

Abstract
The immune system is a very diverse system of the host that evolved during evolution to cope with various pathogens present in the vicinity of environmental surroundings inhabited by multicellular organisms ranging from achordates to chordates (including humans). For example, cells of immune system express various pattern recognition receptors (PRRs) that detect danger via recognizing specific pathogen-associated molecular patterns (PAMPs) and mount a specific immune response. Toll-like receptors (TLRs) are one of these PRRs expressed by various immune cells. However, they were first discovered in the Drosophila melanogaster (common fruit fly) as genes/proteins important in embryonic development and dorso-ventral body patterning/polarity. Till date, 13 different types of TLRs (TLR1-TLR13) have been discovered and described in mammals since the first discovery of TLR4 in humans in late 1997. This discovery of TLR4 in humans revolutionized the field of innate immunity and thus the immunology and host-pathogen interaction. Since then TLRs are found to be expressed on various immune cells and have been targeted for therapeutic drug development for various infectious and inflammatory diseases including cancer. Even, Single nucleotide polymorphisms (SNPs) among various TLR genes have been identified among the different human population and their association with susceptibility/resistance to certain infections and other inflammatory diseases. Thus, in the present review the current and future importance of TLRs in immunity, their pattern of expression among various immune cells along with TLR based therapeutic approach is reviewed.
Keywords: Innate immunity, TLRs, Infection, Inflammation, PAMPs, PRRs
Graphical abstract
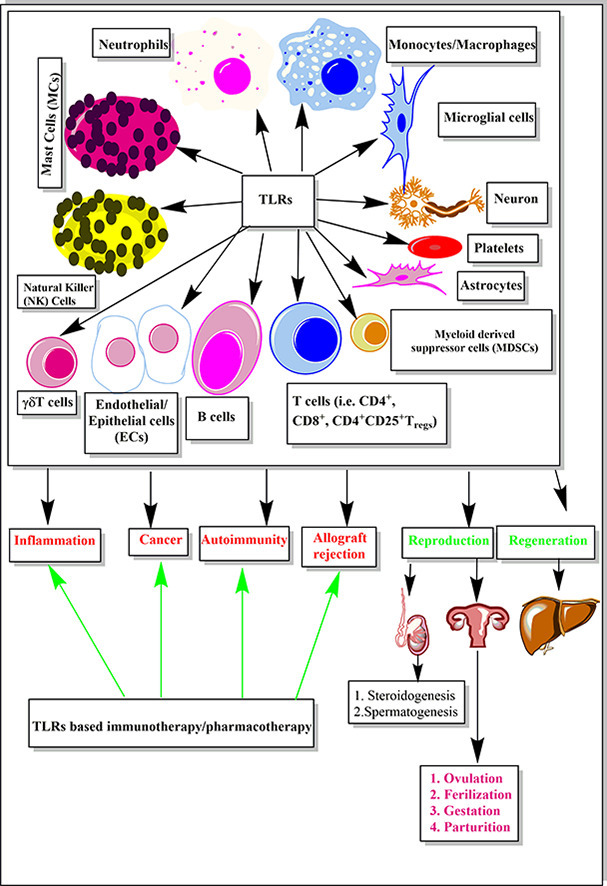
Highlights
-
•
TLRs are first described PRRs that revolutionized the biology of host-pathogen interaction and immune response
-
•
The discovery of different TLRs in humans proved milestone in the field of innate immunity and inflammation
-
•
The pattern of expression of all the TLRs expressed by human immune cells
-
•
An association of various TLR SNPs with different inflammatory diseases
-
•
Currently available drugs or vaccines based on TLRs and their future in drug targeting along with the role in reproduction, and regeneration
1. Introduction
The innate immune system is the primary defense entity of the host to protect against invading pathogens and thought to be evolutionarily conserved and phylogenetically ancient arm of the immune system [1,2]. In humans innate immune system mainly comprises of innate immune cells (i.e. monocytes/macrophages, neutrophils, dendritic cells (DCs), natural killer (NK) cells, mast cells (MCs), eosinophils, basophils along with newly identified innate lymphoid cells (ILCs) and mucosal associated invariant T (MAIT) cells, γδT cells, NKT cells etc.) [[3], [4], [5], [6], [7], [8], [9]] and its humoral components that is circulating complement system proteins/components, cytokines and chemokines secreted by innate immune cells along with various antimicrobial peptides (AMPs) (i.e. LL37, Bactericidal/permeability increasing protein (BPI) etc. [[10], [11], [12], [13], [14], [15], [16]]. Innate immune cells express various pattern recognition receptors (PRRs) including Toll-like receptors (TLRs,) responsible for the recognition of pathogen-associated molecular patterns (PAMPs) and induction of inflammatory immune response [[17], [18], [19], [20], [21]]. Thus this recognition of pathogens by PRRs plays a very important role in the generation of an effective innate immune response. TLRs are one of highly conserved PRRs and have been identified in animals as low as nematodes that is Caenorhabditis elegans (C. elegans) and in ascidian called Ciona intestinalis (C. intestinalis) [[22], [23], [24], [25]]. The first identification of TLRs in 1988 in Drosophila melanogaster or D. melanogaster [26] and then subsequent recognition of its one homolog called TLR4 in humans in 1997 [27] revolutionized the field of innate immunity. This novel discovery of TLR4 in humans filled the great gap stayed long in the field of immunology that is how pathogens and microbes are recognized by host immune system. However, a variation in TLR4 expression and function in different animal species is also observed [28]. In addition to the variation in expression of TLR4 among different animals, a great variation in expression of the number of TLRs in the animal kingdom is observed [29]. For example, Purple sea urchin or Strogylocentrotus purpuratus expresses most that is 222 TLRs, Amphioxus or Branchiostoma floridae expresses 42 TLRs, Xenopus or Xenopus tropicalis expresses 19 TLRs, while Zebra fish or Danio rerio expresses 17 TLRs [30,31]. This can be explained on the basis of the evolutionary primitiveness of the animal. This is because TLRs are involved in the recognition of almost every pathogen including bacteria, viruses, fungi, and parasites in animals as soon as they come in contact with the host via any route of pathogen exposure [[32], [33], [34]]. Thus, TLRs are very important PRRs of immune system required to initiate an effective innate immune response at an early stage of infection [35,36]. While at later stages these TLRs regulate the generation of adaptive immune response [[37], [38], [39]]. Thus, TLRs are still sitting over the top of the immune system pyramid since their first discovery in D. melanogaster in 1988 and will they be remain sitting at this position in the ever-changing and evolving field of innate immunity and immunology. This review is designed to highlight the past, present, and future of TLRs in immunity in terms of their pattern of expression in various immune cells, recognition of various TLR SNPs in humans making them resistant/susceptible to various infections and inflammatory disease and development of various TLR agonists and antagonist as pharmacological therapeutics and/or vaccine adjuvants.
2. Recognition of pathogens by TLRs and generation of inflammatory immune response
2.1. Discovery of TLRs and their recognition as PRRs
The Toll protein was first identified in D. melanogaster or common fruit fly as an integral membrane protein with a cytoplasmic domain and a large extra cytoplasmic domain with a role in dorso-ventral body patterning during embryonic development as a maternal effect gene [26]. The further study established that maternal expression of toll genes plays an important role in the correct spatial organization of lateral and ventral structures of Drosophila embryo [40]. While expression of toll gene in the embryo is an essential factor for the survival of embryo and this zygotic Toll protein exhibits similar biochemical activity as shown by maternal Toll protein [40]. Thus, Toll proteins were first identified as very important proteins responsible for the viability of the insect embryo and their development along with patterning.
In 1991, Gay and Keith showed that cytoplasmic domain of Toll protein of Drosophila was related to interleukin-1receptor (IL-1R) of humans [41]. These Toll proteins were further shown to exert antifungal action in D. melanogaster via regulating the gene responsible for synthesis of an antifungal peptide called drosomycin [42]. Thus, an era of recognition of TLRs as PRRs was about to begin as later in 1997 human homolog of Toll protein was identified by the group led by a prominent immunologist Charles A Janeway Junior, which is now known as toll-like receptor 4 (TLR-4) [27]. Thus, similar to Drosophila Toll, human Toll is also a type I transmembrane protein having an extracellular domain comprising of a leucine-rich repeat (LRR) domain, and a cytoplasmic domain homologous to the cytoplasmic domain of the human IL-1R [27]. Both Drosophila Toll and the IL-1 receptor (IL-1R) signal through NF-κB pathway [27,41].
This identification of human Toll protein revolutionized the field of immunology and led to the development of the concept of PRRs and innate immunity [43]. This is because after their first identification and characterization in humans that is human TLR4, different Drosophila homologs of Toll have been identified, causing an activation of NF-κB upon stimulation. This was further strengthened by the view of Gay and Keith (1991) that TLRs and IL-1Rs are related to each other and trigger similar signaling responsible for the inflammatory pathway [27,41,44]. By 1998, five human TLRs (i.e.TLR1, 2, 3, 4 and 5) had been identified as direct homologs of Drosophila Toll protein along with their genetic location on human chromosomes [44]. For example, tlr 1, 2, and 3 genes for humans are located on chromosome 4, tlr4 gene is located on chromosome 9 and tlr5 gene sits on the chromosome 1 [44]. Subsequently in 1998 the receptor for identification of lipopolysaccharide (LPS) in mouse was identified by a group of researchers, called TLR4 belonging to IL-1R family [45] as a mutation in this gene was making certain mice strains (i.e. C3H/HeJ and C57BL/10ScCr mice) more resistant to LPS but still they were highly prone to get gram-negative bacterial infections [46,47]. Thus, identification of TLRs and the TLR4 as a potential and major receptor for bacterial LPS in mice and humans revolutionized the biology of the mammalian immune system [43,48]. This discovery proved to be a milestone in the evolution of the innate immune system as an important component of the immune system responsible for recognizing potential pathogens, inflammatory immune response and regulating the adaptive immune response. This is because, before the identification of PRRs and concepts regarding innate immunity given by late CA Janeway Junior [49,50], immunologists were mainly focussed on adaptive immunity that is T cell and B cell-based immune response. Thus, the discovery of TLRs as PRRs played an important role in the establishment of the innate immune system as a separate and important branch of immunology. Furthermore, both the arms of immune system regulate each other depending on the cause, duration, and intensity of the associated immune response and its outcome.
3. Current scenario in TLR biology, regulation of innate immune response by TLRs during infection and generation of pro-inflammatory immune response
TLRs have been considered evolutionarily conserved proteins and the oldest TLR has been identified in nematodes (i.e. Caenorhabditis elegans or C. elegans) [51,52]. These are essentially characterized by an extracellular leucine-rich repeat (LRRs) domain, which mediates recognition of PAMPs, a transmembrane domain along with its cytosolic or intracellular Toll/IL-1R-like (TIR) domains required for downstream signaling pathways [27,[51], [52], [53]]. Thus, Toll signaling is present from primitive life that is nematode to the most advanced form of life that is modern human and plays a very important role in the development and immune response [52]. Till date, 10 functional TLRs (i.e. TLR1-TLR10) in humans and 13 active TLRs in laboratory mice have been identified [54]. Whereas, D. melanogaster, has 9 different Toll proteins (i.e. Toll, 18 Wheeler (18W) or Toll-2, Toll-3-Toll-9) [55]. These Toll receptors have two or more characteristic cysteine-rich motifs flanking LRRs. However, TLR-9 of humans and toll-9 of D. melanogaster have only single cysteine-rich motif between the transmembrane domain and LRRs [55]. Even toll-9 from D. melanogaster has a great homology with mammalian TLR1, 2, 4 and 6 [56].
All TLRs expressed by host cells are synthesized in the endoplasmic reticulum (ER) and are transported to Golgi complex and from there these TLRs are transported to either cell membrane or intracellular compartments (i.e. endosomes) [57]. The trafficking of intracellular TLRs (i.e. TLR3, TLR7, TLR8, and TLR9) to endosomes is controlled and regulated by a multi-pass transmembrane protein called UNC93B1 (Unc-93 homolog B1) [57]. The excessive activation of TLR7 is also controlled by UNC93B1 by employing TLR9 to counteract the exaggerated activation of TLR7 [57,58]. Protein associated with TLR4 (PRAT4A) is another ER resident protein molecule controlling TLR trafficking of TLR1, TLR2, TLR4, TLR7 and TLR9 from ER to their site of location that is plasma membrane and endosomes [59]. gp96 (a member of Hsp90 family) in ER acts as a general chaperone for most of TLRs including TLR1, TLR2, TLR4, TLR5, TLR7 and TLR9 [60]. Proteolytic cleavage of nucleic acid-sensing TLRs by Cathepsin B, S, L, H, and K and asparaginyl endopeptidase is required for the functional maturation of TLRs to recognize their competent ligands and mount an effective innate immune response [[61], [62], [63]].
TLRs are either expressed extracellularly on the cell surface (For example, TLR1, TLR2, TLR4, TLR5, TLR6, and probably TLR11 and TLR12 of mice and TLR10 of humans are expressed largely on the cell surface of innate immune cells) or intracellularly in the cytosolic compartment inside vesicles, for example in endosomes (TLR3, TLR7, TLR8, and TLR9 are intracellular TLRs), where they recognize microbial genetic material that is DNA or RNA [[64], [65], [66], [67], [68], [69], [70], [71], [72], [73], [74]]. However, TLR2 and TLR4 are also present as intracellular TLRs in DCs, epithelial and endothelial cells [[75], [76], [77]]. As these intracellular TLRs are present in vesicles or endosomal compartment so they do not come in contact with host cell-derived nucleic acids (DNA or RNA) under most physiologic conditions and do not cause activation of an innate immune response against self DNA or RNA [78]. These intracellular TLRs present in endosomes play an important role in the generation of anti-nucleic acid and rheumatoid factor autoantibodies (for example, IgG autoAbs and IgM rheumatoid factor) in patients suffering from systemic lupus erythematosus (SLE), where autoantibodies are generated against person's own nucleic acids (i.e. DNAs and RNAs) [79]. However, it is beyond the scope of the present article to discuss the detail of each TLR signaling pathway that is described elsewhere [[80], [81], [82], [83], [84], [85]]. Table 1 is summarizing the numbers of TLRs found in mammalian species mainly in humans and laboratory mouse and their role in pathogen recognition, TLR SNPs and disease association and expression pattern.
Table 1.
TLRs, their cellular location, major ligands with their source, TLR SNPs and disease association.
| TLRs | TLR polymorphism (SNP and nucleic acid variant) | TLR polymorphism and disease association | TLR localization | Ligands | Origin of ligands |
|---|---|---|---|---|---|
| TLR1 | T1805G (I602S) R80T | 602S SNP in TLR1 is protective against Leprosy caused by Mycobacterium leprae [397] R80T is associated with increased risk of Aspergillosis [514,515] |
Plasma membrane | Triacyl lipopeptide Soluble factors |
Bacteria and mycobacteria |
| TLR2 | T597C R753Q, SNP-15607A/G of haplotype 2 of TLR2 | T597C SNP Protects against leprosy but predisposes towards tuberculosis infection [516,517] R753Q increases susceptibility to S. aureus infection [518], 15607A/G of haplotype 2 of TLR2 is associated with increased viral shedding and lesional rate in patients with genital herpes simplex virus (HSV) Type 2 infection [519] |
Plasma membrane | PGN, LTA, Lipoproteins or lipopeptides, lipoarabinomannan, A phenol-soluble modulin, Glycoinositolphosp-holipids, glycolipids, porins, zymosan, atypical LPS, Hsp70, EDN | Gram +ve bacteria, mycobacteria, S. epidermidis, Trypanosoma cruzi, Treponema maltophilum, Neisseria, Fungi, Leptospira interrogans, Porphyromonas gingivalis, host, host |
| TLR3 | rs1879026 (G/T), rs13126816, rs3775291, L412F TLR3 polymorphism, 299698T/G, 293248A/A, 299698T/T | rs1879026 (G/T) is linked to increased prevalence of hepatitis B virus infection [520], rs13126816 and rs3775291 are associated with protection against human herpes simplex virus type 2 (HSV-2) infections [521], L412F TLR3 polymorphism is associated with resistance against HIV infection in the Italian population [522], and same SNP in Japanese population is associated with protection against acute graft rejection in adult patients receiving liver transplantation for hepatitis C virus (HCV)-related cirrhosis [523], 299698T/G, 293248A/A, 299698T/T SNPs in the exon of TLR3 are Stevens-Johnson syndrome (SJS), mucocutaneous disease in Japanese patients [524] | Endolysosome | dsRNA | Viruses |
| TLR4 | A896G or D299G C1196T or T399I are non-synonymous SNPs and are linkage disequilibrium [525] | D299G TLR4 polymorphism is associated with increased incidence of gram negative bacteraemia and sepsis [528]. This SNP is also linked to Mediaterranean spotted fever caused by Rickettesia conorii [527]. Humans with T399I TLR4 polymorphism milder hyporesponsive phenotype to LPS [528]. Cosegregation of both D299G and T399I TLR4 SNPs more severe hyporesponsive enss to LPS and higher incidence of sepsis and severe form RSV infection [[529], [530], [531]] | Plasma membrane | LPS, Taxol, Fusion protein, Envelope proteins, HMG-B1, Hsp60, Hsp70, Hsp22, Hsp96, Type III repeat extra domain A of fibronectin, oligosaccharides of hyaluronic acids, polysaccharide fragments of heparin sulphate, fibrinogen, saturated fatty-acids and Fetuin-A | Gram negative bacteria, Plant, RSV, MMTV, Chlamydia pneumoniae, Chlamydia trachomatis, host |
| TLR5 | C1174T (R392) (replacement of sequence encoding Arg392 with a stop codon) Present in 10% of European population [532,533] |
People with this SNP are more prone to develop Legionella pneumophila infections or Legionnaires' disease (Legionellosis) [534] and recurrent cystitis [535]. | Plasma membrane | Flagellin | Bacteria |
| TLR6 | A1401G (a TLR6 promoter SNP) C744T | Associated with high risk of prostate cancer in patients homo or heterozygous for A1401G [394], People with TLR6 C744T exhibit protection against asthma [536] | Plasma membrane | Di-acyl lipopeptides, Zymosan | Mycoplasma |
| TLR7 | Intron I, c.IT-120G | c.1-120G TLR7 allele protects male patients to develop inflammation and fibrosis during chronic HCV-infection [537] | Endolysosome | ssRNA, loxoribine, bropirimine | Viruses, synthetic compounds |
| TLR8 | 129G | Male patients infected with chronic HCV infection exhibit less inflammation due to less IFN-α production [538] | Endolysosome | ssRNA | Viruses |
| TLR9 | G-1174A and A1635G | People with these two SNPs in TLR9 exhibit rapid progressor phenotype for HIV-1 infection [539,540] | Endolysosome | CpG oligodeoxyneucleotide (ODN), Hemozoin pigment | Bacteria and viruses (HSV), Malaria |
| TLR10 | ND | ND | Endolysosome | ND | ND |
| TLR11 | NA | NA | Endolysosome | Profilin-like protein | Toxoplasma gondii |
| TLR12 | NA | NA | Endolysosome | Profilin-like protein | Toxoplasma gondii |
| TLR13 | NA | NA | Endolysosome | 23s ribosomal RNA | Bacteria |
Innate immunity plays a major role in the immunopathogenesis of inflammation and inflammatory diseases including sepsis [17]. As mentioned previously, all the innate immune cells that is macrophages, neutrophils, DCs, mast cells, NK cells (i.e. human NK cells express TLR2, TLR3, TLR5, TLR7/8 and TLR9), epithelial and endothelial cells etc. express various TLRs, which help in the recognition of PAMPs in extracellular as well as intracellular environment [[64], [65], [66], [67], [68], [69], [70], [71], [72],86,87]. Thus, TLR mediated recognition of pathogens by innate immune cells plays a very important role in the induction of pro-inflammatory immune response required to clear the infection. And under certain circumstances, this response can be uncontrolled and exaggerated leading to the development of severe systemic inflammation and sepsis. LPS or endotoxin acts as a major PAMP of gram-negative bacteria and is recognized by TLR4, while lipoteichoic acid (LTA), peptidoglycan (PGN) are the major PAMPs of gram-positive bacteria recognized by TLR2. Additionally, TLR4 is also responsible for cellular recognition of various other molecules including plant dipterene paclitaxel or Taxol (an anticancer agent), fusion proteins (i.e. fusion protein of respiratory syncytial virus (RSV), fibronectin, fibrinogen, heat shock proteins (HSp60 and Hsp70) and hyaluronic acid [32,82,88] (Table 1). However, TLR4 and TLR2 also act as receptors for high-mobility group box protein 1 (HMGB1) that is an alarmin released by cells dying from necrosis or necroptosis but not from apoptosis during acute infections and at later stages of sepsis [[89], [90], [91], [92], [93]]. Thus, apart from PAMPs, TLR signaling can also be activated by molecules generated and released by host cells in the extracellular environment.
4. Different pattern of expression of TLRs on various immune cells playing important role in the pathogenesis of inflammatory diseases
4.1. Endothelial cells (ECs) and their pattern of TLR expression
The TLR mediated activation of innate immune response varies from cell types that is when TLRs (i.e. TLR4 and TLR1/TLR2) expressed on monocytes and macrophages are treated with LPS or Tripalmitoyl-S-glyceryl-cysteine (Pam3Cys, a lipopeptide) they release TNF-α and IL-1β robustly but ECs (i.e. human umbilical vein ECs (HUVECs) and human lung microvascular ECs) under similar condition produce very little or no TNF-α or IL-1β, instead they express IL-6, IL-8, CSF2 (Colony stimulating factor 2), CSF3 (colony stimulating factor 3), ICAM-1 (Intercellular adhesion molecule 1) and SELE (Selectin E or CD62L) genes, which are required for neutrophil chemotaxis and migration [[94], [95], [96]]. While, almost similar amount of IL-6 and IL-8 cytokines are produced by primary human endothelial cells (ECs) and monocytes [94,97]. This may be explained as a divergent role of extracellular signal-regulated kinase1/2 (ERK1/2) during inflammatory activation of ECs and other innate immune cells that is monocytes and macrophages etc. [98]. For example, ERK5 activation plays an important role in HUVECs, primary human lung microvascular ECs and human monocytes activation upon their stimulation with TLR2 agonists [98]. Whereas MEK1 (MAP2K1, mitogen-activated protein kinase kinase 1) negatively impacts the TLR2 signaling in ECs but promotes TLR2 signaling in monocytes [98]. Thus, ERK5 acts as a key regulatory kinase of TLR2 signaling in ECs and monocytes and forms the basis of fundamental differences in TLR signaling pathways between ECs and monocytes [98]. Thus, ECs upon stimulation with TLR signaling pathway act differently from monocytes and macrophages. For example, ECs secrete IFN-β, IL-1α, IL-6, IL-10, IL-28, IL-29, granulocyte-colony stimulating factor (G-CSF), and granulocyte-monocyte-colony stimulating factor (GM-CSF) as major cytokines [77,94,[98], [99], [100], [101], [102], [103], [104]], while leukocytes (that is monocytes, neutrophils and DCs etc.) secrete, G-CSF, IL-1α, IL-1β, IL-2, IL-6, IL-9, IL-10, IL-12 (p35, p40, p70), IL-13, IL-15, IFN-α, IFN-β, IFN-γ, TGF-β1 and TNF-α upon TLR stimulation [[105], [106], [107], [108], [109], [110]].
Another major difference between ECs and leukocytes is that TLR2 and TLR4 are only expressed on cell surface of these leukocytes (i.e. monocytes, macrophages, neutrophils, DCs and NK cells) but in ECs these (i.e. TLR2 and TLR4) are also expressed intracellularly [98,111]. Thus, the LBP (lipid binding protein) mediated formation of LPS-CD14 complexes and their internalization and delivery to intracellular MD2-TLR4 complex initiates intracellular TLR4 activation in these cells [111]. However, the recognition of low to moderate amount of LPS (0.1–10.0 ng/ml) by ECs requires soluble CD14 (sCD14) [111] but it is not required for high levels (≥1 μgm/ml) of LPS detection. CD14 acts as a co-receptor and is present in association with the TLRs in cell surface and for other TLRs also on cell types including macrophages. TLR7 and TLR9 also require CD14 for the uptake of virus and induction of pro-inflammatory cytokines during viral infections including vesicular stomatitis virus infection [112]. Also, human endothelial cells including HUVECs, human coronary artery endothelial cells (HCAECs), human microvessel endothelial cells from brain (HMVEC-Brain), HMVEC-Liver and HMVEC-Lungs do not express TLR8 but human monocytes express higher levels of TLR8 [69]. While HMVECs-liver do not express TLR2 and relatively low levels of TLR2 are found on MHVEC-brain [69]. In addition, the expression of various TLRs (i.e. TLR5, 6, 7, 9, 10) on these ECs is very low as compared to their expression on primary human monocytes [69]. However, human monocytes express very low levels of TLR3 as compared to human endothelial cells [69]. Thus, this variation in expression of TLRs on ECs can impact the pathogenesis and outcome of certain inflammatory disorders where ECs are involved that is sepsis-induced vascular leakage and chronic inflammatory disease [[113], [114], [115]].
4.2. Natural Killer (NK) cells and TLR expression
NK cells are important innate immune cells with an essential role in tumor immunity, antibacterial immune response, anti-viral immune response and human pregnancy [[116], [117], [118], [119], [120], [121], [122]]. The killer immunoglobulin-like receptors (KIRs), the Ly49 receptors, and the CD94/NKG2 (Natural Killer Group 2) receptors are the major families of receptors used by NK cells for their immunological function and education [123,124]. All human and mice NK cells express NKG2D receptors, which are C-type lectin-like, type II transmembrane glycoproteins and regulate NK cell activation during viral infections and different cancers via binding to corresponding ligands [125,126]. Both activating and inhibitory Ly49 receptors are expressed by murine NK cells but not by human cells [127,128]. Although humans have a gene for Ly49 that appears to be pseudogene but baboons express it [129]. KIRs are the major recognition receptors for primates including humans [122]. KIRs regulate NK cell cytotoxic activity via recognizing the human leukocyte antigen (HLA) and binding to these molecules expressed on target cells [[130], [131], [132]]. Till date fourteen receptors in KIR family have been recognized including seven inhibitory receptors (KIR3DL1, 3DL2, 3DL3, 2DL1, 2DL2, 2DL3, and 2DL5), six activating receptors (KIR3DS1, 2DS1, 2DS2, 2DS3, 2DS4, and 2DS5) and one KIR2DL4 (with both inhibitory and activating action) receptor [[133], [134], [135]]. Along with these receptors NK cells also express TLRs that are involved in the recognition of viral and bacterial PAMPs [87,130,136]. However, studies have also shown an interaction between KIRs expressed on NK cells and TLR ligands [137]. The treatment of NK cells with CpG ODN (oligodeoxynucleotide) downregulates KIR3DL2 inhibitory receptors on their surface from the cell surface [137]. Only KIR3DL2+ NK cells release IFN-γ upon stimulation with CpG ODN suggesting that KIR3DL2 plays an important role in the recognition of CpG ODN by NK cells [137]. Furthermore, in human NK cells, extracellular recognition of CpG ODN by surface KIR3DL2 plays a role in the internalization of CpG ODN where it is recognized by endosomal TLR9 expressed in early endosomes [137]. Thus, KIR (i.e.KIR3DL2) expressed on NK cells helps TLRs in recognizing PAMPs and mounting a significant immune response to take care of the infection. However, this stimulation of TLR9 with CpG ODN did not impact the expression of other NK cell surface receptors (i.e. NKG2D, NKG2A, CD16, CD56, DNAM-1, CD2, LFA-1, 2B4, and NTBA) [138,139]. The expression of activating receptors and their ligands on NK cells is also affected by TLR stimulation [140]. This process is seen during the killing of macrophages infected with Mycobacterium tuberculosis by NK cells to clear the pathogen via NKp46, NKp30 and NKG2D overexpression on NK cells and ULBP1 (a ligand of NKG2D) on infected macrophages due to activation of TLR2 [141,143]. Thus, TLR stimulation controls both inhibitory and activating receptors on NK cells.
The pattern of expression of TLRs on human NK cells is also different from monocytes and macrophages, for example, TLR1 is most highly expressed of all TLRs on isolated NK cells at mRNA levels, while mRNAs of other TLRs including TLR2, TLR3, TLR5, and TLR6 are expressed at moderate levels [143,144]. Even stimulation of NK cells via TLR2 and TLR5 activation through outer membrane protein A from Klebsiella pneumoniae (KpOmpA) and flagellin cause synthesis and release of α-defensins and antimicrobial peptides (AMPs) from these cells [144]. Thus, NK cells are capable of direct recognition of bacterial PAMPs and secrete AMPs that exert direct cytotoxic antibacterial action. An increased number of NK cells has also been seen in the joints of patients having rheumatoid arthritis (RA) [145]. Majority of these NK cells are CD56high with higher levels of CD94/NKG2D and lower levels of KIRs and CD16 receptors [145]. Fibrinogen, HSP 60 and 70, EDA (Alternatively spliced domain A) fibronectin, HSP22 (Heat shock protein 22), HMGB1 are various TLR2 and TRL4 ligands that are elevated in synovial tissues of RA patients [146]. NK cells found in inflamed joints of RA patients may act as important inflammatory cells along with synovial macrophages and DCs due to the expression of both TLR2 and TLR4 at the cell surface as well as intracellularly [147].
TLR9 mRNA is absent in CD3−CD56low and CD3−CD56high NK cells [144]. Interestingly, resting human NK cells highly express mRNA of TLR2 but the mRNAs of TLR4 and TLR3 are weekly expressed, while the mRNA of TLR8 is absent [148]. However, upon stimulation with viral antigens, the levels of mRNAs of all these 4 TLRs (TLR2, TLR4, TLR3 and TLR8) were highly elevated [149]. Human uterine NK (uNK) cells express TLR2 at both cell surface and intracellularly [150]. However, the expression of TLR1, TLR2, TLR3, TLR4 and TLR9 at the protein level is reported in adult human NK cells of both CD56high or CD56low subtypes and like ECs, TLR2 and TLR4 are also expressed intracellularly in both human and murine NK cells [148,[151], [152], [153]]. TLR9 is also found to be expressed intracellularly in NK cells [147]. TLR9 is shown to play an important role in the pathogenesis of type 1 diabetes mellitus (T1DM) [154,155]. NK cells are shown to play role in the T1DM [145,156]. Thus, it will be interesting to study the role of TLR9 signaling in NK cells in the pathogenesis of T1D. The mean fluorescent intensity (MFI) of intracellular TLR2 in CD56high and CD56low NK cells increases in patients with sepsis as compared from healthy controls [153]. Thus, the modulation of TLR expression in NK cells has been noticed in patients suffering from sepsis.
The activation of NK cells by TLR ligands is also different from other innate immune cells that are monocytes/macrophages/neutrophils. For example, for cytokine production by both murine and human NK cells along with presence of TLR ligands [i.e. LPS, CpG-DNA or Pam(3)CysSK(4)] they also require accessory cytokines (i.e. IL-15 + IL-18, IL-2 + IL-12, IL-2 + IL-18) to produce IFN-γ or GM-CSF [152,153]. And IL-2 is the only cytokine capable of stimulating NK cells to produce these cytokines in the absence of any TLR ligand [148]. Sometimes TLR ligand-dependent activation of NK cells requires the contact of NK cells with accessory cells that are DCs to produce cytokines, which is mediated by a newly identified interferon regulatory factor-3 (IRF-3)-dependent NK activating molecule [157,158]. The LTA mediated stimulation of human NK cells causes an expression of CD25 and CD69 on these cells along with the release of IFN-γ, indicating the presence of active TLR2 (CD282), causing activation of NK cells in response to TLR2 ligands [159]. Thus, involvement of TLRs expressed on NK cells in the production of IFN-γ upon ligation with their corresponding PAMPs along with the presence of cytokine released by other innate immune cells (i.e. monocytes, neutrophils, mast cells, DC, ECs etc.) plays an important role in the induction of inflammatory immune response including sepsis and its outcome [[160], [161], [162], [163]]. For example, freshly isolated human peripheral blood NK cells in the presence of IL-12 become activated by dsRNA (TLR3 ligand) or unmethylated CpG DNA (ligands for TLR9) [164]. This is indicated by the induction of the expression of CD69 and CD25 (activation markers) [164]. This causes an increased release of IFN-γ and TNF-α and cytolytic action of these NK cells against tumor cells [164]. Thus the use of various TLR agonists to stimulate NK cells has a great potential to serve as adjuvants for NK cell-based immunotherapy against several cancers. On the other hand, this TLR-mediated early recognition of pathogens by NK cells can prove helpful to mount an effective early innate immune response during microbial infections.
4.3. Dendritic cells (DCs) and TLR expression
DCs are another class of innate immune cells, which are classified into two different subtypes: a) CD11c positive myeloid DCs (mDCs) and, b) CD11c negative plasmacytoid DCs (pDCs) in peripheral circulation [165]. pDCs account for 0.1% of all blood mononuclear cells in humans and upon pathogen exposure they secrete number of cytokines including IL-12, TNFα, Regulated on activation normal T cells expressed and secreted (RANTES or CCL5), IL-10, interferon-gamma-induced protein-10 (IP-10 or CXCL10), IL-6, macrophages inflammatory protein (MIP)-1α and MIP-1β [[165], [166], [167], [168], [169]]. In pDCs all TLR7 and TLR9 dependent innate immune responses are mainly mediated by MyD88 activation and the formation of a complex between MyD88, TRAF6 and IRF-7 [170,171]. Formation of the MyD88 complex with IRF7 involves the interaction of death domain of MyD88 with an inhibitory domain of IRF7 causing an activation of IFN-α-dependent promoters [170]. Whereas, adaptor molecule TRAF6 also binds to IRF7 and ubiquitin ligase activity of TRAF6 plays an essential role in activation of IRF7 and release of IFN-α [170]. However, in addition to MyD88, TRAF6 and IRF-7, IRAK4 also plays an important role in activation of IFN genes and secretion of IFN-α and IFN-β via TLR9 pathway activation in pDCs [171]. This complex comprising of MyD88, TRAF6, IRF-7, and IRAK4 has been considered as a cytoplasmic transductional-transcriptional processor (CTTP) [171]. However, in addition to this CTTP complex, an interaction of IRF-7 with IRAK-1 is required for effective production of IFN-α [172]. This is because upon TLR7 and TLR9 activation these pDCs produce only IL-12 and IL-6 with no IFN-α in the absence of IRAK-1 [173]. While in the absence of IRF-5, pDCs secrete normal levels of IFN-α but diminished the amount of IL-12p40 and IL-6 in response to CpG [173]. However, mDCs can be categorized further depending on differential surface expression of CD1c (BDCA1, Blood dendritic cell antigen 1), CD16 and BDCA3 and CD16-mDCs comprise largest population (65–75% of total mDC population) of mDCs in peripheral circulation in humans, followed by CD1c-mDCs (10–20%) and then BDCA3-mDCs (3–5%) [174,175]. Human mDCs express all TLRs except TLR9 that is not expressed by these types of DCs, while pDCs in humans lack TLR2, TLR3, TLR4, TLR5, TLR6, and TLR8 but they express TLR10 and higher levels of TLR9 [166]. It should be noted that mDCs lack TLR9, while pDCs express highest levels of TLR9 [166]. However, human DCs derived by monocytes (moDCs) during in vitro culture express all other TLRs except TLR9 and TLR10 and they express higher levels of TLR4 as compared to CD1c-mDCs [166]. mDCs produce IL-12 and moDCs produce type 1 IFNs upon stimulation with LPS via activation of TLR4-dependent signaling pathway [176,177]. In DCs binding of LPS to CD11b (due to low expression of CD14 on DCs) initiates activation of TLR4 signaling [178,179]. Thereafter, this TLR4 downstream signaling in DCs further requires MAP3K apoptosis signal-regulating kinase 1 (ASK1 or MAP3K5) to activate p38α for optimal secretion of pro-inflammatory cytokines (i.e. TNF-α, IL-1 and IL-6) [180]. Mice lacking ASK1 are resistant to develop LPS induced septic shock and arthritis [181]. ASK1 acts by forming a complex with TRAF2 and TRAF6 in response to LPS mediated generation of reactive oxygen species (ROS) [182]. This is because during steady state ASK1 forms an inactive complex with thioredoxin and TLR4 stimulation breaks this complex and free ASK1 binds to TRAF2 and TRAF6, causing its activation via autophosphorylation [183]. While, stimulation of TLR1, 2 and 6 on DCs causes their maturation and secretion of several cytokines that is IL-6, IL-8, IL-10, IL-12 and TNF-α [166].
Loss of DCs in circulation, spleen, lymph nodes (i.e. mesenteric lymph nodes) and various organs is associated with immunoparalysis observed in patients dying of sepsis [184,185]. Treatment of animal models of sepsis with FMS-related tyrosine kinase 3 ligand (FLT3L) (a DC growth factor) restored the DC function via enhancing releases of certain cytokines that is IL-12, IL-15 and IFN-γ along with augmenting functions of neutrophils, NK and CD4+ T cells and increased the survival of the experimental animals [[186], [187], [188]]. The protective effect of FLT3L was observed even after the adoptive transfer of DCs treated with FLT3L into the septic animals [189]. Intrapulmonary transfer of bone marrow-derived DCs (BMDCs) also prevented the development of fatal pulmonary aspergillosis in mice recovering from sepsis [190]. Animals recovering from sepsis are more prone to develop severe secondary infections due to prolonged immunosuppression via mechanisms involving transforming growth factor β (TGF-β) secreted by paralyzed DCs causing accumulation of regulatory T cells (Tregs) in lungs and the development of tolerogenic DCs expressing Blimp1 [191]. Thus, restoration of normal functioning of host DCs is very important to prevent immunoparalysis or immunosuppression observed in septic patient and further to prevent the patients recovering from sepsis to catch the severe secondary infection. This can be better understood by studying the TLR mediated regulation of DCs as TLR agonists have shown an important improvement in survival of animals suffering from acute pneumonia after hemorrhagic shock [192,193]. TLR agonist treatment might have prevented the generation of tolerogenic DCs or prevented the release of TGF-β from these tolerogenic DCs via increased expression of MHC class II molecules and co-stimulatory receptors that is CD80 and CD86 on DCs [[191], [192], [193]]. Thus, regulation of TLR mediated function of DCs plays an important role in the pathogenesis of both infectious and inflammatory diseases and their outcome in terms of survival. Thus, restoration of DCs during sepsis seems to be one of the primary therapeutic approaches.
4.4. Gamma Delta (γδ) T cells in inflammatory diseases and their expression pattern of TLRs
γδT cells are very important cells of the mammalian innate immune system and play a critical role in early response to invading pathogens, autoimmunity (i.e. Inflammatory Bowel Disease (IBD), psoriasis and asthma etc.) and neurodegeneration along with tissue homeostasis and repair [[194], [195], [196], [197], [198], [199]]. γδT cells constitute minor (2%–10% of CD3+ T cells) but a very important component of the T-cell population in peripheral circulation in humans. Depending on the expression of T cell receptor (TCR) human γδT cells can be classified into two major categories that are Vδ1 and Vδ2 γδT cells [200]. Vδ1 γδT cells constitute the major population of residential T cells in mucosal tissues (i.e. intestines (25–60%), respiratory tract and reproductive tract etc.) and skin environment of humans [[201], [202], [203], [204], [205], [206], [207], [208]], where they provide the first line of defense against invading pathogens and recruitment of inflammatory cells [209,210]. The murine skin Vδ1 γδT cells are also known as Dendritic Epidermal T cells (DETC) [211], which are present in the epidermal layer of the skin [208]. While Vδ2 γδT cells are mainly present in peripheral blood circulation [207]. In patients with sepsis the population of Vδ2 γδT or CD3+CD56+ γδT cells in circulation is significantly reduced [212]. This decrease in CD3+CD56+ γδT cells starts from day second of sepsis and progresses till the death of patient [212]. Only non-proliferative effector subset of γδT cells can be observed in septic patients at very low levels of 0.61% as compared to healthy controls (19.2%) [213]. γδT cells have shown a great potential to mitigate the organ injury and mortality associated with sepsis in animal models [214]. γδT cells also play an important role in modulating the systemic inflammatory response during severe burn injuries [215]. In addition, γδT cells play an important role in the pathogenesis of invasive infections [216], which can later lead to the development of sepsis. Thus, TLR mediated activation of γδT cells should have also played an important role in the pathogenesis of inflammatory diseases including sepsis. The expression of TLRs on non-stimulated or resting γδT cells is hard to detect, but they are expressed once these cells get activated by danger signals like mitochondrial DAMPs (MTDs) (induce TLR2 and TLR4 expression on γδT cells) [217,218], cytokines or alarmins [216]. All the TLRs found in humans have been found to be expressed by activated γδT cells [216].
However, the pattern of expression of TLRs on these γδT cells varies depending on the subtype of these cells. For example, activated Vδ1 and Vδ2 γδT cells both express higher levels of TLR4 and TLR5 [[219], [220], [223]] and are capable of exerting potent antibacterial immune response. While, freshly isolated non-activated human γδT cells do not express TLR3 on their cell surface but get upregulated on the cell surface after 24 h TCR-mediated stimulation [221,222]. But same has not been observed upon incubation of freshly isolated human γδT cells with TLR3 ligands including double-stranded RNA (dsRNA) viruses such as reoviruses, a synthetic analog of dsRNA, polyinosinic-polycytidylic acid [poly(I:C)], and small interfering siRNA [221,222]. TLR1, TLR2, and TLR6 have been found to be expressed on human peripheral blood γδT cells or Vδ2 γδT cells [220,223]. TLR7 and TLR8 are found to be expressed intracellularly in endosomal compartments of both Vδ1 and Vδ2 γδT cells [220]. TLR9 is undetectable in human γδT cells, while TLR10 has been detected on these cells [224,225]. Thus, TLRs are expressed on these innate immune cells and play an active role in the detection of pathogens and associated PAMPs as well as DAMPs directly.
4.5. Myeloid-derived suppressor cells (MDSCs) in inflammatory diseases and the pattern of TLR expression
MDSCs comprise a heterogeneous cell population of immature myeloid cells consisting of myeloid progenitor cells and progenitors of monocytes/macrophages, neutrophils and DCs with strong suppressive action on T cell functions [226]. In mice these MDSCs are categorized in two different populations: (1) monocytic MDSCs or M-MDSCs that are CD11b+ Ly-6G− Ly-6Chigh cells and (2) granulocytic MDSCs or G-MDSCs having CD11b+ Ly-6G+ Ly-6Clow phenotype [227]. However, in cancer patients, MDSCs are defined by the expression of CD33 or Siglec-3 (sialic acid binding Ig-like lectin 3) or gp67 or p67 and lacking phenotypic markers for mature myeloid and lymphoid cells [228]. However, with the advancement of tumor immunology and MDSCs biology different phenotypes of these MDSCs in tumor environment have been described and are discussed in more detail elsewhere [[229], [230], [231]]. In addition to their role in tumor immunology, they also play role in infectious diseases, autoimmunity, obesity and human pregnancy [232]. Thus, MDSCs also comprise a population of important immune cells in various inflammatory processes and diseases.
The major focus of this section is on the pattern of TLR expression on MDSCs. Studies have shown that MDSCs express TLR2, TLR4, TLR1, TLR6, TLR5, TLR3 [[233], [234], [235], [236], [237]]. Thus, expression of these TLRs by MDSCs also made them an important innate immune cells as these PRRs recognize microbial PAMPs and host-derived ligands/DAMPs (i.e. HMGB1, Hsp60, Hsp70 etc.) released during severe infection including sepsis or other potential inflammatory diseases [238]. These MDSCs protect the host from inflammatory tissue/organ damage in the course of acute infection/sepsis at its early/initial stage via suppressing the inflammatory pathway [239,240]. Anti-Gr1 antibody treatment at early stages of acute infection or sepsis indicated their immunosuppressive action as this treatment decreased the effective number of MDSCs and prevented the development of Th2 immune response and suppressed CD4+ and CD8+ T cell associated immune response during sepsis [240]. Further studies in this field established MDSCs as immunoprotective cells rather than immunosuppressive innate immune cells during the early phase of sepsis because their depletion at this stage increased the mortality among experimental animals [241,242]. However, a study further showed that accumulation of these G-MDSCs in the spleen of mice subjected to sepsis at least after 3 days of onset of sepsis and continued to increase even till 12 weeks in mice that survived sepsis [235]. This accumulation of MDSCs was also recognized in other secondary lymphoid organs that are peripheral lymph nodes including inguinal and axillary lymph nodes [235]. This late-stage accumulation and activation of MDSCs in the spleen, bone marrow, and peripheral lymph nodes can have immunosuppressive action on host T cells and may be responsible for the development immunoparalysis due to inhibition of Th1 immune response and skewing it towards Th2 immune response during the late stage of sepsis. However, another study showed that adoptive transfer of MDSCs isolated from septic mice on day 10 into the mice with sepsis at an early stage increased their survival via directly killing the bacteria and inhibiting the release of pro-inflammatory cytokines [241]. Thus, early activation of MDSCs during sepsis can be protective but at late stages may lead to the stage of immunoparalysis that proves detrimental to host. Thus, the exact role of MDSCs in sepsis as a friend or foe is still a topic of debate [[243], [244], [245]]. However, activation of MDSCs via TLR3, TLR7, TLR8 and TLR9 ligands (i.e. poly I:C, CpG ODN, TLR7/8 agonist 3 M-052 etc.) causes inhibition of their immunosuppressive action [236,237,246]. This can serve as a potential cell-specific therapy at later stages of sepsis where immunoparalysis is developed due to increased frequency and overactivation of MDSCs and suppression of T cell-based immunity.
4.6. Platelets and TLR expression pattern
Platelets are mainly characterized in mammalian species as anucleate blood cells responsible for clot formation, atherosclerosis and blood coagulation during trauma to prevent excess blood loss [[247], [248], [249]]. However, further studies in the field of platelet biology established their role in angiogenesis, development, antimicrobial immune response and direct killing of potential pathogens, neurodegeneration, stroke, cancer and metastasis [247,[250], [251], [252], [253]]. TLR4 KO mice have shown defects in their circulating and reticulated platelet counts [254,255]. However, here my major focus is the pattern of TLR expression on platelets and its impact on inflammatory diseases.
Both mouse and human platelets express TLR1, TLR2, TLR4, TLR6 and TLR9 [256,257]. A further study in 2005 showed a weak expression of TLR2, TLR4, and TLR9 on the surface of human CD41+ platelets [258]. But the expression of TLR1, TLR6, and TLR8 on human platelets is very low [258]. Additionally, these CD141+ human platelets highly expressed TLR2, TLR4, and TLR9 in their cytoplasm [258]. These TLRs are functional on platelets as stimulation of platelets with LPS or other TLR-ligands caused thrombocytopenia and production of TNF-α and other potential mediators of inflammation (i.e. thromboxane, tissue factor, pro-inflammatory cytokines etc.) under in vivo conditions [254,256,[259], [260], [261], [262], [263], [264], [265], [266], [267]]. Platelet TLRs are not involved in the recognition of Streptococcus pneumoniae (S. pneumoniae) instead platelets are activated by S. pneumoniae independent of TLR stimulation [268]. Platelet MyD88-dependent TLR signaling does not play any role in host defense and induction of pro-inflammatory immune response against non-capsulated S. pneumoniae in vivo [268]. Thus, platelets mediated TLR signaling and clearance of S. pneumoniae cannot take place in infants and children who are more prone to develop pneumonia and associated sepsis. This is because S. pneumoniae is one of the major cause of childhood pneumonia and associated sepsis all over the world. However, an activation of platelets via TLR stimulation during bacterial infection/sepsis stimulates the clearance of potential pathogens [specifically, Staphylococcus aureus (S. aureus) and Bacillus cereus (B. cereus)] [263].
In addition, platelet TLR2 is involved in histone-induced thrombin generation during sepsis as histones are released from several apoptotic cells [263]. Even careers of TLR4 polymorphism Asp299Gly alone or in a combination of another polymorphism Thr399Ile have shown reduced thromboxane A2 (TXA2) (the main product of platelet arachidonic acid metabolism occurring due to the activation of cyclooxygenase 1 (COX-1), which exerts prothrombotic and proatherogenic effect) [260]. Thus, careers of this TLR4 polymorphism may have protective phenotype against cardiovascular diseases like atherosclerosis and other clot formation disorders due to reduced production of TXA2 by platelets [260]. Hence, the expression pattern of TLRs on platelets is unique in terms of their function and their role in the pathogenesis of inflammatory diseases including sepsis and other chronic inflammatory disorders. This is because platelets and TLRs expressed by these cells play a potential role in the process of inflammation, development of neutrophil extracellular traps (NETs) and several inflammatory diseases including atherosclerosis, sepsis [[269], [270], [271], [272], [273]]. For example, both platelet TLR4 and TLR2 are involved in its interaction with neutrophils causing the formation of a heterotypic complex with neutrophils [259,275].
The stimulation of platelet TLR2 also increases surface expression of P-selectin, activation of integrin alpha(IIb)beta(3) and generation of reactive oxygen species (ROS) playing an important role in inflammation [274]. Furthermore, an in vivo infection with Porphyromonas gingivalis (P. gingivalis) failed to form platelet-neutrophil aggregates in TLR2−/− KO mice [274]. This Platelet–neutrophil interaction involves P-selectin expressed by platelets and neutrophil PSGL1 (P-Selectin Glycoprotein Ligand 1) or CD162 [252]. During hyperlipidemia activation of platelet TLR2 via oxidized phospholipids [oxPCCD36, oxidized phospholipid bound to CD36 or platelet glycoprotein 4 or fatty acid translocase (FAT)] causes hyperactivation of platelets via activation of TIRAP (Toll-interleukin 1 receptor domain containing adaptor protein)-MyD88-IRAK (interleukin-1 receptor-associated kinase)1/4-TRAF6 (TNF receptor-associated factor 6), causing an activation of integrin through Src family kinase (SFK)-spleen tyrosine kinase (Syk)-PLCγ2 (phospholipase Cγ2) pathway [275]. This hyperactivation of platelets via TLR2 during hyperlipidemia is responsible for the development of thrombosis [275]. In addition to TLR2, an activation of TLR9 is also involved in the oxidative stress-induced generation of thrombosis via carboxy(alkylpyrrole) protein adducts (CAPs), that act as novel unconventional ligands for TLR9 [276]. Thus, studies investigating the role of TLRs expressed on platelets in various immune processes during homeostasis and inflammatory diseases have started to highlight their role in platelet biology and inflammatory process. Future studies will further strengthen the importance of TLRs in platelet biology and inflammatory diseases.
4.7. Mast cells and TLR expression
Mast cells are sentinel innate immune cells and regulate both innate and adaptive arms of immune system along with other physiologic and pathophysiologic conditions [[277], [278], [279]]. However, the pattern of expression of TLRs on human and mouse mast cells (i.e. bone marrow-derived mast cells (BMMCs), connective tissue mast cells (CTMCs) and foetal skin-derived mast cells (FSDMCs) etc.) and impact of different TLR-ligands on them has been studied in detail and described elsewhere [71,[280], [281], [282]]. BMMCs from mice and mast cell line MC/9 derived from mice express mRNA for TLR2, TLR4, and TLR6 but not TLR5 and lack surface expression of CD14 [283]. These cells produce pro-inflammatory cytokines in response to LPS and other TLR ligands [283]. While human mast cells express TLR2, TLR4 and respond differently to LPS and LTA or PGN and secrete TNF-α, IL-5, IL-10, and IL-13 respectively [284]. However, the release of TNF-α from human cord blood-derived mast cells requires priming with IL-4 and serum as a source for sCD14 [284]. TLR4 mediated stimulation of murine mast cells via LPS operates through the activation of the only MyD88-dependent pathway, while MyD88-independent or TRIF dependent pathway is not functional [285]. Additionally, TRIF-dependent and MyD88-independent pathways of TLR4 stimulation seem to be operational only when TLR4 gets internalized into endosomes upon its stimulation [[286], [287], [288]]. This TLR4 internalization to endosome upon stimulation with LPS has not been recognized in mast cells and they lack type 1 IFNs release upon LPS stimulation due to the non-existence of TRIF-dependent TLR signaling [285,289]. This can be explained due to a reduced level of expression of TRAM (TRIF-related adaptor molecule) in mice BMMCs that is involved in connecting TRIF to TLR4 receptor complex upon TLR4 stimulation and inducing TRIF-dependent TLR4 signaling pathway and release of type 1 IFNs [285].
Additionally, mice mast cells do not express CD14 that is also required for TLR4 internalization [290,291]. Thus reduced TRAM expression and absence of CD14 on murine mast cells render them to internalize TLR4 upon its stimulation. Human neutrophils also lack TRIF-dependent activation of TLR4 signaling pathway as they also do not release type 1 IFNs upon LPS stimulation [292] but express fewer CD14 (albeit 10 times lesser copy numbers per cell in comparison to monocytes) [293]. Thus CD14 has a potential role in TLR4 internalization and TRIF-dependent release of type 1 IFNs. However more studies are required in this direction. TLR2 stimulation of human mast cells causes the release of histamine, which does not take place by LPS stimulation [284]. In addition to TLR4 and TLR2, human cultured mast cells express TLR-1, TLR-3, TLR-5, TLR-6, TLR-7, TLR-8, TLR-9 and TLR10 (at mRNA levels) [71,294].
Viruses and poly I:C mediated stimulation of TLR3 on human mast cells leads to the release of IFN-α and IFN-β but no TNF-α IL-1β, IL-5, or GM-CSF as released by stimulation with TLR4 or TLR ligands [294]. However mouse BMMCs and FSDMCs behave differently when stimulated with TLR3, TLR7 and TLR9 ligands that are poly I:C, R-848, and CpG ODN [282]. For example, upon stimulation of TLR3, TLR7 and TLR9 with these ligands only FSDMCs release TNF-α, IL-6, RANTES, MIP-1α or CCL3, and MIP-2 (CXCL2) but same molecules are not released by mouse BMMCs [282]. However, both types of mouse mast cells (BMMCs and FSDMCs) fail to degranulate (i.e. histamine, β-hexosaminidase or serotonin release) and release IL-13 in response to TLR3, TLR7 and TLR9 stimulation via poly I:C, R-848, and CpG ODN [282]. Also, mouse BMMCs are less potent than mouse peritoneal cell-derived mast cells (PCDMCs) in terms of their cytokines release upon TLR stimulation [280]. For example, PCDMCs release higher levels of pro-inflammatory cytokines that is IL-1, IL-6, IL-10, IL-17, TNF-α, GM-CSF and IFN-γ upon stimulation with TLR2 ligands LTA and MALP-2 (macrophage-activating lipopeptide-2, a 2-kDa synthetic derivative from Mycoplasma fermentans, which signals through TLR2 and TLR6) as compared to BMMCs [280]. This can be explained as immature BMMCs only express a truncated form of the TLR2 instead of the full-length TLR2 protein [280]. Thus, mast cells located at different places react differently to TLR stimulation. Activation of TLR3 on mast cells via double-stranded RNA (dsRNA) inhibits its attaching property to vitronectin (VN) and fibronectin (FN) and may decrease its response in IgE mediated mast cells stimulation [295]. Activation of TLR2 via LTA downregulates FcepsilonRI expression on human mast cells [296]. Thus, can we rule out that patients recovered from gram-positive bacterial sepsis may have protection against certain allergies caused by over-activation of mast cells due to downregulation of FcepsilonRI expression on mast cells or just this effect lasts only till the presence of TLR2 ligands or for longer duration? An interesting research area needed to be explored.
4.8. T cells and pattern of TLR expression on various subtypes of T cells
T cells [i.e. CD4+ and CD8+ and CD4+CD25+ regulatory T cells (Tregs) etc.] are major cells of the adaptive immune system and are present in peripheral blood circulation as well as in various target organs including skin, liver, lungs, kidneys, spleen etc. Various studies have shown T cells play a very important role in the process of inflammation varying from sterile to infectious to auto-immune inflammation [[297], [298], [299]]. Even these cells programme memory of inflammation depending on previous exposure to potential inflammogens [300]. For example, these CD4+ and CD8+ T cells in both compartments that is peripheral blood circulation and in various organs are affected by sepsis [[301], [302], [303], [304]]. These are significantly decreased to very low levels in patients who succumb to sepsis [[305], [306], [307]]. Even these T cells start to undergo apoptotic cell death within first 24 h of onset of sepsis [303,308,309]. Thus, prevention of this apoptotic T cell death and associated lymphopenia can prove beneficial in restoring protective innate immune response via the release of IFN-γ and IL-17 within first 24 h of sepsis onset [310]. While the number of CD4+CD25+ regulatory T cells (Tregs) in circulation increases and negatively impacts the tissue protective immune response against sepsis (i.e. induce increased apoptotic death of monocytes required to clear to pathogen from circulation) or severe infection leading to the stage of immunoparalysis and poor prognosis [[311], [312], [313], [314]]. However, adoptive transfer of in vitro-stimulated Tregs before or after 6 h of induction of sepsis in a mouse model has increased their survival via enhanced release of TNF-α and clearance of peritoneal bacteria [315]. While depletion of Tregs from circulation post three days of sepsis decreased the mortality among animal subjected sepsis [316]. Thus, the negative and positive impact of Tregs on the outcome of sepsis is determined by the time of Tregs based immunomodulatory approach.
Human peripheral blood T cells express almost all TLRs that is TLR1-TLR10 at mRNA levels but expression at the protein level is detected for only TLR2, TLR3, TLR4, TLR5 and TLR9 [[317], [318], [319]]. CD8+ T cells express higher levels TLR3 and TLR4, while CD4+ T cells express higher levels of TLR1 and TLR9 [318]. However, CD8+T cells isolated from infected person express higher levels of TLR2, where the expression of TLR9 is decreased on CD4+T cells isolated from same person [318]. While, human CD25+CD4+ Tregs express higher levels of TLR8, TLR5, and TLR2 [[320], [321], [322]]. Expression of TLRs on murine T cells also varies depending on the mouse strain for example, CD4+CD45RBhigh T cells isolated from C57BL/6 or B6 mice express TLR1, TLR2, TLR3, TLR6, TLR7 and TLR8 at mRNA levels while mRNAs of TLR4, TLR5 and TLR9 are either expressed at very low levels or are undetectable [323,324]. On the other hand, Naïve CD8+ T cells isolated from B6 mice express TLR1, TLR2, TLR6 and TLR9 at mRNA levels and the expression of TLR4 mRNA is undetectable [325,326]. CD4+ T cells isolated from BALB/c mice express TLR3, TLR4, TLR5 and TLR9 at mRNA levels [327] and only TLR3, TLR4 and TLR9 are detected on protein levels in these cells [328]. On the other hand, CD8+ T cells from these mice express TLR2 at protein levels [324,325].
The TLR expression on T cells is regulated by TCR-signaling, for example, TLR3 and TLR9 expression on CD4+T cells of BALB/s mice get upregulated upon TCR-stimulation [327]. Helper 1 T cells or Th1 cells are shown to be activated by TLR2 signaling upon ligation with Pam3CSK or LTA and release higher levels of IFN-γ in the presence of cytokines IL-2 or IL-12 [329]. Additionally, stimulation of CD4+T cells with mycobacterial proteins via TLR2 causes their differentiation into Th9 cells expressing IL9 mRNA as indicated by differentiation and proliferation of Th9 cells [330]. Furthermore, TLR2 signaling in CD4+T cells causes higher expression of BATF (Basic leucine zipper transcription factor) and PU.1 (a transcription factor that in humans is encoded by SP11 gene), both are the positive regulators of Th9 cell differentiation [330]. TLR signaling on T cells has also been shown to play an important role in Toxoplasma gondii (T. gondii) infections where MyD88 −/− mice exhibited increased mortality [331]. This can be explained due to defective TLR signaling with reduced Th1 effector responses, and reduced levels of IL-12 production required for efficient secretion of IFN-γ from Th1 cells [329,332]. TLR3 and TLR9 signaling via binding to their corresponding ligands that is Poly(I:C) and CpG DNA increase the survival of murine CD4+T cells, which can be explained due to activation of NF-κB and an increased expression of anti-apoptotic protein Bcl-XL [328,332]. Thus, this ability of T cells to directly recognize potential PAMPs via TLRs may play an important role in the pathogenesis of inflammatory disease associated with infections or sterile inflammation where DAMPs are recognized by potential TLRs.
4.9. B cells and pattern of TLR expression
B cells are antibody-producing cells of immune system and serve as a precursor of plasma cells responsible for secreting various antibodies (i.e. IgG, IgM, IgA, IgD, and IgE) as a humoral component of the adaptive immune system. However, these B cells also act as responsible cells for enhancing early innate immune response during bacterial sepsis [333]. For example, Rag1−/− mice deficient in B cells display decreased early pro-inflammatory immune response (for example, decreased levels of cytokines and chemokines (TNF-α, IL-1β, IL-6, IL-10, KC or IL-8, RANTES and MCP-1) during bacterial sepsis and show increased mortality [333]. More interestingly this defect is only shown by B cell-deficient or anti-CD20 B cell-depleted mice, but not by α/β T cell-deficient mice [333]. Further studies have shown that B cells get activated during sepsis as early as 24-h post-CLP-induced sepsis in mice as increased expression of CD69 on the B220+CD19+ cells in spleen, bone marrow (BM), and mesenteric lymph nodes (MLNs) is observed [333]. Whereas the majority of peritoneal B1 (considered as innate-like immune cells and develop early during ontogeny as compared to conventional B-2 cells) cells get depleted from the peritoneal cavity within 36 h after induction of sepsis in mice models and migrate to spleen and lymph nodes after activation [333,334].
Marginal zone (MZ) B cells (also considered as innate-like immune cells similar to B1 cells) are first B cells to encounter blood-borne pathogens during bacteremia due to their primary location at splenic marginal sinuses, a site where macrophages and DCs capture antigens [[334], [335], [336]]. Sepsis robustly activates both MZ and follicular B cells, leading to induction of both germinal cell reaction and plasma cell formation [333]. Also both these MZ (B220+IgM+IgD−CD21+CD23low) and follicular (B220+IgD+IgMlowCD23hi) B cells get decreased during sepsis [333]. However, MZ B cells are worst affected as compared to follicular B cells [333]. The early alteration in B cells number and response is also observed in human patients of severe sepsis and septic shock that is responsible for increased chances of secondary infection in elderly patients with severe sepsis [337,338].
Studying the expression patterns of TLRs and impact of TLR signaling in the functioning of B cells stimulated various researchers to investigate this pathway due to the role of these cells in infection and their potential application in vaccine design [339]. TLR expression on B cells also has a different pattern from other immunological cells, for example, murine B cells express TLR4 even at their normal steady state and are responsive to LPS [340]. This LPS mediated stimulation of TLR4 on murine B cells initiates their proliferation, cytokine release and class switch recombination (CSR) [340]. However, human B cells do not express TLR4 and remain unresponsive to LPS, thus gram-negative bacteria cannot initiate TLR4 signaling at least in human B cells [317,340]. Similarly, TLR2 is expressed on murine B cells and initiates strong immune response upon binding with its cognate ligand that is LTA or PGN. While, human B cells also express TLR2 but here it requires sensitization via cross-linking of the BCR with anti-immunoglobulin or protein A from Staphylococcus aureus (S. aureus) [341,342]. In both human and murine B cells, TLR2 is expressed along with its co-receptors TLR1 and TLR6, but an expression of CD36 is limited to only murine B cells and TLR10 to only human B cells [[343], [344], [345]]. TLR7 and TLR9 are both expressed by murine and human B cells [346]. The CpG ODN mediated activation of TLR9 on B cells increased the magnitude of the antibody response to the protein antigen but inhibited process of antibody affinity maturation both in mice and in human clinical trials [347]. Thus, the activation of TLR9 on B cells has a potential to affect antibody response during various vaccination strategies where various TLR9-agonists are used as adjuvants.
The trafficking of TLR7 to endosomes is independent of UNC-93B1 in human B cells as they do not express UNC-93B1 but not in murine B cells [346,348]. TRAF5 is a negative regulator of TLR signaling in B cells as TRAF5 −/− B cells produce higher levels of IL-6, IL-12p40, IL-10, TNF-α, and IgM via increased activation of MAPKs ERK1/2 and JNK, as compared to normal B cells [349]. The negative regulation by TRAF5 involves blocking the association of TAB2 with TRAF6 during TLR signaling in B cells [349]. While TRAF5 −/− macrophages and DCs remain unaffected in terms of TLR induced cytokine release [349]. Human B cells produce various cytokines and chemokines that are MIP-1α, MIP-1β, macrophage chemotactic protein-1 (MCP-1 or CCL2), IL-1α, IL-1β, IL-6, IL-8, IL-13 and IL-10, IP-10, G-CSF and GM-CSF upon stimulation of TLRs [[350], [351], [352]]. All these cytokines play an important role in the pathogenesis of sepsis and several other inflammatory diseases including rheumatoid arthritis (RA).
Even the germinal center (GC) B cells have higher sensitivity to TLR ligands due to higher expression of MyD88, MAL and IRAK-M, which are essential domains of TLR signaling [353]. For example, patients with inborn errors in the downstream TLR signaling effectors, MyD88 or IRAK4 are susceptible to Streptococcus pneumococcus (S. pneumococcus), Pseudomonas aeruginosa (P. aeruginosa) and S. aureus recurring invasive infections responsible for the induction of sepsis [[354], [355], [356]]. Thus, GC-B cells may play an important role in sepsis due to their increased sensitivity to pathogens as mentioned previously [333,353,357]. In addition to their active role in the pathogenesis of sepsis, Type 2 Diabetes (T2D) and periodontal disease (PD) are two inflammatory diseases that occur conjointly and involve disruption of TLR-dependent B cell function and release of pro-inflammatory cytokines [358]. B cells that play an important role in the negative regulation of inflammation and inflammatory diseases are called as regulatory B cells [359]. Among several regulatory B cell subsets, IL-10 producing regulatory B cells are most studied and have been found to be associated with several inflammatory diseases including autoimmunity (i.e. RA, SLE, T1DM, inflammatory bowel disease (IBD), multiple sclerosis (MS) and pemphigus vulgaris), where their levels are usually decreased [359,360]. Hence B cells are very crucial immune cells for different inflammatory conditions and TLR signaling is one of the several IRAK pathways involved in inflammation. Thus a future study in this direction will prove beneficial to identify novel mechanisms of various inflammatory disease and their therapeutic targets.
4.10. Microglia, astrocytes, and neurons also express TLRs with specific pattern
Microglia and astrocytes are innate immune cells of brain expressing TLRs that play important role in the recognition of pathogen entering into the brain, inflammation associated with neurodegeneration and the pathogenesis of stroke [[361], [362], [363], [364], [365]]. Microglia are found to express TLR1, TLR2, TLR3, TLR4, TLR5, TLR7, TLR8, TLR9 and co-receptor CD14 and their expression increases following exposure to bacterial pathogens [366,367]. Whereas human brain astrocytes also express TLR1, TLR2, TLR3, TLR5, TLR7, TLR8, and TLR9 but the expression of TLR4 on astrocytes is controversial some reports agree with its expression on astrocytes while others have not found the expression of TLR4 on astrocytes [[368], [369], [370], [371], [372], [373], [374], [375], [376]]. Both isoforms of MyD88 that is full length and short form, TIRAP/Mal, and TICAM-1/TRIF, are all present in astrocytes, which are the primary requirement for the initiation and completion of TLR signaling [369]. Thus, expression of TLRs in astrocytes may explain their role as innate immune cells of the brain in addition to microglia for the recognition of pathogens and complications of the central nervous system (CNS) observed during sepsis and other inflammatory diseases.
Human and mouse neurons also express TLR3 and mount a strong pro-inflammatory immune response that is the release of TNF-α, IL-6, CCL-5, CXCL-10 and IFN-β upon treatment with dsRNA (a ligand for TLR3) [377,378]. TLR4 and CD14 receptor expression have also been reported in human and rat neurons [379]. TLR7 has also been shown to be expressed by human neurons [380]. Mouse neurons also express TLR8 but here it is not involved in the canonical TLR-NF-κB signaling pathway and does not mount a classic TLR mediated immune response but functions as a negative regulator of neurite outgrowth and induces apoptosis of neurons [381]. Thus, expression of TLRs including TLR4 on neurons and microglia indicate that they may also be activated by different PAMPs and play an active role in sepsis/infection associated neuropathological and neurobehavioral conditions called sepsis-associated delirium (SAD) [[382], [383], [384], [385]]. In addition to stimulation of TLRs expressed in brain environment that is neurons, astrocytes and microglia under microbial infections these can also be stimulated under sterile inflammatory conditions (i.e. neurodegenerative disorders, stroke, multiple sclerosis (MS) and spinal cord injury etc.) causing inflammatory damage to neurons or brain [376]. Thus, further studies in context to TLRs and their role in neurons and brain innate immune cells may explore the unknown adventure of these PRRs in various inflammatory disorders of the brain.
5. TLR polymorphism in humans and their disease association
TLRs are found to be expressed on almost every immune cell type and are involved in the recognition of various PAMPs and host-derived DAMPs involved in the pathogenesis of various inflammatory diseases [[386], [387], [388], [389]]. However, polymorphic variants of these TLR genes in humans have been identified to be associated with several inflammatory disorders. This gene polymorphisms can occur due to the various process including duplication, deletion or a mutation causing triplication of the high quantity of DNA base pairs sequences or changes in intron regions. A total of 13 introns are found in the TIR domains of TLR genes [390], suggesting that TIR domain faced many challenges of invasion by introns during the evolution of the innate immune system that serves as a conclusive support of intron-latent theory [391,392]. Thus, the presence of introns in TIR domain of TLRs makes them to undergo intron-specific polymorphism in addition to other forms of gene polymorphism including single nucleotide polymorphism (SNP), small-scale insertions and deletions, polymorphic repetitive elements and microsatellite variation. The first case of TLR polymorphism was observed in mouse strain C3H/HeJ with LPS hyporesponsive phenotype due to the substitution of proline with histidine at amino acid 712 in TLR4 protein [46,393]. Thereafter lots of studies are performed in this direction and several SNPs in human TLR genes are identified. For example, 3 SNPs (rs77590742, rs574356 and rs5743604) from TLR1 allele (−7202G, −6399T, and −833T) are associated with higher risk of prostate cancer in Swedish population [394]. In addition to TLR1 polymorphism, a gene cluster of TLR6 and TLR10 are also associated with prostate cancer development along with IRAK-1 and 4 [395]. While T1805G SNP in the coding region of TLR1 occurring due to shifting of isoleucine into serine at 602 aa (I602S) in the extracellular region of TLR1 is associated with diminished activation of NF-κB and IL-6 synthesis upon stimulation with TLR1 ligands [396]. This I602S SNP may have an association with protection against Mycobacteria leprae induced leprosy and this 602S allele comprises the most common SNP affecting TLR function identified to date among white European population [397].
TLR2 gene has two exons and is located on chromosome 4q32 [398]. The TLR2 SNP Arg753Gln originated due to the substitution of arginine to glutamine at 753 amino acid residue is associated with higher risk of development of tuberculosis [399]. In children same SNP is associated with higher incidence of urinary tract infections caused by gram-negative bacteria, increased risk of febrile recurrent febrile infections, rheumatic fever and in adults increases the risk of development of familial Mediterranean fever [[400], [401], [402], [403]]. This SNP is also associated with non-infectious diseases like coronary restenosis, type 1 diabetes (T1D) and allograft failure and associated death [[404], [405], [406], [407]].
TLR3 gene has 6 exons and is located on chromosome 4q35.1 in humans. L412F polymorphism in the TLR3 gene may serve as a genetic risk factor for human cytomegalovirus (HCMV) infection in children [408]. In addition, variant TLR3 genotype rs5743312 (C9948T, intron 3, C > T) is also shown to be significantly associated with increased cancer risk [409]. Another polymorphism in TLR3 gene is associated with age-related macular degeneration (AMD) in Caucasian but not in Asian population [410]. Leu412Phe polymorphism in TLR3 genes is also associated with enteroviral myocarditis/cardiomyopathy in the Caucasian population [411]. TLR4 gene has four exons and is located on chromosome 9q33.1 in humans. TLR4 Thr399Ile polymorphism is shown to exert protection against visceral leishmaniasis (VL) in Mediterranean basin [412]. While g.13726T > C and g.15090G > A genetic polymorphisms of TLR4 gene make Chinese Han Population more susceptible to develop type 2 diabetes mellitus (T2DM) [413]. TLR4 gene polymorphism rs10759932 is found to be associated with lifetime cigarette smoking in bipolar disorder [414]. TLR4 codon 299 heterozygous gene mutation (Asp299Gly) is shown to be protective against nonalcoholic fatty liver disease (NAFLD) in humans [415].
TLR5 gene has seven exons and is located on chromosome 1q41. Two TLR5 gene variants mainly R392X and N592S are found to be associated with increased risk of ulcerative colitis (UC) in a population of Northern India [416]. TLR5 polymorphism rs5744174 is shown to exert protection against Grave's disease in females of Chinese Cantonese population [417]. TLR9 gene with its 2 exons is located on chromosome 3p21.2 in humans. TLR9 gene polymorphism rs352139 is associated with increased risk of Tuberculosis (TB) in Mexican population but it is associated with decreased risk of TB in Indonesian population [418]. A link between TLR9-1237 T/C gene polymorphism and molecular risk for diabetes foot (DF) disease in patients with T2DM is also observed [419].
It is impossible to include all TLR (TLR1-TLR10) SNPs in humans and their association with the disease susceptibility here in this manuscript, therefore, this information can be found in detail in references mentioned [398,[420], [421], [422]]. Table 1 is shown with different TLRs and their associated SNPs and disease association. Thus, genetic variation in TLR expression and associated SNPs have been found to be associated with certain inflammatory and infectious diseases in different population all over the world. This may lead to the development of TLR based gene therapy as well as the development of certain molecules to target these TLRs responsible for disease pathogenesis. These days various cell lines derived from patients with different diseases are used to carry out in vitro studies relevant to human diseases including cancers, neurodegenerative diseases etc. However, this experimental approach can also be affected by the phenomenon of TLR SNPs as these cell line may have specific SNP for TLRs depending on the population frequency and for that person's ancestral background [423]. It is more common that SNPs for genes associated with inflammation are not commonly screened in common human cells, for example, HeLa and HEK293 cell lines [423]. Thus, the presence of TLR SNPs (homozygous or heterozygous alleles) on these cell lines has a great potential to alter the TLR mediated inflammatory pathway and its associated impact on in vitro studies [423]. Thus, this line should be screened for any TLR SNPs before their use in experiments where TLR signaling plays an important role.
6. Current and future TLR based therapeutics
Due to their potential role in the recognition of PAMPs of various origin and generation of pro-inflammatory response during different inflammatory conditions including sepsis, TLRs are considered a novel therapeutic drug target. For example, a TLR4 antagonist known as Eritoran (E 5564) was developed for the treatment of sepsis initially by a Japanese pharmaceutical company Eisai Co but in 2013 phase III clinical trial showed it as effective as placebo in 28-day mortality study during sepsis [17,424]. However, it proved to be effective against lethal influenza infection-associated acute lung injury (ALI) in mice and can be used a potential TLR4 based therapeutic approach to target influenza-associated ALI [425]. In addition, several TLR4 agonists including monophosphoryl lipid A (MPLA) is 1000 times less toxic than bacterial LPS and can be used as an adjuvant with safety profile as similar to alum in humans [[426], [427], [428]]. The MPLA based adjuvant also called AS04 has successfully been used in hepatitis B vaccine (FENDrix®) in humans [429]. In addition, AS04 has also been used as an adjuvant with human papillomavirus (HPV) vaccine called Cervarix® [430]. AS04 is also used as an adjuvant with certain cancer vaccines, for example, Melacine® comprises of lysates obtained from two allogeneic melanoma cell lines combined with DETOX® (an adjuvant containing AS04 and a component of cell wall of Mycobacterium phlei), is now approved by Canadian FDA for the treatment of metastatic melanoma [431]. This TLR4 agonist has also been used as an immunostimulator along with a vaccine called Stimuvax®, used in certain adenocarcinoma including non-small cell lung cancer [432]. Thus, TLR4 based agonists and antagonists have opened a new gateway to design potential immunomodulators that can either be used alone or in combination as potent adjuvants for increasing the efficacy of vaccines.
Additionally, other TLRs have also been used in drug designing. For example, TLR2 agonist called SMP-105, a derivative of mycolic acid (MA), arabinogalactan (consisting of arabinose, galactose, and rhamnose) and PGN (comprising of alanine, glutamic acid, diaminopimelic acid, muramic acid, glucosamine, and galactosamine) are isolated from Mycobacterium bovis bacillus Calmette-Guerin (BCG) Tokyo 172 [433]. Mycobacterium bovis BCG is used to treat bladder cancer as a potential immunotherapeutic agent since 1976 [434,435] and SMP-105 has also shown its anti-tumor effect against bladder carcinoma cell lines and against other cancers including lung cancer via activating TLR2 [436,437]. TLR3 agonist IPH-3102 that mimics dsRNA has been shown to destroy breast cancer and melanoma cancer cells via activating NF-κB and type 1 IFNs in vitro [438]. Further research has also shown its efficacy in animal models of cancers as a potential therapeutic approach in the field of cancer immunotherapy [439].
The synthetic TLR3 agonist called poly I:C12U (Ampligen), when given as an adjuvant in an animal model of influenza virus H5N1 infection with H5N1 vaccine caused an increased production of antigen-specific IgA production causing a significant decrease in virus titer and protecting all animals [440]. Ampligen or poly I:C12U acts as a potent inducer of maturation of human monocyte-derived DC with sustained release and production of IL-12 cytokine in vitro [441]. These are capable of generating Th1 specific anti-cancer responses in peripheral blood T-cells derived from cancer patients in the presence of ascites medium containing immunosuppressive cytokines [441]. Rintatolimod, a TLR3 agonist that is sold under the name of Ampligen is approved for chronic fatigue syndrome (CFS), hepatitis B and hepatitis C infection, HIV, Influenza and severe acute respiratory syndrome (SARS) [441]. Thus, TLR agonists can be used as potent immunotherapeutic agents during certain infections including influenza and cancers.
TLR7, TLR8, and TLR9 based therapeutic approaches have also been established in recent years. For example, Imiquimod (Aldara, 3M) is designated as one of the first and most successful TLR based therapeutic approaches that targets TLR7/TLR8-MyD88 pathway of TLR signaling and causes induction of release of IL-6, TNF-α and IFN-α (exerting antiviral and antitumor effect) [438,442]. Imiquimod (Aldara, 3 M) is now approved for warts caused by HPV, basal cell carcinoma, and actinic keratosis [[443], [444], [445]]. Even ssRNA based therapeutics targeting of TLR7 and TLR8 has potent antitumor action and reverses the immunosuppressive action of regulatory T cells (Tregs) via activating DCs [446] and via inducing Th1 immune response [438]. For example, another TLR7 agonist called 852A (N-[4-(4-amino-2-ethyl-1 Himidazo[4,5c]quinolin-1-yl) butyl] methanesulfonamide, 3 M-001) that is a small-molecule imidazoquinoline similar to imiquimod with highly selective potential to activate TLR-7 is currently under investigation for the treatment of inoperable melanoma, renal cell cancer, breast cancer, ovarian tumor and cancer of cervix etc. [[447], [448], [449]]. 852A (N-[4-(4-amino-2-ethyl-1 Himidazo[4,5c]quinolin-1-yl) butyl] methanesulfonamide, 3M-001) has potent stimulatory action on DCs and stimulates them to release IFN-α along with other potent cytokines having an anti-tumor effect [[449], [450], [451]]. Similarly, Immunomodulatory oligonucleotides (IMOs), called IMO-2055 and IMO-2125, targeting TLR9 have also been developed and are used as single therapeutic agents or as in combination with other cancer chemotherapeutic drugs in certain cancers including non-small cell lung cancer (NSCLC) and colorectal cancer along with viral infections (i.e. Hepatitis C virus-infected patients) [[452], [453], [454]]. Thus, so far only three TLR agonists have been approved in different countries for different diseases [455]. These include: (1) Picibanil, a lyophilized formulation of the bacteria called Streptococcus pyogenes (S. pyogenes) is approved in Japan for various cancers [456,457], (2) monophosphoryl lipid A (MPL) derived from Salmonella minnesota (S. minnesota) LPS is used as an adjuvant in peptide-based vaccine called HPV-16/18 AS04-adjuvanted vaccine specifically in cervical intraepithelial neoplasia 2+ (CIN2+) caused by HPV-16 and HPV-18 [458,459], and (3) imiquimod, derived from imidazoquinoline targeting TLR7/TLR8-MyD88 pathway is approved for the topical treatment of actinic keratosis, superficial basal cell carcinoma, and external anogenital warts (Condylomata acuminata) [460,461]. In addition, several new clinical trials with various TLR agonists and antagonist have been initiated for various types of cancers [455]. Thus, due to their multi-action varying from vaccine targets, adjuvant use and their use as biologics to treat various inflammatory conditions, TLRs have been considered as “Swiss army knife” of the human immune system that have potential to act in various infections and sterile inflammatory conditions as well [462].
In addition to IMO-2055 and 2125, other TLR9 agonists called MGN9-1703 and MGN-1706 that are double stem-loop immunomodulating agents consist of non-coding DNA molecules are also being developed as anticancer agents and are under investigation in animal models and in prostate and colorectal cancer patients [438,463]. Thus, various TLR antagonists and agonists have been developed and are under investigation in several diseases (i.e. allergy, asthma, autoimmunity, and cancers) other than infectious diseases, where TLR signaling is the major contributing factor and was discovered in relevance. For example, TLR9 antagonist called hydroxychloroquine (HCQ), an antimalarial drug is now used as a therapeutic agent to treat autoimmune diseases (i.e. SLE and RA) [438]. Furthermore, quinazoline derivative called CPG-52364 is a specific inhibitor of TLR7, TLR8, and TLR9 that has shown efficacy in SLE and other cancers [464]. Most recently, an aminoguanidine compound called guanabenz (2,6-dichlorobenzylidene aminoguanidine acetate) that is used as an antihypertensive drug is shown to prevent the activation of TLR9 upon stimulation with CpG ODNs or DNA immunoglobulin complexes in endosomes [465]. The drug inhibited TLR9 activation in both murine and human DCs and B cells in vitro [465]. Under in vivo conditions, guanabenz showed protection against CpG ODN induced systemic cytokine shock and a chemically induced autoimmune disease called SLE in mice [465]. In addition to these direct agonists and antagonists of TLR signaling, future TLR based therapeutics will be comprised of miRNAs regulating TLR signaling and the host-derived negative regulators of TLR signaling (i.e. certain kinases, phosphatases, cytokines etc.) which have potential to directly degrade the TLR-signaling molecules involved in MyD88-dependent or TRIF-dependent and MyD88-independent TLR signaling. However, it is impossible to describe all the endogenous negative regulators of TLR signaling here in this manuscript whose detail can be found somewhere else [[466], [467], [468], [469], [470]].
7. Future perspective
TLRs are the relatively new class of innate immune receptors in context to mammalian/human immunology as were first discovered to be expressed on human macrophages in 1997. Further studies in the field of immunology and TLR expression established that the TLR expression is not limited to innate immune cells but are also expressed by cells of adaptive immunity that is T cells (i.e. CD4+T cells, CD8+T cells, regulatory CD4+ CD25+ T cells etc.) and B cells [37,323,340,357,471]. In addition to directly impacting the biology of innate immune response, TLRs have also been shown to impact the adaptive immune response mediated by different types of T cells and B cells that is their development, activation, survival, death and associated immune response [327,331,339,472]. Even TLR2−/− mice, unlike TLR4−/− mice, contain significantly fewer CD4+CD25+ regulatory T cells (Tregs) as compared to corresponding wild-type (WT) mice [473]. Thus, TLRs serve as an important PRRs for the cells of both innate and adaptive immune system. However, TLRs have also been found to be expressed by newly identified cells called innate lymphoid cells (ILCs) that appear morphologically as lymphoid cells but represent a heterogeneous group of hematopoietic innate immune cells [3,474,475]. For example, these ILCs express TLR2, TLR3, and TLR9 but TLR4 and TLR7 are not expressed by these cells [474]. In addition, c-Kit+ ILCs (type 2 and 3 ILCs) are a specific population of ILCs that are found to expand in humans during filarial worm or helminthic infection [476]. These c-Kit+ ILCs (type 2 and 3 ILCs) also express TLR5, TLR6, TLR7 and TLR9 [476]. This purified c-Kit+ ILCs produce a higher amount of GM-CSF in response to TLR6 ligand [476]. However, TLR-signaling in the c-Kit+ ILCs did not induce more prototypic cytokines including IL-5, IL-17A, IFN-γ [476]. Human conventional type 3 ILCs express mRNA transcripts of TLR1, TLR2, TLR5, TLR6, TLR7, and TLR9 [477]. But only TLR2 agonists are capable to induce cytokine production from human type 3 ILCs in the presence of IL-2, IL-15, and IL-23 [477]. This specific expression of TLR2 on type 3 ILCs or lymphoid tissue inducer (LTi) cells is involved in the depletion of these cells via induction of their apoptosis in simian immunodeficiency virus (SIV)-infected macaques [478]. This apoptosis of type 3 ILCs or lymphoid tissue inducer (LTi) cells causes a breach in intestinal barrier leading to microbial translocation and development of bacteremia, systemic immune activation and disease progression in patients infected with HIV-1 infection [3,478]. Thus, expression of certain TLRs by ILCs has opened a new direction in the field of TLRs and ILCs research.
TLRs also play important role in the sterile inflammatory conditions ranging from cancer to autoimmunity. For example, they have been implicated in the pathogenesis of several auto-immune diseases including SLE, RA, seronegative spondylarthritides (SpA), multiple sclerosis (MS), experimental autoimmune encephalitis (EAE), type 2 diabetes mellitus (T2DM), systemic sclerosis, Sjogren's syndrome (SS), myositis (a chronic inflammatory autoimmune disease primarily affecting skeletal muscles, causing severe muscle weakness and fatigue [[479], [480], [481], [482], [483], [484]]. They have also been associated with the pathogenesis of several human cancers including B cell malignancies, colorectal cancer, basal cell carcinoma, bladder cancer, including several other cancers [[485], [486], [487], [488], [489]]. This involvement of TLRs in the pathogenesis of autoimmune diseases and cancers has led to the development novel immunotherapies for these diseases [[490], [491], [492], [493], [494]]. Even TLR mediated immune stimulation approach is now being used to enhance radiation therapy to control tumor growth [495]. For example, various TLRs have now also been implicated in the pathogenesis of neurodegenerative disorders including Parkinson's disease (PD), Alzheimer's disease (AD), Amyotrophic lateral sclerosis (ALS) and repair mechanism is brain disorders [[496], [497], [498], [499], [500]]. Thus, TLRs are playing an important role in infectious and non-infectious disease and are targeted as therapeutic approaches in these diseases [501,502]. Even studies have shown that these TLRs are controlling mammalian reproduction via regulating the process of steroidogenesis, spermatogenesis in males and ovulation, fertilization, gestation, and parturition in females [[503], [504], [505], [506]]. TLRs have also been implicated in the process of regeneration including liver regeneration in mammals [[507], [508], [509]]. They are also involved in allograft inflammation and promote acute and chronic organ transplant rejection [[510], [511], [512], [513]].
In summary, the story of TLRs started from their first discovery in D. melanogaster as genes controlling body patterning during embryonic development. From this stage of embryonic development, they established their role in the immune system as important PRRs responsible for the recognition of pathogens. This discovery by the research group led by late Charles A. Janeway Jr. led to the development of the concept of PRRs and PAMPs and revolutionized the field of innate immunity. Now after 20 years of their discovery in humans, TLRs are ruling the world of human biology from their role in infections, inflammation, reproduction, development, autoimmunity, cancer, allograft inflammation/rejection and regeneration. Still, the continuous research in the field of TLRs has great potential to explore the unexplored role of TLR in the very complex system of mammalian/human biology. TLRs are awesome and need greater attention in terms of their evolution, understanding the mammalian biology under both homeostatic and inflammatory disease conditions. Thus, along with controlling various biological functions, different TLR agonists and antagonists have been designed to target various inflammatory conditions including cancers and autoimmunity.
References
- 1.Kimbrell D.A., Beutler B. The evolution and genetics of innate immunity. Nat. Rev. Genet. 2001;2(4):256–267. doi: 10.1038/35066006. [DOI] [PubMed] [Google Scholar]
- 2.Buchmann K. Evolution of Innate Immunity: Clues from Invertebrates via Fish to Mammals. Front. Immunol. 2014;5:459. doi: 10.3389/fimmu.2014.00459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kumar V. Innate lymphoid cells: new paradigm in immunology of inflammation. Immunol. Lett. 2014;157(1–2):23–37. doi: 10.1016/j.imlet.2013.11.003. [DOI] [PubMed] [Google Scholar]
- 4.Kumar V., Sharma A. Neutrophils: Cinderella of innate immune system. Int. Immunopharmacol. 2010;10(11):1325–1334. doi: 10.1016/j.intimp.2010.08.012. [DOI] [PubMed] [Google Scholar]
- 5.Silva M.T. When two is better than one: macrophages and neutrophils work in concert in innate immunity as complementary and cooperative partners of a myeloid phagocyte system. J. Leukoc. Biol. 2010;87(1):93–106. doi: 10.1189/jlb.0809549. [DOI] [PubMed] [Google Scholar]
- 6.Galli S.J., Borregaard N., Wynn T.A. Phenotypic and functional plasticity of cells of innate immunity: macrophages, mast cells and neutrophils. Nat. Immunol. 2011;12(11):1035–1044. doi: 10.1038/ni.2109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kolaczkowska E., Kubes P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013;13(3):159–175. doi: 10.1038/nri3399. [DOI] [PubMed] [Google Scholar]
- 8.de Oliveira S., Rosowski E.E., Huttenlocher A. Neutrophil migration in infection and wound repair: going forward in reverse. Nat. Rev. Immunol. 2016;16(6):378–391. doi: 10.1038/nri.2016.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kumar V., Ahmad A. Role of MAIT cells in the immunopathogenesis of inflammatory diseases: New players in old game. Int. Rev. Immunol. 2017:1–21. doi: 10.1080/08830185.2017.1380199. [DOI] [PubMed] [Google Scholar]
- 10.Morgan B.P., Harris C.L. Complement, a target for therapy in inflammatory and degenerative diseases. Nat. Rev. Drug Discov. 2015;14(12):857–877. doi: 10.1038/nrd4657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pasupuleti M., Schmidtchen A., Malmsten M. Antimicrobial peptides: key components of the innate immune system. Crit. Rev. Biotechnol. 2012;32(2):143–171. doi: 10.3109/07388551.2011.594423. [DOI] [PubMed] [Google Scholar]
- 12.Wiesner J., Vilcinskas A. Antimicrobial peptides: the ancient arm of the human immune system. Virulence. 2010;1(5):440–464. doi: 10.4161/viru.1.5.12983. [DOI] [PubMed] [Google Scholar]
- 13.Diamond G., Beckloff N., Weinberg A., Kisich K.O. The Roles of Antimicrobial Peptides in Innate Host Defense. Curr. Pharm. Des. 2009;15(21):2377–2392. doi: 10.2174/138161209788682325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Radek K., Gallo R. Antimicrobial peptides: natural effectors of the innate immune system. Semin. Immunopathol. 2007;29(1):27–43. doi: 10.1007/s00281-007-0064-5. [DOI] [PubMed] [Google Scholar]
- 15.Wagner E., Frank M.M. Therapeutic potential of complement modulation. Nat. Rev. Drug Discov. 2010;9(1):43–56. doi: 10.1038/nrd3011. [DOI] [PubMed] [Google Scholar]
- 16.Dunkelberger J.R., Song W.-C. Complement and its role in innate and adaptive immune responses. Cell Res. 2009;20(1):34–50. doi: 10.1038/cr.2009.139. [DOI] [PubMed] [Google Scholar]
- 17.Kumar V., Sharma A. Innate immunity in sepsis pathogenesis and its modulation: new immunomodulatory targets revealed. J. Chemother. 2008;20(6):672–683. doi: 10.1179/joc.2008.20.6.672. [DOI] [PubMed] [Google Scholar]
- 18.Romani L. Immunity to fungal infections. Nat. Rev. Immunol. 2011;11(4):275–288. doi: 10.1038/nri2939. [DOI] [PubMed] [Google Scholar]
- 19.Areschoug T., Gordon S. Pattern recognition receptors and their role in innate immunity: focus on microbial protein ligands. Contrib. Microbiol. 2008;15:45–60. doi: 10.1159/000135685. [DOI] [PubMed] [Google Scholar]
- 20.Kumagai Y., Akira S. Identification and functions of pattern-recognition receptors. J. Allergy Clin. Immunol. 2010;125(5):985–992. doi: 10.1016/j.jaci.2010.01.058. [DOI] [PubMed] [Google Scholar]
- 21.Takeuchi O., Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–820. doi: 10.1016/j.cell.2010.01.022. [DOI] [PubMed] [Google Scholar]
- 22.Azumi K., De Santis R., De Tomaso A., Rigoutsos I., Yoshizaki F., Pinto M.R., Marino R., Shida K., Ikeda M., Ikeda M., Arai M., Inoue Y., Shimizu T., Satoh N., Rokhsar D.S., Du Pasquier L., Kasahara M., Satake M., Nonaka M. Genomic analysis of immunity in a Urochordate and the emergence of the vertebrate immune system: "waiting for Godot". Immunogenetics. 2003;55(8):570–581. doi: 10.1007/s00251-003-0606-5. [DOI] [PubMed] [Google Scholar]
- 23.Tenor J.L., Aballay A. A conserved Toll-like receptor is required for Caenorhabditis elegans innate immunity. EMBO Rep. 2008;9(1):103–109. doi: 10.1038/sj.embor.7401104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Irazoqui J.E., Urbach J.M., Ausubel F.M. Evolution of host innate defence: insights from C. elegans and primitive invertebrates. Nat. Rev. Immunol. 2010;10(1):47–58. doi: 10.1038/nri2689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Brandt J.P., Ringstad N. Toll-like Receptor Signaling Promotes Development and Function of Sensory Neurons Required for a C. elegans Pathogen-Avoidance Behavior. Curr. Biol. 2015;25(17):2228–2237. doi: 10.1016/j.cub.2015.07.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hashimoto C., Hudson K.L., Anderson K.V. The Toll gene of Drosophila, required for dorsal-ventral embryonic polarity, appears to encode a transmembrane protein. Cell. 1988;52(2):269–279. doi: 10.1016/0092-8674(88)90516-8. [DOI] [PubMed] [Google Scholar]
- 27.Medzhitov R., Preston-Hurlburt P., Janeway C.A., Jr. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature. 1997;388(6640):394–397. doi: 10.1038/41131. [DOI] [PubMed] [Google Scholar]
- 28.Vaure C., Liu Y. A comparative review of toll-like receptor 4 expression and functionality in different animal species. Front. Immunol. 2014;5:316. doi: 10.3389/fimmu.2014.00316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Leulier F., Lemaitre B. Toll-like receptors—taking an evolutionary approach. Nat. Rev. Genet. 2008;9(3):165–178. doi: 10.1038/nrg2303. [DOI] [PubMed] [Google Scholar]
- 30.Roach J.C., Glusman G., Rowen L., Kaur A., Purcell M.K., Smith K.D., Hood L.E., Aderem A. The evolution of vertebrate Toll-like receptors. Proc. Natl. Acad. Sci. U. S. A. 2005;102(27):9577–9582. doi: 10.1073/pnas.0502272102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rast J.P., Smith L.C., Loza-Coll M., Hibino T., Litman G.W. Genomic insights into the immune system of the sea urchin. Science. 2006;314(5801):952–956. doi: 10.1126/science.1134301. (New York, N.Y.) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Akira S., Uematsu S., Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124(4):783–801. doi: 10.1016/j.cell.2006.02.015. [DOI] [PubMed] [Google Scholar]
- 33.Kawai T., Akira S. Antiviral signaling through pattern recognition receptors. J. Biochem. 2007;141(2):137–145. doi: 10.1093/jb/mvm032. [DOI] [PubMed] [Google Scholar]
- 34.Coban C., Ishii K.J., Uematsu S., Arisue N., Sato S., Yamamoto M., Kawai T., Takeuchi O., Hisaeda H., Horii T., Akira S. Pathological role of Toll-like receptor signaling in cerebral malaria. Int. Immunol. 2007;19(1):67–79. doi: 10.1093/intimm/dxl123. [DOI] [PubMed] [Google Scholar]
- 35.Akira S. TLR signaling. Curr. Top. Microbiol. Immunol. 2006;311:1–16. doi: 10.1007/3-540-32636-7_1. [DOI] [PubMed] [Google Scholar]
- 36.Uematsu S., Akira S. Toll-like receptors and innate immunity. J. Mol. Med. 2006;84(9):712–725. doi: 10.1007/s00109-006-0084-y. (Berlin, Germany) [DOI] [PubMed] [Google Scholar]
- 37.Liu G., Zhao Y. Toll-like receptors and immune regulation: their direct and indirect modulation on regulatory CD4(+) CD25(+) T cells. Immunology. 2007;122(2):149–156. doi: 10.1111/j.1365-2567.2007.02651.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Manicassamy S., Pulendran B. Modulation of adaptive immunity with Toll-like receptors. Semin. Immunol. 2009;21(4):185–193. doi: 10.1016/j.smim.2009.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Majewska M., Szczepanik M. The role of Toll-like receptors (TLR) in innate and adaptive immune responses and their function in immune response regulation. Postepy Hig. Med. Dosw. 2006;60:52–63. (Online) [PubMed] [Google Scholar]
- 40.Gerttula S., Jin Y.S., Anderson K.V. Zygotic expression and activity of the Drosophila Toll gene, a gene required maternally for embryonic dorsal-ventral pattern formation. Genetics. 1988;119(1):123–133. doi: 10.1093/genetics/119.1.123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Gay N.J., Keith F.J. Drosophila Toll and IL-1 receptor. Nature. 1991;351(6325):355–356. doi: 10.1038/351355b0. [DOI] [PubMed] [Google Scholar]
- 42.Lemaitre B., Nicolas E., Michaut L., Reichhart J.M., Hoffmann J.A. The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell. 1996;86(6):973–983. doi: 10.1016/s0092-8674(00)80172-5. [DOI] [PubMed] [Google Scholar]
- 43.Medzhitov R., Janeway C.A., Jr. Innate immunity: the virtues of a nonclonal system of recognition. Cell. 1997;91(3):295–298. doi: 10.1016/s0092-8674(00)80412-2. [DOI] [PubMed] [Google Scholar]
- 44.Rock F.L., Hardiman G., Timans J.C., Kastelein R.A., Bazan J.F. A family of human receptors structurally related to Drosophila Toll. Proc. Natl. Acad. Sci. U. S. A. 1998;95(2):588–593. doi: 10.1073/pnas.95.2.588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Poltorak A., Smirnova I., He X., Liu M.Y., Van Huffel C., McNally O., Birdwell D., Alejos E., Silva M., Du X., Thompson P., Chan E.K., Ledesma J., Roe B., Clifton S., Vogel S.N., Beutler B. Genetic and physical mapping of the Lps locus: identification of the toll-4 receptor as a candidate gene in the critical region. Blood Cells Mol. Dis. 1998;24(3):340–355. doi: 10.1006/bcmd.1998.0201. [DOI] [PubMed] [Google Scholar]
- 46.Poltorak A., He X., Smirnova I., Liu M.Y., Van Huffel C., Du X., Birdwell D., Alejos E., Silva M., Galanos C., Freudenberg M., Ricciardi-Castagnoli P., Layton B., Beutler B. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science (New York, N.Y.) 1998;282(5396):2085–2088. doi: 10.1126/science.282.5396.2085. [DOI] [PubMed] [Google Scholar]
- 47.Beutler B., Poltorak A. The search for Lps: 1993–1998. J. Endotoxin Res. 2000;6(4):269–293. [PubMed] [Google Scholar]
- 48.Beutler B. Tlr4: central component of the sole mammalian LPS sensor. Curr. Opin. Immunol. 2000;12(1):20–26. doi: 10.1016/s0952-7915(99)00046-1. [DOI] [PubMed] [Google Scholar]
- 49.Janeway C.A., Jr. The immune system evolved to discriminate infectious nonself from noninfectious self. Immunol. Today. 1992;13(1):11–16. doi: 10.1016/0167-5699(92)90198-G. [DOI] [PubMed] [Google Scholar]
- 50.Medzhitov R., Janeway C.A., Jr. Innate immunity: impact on the adaptive immune response. Curr. Opin. Immunol. 1997;9(1):4–9. doi: 10.1016/s0952-7915(97)80152-5. [DOI] [PubMed] [Google Scholar]
- 51.Rich T., Allen R., Trowsdale J. How low can toll go? Trends Genet. 2000;16(7):292–294. doi: 10.1016/s0168-9525(00)02026-6. [DOI] [PubMed] [Google Scholar]
- 52.Fallon P.G., Allen R.L., Rich T. Primitive Toll signalling: bugs, flies, worms and man. Trends Immunol. 2001;22(2):63–66. doi: 10.1016/s1471-4906(00)01800-7. [DOI] [PubMed] [Google Scholar]
- 53.Wasserman S.A. Toll signaling: the enigma variations. Curr. Opin. Genet. Dev. 2000;10(5):497–502. doi: 10.1016/s0959-437x(00)00118-0. [DOI] [PubMed] [Google Scholar]
- 54.Kawai T., Akira S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity. 2011;34(5):637–650. doi: 10.1016/j.immuni.2011.05.006. [DOI] [PubMed] [Google Scholar]
- 55.Imler J.L., Hoffmann J.A. Toll receptors in innate immunity. Trends Cell Biol. 2001;11(7):304–311. doi: 10.1016/s0962-8924(01)02004-9. [DOI] [PubMed] [Google Scholar]
- 56.Du X., Poltorak A., Wei Y., Beutler B. Three novel mammalian toll-like receptors: gene structure, expression, and evolution. Eur. Cytokine Netw. 2000;11(3):362–371. [PubMed] [Google Scholar]
- 57.Kawasaki T., Kawai T. Toll-like receptor signaling pathways. Front. Immunol. 2014;5:461. doi: 10.3389/fimmu.2014.00461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Fukui R., Saitoh S., Matsumoto F., Kozuka-Hata H., Oyama M., Tabeta K., Beutler B., Miyake K. Unc93B1 biases Toll-like receptor responses to nucleic acid in dendritic cells toward DNA- but against RNA-sensing. J. Exp. Med. 2009;206(6):1339–1350. doi: 10.1084/jem.20082316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Takahashi K., Shibata T., Akashi-Takamura S., Kiyokawa T., Wakabayashi Y., Tanimura N., Kobayashi T., Matsumoto F., Fukui R., Kouro T., Nagai Y., Takatsu K., Saitoh S., Miyake K. A protein associated with Toll-like receptor (TLR) 4 (PRAT4A) is required for TLR-dependent immune responses. J. Exp. Med. 2007;204(12):2963–2976. doi: 10.1084/jem.20071132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Yang Y., Liu B., Dai J., Srivastava P.K., Zammit D.J., Lefrancois L., Li Z. Heat shock protein gp96 is a master chaperone for toll-like receptors and is important in the innate function of macrophages. Immunity. 2007;26(2):215–226. doi: 10.1016/j.immuni.2006.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Park B., Brinkmann M.M., Spooner E., Lee C.C., Kim Y.M., Ploegh H.L. Proteolytic cleavage in an endolysosomal compartment is required for activation of Toll-like receptor 9. Nat. Immunol. 2008;9(12):1407–1414. doi: 10.1038/ni.1669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ewald S.E., Engel A., Lee J., Wang M., Bogyo M., Barton G.M. Nucleic acid recognition by Toll-like receptors is coupled to stepwise processing by cathepsins and asparagine endopeptidase. J. Exp. Med. 2011;208(4):643–651. doi: 10.1084/jem.20100682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Garcia-Cattaneo A., Gobert F.X., Muller M., Toscano F., Flores M., Lescure A., Del Nery E., Benaroch P. Cleavage of Toll-like receptor 3 by cathepsins B and H is essential for signaling. Proc. Natl. Acad. Sci. U. S. A. 2012;109(23):9053–9058. doi: 10.1073/pnas.1115091109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kawai T., Akira S. The roles of TLRs, RLRs and NLRs in pathogen recognitionInt Immunol. 21:313. Int. Immunol. 2009;21(4):317–337. doi: 10.1093/intimm/dxp017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kumar H., Kawai T., Akira S. Toll-like receptors and innate immunity. Biochem. Biophys. Res. Commun. 2009;388(4):621–625. doi: 10.1016/j.bbrc.2009.08.062. [DOI] [PubMed] [Google Scholar]
- 66.McClure R., Massari P. TLR-Dependent Human Mucosal Epithelial Cell Responses to Microbial Pathogens. Front. Immunol. 2014;5:386. doi: 10.3389/fimmu.2014.00386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Fitzner N., Clauberg S., Essmann F., Liebmann J., Kolb-Bachofen V. Human Skin Endothelial Cells Can Express All 10 TLR Genes and Respond to Respective Ligands. Clin. Vaccine Immunol. 2008;15(1):138–146. doi: 10.1128/CVI.00257-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Hemmi H., Akira S. TLR signalling and the function of dendritic cells. Chem. Immunol. Allergy. 2005;86:120–135. doi: 10.1159/000086657. [DOI] [PubMed] [Google Scholar]
- 69.Khakpour S., Wilhelmsen K., Hellman J. Vascular endothelial cell Toll-like receptor pathways in sepsis. Innate Immun. 2015;21(8):827–846. doi: 10.1177/1753425915606525. [DOI] [PubMed] [Google Scholar]
- 70.Nagyoszi P., Wilhelm I., Farkas A.E., Fazakas C., Dung N.T., Hasko J., Krizbai I.A. Expression and regulation of toll-like receptors in cerebral endothelial cells. Neurochem. Int. 2010;57(5):556–564. doi: 10.1016/j.neuint.2010.07.002. [DOI] [PubMed] [Google Scholar]
- 71.Sandig H., Bulfone-Paus S. TLR signaling in mast cells: common and unique features. Front. Immunol. 2012;3:185. doi: 10.3389/fimmu.2012.00185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Young S.L., Lyddon T.D., Jorgenson R.L., Misfeldt M.L. Expression of Toll-like Receptors in Human Endometrial Epithelial Cells and Cell Lines. Am. J. Reprod. Immunol. 2004;52(1):67–73. doi: 10.1111/j.1600-0897.2004.00189.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Trudler D., Farfara D., Frenkel D. Toll-Like Receptors Expression and Signaling in Glia Cells in Neuro-Amyloidogenic Diseases: Towards Future Therapeutic Application. Mediat. Inflamm. 2010;2010:12. doi: 10.1155/2010/497987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Jack C.S., Arbour N., Manusow J., Montgrain V., Blain M., McCrea E., Shapiro A., Antel J.P. TLR signaling tailors innate immune responses in human microglia and astrocytes. J. Immunol. 2005;175(7):4320–4330. doi: 10.4049/jimmunol.175.7.4320. (Baltimore, Md. : 1950) [DOI] [PubMed] [Google Scholar]
- 75.Hornef M.W., Normark B.H., Vandewalle A., Normark S. Intracellular recognition of lipopolysaccharide by toll-like receptor 4 in intestinal epithelial cells. J. Exp. Med. 2003;198(8):1225–1235. doi: 10.1084/jem.20022194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Uronen-Hansson H., Allen J., Osman M., Squires G., Klein N., Callard R.E. Toll-like receptor 2 (TLR2) and TLR4 are present inside human dendritic cells, associated with microtubules and the Golgi apparatus but are not detectable on the cell surface: integrity of microtubules is required for interleukin-12 production in response to internalized bacteria. Immunology. 2004;111(2):173–178. doi: 10.1111/j.0019-2805.2003.01803.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Shuang C., Wong M.H., Schulte D.J., Arditi M., Michelsen K.S. Differential expression of Toll-like receptor 2 (TLR2) and responses to TLR2 ligands between human and murine vascular endothelial cells. J. Endotoxin Res. 2007;13(5):281–296. doi: 10.1177/0968051907085096. [DOI] [PubMed] [Google Scholar]
- 78.Blasius A.L., Beutler B. Intracellular toll-like receptors. Immunity. 2010;32(3):305–315. doi: 10.1016/j.immuni.2010.03.012. [DOI] [PubMed] [Google Scholar]
- 79.Kono D.H., Haraldsson M.K., Lawson B.R., Pollard K.M., Koh Y.T., Du X., Arnold C.N., Baccala R., Silverman G.J., Beutler B.A., Theofilopoulos A.N. Endosomal TLR signaling is required for anti-nucleic acid and rheumatoid factor autoantibodies in lupus. Proc. Natl. Acad. Sci. U. S. A. 2009;106(29):12061–12066. doi: 10.1073/pnas.0905441106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Akira S., Takeda K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004;4(7):499–511. doi: 10.1038/nri1391. [DOI] [PubMed] [Google Scholar]
- 81.Pandey S., Kawai T., Akira S. Microbial sensing by Toll-like receptors and intracellular nucleic acid sensors. Cold Spring Harb. Perspect. Biol. 2014;7(1):a016246. doi: 10.1101/cshperspect.a016246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Tartey S., Takeuchi O. Pathogen recognition and Toll-like receptor targeted therapeutics in innate immune cells. Int. Rev. Immunol. 2017;36(2):57–73. doi: 10.1080/08830185.2016.1261318. [DOI] [PubMed] [Google Scholar]
- 83.Vidya M.K., Kumar V.G., Sejian V., Bagath M., Krishnan G., Bhatta R. Toll-like receptors: Significance, ligands, signaling pathways, and functions in mammals. Int. Rev. Immunol. 2017:1–17. doi: 10.1080/08830185.2017.1380200. [DOI] [PubMed] [Google Scholar]
- 84.Goulopoulou S., McCarthy C.G., Webb R.C. Toll-like Receptors in the Vascular System: Sensing the Dangers Within. Pharmacol. Rev. 2016;68(1):142–167. doi: 10.1124/pr.114.010090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kawai T., Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat. Immunol. 2010;11(5):373–384. doi: 10.1038/ni.1863. [DOI] [PubMed] [Google Scholar]
- 86.O'Connor G.M., Hart O.M., Gardiner C.M. Putting the natural killer cell in its place. Immunology. 2006;117(1):1–10. doi: 10.1111/j.1365-2567.2005.02256.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Sivori S., Carlomagno S., Pesce S., Moretta A., Vitale M., Marcenaro E. TLR/NCR/KIR: Which One to Use and When? Front. Immunol. 2014;5(105) doi: 10.3389/fimmu.2014.00105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Szajnik M., Szczepanski M.J., Czystowska M., Elishaev E., Mandapathil M., Nowak-Markwitz E., Spaczynski M., Whiteside T.L. TLR4 signaling induced by lipopolysacharide or paclitaxel regulates tumor survival and chemoresistance in ovarian cancer. Oncogene. 2009;28(49):4353–4363. doi: 10.1038/onc.2009.289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Pisetsky D.S., Erlandsson-Harris H., Andersson U. High-mobility group box protein 1 (HMGB1): an alarm in mediating the pathogenesis of rheumatic disease. Arthritis Res. Ther. 2008;10(3):209. doi: 10.1186/ar2440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Wang H., Ward M.F., Sama A.E. Targeting HMGB1 in the treatment of sepsis. Expert Opin. Ther. Targets. 2014;18(3):257–268. doi: 10.1517/14728222.2014.863876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Huang W., Tang Y., Li L. HMGB1, a potent proinflammatory cytokine in sepsis. Cytokine. 2010;51(2):119–126. doi: 10.1016/j.cyto.2010.02.021. [DOI] [PubMed] [Google Scholar]
- 92.Rittirsch D., Flierl M.A., Ward P.A. Harmful molecular mechanisms in sepsis. Nat. Rev. Immunol. 2008;8(10):776–787. doi: 10.1038/nri2402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Dumitriu I.E., Baruah P., Manfredi A.A., Bianchi M.E., Rovere-Querini P. HMGB1: guiding immunity from within. Trends Immunol. 2005;26(7):381–387. doi: 10.1016/j.it.2005.04.009. [DOI] [PubMed] [Google Scholar]
- 94.Wilhelmsen K., Mesa K.R., Prakash A., Xu F., Hellman J. Activation of endothelial TLR2 by bacterial lipoprotein upregulates proteins specific for the neutrophil response. Innate Immun. 2012;18(4):602–616. doi: 10.1177/1753425911429336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Imaizumi T., Itaya H., Fujita K., Kudoh D., Kudoh S., Mori K., Fujimoto K., Matsumiya T., Yoshida H., Satoh K. Expression of tumor necrosis factor-alpha in cultured human endothelial cells stimulated with lipopolysaccharide or interleukin-1alpha. Arterioscler. Thromb. Vasc. Biol. 2000;20(2):410–415. doi: 10.1161/01.atv.20.2.410. [DOI] [PubMed] [Google Scholar]
- 96.Ranta V., Orpana A., Carpen O., Turpeinen U., Ylikorkala O., Viinikka L. Human vascular endothelial cells produce tumor necrosis factor-alpha in response to proinflammatory cytokine stimulation. Crit. Care Med. 1999;27(10):2184–2187. doi: 10.1097/00003246-199910000-00019. [DOI] [PubMed] [Google Scholar]
- 97.Ojeda M.O., van't Veer C., Fernandez Ortega C.B., Arana Rosainz Mde J., Buurman W.A. Dialyzable leukocyte extract differentially regulates the production of TNFalpha, IL-6, and IL-8 in bacterial component-activated leukocytes and endothelial cells. Inflamm. Res. 2005;54(2):74–81. doi: 10.1007/s00011-004-1326-5. (et al.) [DOI] [PubMed] [Google Scholar]
- 98.Wilhelmsen K., Mesa K.R., Lucero J., Xu F., Hellman J. ERK5 protein promotes, whereas MEK1 protein differentially regulates, the Toll-like receptor 2 protein-dependent activation of human endothelial cells and monocytes. J. Biol. Chem. 2012;287(32):26478–26494. doi: 10.1074/jbc.M112.359489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Pegu A., Qin S., Fallert Junecko B.A., Nisato R.E., Pepper M.S., Reinhart T.A. Human lymphatic endothelial cells express multiple functional TLRs. J. Immunol. 2008;180(5):3399–3405. doi: 10.4049/jimmunol.180.5.3399. (Baltimore, Md. : 1950) [DOI] [PubMed] [Google Scholar]
- 100.Talreja J., Kabir M.H., B.F. M, Stechschulte D.J., Dileepan K.N. Histamine induces Toll-like receptor 2 and 4 expression in endothelial cells and enhances sensitivity to Gram-positive and Gram-negative bacterial cell wall components. Immunology. 2004;113(2):224–233. doi: 10.1111/j.1365-2567.2004.01946.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Garrafa E., Imberti L., Tiberio G., Prandini A., Giulini S.M., Caimi L. Heterogeneous expression of toll-like receptors in lymphatic endothelial cells derived from different tissues. Immunol. Cell Biol. 2011;89(3):475–481. doi: 10.1038/icb.2010.111. [DOI] [PubMed] [Google Scholar]
- 102.Shin H.S., Xu F., Bagchi A., Herrup E., Prakash A., Valentine C., Kulkarni H., Wilhelmsen K., Warren S., Hellman J. Bacterial lipoprotein TLR2 agonists broadly modulate endothelial function and coagulation pathways in vitro and in vivo. J. Immunol. 2011;186(2):1119–1130. doi: 10.4049/jimmunol.1001647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Zeuke S., Ulmer A.J., Kusumoto S., Katus H.A., Heine H. TLR4-mediated inflammatory activation of human coronary artery endothelial cells by LPS. Cardiovasc. Res. 2002;56(1):126–134. doi: 10.1016/s0008-6363(02)00512-6. [DOI] [PubMed] [Google Scholar]
- 104.Verma S., Nakaoke R., Dohgu S., Banks W.A. Release of cytokines by brain endothelial cells: A polarized response to lipopolysaccharide. Brain Behav. Immun. 2006;20(5):449–455. doi: 10.1016/j.bbi.2005.10.005. [DOI] [PubMed] [Google Scholar]
- 105.Lundberg A.M., Drexler S.K., Monaco C., Williams L.M., Sacre S.M., Feldmann M., Foxwell B.M. Key differences in TLR3/poly I:C signaling and cytokine induction by human primary cells: a phenomenon absent from murine cell systems. Blood. 2007;110(9):3245–3252. doi: 10.1182/blood-2007-02-072934. [DOI] [PubMed] [Google Scholar]
- 106.Huang H., Ostroff G.R., Lee C.K., Wang J.P., Specht C.A., Levitz S.M. Distinct patterns of dendritic cell cytokine release stimulated by fungal beta-glucans and toll-like receptor agonists. Infect. Immun. 2009;77(5):1774–1781. doi: 10.1128/IAI.00086-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Long E.M., Millen B., Kubes P., Robbins S.M. Lipoteichoic acid induces unique inflammatory responses when compared to other toll-like receptor 2 ligands. PLoS One. 2009;4(5) doi: 10.1371/journal.pone.0005601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Hartmann G., Krieg A.M. CpG DNA and LPS induce distinct patterns of activation in human monocytes. Gene Ther. 1999;6(5):893–903. doi: 10.1038/sj.gt.3300880. [DOI] [PubMed] [Google Scholar]
- 109.Boonstra A., Rajsbaum R., Holman M., Marques R., Asselin-Paturel C., Pereira J.P., Bates E.E., Akira S., Vieira P., Liu Y.J., Trinchieri G., O'Garra A. Macrophages and myeloid dendritic cells, but not plasmacytoid dendritic cells, produce IL-10 in response to MyD88- and TRIF-dependent TLR signals, and TLR-independent signals. J. Immunol. 2006;177(11):7551–7558. doi: 10.4049/jimmunol.177.11.7551. (Baltimore, Md. : 1950) [DOI] [PubMed] [Google Scholar]
- 110.Guiducci C., Ott G., Chan J.H., Damon E., Calacsan C., Matray T., Lee K.D., Coffman R.L., Barrat F.J. Properties regulating the nature of the plasmacytoid dendritic cell response to Toll-like receptor 9 activation. J. Exp. Med. 2006;203(8):1999–2008. doi: 10.1084/jem.20060401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Dunzendorfer S., Lee H.K., Soldau K., Tobias P.S. Toll-like receptor 4 functions intracellularly in human coronary artery endothelial cells: roles of LBP and sCD14 in mediating LPS responses. FASEB J. 2004;18(10):1117–1119. doi: 10.1096/fj.03-1263fje. [DOI] [PubMed] [Google Scholar]
- 112.Baumann C.L., Aspalter I.M., Sharif O., Pichlmair A., Bluml S., Grebien F., Bruckner M., Pasierbek P., Aumayr K., Planyavsky M., Bennett K.L., Colinge J., Knapp S., Superti-Furga G. CD14 is a coreceptor of Toll-like receptors 7 and 9. J. Exp. Med. 2010;207(12):2689–2701. doi: 10.1084/jem.20101111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Sutton N.R., Baek A., Pinsky D.J. Endothelial Cells and Inflammation. In: Mackay I.R., Rose N.R., Diamond B., Davidson A., editors. Encyclopedia of Medical Immunology: Autoimmune Diseases. Springer New York; New York, NY: 2014. pp. 367–381. [Google Scholar]
- 114.Pober J.S., Sessa W.C. Vol. 7. 2007. Evolving functions of endothelial cells in inflammation; p. 803. [DOI] [PubMed] [Google Scholar]
- 115.Castellon X., Bogdanova V. Chronic Inflammatory Diseases and Endothelial Dysfunction. Aging Dis. 2016;7(1):81–89. doi: 10.14336/AD.2015.0803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Guillerey C., Huntington N.D., Smyth M.J. Targeting natural killer cells in cancer immunotherapy. Nat. Immunol. 2016;17:1025. doi: 10.1038/ni.3518. [DOI] [PubMed] [Google Scholar]
- 117.Waggoner S.N., Reighard S.D., Gyurova I.E., Cranert S.A., Mahl S.E., Karmele E.P., McNally J.P., Moran M.T., Brooks T.R., Yaqoob F., Rydyznski C.E. Roles of natural killer cells in antiviral immunity. Curr. Opin. Virol. 2016;16(Supplement C):15–23. doi: 10.1016/j.coviro.2015.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Kumar V., Medhi B. Emerging role of uterine natural killer cells in establishing pregnancy. Iran. J. Immunol. 2008;5(2):71–81. doi: 10.22034/iji.2008.48555. [DOI] [PubMed] [Google Scholar]
- 119.Gaynor L.M., Colucci F. Uterine Natural Killer Cells: Functional Distinctions and Influence on Pregnancy in Humans and Mice. Front. Immunol. 2017;8:467. doi: 10.3389/fimmu.2017.00467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Vacca P., Mingari M.C., Moretta L. Natural killer cells in human pregnancy. J. Reprod. Immunol. 2013;97(1):14–19. doi: 10.1016/j.jri.2012.10.008. [DOI] [PubMed] [Google Scholar]
- 121.Lee Y.-C., Lin S.-J. Neonatal Natural Killer Cell Function: Relevance to Antiviral Immune Defense. Clin. Dev. Immunol. 2013;2013:6. doi: 10.1155/2013/427696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Boyton R.J., Altmann D.M. Natural killer cells, killer immunoglobulin-like receptors and human leucocyte antigen class I in disease. Clin. Exp. Immunol. 2007;149(1):1–8. doi: 10.1111/j.1365-2249.2007.03424.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Boudreau J.E., Hsu K.C. Natural killer cell education in human health and disease. Curr. Opin. Immunol. 2018;50:102–111. doi: 10.1016/j.coi.2017.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Jamieson A.M., Diefenbach A., McMahon C.W., Xiong N., Carlyle J.R., Raulet D.H. The role of the NKG2D immunoreceptor in immune cell activation and natural killing. Immunity. 2002;17(1):19–29. doi: 10.1016/s1074-7613(02)00333-3. [DOI] [PubMed] [Google Scholar]
- 125.Spear P., Wu M.-R., Sentman M.-L., Sentman C.L. NKG2D ligands as therapeutic targets. Cancer Immun. 2013;13:8. [PMC free article] [PubMed] [Google Scholar]
- 126.Berry R., Rossjohn J., Brooks A.G. The Ly49 natural killer cell receptors: a versatile tool for viral self-discrimination. Immunol. Cell Biol. 2014;92:214. doi: 10.1038/icb.2013.100. [DOI] [PubMed] [Google Scholar]
- 127.Rahim M.M.A., Tu M.M., Mahmoud A.B., Wight A., Abou-Samra E., Lima P.D.A., Makrigiannis A.P. Ly49 Receptors: Innate and Adaptive Immune Paradigms. Front. Immunol. 2014;5:145. doi: 10.3389/fimmu.2014.00145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Mager D.L., McQueen K.L., Wee V., Freeman J.D. Evolution of natural killer cell receptors: coexistence of functional Ly49 and KIR genes in baboons. Curr. Biol. 2001;11(8):626–630. doi: 10.1016/s0960-9822(01)00148-8. [DOI] [PubMed] [Google Scholar]
- 129.Vivier E., Tomasello E., Baratin M., Walzer T., Ugolini S. Functions of natural killer cells. Nat. Immunol. 2008;9(5):503–510. doi: 10.1038/ni1582. [DOI] [PubMed] [Google Scholar]
- 130.Thielens A., Vivier E., Romagné F. NK cell MHC class I specific receptors (KIR): from biology to clinical intervention. Curr. Opin. Immunol. 2012;24(2):239–245. doi: 10.1016/j.coi.2012.01.001. [DOI] [PubMed] [Google Scholar]
- 131.Boudreau J.E., Hsu K.C. Natural Killer Cell Education and the Response to Infection and Cancer Therapy: Stay Tuned. Trends Immunol. 2018;39(3):222–239. doi: 10.1016/j.it.2017.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Ashouri E., Dabbaghmanesh M.H., Ranjbar Omrani G. Presence of more activating KIR genes is associated with Hashimoto's thyroiditis. Endocrine. 2014;46(3):519–525. doi: 10.1007/s12020-013-0080-2. [DOI] [PubMed] [Google Scholar]
- 133.Li J.T., Guo C., Li M.L., Wei Y.Q., Hou Y.F., Jiao Y.L., Zhao Y.R., Sun H., Xu J., Cao M.F., Feng L., Yu G.N., Gao L., Liu Y.Q., Zhang B.C., Zhao J.J., Zhang H.Q. Killer cell immunoglobulin-like receptor genes and their hla-c ligands in hashimoto thyroiditis in a chinese population. Endocr. Pract. 2016;22(8):935–940. doi: 10.4158/EP151175.OR. [DOI] [PubMed] [Google Scholar]
- 134.Vilches C., Parham P. KIR: diverse, rapidly evolving receptors of innate and adaptive immunity. Annu. Rev. Immunol. 2002;20:217–251. doi: 10.1146/annurev.immunol.20.092501.134942. [DOI] [PubMed] [Google Scholar]
- 135.Adib-Conquy M., Scott-Algara D., Cavaillon J.-M., Souza-Fonseca-Guimaraes F. TLR-mediated activation of NK cells and their role in bacterial/viral immune responses in mammals. Immunol. Cell Biol. 2013;92:256. doi: 10.1038/icb.2013.99. [DOI] [PubMed] [Google Scholar]
- 136.Sivori S., Falco M., Carlomagno S., Romeo E., Soldani C., Bensussan A., Viola A., Moretta L., Moretta A. A novel KIR-associated function: evidence that CpG DNA uptake and shuttling to early endosomes is mediated by KIR3DL2. Blood. 2010;116(10):1637–1647. doi: 10.1182/blood-2009-12-256586. [DOI] [PubMed] [Google Scholar]
- 137.Lodoen M.B., Lanier L.L. Natural killer cells as an initial defense against pathogens. Curr. Opin. Immunol. 2006;18(4):391–398. doi: 10.1016/j.coi.2006.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Sivori S., Falco M., Moretta L., Moretta A. Extending killer Ig-like receptor function: from HLA class I recognition to sensors of microbial products. Trends Immunol. 2010;31(8):289–294. doi: 10.1016/j.it.2010.05.007. [DOI] [PubMed] [Google Scholar]
- 139.Guo Q., Zhang C. Critical role of Toll-like receptor signaling in NK cell activation. Chin. Sci. Bull. 2012;57(24):3192–3202. [Google Scholar]
- 140.Vankayalapati R., Garg A., Porgador A., Griffith D.E., Klucar P., Safi H., Girard W.M., Cosman D., Spies T., Barnes P.F. Role of NK cell-activating receptors and their ligands in the lysis of mononuclear phagocytes infected with an intracellular bacterium. J. Immunol. 2005;175(7):4611–4617. doi: 10.4049/jimmunol.175.7.4611. (Baltimore, Md. : 1950) [DOI] [PubMed] [Google Scholar]
- 141.Guerra C., Johal K., Morris D., Moreno S., Alvarado O., Gray D., Tanzil M., Pearce D., Venketaraman V. Control of Mycobacterium tuberculosis growth by activated natural killer cells. Clin. Exp. Immunol. 2012;168(1):142–152. doi: 10.1111/j.1365-2249.2011.04552.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Lauzon N.M., Mian F., MacKenzie R., Ashkar A.A. The direct effects of Toll-like receptor ligands on human NK cell cytokine production and cytotoxicity. Cell. Immunol. 2006;241(2):102–112. doi: 10.1016/j.cellimm.2006.08.004. [DOI] [PubMed] [Google Scholar]
- 143.Chalifour A., Jeannin P., Gauchat J.F., Blaecke A., Malissard M., N'Guyen T., Thieblemont N., Delneste Y. Direct bacterial protein PAMP recognition by human NK cells involves TLRs and triggers alpha-defensin production. Blood. 2004;104(6):1778–1783. doi: 10.1182/blood-2003-08-2820. [DOI] [PubMed] [Google Scholar]
- 144.Popko K., Górska E. The role of natural killer cells in pathogenesis of autoimmune diseases. Cent. Eur. J. Immunol. 2015;40(4):470–476. doi: 10.5114/ceji.2015.56971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Huang Q.-Q., Pope R.M. Role of toll like receptors in rheumatoid arthritis. Curr. Rheumatol. Rep. 2009;11(5):357–364. doi: 10.1007/s11926-009-0051-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Tadema H., Abdulahad W.H., Stegeman C.A., Kallenberg C.G., Heeringa P. Increased expression of Toll-like receptors by monocytes and natural killer cells in ANCA-associated vasculitis. PLoS One. 2011;6(9) doi: 10.1371/journal.pone.0024315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Adib-Conquy M., Scott-Algara D., Cavaillon J.M., Souza-Fonseca-Guimaraes F. TLR-mediated activation of NK cells and their role in bacterial/viral immune responses in mammals. Immunol. Cell Biol. 2014;92(3):256–262. doi: 10.1038/icb.2013.99. [DOI] [PubMed] [Google Scholar]
- 148.Saikh K.U., Lee J.S., Kissner T.L., Dyas B., Ulrich R.G. Toll-like receptor and cytokine expression patterns of CD56+ T cells are similar to natural killer cells in response to infection with Venezuelan equine encephalitis virus replicons. J. Infect. Dis. 2003;188(10):1562–1570. doi: 10.1086/379196. [DOI] [PubMed] [Google Scholar]
- 149.Eriksson M., Meadows S.K., Basu S., Mselle T.F., Wira C.R., Sentman C.L. TLRs mediate IFN-gamma production by human uterine NK cells in endometrium. J. Immunol. 2006;176(10):6219–6224. doi: 10.4049/jimmunol.176.10.6219. [DOI] [PubMed] [Google Scholar]
- 150.Comin F., Speziali E., Martins-Filho O.A., Caldas I.R., Moura V., Gazzinelli A., Correa-Oliveira R., Faria A.M. Ageing and Toll-like receptor expression by innate immune cells in chronic human schistosomiasis. Clin. Exp. Immunol. 2007;149(2):274–284. doi: 10.1111/j.1365-2249.2007.03403.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Souza-Fonseca-Guimaraes F., Parlato M., Fitting C., Cavaillon J.M., Adib-Conquy M. NK cell tolerance to TLR agonists mediated by regulatory T cells after polymicrobial sepsis. J. Immunol. 2012;188(12):5850–5858. doi: 10.4049/jimmunol.1103616. [DOI] [PubMed] [Google Scholar]
- 152.Souza-Fonseca-Guimaraes F., Parlato M., Philippart F., Misset B., Cavaillon J.M., Adib-Conquy M. Toll-like receptors expression and interferon-gamma production by NK cells in human sepsis. Crit. Care. 2012;16(5):R206. doi: 10.1186/cc11838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Zipris D. Toll-Like Receptors and Type 1 Diabetes. In: Islam M.S., editor. The Islets of Langerhans. Springer Netherlands; Dordrecht: 2010. pp. 585–610. [Google Scholar]
- 154.Zipris D., Lien E., Nair A., Xie J.X., Greiner D.L., Mordes J.P., Rossini A.A. TLR9-signaling pathways are involved in Kilham rat virus-induced autoimmune diabetes in the biobreeding diabetes-resistant rat. J. Immunol. 2007;178(2):693–701. doi: 10.4049/jimmunol.178.2.693. [DOI] [PubMed] [Google Scholar]
- 155.Flodstrom M., Maday A., Balakrishna D., Cleary M.M., Yoshimura A., Sarvetnick N. Target cell defense prevents the development of diabetes after viral infection. Nat. Immunol. 2002;3(4):373–382. doi: 10.1038/ni771. [DOI] [PubMed] [Google Scholar]
- 156.Souza-Fonseca-Guimaraes F., Adib-Conquy M., Cavaillon J.M. Natural killer (NK) cells in antibacterial innate immunity: angels or devils? Mol. Med. 2012;18:270–285. doi: 10.2119/molmed.2011.00201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Ebihara T., Azuma M., Oshiumi H., Kasamatsu J., Iwabuchi K., Matsumoto K., Saito H., Taniguchi T., Matsumoto M., Seya T. Identification of a polyI:C-inducible membrane protein that participates in dendritic cell-mediated natural killer cell activation. J. Exp. Med. 2010;207(12):2675–2687. doi: 10.1084/jem.20091573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Millard A.L., Spirig R., Mueller N.J., Seebach J.D., Rieben R. Inhibition of direct and indirect TLR-mediated activation of human NK cells by low molecular weight dextran sulfate. Mol. Immunol. 2010;47(14):2349–2358. doi: 10.1016/j.molimm.2010.05.284. [DOI] [PubMed] [Google Scholar]
- 159.Newman K.C., Riley E.M. Whatever turns you on: accessory-cell-dependent activation of NK cells by pathogens. Nat. Rev. Immunol. 2007;7(4):279–291. doi: 10.1038/nri2057. [DOI] [PubMed] [Google Scholar]
- 160.Etogo A.O., Nunez J., Lin C.Y., Toliver-Kinsky T.E., Sherwood E.R. NK but not CD1-restricted NKT cells facilitate systemic inflammation during polymicrobial intra-abdominal sepsis. J. Immunol. 2008;180(9):6334–6345. doi: 10.4049/jimmunol.180.9.6334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Badgwell B., Parihar R., Magro C., Dierksheide J., Russo T., Carson W.E., 3rd Natural killer cells contribute to the lethality of a murine model of Escherichia coli infection. Surgery. 2002;132(2):205–212. doi: 10.1067/msy.2002.125311. [DOI] [PubMed] [Google Scholar]
- 162.Andaluz-Ojeda D., Iglesias V., Bobillo F., Almansa R., Rico L., Gandia F., Loma A.M., Nieto C., Diego R., Ramos E., Nocito M., Resino S., Eiros J.M., Tamayo E., de Lejarazu R.O., Bermejo-Martin J.F. Early natural killer cell counts in blood predict mortality in severe sepsis. Crit. Care. 2011;15(5):R243. doi: 10.1186/cc10501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Sivori S., Falco M., Chiesa M.D., Carlomagno S., Vitale M., Moretta L., Moretta A. CpG and double-stranded RNA trigger human NK cells by Toll-like receptors: Induction of cytokine release and cytotoxicity against tumors and dendritic cells. Proc. Natl. Acad. Sci. U. S. A. 2004;101(27):10116–10121. doi: 10.1073/pnas.0403744101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Shortman K., Liu Y.J. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2002;2(3):151–161. doi: 10.1038/nri746. [DOI] [PubMed] [Google Scholar]
- 165.Schreibelt G., Tel J., Sliepen K.H., Benitez-Ribas D., Figdor C.G., Adema G.J., de Vries I.J. Toll-like receptor expression and function in human dendritic cell subsets: implications for dendritic cell-based anti-cancer immunotherapy. Cancer Immunol. Immunother. 2010;59(10):1573–1582. doi: 10.1007/s00262-010-0833-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Hochrein H., O'Keeffe M., Wagner H. Human and mouse plasmacytoid dendritic cells. Hum. Immunol. 2002;63(12):1103–1110. doi: 10.1016/s0198-8859(02)00748-6. [DOI] [PubMed] [Google Scholar]
- 167.Rothenfusser S., Hornung V., Krug A., Towarowski A., Krieg A.M., Endres S., Hartmann G. Distinct CpG oligonucleotide sequences activate human gamma delta T cells via interferon-alpha/-beta. Eur. J. Immunol. 2001;31(12):3525–3534. doi: 10.1002/1521-4141(200112)31:12<3525::aid-immu3525>3.0.co;2-5. [DOI] [PubMed] [Google Scholar]
- 168.Proietto A.I., O'Keeffe M., Gartlan K., Wright M.D., Shortman K., Wu L., Lahoud M.H. Differential production of inflammatory chemokines by murine dendritic cell subsets. Immunobiology. 2004;209(1–2):163–172. doi: 10.1016/j.imbio.2004.03.002. [DOI] [PubMed] [Google Scholar]
- 169.Kawai T., Sato S., Ishii K.J., Coban C., Hemmi H., Yamamoto M., Terai K., Matsuda M., Inoue J., Uematsu S., Takeuchi O., Akira S. Interferon-alpha induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88 and TRAF6. Nat. Immunol. 2004;5(10):1061–1068. doi: 10.1038/ni1118. [DOI] [PubMed] [Google Scholar]
- 170.Honda K., Yanai H., Mizutani T., Negishi H., Shimada N., Suzuki N., Ohba Y., Takaoka A., Yeh W.C., Taniguchi T. Role of a transductional-transcriptional processor complex involving MyD88 and IRF-7 in Toll-like receptor signaling. Proc. Natl. Acad. Sci. U. S. A. 2004;101(43):15416–15421. doi: 10.1073/pnas.0406933101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Fuchsberger M., Hochrein H., O'Keeffe M. Activation of plasmacytoid dendritic cells. Immunol. Cell Biol. 2005;83(5):571–577. doi: 10.1111/j.1440-1711.2005.01392.x. [DOI] [PubMed] [Google Scholar]
- 172.Uematsu S., Sato S., Yamamoto M., Hirotani T., Kato H., Takeshita F., Matsuda M., Coban C., Ishii K.J., Kawai T., Takeuchi O., Akira S. Interleukin-1 receptor-associated kinase-1 plays an essential role for Toll-like receptor (TLR)7- and TLR9-mediated interferon-{alpha} induction. J. Exp. Med. 2005;201(6):915–923. doi: 10.1084/jem.20042372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.MacDonald K.P., Munster D.J., Clark G.J., Dzionek A., Schmitz J., Hart D.N. Characterization of human blood dendritic cell subsets. Blood. 2002;100(13):4512–4520. doi: 10.1182/blood-2001-11-0097. [DOI] [PubMed] [Google Scholar]
- 174.Piccioli D., Tavarini S., Borgogni E., Steri V., Nuti S., Sammicheli C., Bardelli M., Montagna D., Locatelli F., Wack A. Functional specialization of human circulating CD16 and CD1c myeloid dendritic-cell subsets. Blood. 2007;109(12):5371–5379. doi: 10.1182/blood-2006-08-038422. [DOI] [PubMed] [Google Scholar]
- 175.Ito T., Amakawa R., Kaisho T., Hemmi H., Tajima K., Uehira K., Ozaki Y., Tomizawa H., Akira S., Fukuhara S. Interferon-alpha and interleukin-12 are induced differentially by Toll-like receptor 7 ligands in human blood dendritic cell subsets. J. Exp. Med. 2002;195(11):1507–1512. doi: 10.1084/jem.20020207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Means T.K., Hayashi F., Smith K.D., Aderem A., Luster A.D. The Toll-like receptor 5 stimulus bacterial flagellin induces maturation and chemokine production in human dendritic cells. J. Immunol. 2003;170(10):5165–5175. doi: 10.4049/jimmunol.170.10.5165. [DOI] [PubMed] [Google Scholar]
- 177.Perera P.Y., Mayadas T.N., Takeuchi O., Akira S., Zaks-Zilberman M., Goyert S.M., Vogel S.N. CD11b/CD18 acts in concert with CD14 and Toll-like receptor (TLR) 4 to elicit full lipopolysaccharide and taxol-inducible gene expression. J. Immunol. 2001;166(1):574–581. doi: 10.4049/jimmunol.166.1.574. [DOI] [PubMed] [Google Scholar]
- 178.Ling G.S., Bennett J., Woollard K.J., Szajna M., Fossati-Jimack L., Taylor P.R., Scott D., Franzoso G., Cook H.T., Botto M. Integrin CD11b positively regulates TLR4-induced signalling pathways in dendritic cells but not in macrophages. Nat. Commun. 2014;5:3039. doi: 10.1038/ncomms4039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Matsuzawa A., Saegusa K., Noguchi T., Sadamitsu C., Nishitoh H., Nagai S., Koyasu S., Matsumoto K., Takeda K., Ichijo H. ROS-dependent activation of the TRAF6-ASK1-p38 pathway is selectively required for TLR4-mediated innate immunity. Nat. Immunol. 2005;6(6):587–592. doi: 10.1038/ni1200. [DOI] [PubMed] [Google Scholar]
- 180.Mnich S.J., Blanner P.M., Hu L.G., Shaffer A.F., Happa F.A., O'Neil S., Ukairo O., Weiss D., Welsh E., Storer C., Mbalaviele G., Ichijo H., Monahan J.B., Hardy M.M., Eda H. Critical role for apoptosis signal-regulating kinase 1 in the development of inflammatory K/BxN serum-induced arthritis. Int. Immunopharmacol. 2010;10(10):1170–1176. doi: 10.1016/j.intimp.2010.06.023. [DOI] [PubMed] [Google Scholar]
- 181.Fujino G., Noguchi T., Matsuzawa A., Yamauchi S., Saitoh M., Takeda K., Ichijo H. Thioredoxin and TRAF family proteins regulate reactive oxygen species-dependent activation of ASK1 through reciprocal modulation of the N-terminal homophilic interaction of ASK1. Mol. Cell. Biol. 2007;27(23):8152–8163. doi: 10.1128/MCB.00227-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Noguchi T., Takeda K., Matsuzawa A., Saegusa K., Nakano H., Gohda J., Inoue J., Ichijo H. Recruitment of tumor necrosis factor receptor-associated factor family proteins to apoptosis signal-regulating kinase 1 signalosome is essential for oxidative stress-induced cell death. J. Biol. Chem. 2005;280(44):37033–37040. doi: 10.1074/jbc.M506771200. [DOI] [PubMed] [Google Scholar]
- 183.Fan X., Liu Z., Jin H., Yan J., Liang H.P. Alterations of dendritic cells in sepsis: featured role in immunoparalysis. Biomed. Res. Int. 2015;2015:903720. doi: 10.1155/2015/903720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Hotchkiss R.S., Tinsley K.W., Swanson P.E., Grayson M.H., Osborne D.F., Wagner T.H., Cobb J.P., Coopersmith C., Karl I.E. Depletion of dendritic cells, but not macrophages, in patients with sepsis. J. Immunol. 2002;168(5):2493–2500. doi: 10.4049/jimmunol.168.5.2493. [DOI] [PubMed] [Google Scholar]
- 185.Toliver-Kinsky T.E., Lin C.Y., Herndon D.N., Sherwood E.R. Stimulation of hematopoiesis by the Fms-like tyrosine kinase 3 ligand restores bacterial induction of Th1 cytokines in thermally injured mice. Infect. Immun. 2003;71(6):3058–3067. doi: 10.1128/IAI.71.6.3058-3067.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Toliver-Kinsky T.E., Cui W., Murphey E.D., Lin C., Sherwood E.R. Enhancement of dendritic cell production by fms-like tyrosine kinase-3 ligand increases the resistance of mice to a burn wound infection. J. Immunol. 2005;174(1):404–410. doi: 10.4049/jimmunol.174.1.404. [DOI] [PubMed] [Google Scholar]
- 187.Bohannon J., Cui W., Sherwood E., Toliver-Kinsky T. Dendritic cell modification of neutrophil responses to infection after burn injury. J. Immunol. 2010;185(5):2847–2853. doi: 10.4049/jimmunol.0903619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Hotchkiss R.S., Monneret G., Payen D. Sepsis-induced immunosuppression: from cellular dysfunctions to immunotherapy. Nat. Rev. Immunol. 2013;13(12):862–874. doi: 10.1038/nri3552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Benjamim C.F., Hogaboam C.M., Lukacs N.W., Kunkel S.L. Septic mice are susceptible to pulmonary aspergillosis. Am. J. Pathol. 2003;163(6):2605–2617. doi: 10.1016/S0002-9440(10)63615-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Roquilly A., McWilliam H.E.G., Jacqueline C., Tian Z., Cinotti R., Rimbert M., Wakim L., Caminschi I., Lahoud M.H., Belz G.T., Kallies A., Mintern J.D., Asehnoune K., Villadangos J.A. Local Modulation of Antigen-Presenting Cell Development after Resolution of Pneumonia Induces Long-Term Susceptibility to Secondary Infections. Immunity. 2017;47(1):135-147.e5. doi: 10.1016/j.immuni.2017.06.021. [DOI] [PubMed] [Google Scholar]
- 191.Benjamim C.F., Lundy S.K., Lukacs N.W., Hogaboam C.M., Kunkel S.L. Reversal of long-term sepsis-induced immunosuppression by dendritic cells. Blood. 2005;105(9):3588–3595. doi: 10.1182/blood-2004-08-3251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Roquilly A., Broquet A., Jacqueline C., Gautreau L., Segain J.P., de Coppet P., Caillon J., Altare F., Josien R., Asehnoune K. Toll-like receptor-4 agonist in post-haemorrhage pneumonia: role of dendritic and natural killer cells. Eur. Respir. J. 2013;42(5):1365–1378. doi: 10.1183/09031936.00152612. [DOI] [PubMed] [Google Scholar]
- 193.Dar A.A., Patil R.S., Chiplunkar S.V. Insights into the Relationship between Toll Like Receptors and Gamma Delta T Cell Responses. Front. Immunol. 2014;5:366. doi: 10.3389/fimmu.2014.00366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Fay N.S., Larson E.C., Jameson J.M. Chronic Inflammation and γδ T Cells. Front. Immunol. 2016;7:210. doi: 10.3389/fimmu.2016.00210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Catalan-Serra I., Sandvik A.K., Bruland T., Andreu-Ballester J.C. Gammadelta T Cells in Crohn's Disease: A New Player in the Disease Pathogenesis? J. Crohns Colitis. 2017;11(9):1135–1145. doi: 10.1093/ecco-jcc/jjx039. [DOI] [PubMed] [Google Scholar]
- 196.Papotto P.H., Reinhardt A., Prinz I., Silva-Santos B. Innately versatile: γδ17 T cells in inflammatory and autoimmune diseases. J. Autoimmun. 2018;87:26–37. doi: 10.1016/j.jaut.2017.11.006. [DOI] [PubMed] [Google Scholar]
- 197.Gelderblom M., Arunachalam P., Magnus T. γδ T cells as early sensors of tissue damage and mediators of secondary neurodegeneration. Front. Cell. Neurosci. 2014;8:368. doi: 10.3389/fncel.2014.00368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Nielsen M.M., Witherden D.A., Havran W.L. γδ T cells in homeostasis and host defence of epithelial barrier tissues. Nat. Rev. Immunol. 2017;17:733. doi: 10.1038/nri.2017.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Jin Y., Xia M., Saylor C.M., Narayan K., Kang J., Wiest D.L., Wang Y., Xiong N. Cutting edge: Intrinsic programming of thymic gammadeltaT cells for specific peripheral tissue localization. J. Immunol. 2010;185(12):7156–7160. doi: 10.4049/jimmunol.1002781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Born W.K., Reardon C.L., O'Brien R.L. The function of gammadelta T cells in innate immunity. Curr. Opin. Immunol. 2006;18(1):31–38. doi: 10.1016/j.coi.2005.11.007. [DOI] [PubMed] [Google Scholar]
- 201.Hayday A.C., Roberts S., Ramsburg E. gammadelta cells and the regulation of mucosal immune responses. Am. J. Respir. Crit. Care Med. 2000;162(4 Pt 2):S161–3. doi: 10.1164/ajrccm.162.supplement_3.15tac4. [DOI] [PubMed] [Google Scholar]
- 202.Hayday A.C. [gamma][delta] cells: a right time and a right place for a conserved third way of protection. Annu. Rev. Immunol. 2000;18:975–1026. doi: 10.1146/annurev.immunol.18.1.975. [DOI] [PubMed] [Google Scholar]
- 203.Alcaide M.L., Strbo N., Romero L., Jones D.L., Rodriguez V.J., Arheart K., Martinez O., Bolivar H., Podack E.R., Fischl M.A. Bacterial Vaginosis Is Associated with Loss of Gamma Delta T Cells in the Female Reproductive Tract in Women in the Miami Women Interagency HIV Study (WIHS): A Cross Sectional Study. PLoS ONE. 2016;11(4) doi: 10.1371/journal.pone.0153045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Vroom T.M., Scholte G., Ossendorp F., Borst J. Tissue distribution of human gamma delta T cells: no evidence for general epithelial tropism. J. Clin. Pathol. 1991;44(12):1012–1017. doi: 10.1136/jcp.44.12.1012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Sheridan Brian S., Romagnoli Pablo A., Pham Q.-M., Fu H.-H., Alonzo F., Schubert W.-D., Freitag Nancy E., Lefrançois L. γδ T Cells Exhibit Multifunctional and Protective Memory in Intestinal Tissues. Immunity. 2013;39(1):184–195. doi: 10.1016/j.immuni.2013.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Chennupati V., Worbs T., Liu X., Malinarich F.H., Schmitz S., Haas J.D., Malissen B., Forster R., Prinz I. Intra- and intercompartmental movement of gammadelta T cells: intestinal intraepithelial and peripheral gammadelta T cells represent exclusive nonoverlapping populations with distinct migration characteristics. J. Immunol. 2010;185(9):5160–5168. doi: 10.4049/jimmunol.1001652. [DOI] [PubMed] [Google Scholar]
- 207.Vantourout P., Hayday A. Six-of-the-best: unique contributions of γδ T cells to immunology. Nat. Rev. Immunol. 2013;13(2):88–100. doi: 10.1038/nri3384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Cho J.S., Pietras E.M., Garcia N.C., Ramos R.I., Farzam D.M., Monroe H.R., Magorien J.E., Blauvelt A., Kolls J.K., Cheung A.L., Cheng G., Modlin R.L., Miller L.S. IL-17 is essential for host defense against cutaneous Staphylococcus aureus infection in mice. J. Clin. Invest. 2010;120(5):1762–1773. doi: 10.1172/JCI40891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Molne L., Corthay A., Holmdahl R., Tarkowski A. Role of gamma/delta T cell receptor-expressing lymphocytes in cutaneous infection caused by Staphylococcus aureus. Clin. Exp. Immunol. 2003;132(2):209–215. doi: 10.1046/j.1365-2249.2003.02151.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Asarnow D.M., Kuziel W.A., Bonyhadi M., Tigelaar R.E., Tucker P.W., Allison J.P. Limited diversity of gamma delta antigen receptor genes of Thy-1+ dendritic epidermal cells. Cell. 1988;55(5):837–847. doi: 10.1016/0092-8674(88)90139-0. [DOI] [PubMed] [Google Scholar]
- 211.Andreu-Ballester J.C., Tormo-Calandin C., Garcia-Ballesteros C., Perez-Griera J., Amigo V., Almela-Quilis A., Ruiz del Castillo J., Penarroja-Otero C., Ballester F. Association of gammadelta T cells with disease severity and mortality in septic patients. Clin. Vaccine Immunol. 2013;20(5):738–746. doi: 10.1128/CVI.00752-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Galley H.F., Lowes D.A., Thompson K., Wilson N.D., Wallace C.A., Webster N.R. Characterisation of gamma delta (gammadelta) T cell populations in patients with sepsis. Cell Biol. Int. 2015;39(2):210–216. doi: 10.1002/cbin.10361. [DOI] [PubMed] [Google Scholar]
- 213.Tschop J., Martignoni A., Goetzman H.S., Choi L.G., Wang Q., Noel J.G., Ogle C.K., Pritts T.A., Johannigman J.A., Lentsch A.B., Caldwell C.C. Gammadelta T cells mitigate the organ injury and mortality of sepsis. J. Leukoc. Biol. 2008;83(3):581–588. doi: 10.1189/jlb.0707507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Kim A., Lang T., Xue M., Wijewardana A., Jackson C., Vandervord J. The Role of Th-17 Cells and gammadelta T-Cells in Modulating the Systemic Inflammatory Response to Severe Burn Injury. Int. J. Mol. Sci. 2017;18(4) doi: 10.3390/ijms18040758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Zheng J., Liu Y., Lau Y.L., Tu W. gammadelta-T cells: an unpolished sword in human anti-infection immunity. Cell. Mol. Immunol. 2013;10(1):50–57. doi: 10.1038/cmi.2012.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Schwacha M.G., Rani M., Zhang Q., Nunez-Cantu O., Cap A.P. Mitochondrial damage-associated molecular patterns activate gammadelta T-cells. Innate Immun. 2014;20(3):261–268. doi: 10.1177/1753425913488969. [DOI] [PubMed] [Google Scholar]
- 217.Schwacha M.G., Rani M., Nicholson S.E., Lewis A.M., Holloway T.L., Sordo S., Cap A.P. Dermal gammadelta T-Cells Can Be Activated by Mitochondrial Damage-Associated Molecular Patterns. PLoS One. 2016;11(7) doi: 10.1371/journal.pone.0158993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Devilder M.C., Allain S., Dousset C., Bonneville M., Scotet E. Early triggering of exclusive IFN-gamma responses of human Vgamma9Vdelta2 T cells by TLR-activated myeloid and plasmacytoid dendritic cells. J. Immunol. 2009;183(6):3625–3633. doi: 10.4049/jimmunol.0901571. [DOI] [PubMed] [Google Scholar]
- 219.Pietschmann K., Beetz S., Welte S., Martens I., Gruen J., Oberg H.H., Wesch D., Kabelitz D. Toll-like receptor expression and function in subsets of human gammadelta T lymphocytes. Scand. J. Immunol. 2009;70(3):245–255. doi: 10.1111/j.1365-3083.2009.02290.x. [DOI] [PubMed] [Google Scholar]
- 220.Wesch D., Peters C., Oberg H.H., Pietschmann K., Kabelitz D. Modulation of gammadelta T cell responses by TLR ligands. Cell. Mol. Life Sci. 2011;68(14):2357–2370. doi: 10.1007/s00018-011-0699-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Wesch D., Beetz S., Oberg H.H., Marget M., Krengel K., Kabelitz D. Direct costimulatory effect of TLR3 ligand poly(I:C) on human gamma delta T lymphocytes. J. Immunol. 2006;176(3):1348–1354. doi: 10.4049/jimmunol.176.3.1348. [DOI] [PubMed] [Google Scholar]
- 222.Beetz S., Wesch D., Marischen L., Welte S., Oberg H.H., Kabelitz D. Innate immune functions of human gammadelta T cells. Immunobiology. 2008;213(3–4):173–182. doi: 10.1016/j.imbio.2007.10.006. [DOI] [PubMed] [Google Scholar]
- 223.Deetz C.O., Hebbeler A.M., Propp N.A., Cairo C., Tikhonov I., Pauza C.D. Gamma interferon secretion by human Vgamma2Vdelta2 T cells after stimulation with antibody against the T-cell receptor plus the Toll-Like receptor 2 agonist Pam3Cys. Infect. Immun. 2006;74(8):4505–4511. doi: 10.1128/IAI.00088-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Hedges J.F., Lubick K.J., Jutila M.A. Gamma delta T cells respond directly to pathogen-associated molecular patterns. J. Immunol. 2005;174(10):6045–6053. doi: 10.4049/jimmunol.174.10.6045. [DOI] [PubMed] [Google Scholar]
- 225.Gabrilovich D.I., Nagaraj S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009;9(3):162–174. doi: 10.1038/nri2506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Condamine T., Gabrilovich D.I. Molecular mechanisms regulating myeloid-derived suppressor cell differentiation and function. Trends Immunol. 2011;32(1):19–25. doi: 10.1016/j.it.2010.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Almand B., Clark J.I., Nikitina E., van Beynen J., English N.R., Knight S.C., Carbone D.P., Gabrilovich D.I. Increased production of immature myeloid cells in cancer patients: a mechanism of immunosuppression in cancer. J. Immunol. 2001;166(1):678–689. doi: 10.4049/jimmunol.166.1.678. [DOI] [PubMed] [Google Scholar]
- 228.Peranzoni E., Zilio S., Marigo I., Dolcetti L., Zanovello P., Mandruzzato S., Bronte V. Myeloid-derived suppressor cell heterogeneity and subset definition. Curr. Opin. Immunol. 2010;22(2):238–244. doi: 10.1016/j.coi.2010.01.021. [DOI] [PubMed] [Google Scholar]
- 229.Nagaraj S., Gabrilovich D.I. Myeloid-derived suppressor cells in human cancer. Cancer J. 2010;16(4):348–353. doi: 10.1097/PPO.0b013e3181eb3358. [DOI] [PubMed] [Google Scholar]
- 230.Ribechini E., Greifenberg V., Sandwick S., Lutz M.B. Subsets, expansion and activation of myeloid-derived suppressor cells. Med. Microbiol. Immunol. 2010;199(3):273–281. doi: 10.1007/s00430-010-0151-4. [DOI] [PubMed] [Google Scholar]
- 231.Veglia F., Perego M., Gabrilovich D. Myeloid-derived suppressor cells coming of age. Nat. Immunol. 2018;19(2):108–119. doi: 10.1038/s41590-017-0022-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Maruyama A., Shime H., Takeda Y., Azuma M., Matsumoto M., Seya T. Pam2 lipopeptides systemically increase myeloid-derived suppressor cells through TLR2 signaling. Biochem. Biophys. Res. Commun. 2015;457(3):445–450. doi: 10.1016/j.bbrc.2015.01.011. [DOI] [PubMed] [Google Scholar]
- 233.Bunt S.K., Clements V.K., Hanson E.M., Sinha P., Ostrand-Rosenberg S. Inflammation enhances myeloid-derived suppressor cell cross-talk by signaling through Toll-like receptor 4. J. Leukoc. Biol. 2009;85(6):996–1004. doi: 10.1189/jlb.0708446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Delano M.J., Scumpia P.O., Weinstein J.S., Coco D., Nagaraj S., Kelly-Scumpia K.M., O'Malley K.A., Wynn J.L., Antonenko S., Al-Quran S.Z., Swan R., Chung C.S., Atkinson M.A., Ramphal R., Gabrilovich D.I., Reeves W.H., Ayala A., Phillips J., Laface D., Heyworth P.G., Clare-Salzler M., Moldawer L.L. MyD88-dependent expansion of an immature GR-1(+)CD11b(+) population induces T cell suppression and Th2 polarization in sepsis. J. Exp. Med. 2007;204(6):1463–1474. doi: 10.1084/jem.20062602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Zoglmeier C., Bauer H., Norenberg D., Wedekind G., Bittner P., Sandholzer N., Rapp M., Anz D., Endres S., Bourquin C. CpG blocks immunosuppression by myeloid-derived suppressor cells in tumor-bearing mice. Clin. Cancer Res. 2011;17(7):1765–1775. doi: 10.1158/1078-0432.CCR-10-2672. [DOI] [PubMed] [Google Scholar]
- 236.Shirota Y., Shirota H., Klinman D.M. Intratumoral injection of CpG oligonucleotides induces the differentiation and reduces the immunosuppressive activity of myeloid-derived suppressor cells. J. Immunol. 2012;188(4):1592–1599. doi: 10.4049/jimmunol.1101304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Ost M., Singh A., Peschel A., Mehling R., Rieber N., Hartl D. Myeloid-Derived Suppressor Cells in Bacterial Infections. Front. Cell. Infect. Microbiol. 2016;6:37. doi: 10.3389/fcimb.2016.00037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Brudecki L., Ferguson D.A., McCall C.E., El Gazzar M. Myeloid-derived suppressor cells evolve during sepsis and can enhance or attenuate the systemic inflammatory response. Infect. Immun. 2012;80(6):2026–2034. doi: 10.1128/IAI.00239-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Cuenca A.G., Delano M.J., Kelly-Scumpia K.M., Moreno C., Scumpia P.O., Laface D.M., Heyworth P.G., Efron P.A., Moldawer L.L. A paradoxical role for myeloid-derived suppressor cells in sepsis and trauma. Mol. Med. 2011;17(3–4):281–292. doi: 10.2119/molmed.2010.00178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Derive M., Bouazza Y., Alauzet C., Gibot S. Myeloid-derived suppressor cells control microbial sepsis. Intensive Care Med. 2012;38(6):1040–1049. doi: 10.1007/s00134-012-2574-4. [DOI] [PubMed] [Google Scholar]
- 241.Sander L.E., Sackett S.D., Dierssen U., Beraza N., Linke R.P., Muller M., Blander J.M., Tacke F., Trautwein C. Hepatic acute-phase proteins control innate immune responses during infection by promoting myeloid-derived suppressor cell function. J. Exp. Med. 2010;207(7):1453–1464. doi: 10.1084/jem.20091474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Cuenca A.G., Moldawer L.L. Myeloid-derived suppressor cells in sepsis: friend or foe? Intensive Care Med. 2012;38(6):928–930. doi: 10.1007/s00134-012-2575-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Dai J., El Gazzar M., Li G.Y., Moorman J.P., Yao Z.Q. Myeloid-derived suppressor cells: paradoxical roles in infection and immunity. J. Innate Immun. 2015;7(2):116–126. doi: 10.1159/000368233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Lai D., Qin C., Shu Q. Myeloid-derived suppressor cells in sepsis. Biomed. Res. Int. 2014;2014:598654. doi: 10.1155/2014/598654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Zhao B.G., Vasilakos J.P., Tross D., Smirnov D., Klinman D.M. Combination therapy targeting toll like receptors 7, 8 and 9 eliminates large established tumors. J. Immunother. Cancer. 2014;2:12. doi: 10.1186/2051-1426-2-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Li J.L., Zarbock A., Hidalgo A. Platelets as autonomous drones for hemostatic and immune surveillance. J. Exp. Med. 2017;214(8):2193–2204. doi: 10.1084/jem.20170879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Lievens D., von Hundelshausen P. Platelets in atherosclerosis. Thromb. Haemost. 2011;106(5):827–838. doi: 10.1160/TH11-08-0592. [DOI] [PubMed] [Google Scholar]
- 248.Schulz C., Massberg S. Platelets in atherosclerosis and thrombosis. Handb. Exp. Pharmacol. 2012;210:111–133. doi: 10.1007/978-3-642-29423-5_5. [DOI] [PubMed] [Google Scholar]
- 249.Li J.L., Zarbock A., Hidalgo A. Platelets as autonomous drones for hemostatic and immune surveillance. J. Exp. Med. 2017 doi: 10.1084/jem.20170879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Semple J.W., Freedman J. Platelets and innate immunity. Cell. Mol. Life Sci. 2010;67(4):499–511. doi: 10.1007/s00018-009-0205-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Mancuso M.E., Santagostino E. Platelets: much more than bricks in a breached wall. Br. J. Haematol. 2017;178(2):209–219. doi: 10.1111/bjh.14653. [DOI] [PubMed] [Google Scholar]
- 252.Koupenova M., Clancy L., Corkrey H.A., Freedman J.E. Circulating Platelets as Mediators of Immunity, Inflammation, and Thrombosis. Circ. Res. 2018;122(2):337–351. doi: 10.1161/CIRCRESAHA.117.310795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Morrell C.N., Aggrey A.A., Chapman L.M., Modjeski K.L. Emerging roles for platelets as immune and inflammatory cells. Blood. 2014;123(18):2759–2767. doi: 10.1182/blood-2013-11-462432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Jayachandran M., Brunn G.J., Karnicki K., Miller R.S., Owen W.G., Miller V.M. In vivo effects of lipopolysaccharide and TLR4 on platelet production and activity: implications for thrombotic risk. J. Appl. Physiol. 2007;102(1):429–433. doi: 10.1152/japplphysiol.01576.2005. [DOI] [PubMed] [Google Scholar]
- 255.Andonegui G., Kerfoot S.M., McNagny K., Ebbert K.V., Patel K.D., Kubes P. Platelets express functional Toll-like receptor-4. Blood. 2005;106(7):2417–2423. doi: 10.1182/blood-2005-03-0916. [DOI] [PubMed] [Google Scholar]
- 256.Aslam R., Speck E.R., Kim M., Crow A.R., Bang K.W., Nestel F.P., Ni H., Lazarus A.H., Freedman J., Semple J.W. Platelet Toll-like receptor expression modulates lipopolysaccharide-induced thrombocytopenia and tumor necrosis factor-alpha production in vivo. Blood. 2006;107(2):637–641. doi: 10.1182/blood-2005-06-2202. [DOI] [PubMed] [Google Scholar]
- 257.Shiraki R., Inoue N., Kawasaki S., Takei A., Kadotani M., Ohnishi Y., Ejiri J., Kobayashi S., Hirata K., Kawashima S., Yokoyama M. Expression of Toll-like receptors on human platelets. Thromb. Res. 2004;113(6):379–385. doi: 10.1016/j.thromres.2004.03.023. [DOI] [PubMed] [Google Scholar]
- 258.Cognasse F., Hamzeh H., Chavarin P., Acquart S., Genin C., Garraud O. Evidence of Toll-like receptor molecules on human platelets. Immunol. Cell Biol. 2005;83(2):196–198. doi: 10.1111/j.1440-1711.2005.01314.x. [DOI] [PubMed] [Google Scholar]
- 259.Clark S.R., Ma A.C., Tavener S.A., McDonald B., Goodarzi Z., Kelly M.M., Patel K.D., Chakrabarti S., McAvoy E., Sinclair G.D., Keys E.M., Allen-Vercoe E., Devinney R., Doig C.J., Green F.H., Kubes P. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 2007;13(4):463–469. doi: 10.1038/nm1565. [DOI] [PubMed] [Google Scholar]
- 260.Patrignani P., Di Febbo C., Tacconelli S., Moretta V., Baccante G., Sciulli M.G., Ricciotti E., Capone M.L., Antonucci I., Guglielmi M.D., Stuppia L., Porreca E. Reduced thromboxane biosynthesis in carriers of toll-like receptor 4 polymorphisms in vivo. Blood. 2006;107(9):3572–3574. doi: 10.1182/blood-2005-12-4811. [DOI] [PubMed] [Google Scholar]
- 261.Fabrice C., John W.S., Olivier G. Platelets as Potential Immunomodulators: Is There a Role for Platelet Toll-Like Receptors? Curr. Immunol. Rev. 2007;3(2):109–115. [Google Scholar]
- 262.Rondina M.T., Schwertz H., Harris E.S., Kraemer B.F., Campbell R.A., Mackman N., Grissom C.K., Weyrich A.S., Zimmerman G.A. The septic milieu triggers expression of spliced tissue factor mRNA in human platelets. J. Thromb. Haemost. 2011;9(4):748–758. doi: 10.1111/j.1538-7836.2011.04208.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Cognasse F., Nguyen K.A., Damien P., McNicol A., Pozzetto B., Hamzeh-Cognasse H., Garraud O. The Inflammatory Role of Platelets via Their TLRs and Siglec Receptors. Front. Immunol. 2015;6:83. doi: 10.3389/fimmu.2015.00083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Hamzeh-Cognasse H., Damien P., Chabert A., Pozzetto B., Cognasse F., Garraud O. Platelets and infections - complex interactions with bacteria. Front. Immunol. 2015;6:82. doi: 10.3389/fimmu.2015.00082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Cognasse F., Hamzeh-Cognasse H., Garraud O. Platelets "Toll-like receptor" engagement stimulates the release of immunomodulating molecules. Transfus. Clin. Biol. 2008;15(4):139–147. doi: 10.1016/j.tracli.2008.07.010. [DOI] [PubMed] [Google Scholar]
- 266.Cognasse F., Hamzeh-Cognasse H., Lafarge S., Delezay O., Pozzetto B., McNicol A., Garraud O. Toll-like receptor 4 ligand can differentially modulate the release of cytokines by human platelets. Br. J. Haematol. 2008;141(1):84–91. doi: 10.1111/j.1365-2141.2008.06999.x. [DOI] [PubMed] [Google Scholar]
- 267.Cognasse F., Osselaer J.C., Payrat J.M., Chavarin P., Corash L., Garraud O. Release of immune modulation factors from platelet concentrates during storage after photochemical pathogen inactivation treatment. Transfusion. 2008;48(5):809–813. doi: 10.1111/j.1537-2995.2008.01655.x. [DOI] [PubMed] [Google Scholar]
- 268.de Stoppelaar S.F., Claushuis T.A., Schaap M.C., Hou B., van der Poll T., Nieuwland R., van 't Veer C. Toll-Like Receptor Signalling Is Not Involved in Platelet Response to Streptococcus pneumoniae In Vitro or In Vivo. PLoS One. 2016;11(6) doi: 10.1371/journal.pone.0156977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Senturk T. Platelet function in inflammatory diseases: insights from clinical studies. Inflamm. Allergy Drug Targets. 2010;9(5):355–363. doi: 10.2174/187152810793938026. [DOI] [PubMed] [Google Scholar]
- 270.Ferrer-Acosta Y., González M., Fernández M., Valance W.A. Emerging Roles for Platelets in Inflammation and Disease. J. Infect. Dis. Ther. 2014;2(4):149. doi: 10.4172/2332-0877.1000149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Thomas M.R., Storey R.F. The role of platelets in inflammation. Thromb. Haemost. 2015;114(3):449–458. doi: 10.1160/TH14-12-1067. [DOI] [PubMed] [Google Scholar]
- 272.Carestia A., Kaufman T., Schattner M. Platelets: New Bricks in the Building of Neutrophil Extracellular Traps. Front. Immunol. 2016;7:271. doi: 10.3389/fimmu.2016.00271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273.Rondina M.T., Garraud O. Emerging evidence for platelets as immune and inflammatory effector cells. Front. Immunol. 2014;5:653. doi: 10.3389/fimmu.2014.00653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Blair P., Rex S., Vitseva O., Beaulieu L., Tanriverdi K., Chakrabarti S., Hayashi C., Genco C.A., Iafrati M., Freedman J.E. Stimulation of Toll-like receptor 2 in human platelets induces a thromboinflammatory response through activation of phosphoinositide 3-kinase. Circ. Res. 2009;104(3):346–354. doi: 10.1161/CIRCRESAHA.108.185785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275.Biswas S., Zimman A., Gao D., Byzova T.V., Podrez E.A. TLR2 Plays a Key Role in Platelet Hyperreactivity and Accelerated Thrombosis Associated With Hyperlipidemia. Circ. Res. 2017;121(8):951–962. doi: 10.1161/CIRCRESAHA.117.311069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276.Panigrahi S., Ma Y., Hong L., Gao D., West X.Z., Salomon R.G., Byzova T.V., Podrez E.A. Engagement of Platelet Toll-Like Receptor 9 by Novel Endogenous Ligands Promotes Platelet Hyper-Reactivity and Thrombosis. Circ. Res. 2013;112(1):103–112. doi: 10.1161/CIRCRESAHA.112.274241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277.Kumar V., Sharma A. Mast cells: emerging sentinel innate immune cells with diverse role in immunity. Mol. Immunol. 2010;48(1–3):14–25. doi: 10.1016/j.molimm.2010.07.009. [DOI] [PubMed] [Google Scholar]
- 278.Krystel-Whittemore M., Dileepan K.N., Wood J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2015;6:620. doi: 10.3389/fimmu.2015.00620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 279.Frossi B., Mion F., Tripodo C., Colombo M.P., Pucillo C.E. Rheostatic Functions of Mast Cells in the Control of Innate and Adaptive Immune Responses. Trends Immunol. 2017;38(9):648–656. doi: 10.1016/j.it.2017.04.001. [DOI] [PubMed] [Google Scholar]
- 280.Mrabet-Dahbi S., Metz M., Dudeck A., Zuberbier T., Maurer M. Murine mast cells secrete a unique profile of cytokines and prostaglandins in response to distinct TLR2 ligands. Exp. Dermatol. 2009;18(5):437–444. doi: 10.1111/j.1600-0625.2009.00878.x. [DOI] [PubMed] [Google Scholar]
- 281.Li G., Domenico J., Jia Y., Lucas J.J., Gelfand E.W. NF-kappaB-dependent induction of cathelicidin-related antimicrobial peptide in murine mast cells by lipopolysaccharide. Int. Arch. Allergy Immunol. 2009;150(2):122–132. doi: 10.1159/000218115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Matsushima H., Yamada N., Matsue H., Shimada S. TLR3-, TLR7-, and TLR9-mediated production of proinflammatory cytokines and chemokines from murine connective tissue type skin-derived mast cells but not from bone marrow-derived mast cells. J. Immunol. 2004;173(1):531–541. doi: 10.4049/jimmunol.173.1.531. [DOI] [PubMed] [Google Scholar]
- 283.McCurdy J.D., Lin T.J., Marshall J.S. Toll-like receptor 4-mediated activation of murine mast cells. J. Leukoc. Biol. 2001;70(6):977–984. [PubMed] [Google Scholar]
- 284.Varadaradjalou S., Feger F., Thieblemont N., Hamouda N.B., Pleau J.M., Dy M., Arock M. Toll-like receptor 2 (TLR2) and TLR4 differentially activate human mast cells. Eur. J. Immunol. 2003;33(4):899–906. doi: 10.1002/eji.200323830. [DOI] [PubMed] [Google Scholar]
- 285.Keck S., Muller I., Fejer G., Savic I., Tchaptchet S., Nielsen P.J., Galanos C., Huber M., Freudenberg M.A. Absence of TRIF signaling in lipopolysaccharide-stimulated murine mast cells. J. Immunol. 2011;186(9):5478–5488. doi: 10.4049/jimmunol.1000458. [DOI] [PubMed] [Google Scholar]
- 286.Kagan J.C., Su T., Horng T., Chow A., Akira S., Medzhitov R. TRAM couples endocytosis of Toll-like receptor 4 to the induction of interferon-beta. Nat. Immunol. 2008;9(4):361–368. doi: 10.1038/ni1569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287.Kagan J.C. Signaling organelles of the innate immune system. Cell. 2012;151(6):1168–1178. doi: 10.1016/j.cell.2012.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288.Kagan J.C. Defining the subcellular sites of innate immune signal transduction. Trends Immunol. 2012;33(9):442–448. doi: 10.1016/j.it.2012.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Dietrich N., Rohde M., Geffers R., Kroger A., Hauser H., Weiss S., Gekara N.O. Mast cells elicit proinflammatory but not type I interferon responses upon activation of TLRs by bacteria. Proc. Natl. Acad. Sci. U. S. A. 2010;107(19):8748–8753. doi: 10.1073/pnas.0912551107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290.Lee C.C., Avalos A.M., Ploegh H.L. Accessory molecules for Toll-like receptors and their function. Nat. Rev. Immunol. 2012;12(3):168–179. doi: 10.1038/nri3151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Zanoni I., Ostuni R., Marek L.R., Barresi S., Barbalat R., Barton G.M., Granucci F., Kagan J.C. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Cell. 2011;147(4):868–880. doi: 10.1016/j.cell.2011.09.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Tamassia N., Le Moigne V., Calzetti F., Donini M., Gasperini S., Ear T., Cloutier A., Martinez F.O., Fabbri M., Locati M., Mantovani A., McDonald P.P., Cassatella M.A. The MyD88-independent pathway is not mobilized in human neutrophils stimulated via TLR4. J. Immunol. 2007;178(11):7344–7356. doi: 10.4049/jimmunol.178.11.7344. [DOI] [PubMed] [Google Scholar]
- 293.Jersmann H.P.A. Time to abandon dogma: CD14 is expressed by non-myeloid lineage cells. Immunol. Cell Biol. 2005;83(5):462–467. doi: 10.1111/j.1440-1711.2005.01370.x. [DOI] [PubMed] [Google Scholar]
- 294.Kulka M., Alexopoulou L., Flavell R.A., Metcalfe D.D. Activation of mast cells by double-stranded RNA: evidence for activation through Toll-like receptor 3. J. Allergy Clin. Immunol. 2004;114(1):174–182. doi: 10.1016/j.jaci.2004.03.049. [DOI] [PubMed] [Google Scholar]
- 295.Kulka M., Metcalfe D.D. TLR3 activation inhibits human mast cell attachment to fibronectin and vitronectin. Mol. Immunol. 2006;43(10):1579–1586. doi: 10.1016/j.molimm.2005.09.019. [DOI] [PubMed] [Google Scholar]
- 296.Yoshioka M., Fukuishi N., Iriguchi S., Ohsaki K., Yamanobe H., Inukai A., Kurihara D., Imajo N., Yasui Y., Matsui N., Tsujita T., Ishii A., Seya T., Takahama M., Akagi M. Lipoteichoic acid downregulates FcepsilonRI expression on human mast cells through Toll-like receptor 2. J. Allergy Clin. Immunol. 2007;120(2):452–461. doi: 10.1016/j.jaci.2007.03.027. [DOI] [PubMed] [Google Scholar]
- 297.Ross R. The role of T lymphocytes in inflammation. Proc. Natl. Acad. Sci. U. S. A. 1994;91(8):2879. doi: 10.1073/pnas.91.8.2879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298.Li B., Zheng S.G. How regulatory T cells sense and adapt to inflammation. Cell. Mol. Immunol. 2015;12:519. doi: 10.1038/cmi.2015.65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299.Skapenko A., Leipe J., Lipsky P.E., Schulze-Koops H. The role of the T cell in autoimmune inflammation. Arthritis Res. Ther. 2005;7(2):S4. doi: 10.1186/ar1703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300.van der Veeken J., Gonzalez A.J., Cho H., Arvey A., Hemmers S., Leslie C.S., Rudensky A.Y. Memory of Inflammation in Regulatory T Cells. Cell. 2016;166(4):977–990. doi: 10.1016/j.cell.2016.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Hotchkiss R.S., Osmon S.B., Chang K.C., Wagner T.H., Coopersmith C.M., Karl I.E. Accelerated lymphocyte death in sepsis occurs by both the death receptor and mitochondrial pathways. J. Immunol. 2005;174(8):5110–5118. doi: 10.4049/jimmunol.174.8.5110. [DOI] [PubMed] [Google Scholar]
- 302.Le Tulzo Y., Pangault C., Gacouin A., Guilloux V., Tribut O., Amiot L., Tattevin P., Thomas R., Fauchet R., Drenou B. Early circulating lymphocyte apoptosis in human septic shock is associated with poor outcome. Shock. 2002;18(6):487–494. doi: 10.1097/00024382-200212000-00001. [DOI] [PubMed] [Google Scholar]
- 303.Inoue S., Suzuki-Utsunomiya K., Okada Y., Taira T., Iida Y., Miura N., Tsuji T., Yamagiwa T., Morita S., Chiba T., Sato T., Inokuchi S. Reduction of immunocompetent T cells followed by prolonged lymphopenia in severe sepsis in the elderly. Crit. Care Med. 2013;41(3):810–819. doi: 10.1097/CCM.0b013e318274645f. [DOI] [PubMed] [Google Scholar]
- 304.Cao C., Ma T., Chai Y.-f., Shou S.-t. The role of regulatory T cells in immune dysfunction during sepsis. World J Emerg Med. 2015;6(1):5–9. doi: 10.5847/wjem.j.1920-8642.2015.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Markwart R., Condotta S.A., Requardt R.P., Borken F., Schubert K., Weigel C., Bauer M., Griffith T.S., Förster M., Brunkhorst F.M., Badovinac V.P., Rubio I. Immunosuppression after Sepsis: Systemic Inflammation and Sepsis Induce a Loss of Naïve T-Cells but No Enduring Cell-Autonomous Defects in T-Cell Function. PLoS ONE. 2014;9(12) doi: 10.1371/journal.pone.0115094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 306.Condotta S.A., Cabrera-Perez J., Badovinac V.P., Griffith T.S. T-cell mediated immunity and the role of TRAIL in sepsis-induced immunosuppression. Crit. Rev. Immunol. 2013;33(1):23–40. doi: 10.1615/critrevimmunol.2013006721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Cabrera-Perez J., Condotta S.A., Badovinac V.P., Griffith T.S. Impact of sepsis on CD4 T cell immunity. J. Leukoc. Biol. 2014;96(5):767–777. doi: 10.1189/jlb.5MR0114-067R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308.Wang S.D., Huang K.J., Lin Y.S., Lei H.Y. Sepsis-induced apoptosis of the thymocytes in mice. J. Immunol. 1994;152(10):5014–5021. [PubMed] [Google Scholar]
- 309.Boomer J.S., To K., Chang K.C., Takasu O., Osborne D.F., Walton A.H., Bricker T.L., Jarman S.D., II, Kreisel D., Krupnick A.S., Srivastava A., Swanson P.E., Green J.M., Hotchkiss R.S. Immunosuppression in patients who die of sepsis and multiple organ failure. JAMA. 2011;306(23):2594–2605. doi: 10.1001/jama.2011.1829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Kasten K.R., Tschop J., Adediran S.G., Hildeman D.A., Caldwell C.C. T cells are potent early mediators of the host response to sepsis. Shock. 2010;34(4):327–336. doi: 10.1097/SHK.0b013e3181e14c2e. [DOI] [PubMed] [Google Scholar]
- 311.Venet F., Pachot A., Debard A.L., Bohe J., Bienvenu J., Lepape A., Monneret G. Increased percentage of CD4+CD25+ regulatory T cells during septic shock is due to the decrease of CD4+CD25- lymphocytes. Crit. Care Med. 2004;32(11):2329–2331. doi: 10.1097/01.ccm.0000145999.42971.4b. [DOI] [PubMed] [Google Scholar]
- 312.Taylor A.L., Llewelyn M.J. Superantigen-induced proliferation of human CD4+CD25- T cells is followed by a switch to a functional regulatory phenotype. J. Immunol. 2010;185(11):6591–6598. doi: 10.4049/jimmunol.1002416. [DOI] [PubMed] [Google Scholar]
- 313.Wisnoski N., Chung C.S., Chen Y., Huang X., Ayala A. The contribution of CD4+ CD25+ T-regulatory-cells to immune suppression in sepsis. Shock. 2007;27(3):251–257. doi: 10.1097/01.shk.0000239780.33398.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314.Venet F., Pachot A., Debard A.L., Bohe J., Bienvenu J., Lepape A., Powell W.S., Monneret G. Human CD4+CD25+ regulatory T lymphocytes inhibit lipopolysaccharide-induced monocyte survival through a Fas/Fas ligand-dependent mechanism. J. Immunol. 2006;177(9):6540–6547. doi: 10.4049/jimmunol.177.9.6540. [DOI] [PubMed] [Google Scholar]
- 315.Heuer J.G., Zhang T., Zhao J., Ding C., Cramer M., Justen K.L., Vonderfecht S.L., Na S. Adoptive transfer of in vitro-stimulated CD4+CD25+ regulatory T cells increases bacterial clearance and improves survival in polymicrobial sepsis. J. Immunol. 2005;174(11):7141–7146. doi: 10.4049/jimmunol.174.11.7141. [DOI] [PubMed] [Google Scholar]
- 316.Chen X., Baumel M., Mannel D.N., Howard O.M., Oppenheim J.J. Interaction of TNF with TNF receptor type 2 promotes expansion and function of mouse CD4+CD25+ T regulatory cells. J. Immunol. 2007;179(1):154–161. doi: 10.4049/jimmunol.179.1.154. [DOI] [PubMed] [Google Scholar]
- 317.Hornung V., Rothenfusser S., Britsch S., Krug A., Jahrsdorfer B., Giese T., Endres S., Hartmann G. Quantitative expression of toll-like receptor 1–10 mRNA in cellular subsets of human peripheral blood mononuclear cells and sensitivity to CpG oligodeoxynucleotides. J. Immunol. 2002;168(9):4531–4537. doi: 10.4049/jimmunol.168.9.4531. [DOI] [PubMed] [Google Scholar]
- 318.Mansson A., Adner M., Cardell L.O. Toll-like receptors in cellular subsets of human tonsil T cells: altered expression during recurrent tonsillitis. Respir. Res. 2006;7:36. doi: 10.1186/1465-9921-7-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319.Zarember K.A., Godowski P.J. Tissue expression of human Toll-like receptors and differential regulation of Toll-like receptor mRNAs in leukocytes in response to microbes, their products, and cytokines. J. Immunol. 2002;168(2):554–561. doi: 10.4049/jimmunol.168.2.554. [DOI] [PubMed] [Google Scholar]
- 320.Peng G., Guo Z., Kiniwa Y., Voo K.S., Peng W., Fu T., Wang D.Y., Li Y., Wang H.Y., Wang R.F. Toll-like receptor 8-mediated reversal of CD4+ regulatory T cell function. Science. 2005;309(5739):1380–1384. doi: 10.1126/science.1113401. (New York, N.Y.) [DOI] [PubMed] [Google Scholar]
- 321.Zanin-Zhorov A., Cahalon L., Tal G., Margalit R., Lider O., Cohen I.R. Heat shock protein 60 enhances CD4+ CD25+ regulatory T cell function via innate TLR2 signaling. J. Clin. Invest. 2006;116(7):2022–2032. doi: 10.1172/JCI28423. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 322.Crellin N.K., Garcia R.V., Hadisfar O., Allan S.E., Steiner T.S., Levings M.K. Human CD4+ T cells express TLR5 and its ligand flagellin enhances the suppressive capacity and expression of FOXP3 in CD4+CD25+ T regulatory cells. J. Immunol. 2005;175(12):8051–8059. doi: 10.4049/jimmunol.175.12.8051. [DOI] [PubMed] [Google Scholar]
- 323.Caramalho I., Lopes-Carvalho T., Ostler D., Zelenay S., Haury M., Demengeot J. Regulatory T Cells Selectively Express Toll-like Receptors and Are Activated by Lipopolysaccharide. J. Exp. Med. 2003;197(4):403–411. doi: 10.1084/jem.20021633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324.Tomita T., Kanai T., Fujii T., Nemoto Y., Okamoto R., Tsuchiya K., Totsuka T., Sakamoto N., Akira S., Watanabe M. MyD88-dependent pathway in T cells directly modulates the expansion of colitogenic CD4+ T cells in chronic colitis. J. Immunol. 2008;180(8):5291–5299. doi: 10.4049/jimmunol.180.8.5291. [DOI] [PubMed] [Google Scholar]
- 325.Sobek V., Birkner N., Falk I., Wurch A., Kirschning C.J., Wagner H., Wallich R., Lamers M.C., Simon M.M. Direct Toll-like receptor 2 mediated co-stimulation of T cells in the mouse system as a basis for chronic inflammatory joint disease. Arthritis Res. Ther. 2004;6(5):R433–46. doi: 10.1186/ar1212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 326.Cottalorda A., Verschelde C., Marcais A., Tomkowiak M., Musette P., Uematsu S., Akira S., Marvel J., Bonnefoy-Berard N. TLR2 engagement on CD8 T cells lowers the threshold for optimal antigen-induced T cell activation. Eur. J. Immunol. 2006;36(7):1684–1693. doi: 10.1002/eji.200636181. [DOI] [PubMed] [Google Scholar]
- 327.Gelman A.E., Zhang J., Choi Y., Turka L.A. Toll-like receptor ligands directly promote activated CD4+ T cell survival. J. Immunol. 2004;172(10):6065–6073. doi: 10.4049/jimmunol.172.10.6065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Fukata M., Breglio K., Chen A., Vamadevan A.S., Goo T., Hsu D., Conduah D., Xu R., Abreu M.T. The myeloid differentiation factor 88 (MyD88) is required for CD4+ T cell effector function in a murine model of inflammatory bowel disease. J. Immunol. 2008;180(3):1886–1894. doi: 10.4049/jimmunol.180.3.1886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 329.MacLeod H., Wetzler L.M. T cell activation by TLRs: a role for TLRs in the adaptive immune response. Sci. STKE. 2007;2007(402) doi: 10.1126/stke.4022007pe48. [DOI] [PubMed] [Google Scholar]
- 330.Karim A.F., Reba S.M., Li Q., Boom W.H., Rojas R.E. Toll like Receptor 2 engagement on CD4+ T cells promotes TH9 differentiation and function. Eur. J. Immunol. 2017;47(9):1513–1524. doi: 10.1002/eji.201646846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 331.Rahman A.H., Taylor D.K., Turka L.A. The contribution of direct TLR signaling to T cell responses. Immunol. Res. 2009;45(1):25–36. doi: 10.1007/s12026-009-8113-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 332.Kabelitz D. Expression and function of Toll-like receptors in T lymphocytes. Curr. Opin. Immunol. 2007;19(1):39–45. doi: 10.1016/j.coi.2006.11.007. [DOI] [PubMed] [Google Scholar]
- 333.Kelly-Scumpia K.M., Scumpia P.O., Weinstein J.S., Delano M.J., Cuenca A.G., Nacionales D.C., Wynn J.L., Lee P.Y., Kumagai Y., Efron P.A., Akira S., Wasserfall C., Atkinson M.A., Moldawer L.L. B cells enhance early innate immune responses during bacterial sepsis. J. Exp. Med. 2011;208(8):1673–1682. doi: 10.1084/jem.20101715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 334.Rawlings D.J., Schwartz M.A., Jackson S.W., Meyer-Bahlburg A. Integration of B cell responses through Toll-like receptors and antigen receptors. Nat. Rev. Immunol. 2012;12(4):282–294. doi: 10.1038/nri3190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 335.Martin F., Oliver A.M., Kearney J.F. Marginal zone and B1 B cells unite in the early response against T-independent blood-borne particulate antigens. Immunity. 2001;14(5):617–629. doi: 10.1016/s1074-7613(01)00129-7. [DOI] [PubMed] [Google Scholar]
- 336.McHeyzer-Williams M.G. B cells as effectors. Curr. Opin. Immunol. 2003;15(3):354–361. doi: 10.1016/s0952-7915(03)00046-3. [DOI] [PubMed] [Google Scholar]
- 337.Monserrat J., de Pablo R., Diaz-Martín D., Rodríguez-Zapata M., de la Hera A., Prieto A., Alvarez-Mon M. Early alterations of B cells in patients with septic shock. Crit. Care. 2013;17(3):R105. doi: 10.1186/cc12750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 338.Suzuki K., Inoue S., Kametani Y., Komori Y., Chiba S., Sato T., Inokuchi S., Ogura S. Reduced Immunocompetent B Cells and Increased Secondary Infection in Elderly Patients With Severe Sepsis. Shock. 2016;46(3):270–278. doi: 10.1097/SHK.0000000000000619. [DOI] [PubMed] [Google Scholar]
- 339.Hua Z., Hou B. TLR signaling in B-cell development and activation. Cell. Mol. Immunol. 2013;10(2):103–106. doi: 10.1038/cmi.2012.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 340.Bekeredjian-Ding I., Jego G. Toll-like receptors – sentries in the B-cell response. Immunology. 2009;128(3):311–323. doi: 10.1111/j.1365-2567.2009.03173.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 341.Bekeredjian-Ding I., Inamura S., Giese T., Moll H., Endres S., Sing A., Zahringer U., Hartmann G. Staphylococcus aureus protein A triggers T cell-independent B cell proliferation by sensitizing B cells for TLR2 ligands. J. Immunol. 2007;178(5):2803–2812. doi: 10.4049/jimmunol.178.5.2803. [DOI] [PubMed] [Google Scholar]
- 342.Ruprecht C.R., Lanzavecchia A. Toll-like receptor stimulation as a third signal required for activation of human naive B cells. Eur. J. Immunol. 2006;36(4):810–816. doi: 10.1002/eji.200535744. [DOI] [PubMed] [Google Scholar]
- 343.Hasan U., Chaffois C., Gaillard C., Saulnier V., Merck E., Tancredi S., Guiet C., Briere F., Vlach J., Lebecque S., Trinchieri G., Bates E.E. Human TLR10 is a functional receptor, expressed by B cells and plasmacytoid dendritic cells, which activates gene transcription through MyD88. J. Immunol. 2005;174(5):2942–2950. doi: 10.4049/jimmunol.174.5.2942. [DOI] [PubMed] [Google Scholar]
- 344.Won W.J., Bachmann M.F., Kearney J.F. CD36 is differentially expressed on B cell subsets during development and in responses to antigen. J. Immunol. 2008;180(1):230–237. doi: 10.4049/jimmunol.180.1.230. [DOI] [PubMed] [Google Scholar]
- 345.Buchta C.M., Bishop G.A. Toll-like receptors and B cells: functions and mechanisms. Immunol. Res. 2014;59(1–3):12–22. doi: 10.1007/s12026-014-8523-2. [DOI] [PubMed] [Google Scholar]
- 346.Tabeta K., Hoebe K., Janssen E.M., Du X., Georgel P., Crozat K., Mudd S., Mann N., Sovath S., Goode J., Shamel L., Herskovits A.A., Portnoy D.A., Cooke M., Tarantino L.M., Wiltshire T., Steinberg B.E., Grinstein S., Beutler B. The Unc93b1 mutation 3d disrupts exogenous antigen presentation and signaling via Toll-like receptors 3, 7 and 9. Nat. Immunol. 2006;7(2):156–164. doi: 10.1038/ni1297. [DOI] [PubMed] [Google Scholar]
- 347.Akkaya M., Akkaya B., Kim A.S., Miozzo P., Sohn H., Pena M., Roesler A.S., Theall B.P., Henke T., Kabat J., Lu J., Dorward D.W., Dahlstrom E., Skinner J., Miller L.H., Pierce S.K. Toll-like receptor 9 antagonizes antibody affinity maturation. Nat. Immunol. 2018;19(3):255–266. doi: 10.1038/s41590-018-0052-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 348.Isnardi I., Ng Y.S., Srdanovic I., Motaghedi R., Rudchenko S., von Bernuth H., Zhang S.Y., Puel A., Jouanguy E., Picard C., Garty B.Z., Camcioglu Y., Doffinger R., Kumararatne D., Davies G., Gallin J.I., Haraguchi S., Day N.K., Casanova J.L., Meffre E. IRAK-4- and MyD88-dependent pathways are essential for the removal of developing autoreactive B cells in humans. Immunity. 2008;29(5):746–757. doi: 10.1016/j.immuni.2008.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 349.Buchta C.M., Bishop G.A. TRAF5 negatively regulates TLR signaling in B lymphocytes. J. Immunol. 2014;192(1):145–150. doi: 10.4049/jimmunol.1301901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 350.Agrawal S., Gupta S. TLR1/2, TLR7, and TLR9 signals directly activate human peripheral blood naive and memory B cell subsets to produce cytokines, chemokines, and hematopoietic growth factors. J. Clin. Immunol. 2011;31(1):89–98. doi: 10.1007/s10875-010-9456-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 351.Jagannathan M., Hasturk H., Liang Y., Shin H., Hetzel J.T., Kantarci A., Rubin D., McDonnell M.E., Van Dyke T.E., Ganley-Leal L.M., Nikolajczyk B.S. TLR cross-talk specifically regulates cytokine production by B cells from chronic inflammatory disease patients. J. Immunol. 2009;183(11):7461–7470. doi: 10.4049/jimmunol.0901517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 352.Booth J., Wilson H., Jimbo S., Mutwiri G. Modulation of B cell responses by Toll-like receptors. Cell Tissue Res. 2011;343(1):131–140. doi: 10.1007/s00441-010-1031-3. [DOI] [PubMed] [Google Scholar]
- 353.Meyer-Bahlburg A., Khim S., Rawlings D.J. B cell intrinsic TLR signals amplify but are not required for humoral immunity. J. Exp. Med. 2007;204(13):3095–3101. doi: 10.1084/jem.20071250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 354.von Bernuth H., Picard C., Jin Z., Pankla R., Xiao H., Ku C.L., Chrabieh M., Mustapha I.B., Ghandil P., Camcioglu Y., Vasconcelos J., Sirvent N., Guedes M., Vitor A.B., Herrero-Mata M.J., Arostegui J.I., Rodrigo C., Alsina L., Ruiz-Ortiz E., Juan M., Fortuny C., Yague J., Anton J., Pascal M., Chang H.H., Janniere L., Rose Y., Garty B.Z., Chapel H., Issekutz A., Marodi L., Rodriguez-Gallego C., Banchereau J., Abel L., Li X., Chaussabel D., Puel A., Casanova J.L. Pyogenic bacterial infections in humans with MyD88 deficiency. Science. 2008;321(5889):691–696. doi: 10.1126/science.1158298. (New York, N.Y.) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 355.Picard C., Puel A., Bonnet M., Ku C.L., Bustamante J., Yang K., Soudais C., Dupuis S., Feinberg J., Fieschi C., Elbim C., Hitchcock R., Lammas D., Davies G., Al-Ghonaium A., Al-Rayes H., Al-Jumaah S., Al-Hajjar S., Al-Mohsen I.Z., Frayha H.H., Rucker R., Hawn T.R., Aderem A., Tufenkeji H., Haraguchi S., Day N.K., Good R.A., Gougerot-Pocidalo M.A., Ozinsky A., Casanova J.L. Pyogenic bacterial infections in humans with IRAK-4 deficiency. Science. 2003;299(5615):2076–2079. doi: 10.1126/science.1081902. (New York, N.Y.) [DOI] [PubMed] [Google Scholar]
- 356.Picard C., von Bernuth H., Ghandil P., Chrabieh M., Levy O., Arkwright P.D., McDonald D., Geha R.S., Takada H., Krause J.C., Creech C.B., Ku C.L., Ehl S., Marodi L., Al-Muhsen S., Al-Hajjar S., Al-Ghonaium A., Day-Good N.K., Holland S.M., Gallin J.I., Chapel H., Speert D.P., Rodriguez-Gallego C., Colino E., Garty B.Z., Roifman C., Hara T., Yoshikawa H., Nonoyama S., Domachowske J., Issekutz A.C., Tang M., Smart J., Zitnik S.E., Hoarau C., Kumararatne D.S., Thrasher A.J., Davies E.G., Bethune C., Sirvent N., de Ricaud D., Camcioglu Y., Vasconcelos J., Guedes M., Vitor A.B., Rodrigo C., Almazan F., Mendez M., Arostegui J.I., Alsina L., Fortuny C., Reichenbach J., Verbsky J.W., Bossuyt X., Doffinger R., Abel L., Puel A., Casanova J.L. Clinical features and outcome of patients with IRAK-4 and MyD88 deficiency. Medicine. 2010;89(6):403–425. doi: 10.1097/MD.0b013e3181fd8ec3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 357.Browne E.P. Regulation of B-cell responses by Toll-like receptors. Immunology. 2012;136(4):370–379. doi: 10.1111/j.1365-2567.2012.03587.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 358.Nikolajczyk B.S. B cells as under-appreciated mediators of non-autoimmune inflammatory disease. Cytokine. 2010;50(3):234–242. doi: 10.1016/j.cyto.2010.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 359.Miyagaki T., Fujimoto M., Sato S. Regulatory B cells in human inflammatory and autoimmune diseases: from mouse models to clinical research. Int. Immunol. 2015;27(10):495–504. doi: 10.1093/intimm/dxv026. [DOI] [PubMed] [Google Scholar]
- 360.Ray A., Dittel B.N. Mechanisms of Regulatory B cell Function in Autoimmune and Inflammatory Diseases beyond IL-10. J. Clin. Med. 2017;6(1):12. doi: 10.3390/jcm6010012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 361.Patel A.R., Ritzel R., McCullough L.D., Liu F. Microglia and ischemic stroke: a double-edged sword. Int. J. Physiol. Pathophysiol. Pharmacol. 2013;5(2):73–90. [PMC free article] [PubMed] [Google Scholar]
- 362.Guruswamy R., ElAli A. Complex Roles of Microglial Cells in Ischemic Stroke Pathobiology: New Insights and Future Directions. Int. J. Mol. Sci. 2017;18(3):496. doi: 10.3390/ijms18030496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 363.Mariani M.M., Kielian T. Microglia in Infectious Diseases of the Central Nervous System. J. NeuroImmune Pharmacol. 2009;4(4):448–461. doi: 10.1007/s11481-009-9170-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 364.Rock R.B., Gekker G., Hu S., Sheng W.S., Cheeran M., Lokensgard J.R., Peterson P.K. Role of Microglia in Central Nervous System Infections. Clin. Microbiol. Rev. 2004;17(4):942–964. doi: 10.1128/CMR.17.4.942-964.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 365.Perry V.H., Holmes C. Microglial priming in neurodegenerative disease. Nat. Rev. Neurol. 2014;10:217. doi: 10.1038/nrneurol.2014.38. [DOI] [PubMed] [Google Scholar]
- 366.Bsibsi M., Ravid R., Gveric D., van Noort J.M. Broad expression of Toll-like receptors in the human central nervous system. J. Neuropathol. Exp. Neurol. 2002;61(11):1013–1021. doi: 10.1093/jnen/61.11.1013. [DOI] [PubMed] [Google Scholar]
- 367.Olson J.K., Miller S.D. Microglia initiate central nervous system innate and adaptive immune responses through multiple TLRs. J. Immunol. 2004;173(6):3916–3924. doi: 10.4049/jimmunol.173.6.3916. [DOI] [PubMed] [Google Scholar]
- 368.Farina C., Aloisi F., Meinl E. Astrocytes are active players in cerebral innate immunity. Trends Immunol. 2007;28(3):138–145. doi: 10.1016/j.it.2007.01.005. [DOI] [PubMed] [Google Scholar]
- 369.Farina C., Krumbholz M., Giese T., Hartmann G., Aloisi F., Meinl E. Preferential expression and function of Toll-like receptor 3 in human astrocytes. J. Neuroimmunol. 2005;159(1-2):12–19. doi: 10.1016/j.jneuroim.2004.09.009. [DOI] [PubMed] [Google Scholar]
- 370.Laflamme N., Rivest S. Toll-like receptor 4: the missing link of the cerebral innate immune response triggered by circulating gram-negative bacterial cell wall components. FASEB J. 2001;15(1):155–163. doi: 10.1096/fj.00-0339com. [DOI] [PubMed] [Google Scholar]
- 371.Lehnardt S., Lachance C., Patrizi S., Lefebvre S., Follett P.L., Jensen F.E., Rosenberg P.A., Volpe J.J., Vartanian T. The toll-like receptor TLR4 is necessary for lipopolysaccharide-induced oligodendrocyte injury in the CNS. J. Neurosci. 2002;22(7):2478–2486. doi: 10.1523/JNEUROSCI.22-07-02478.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 372.Lehnardt S., Massillon L., Follett P., Jensen F.E., Ratan R., Rosenberg P.A., Volpe J.J., Vartanian T. Activation of innate immunity in the CNS triggers neurodegeneration through a Toll-like receptor 4-dependent pathway. Proc. Natl. Acad. Sci. U. S. A. 2003;100(14):8514–8519. doi: 10.1073/pnas.1432609100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 373.Bowman C.C., Rasley A., Tranguch S.L., Marriott I. Cultured astrocytes express toll-like receptors for bacterial products. Glia. 2003;43(3):281–291. doi: 10.1002/glia.10256. [DOI] [PubMed] [Google Scholar]
- 374.Carpentier P.A., Begolka W.S., Olson J.K., Elhofy A., Karpus W.J., Miller S.D. Differential activation of astrocytes by innate and adaptive immune stimuli. Glia. 2005;49(3):360–374. doi: 10.1002/glia.20117. [DOI] [PubMed] [Google Scholar]
- 375.Crack P.J., Bray P.J. Toll-like receptors in the brain and their potential roles in neuropathology. Immunol. Cell Biol. 2007;85(6):476–480. doi: 10.1038/sj.icb.7100103. [DOI] [PubMed] [Google Scholar]
- 376.Hanke M.L., Kielian T. Toll-like receptors in health and disease in the brain: mechanisms and therapeutic potential. Clin. Sci. 2011;121(9):367–387. doi: 10.1042/CS20110164. (Lond.) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 377.Lafon M., Megret F., Lafage M., Prehaud C. The innate immune facet of brain: human neurons express TLR-3 and sense viral dsRNA. J. Mol. Neurosci. 2006;29(3):185–194. doi: 10.1385/JMN:29:3:185. [DOI] [PubMed] [Google Scholar]
- 378.Cameron J.S., Alexopoulou L., Sloane J.A., DiBernardo A.B., Ma Y., Kosaras B., Flavell R., Strittmatter S.M., Volpe J., Sidman R., Vartanian T. Toll-like receptor 3 is a potent negative regulator of axonal growth in mammals. J. Neurosci. 2007;27(47):13033–13041. doi: 10.1523/JNEUROSCI.4290-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 379.Wadachi R., Hargreaves K.M. Trigeminal nociceptors express TLR-4 and CD14: a mechanism for pain due to infection. J. Dent. Res. 2006;85(1):49–53. doi: 10.1177/154405910608500108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 380.Barajon I., Serrao G., Arnaboldi F., Opizzi E., Ripamonti G., Balsari A., Rumio C. Toll-like receptors 3, 4, and 7 are expressed in the enteric nervous system and dorsal root ganglia. J. Histochem. Cytochem. 2009;57(11):1013–1023. doi: 10.1369/jhc.2009.953539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 381.Ma Y., Li J., Chiu I., Wang Y., Sloane J.A., Lu J., Kosaras B., Sidman R.L., Volpe J.J., Vartanian T. Toll-like receptor 8 functions as a negative regulator of neurite outgrowth and inducer of neuronal apoptosis. J. Cell Biol. 2006;175(2):209–215. doi: 10.1083/jcb.200606016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 382.MacLullich A.M.J., Ferguson K.J., Miller T., de Rooij S.E.J.A., Cunningham C. Unravelling the pathophysiology of delirium: a focus on the role of aberrant stress responses. J. Psychosom. Res. 2008;65(3):229–238. doi: 10.1016/j.jpsychores.2008.05.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 383.Piva S., McCreadie V.A., Latronico N. Neuroinflammation in sepsis: sepsis associated delirium. Cardiovasc. Hematol. Disord. Drug Targets. 2015;15(1):10–18. doi: 10.2174/1871529x15666150108112452. [DOI] [PubMed] [Google Scholar]
- 384.Jalleh R., Koh K., Choi B., Liu E., Maddison J., Hutchinson M.R. Role of microglia and toll-like receptor 4 in the pathophysiology of delirium. Med. Hypotheses. 2012;79(6):735–739. doi: 10.1016/j.mehy.2012.08.013. [DOI] [PubMed] [Google Scholar]
- 385.Tsuruta R., Oda Y. A clinical perspective of sepsis-associated delirium. J. Intensive Care. 2016;4:18. doi: 10.1186/s40560-016-0145-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 386.Land W.G. The Role of Damage-Associated Molecular Patterns (DAMPs) in Human Diseases: Part II: DAMPs as diagnostics, prognostics and therapeutics in clinical medicine. Sultan Qaboos Univ. Med. J. 2015;15(2):e157–e170. [PMC free article] [PubMed] [Google Scholar]
- 387.Vénéreau E., Ceriotti C., Bianchi M.E. DAMPs from Cell Death to New Life. Front. Immunol. 2015;6(422) doi: 10.3389/fimmu.2015.00422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 388.Tang D., Kang R., Coyne C.B., Zeh H.J., Lotze M.T. PAMPs and DAMPs: Signal 0s that Spur Autophagy and Immunity. Immunol. Rev. 2012;249(1):158–175. doi: 10.1111/j.1600-065X.2012.01146.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 389.Rubartelli A., Lotze M.T. Inside, outside, upside down: damage-associated molecular-pattern molecules (DAMPs) and redox. Trends Immunol. 2007;28(10):429–436. doi: 10.1016/j.it.2007.08.004. [DOI] [PubMed] [Google Scholar]
- 390.Kanzok S.M., Hoa N.T., Bonizzoni M., Luna C., Huang Y., Malacrida A.R., Zheng L. Origin of Toll-like receptor-mediated innate immunity. J. Mol. Evol. 2004;58(4):442–448. doi: 10.1007/s00239-003-2565-8. [DOI] [PubMed] [Google Scholar]
- 391.Palmer J.D., Logsdon J.M., Jr. The recent origins of introns. Curr. Opin. Genet. Dev. 1991;1(4):470–477. doi: 10.1016/s0959-437x(05)80194-7. [DOI] [PubMed] [Google Scholar]
- 392.Koonin E.V. The origin of introns and their role in eukaryogenesis: a compromise solution to the introns-early versus introns-late debate? Biol. Direct. 2006;1:22. doi: 10.1186/1745-6150-1-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 393.Hoshino K., Takeuchi O., Kawai T., Sanjo H., Ogawa T., Takeda Y., Takeda K., Akira S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: evidence for TLR4 as the Lps gene product. J. Immunol. 1999;162(7):3749–3752. [PubMed] [Google Scholar]
- 394.Sun J., Wiklund F., Hsu F.C., Balter K., Zheng S.L., Johansson J.E., Chang B., Liu W., Li T., Turner A.R., Li L., Li G., Adami H.O., Isaacs W.B., Xu J., Gronberg H. Interactions of sequence variants in interleukin-1 receptor-associated kinase4 and the toll-like receptor 6-1-10 gene cluster increase prostate cancer risk. Cancer Epidemiol. Biomark. Prev. 2006;15(3):480–485. doi: 10.1158/1055-9965.EPI-05-0645. [DOI] [PubMed] [Google Scholar]
- 395.Sun J., Wiklund F., Zheng S.L., Chang B., Balter K., Li L., Johansson J.E., Li G., Adami H.O., Liu W., Tolin A., Turner A.R., Meyers D.A., Isaacs W.B., Xu J., Gronberg H. Sequence variants in Toll-like receptor gene cluster (TLR6-TLR1-TLR10) and prostate cancer risk. J. Natl. Cancer Inst. 2005;97(7):525–532. doi: 10.1093/jnci/dji070. [DOI] [PubMed] [Google Scholar]
- 396.Hawn T.R., Misch E.A., Dunstan S.J., Thwaites G.E., Lan N.T., Quy H.T., Chau T.T., Rodrigues S., Nachman A., Janer M., Hien T.T., Farrar J.J., Aderem A. A common human TLR1 polymorphism regulates the innate immune response to lipopeptides. Eur. J. Immunol. 2007;37(8):2280–2289. doi: 10.1002/eji.200737034. [DOI] [PubMed] [Google Scholar]
- 397.Johnson C.M., Lyle E.A., Omueti K.O., Stepensky V.A., Yegin O., Alpsoy E., Hamann L., Schumann R.R., Tapping R.I. Cutting edge: A common polymorphism impairs cell surface trafficking and functional responses of TLR1 but protects against leprosy. J. Immunol. 2007;178(12):7520–7524. doi: 10.4049/jimmunol.178.12.7520. [DOI] [PubMed] [Google Scholar]
- 398.Trejo-de la O.A., Hernandez-Sancen P., Maldonado-Bernal C. Relevance of single-nucleotide polymorphisms in human TLR genes to infectious and inflammatory diseases and cancer. Genes Immun. 2014;15(4):199–209. doi: 10.1038/gene.2014.10. [DOI] [PubMed] [Google Scholar]
- 399.Ogus A.C., Yoldas B., Ozdemir T., Uguz A., Olcen S., Keser I., Coskun M., Cilli A., Yegin O. The Arg753GLn polymorphism of the human toll-like receptor 2 gene in tuberculosis disease. Eur. Respir. J. 2004;23(2):219–223. doi: 10.1183/09031936.03.00061703. [DOI] [PubMed] [Google Scholar]
- 400.Tabel Y., Berdeli A., Mir S. Association of TLR2 gene Arg753Gln polymorphism with urinary tract infection in children. Int. J. Immunogenet. 2007;34(6):399–405. doi: 10.1111/j.1744-313X.2007.00709.x. [DOI] [PubMed] [Google Scholar]
- 401.Kutukculer N., Yeniay B.S., Aksu G., Berdeli A. Arg753Gln polymorphism of the human toll-like receptor-2 gene in children with recurrent febrile infections. Biochem. Genet. 2007;45(7–8):507–514. doi: 10.1007/s10528-007-9091-0. [DOI] [PubMed] [Google Scholar]
- 402.Berdeli A., Celik H.A., Ozyurek R., Dogrusoz B., Aydin H.H. TLR-2 gene Arg753Gln polymorphism is strongly associated with acute rheumatic fever in children. J. Mol. Med. 2005;83(7):535–541. doi: 10.1007/s00109-005-0677-x. [DOI] [PubMed] [Google Scholar]
- 403.Ozen S., Berdeli A., Turel B., Kutlay S., Yalcinkaya F., Arici M., Besbas N., Bakkaloglu A., Yilmaz E. Arg753Gln TLR-2 polymorphism in familial mediterranean fever: linking the environment to the phenotype in a monogenic inflammatory disease. J. Rheumatol. 2006;33(12):2498–2500. [PubMed] [Google Scholar]
- 404.Hamann L., Gomma A., Schroder N.W., Stamme C., Glaeser C., Schulz S., Gross M., Anker S.D., Fox K., Schumann R.R. A frequent toll-like receptor (TLR)-2 polymorphism is a risk factor for coronary restenosis. J. Mol. Med. 2005;83(6):478–485. doi: 10.1007/s00109-005-0643-7. [DOI] [PubMed] [Google Scholar]
- 405.Park Y., Park S., Yoo E., Kim D., Shin H. Association of the polymorphism for Toll-like receptor 2 with type 1 diabetes susceptibility. Ann. N. Y. Acad. Sci. 2004;1037:170–174. doi: 10.1196/annals.1337.028. [DOI] [PubMed] [Google Scholar]
- 406.Eid A.J., Brown R.A., Paya C.V., Razonable R.R. Association between toll-like receptor polymorphisms and the outcome of liver transplantation for chronic hepatitis C virus. Transplantation. 2007;84(4):511–516. doi: 10.1097/01.tp.0000276960.35313.bf. [DOI] [PubMed] [Google Scholar]
- 407.Kijpittayarit S., Eid A.J., Brown R.A., Paya C.V., Razonable R.R. Relationship between Toll-like receptor 2 polymorphism and cytomegalovirus disease after liver transplantation. Clin. Infect. Dis. 2007;44(10):1315–1320. doi: 10.1086/514339. [DOI] [PubMed] [Google Scholar]
- 408.Studzińska M., Jabłońska A., Wiśniewska-Ligier M., Nowakowska D., Gaj Z., Leśnikowski Z.J., Woźniakowska-Gęsicka T., Wilczyński J., Paradowska E. Association of TLR3 L412F Polymorphism with Cytomegalovirus Infection in Children. PLoS ONE. 2017;12(1) doi: 10.1371/journal.pone.0169420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 409.Wang B.-G., Yi D.-H., Liu Y.-F. TLR3 gene polymorphisms in cancer: a systematic review and meta-analysis. Chin. J. Cancer. 2015;34:19. doi: 10.1186/s40880-015-0020-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 410.Ma L., Tang F.Y., Chu W.K., Young A.L., Brelen M.E., Pang C.P., Chen L.J. Association of toll-like receptor 3 polymorphism rs3775291 with age-related macular degeneration: a systematic review and meta-analysis. Sci. Rep. 2016;6:19718. doi: 10.1038/srep19718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 411.Gorbea C., Makar K.A., Pauschinger M., Pratt G., Bersola J.L., Varela J., David R.M., Banks L., Huang C.H., Li H., Schultheiss H.P., Towbin J.A., Vallejo J.G., Bowles N.E. A role for Toll-like receptor 3 variants in host susceptibility to enteroviral myocarditis and dilated cardiomyopathy. J. Biol. Chem. 2010;285(30):23208–23223. doi: 10.1074/jbc.M109.047464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 412.Ejghal R., Hida M., Bennani M.L., Meziane M., Aurag R., Lemrani M. The TLR2 and TLR4 gene polymorphisms in Moroccan visceral leishmaniasis patients. Acta Trop. 2016;158:77–82. doi: 10.1016/j.actatropica.2016.02.020. [DOI] [PubMed] [Google Scholar]
- 413.Lei T., Tang W., Xiong Y., Di Y., Zhang K., Shu X. Association of TLR4 gene polymorphisms with susceptibility to type 2 diabetes mellitus in the Chinese Han population. Int. Immunopharmacol. 2015;24(1):68–71. doi: 10.1016/j.intimp.2014.11.007. [DOI] [PubMed] [Google Scholar]
- 414.Zerdazi E.H., Oliveira J., Vorspan F., Bennabi M., Jamain S., Etain B., Leboyer M., Tamouza R., Bellivier F. TLR4 gene polymorphism associated with lifetime cigarette smoking in bipolar disorder. J. Neuroimmunol. 2017;305:96–101. doi: 10.1016/j.jneuroim.2017.01.021. [DOI] [PubMed] [Google Scholar]
- 415.Kiziltas S., Ata P., Colak Y., Mesci B., Senates E., Enc F., Ulasoglu C., Tuncer I., Oguz A. TLR4 gene polymorphism in patients with nonalcoholic fatty liver disease in comparison to healthy controls. Metab. Syndr. Relat. Disord. 2014;12(3):165–170. doi: 10.1089/met.2013.0120. [DOI] [PubMed] [Google Scholar]
- 416.Meena N.K., Ahuja V., Meena K., Paul J. Association of TLR5 Gene Polymorphisms in Ulcerative Colitis Patients of North India and Their Role in Cytokine Homeostasis. PLoS ONE. 2015;10(3) doi: 10.1371/journal.pone.0120697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 417.Xiao W., Liu Z., Lin J., Xiong C., Li J., Wu K., Ma Y., Gong Y., Liu Z. Association of TLR4 and TLR5 gene polymorphisms with Graves’ disease in Chinese Cantonese population. Hum. Immunol. 2014;75(7):609–613. doi: 10.1016/j.humimm.2014.05.001. [DOI] [PubMed] [Google Scholar]
- 418.Chen Z., Wang W., Liang J., Wang J., Feng S., Zhang G. Association between toll-like receptors 9 (TLR9) gene polymorphism and risk of pulmonary tuberculosis: meta-analysis. BMC Pulm. Med. 2015;15:57. doi: 10.1186/s12890-015-0049-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 419.Wifi M.-N.A., Assem M., Elsherif R.H., El-Azab H.A.-F., Saif A. Toll-like receptors-2 and -9 (TLR2 and TLR9) gene polymorphism in patients with type 2 diabetes and diabetic foot. Medicine. 2017;96(17) doi: 10.1097/MD.0000000000006760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 420.Noreen M., Arshad M. Association of TLR1, TLR2, TLR4, TLR6, and TIRAP polymorphisms with disease susceptibility. Immunol. Res. 2015;62(2):234–252. doi: 10.1007/s12026-015-8640-6. [DOI] [PubMed] [Google Scholar]
- 421.Sharma S., Garg I., Ashraf M.Z. TLR signalling and association of TLR polymorphism with cardiovascular diseases. Vasc. Pharmacol. 2016;87:30–37. doi: 10.1016/j.vph.2016.10.008. [DOI] [PubMed] [Google Scholar]
- 422.Schwartz D.A., Cook D.N. Polymorphisms of the Toll-like receptors and human disease. Clin. Infect. Dis. 2005;41(Suppl 7):S403–7. doi: 10.1086/431985. [DOI] [PubMed] [Google Scholar]
- 423.Tokarz S.A., DeValk J., Luo W., Pattnaik B.R., Schrodi S.J., Pillers D.A. Cell line donor genotype and its influence on experimental phenotype: Toll-like receptor SNPs and potential variability in innate immunity. Mol. Genet. Metab. 2016;118(3):147–152. doi: 10.1016/j.ymgme.2016.04.011. [DOI] [PubMed] [Google Scholar]
- 424.Opal S.M., Laterre P.F., Francois B., LaRosa S.P., Angus D.C., Mira J.P., Wittebole X., Dugernier T., Perrotin D., Tidswell M., Jauregui L., Krell K., Pachl J., Takahashi T., Peckelsen C., Cordasco E., Chang C.S., Oeyen S., Aikawa N., Maruyama T., Schein R., Kalil A.C., Van Nuffelen M., Lynn M., Rossignol D.P., Gogate J., Roberts M.B., Wheeler J.L., Vincent J.L. Effect of eritoran, an antagonist of MD2-TLR4, on mortality in patients with severe sepsis: the ACCESS randomized trial. JAMA. 2013;309(11):1154–1162. doi: 10.1001/jama.2013.2194. [DOI] [PubMed] [Google Scholar]
- 425.Shirey K.A., Lai W., Scott A.J., Lipsky M., Mistry P., Pletneva L.M., Karp C.L., McAlees J., Gioannini T.L., Weiss J., Chen W.H., Ernst R.K., Rossignol D.P., Gusovsky F., Blanco J.C.G., Vogel S.N. The TLR4 antagonist Eritoran protects mice from lethal influenza infection. Nature. 2013;497(7450):498–502. doi: 10.1038/nature12118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 426.Ulrich J.T., Myers K.R. Monophosphoryl lipid A as an adjuvant. Past experiences and new directions. Pharm. Biotechnol. 1995;6:495–524. [PubMed] [Google Scholar]
- 427.Johnson A.G. Molecular adjuvants and immunomodulators: new approaches to immunization. Clin. Microbiol. Rev. 1994;7(3):277–289. doi: 10.1128/cmr.7.3.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 428.Baldridge J.R., McGowan P., Evans J.T., Cluff C., Mossman S., Johnson D., Persing D. Taking a Toll on human disease: Toll-like receptor 4 agonists as vaccine adjuvants and monotherapeutic agents. Expert. Opin. Biol. Ther. 2004;4(7):1129–1138. doi: 10.1517/14712598.4.7.1129. [DOI] [PubMed] [Google Scholar]
- 429.Steinhagen F., Kinjo T., Bode C., Klinman D.M. TLR-based immune adjuvants. Vaccine. 2011;29(17):3341–3355. doi: 10.1016/j.vaccine.2010.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 430.Garcon N., Chomez P., Van Mechelen M. GlaxoSmithKline Adjuvant Systems in vaccines: concepts, achievements and perspectives. Expert Rev. Vaccines. 2007;6(5):723–739. doi: 10.1586/14760584.6.5.723. [DOI] [PubMed] [Google Scholar]
- 431.Sondak V.K., Sosman J.A. Results of clinical trials with an allogenic melanoma tumor cell lysate vaccine: Melacine. Semin. Cancer Biol. 2003;13(6):409–415. doi: 10.1016/j.semcancer.2003.09.004. [DOI] [PubMed] [Google Scholar]
- 432.North S., Butts C. Vaccination with BLP25 liposome vaccine to treat non-small cell lung and prostate cancers. Expert Rev. Vaccines. 2005;4(3):249–257. doi: 10.1586/14760584.4.3.249. [DOI] [PubMed] [Google Scholar]
- 433.Uenishi Y., Okada T., Okabe S., Sunagawa M. Study on the cell wall skeleton derived from Mycobacterium bovis BCG Tokyo 172 (SMP-105): establishment of preparation and analytical methods. Chem. Pharm. Bull. 2007;55(6):843–852. doi: 10.1248/cpb.55.843. [DOI] [PubMed] [Google Scholar]
- 434.Morales A., Eidinger D., Bruce A.W. Intracavitary Bacillus Calmette-Guerin in the treatment of superficial bladder tumors. J. Urol. 1976;116(2):180–183. doi: 10.1016/s0022-5347(17)58737-6. [DOI] [PubMed] [Google Scholar]
- 435.Simons M.P., O’Donnell M.A., Griffith T.S. Role of Neutrophils in BCG Immunotherapy for Bladder Cancer. Urol. Oncol. 2008;26(4):341–345. doi: 10.1016/j.urolonc.2007.11.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 436.Murata M. Activation of Toll-like receptor 2 by a novel preparation of cell wall skeleton from Mycobacterium bovis BCG Tokyo (SMP-105) sufficiently enhances immune responses against tumors. Cancer Sci. 2008;99(7):1435–1440. doi: 10.1111/j.1349-7006.2008.00832.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 437.Kato T., Bilim V., Yuuki K., Naito S., Yamanobe T., Nagaoka A., Yano I., Akaza H., Tomita Y. Bacillus Calmette-Guerin and BCG cell wall skeleton suppressed viability of bladder cancer cells in vitro. Anticancer Res. 2010;30(10):4089–4096. [PubMed] [Google Scholar]
- 438.Hennessy E.J., Parker A.E., O'Neill L.A. Targeting Toll-like receptors: emerging therapeutics? Nat. Rev. Drug Discov. 2010;9(4):293–307. doi: 10.1038/nrd3203. [DOI] [PubMed] [Google Scholar]
- 439.Panter G., Kuznik A., Jerala R. Therapeutic applications of nucleic acids as ligands for Toll-like receptors. Curr. Opin. Mol. Ther. 2009;11(2):133–145. [PubMed] [Google Scholar]
- 440.Navabi H., Jasani B., Reece A., Clayton A., Tabi Z., Donninger C., Mason M., Adams M. A clinical grade poly I:C-analogue (Ampligen) promotes optimal DC maturation and Th1-type T cell responses of healthy donors and cancer patients in vitro. Vaccine. 2009;27(1):107–115. doi: 10.1016/j.vaccine.2008.10.024. [DOI] [PubMed] [Google Scholar]
- 441.Jasani B., Navabi H., Adams M. Ampligen: a potential toll-like 3 receptor adjuvant for immunotherapy of cancer. Vaccine. 2009;27(25–26):3401–3404. doi: 10.1016/j.vaccine.2009.01.071. [DOI] [PubMed] [Google Scholar]
- 442.Hemmi H., Kaisho T., Takeuchi O., Sato S., Sanjo H., Hoshino K., Horiuchi T., Tomizawa H., Takeda K., Akira S. Small anti-viral compounds activate immune cells via the TLR7 MyD88-dependent signaling pathway. Nat. Immunol. 2002;3(2):196–200. doi: 10.1038/ni758. [DOI] [PubMed] [Google Scholar]
- 443.Beutner K.R., Spruance S.L., Hougham A.J., Fox T.L., Owens M.L., Douglas J.M., Jr. Treatment of genital warts with an immune-response modifier (imiquimod) J. Am. Acad. Dermatol. 1998;38(2 Pt 1):230–239. doi: 10.1016/s0190-9622(98)70243-9. [DOI] [PubMed] [Google Scholar]
- 444.Stanley M.A. Imiquimod and the imidazoquinolones: mechanism of action and therapeutic potential. Clin. Exp. Dermatol. 2002;27(7):571–577. doi: 10.1046/j.1365-2230.2002.01151.x. [DOI] [PubMed] [Google Scholar]
- 445.Schulze H.J., Cribier B., Requena L., Reifenberger J., Ferrandiz C., Garcia Diez A., Tebbs V., McRae S. Imiquimod 5% cream for the treatment of superficial basal cell carcinoma: results from a randomized vehicle-controlled phase III study in Europe. Br. J. Dermatol. 2005;152(5):939–947. doi: 10.1111/j.1365-2133.2005.06486.x. [DOI] [PubMed] [Google Scholar]
- 446.O'Neill L.A., Bryant C.E., Doyle S.L. Therapeutic targeting of Toll-like receptors for infectious and inflammatory diseases and cancer. Pharmacol. Rev. 2009;61(2):177–197. doi: 10.1124/pr.109.001073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 447.Harrison L.I., Astry C., Kumar S., Yunis C. Pharmacokinetics of 852A, an Imidazoquinoline Toll-Like Receptor 7-Specific Agonist, Following Intravenous, Subcutaneous, and Oral Administrations in Humans. J. Clin. Pharmacol. 2007;47(8):962–969. doi: 10.1177/0091270007303766. [DOI] [PubMed] [Google Scholar]
- 448.Geller M.A., Cooley S., Argenta P.A., Downs L.S., Carson L.F., Judson P.L., Ghebre R., Weigel B., Panoskaltsis-Mortari A., Curtsinger J., Miller J.S. Toll-Like Receptor-7 (TLR-7) Agonist Administered Subcutaneously in a Prolonged Dosing Schedule in Heavily Pretreated Recurrent Breast, Ovarian, and Cervix Cancers. Cancer Immunol. Immunother. 2010;59(12):1877–1884. doi: 10.1007/s00262-010-0914-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 449.Engel A.L., Holt G.E., Lu H. The pharmacokinetics of Toll-like receptor agonists and the impact on the immune system. Expert. Rev. Clin. Pharmacol. 2011;4(2):275–289. doi: 10.1586/ecp.11.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 450.Dudek A.Z., Yunis C., Harrison L.I., Kumar S., Hawkinson R., Cooley S., Vasilakos J.P., Gorski K.S., Miller J.S. First in human phase I trial of 852A, a novel systemic toll-like receptor 7 agonist, to activate innate immune responses in patients with advanced cancer. Clin. Cancer Res. 2007;13(23):7119–7125. doi: 10.1158/1078-0432.CCR-07-1443. [DOI] [PubMed] [Google Scholar]
- 451.Dummer R., Hauschild A., Becker J.C., Grob J.J., Schadendorf D., Tebbs V., Skalsky J., Kaehler K.C., Moosbauer S., Clark R., Meng T.C., Urosevic M. An exploratory study of systemic administration of the toll-like receptor-7 agonist 852A in patients with refractory metastatic melanoma. Clin. Cancer Res. 2008;14(3):856–864. doi: 10.1158/1078-0432.CCR-07-1938. [DOI] [PubMed] [Google Scholar]
- 452.Agrawal S., Kandimalla E.R. Synthetic agonists of Toll-like receptors 7, 8 and 9. Biochem. Soc. Trans. 2007;35(Pt 6):1461–1467. doi: 10.1042/BST0351461. [DOI] [PubMed] [Google Scholar]
- 453.Goodchild A., Nopper N., Craddock A., Law T., King A., Fanning G., Rivory L., Passioura T. Primary leukocyte screens for innate immune agonists. J. Biomol. Screen. 2009;14(6):723–730. doi: 10.1177/1087057109335325. [DOI] [PubMed] [Google Scholar]
- 454.Krieg A.M. Toll-like receptor 9 (TLR9) agonists in the treatment of cancer. Oncogene. 2008;27(2):161–167. doi: 10.1038/sj.onc.1210911. [DOI] [PubMed] [Google Scholar]
- 455.Iribarren K., Bloy N., Buqué A., Cremer I., Eggermont A., Fridman W.H., Fucikova J., Galon J., Špíšek R., Zitvogel L., Kroemer G., Galluzzi L. Trial Watch: Immunostimulation with Toll-like receptor agonists in cancer therapy. Oncoimmunology. 2016;5(3) doi: 10.1080/2162402X.2015.1088631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 456.Rakoff-Nahoum S., Medzhitov R. Toll-like receptors and cancer. Nat. Rev. Cancer. 2009;9(1):57–63. doi: 10.1038/nrc2541. [DOI] [PubMed] [Google Scholar]
- 457.Okamoto H., Shoin S., Koshimura S., Shimizu R. Studies on the anticancer and streptolysin S-forming abilities of hemolytic streptococci. Jpn. J. Microbiol. 1967;11(4):323–326. doi: 10.1111/j.1348-0421.1967.tb00350.x. [DOI] [PubMed] [Google Scholar]
- 458.Paavonen J., Naud P., Salmeron J., Wheeler C.M., Chow S.N., Apter D., Kitchener H., Castellsague X., Teixeira J.C., Skinner S.R., Hedrick J., Jaisamrarn U., Limson G., Garland S., Szarewski A., Romanowski B., Aoki F.Y., Schwarz T.F., Poppe W.A., Bosch F.X., Jenkins D., Hardt K., Zahaf T., Descamps D., Struyf F., Lehtinen M., Dubin G. Efficacy of human papillomavirus (HPV)-16/18 AS04-adjuvanted vaccine against cervical infection and precancer caused by oncogenic HPV types (PATRICIA): final analysis of a double-blind, randomised study in young women. Lancet. 2009;374(9686):301–314. doi: 10.1016/S0140-6736(09)61248-4. (London, England) [DOI] [PubMed] [Google Scholar]
- 459.Lehtinen M., Paavonen J. Sound efficacy of prophylactic HPV vaccination: Basics and implications. Oncoimmunology. 2012;1(6):995–996. doi: 10.4161/onci.20011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 460.Huang S.W., Kao J.K., Wu C.Y., Wang S.T., Lee H.C., Liang S.M., Chen Y.J., Shieh J.J. Targeting aerobic glycolysis and HIF-1alpha expression enhance imiquimod-induced apoptosis in cancer cells. Oncotarget. 2014;5(5):1363–1381. doi: 10.18632/oncotarget.1734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 461.Hoffman E.S., Smith R.E., Renaud R.C., Jr. From the analyst's couch: TLR-targeted therapeutics. Nat. Rev. Drug Discov. 2005;4(11):879–880. doi: 10.1038/nrd1880. [DOI] [PubMed] [Google Scholar]
- 462.Dowling J.K., Mansell A. Toll-like receptors: the swiss army knife of immunity and vaccine development. Clin. Transl. Immunology. 2016;5 doi: 10.1038/cti.2016.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 463.Kochling J., Prada J., Bahrami M., Stripecke R., Seeger K., Henze G., Wittig B., Schmidt M. Anti-tumor effect of DNA-based vaccination and dSLIM immunomodulatory molecules in mice with Ph+ acute lymphoblastic leukaemia. Vaccine. 2008;26(36):4669–4675. doi: 10.1016/j.vaccine.2008.06.094. [DOI] [PubMed] [Google Scholar]
- 464.Parkinson T. The future of toll-like receptor therapeutics. Curr. Opin. Mol. Ther. 2008;10(1):21–31. [PubMed] [Google Scholar]
- 465.Perego J., Mendes A., Bourbon C., Camosseto V., Combes A., Liu H., Manh T.V., Dalet A., Chasson L., Spinelli L., Bardin N., Chiche L., Santos M.A.S., Gatti E., Pierre P. Guanabenz inhibits TLR9 signaling through a pathway that is independent of eIF2alpha dephosphorylation by the GADD34/PP1c complex. Sci. Signal. 2018;11(514) doi: 10.1126/scisignal.aam8104. [DOI] [PubMed] [Google Scholar]
- 466.Negishi H., Ohba Y., Yanai H., Takaoka A., Honma K., Yui K., Matsuyama T., Taniguchi T., Honda K. Negative regulation of Toll-like-receptor signaling by IRF-4. Proc. Natl. Acad. Sci. U. S. A. 2005;102(44):15989–15994. doi: 10.1073/pnas.0508327102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 467.O'Neill L.A. When signaling pathways collide: positive and negative regulation of toll-like receptor signal transduction. Immunity. 2008;29(1):12–20. doi: 10.1016/j.immuni.2008.06.004. [DOI] [PubMed] [Google Scholar]
- 468.Kondo T., Kawai T., Akira S. Dissecting negative regulation of Toll-like receptor signaling. Trends Immunol. 2012;33(9):449–458. doi: 10.1016/j.it.2012.05.002. [DOI] [PubMed] [Google Scholar]
- 469.Rodriguez-Calvo R., Tajes M., Vazquez-Carrera M. The NR4A subfamily of nuclear receptors: potential new therapeutic targets for the treatment of inflammatory diseases. Expert Opin. Ther. Targets. 2017;21(3):291–304. doi: 10.1080/14728222.2017.1279146. [DOI] [PubMed] [Google Scholar]
- 470.Sun H., Gong S., Carmody R.J., Hilliard A., Li L., Sun J., Kong L., Xu L., Hilliard B., Hu S., Shen H., Yang X., Chen Y.H. TIPE2, a novel negative regulator of innate and adaptive immunity that maintains immune homeostasis. Cell. 2008;133(3):415–426. doi: 10.1016/j.cell.2008.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 471.Mokuno Y., Matsuguchi T., Takano M., Nishimura H., Washizu J., Ogawa T., Takeuchi O., Akira S., Nimura Y., Yoshikai Y. Expression of Toll-Like Receptor 2 on γδ T Cells Bearing Invariant Vγ6/Vδ1 Induced by Escherichia coli Infection in Mice. J. Immunol. 2000;165(2):931–940. doi: 10.4049/jimmunol.165.2.931. [DOI] [PubMed] [Google Scholar]
- 472.Funderburg N., Luciano A.A., Jiang W., Rodriguez B., Sieg S.F., Lederman M.M. Toll-Like Receptor Ligands Induce Human T Cell Activation and Death, a Model for HIV Pathogenesis. PLoS ONE. 2008;3(4) doi: 10.1371/journal.pone.0001915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 473.Sutmuller R.P.M., den Brok M.H., Kramer M., Bennink E.J., Toonen L.W.J., Kullberg B.-J., Joosten L.A., Akira S., Netea M.G., Adema G.J. Toll-like receptor 2 controls expansion and function of regulatory T cells. J. Clin. Investig. 2006;116(2):485–494. doi: 10.1172/JCI25439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 474.Marafini I., Monteleone I., Di Fusco D., Cupi M.L., Paoluzi O.A., Colantoni A., Ortenzi A., Izzo R., Vita S., De Luca E., Sica G., Pallone F., Monteleone G. TNF-α Producing Innate Lymphoid Cells (ILCs) Are Increased in Active Celiac Disease and Contribute to Promote Intestinal Atrophy in Mice. PLoS ONE. 2015;10(5) doi: 10.1371/journal.pone.0126291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 475.Kumar V. Innate Lymphoid Cells: Immunoregulatory Cells of Mucosal Inflammation. Eur. J. Inflamm. 2014;12(1):11–20. [Google Scholar]
- 476.Boyd A., Ribeiro J.M.C., Nutman T.B. Human CD117 (cKit)+ Innate Lymphoid Cells Have a Discrete Transcriptional Profile at Homeostasis and Are Expanded during Filarial Infection. PLoS ONE. 2014;9(9) doi: 10.1371/journal.pone.0108649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 477.Crellin N.K., Trifari S., Kaplan C.D., Satoh-Takayama N., Di Santo J.P., Spits H. Regulation of Cytokine Secretion in Human CD127+ LTi-like Innate Lymphoid Cells by Toll-like Receptor 2. Immunity. 2010;33(5):752–764. doi: 10.1016/j.immuni.2010.10.012. [DOI] [PubMed] [Google Scholar]
- 478.Xu H., Wang X., Lackner A.A., Veazey R.S. Type 3 innate lymphoid cell depletion is mediated by TLRs in lymphoid tissues of simian immunodeficiency virus-infected macaques. FASEB J. 2015;29(12):5072–5080. doi: 10.1096/fj.15-276477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 479.Roelofs M.F., Abdollahi-Roodsaz S., Joosten L.A., van den Berg W.B., Radstake T.R. The orchestra of Toll-like receptors and their potential role in frequently occurring rheumatic conditions. Arthritis Rheum. 2008;58(2):338–348. doi: 10.1002/art.23217. [DOI] [PubMed] [Google Scholar]
- 480.Mohammad Hosseini A., Majidi J., Baradaran B., Yousefi M. Toll-Like Receptors in the Pathogenesis of Autoimmune Diseases. Adv. Pharm. Bull. 2015;5(Suppl. 1):605–614. doi: 10.15171/apb.2015.082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 481.Farrugia M., Baron B. The Role of Toll-Like Receptors in Autoimmune Diseases through Failure of the Self-Recognition Mechanism. Int. J. Inflamm. 2017;2017:8391230. doi: 10.1155/2017/8391230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 482.Duffy L., O’Reilly S.C. Toll-like receptors in the pathogenesis of autoimmune diseases: recent and emerging translational developments. Immunotargets Ther. 2016;5:69–80. doi: 10.2147/ITT.S89795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 483.O'Reilly S. Toll Like Receptors in systemic sclerosis: An emerging target. Immunol. Lett. 2018;195:2–8. doi: 10.1016/j.imlet.2017.09.001. [DOI] [PubMed] [Google Scholar]
- 484.Jimenez-Dalmaroni M.J., Gerswhin M.E., Adamopoulos I.E. The critical role of toll-like receptors--From microbial recognition to autoimmunity: A comprehensive review. Autoimmun. Rev. 2016;15(1):1–8. doi: 10.1016/j.autrev.2015.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 485.Isaza-Correa J.M., Liang Z., van den Berg A., Diepstra A., Visser L. Toll-like receptors in the pathogenesis of human B cell malignancies. J. Hematol. Oncol. 2014;7:57. doi: 10.1186/s13045-014-0057-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 486.O'Neill L.A.J. Toll-like receptors in cancer. Oncogene. 2008;27:158. [Google Scholar]
- 487.Huang B., Zhao J., Unkeless J.C., Feng Z.H., Xiong H. TLR signaling by tumor and immune cells: a double-edged sword. Oncogene. 2008;27:218. doi: 10.1038/sj.onc.1210904. [DOI] [PubMed] [Google Scholar]
- 488.Li T.T., Ogino S., Qian Z.R. Toll-like receptor signaling in colorectal cancer: carcinogenesis to cancer therapy. World J. Gastroenterol. 2014;20(47):17699–17708. doi: 10.3748/wjg.v20.i47.17699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 489.Basith S., Manavalan B., Yoo T.H., Kim S.G., Choi S. Roles of toll-like receptors in Cancer: A double-edged sword for defense and offense. Arch. Pharm. Res. 2012;35(8):1297–1316. doi: 10.1007/s12272-012-0802-7. [DOI] [PubMed] [Google Scholar]
- 490.Shi M., Chen X., Ye K., Yao Y., Li Y. Application potential of toll-like receptors in cancer immunotherapy: Systematic review. Medicine. 2016;95(25) doi: 10.1097/MD.0000000000003951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 491.Barrat F.J., Coffman R.L. Development of TLR inhibitors for the treatment of autoimmune diseases. Immunol. Rev. 2008;223:271–283. doi: 10.1111/j.1600-065X.2008.00630.x. [DOI] [PubMed] [Google Scholar]
- 492.Kaczanowska S., Joseph A.M., Davila E. TLR agonists: our best frenemy in cancer immunotherapy. J. Leukoc. Biol. 2013;93(6):847–863. doi: 10.1189/jlb.1012501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 493.Li K., Qu S., Chen X., Wu Q., Shi M. Promising Targets for Cancer Immunotherapy: TLRs, RLRs, and STING-Mediated Innate Immune Pathways. Int. J. Mol. Sci. 2017;18(2):404. doi: 10.3390/ijms18020404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 494.Du B., Jiang Q.-L., Cleveland J., Liu B.-R., Zhang D. Targeting Toll-like receptors against cancer. J. Cancer Metastasis Treat. 2016;2(12):463–470. [Google Scholar]
- 495.Baird J.R., Monjazeb A.M., Shah O., McGee H., Murphy W.J., Crittenden M.R., Gough M.J. Stimulating Innate Immunity to Enhance Radiation Therapy-Induced Tumor Control. Int. J. Radiat. Oncol. Biol. Phys. 2017;99(2):362–373. doi: 10.1016/j.ijrobp.2017.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 496.Okun E., Griffioen K.J., Lathia J.D., Tang S.-C., Mattson M.P., Arumugam T.V. Toll-Like Receptors in Neurodegeneration. Brain Res. Rev. 2009;59(2):278–292. doi: 10.1016/j.brainresrev.2008.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 497.Arroyo D.S., Soria J.A., Gaviglio E.A., Rodriguez-Galan M.C., Iribarren P. Toll-like receptors are key players in neurodegeneration. Int. Immunopharmacol. 2011;11(10):1415–1421. doi: 10.1016/j.intimp.2011.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 498.Drouin-Ouellet J., Cicchetti F. Inflammation and neurodegeneration: the story 'retolled'. Trends Pharmacol. Sci. 2012;33(10):542–551. doi: 10.1016/j.tips.2012.07.002. [DOI] [PubMed] [Google Scholar]
- 499.van Noort J.M., Bsibsi M. Toll-like receptors in the CNS: implications for neurodegeneration and repair. Prog. Brain Res. 2009;175:139–148. doi: 10.1016/S0079-6123(09)17509-X. [DOI] [PubMed] [Google Scholar]
- 500.Rosenberger K., Derkow K., Dembny P., Krüger C., Schott E., Lehnardt S. The impact of single and pairwise Toll-like receptor activation on neuroinflammation and neurodegeneration. J. Neuroinflammation. 2014;11(1):166. doi: 10.1186/s12974-014-0166-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 501.Gao W., Xiong Y., Li Q., Yang H. Inhibition of Toll-Like Receptor Signaling as a Promising Therapy for Inflammatory Diseases: A Journey from Molecular to Nano Therapeutics. Front. Physiol. 2017;8:508. doi: 10.3389/fphys.2017.00508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 502.Achek A., Yesudhas D., Choi S. Toll-like receptors: promising therapeutic targets for inflammatory diseases. Arch. Pharm. Res. 2016;39(8):1032–1049. doi: 10.1007/s12272-016-0806-9. [DOI] [PubMed] [Google Scholar]
- 503.Kannaki T.R., Shanmugam M., Verma P.C. Toll-like receptors and their role in animal reproduction. Anim. Reprod. Sci. 2011;125(1–4):1–12. doi: 10.1016/j.anireprosci.2011.03.008. [DOI] [PubMed] [Google Scholar]
- 504.Girling J.E., Hedger M.P. Toll-like receptors in the gonads and reproductive tract: emerging roles in reproductive physiology and pathology. Immunol. Cell Biol. 2007;85(6):481–489. doi: 10.1038/sj.icb.7100086. [DOI] [PubMed] [Google Scholar]
- 505.Aflatoonian R., Fazeli A. Toll-like receptors in female reproductive tract and their menstrual cycle dependent expression. J. Reprod. Immunol. 2008;77(1):7–13. doi: 10.1016/j.jri.2007.03.014. [DOI] [PubMed] [Google Scholar]
- 506.Hedger M.P. Toll-like receptors and signalling in spermatogenesis and testicular responses to inflammation—a perspective. J. Reprod. Immunol. 2011;88(2):130–141. doi: 10.1016/j.jri.2011.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 507.Iimuro Y., Fujimoto J. TLRs, NF-κB, JNK, and Liver Regeneration. Gastroenterol. Res. Pract. 2010;2010:598109. doi: 10.1155/2010/598109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 508.Seki E., Tsutsui H., Iimuro Y., Naka T., Son G., Akira S., Kishimoto T., Nakanishi K., Fujimoto J. Contribution of Toll-like receptor/myeloid differentiation factor 88 signaling to murine liver regeneration. Hepatology. 2005;41(3):443–450. doi: 10.1002/hep.20603. (Baltimore, Md.) [DOI] [PubMed] [Google Scholar]
- 509.Chen Y., Sun R. Toll-like receptors in acute liver injury and regeneration. Int. Immunopharmacol. 2011;11(10):1433–1441. doi: 10.1016/j.intimp.2011.04.023. [DOI] [PubMed] [Google Scholar]
- 510.Braza F., Brouard S., Chadban S., Goldstein D.R. Role of TLRs and DAMPs in allograft inflammation and transplant outcomes. Nat. Rev. Nephrol. 2016;12(5):281–290. doi: 10.1038/nrneph.2016.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 511.Wu H., Chadban S.J. Roles of Toll-like receptors in transplantation. Curr. Opin. Organ Transplant. 2014;19(1):1–7. doi: 10.1097/MOT.0000000000000038. [DOI] [PubMed] [Google Scholar]
- 512.Goldstein D.R., Tesar B.M., Akira S., Lakkis F.G. Critical role of the Toll-like receptor signal adaptor protein MyD88 in acute allograft rejection. J. Clin. Invest. 2003;111(10):1571–1578. doi: 10.1172/JCI17573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 513.Leventhal J.S., Schroppel B. Toll-like receptors in transplantation: sensing and reacting to injury. Kidney Int. 2012;81(9):826–832. doi: 10.1038/ki.2011.498. [DOI] [PubMed] [Google Scholar]
- 514.Kesh S., Mensah N.Y., Peterlongo P., Jaffe D., Hsu K., V.D.B. M, O'Reilly R., Pamer E., Satagopan J., Papanicolaou G.A. TLR1 and TLR6 polymorphisms are associated with susceptibility to invasive aspergillosis after allogeneic stem cell transplantation. Ann. N. Y. Acad. Sci. 2005;1062:95–103. doi: 10.1196/annals.1358.012. [DOI] [PubMed] [Google Scholar]
- 515.Mezger M., Einsele H., Loeffler J. Genetic susceptibility to infections with Aspergillus fumigatus. Crit. Rev. Microbiol. 2010;36(2):168–177. doi: 10.3109/10408410903530619. [DOI] [PubMed] [Google Scholar]
- 516.Bochud P.Y., Hawn T.R., Siddiqui M.R., Saunderson P., Britton S., Abraham I., Argaw A.T., Janer M., Zhao L.P., Kaplan G., Aderem A. Toll-like receptor 2 (TLR2) polymorphisms are associated with reversal reaction in leprosy. J. Infect. Dis. 2008;197(2):253–261. doi: 10.1086/524688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 517.Thuong N.T., Hawn T.R., Thwaites G.E., Chau T.T., Lan N.T., Quy H.T., Hieu N.T., Aderem A., Hien T.T., Farrar J.J., Dunstan S.J. A polymorphism in human TLR2 is associated with increased susceptibility to tuberculous meningitis. Genes Immun. 2007;8(5):422–428. doi: 10.1038/sj.gene.6364405. [DOI] [PubMed] [Google Scholar]
- 518.Moore C.E., Segal S., Berendt A.R., Hill A.V., Day N.P. Lack of association between Toll-like receptor 2 polymorphisms and susceptibility to severe disease caused by Staphylococcus aureus. Clin. Diagn. Lab. Immunol. 2004;11(6):1194–1197. doi: 10.1128/CDLI.11.6.1194-1197.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 519.Bochud P.Y., Magaret A.S., Koelle D.M., Aderem A., Wald A. Polymorphisms in TLR2 are associated with increased viral shedding and lesional rate in patients with genital herpes simplex virus Type 2 infection. J. Infect. Dis. 2007;196(4):505–509. doi: 10.1086/519693. [DOI] [PubMed] [Google Scholar]
- 520.Al-Qahtani A., Al-Ahdal M., Abdo A., Sanai F., Al-Anazi M., Khalaf N., Viswan N.A., Al-Ashgar H., Al-Humaidan H., Al-Suwayeh R., Hussain Z., Alarifi S., Al-Okail M., Almajhdi F.N. Toll-like receptor 3 polymorphism and its association with hepatitis B virus infection in Saudi Arabian patients. J. Med. Virol. 2012;84(9):1353–1359. doi: 10.1002/jmv.23271. [DOI] [PubMed] [Google Scholar]
- 521.Svensson A., Tunback P., Nordstrom I., Padyukov L., Eriksson K. Polymorphisms in Toll-like receptor 3 confer natural resistance to human herpes simplex virus type 2 infection. J. Gen. Virol. 2012;93(Pt 8):1717–1724. doi: 10.1099/vir.0.042572-0. [DOI] [PubMed] [Google Scholar]
- 522.Sironi M., Biasin M., Cagliani R., Forni D., De Luca M., Saulle I., Lo Caputo S., Mazzotta F., Macias J., Pineda J.A., Caruz A., Clerici M. A common polymorphism in TLR3 confers natural resistance to HIV-1 infection. J. Immunol. 2012;188(2):818–823. doi: 10.4049/jimmunol.1102179. [DOI] [PubMed] [Google Scholar]
- 523.Citores M.J., Banos I., Noblejas A., Rosado S., Castejon R., Cuervas-Mons V. Toll-like receptor 3 L412F polymorphism may protect against acute graft rejection in adult patients undergoing liver transplantation for hepatitis C-related cirrhosis. Transplant. Proc. 2011;43(6):2224–2226. doi: 10.1016/j.transproceed.2011.05.011. [DOI] [PubMed] [Google Scholar]
- 524.Ueta M., Sotozono C., Inatomi T., Kojima K., Tashiro K., Hamuro J., Kinoshita S. Toll-like receptor 3 gene polymorphisms in Japanese patients with Stevens-Johnson syndrome. Br. J. Ophthalmol. 2007;91(7):962–965. doi: 10.1136/bjo.2006.113449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 525.Misch E.A., Hawn T.R. Toll-like receptor polymorphisms and susceptibility to human disease. Clin. Sci. 2008;114(5):347–360. doi: 10.1042/CS20070214. (Lond.) [DOI] [PubMed] [Google Scholar]
- 526.Agnese D.M., Calvano J.E., Hahm S.J., Coyle S.M., Corbett S.A., Calvano S.E., Lowry S.F. Human toll-like receptor 4 mutations but not CD14 polymorphisms are associated with an increased risk of gram-negative infections. J. Infect. Dis. 2002;186(10):1522–1525. doi: 10.1086/344893. [DOI] [PubMed] [Google Scholar]
- 527.Balistreri C.R., Candore G., Lio D., Colonna-Romano G., Di Lorenzo G., Mansueto P., Rini G., Mansueto S., Cillari E., Franceschi C., Caruso C. Role of TLR4 receptor polymorphisms in Boutonneuse fever. Int. J. Immunopathol. Pharmacol. 2005;18(4):655–660. doi: 10.1177/039463200501800406. [DOI] [PubMed] [Google Scholar]
- 528.Arbour N.C., Lorenz E., Schutte B.C., Zabner J., Kline J.N., Jones M., Frees K., Watt J.L., Schwartz D.A. TLR4 mutations are associated with endotoxin hyporesponsiveness in humans. Nat. Genet. 2000;25(2):187–191. doi: 10.1038/76048. [DOI] [PubMed] [Google Scholar]
- 529.Tal G., Mandelberg A., Dalal I., Cesar K., Somekh E., Tal A., Oron A., Itskovich S., Ballin A., Houri S., Beigelman A., Lider O., Rechavi G., Amariglio N. Association between common Toll-like receptor 4 mutations and severe respiratory syncytial virus disease. J. Infect. Dis. 2004;189(11):2057–2063. doi: 10.1086/420830. [DOI] [PubMed] [Google Scholar]
- 530.Awomoyi A.A., Rallabhandi P., Pollin T.I., Lorenz E., Sztein M.B., Boukhvalova M.S., Hemming V.G., Blanco J.C., Vogel S.N. Association of TLR4 polymorphisms with symptomatic respiratory syncytial virus infection in high-risk infants and young children. J. Immunol. 2007;179(5):3171–3177. doi: 10.4049/jimmunol.179.5.3171. [DOI] [PubMed] [Google Scholar]
- 531.Lorenz E., Mira J.P., Frees K.L., Schwartz D.A. Relevance of mutations in the TLR4 receptor in patients with gram-negative septic shock. Arch. Intern. Med. 2002;162(9):1028–1032. doi: 10.1001/archinte.162.9.1028. [DOI] [PubMed] [Google Scholar]
- 532.Netea M.G., Wijmenga C., O'Neill L.A. Genetic variation in Toll-like receptors and disease susceptibility. Nat. Immunol. 2012;13(6):535–542. doi: 10.1038/ni.2284. [DOI] [PubMed] [Google Scholar]
- 533.Wlasiuk G., Khan S., Switzer W.M., Nachman M.W. A history of recurrent positive selection at the toll-like receptor 5 in primates. Mol. Biol. Evol. 2009;26(4):937–949. doi: 10.1093/molbev/msp018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 534.Hawn T.R., Verbon A., Lettinga K.D., Zhao L.P., Li S.S., Laws R.J., Skerrett S.J., Beutler B., Schroeder L., Nachman A., Ozinsky A., Smith K.D., Aderem A. A common dominant TLR5 stop codon polymorphism abolishes flagellin signaling and is associated with susceptibility to legionnaires' disease. J. Exp. Med. 2003;198(10):1563–1572. doi: 10.1084/jem.20031220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 535.Hawn T.R., Scholes D., Li S.S., Wang H., Yang Y., Roberts P.L., Stapleton A.E., Janer M., Aderem A., Stamm W.E., Zhao L.P., Hooton T.M. Toll-like receptor polymorphisms and susceptibility to urinary tract infections in adult women. PLoS One. 2009;4(6) doi: 10.1371/journal.pone.0005990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 536.Tantisira K., Klimecki W.T., Lazarus R., Palmer L.J., Raby B.A., Kwiatkowski D.J., Silverman E., Vercelli D., Martinez F.D., Weiss S.T. Toll-like receptor 6 gene (TLR6): single-nucleotide polymorphism frequencies and preliminary association with the diagnosis of asthma. Genes Immun. 2004;5(5):343–346. doi: 10.1038/sj.gene.6364096. [DOI] [PubMed] [Google Scholar]
- 537.Schott E., Witt H., Neumann K., Taube S., Oh D.Y., Schreier E., Vierich S., Puhl G., Bergk A., Halangk J., Weich V., Wiedenmann B., Berg T. A Toll-like receptor 7 single nucleotide polymorphism protects from advanced inflammation and fibrosis in male patients with chronic HCV-infection. J. Hepatol. 2007;47(2):203–211. doi: 10.1016/j.jhep.2007.03.021. [DOI] [PubMed] [Google Scholar]
- 538.Wang C.H., Eng H.L., Lin K.H., Chang C.H., Hsieh C.A., Lin Y.L., Lin T.M. TLR7 and TLR8 gene variations and susceptibility to hepatitis C virus infection. PLoS One. 2011;6(10) doi: 10.1371/journal.pone.0026235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 539.Bochud P.Y., Hersberger M., Taffe P., Bochud M., Stein C.M., Rodrigues S.D., Calandra T., Francioli P., Telenti A., Speck R.F., Aderem A. Polymorphisms in Toll-like receptor 9 influence the clinical course of HIV-1 infection. AIDS. 2007;21(4):441–446. doi: 10.1097/QAD.0b013e328012b8ac. (London, England) [DOI] [PubMed] [Google Scholar]
- 540.Medvedev A.E. Toll-like receptor polymorphisms, inflammatory and infectious diseases, allergies, and cancer. J. Interf. Cytokine Res. 2013;33(9):467–484. doi: 10.1089/jir.2012.0140. [DOI] [PMC free article] [PubMed] [Google Scholar]