjuvanted control.
These studies support the concept that chimeric VLPs or adjuvanted influenza VLP vaccines can be more effective in inducing antibody responses and protection than standard VLP controls. However, most of these chimeric VLPs or adjuvanted influenza VLP vaccines were tested in mouse models and remain unclear for efficacy and safety in translational application to humans.
3. RSV VLP vaccines
3.1. RSV VLP vaccine components and platforms
Human respiratory syncytial virus (RSV) belongs to the pneumoviridae family, containing fusion protein (F) and attachment (G) glycoprotein on the surface of the virion, which are directed toward outside from the virion membrane. The F glycoproteins are relatively conserved among the different RSV A and B strains. RSV F and G glycoproteins contain B cell and T cell epitopes to generate a protective-immune response in hosts after natural infection [90–92]. Although RSV contains another surface glycoprotein, SH glycoprotein, but naturally it is weakly immunogenic [93]. RSV VLP vaccines have been generated by expressing RSV G or F glycoproteins. As seen in Figure 1(b) and Table 3, Newcastle diseases virus (NDV) VLPs containing the ectodomain of RSV G and F proteins have been generated. Recombinant chimeric RSV-NDV constructs of RSV G and F protein fused to the cytoplasmic tail (CT) and the transmembrane (TM) domain of NDV hemagglutinin-neuraminidase (HN) and F protein were generated to produce RSV VLP vaccines based on NDV structural proteins [94,95]. Interaction between the TM-CT of chimeric RSV-NDV glycoproteins and NDV – nucleoprotein (NP) and matrix M protein was necessary for efficient production of chimeric RSV-NDV VLP vaccines in avian cells by DNA transfection [95,96]. Both chimeric F(F/F) and G (H/G) proteins were together co-presented on the surface of NDV-VLP with the help of NDV structural proteins for VLP assembly and budding [94]. Chimeric RSV – NDV VLP vaccines containing RSV G or RSV G + F proteins were immunogenic and prevented the replication of RSV in mice without causing vaccine-enhanced RSV lung histopathology [94,95].
Table 3.
Pneumovirus VLP vaccines in animal models.
| Source | Component | Protective immunity | Host | Reference |
|---|---|---|---|---|
| RSV A2 | RSV G/NDV (NP, M) | Lung virus replication inhibited, neutralizing antibody induced, no immunopathology | Mice | [94,95] |
| RSV F DNA + F VLP + G VLP (FFG-VLP) | Lung viral replication below the detection limit, neutralizing activity, ASC response, no weight loss, pulmonary inflammation prevented | Cotton rats | [118] | |
| RSV F + G + DNA-F Influenza M1 (FFG-VLP) | Clearance of lung virus loads, the absence of eosinophil infiltrates, no inflammatory cytokine-secreting cells | Mice | [112,113] | |
| RSV-G + F + M/alum/MPLA | Neutralizing antibody response, virus clearance | Cotton rats | [102] | |
| hMPV-M + RSV Pre-F/F + RSV Post F/F + Alum | Prevented detectable viral replication, Th1-mediated immune response, no severe bronchiolitis, neutralizing antibody elicited | Mice | [110] | |
| RSV F/influenza M1 | Least weight loss, no sign of histopathology and eosinophilia IgG2a antibody response, Th1 immune response |
Mice | [109,114,115] | |
| RSV G (1-780bp, 450-780bp)/influenza M1 | Lung virus replication inhibited, no enhanced inflammatory response | Mice | [105] | |
| RSV G, RSV F/M1 | IgG2a dominant antibody response, lung virus loads decreased | Mice | [101] | |
| RSV G + RSV F/M1 | Mixed VLP F+ VLP G provided a high level of protection, no vaccine-induced immunopathology | Mice | [111] | |
| RSV F/M1 RSV G/M1 RSVFG/M1 |
FG VLP and F VLP: no pulmonary inflammation, G VLP: moderate lung inflammation, F VLP and FG VLP: antibody secreting cell responses |
Cotton rat | [117] | |
| NDV-RSV H/G+ Pre- F/F, H/G+ post-F/F | Lung virus replication inhibited, neutralizing antibody induced | Mice | [97] | |
| H/G + RSV pre-F + NDV-NP + NDV-M,H/G + post-F + NDV-NP + NDV-M | Neutralizing antibody induced, pulmonary inflammation reduced | Cotton rat | [99,120] | |
| RSV F nanoparticle | Neutralizing antibodies induced, virus replication inhibited, no sign of disease enhancement | Cotton rat | [106] | |
| HMPV | HMPV F, G | Homologous and heterologous virus, neutralizing antibody induced, lung virus titer reduced | Mice | [13] |
| HMPV F, G, M | NA | NA | [123] | |
| HMPV F, M | Neutralizing antibody induced, lung virus titer reduced | Mice | [14] | |
| HMPV F, M | Lung virus replication inhibited | Mice | [124] |
RSV F proteins have two forms of conformation, one is a metastable pre-fusion F protein conformation and another is a stable post-fusion F protein form. A pre-fusion form of RSV-F protein contains potent neutralizing epitopes of sites Ø and V, which are not exposed in the post-fusion form [90,98]. Pre-fusion F conformation-stabilized protein was reported by introducing pre-fusion stabilizing mutations (DS-Cav1) of 2 cystine disulfide bond formation and 2 point mutations of cavity filling as well as a foldon oligomer stabilizing domain [90]. For presentation of pre-fusion F protein on the surfaces of VLPs, the pre-fusion stabilizing DS-Cav1 pre-fusion F with or without foldon domain was fused to the TM of NDV F protein [99]. A recent study reported that NDV VLPs containing pre-fusion F stabilized single chain F with deletions of the peptide 27 sequence and cleavage sites and point mutations could induce higher RSV neutralizing antibodies in cotton rats, compared to NDV VLP with DS-Cav1 F [100].
RSV VLPs were also generated by using influenza matrix protein M1 as a core protein [101]. RSV G and F full-length glycoproteins were expressed in insect cells via rBVs containing genes encoding G or F proteins [101]. Both RSV-F and RSV-G VLPs were observed in spherical shapes with spikes on their surfaces in morphology. RSV F VLPs displayed a range of size distribution at approximate 80–100 nm in diameter, whereas G-VLPs were similar but somewhat more heterogeneous in size [101]. RSV VLPs were also reported to be generated by transfection of mammalian HEK293 cells with three types of expression plasmids, containing codon optimized RSV-G, F and M proteins [102]. RSV VLPs appear to mimic native RSV in morphology and structure, as examined by transmission electron microscopy.
A region within the central RSV G protein located between amino acids 131 to 230 was reported to contain protective epitopes, inducing T helper cell (Th cell) responses and eosinophilia [103,104]. A tandem repeat gene encoding this RSV G protein region containing T cell and B cell epitopes and CX3C motif was utilized to generate RSV G fragment VLPs [105]. G VLPs containing tandem repeat G fragment proteins could provide protection against RSV after vaccination of mice and challenge with RSV [105]. A different platform of RSV F protein nanoparticle vaccines was generated for advanced clinical trials. A near full-length RSV F protein with transmembrane domains, containing mutations to enhance the expression on insect cell surfaces using the rBV expression system, was purified under detergent conditions and formulated into nanoparticles, and found to be highly immunogenic in inducing neutralizing antibodies in cotton rats [106]. In a follow up study, prefusogenic F nanoparticle vaccine was found to induce neutralizing antibodies competitive with monoclonal antibodies targeting to multiple antigenic sites present on pre-F and post-F conformation [107]. Taken together, RSV G or F proteins could be presented on VLPs or formulated into nanoparticles utilizing different strategies, which were immunogenic and could induce protective immune responses in animal models.
3.2. Vaccine efficacy of RSV VLP vaccines
Alum-adjuvanted formalin-inactivated RSV (FI-RSV) vaccines failed to provide protection in children in clinical trials because vaccine-enhanced respiratory disease upon RSV infection was observed during winter epidemic season [108]. There are safety concerns in applying inactivated or subunit RSV vaccines to naïve young children. As implied by the absence of licensed RSV vaccines until now, it is challenging difficult to develop effective and safe RSV vaccines inducing protective RSV neutralizing immunity but avoiding RSV vaccination-induced respiratory disease after RSV infection. In a comparison study of RSV F VLP and alum-adjuvanted FI-RSV, F VLP intranasal immune mice induced T helper type 1 (Th1)-biased IgG2a antibodies, neutralizing immunity against RSV, CD8α+ CD103+ dendritic cells, F-specific IFN-γ+ and TNF-α+ CD8+ T cells, displaying least weight loss and no sign of histopathology and eosinophilia [109]. In contrast, alum-adjuvanted FI-RSV immune mice showed severe lung histopathology and eosinophilia together with high levels of IL-4+ and TNF α+ CD4+ T cell responses despite lung viral control after RSV challenge [109].
Chimeric RSV-NDV VLPs containing ectodomains of RSV F and G or G protein alone confer protection against RSV challenge infection in mice without displaying enhanced respiratory lung inflammation [94,95]. Immunization of mice with RSV VLP vaccines containing either F or G proteins induced RSV A2 -specific IgG, IgG1, and IgG2a antibodies in sera and in lung extracts [101]. RSV VLP vaccination of mice induced neutralizing antibodies, correlating with reducing lung viral loads and protection in mice after RSV challenge infection [101]. RSV VLPs composed of RSV G, F, M proteins elicited high titers of IgG antibody responses to homologous RSV A2 and heterologous RSV B strain challenge [102]. Mammalian cell-derived RSV VLP vaccines adjuvanted with combination of alum and monophosphoryl lipid A induced higher levels of IgG titers than non-adjuvanted VLPs [102].
RSV VLP vaccines with combined post-fusion and pre-fusion F glycoproteins afforded higher neutralizing titers, T helper type 1 (Th1) immune responses, and more effective prevention of lung viral replication, compared to either post-fusion F or pre-fusion F VLP vaccination after challenge in mice [110]. Vaccination of mice with combined RSV F VLP F + G VLP induced Th1 CD8 T cell responses and provided additive protection and prevented lung immunopathology after challenge but RSV G alone VLP vaccination resulted in substantial lung histopathology after RSV challenge [111]. VLP vaccine containing tandem repetitive RSV G fragment was able to provide recall B cell responses and protection against lung viral replication after RSV challenge [105]. Also, immunization with combined RSV VLP vaccines (F + G) with F DNA (FFG VLP) induced Th1-biased IgG2a isotype antibody responses to RSV F and more effective in preventing lung histopathogloy after RSV challenge than immunity induced by live RSV prior infection [112]. In addition, combined FFG VLP vaccination induced higher ratios of CD11c+ versus CD11b+ and IFN-γ+ CD8 versus CD4 T cells in addition to effective clearing lung viral loads [113]. Interestingly, prime immunization of mice with RSV F VLP vaccine induced Th1-biased immune response, preventing the induction of lung histopathology due to subsequent FI-RSV vaccination [114]. Unique immunogenic properties of RSV F VLP platforms were evident by a study reporting that soluble F protein vaccination exacerbated pulmonary histopathology upon RSV challenge but not when presented on VLPs in a mouse model [115]. Most individuals are considered to be infected with RSV by ages two to three years old but live RSV infection did not provide long-term protective immunity. A recent study demonstrated that avian cell-derived VLP vaccines containing ectodomains of RSV G and pre-F/F proteins were highly effective in inducing higher levels of RSV neutralizing antibodies even in the presence of prior RSV infection in a mouse model compared to the RSV re-infected mice [116]. Cotton rats are known to be more permissive to RSV infection and thus considered to be a preferred animal model than mice for RSV pathogenesis and vaccine efficacy studies. VLP vaccines containing two chimeric RSV ectodomain glycoproteins (chimeric pre-fusion F/F and H/G) were shown to confer protection against RSV replication upon RSV challenge in cotton rats [99]. NDV-RSV VLPs containing stabilized pre-fusion F/F chimera protein were more effective in inducing higher titers of serum neutralizing activity, compared to low dose live RSV prior infection [99]. Pre-F/F VLPs immunization protected cotton rats and did not result in enhanced lung inflammation after RSV challenge. Combined F + G VLP with F DNA vaccination of cotton rats was more effective in inducing RSV neutralizing antibody titers than RSV F but both combined FFG VLP and F VLP vaccination prevented the induction of vaccine-induced inflammation in the lungs, in contrast to FI-RSV [117,118].
Maternal vaccination with RSV VLP or F nanoparticle vaccines provided insight into protecting infants from RSV infection. RSV F nanoparticle protein or chimeric NDV-RSV VLP-induced immunity could be transferred from mothers to the infants [119]. Pregnant cotton rats were intramuscularly immunized with RSV pre-fusion F/G VLP or RSV F nanoparticle protein vaccines to evaluate the protective role of maternally transferred immunity in infants through the placental barrier [119,120]. The offspring pups born to vaccinated cotton rats were protected against RSV infection and pulmonary inflammation [120]. RSV F nanoparticle protein vaccine induced polyclonal palivizumab-competitive high neutralizing activities against RSV A and B viruses after active and passive immunization of cotton rats [119]. These results indicate that studies on RSV vaccines based on VLP and nanoparticle platforms are expected to provide important insight into developing effective and safe RSV vaccines.
4. HMPV VLP vaccines
4.1. Production of HMPV VLP vaccines
Human metapneumovirus (HMPV) is also an enveloped virus belonging to the pneumoviridae family and contains M protein as core protein which is encircled by lipid membrane expressing three HMPV surface antigens (F, G, SH). The F protein is required for virus assembly, fusion, and entry and the primary target of neutralizing antibodies [121,122]. HMPV-VLP was produced by co-expression of F and G proteins along with the core matrix protein M through the mammalian cell-based DNA transfection [123]. This study has also revealed some new findings about HMPV VLP assembly, that HMPV-G protein itself can independently facilitate the formation of VLP and additionally, that expression of HMPV-F protein on the surface of VLP is largely dependent on its association with the HMPV-G [123]. Another group of researchers has produced HMPV VLP by using retrovirus-derived Gag core protein [13]. Murine leukemia virus Gag protein was used as a core particle to generate HMPV VLPs containing HMPV- G and F glycoproteins from either lineage A or B [118]. HMPV VLPs have been also generated by expressing viral proteins matrix (M) and F proteins in suspension-adapted human embryonic kidney epithelial (293-F) cells through mammalian cell-based expression system [14]. In this study, full-length M and F viral protein sequences were derived from pathogenic clinical HMPV (genotype A2) isolates TN/94-49 and TN/92-4, respectively. Recently, another group of researchers reported the generation of HMPV VLPs by the mammalian cell-based expression system which expressed RSV prefusion F protein (deletion of p27 and introduction of disulfide bonds to stabilize pre-fusion F) [99] or post fusion F protein [110]. Metapneumovirus M protein was used as core protein and both codon optimized RSV Pre-fusion F and post-fusion F sequences were fused with the TM of hMPV-F protein to generate RSV-HMPV VLPs [110].
4.2. Vaccine efficacy of HMPV VLP vaccines
HMPV VLP vaccines displaying F and G surface glycoproteins of lineage A or B provided protection against HMPV challenge in mice [13] (Table 3). HMPV VLP immunized mice showed serum neutralizing antibodies and significant reduction of lung viral titers after challenge. Adjuvanted HMPV VLP expressing HMPV F and matrix (M) could provide complete protection against HMPV replication in the lungs of mice [14]. HMPV VLP immunization induced F-specific IgG antibody and CD8+ T cell response recognizing an F protein epitope. HMPV VLP vaccines adjuvanted with either TiterMax Gold or α-galactosylceramide enhanced neutralizing-antibody responses in mice [107]. Other studies demonstrated that serum neutralizing antibodies alone might not be sufficient for long-lasting protection against reinfection with HMPV [114] and that T cell immunity together would be important for protection against reinfection or long-term protection [124]. Importantly, HMPV VLP vaccination did not cause enhanced respiratory disease upon viral challenge [124]. These previous studies indicate that HMPV vaccines based on VLPs produced in mammalian cells could be effective in inducing protective immunity against HMPV.
5. Clinical studies of VLP vaccines against respiratory viruses
5.1. Clinical studies of influenza VLP vaccines
Seasonal influenza VLP vaccines might provide comparable or more effective protection over inactivated split vaccines produced in egg substrates. A phase-I/II, double-blind, dose dependent vaccine efficacy study was conducted in Mexico against 2009 pandemic H1N1 virus to evaluate the vaccine safety and immunogenicity of influenza HA-VLPs produced in insect cells via the rBV expression system [125] (Table 4). H1N1 VLP vaccine was safe and well tolerated in 4500 healthy adults at 18–64 ages after vaccination with VLP vaccines (5-, 15-, or 45-μg HA dose) [125]. The low dose (5 μg of HA) HA VLP vaccine was found to be immunogenic and safe after vaccination. The local adverse events were observed in the moderate (15-μg HA) or high dose (45-μg HA) groups. The low dose (5 μg of HA) subjects or placebo group did not display local and serious adverse events after vaccination. Notably, in the follow up clinical studies, 82-92% of all influenza VLP vaccinated subjects were observed to have sero-protective antibody levels (≥40 HAI titer) after boost [125]. Another phase I/II dose dependent clinical trial approved by Food and Drug administration was conducted in healthy adults to determine the efficacy of rBV/insect cell-derived A/Indonesia/05/2005 (H5N1) VLP vaccines [126]. In this clinical study, different doses of H5 HA (15, 45, or 90 μg of HA) were tested and found to induce functional antibodies, preferentially recognizing the oligomeric form of hemagglutinin [126]. Low HAI titers were observed in the 15 μg dose-immunized subjects whereas high HAI titers and cross-reactivity against other clade 2 viruses were induced in individuals with 45- and 90-μg H5 HA VLP dose immunization. There were no serious local or systemic adverse events, and only a single subject was discontinued due to an adverse event. An additional clinical study on avian influenza VLP vaccines, unadjuvanted H7N9 VLPs vaccine exhibited poor antibody responses [80]. In contrast, ISCOMATRIXTM adjuvanted VLP vaccines elicited HAI responses and raised high-quality antibody immune responses against avian influenza in naïve humans [80].
The safety and immunogenicity of the N. benthamiana plant cell-derived influenza H5 HA (A/Indonesia/5/05) VLP vaccines were assessed in a phase I human clinical trial [127]. To license an influenza vaccine, the immunogenic properties of vaccines should exhibit remarkable serologic responses as measured by HAI titers, single radial hemolysis (SRH) and microneutralization titers. A randomized, double-blind, placebo-controlled phase I clinical trial was performed among 18–60 years old healthy adults at the McGill University Health Center to assess the efficacy of plant cells derived, alum-adjuvanted A/Indonesia/5/05 H5N1 VLP by intramuscular immunization with 5, 10, or 20 µg vaccine dose [128]. The immunogenicity of vaccine (H5 HA VLP) was dose dependent, and 96% of vaccinated subjects with a high dose showed detectable microneutralization titers. In the clinical trials above, it has been reported that in the high dose 20 µg HA group after getting the first dose and the 10-µg HA group after receiving the second dose, 8.3% of the vaccinated subjects experienced transient headaches. However, the adverse events were mild-to-moderate and self-limited. Another phase I clinical trial has been completed with plant cell derived HA seasonal influenza VLP vaccine to evaluate the immunogenicity, safety, and efficacy [43]. In this clinical study, 18–49 years old subjects were intramuscularly administered H1-VLPs at 5-, 13-, and 28-μg H1 HA-VLPs. The clinical results were found to be safe and well tolerated at all dose levels and even 5-µg dose of VLPs induced greater than 1:40 HAI titers in 83% of the subjects in this low-vaccine dose group [43] (Table 4). Two randomized Phase II clinical trials in 18 to 49 and over 50 years old adults have shown that the 30-µg dose of quadrivalent plant-derived influenza VLP vaccine produced consistent humoral and cellular responses [129]. Recently, phase III clinical trials are ongoing to evaluate the efficacy, safety, and immunogenicity of plant-made influenza quadrivalent VLP vaccines during the 2018–2019 influenza season in elderly adults 65 years of age and older (ClinicalTrials.gov Identifier: NCT03739112). In this phase III trial, quadrivalent VLP influenza Vaccine (30-μg HA/strain) will be compared with standard seasonal vaccine (15-μg HA/strain).
A phase I double-blinded, randomized clinical trial has been conducted in healthy volunteer with bacteriophage gH1-Qbeta VLP vaccine with and without alhydrogel adjuvant to determine the efficacy and immunogenicity of bacteriophage-derived influenza VLP vaccine. The healthy subjects were intramuscularly immunized with 100-μg gH1-Qbeta VLP vaccine containing 42 μg HA antigen showed similar antibody-mediated immunogenicity and a comparable safety profile to commercially available vaccines [130].
These clinical studies support that plant cell-derived influenza VLP vaccines would be safe and immunogenic. Further clinical studies on quadrivalent VLP vaccine safety and dose-escalating studies in comparison with conventional egg-derived influenza vaccines are being planned.
5.2. Clinical studies of RSV nanoparticle vaccines
RSV F protein nanoparticle vaccines produced from insect cells via the rBV expression system are in phase III clinical trials [131,132]. A study of a RSV recombinant fusion (F) nanoparticle vaccine in healthy women of childbearing age demonstrated that the vaccine was safe, immunogenic, and appeared to be effective in reducing RSV infections (Clinical Trials Registration: NCT01704365) [133]. RSV F protein nanoparticle vaccination in older adults induces functional immunity to RSV (ClinicalTrials.gov number NCT01709019) [134].
In a phase III study with the elderly (≥60 years), the insect cell-derived recombinant RSV F nanoparticle vaccine failed to demonstrate significant efficacy of protection against RSV moderate to severe lower respiratory disease in older adults, partially due to a low-attack rate.
There are no clinical trials yet to evaluate the efficacy of RSV and HMPV vaccines based on VLP platforms, suggesting technological challenges in manufacturing clinical-grade RSV and HMPV VLP vaccines stabilizing pre-fusion F conformation and the scale-up processes of VLP vaccine production.
6. Expert opinion
6.1. Influenza VLP vaccines
VLPs provide an alternative platform to generate an effective vaccine probably due to its multifarious desirable properties. VLPs can be self-assembled, expressing one or multi proteins with dense epitopes or chimeric proteins being presented on the VLP surface. VLP vaccines can be produced in cell cultures with different expression systems and are compatible for formulating with various adjuvant systems. Influenza VLP vaccines present individual HA or NA or in combinations on the VLP surface as native-like functional glycoproteins. Both HA and NA proteins on VLPs can elicit HA and NA specific antibodies. However, due to the immune-dominant properties of HA protein, it could induce more HA-specific antibody responses than NA. To avoid immunodominant effects of HA, a separate individual NA VLP or multi-valent NA (N1, N2) VLP vaccine could be an alternative approach.
Influenza VLP vaccine production can be scalable through the insect or plant cell-based expression system. Compared to conventional influenza split vaccine or attenuated vaccines produced in embryonated egg substrates, influenza VLP vaccines would be stable in antigenic properties due to the lack of growth adaptation. An additional advantage is that manufacture of influenza VLP vaccine does not require handling live viruses and growth adaptation. In the case of pandemic outbreaks, it will be possible to produce and distribute insect cell-derived pandemic VLP vaccines within a short time period than egg-growth vaccines. Egg cell-based live attenuated influenza vaccines and inactivated split influenza vaccines would have a limitation in manufacturing capacity to produce sufficient vaccine doses to meet the surge in high vaccine demand during a pandemic outbreak.
In contrast to the plant cell expressed VLPs, rBV/insect cell-based expression system has flexibility to co-express various influenza viral structural proteins on the surface of the VLPs. Preclinical as well as clinical studies reported that immunization with insect cell-derived HA-NA-M1 VLPs can induce effective serum neutralizing antibodies with high HAI titers and NA inhibiting antibodies. Although HA immunity plays a major role in providing homologous protection, NA immunity would contribute to providing heterologous cross protection. In the clinical trials with influenza VLP vaccines produced in plant cells or in the insect cells, there were no serious adverse events reported so far. The combined VLP technology and innovative approaches of plant and insect cell expression systems will lead to developing effective influenza VLP vaccines in near future.
Production of recombinant purified protein subunit vaccines would cost high prices due to multiple purification steps but provide high safety features. To overcome low immunogenicity of soluble proteins, a technology of formulating protein nanoparticle vaccines would be highly desirable. Nanoparticle preparations of recombinant proteins (influenza virus HA conserved stalk domains, tandem repeat M2e proteins) expressed in insect cells, and purified to high purity, were immunogenic and effective in inducing cross-protective immunity in mice [135]. Combining the technology of VLP vaccine and protein nanoparticle vaccine technology would provide a promising vaccination strategy. Utilizing desirable properties of non-replication and high immunogenicity, VLP-based vaccine technologies are worthwhile to be further developed toward more effective and safe vaccines against viral pathogens difficult to control. Cross-protective influenza conserved antigens (M2e, conserved stalks, NA) are poorly immunogenic and thus developing effective VLP vaccines to present these cross protective antigens should be a future direction. VLP vaccines containing conserved cross-protective epitopes but inducing non-neutralizing immunity such as M2e would not induce high protective immunity, a strategy of supplementing HA-based vaccines with VLPs containing conserved epitopes might provide new vaccination enhancing cross-protective efficacy of current influenza vaccines. With encouraging outcomes from preclinical mouse studies and clinical trials of influenza VLP vaccine platforms, further development of VLP vaccines presenting influenza-conserved antigens in an immunogenic form would provide new approaches to develop cross-protective and universal influenza vaccines against seasonal and pandemic viruses.
6.2. RSV and HMPV VLP vaccines
RSV VLP vaccines were demonstrated to be unique in conferring protection against RSV while preventing vaccine-enhanced respiratory disease upon RSV infection [93,131]. These preclinical outcomes of RSV VLP vaccines in mice and cotton rat animal studies provide evidence warrantying further development of effective and safe RSV vaccines. It would be a future direction to develop RSV VLP platforms more stably presenting pre-fusion F proteins, compared to the current pre-F version with DS-Cav1. RSV pre-fusion stabilizing mutations (DS-Cav1) with a foldon stabilizer retained pre-F site Ø epitopes and site II neutralizing epitopes in soluble proteins [136]. It is likely that additional mutations together with DS-Cav1 or alternative mutations in RSV F will be required for further stabilization of pre-fusion F protein antigens on VLP platforms [99,100]. RSV F nanoparticle formulations of recombinant F proteins produced in the insect cell expression system are under advanced clinical trials of maternal vaccination to protect infants although the vaccine phase III clinical trial in the elderly was not successful [132,133,137]. An alternative vaccination strategy of heterologous prime with RSV pre-fusion F VLP and boost with pre-F protein nanoparticles might be a future study to develop effective and safe RSV vaccination. HMPV VLP vaccines were reported to provide protection against subgroups A and B in a mouse model but the prevention of vaccine-enhanced disease by HMPV VLPs remain to be further tested in more relevant animal models.
6.3. General consideration in VLP vaccine technology
A consideration in VLP vaccine technology is that host cell- and vector-derived components are randomly packed together into the assembled VLPs and sometimes abortive packaging can happen which would have impact on generating immune responses to vaccine antigens. There is a probability that protective immune responses would be dependent on the manufacturing process and resulting different batches of VLP vaccines. It is also expected that host immune responses to non-vaccine components in VLP vaccines would be induced after VLP vaccination although immune responses to non-vaccine antigen vector components would not have adverse effects since most enveloped viruses (influenza, RSV, HMPV) would contain host cell-derived non-viral components. Since VLP platform vaccines are effective in priming immune responses in naïve hosts, a strategy of heterologous VLP prime and protein boost vaccination would enhance immune responses to vaccine antigens, minimizing the immune responses to non-vaccine vector components. Advancing VLP vaccine technology should be continued to maximize the protective vaccine antigen contents in immunogenic conformation as for future research efforts.
Funding Statement
This work was supported by grants from the National Research Foundation of Korea (NRF) (2018R1A2B6003535, 2018R1A6A1A03025124) and Cooperative Research Program for Agriculture Science & Technology Development (PJ01320501), Rural Development Administration, Republic of Korea, and by National Institutes of Health (NIH)/National Institute of Allergy and Infectious Diseases (NIAID) grants AI093772 (SMK), AI105170 (SMK), and AI134132 (SMK).
Article highlights
Recombinant virus-like particles (VLPs) containing antigenic proteins from influenza virus, human respiratory syncytial virus (RSV), and human metapneumovirus (HMPV) can be generated in various expression systems (insect cells, mammalian cells, plant cells).
Influenza VLP vaccines are able to induce protective immune responses against seasonal influenza and pandemic influenza viruses in preclinical and clinical studies.
Influenza M2e VLPs, adjuvanted VLP vaccines could be candidates as universal influenza vaccines for broader immunity. However, cross-protective influenza VLP vaccines inducing higher efficacy remain to be developed. RSV and HMPV vaccines based on VLP platforms could be developed as promising candidates in providing protective immunity while avoiding the induction of enhanced respiratory disease. Heterologous prime with VLP vaccines and boost with protein nanoparticle vaccines would provide protective immunity against RSV and HMPV.
Further studies are required to develop VLP vaccine technology to maximize the vaccine components and to minimize immune responses to non-vaccine vector components.
Declaration of interest
The authors have no relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript. This includes employment, consultancies, honoraria, stock ownership or options, expert testimony, grants or patents received or pending, or royalties.
Reviewer disclosures
A reviewer has declared that they own a patent on plant-made H5 HA and that they are funded by Medicago Inc. for research on emerging viruses. All other peer reviewers on this manuscript have no relevant financial or other relationships to disclose.
References
Papers of special note have been highlighted as either of interest (•) or of considerable interest (••) to readers.
- 1.Pavia AT. Viral infections of the lower respiratory tract: old viruses, new viruses, and the role of diagnosis. Clin Infect Dis. 2011;52(Suppl4):S284–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jain S, Self WH, Wunderink RG, et al. Community-acquired pneumonia requiring hospitalization among US adults. N Engl J Med. 2015;373(5):415–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tregoning JS, Schwarze J.. Respiratory viral infections in infants: causes, clinical symptoms, virology, and immunology. Clin Microbiol Rev. 2010;23(1):74–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nakano K, Shiroma A, Shimoji M, et al. Advantages of genome sequencing by long-read sequencer using SMRT technology in medical area. Hum Cell. 2017;30(3):149–161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Quan F, Lee Y, Kim K, et al. Progress in developing virus-like particle influenza vaccines. Expert Rev Vaccines. 2016;15(10):1281–1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Pham D, Grégoire N, Couet W, et al. Pulmonary delivery of pyrazinamide-loaded large porous particles. Eur J Pharm Biopharm. 2015;94:241–250. [DOI] [PubMed] [Google Scholar]
- 7.Schweitzer JW, Justice NA. Respiratory Syncytial Virus Infection (RSV). StatPearls. Anonymous StatPearls Publishing LLC, St. Petersburg (FL); 2019. [PubMed] [Google Scholar]
- 8.Seale AC, Bianchi-Jassir F, Russell NJ, et al. Estimates of the burden of group B streptococcal disease worldwide for pregnant women, stillbirths, and children. Clin Infect Dis. 2017;65(Suppl2):S200–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Resch B. Product review on the monoclonal antibody palivizumab for prevention of respiratory syncytial virus infection. Hum Vaccin Immunother. 2017;13(9):2138–2149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Leung DY, Sampson H, Geha R, et al. Pediatric allergy: principles and practice E-Book. Amsterdam, Netherlands: Elsevier Health Sciences; 2010. [Google Scholar]
- 11.Wu N, Yang W, Beineke A, et al. The differentiated airway epithelium infected by influenza viruses maintains the barrier function despite a dramatic loss of ciliated cells. Sci Rep. 2016;6:39668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kim JI, Park S, Lee I, et al. Genome-wide analysis of human metapneumovirus evolution. PLoS One. 2016;11(4):e0152962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lévy C, Aerts L, Hamelin M, et al. Virus-like particle vaccine induces cross-protection against human metapneumovirus infections in mice. Vaccine. 2013;31(25):2778–2785. [DOI] [PubMed] [Google Scholar]
- 14.Cox RG, Erickson JJ, Hastings AK, et al. Human metapneumovirus virus-like particles induce protective B and T cell responses in a mouse model. J Virol. 2014;88(11):6368–6379. [DOI] [PMC free article] [PubMed] [Google Scholar]; •• A study reporting protective immunity induced by human metapneumovirus virus-like particles in mice.
- 15.Quan FS, Kim YC, Vunnava A, et al. Intradermal vaccination with influenza virus-like particles by using microneedles induces protection superior to that with intramuscular immunization. J Virol. 2010;84(15):7760–7769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Quan FS, Huang C, Compans RW, et al. Virus-like particle vaccine induces protective immunity against homologous and heterologous strains of influenza virus. J Virol. 2007;81(7):3514–3524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kushnir N, Streatfield SJ, Yusibov V. Virus-like particles as a highly efficient vaccine platform: diversity of targets and production systems and advances in clinical development. Vaccine. 2012;31(1):58–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Krammer F, Palese P. Advances in the development of influenza virus vaccines. Nat Rev Drug Discov. 2015;14(3):167. [DOI] [PubMed] [Google Scholar]
- 19.Akahata W, Yang Z, Andersen H, et al. A virus-like particle vaccine for epidemic Chikungunya virus protects nonhuman primates against infection. Nat Med. 2010;16(3):334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bachmann MF, Rohrer UH, Kundig TM, et al. The influence of antigen organization on B cell responsiveness. Science. 1993;262(5138):1448–1451. [DOI] [PubMed] [Google Scholar]
- 21.Latham T, Galarza JM. Formation of wild-type and chimeric influenza virus-like particles following simultaneous expression of only four structural proteins. J Virol. 2001;75(13):6154–6165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bright RA, Carter DM, Daniluk S, et al. Influenza virus-like particles elicit broader immune responses than whole virion inactivated influenza virus or recombinant hemagglutinin. Vaccine. 2007;25(19):3871–3878. [DOI] [PubMed] [Google Scholar]
- 23.Pushko P, Tumpey TM, Bu F, et al. Influenza virus-like particles comprised of the HA, NA, and M1 proteins of H9N2 influenza virus induce protective immune responses in BALB/c mice. Vaccine. 2005;23(50):5751–5759. [DOI] [PubMed] [Google Scholar]
- 24.Pushko P, Tumpey TM, Van Hoeven N, et al. Evaluation of influenza virus-like particles and Novasome adjuvant as candidate vaccine for avian influenza. Vaccine. 2007;25(21):4283–4290. [DOI] [PubMed] [Google Scholar]
- 25.Galarza JM, Latham T, Cupo A. Virus-like particle (VLP) vaccine conferred complete protection against a lethal influenza virus challenge. Viral Immunol. 2005;18(1):244–251. [DOI] [PubMed] [Google Scholar]
- 26.Park YC, Song JM. Preparation and immunogenicity of influenza virus-like particles using nitrocellulose membrane filtration. Clin Exp Vaccine Res. 2017;6(1):61–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Matassov D, Cupo A, Galarza JM. A novel intranasal virus-like particle (VLP) vaccine designed to protect against the pandemic 1918 influenza A virus (H1N1). Viral Immunol. 2007;20(3):441–452. [DOI] [PubMed] [Google Scholar]
- 28.Rezaei F, Mirshafiey A, Shahmahmoodi S, et al. Influenza Virus-like Particle Containing Two Different Subtypes of Hemagglutinin Confers Protection in Mice Against Lethal Challenge With A/PR8 (H1N1) and A/HK (H3N2) Viruses. Iran Red Crescent Med J. 2013;15(1):75–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lee D, Bae S, Park J, et al. Virus-like particle vaccine protects against H3N2 canine influenza virus in dog. Vaccine. 2013;31(32):3268–3273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Giles BM, Ross TM. A computationally optimized broadly reactive antigen (COBRA) based H5N1 VLP vaccine elicits broadly reactive antibodies in mice and ferrets. Vaccine. 2011;29(16):3043–3054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bright RA, Carter DM, Crevar CJ, et al. Cross-clade protective immune responses to influenza viruses with H5N1 HA and NA elicited by an influenza virus-like particle. PLoS One. 2008;3(1):e1501. [DOI] [PMC free article] [PubMed] [Google Scholar]; • A study that describes details on homologous and heterologous protection by H5N1 influenza VLPs.
- 32.Mahmood K, Bright RA, Mytle N, et al. H5N1 VLP vaccine induced protection in ferrets against lethal challenge with highly pathogenic H5N1 influenza viruses. Vaccine. 2008;26(42):5393–5399. [DOI] [PubMed] [Google Scholar]
- 33.Park J, Lee D, Youn H, et al. Protective efficacy of crude virus‐like particle vaccine against HPAI H5N1 in chickens and its application on DIVA strategy. Influenza Other Respir Viruses. 2013;7(3):340–348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Choi J, Kim M, Kang H, et al. Protective efficacy of baculovirus-derived influenza virus-like particles bearing H5 HA alone or in combination with M1 in chickens. Vet Microbiol. 2013;162(2–4):623–630. [DOI] [PubMed] [Google Scholar]
- 35.Wu C, Yeh Y, Yang Y, et al. Mammalian expression of virus-like particles for advanced mimicry of authentic influenza virus. PLoS One. 2010;5(3):e9784. [DOI] [PMC free article] [PubMed] [Google Scholar]; • A study reporting the production and characterization of influenza VLPs in mammalian cells.
- 36.Haynes JR, Dokken L, Wiley JA, et al. Influenza-pseudotyped Gag virus-like particle vaccines provide broad protection against highly pathogenic avian influenza challenge. Vaccine. 2009;27(4):530–541. [DOI] [PubMed] [Google Scholar]
- 37.Ren Z, Ji X, Meng L, et al. H5N1 influenza virus-like particle vaccine protects mice from heterologous virus challenge better than whole inactivated virus. Virus Res. 2015;200:9–18. [DOI] [PubMed] [Google Scholar]
- 38.Xue C, Tian G, Chen X, et al. Incorporation of conserved nucleoprotein into influenza virus-like particles could provoke a broad protective immune response in BALB/c mice and chickens. Virus Res. 2015;195:35–42. [DOI] [PubMed] [Google Scholar]
- 39.Pillet S, Racine T, Nfon C, et al. Plant-derived H7 VLP vaccine elicits protective immune response against H7N9 influenza virus in mice and ferrets. Vaccine. 2015;33(46):6282–6289. [DOI] [PubMed] [Google Scholar]
- 40.Liu YV, Massare MJ, Pearce MB, et al. Recombinant virus-like particles elicit protective immunity against avian influenza A (H7N9) virus infection in ferrets. Vaccine. 2015;33(18):2152–2158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lee D, Park J, Lee Y, et al. H9N2 avian influenza virus-like particle vaccine provides protective immunity and a strategy for the differentiation of infected from vaccinated animals. Vaccine. 2011;29(23):4003–4007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kang H, Chu K, Lee D, et al. Influenza M2 virus-like particle vaccination enhances protection in combination with avian influenza HA VLPs. PLoS One. 2019;14(6):e0216871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Redkiewicz P, Sirko A, Kamel KA, et al. Plant expression systems for production of hemagglutinin as a vaccine against influenza virus. Acta Biochim Pol. 2014;61:3. [PubMed] [Google Scholar]
- 44.Lindsay BJ, Bonar MM, Costas-Cancelas IN, et al. Morphological characterization of a plant-made virus-like particle vaccine bearing influenza virus hemagglutinins by electron microscopy. Vaccine. 2018;36(16):2147–2154. [DOI] [PubMed] [Google Scholar]
- 45.Hodgins B, Pillet S, Landry N, et al. A plant-derived VLP influenza vaccine elicits a balanced immune response even in very old mice with co-morbidities. PLoS One. 2019;14(1):e0210009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kang S, Song J, Quan F, et al. Influenza vaccines based on virus-like particles. Virus Res. 2009;143(2):140–146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Quan FS, Steinhauer D, Huang C, et al. A bivalent influenza VLP vaccine confers complete inhibition of virus replication in lungs. Vaccine. 2008;26(26):3352–3361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Margine I, Krammer F. Animal models for influenza viruses: implications for universal vaccine development. Pathogens. 2014;3(4):845–874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Grgacic EV, Anderson DA. Virus-like particles: passport to immune recognition. Methods. 2006;40(1):60–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Schwartzman LM, Cathcart AL, Pujanauski LM, et al. An intranasal virus-like particle vaccine broadly protects mice from multiple subtypes of influenza A virus. MBio. 2015;6(4):e01044–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang BZ, Quan FS, Kang SM, et al. Incorporation of membrane-anchored flagellin into influenza virus-like particles enhances the breadth of immune responses. J Virol. 2008;82(23):11813–11823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Quan F, Kim M, Lee B, et al. Influenza M1 VLPs containing neuraminidase induce heterosubtypic cross-protection. Virology. 2012;430(2):127–135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Ross TM, Mahmood K, Crevar CJ, et al. A trivalent virus-like particle vaccine elicits protective immune responses against seasonal influenza strains in mice and ferrets. PLoS One. 2009;4(6):e6032. [DOI] [PMC free article] [PubMed] [Google Scholar]; • A well comparison study of immunogenicity and efficacies between trivalent VLP and commercial trivalent inactivated vaccines.
- 54.Quan F, Vunnava A, Compans RW, et al. Virus-like particle vaccine protects against 2009 H1N1 pandemic influenza virus in mice. PLoS One. 2010;5(2):e9161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Pushko P, Kort T, Nathan M, et al. Recombinant H1N1 virus-like particle vaccine elicits protective immunity in ferrets against the 2009 pandemic H1N1 influenza virus. Vaccine. 2010;28(30):4771–4776. [DOI] [PubMed] [Google Scholar]
- 56.Wang B, Xu R, Quan F, et al. Intranasal immunization with influenza VLPs incorporating membrane-anchored flagellin induces strong heterosubtypic protection. PLoS One. 2010;5(11):e13972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Szécsi J, Boson B, Johnsson P, et al. Induction of neutralising antibodies by virus-like particles harbouring surface proteins from highly pathogenic H5N1 and H7N1 influenza viruses. Virol J. 2006;3(1):70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Song JM, Van Rooijen N, Bozja J, et al. Vaccination inducing broad and improved cross protection against multiple subtypes of influenza A virus. Proc Natl Acad Sci U S A. 2011;108(2):757–761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Song J, Hossain J, Yoo D, et al. Protective immunity against H5N1 influenza virus by a single dose vaccination with virus-like particles. Virology. 2010;405(1):165–175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Qin J, Zhang Y, Shen X, et al. H7 virus-like particles assembled by hemagglutinin containing H3N2 transmembrane domain and M1 induce broad homologous and heterologous protection in mice. Vaccine. 2018;36(33):5030–5036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Jiang W, Wang S, Chen H, et al. A bivalent heterologous DNA virus-like-particle prime-boost vaccine elicits broad protection against both group 1 and 2 influenza A viruses. J Virol. 2017;91(9):e02052–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Tretyakova I, Hidajat R, Hamilton G, et al. Preparation of quadri-subtype influenza virus-like particles using bovine immunodeficiency virus gag protein. Virology. 2016;487:163–171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kim Y, Lee Y, Kim M, et al. Cross-protective efficacy of influenza virus M2e containing virus-like particles is superior to hemagglutinin vaccines and variable depending on the genetic backgrounds of mice. Front Immunol. 2017;8:1730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kim M, Song J, Eunju O, et al. Virus-like particles containing multiple M2 extracellular domains confer improved cross-protection against various subtypes of influenza virus. Mol Ther. 2013;21(2):485–492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lee Y, Kim K, Ko E, et al. New vaccines against influenza virus. Clin Exp Vaccine Res. 2014;3(1):12–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Music N, Reber AJ, Kim M, et al. Supplementation of H1N1pdm09 split vaccine with heterologous tandem repeat M2e5x virus-like particles confers improved cross-protection in ferrets. Vaccine. 2016;34(4):466–473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lee Y, Lee Y, Kim M, et al. A novel vaccination strategy mediating the induction of lung-resident memory CD8 T cells confers heterosubtypic immunity against future pandemic influenza virus. J Immunol. 2016;196(6):2637–2645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Song B, Kang H, Lee E, et al. Supplemented vaccination with tandem repeat M2e virus-like particles enhances protection against homologous and heterologous HPAI H5 viruses in chickens. Vaccine. 2016;34(5):678–686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kim M, Lee Y, Ko E, et al. Supplementation of influenza split vaccines with conserved M2 ectodomains overcomes strain specificity and provides long-term cross protection. Mol Ther. 2014;22(7):1364–1374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Lee Y, Kim K, Ko E, et al. Enhancing the cross protective efficacy of live attenuated influenza virus vaccine by supplemented vaccination with M2 ectodomain virus-like particles. Virology. 2019;529:111–121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Andersson AC, Håkansson KO, Jensen BAH, et al. Increased immunogenicity and protective efficacy of influenza M2e fused to a tetramerizing protein. PLoS One. 2012;7(10):e46395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Eliasson DG, El Bakkouri K, Schön K, et al. CTA1-M2e-DD: a novel mucosal adjuvant targeted influenza vaccine. Vaccine. 2008;26(9):1243–1252. [DOI] [PubMed] [Google Scholar]
- 73.Wu F, Yuan X, Li J, et al. The co-administration of CpG-ODN influenced protective activity of influenza M2e vaccine. Vaccine. 2009;27(32):4320–4324. [DOI] [PubMed] [Google Scholar]
- 74.Ernst WA, Kim HJ, Tumpey TM, et al. Protection against H1, H5, H6 and H9 influenza A infection with liposomal matrix 2 epitope vaccines. Vaccine. 2006;24(24):5158–5168. [DOI] [PubMed] [Google Scholar]
- 75.Jefferson T, Rivetti D, Rivetti A, et al. Efficacy and effectiveness of influenza vaccines in elderly people: a systematic review. Lancet. 2005;366(9492):1165–1174. [DOI] [PubMed] [Google Scholar]
- 76.Wei H, Chang W, Lin S, et al. Fabrication of influenza virus-like particles using M2 fusion proteins for imaging single viruses and designing vaccines. Vaccine. 2011;29(41):7163–7172. [DOI] [PubMed] [Google Scholar]
- 77.Wang L, Wang YC, Feng H, et al. Virus-like particles containing the tetrameric ectodomain of influenza matrix protein 2 and flagellin induce heterosubtypic protection in mice. Biomed Res Int. 2013;686549:2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kim K, Kwon Y, Lee Y, et al. Virus-like particles presenting flagellin exhibit unique adjuvant effects on eliciting T helper type 1 humoral and cellular immune responses to poor immunogenic influenza virus M2e protein vaccine. Virology. 2018;524:172–181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Silva A, Mount A, Krstevska K, et al. The combination of ISCOMATRIX adjuvant and TLR agonists induces regression of established solid tumors in vivo. J Immunol. 2015;194(5):2199–2207. [DOI] [PubMed] [Google Scholar]
- 80.Chung KY, Coyle EM, Jani D, et al. ISCOMATRIX™ adjuvant promotes epitope spreading and antibody affinity maturation of influenza A H7N9 virus like particle vaccine that correlate with virus neutralization in humans. Vaccine. 2015;33(32):3953–3962. [DOI] [PubMed] [Google Scholar]; •• A study of clinical trials proving the advantages of timing and efficacy of pandemic potential H7N9 influenza VLP vaccines.
- 81.Smith GE, Flyer DC, Raghunandan R, et al. Development of influenza H7N9 virus like particle (VLP) vaccine: homologous A/Anhui/1/2013 (H7N9) protection and heterologous A/chicken/Jalisco/CPA1/2012 (H7N3) cross-protection in vaccinated mice challenged with H7N9 virus. Vaccine. 2013;31(40):4305–4313. [DOI] [PubMed] [Google Scholar]
- 82.Ko E, Lee Y, Lee Y, et al. Flagellin-expressing virus-like particles exhibit adjuvant effects on promoting IgG isotype-switched long-lasting antibody induction and protection of influenza vaccines in CD4-deficient mice. Vaccine. 2019;37(26):3426–3434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Patel JM, Kim M, Vartabedian VF, et al. Protein transfer-mediated surface engineering to adjuvantate virus-like nanoparticles for enhanced anti-viral immune responses. Nanomedicine. 2015;11(5):1097–1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Mohan T, Berman Z, Luo Y, et al. Chimeric virus-like particles containing influenza HA antigen and GPI-CCL28 induce long-lasting mucosal immunity against H3N2 viruses. Sci Rep. 2017;7:40226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kazaks A, Lu I, Farinelle S, et al. Production and purification of chimeric HBc virus-like particles carrying influenza virus LAH domain as vaccine candidates. BMC Biotechnol. 2017;17(1):79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Chiang C, Beljanski V, Yin K, et al. Sequence-specific modifications enhance the broad-spectrum antiviral response activated by RIG-I agonists. J Virol. 2015;89(15):8011–8025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Beljanski V, Chiang C, Kirchenbaum GA, et al. Enhanced influenza virus-like particle vaccination with a structurally optimized RIG-I agonist as adjuvant. J Virol. 2015;89(20):10612–10624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Li X, Ju H, Liu J, et al. Influenza virus‐like particles harboring H9N2 HA and NA proteins induce a protective immune response in chicken. Influenza Other Respir Viruses. 2017;11(6):518–524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Quan F, Ko E, Kwon Y, et al. Mucosal adjuvants for influenza virus-like particle vaccine. Viral Immunol. 2013;26(6):385–395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.McLellan JS, Yang Y, Graham BS, et al. Structure of respiratory syncytial virus fusion glycoprotein in the postfusion conformation reveals preservation of neutralizing epitopes. J Virol. 2011;85(15):7788–7796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Lee H, Lee J, Park M, et al. Monoclonal antibody against G glycoprotein increases respiratory syncytial virus clearance in vivo and prevents vaccine-enhanced diseases. PLoS One. 2017;12(1):e0169139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Olson MR, Varga SM. Pulmonary immunity and immunopathology: lessons from respiratory syncytial virus. Expert Rev Vaccines. 2008;7(8):1239–1255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Schepens B, Schotsaert M, Saelens X. Small hydrophobic protein of respiratory syncytial virus as a novel vaccine antigen. Immunotherapy. 2015;7(3):203–206. [DOI] [PubMed] [Google Scholar]
- 94.McGinnes LW, Gravel KA, Finberg RW, et al. Assembly and immunological properties of Newcastle disease virus-like particles containing the respiratory syncytial virus F and G proteins. J Virol. 2011;85(1):366–377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Murawski MR, McGinnes LW, Finberg RW, et al. Newcastle disease virus-like particles containing respiratory syncytial virus G protein induced protection in BALB/c mice, with no evidence of immunopathology. J Virol. 2010;84(2):1110–1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Chaiwatpongsakorn S, Epand RF, Collins PL, et al. Soluble respiratory syncytial virus fusion protein in the fully cleaved, pretriggered state is triggered by exposure to low-molarity buffer. J Virol. 2011;85(8):3968–3977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.McGinnes Cullen L, Schmidt MR, Morrison TG. Effect of previous respiratory syncytial virus infection on murine immune responses to F and G protein-containing virus-like particles. J Virol. 2019;93(9):e00087–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.McLellan JS, Chen M, Leung S, et al. Structure of RSV fusion glycoprotein trimer bound to a prefusion-specific neutralizing antibody. Science. 2013;340(6136):1113–1117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.McGinnes Cullen L, Schmidt MR, Kenward SA, et al. Murine immune responses to virus-like particle-associated pre- and postfusion forms of the respiratory syncytial virus F protein. J Virol. 2015;89(13):6835–6847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Cullen LM, Schmidt MR, Torres GM, et al. Comparison of immune responses to different versions of VLP associated stabilized RSV pre-fusion F protein. Vaccines (Basel). 2019;7(1):21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Quan F, Kim Y, Lee S, et al. Viruslike particle vaccine induces protection against respiratory syncytial virus infection in mice. J Infect Dis. 2011;204(7):987–995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Walpita P, Johns LM, Tandon R, et al. Mammalian cell-derived respiratory syncytial virus-like particles protect the lower as well as the upper respiratory tract. PLoS One. 2015;10(7):e0130755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Tebbey PW, Hagen M, Hancock GE. Atypical pulmonary eosinophilia is mediated by a specific amino acid sequence of the attachment (G) protein of respiratory syncytial virus. J Exp Med. 1998;188(10):1967–1972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Sparer TE, Matthews S, Hussell T, et al. Eliminating a region of respiratory syncytial virus attachment protein allows induction of protective immunity without vaccine-enhanced lung eosinophilia. J Exp Med. 1998;187(11):1921–1926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Kim A, Lee D, Lee S, et al. Protection induced by virus-like particle vaccine containing tandem repeat gene of respiratory syncytial virus G protein. PLoS One. 2018;13(1):e0191277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Smith G, Raghunandan R, Wu Y, et al. Respiratory syncytial virus fusion glycoprotein expressed in insect cells form protein nanoparticles that induce protective immunity in cotton rats. PLoS One. 2012;7(11):e50852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Gilbert BE, Patel N, Lu H, et al. Respiratory syncytial virus fusion nanoparticle vaccine immune responses target multiple neutralizing epitopes that contribute to protection against wild-type and palivizumab-resistant mutant virus challenge. Vaccine. 2018;36(52):8069–8078. [DOI] [PubMed] [Google Scholar]
- 108.Kim HW, Canchola JG, Brandt CD, et al. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am J Epidemiol. 1969;89(4):422–434. [DOI] [PubMed] [Google Scholar]
- 109.Kim KH, Lee YT, Hwang HS, et al. Virus-like particle vaccine containing the f protein of respiratory syncytial virus confers protection without pulmonary disease by modulating specific subsets of dendritic cells and effector T cells. J Virol. 2015;89(22):11692–11705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Cimica V, Boigard H, Bhatia B, et al. Novel respiratory syncytial virus-like particle vaccine composed of the postfusion and prefusion conformations of the F glycoprotein. Clin Vaccine Immunol. 2016;23(6):451–459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Lee S, Quan F, Kwon Y, et al. Additive protection induced by mixed virus-like particles presenting respiratory syncytial virus fusion or attachment glycoproteins. Antiviral Res. 2014;111:129–135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Hwang HS, Kwon Y, Lee JS, et al. Co-immunization with virus-like particle and DNA vaccines induces protection against respiratory syncytial virus infection and bronchiolitis. Antiviral Res. 2014;110:115–123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Ko E, Kwon Y, Lee JS, et al. Virus-like nanoparticle and DNA vaccination confers protection against respiratory syncytial virus by modulating innate and adaptive immune cells. Nanomedicine. 2015;11(1):99–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Hwang HS, Lee Y, Kim K, et al. Virus-like particle vaccine primes immune responses preventing inactivated-virus vaccine-enhanced disease against respiratory syncytial virus. Virology. 2017;511:142–151. [DOI] [PubMed] [Google Scholar]
- 115.Lee Y, Lee Y, Ko E, et al. Soluble F proteins exacerbate pulmonary histopathology after vaccination upon respiratory syncytial virus challenge but not when presented on virus-like particles. Hum Vaccin Immunother. 2017;13(11):2594–2605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Cullen LM, Schmidt MR, Morrison TG. Effect of previous respiratory syncytial virus infection on murine immune responses to F and G protein containing virus-like particles. J Virol. 2019;93(9):e00087–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Hwang HS, Kim K, Lee Y, et al. Virus-like particle vaccines containing F or F and G proteins confer protection against respiratory syncytial virus without pulmonary inflammation in cotton rats. Hum Vaccin Immunother. 2017;13(5):1031–1039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Hwang HS, Lee Y, Kim K, et al. Combined virus-like particle and fusion protein-encoding DNA vaccination of cotton rats induces protection against respiratory syncytial virus without causing vaccine-enhanced disease. Virology. 2016;494:215–224. [DOI] [PubMed] [Google Scholar]
- 119.Raghunandan R, Lu H, Zhou B, et al. An insect cell derived respiratory syncytial virus (RSV) F nanoparticle vaccine induces antigenic site II antibodies and protects against RSV challenge in cotton rats by active and passive immunization. Vaccine. 2014;32(48):6485–6492. [DOI] [PMC free article] [PubMed] [Google Scholar]; • A study reporting F nanoparticle vaccine efficacy in cotton rats.
- 120.Blanco JC, Pletneva LM, McGinnes-Cullen L, et al. Efficacy of a respiratory syncytial virus vaccine candidate in a maternal immunization model. Nat Commun. 2018;9(1):1904. [DOI] [PMC free article] [PubMed] [Google Scholar]; • A study reporting vaccine efficacy in a maternal immunization model.
- 121.Leyrat C, Paesen GC, Charleston J, et al. Structural insights into the human metapneumovirus glycoprotein ectodomain. J Virol. 2014;88(19):11611–11616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Cseke G, Wright DW, Tollefson SJ, et al. Human metapneumovirus fusion protein vaccines that are immunogenic and protective in cotton rats. J Virol. 2007;81(2):698–707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Loo LH, Jumat MR, Fu Y, et al. Evidence for the interaction of the human metapneumovirus G and F proteins during virus-like particle formation. Virol J. 2013;10(1):294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Wen SC, Schuster JE, Gilchuk P, et al. Lung CD8+ T cell impairment occurs during human metapneumovirus infection despite virus-like particle induction of functional CD8+ T cells. J Virol. 2015;89(17):8713–8726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.López-Macías C, Ferat-Osorio E, Tenorio-Calvo A, et al. Safety and immunogenicity of a virus-like particle pandemic influenza A (H1N1) 2009 vaccine in a blinded, randomized, placebo-controlled trial of adults in Mexico. Vaccine. 2011;29(44):7826–7834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Khurana S, Wu J, Verma N, et al. H5N1 virus-like particle vaccine elicits cross-reactive neutralizing antibodies that preferentially bind to the oligomeric form of influenza virus hemagglutinin in humans. J Virol. 2011;85(21):10945–10954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.D’Aoust M, Couture MM, Charland N, et al. The production of hemagglutinin‐based virus‐like particles in plants: a rapid, efficient and safe response to pandemic influenza. Plant Biotechnol J. 2010;8(5):607–619. [DOI] [PubMed] [Google Scholar]
- 128.Landry N, Ward BJ, Trépanier S, et al. Preclinical and clinical development of plant-made virus-like particle vaccine against avian H5N1 influenza. PLoS One. 2010;5(12):e15559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Pillet S, Couillard J, Trépanier S, et al. Immunogenicity and safety of a quadrivalent plant-derived virus like particle influenza vaccine candidate—two randomized Phase II clinical trials in 18 to 49 and≥ 50 years old adults. PLoS One. 2019;14(6):e0216533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Low JG, Lee LS, Ooi EE, et al. Safety and immunogenicity of a virus-like particle pandemic influenza A (H1N1) 2009 vaccine: results from a double-blinded, randomized Phase I clinical trial in healthy Asian volunteers. Vaccine. 2014;32(39):5041–5048. [DOI] [PubMed] [Google Scholar]
- 131.Broadbent L, Groves H, Shields MD, et al. Respiratory syncytial virus, an ongoing medical dilemma: an expert commentary on respiratory syncytial virus prophylactic and therapeutic pharmaceuticals currently in clinical trials. Influenza Other Respir Viruses. 2015;9(4):169–178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Mazur NI, Higgins D, Nunes MC, et al. The respiratory syncytial virus vaccine landscape: lessons from the graveyard and promising candidates. Lancet Infect Dis. 2018;18(10):e295–311. [DOI] [PubMed] [Google Scholar]
- 133.Glenn GM, Fries LF, Thomas DN, et al. A randomized, blinded, controlled, dose-ranging study of a respiratory syncytial virus recombinant fusion (F) nanoparticle vaccine in healthy women of childbearing age. J Infect Dis. 2015;213(3):411–422. [DOI] [PubMed] [Google Scholar]
- 134.Patel N, Massare MJ, Tian J, et al. Respiratory syncytial virus prefusogenic fusion (F) protein nanoparticle vaccine: structure, antigenic profile, immunogenicity, and protection. Vaccine. 2019;37:6112–6124. [DOI] [PubMed] [Google Scholar]
- 135.Deng L, Mohan T, Chang TZ, et al. Double-layered protein nanoparticles induce broad protection against divergent influenza A viruses. Nat Commun. 2018;9(1):359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.McLellan JS, Chen M, Joyce MG, et al. Structure-based design of a fusion glycoprotein vaccine for respiratory syncytial virus. Science. 2013;342(6158):592–598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Fries L, Shinde V, Stoddard JJ, et al. Immunogenicity and safety of a respiratory syncytial virus fusion protein (RSV F) nanoparticle vaccine in older adults. Immun Ageing. 2017;14(1):8. [DOI] [PMC free article] [PubMed] [Google Scholar]