

Abstract
Recombinant fragments of S proteins from the Severe Acute Respiratory Syndrome (SARS) coronavirus (SARA-CoV) were generated and used in a Western blot (WB) assay that was compared to a commercial SARS ELISA method. In 85% of confirmed SARS cases (n = 20), the S2 recombinant fragment based WB was positive and this was comparable to the commercial ELISA using heat killed SARS-CoV. WB using the other four recombinant fragments in confirmed SARS cases generated lower rates of detection (S1—75%, S1-N—25%, S1-C—55%). Evaluation of sera from healthy controls (n = 60) resulted in two weakly positive ELISA results with the remainder being negative while the S2 protein WB demonstrated three positive results from the 20 controls with a history of SARS contact and no positive results in 40 noncontact controls. A discrepancy between the ELISA and S2 WB arose when evaluating per-2003 sera from individuals (n = 10) with SARS-like symptoms (ELISA—100% positive, S2 WB—30% positive). These data suggest that the S2 WB assay may be particularly useful in ELISA-negative SARS cases and in some ELISA-positive non-SARS cases.
Keywords: SARS, ELISA, WB, Antibody, Immune response
Introduction
Severe Acute Respiratory Syndrome (SARS) was detected in China during the spring of 2004 [1]. A total of nine cases (including three suspected cases) were reported and a female patient died. SARS is caused by a new strain of human coronavirus (CoV) that has been isolated from SARS patients [2], [3]. Unlike other human CoVs, which cause mild respiratory infection [4], [5], [6], [7] comprising approximately 15–30% of common colds [8], [9], the SARS-CoV causes severe respiratory infection, which progresses to acute lung injury or acute respiratory distress syndrome (ARDS) and the mortality around 10% [10]. In the early stage of SARS or in atypical SARS, patients usually demonstrate symptoms similar to the common cold or influenza. In this situation, the detection of antibodies against SARS-CoV has been used to confirm the diagnosis of SARS. It is also important to analyze the immunogens in SARS-CoV and their capacity to elicit protective immunity, to better characterize the immune response for early diagnosis, and to understand the immunopathogenesis of the SARS.
The spike (S) protein from the SARS-CoV is a heterodimer, a characteristic of coronaviruses, and consists of two noncovalently bound S protein subunits, S1 and S2, derived from the N and C terminal halves of the S protein. During a coronavirus infection, the S1 subunit binds to its specific receptors on host cells [11], [12], followed by the S2 subunit mediating fusion between the virus and host cell membrane leading to the entry of virus into the cells. In previous studies of coronavirus infection, the S protein is a potent immunogen, which induces neutralizing antibodies [13]. After infection, neutralizing antibodies are typically directed against certain fragments of S1 or S2, which can block the receptor recognition and membrane fusion of the coronavirus [14]. Recent reports showed that the viral receptor, ACE2, binding site was linked to the fragment of the C terminal of S1 and S2 [15], [16], and this suggests that antibodies against S1-C and S2 may be associated with the generation of protective immune response.
In order to understand the immunogenecity of fragments of the S protein of SARS-CoV and to evaluate their potential use as target antigens for a diagnostic ELISA assay. We prepared recombinant S1 and S2 subunits and S1 fragments (Fig. 1 ) to evaluate the serum from SARS-recovered patients for possible antibodies to these antigens.
Fig. 1.
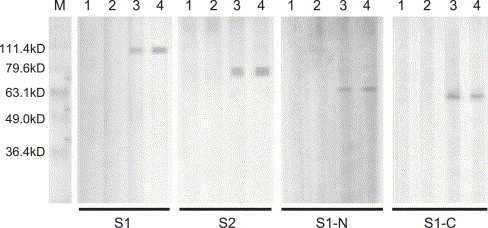
Western blotting analysis of expressed recombinant SARS virus protein. M: protein MW marker, the molecule weights of S1, S1-C, S1-N, and S2 recombinant proteins are 108, 64, 59, and 79.9 kDa, respectively. The numbers of patients are showed above the figures of W.B. 1 and 2: sera from health donor (with SARS contact history); 3 and 4: sera from patients recovered from SARS.
Materials and methods
Patients and sera
In order to compare the utility of ELISA and WB assay with the two recombinant fragments of S protein, we used sera obtained from SARS patient samples at The First Hospital of Peking University. The sera were collected from a group of medical workers who were infected with SARS-CoV when they cared for SARS patients (Table 1 ). All were diagnosed with SARS by following the diagnostic standards including a clear SARS-CoV contact history, fever, and X-ray changes. All of these cases emerged at approximately the same time, the end of April 2003. The control sera from healthy donors come from three groups, 20 samples collected prior to the outbreak of SARS, 20 samples collected from medical workers in The First Hospital of Peking University during the epidemic period of SARS, and 20 samples collected from the healthy donors who are medical workers in The First Hospital of Peking University and had occasionally contacted the SARS patients.
Table 1.
The data of patients studied
| Number | Age | Gender | Date of fever beginninga | Date of leaving hospitalb | Date of sample collectingc | ELISA | S1 | S2 | S1-N | S1-C |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 21 | Female | 04/23 | 06/01 | 06/25 | +(1.667) | + | + | − | + |
| 2 | 28 | Male | 04/23 | 06/01 | 06/25 | +(1.437) | + | + | − | − |
| 3 | 21 | Female | 04/21 | 06/01 | 06/25 | +(1.658) | + | + | − | + |
| 4 | 24 | Female | 05/03 | 06/01 | 06/25 | +(1.624) | − | − | − | − |
| 5 | 28 | Male | 05/10 | 06/01 | 06/25 | +(0.779) | + | + | + | − |
| 6 | 47 | Female | 05/03 | 06/01 | 06/25 | +(0.726) | − | + | − | − |
| 7 | 34 | Female | 04/23 | 06/01 | 06/25 | −(0.007) | − | − | − | − |
| 8 | 22 | Female | 04/18 | 06/01 | 06/25 | +(1.432) | + | − | − | − |
| 9 | 30 | Female | 05/03 | 06/01 | 06/25 | −(0.045) | − | + | − | − |
| 10 | 23 | Female | 04/23 | 06/01 | 06/25 | +(1.210) | + | + | − | − |
| 11 | 33 | Female | 04/29 | 06/01 | 06/25 | +(1.587) | + | + | + | + |
| 12 | 21 | Female | 04/23 | 06/01 | 06/25 | +(1.658) | + | + | − | + |
| 13 | 23 | Female | 03/24 | 06/01 | 06/25 | +(1.680) | + | + | − | + |
| 14 | 40 | Female | 04/24 | 06/01 | 06/25 | +(1.420) | + | + | − | + |
| 15 | 25 | Female | 04/23 | 06/01 | 06/25 | +(1.709) | + | + | − | + |
| 16 | 39 | Female | 04/23 | 06/01 | 06/25 | −(0.068) | − | + | + | − |
| 17 | 38 | Female | 04/29 | 06/01 | 06/25 | +(1.394) | + | + | − | + |
| 18 | 20 | Female | 04/27 | 06/01 | 06/25 | +(1.575) | + | + | − | + |
| 19 | 52 | Female | 04/26 | 06/01 | 06/25 | +(1.457) | + | + | + | + |
| 20 | 21 | Female | 04/29 | 06/01 | 06/25 | +(1.287) | + | + | + | + |
All patients in our study are medical workers exposed to SARS during their work.
We used the date (MM/DD) of fever initiation as the date of the SARS initiation.
In this study, we used the date of leaving the hospital as the date of SARS recovery.
After leaving from the hospital, all patients had 3 weeks of medical observation period and we collected the samples at the end of this isolation period. Antibodies in serum were detected with ELISA kit (OD value at 450 nm), value of cutoff (CO) is 0.135.
In order to test the utility of the WB with recombinant S2 subunit, 31 serum samples from SARS-suspected patients (12 from Princess Margaret Hospital, Hong Kong, and 19 from Beijing SARS control network laboratories) and 10 serum samples from patients with SARS-similar fever (all collected before 2003, stored at −80°C, and found seropositive when evaluated with a SARS ELISA kit).
The patients gave written consent for blood collection and Hospital Ethics Review Committee approved this study. Testing for antibodies against GST or thioredoxin (TRX) was negative in all of the serum samples used in this study, and all of the samples were stored at −80°C before evaluation.
Expression of recombinant SARS-CoV S fragments and Western blot
Four fragments of SARS-CoV S protein, S1 (a TRX fusion recombinant protein of S1 subunit), S2 (a recombinant protein of S2 subunit with 6X His tail), S1-N (a GST fusion recombinant protein of the 344 amino acid residues of S1 N terminal), and S1-C (a GST fusion recombinant protein of the 311 amino acid residues of S1-C terminal) were expressed in Escherichia coli as described previously [17]. Briefly, each expressed fragment of S protein was amplified with the template of plasmid containing corresponding full-length cDNA of SARS-CoV S gene (Gifts from Dr. Yong Xie, Hong Kong University of Science and Technology) and specific primers (Table 2 ). The amplified products were digested with restriction endonuclease BamHI and KpnI. The fragments of S1 and S2 cDNAs were inserted into the pET32a-TRX and pET32a expression plasmids, respectively, whereas the fragments of S1-N and S1-C cDNAs were inserted into the pGEX-4T-2. After verification of the correct sequences, the plasmids were transformed into E. coli BL-21 (pET32a-TRX/S1 and pET32a/S2) and DH5α (pGEX-4T-2/S1-N and pGEX-4T-2/S1-C), respectively. The E. coli was cultured in 2-YT medium at 37° and induced with IPTG for 6 h. The E. coli was harvested and lysed by ultrasonication. The inclusion bodies (IB) were collected and thoroughly washed. The recombinant S1-TRX fusion protein and S2 protein, both of which contain a 6xHis tag, were purified with Ni-NTA resin (Qiagen QIAexpress). As the S1-N and S1-C proteins were fused with GST proteins on N terminus, they were purified with GST-glutathione sepharose 4B affinity system.
Table 2.
The sequences of primers for amplification of S fragments
| Fragment | Sense | Antisense |
|---|---|---|
| S1 | 5′-ggggatccagtgaccttgaccggtgcaccac-3′ | 5′-ccggtaccaactgtatggtaactagcacaaatgcc-3′ |
| S2 | 5′-ggggatcctctttattacgtagtactagcc-3′ | 5′-ccggtacctgtgtaatgtaatttgacacc-3′ |
| S1-N | 5′-ggggatcctttattttcttattatttcttactc-3′ | 5′-ccggtaccaactgtatggtaactagcacaaatgc-3′ |
| S1-C | 5′-ggggatcctcaacatttttttcaacctttaagtgc-3′ | 5′-ccggtacctgtatggtaactagcacaaatgccagc-3′ |
Western blotting
The screening of the S protein of SARS-CoV against sera from SARS patients or from healthy donors was performed by Western blotting (WB). Briefly, every purified SARS recombinant protein was separated on 10% SDS-polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes. After blocking with 5% milk, the membranes were sequentially incubated with sera diluted in 1:500, goat anti-human IgG (H + L) alkaline phosphatase conjugate (1:10,000, Promega), and the substrate NBT/BCIP (Promega). The antibody-positive sera were tested twice to verify the immunoreaction. Mock-transfected E. coli lysate made in the same vectors were used in WB analysis as negative controls.
ELISA
IgG antibodies against sera from SARS patients were detected by ELISA using ELISA kit (Huada Bioscience, China), which was made up with the killed SARS virus and served as the main antigens. In brief, 100 μl/well of diluted serum (1:200) was added and incubated for 2 h at room temperature (RT). Plates were washed, followed by the addition of 100 μl/well diluted goat anti-human IgG (H + L) alkaline phosphatase conjugate (1:10,000, Promega) for 1 h at RT. Plates were washed, incubated with 100 μl/well of substrate solution for 30 min at RT, and immediately read with a microtiter plate reader (Anthos 2001, Anthos, Austria) at 450 nm. Samples with an optical density (OD) over the cutoff (CO) value were identified as positive. CO value was defined as CO value = [0.13 + NOD], and this translated into 0.135 for IgG in our study (NOD < 0.005, so the CO was set at 0.135).
Results
Anti-SARS-CoV-specific antibodies could be detected in serum by WB with recombinant SARS-CoV S protein fragments as well as by ELISA with killed SARS-CoV
The recombinant S protein fragments were recognized by the sera from SARS-recovered donors, and S1, S2, S1-N, and S1-C recombinant proteins were estimated to be around 108 kDa (TRX-S1), 90 kDa (S2), 64 kDa (S1-N-GST), and 59 kDa (S1-C-GST), respectively (Fig. 1). Seroreactivity analysis was performed using WB and ELISA. In the 20 sera collected from the donors who have fully recovered from SARS infection, there were 17 sera reactive to killed SARS virus as measured by an ELISA kit (Table 1). In these 20 sera from SARS-recovered donors, there were 15, 17, 11, and 5 sera reactive to S1, S2, S1-N, and S1-C recombinant proteins, respectively, as detected by WB (details in Table 1). None of the sera from control healthy donors was reactive using the killed SARS virus ELISA kit, with the exception of a very weak reaction found in two samples from sera collected prior to the outbreak of SARS (OD 0.172 and 0.136, respectively). Antibodies against S1 and S1-N were also detected by WB in sera from the three groups of healthy donors (the positive rate is shown in Table 2). In contrast, antibodies against S1-C and S2 fragments by WB were only detected in the sera from the healthy donors with SARS contact history, but not in the sera from the other healthy donors (Table 3 ).
Table 3.
The antibody response to S antigen subunits in donors fully recovered from SARS infection and different groups of healthy donors
| SARS patients | Healthy donors with SARS patients contact history | Healthy donors during SARS epidemic | Healthy donors before SARS epidemic | |
|---|---|---|---|---|
| S1 | 75% (15/20) | 30% (6/20) | 10% (2/20) | 20% (4/20) |
| S2 | 85%(17/20) | 15% (3/20) | 0% (0/20) | 0% (0/20) |
| S1-N | 25% (5/20) | 10% (2/20) | 15% (3/20) | 20% (4/20) |
| S1-C | 55% (11/20) | 15% (3/20) | 0% (0/20) | 0% (0/20) |
| ELISA | 85% (17/20) | 0%(0/20) | 0%(0/20) | 10%(2/20)a |
The very weak reaction was found with ELISA kit in two samples in which OD were 0.172 and 0.136, respectively, and CO is 0.135.
WB with recombinant S2 subunit could be useful in diagnosis of SARS
Anti-SARS-CoV antibodies and anti-S2-subunit antibodies could be detected in sera from SARS-suspected patients from Hong Kong and Beijing by ELISA and WB, respectively (Table 4 ). The positive rate of the antibodies in these sera is less than that in sera from SARS patients in The First Hospital of Peking University who were diagnosed as SARS confirmed. Furthermore, in 10 selected ELISA-positive serum samples from patients with a compatible history of disease prior to 2003, antibodies against S2 recombinant protein were detected in three samples (Table 4).
Table 4.
The positive rate of serum from different donors detected by ELISA and WB
| Serum from Hong Konga | Serum from Beijingb | Serum from SARS-like patientsc | |
|---|---|---|---|
| S2 | 67% (8/12) | 37% (7/19) | 30% (3/10) |
| ELISA | 75% (9/12) | 53% (9/19) | 100% (10/10) |
Serum samples from Princess Margaret Hospital, Hong Kong.
Serum samples from Beijing SARS control network laboratories.
Serum samples from fever patients collected before 2003.
Discussion
In order to evaluate the diagnostic utility of a WB assay using recombinant S protein fragments, sera from 20 SARS-recovered donors were tested in this study. All patients were diagnosed as SARS cases and infected in the same time interval. The samples were collected at the same time to avoid any possible error caused by sample collection differences. In the 85% of these serum samples, antibodies against SARS-CoV were detected with a commercial ELISA assay were also positive using the WB with recombinant S2 protein that we developed. Antibodies were not detected using either test method in the healthy control sera with the exception of two weakly reactive samples. Furthermore, only serum from one patient was negative with both the commercial ELISA and WB based on the S2 recombinant protein. This suggests that the commercial ELISA kit for SARS combined with the WB assay using the recombinant S2 subunit may provide improved diagnostic capability.
Antibodies against S1 and S1-N were detected by WB in sera both from SARS-recovered donors and all the control groups. Thus, it appears that antibodies directed against S1 or S1-N are less useful for making a diagnosis of SARS. On the other hand, antibodies against S1-C and S2 by WB were detected only in sera from SARS-recovered donors and some donors with SARS contact history. In some commercial ELISA-negative serum samples, antibodies to S1-C and S2 by WB were positive possibly due to the destruction or alteration of SARS-CoV surface protein during the viral preparation process for the ELISA kits. This suggests that detecting antibodies to S1-C or S2 by WB may be useful in making a diagnosis of SARS exposure in the setting of a negative SARS ELISA assay.
However, clinicians should cautious in interpreting a weakly positive ELISA result (OD < 0.5). This could represent an antibody to a non-SARS-CoV that cross-reacts with the SARS-CoV antigen preparation in the ELISA kit, a phenomenon that has been reported previously [18], [19]. As shown above, recombinant S1 and its fragments as a target for antibody detection do not appear to provide additional useful diagnostic information. However, 10 ELISA-positive (OD < 0.5) sera were identified from patients collected before SARS outbreak in 2003 with symptoms that included fever. These sera were tested using the WB assay with recombinant S2 subunit and three of these were positive. Additional testing with the recombinant S2 protein are required to determine if the WB to recombinant S2 protein will prove to be useful in evaluating ELISA-positive sera from patients who do not fulfill the criteria for diagnosing SARS.
In order to evaluate the utility of the WB assay with recombinant S2 subunit in aiding in the diagnosis of SARS, sera from SARS-suspected patients were tested with both the S2-WB and a commercial SARS ELISA assay. Antibodies against SARS-CoV were detected in some of the serum samples from two different groups of suspected SARS patients using the combination of SARS ELISA and S2-WB assays. As might be expected, the positive rate in these serum samples was lower than that found in SARS-confirmed cases obtained from The First Hospital of Peking University. This may be due to non-SARS patients being present within these two groups and/or the absence of a significant antibody response early in the course of SARS. If the latter circumstance is correct, then the evaluation of the diagnostic effectiveness of a test method including ELISA or WB should be established using sera from SARS-confirmed cases rather than SARS-suspected cases. The time course of antibody generation to SARS-CoV following the onset of symptoms is an important diagnostic issue and has been previously reported to vary widely [20]—from 3 days to more than 1 month following the onset of fever (data not shown).
SARS-CoV recombinant proteins expressed in E. coli have been shown to be recognized by antibodies in the serum from coronavirus-infected animals [21], [22]. Furthermore, recombinant coronavirus proteins expressed in E. coli has been demonstrated to have antigenicity and receptor binding ability [23], [24]. Our data suggest that the recombinant S1-C and S2 may have potential use in the production of SARS vaccines. This is based in part on the observation that antibodies against S protein were only detected rarely in samples from healthy donors who had been in contact with SARS patients, suggesting that infection and/or immune response may not be consistently generated by common contact. This could explain why the researchers are infected even after long-term contact with SARS-CoV prior to infection.
In conclusion, our findings suggest that diagnostic approaches using SARS-CoV-specific component proteins may increase the diagnostic accuracy of testing when combined with conventional ELISA tests based on killed virus as the target antigen.
Acknowledgments
This work is supported by Ministry of Education, China, National Nature Science Foundation of China (NSFC, No. 30340011), and Ministry of Science and Technology, China (No. 2003CB514100), for funding this research work.
Contributor Information
Yue-Dan Wang, Email: wangyuedan@hotmail.com.
Wei Feng Chen, Email: wfchen@public.bta.net.cn.
References
- 1.Normile D. Mounting lab accidents raise SARS fears. Science. 2004;304:659–661. doi: 10.1126/science.304.5671.659. [DOI] [PubMed] [Google Scholar]
- 2.Marra M.A., Jones S.J., Astell C.R., Holt R.A., Brooks-Wilson A., Butterfield Y.S., Khattra J., Asano J.K., Barber S.A., Chan S.Y., Cloutier A., Coughlin S.M., Freeman D., Girn N., Griffith O.L., Leach S.R., Mayo M., McDonald H., Montgomery S.B., Pandoh P.K., Petrescu A.S., Robertson A.G., Schein J.E., Siddiqui A., Smailus D.E., Stott J.M., Yang G.S., Plummer F., Andonov A., Artsob H., Bastien N., Bernard K., Booth T.F., Bowness D., Czub M., Drebot M., Fernando L., Flick R., Garbutt M., Gray M., Grolla A., Jones S., Feldmann H., Meyers A., Kabani A., Li Y., Normand S., Stroher U., Tipples G.A., Tyler S., Vogrig R., Ward D., Watson B., Brunham R.C., Krajden M., Petric M., Skowronski D.M., Upton C., Roper R.L. The genome sequence of the SARS-associated coronavirus. Science. 2003;300:1399–1404. doi: 10.1126/science.1085953. [DOI] [PubMed] [Google Scholar]
- 3.Holmes K.V. SARS-associated coronavirus. N. Engl. J. Med. 2003;348:1948. doi: 10.1056/NEJMp030078. [DOI] [PubMed] [Google Scholar]
- 4.Wentworth D.E., Holmes K.V. Molecular determinants of species specificity in the coronavirus receptor aminopeptidase N (CD13): influence of N-linked glycosylation. J. Virol. 2001;75:9741. doi: 10.1128/JVI.75.20.9741-9752.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lachance C., Arbour N., Cashman N.R., Talbot P.J. Involvement of aminopeptidase N (CD13) in infection of human neural cells by human coronavirus 229E. J. Virol. 1998;72:6511. doi: 10.1128/jvi.72.8.6511-6519.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Stewart J.N., Mounir S., Talbot P.J. Human coronavirus gene expression in the brains of multiple sclerosis patients. Virology. 1992;191:502. doi: 10.1016/0042-6822(92)90220-J. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Talbot P.J., Ekande S., Cashman N.R., Mounir S., Stewart J.N. Neurotropism of human coronavirus 229E. Adv. Exp. Med. Biol. 1993;342:339. doi: 10.1007/978-1-4615-2996-5_52. [DOI] [PubMed] [Google Scholar]
- 8.Holmes K.V., Lai M.M. Coronaviridae: the viruses and their replication. In: Fields B.N., Knipe D.M., Howley P.M., editors. Fields Virology. third ed. Lippincott-Raven; Philadelphia, PA: 1996. pp. 1075–1094. [Google Scholar]
- 9.Vabret A., Mourez T., Gouarin S., Petitjean J., Freymuth F. An outbreak of coronavirus OC43 respiratory infection in Normandy, France. Clin. Infect. Dis. 2003;36:985. doi: 10.1086/374222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lew T.W., Kwek T.K., Tai D., Earnest A., Loo S., Singh K., Kwan K.M., Chan Y., Yim C.F., Bek S.L., Kor A.C., Yap W.S., Chelliah Y.R., Lai Y.C., Goh S.K. Acute respiratory distress syndrome in critically ill patients with severe acute respiratory syndrome. JAMA. 2003;290:374. doi: 10.1001/jama.290.3.374. [DOI] [PubMed] [Google Scholar]
- 11.Kubo H., Yamada Y.K., Taguchi F. Localization of neutralizing epitopes and the receptor-binding site within the amino-terminal 330 amino acids of the murine coronavirus spike protein. J. Virol. 1994;68:5403–5410. doi: 10.1128/jvi.68.9.5403-5410.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Taguchi F. The S2 subunit of the murine coronavirus spike protein is not involved in receptor binding. J. Virol. 1995;69:7260–7263. doi: 10.1128/jvi.69.11.7260-7263.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Collins A., Knobler R.L., Powell H., Buchmeier M.J. Monoclonal antibodies to murine hepatitis virus-4 (strain JHM) define the viral glycoprotein responsible for attachment and cell fusion. Virology. 1982;119:358–371. doi: 10.1016/0042-6822(82)90095-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Godet M., Grosclaude J., Delmas B., Laude H. Major receptor binding and neutralization determinants are located within the same domain of the transmissible gastroenteritis virus (coronavirus) spike protein. J. Virol. 1984;68:8008–8016. doi: 10.1128/jvi.68.12.8008-8016.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Xiao X., Chakraborti S., Dimitrov A.S., Gramatikoff K., Dimitrov D.S. The SARS-CoV S glycoprotein: expression and functional characterization. Biochem. Biophys. Res. Commun. 2003;312:1159–1164. doi: 10.1016/j.bbrc.2003.11.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Babcock G.J., Esshaki D.J., Thomas W.D., Jr., Ambrosino D.M. Amino acids 270 to 510 of the severe acute respiratory syndrome coronavirus spike protein are required for interaction with receptor. J. Virol. 2004;8(9):4552–4560. doi: 10.1128/JVI.78.9.4552-4560.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Yang X.A., Dong X.Y., Li Y., Li Y.Y., Xu G.B., Feng Z.R., Wang Y.D., Chen W.F. Expression and purification of recombinant N-terminal protein of SARS S1 subunit expressed in E. coli Beijing. Da Xue Xue Bao. 2004;36(1):79–81. [PubMed] [Google Scholar]
- 18.Li L.H., Shi Y.L., Li P., Xu D.X., Wan G.P., Gu X.Q., Zhang X.L., Ma Q.J., Cao C. Detection and analysis of SARS coronavirus-specific antibodies in sera from non-SARS children. Di Yi Jun Yi Da Xue Xue Bao. 2003;23(10):1085–1087. [PubMed] [Google Scholar]
- 19.Stavrinides J., Guttman D.S. Mosaic evolution of the severe acute respiratory syndrome coronavirus. J. Virol. 2004;78(1):76–82. doi: 10.1128/JVI.78.1.76-82.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liu X., Shi Y., Li P., Li L., Yi Y., Ma Q., Cao C. Profile of antibodies to the nucleocapsid protein of the severe acute respiratory syndrome (SARS)-associated coronavirus in probable SARS patients. Clin. Diagn. Lab. Immunol. 2004;11(1):227–228. doi: 10.1128/CDLI.11.1.227-228.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wang C.H., Hong C.C., Seak J.C. An ELISA for antibodies against infectious bronchitis virus using an S1 spike polypeptide. Vet. Microbiol. 2002;85:333–342. doi: 10.1016/s0378-1135(01)00525-9. [DOI] [PubMed] [Google Scholar]
- 22.Suiter B.T., Pfeiffer N.E., Jones E.V., Reed A.P., Klepfer S.R., Miller T.J., Srikumaran S. Serological recognition of feline infectious peritonitis virus spike gene regions expressed as synthetic peptides and E. coli fusion protein. Arch. Virol. 1995;140:687–702. doi: 10.1007/BF01309958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bosch B.J., van der Zee R., de Haan C.A., Rottier PJ. The coronavirus spike protein is a class I virus fusion protein: structural and functional characterization of the fusion core complex. J. Virol. 2003;77:8801–8811. doi: 10.1128/JVI.77.16.8801-8811.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ho T.Y., Wu S.L., Cheng S.E., Wei Y.C., Huang S.P., Hsiang C.Y. Antigenicity and receptor-binding ability of recombinant SARS coronavirus spike protein. Biochem. Biophys. Res. Commun. 2004;313:938–947. doi: 10.1016/j.bbrc.2003.11.180. [DOI] [PMC free article] [PubMed] [Google Scholar]