

Highlights
-
•
Human DC subsets can exert tumoricidal activity.
-
•
Killer DCs exploit several mechanisms for direct killing of target cells, including TRAIL and granzyme B.
-
•
Antigen presentation and/or IFN production are important additional effector functions.
-
•
Killer DCs are promising targets for immunotherapeutic strategies.
Keywords: plasmacytoid dendritic cells, myeloid dendritic cells, cytotoxicity, TRAIL, granzyme B, antitumor therapy
Abstract
Dendritic cells (DCs) are a family of professional antigen-presenting cells (APCs) that are able to initiate innate and adaptive immune responses against pathogens and tumor cells. The DC family is heterogeneous and is classically divided into two main subsets, each with its unique phenotypic and functional characteristics: myeloid DCs (mDCs) and plasmacytoid DCs (pDCs). Recent results have provided intriguing evidence that both DC subsets can also function as direct cytotoxic effector cells; in particular, against cancer cells. In this review, we delve into this understudied function of human DCs and discuss why these so-called killer DCs might become important tools in future cancer immunotherapies.
DCs: commanders-in-chief of the immune army
Forty years after their discovery by Zanvil Cohn and Ralph Steinman, DCs continue to fascinate and intrigue immunologists. Although the nomenclature of the DC system is still evolving and novel markers to identify and subclassify DCs are being continually identified, it is well established that the DC family constitutes a heterogeneous group of cells that can be categorized in two main subtypes: mDCs and pDCs. Despite the considerable heterogeneity between the different DC types in terms of phenotype, gene expression profile, and function, a common characteristic of all DCs is their capacity for antigen presentation and their unique ability to prime and activate naïve T lymphocytes. As the primary APCs of the immune system, DCs are pivotal in eliciting adaptive immune responses and, as such, in determining the balance between immunity and tolerance [1]. In addition to their central role in adaptive immunity, DCs also occupy a pre-eminent place within the innate immune system. In this context, DCs express Toll-like receptors (TLRs); a family of innate immune receptors involved in sensing viruses and other microbial stimuli. DCs are also capable of activating other innate immune cells, including natural killer (NK) cells.
Given their key role in regulating innate and adaptive immunity, DCs are critical for the induction of antitumor immunity [1]. Through their role in the induction of antigen-specific cytotoxic T lymphocytes (CTLs) and through their capacity to harness the cytotoxic activity of innate immune cells (NK cells, NKT cells, and γδ T cells), DCs can elicit potent cytotoxic immune responses towards tumor cells [2]. Evidence from animal and human studies indicates that DCs themselves can initiate cytotoxic effector function through which they directly contribute to tumor cell killing. These so-called killer DCs were first described in the mid-1990s, when a population of murine DCs was identified with the capacity to lyse CD4+ T cells in a FAS−FAS ligand (FAS-L)-dependent fashion [3]. Almost one decade later, three research groups independently reported on the existence of a novel DC type within the murine immune system that bore phenotypic, molecular, and functional characteristics of both DCs and NK cells 4, 5, 6. This DC subtype was termed natural killer dendritic cell (NKDC) or, alternatively, interferon-producing killer DC (IKDC) [4] because of its NK cell-like properties such as cytotoxic activity and capacity to produce high amounts of interferon (IFN)-γ 4, 5, 6. Although NKDCs were capable of antigen processing and presentation, it soon became apparent that this term was actually a misnomer because NKDCs belong to the NK cell lineage and not to the DC lineage 7, 8, 9. This apparently erroneous terminology has led to confusion and even skepticism over the actual existence of killer DCs.
Recent studies, however, have provided substantial evidence for direct cytotoxic effector function in DCs. These killer DCs, which have been identified in both rodents and humans, appear to constitute a heterogeneous population of cells that have the following characteristics in common: (i) they are endowed with direct cytolytic potential; (ii) they fulfill the phenotypic and functional criteria to be classified as bona fide DCs; and (iii) they cannot be defined as NK cells despite their cytotoxic activity (e.g., absence of classical NK cell surface markers, and target cell profile different from that of NK cells). Here, we summarize the findings accumulated in recent years on killer DCs and discuss the potential relevance of these cells to future immunotherapy strategies. No major differences in mode of action have been identified between rodent and human killer DCs, and because the existence and function of rodent killer DCs has been excellently reviewed elsewhere 10, 11, we focus this discussion on killer DCs in humans.
Killer DCs in humans
Monocytes and monocyte-derived DCs (MoDCs) as killer cells
Immature DCs reside mostly in parts of the body that are in close contact with the outside world, such as skin and mucosal tissue, and are able quickly to sense and take up pathogens that could harm the host. After pathogen recognition, the DCs mature and migrate to lymphoid tissues to present the pathogenic peptides to T cells. MoDCs are by far the most widely used cell type for the study of human DCs ex vivo (Table 1 ). Although a variety of protocols exist to generate MoDCs, the basic procedure consists of two phases: (i) monocytes are differentiated into immature DCs using a combination of cytokines [e.g., granulocyte–macrophage colony-stimulating factor (GM-CSF) and interleukin (IL)-4, IL-15 or IFNα]; and (ii) MoDC maturation is induced with activation stimuli (e.g., proinflammatory cytokines or TLR ligands). Immature DCs exploit a wide variety of pattern recognition receptors to recognize and take up antigens. Several of these differentiating and maturating agents can trigger human MoDCs to acquire a cytotoxic effector function. Different studies have suggested that this cytolytic potential already resides in the monocytic precursor cell compartment (Table 1). Indeed, human CD14+ as well as CD16+ monocytes stimulated with type I or II IFN, R848 a ligand for TLR7 and TLR8, or lipopolysaccharide (LPS) a ligand for TLR4, were shown to exert antitumor activity against a range of cancer cell lines 12, 13, 14, 15, 16, 17 (Table 1). In three of these studies, TNF-α-related apoptosis-inducing ligand (TRAIL) was implicated in the direct tumoricidal activity of human monocytes 12, 13, 15. TRAIL-dependent apoptosis has also been reported to play a role in the cytotoxic activity of human MoDCs. Viruses, including measles virus 18, 19, cytomegalovirus (CMV) [20], and HIV-1 21, 22, 23, TLR ligands (double-stranded RNA), type I IFNs 24, 25, 26, IL-15 [27], and the maturating agent CD40L [28], are all capable of inducing TRAIL expression in MoDCs (Table 1). Conversely, another study found that CD40 ligation inhibits TRAIL expression in MoDCs, but the cytotoxic ability of these CD40L-matured MoDCs was preserved, suggesting TRAIL-independent mechanisms for induction of cell death [28]. Indeed, as outlined in Table 1, both cell contact-dependent and -independent mechanisms have been implicated in MoDC-mediated killing and their cytolytic armamentarium includes, in addition to TRAIL, a broad range of cytotoxic effector molecules such as TNFα 21, 22, 23, 24, 29, 30, FAS-L 20, 21, 22, 29, 31, 32, caspase-8 19, 33, 34, 35, lymphotoxin (LT)-α1β2 [29], TNF-like weak inducer of apoptosis (TWEAK) [21], IFN-γ [28], granzyme B 27, 36, and programmed death ligand (PD-L1/2) 32, 37.
Table 1.
Human killer monocytes and MoDCsc
| Cell type | Stimulation | Target | E:T ratio | Killing mechanism | Refs |
|---|---|---|---|---|---|
| Monocytes | |||||
| Monocytes | HIV-1 | T cells | 1:1 | Contact dependent | [38] |
| Monocytes | IFNα, IFNγ | OVCAR3 (ovarian cancer) WM793 (melanoma) MDA231 (breast cancer) Colo205 (colon cancer) PC-3 (prostate cancer) H2126 (lung cancer) |
50:1 | TRAIL | [12] |
| Monocytes | IFNγ | HSC3 (oral squamous cell carcinoma) | 50:1 | TWEAK TRAIL |
[15] |
| Monocytes | LPS | K562 (leukemia) | 10:1 | No involvement of Fas-L, TNF-α or TGF-β | [14] |
| Monocytes | IFNα | K562 PC-3 Jurkat (T cell leukemia) |
? | TRAILa | [13] |
| CD16+ slan DCs | Unstimulated | Colo205 SkBr3 |
40:1 | ADCC by 17-1A (colo205) or by herceptin (SkBr3) Effector molecule: TNF-α |
[17] |
| CD16+ slan DCs M-DC8+ |
IFN-γ | Capan 1 (pancreatic cancer) MCF-7 Colo205 HT-29 (colon cancer) |
40:1 | ND | [16] |
| CD16+ slan DCs (CHC) | IMQ or resiquimod | Capan-1 HT-29 |
40:1 | ND | [77] |
| MoDCs | |||||
| MoDCs (measles) |
Measles | T cells (autologous) | 1:1 | TNF family member (postulated) | [19] |
| MoDCs | LPS | Jurkat Molt (T cell leukemia) K562 THP-1 (monocytic cancer) U937 (lymphoma) |
1:1 | Contact dependent | [78] |
| MoDCs | Measles virus | MDA231 | 50:1 | TRAIL | [18] |
| MoDCs | Immature | OVA.1 (ovarian cancer) SW626 (ovarian cancer) |
40:1 | FAS-L | [31] |
| MoDCs | Immature | SiHA, Caski (HPV+ cervical cancer) | 8:1 | Contact-dependent no involvement of TRAIL or FAS-L |
[79] |
| MoDCs CD14+, CD34+ |
IFNβ | HL60 (leukemia) | 20:1 | TRAILa | [25] |
| MoDCs | dsRNA → CD40L → |
MDA231 MDA231 |
50:1 50:1 |
TRAILa TNF-α |
[24] |
| MoDCs | CMV | CMV-reactive T cells | 1:53 | FAS-L, TRAIL | [20] |
| MoDCs | Immature | Jurkat Molt MCF-7 (breast cancer) U87 (glioblastoma) HCT-5 (colorectal cancer) A498 (renal cancer) 786.O (renal cancer) Caki.2 (renal cancer) |
10:1 | Caspase 8 (FADD independent) | [33] |
| MoDCs | CD40L | PCI-13 SSCHN (head and neck cancer) |
1:1 | TNF-α, LT-α, LT-β, FAS-L, TRAILa | [29] |
| MoDCs | CD40L, LPS | MCF-7 MDA-MB-468 (breast cancer) SK-BR-3 (breast cancer) |
? | TNF-αa Contact independent |
[30] |
| MoDCs | HIV-1 IFNα |
MDA231 CD4+ T cell line HIV-H9 CD4+ T cells (HIV-1 viremic patients) |
? | TRAILa, FAS-La, TNF-αa TWEAKa |
[21] |
| MoDCs | HIV-1 (nef) | CD8+ T cells | ? | sTNF-α FAS-L |
[22] |
| MoDCs | Immature | Jurkat | 10:1 | Caspase 8/Bcl-2 (FADD independent) | [35] |
| Cordblood MoDCs |
IFNγ → LPS → |
HL60, Jurkat Daudi, Jurkat |
20:1 | ND | [80] |
| MoDCs | IFNα | K562 | 20:1 | ND | [81] |
| MoDCs | HIV-1 (Vpr) + LPS | Allogeneic CD8+ T cells | 1:20 | sTNF-α | [23] |
| MoDCs | U251 (glioma) Jurkat |
20:1 | Contact dependent FADD and caspase-8 dependent |
[34] | |
| MoDCs | IFNα | K562 | 50:1 | TRAILa | [26] |
| MoDCs | OK432 | T2 K562 EJ 253J |
10:1 | Contact dependent (CD40–CD40L | [82] |
| MoDCs | Immature (CD123+) | U937 Jurkat HL-60 |
40:1 | TRAIL | [83] |
| MoDCs | CD40L | OSC-70 (oral squamous cell carcinoma) |
1:8 | TRAILa IFN-γ |
[28] |
| MoDCs (PB and CB) |
SARS coronavirus | ND | ND | TRAIL | [84] |
| MoDCs | LPS | SkBr3 (Her2-neu+ breast cancer) | 100:1 | ADCC by trastuzumab | [85] |
| MoDCs | LPS | MCF7 (breast) HeLa (cervix) HT29, HCT116, SW480 (colorectal) no killing of T lymphocytes |
5:1 | peroxynitrites | [86] |
| MoDCs (TBC) |
IFNα, LPS | CD4+ T cells CD8+ T cells |
10:1 | PD-L1 | [37] |
| MoDCs | IFNγ, LPS | T47D (breast cancer) | ND | ND | [87] |
| MoDCs | IL-15 | K562 | 50:1 | Granzyme B, TRAILa | [27] |
| MoDCs (CHC patients)b |
LPS | Allogeneic healthy CD4+ T cells Autologous CD4+ T cells |
4:1 | FAS-L, PD-L2, Contact dependent |
[32] |
| MoDCs | γ-irradiated HT-29 | HT-29 (colon cancer) | 20:1 | Perforin/granzyme B | [36] |
Partially dependent, other mechanisms (shown when possible) may be involved.
Chronic hepatitis C patients.
Abbreviations: ADCC, antibody-dependent cell-mediated cytotoxicity; CB, cord blood; E: T ratio, effector cell to target cell ratio; FADD, FAS-associated protein with death domain; HPV, human papillomavirus; ND, not determined; PB, peripheral blood; TGF, transforming growth factor.
The majority of studies on killer MoDCs have focused on their tumoricidal potential. MoDCs are capable of exerting cytotoxicity against a broad range of tumor cell lines, while sparing normal cells (Table 1). The reasons for this apparent tumor-selective action remain incompletely understood, although the expression of decoy receptors (e.g., TRAIL decoy receptors DR1 and DR2) and the activation of antiapoptotic mechanisms [e.g., upregulation of cellular FADD-like IL-1β-converting enzyme protease-inhibitory protein (c-FLIP)] may help to explain why normal cells are largely resistant to killing by MoDCs [25].
Despite their seemingly preferential tumor-directed action, under certain circumstances, MoDCs can induce T cell death 21, 32, 37. This observation was made in several studies (Table 1); all of which were performed in the context of infectious diseases 21, 32, 37. The first study demonstrated that MoDCs infected with measles virus can induce paracrine killing of autologous T cells [19]. Monocytes [38] and MoDCs [21] exposed to HIV-1 were capable of inducing apoptosis of HIV-1-infected as well as noninfected CD4+ T cells from either allogeneic or autologous origin. In another study, it was shown that LPS-matured MoDCs derived from tuberculosis patients had an increased expression of PD-L1, which underlined their antiproliferative and proapoptotic activity towards both CD4+ and CD8+ T cells [37]. Similarly, LPS-matured MoDCs generated from chronic hepatitis C patients were found to lyse both healthy (allogeneic) as well as patient-derived (autologous) CD4+ T cells in a PD-L2- and FAS-L-dependent fashion [32]. Taken together, these findings indicate that human monocytes and MoDCs, when appropriately stimulated, can function as cytotoxic antitumor effectors and, in the context of chronic infection, also as immunoregulatory cells with T cell killing activity.
Blood mDCs as killer cells
Although the observation that ex vivo generated MoDCs can act as direct killer cells is interesting, another relevant question is whether this is also applicable to naturally circulating DCs. Although most studies used MoDCs, several studies have delved into the killer function of blood DCs and have shown that the two main blood DC subsets, mDCs and pDCs, can be cytotoxic. The human blood mDC subset is usually defined as lineage (Lin)–HLA-DR+CD11c+CD123(IL-3Rα)dim cells. Blood mDCs can be further subdivided in nonoverlapping subsets based on the expression of blood dendritic cell antigen (BDCA)-1 (CD1c) and BDCA-3 (CD141) [39]. Classically, mDCs remain in an immature state and migrate from peripheral tissues to lymph nodes after maturation where they can activate T cells. Cytotoxic potential has hitherto been reported for the total CD11c+ mDC population, as well as for the CD1c+ subset (Table 2 ). The first description of the direct cytotoxic activity of human blood mDCs dates back to the late 1990s, where CD11c+ mDCs, stimulated with IFNα or IFNγ, directly lysed various tumor cell lines in a TRAIL-dependent fashion [40]. TRAIL has also been implicated in blood DC-mediated cytotoxicity in two other studies 41, 42 (Table 2). By contrast, TRAIL seems not to be an important mediator of cytotoxicity by TLR-activated mDCs. A recent study showed that neither BDCA-1+ nor BDCA-3+ mDCs produce TRAIL after exposure to TLR ligands [43]. In line with this, it was observed that tumor-infiltrating CD11c+ mDCs express and secrete perforin and granzyme B, but not TRAIL, upon TLR7 and TLR8 stimulation [44]. Granzyme B has also been implicated in mDC-mediated apoptosis in IL-15-activated CD11c+ mDCs [45]. Taken together, these data illustrate that killer mDCs, like their in vitro MoDC counterparts, can exploit a variety of cytotoxic effector mechanisms to exert killing function (Table 2). Similar to MoDCs, most studies on the killer function of native blood mDCs have been performed using tumor cell lines as target cells. The ability of blood mDCs to kill T cells was examined in one study that showed LPS-activated BDCA-1+ mDCs from chronic hepatitis C patients induced lysis of autologous patient T cells as well as allogeneic healthy T cells in a FAS-L- and PD-L2-dependent fashion (Table 2) [32]. This study provides evidence for the ability of native blood mDCs to kill T cells during chronic viral infection; a mechanism that may be exploited by viruses to escape antiviral T cells.
Table 2.
Human killer DCs divided into major subsetsc
| Subset of DC | Stimulation | Target | E:T ratio | Killing mechanism | Refs |
|---|---|---|---|---|---|
| mDCs | |||||
| CD11c+ blood mDCs | IFNα, IFNγ | Jurkat OVCAR3 PC-3 WM793 |
50:1 | TRAIL | [40] |
| CD11c+ blood mDCs | IFNγ, IL-15, LPS |
MCF-7 HBL-100 (breast cancer) MDA-MB-231 (breast cancer) MDA-MB-415 (breast cancer) |
10:1 | ND | [88] |
| CD11c+ mDCs | IMQ | K562 | 25:1 | Perforin/granzyme B | [44] |
| CD1c+ mDCs (CHC patients)b | Unstimulated | K562 U937 Jurkat |
50:1 | TRAIL | [42] |
| CD11c+ blood mDCs | IL-15 | Human aortic endothelial cells Porcine aortic endothelial cells |
10:1 | Granzyme Ba /caspase 8 |
[45] |
| Blood mDCs BDCA-1+ |
LPS | Allogeneic healthy CD4+ T cells Autologous CD4+ T cells |
4:1 | FAS-L PD-L2 Contact dependent |
[32] |
| pDCs | |||||
| pDC cell line GEN2.2 Blood pDCs → |
Influenza virus, Type I IFNs Influenza virus |
A549 (epithelial cancer) A549 |
25:1 15:1 |
TRAILa TRAIL |
[49] |
| Blood pDCs | IMQ | Jurkat | 25:1 | TRAIL | [44] |
| Blood pDCs | HIV-1 | SupT1 (CD4+ T cell line) | 20:1 | TRAILa | [56] |
| Blood pDCs | HIV-1 | CD4+ T cells (HIV-1 viremic patients) | 10:1 | TRAIL IFNα |
[89] |
| Blood pDCs | IL-3/CD40L → CpG → |
K562 K562 1806 (breast cancer) Colo829 (melanoma) |
100:1 | Contact dependent ND |
[52] |
| Blood pDCs | Flu CpG |
Jurkat J32 | Culture Sup |
TRAIL | [90] |
| Blood pDCs | HTLV-1 | DR5+ T cells | 1:2 | TRAIL | [55] |
| Blood pDCs | HIV | HIV-infected Sup-T1 cell line | 10:1 | TRAIL | [58] |
| Blood pDCs | IL-3/IL-10 | T cells | 1:250 | Granzyme B | [54] |
| Blood pDCs | IL-3 | K562 | 10:1 | Granzyme B /Caspase |
[53] |
| Blood pDCs | CpG | H9 (CD4+ T cell line) | 2:1 | TRAILa | [57] |
| Blood pDCs | IMQ CpG IFNα |
Jurkat WM793 SKMel2 (melanoma) Jurkat Jurkat WM793 SKMel2 |
20:1 | TRAIL | [51] |
| Blood pDCs | IL-3 → R848 → FSME → |
K562 K562 K562 Daudi (lymphatic cancer) |
20:1 | ND ND Contact dependent |
[50] |
Partially dependent, other mechanisms (shown when possible) may be involved.
Chronic hepatitis C patients.
Abbreviation: IMQ, imiquimod.
pDCs as killer cells
pDCs are defined as Lin–D11c–CD4+CD45RA+IL-3Rα(CD123)+ILT3+ cells. Additionally, the markers BDCA-2 (CD303), BDCA-4 (CD304), and immunoglobulin-like transcript (ILT)7 are restricted to pDCs both in peripheral blood and bone marrow [46]. On the functional level, human pDCs differ from other DC subsets by their ability to produce large amounts of type I IFNs upon TLR7 or TLR9 ligation by viral or bacterial components [47]. pDCs are generally found to circulate in the periphery, however they can infiltrate tissue in case of infection, inflammation, or tumor 44, 48. Like their myeloid counterparts, pDCs are also described to exert a direct cytotoxic function (Figure 1 and Table 2). For example, the human pDC cell line GEN2.2 is capable of lysing tumor cells in a partly TRAIL-dependent manner after stimulation with inactivated influenza virus or type I IFNs (Table 2). This pDC cell line expresses the NK cell surface marked CD56, whereas other NK cell markers are absent [49]. pDCs activated by the tick-borne encephalitis vaccine FMSE also upregulate CD56, whereas IL-3 or several TLR agonists do not induce upregulation [50]. The CD56+ pDCs express high amounts of TRAIL and granzyme B but neither the cytotoxic molecules nor CD56 are required for the observed cytotoxicity. Nevertheless, the killing capacity is dependent on cell-to-cell contact, whereby FSME-pDCs specifically lyse MHC-class-I-negative tumor cell lines Daudi and K562, but not MHC class I positive cell lines [50]. By contrast, pDCs stimulated with imiquimod (a TLR7 agonist and to lesser extend TLR8 agonist), CpG, or IFN-α kill MHC-class-I-positive tumor cells in a TRAIL- and contact-dependent manner 44, 51. Although TRAIL appears to be an important mediator of pDC-mediated cytotoxicity, other cytotoxic effector molecules are implicated (Table 2). It has been shown that different cytotoxic effector molecules are expressed by blood pDCs, including TRAIL, granzyme B, and lysozyme. High lysozyme expression by the CD2high pDC subset has been observed, although this is not related to the increased cytotoxic activity of the CD2high pDCs as compared to their CD2low counterparts. It is important to note that the high lysozyme expression in the CD2high pDC subset could not be confirmed in another independently performed study [51], indicating that other mechanisms, such as the superior ability of CD2high pDCs over CD2low pDCs to bind their targets, might be involved in the cytotoxic action of CD2high pDCs [52]. Expression of granzyme B has been observed in unstimulated pDCs 51, 52 as well as in pDCs stimulated with IL-3 and/or IL-10 50, 53, 54, but this molecule only seems to contribute to cytotoxicity in the stimulated pDCs. These granzyme-B-producing, IL-3- and IL-10-activated pDCs target T cells in a granzyme-B-dependent, but perforin-independent manner [54]. This confirms the findings by others that killer DCs can mediate cytotoxicity through the granzyme pathway while being completely devoid of perforin 27, 51. Apart from granzyme B, TRAIL has also been implicated as a mediator of T cell lysis by pDCs. pDCs stimulated with purified human T cell leukemia virus (HTLV)-1 or HIV-1 upregulate TRAIL and induced TRAIL-dependent apoptosis in primary CD4+ T cells or CD4+ T cell lines 55, 56. Both the TLR7 pathway and endosomal degradation are involved in the transformation of pDCs into their killer variant by HLTV-1 [55]. In agreement with this, pDCs isolated from viremic HIV patients express TRAIL and induce apoptosis of HIV-infected CD4+ T cells [57]. In another study, TRAIL+ pDCs displayed no cytotoxic responsiveness to HIV-infected autologous CD4+ T cells, but were capable of inducing apoptosis of an HIV-infected CD4+ T cell line [58]. The ability of human killer pDCs to induce apoptosis in virus-infected cells may be a protective mechanism by which the host immune system controls virus spread [59].
Figure 1.
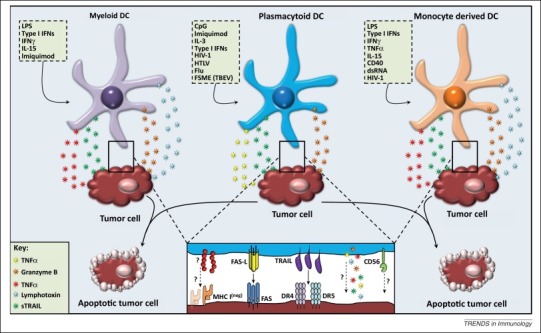
Direct cytotoxicity of human killer DC subsets. Activation of human DCs with various stimuli, for example, viruses, cytokines, TLR ligands, induces a cytotoxic function. DCs can exert their cytotoxic ability either by the secretion of soluble factors or by the expression of apoptosis inducing molecules. It is unclear whether soluble factors play a role at the contact site between target cells and DCs, how killer DCs recognize MHC-class-I-negative target cells, or if there is a role for CD56 in the cytotoxic function of killer DCs. Abbreviations: CpG, CpG oligodeoxynucleotides; DC, dendritic cell; dsRNA, double-stranded RNA; Flu, influenza; FSME, früh sommer meningo-encephalitis; HIV-1, human immunodeficiency virus 1; HTLV, human T lymphotrophic virus; IFN, interferon; IL, interleukin; LPS, lipopolysaccharide; sTRAIL, soluble tumor necrosis factor-related apoptosis-inducing ligand; TBEV, tick-borne encephalitis virus; TNF, tumor necrosis factor.
Killer DCs in the war on cancer
Indirect cytotoxic effects of pDC-derived type I IFNs
pDCs are generally accepted as the major type I IFN-producing cells of the immune system. These type I IFNs can initiate protective immunity through maturation of mDCs and subsequent activation of T cells and NK cells (reviewed in [60]). Thereby, pDCs may play a central role in inducing indirect cytotoxic activity against tumors via various pathways, for example, apoptosis induction and anti-angiogenesis via signaling through a common IFNα receptor. Furthermore, the direct inhibitory effects on tumor cell growth/functions were thought to be the major mechanisms in the antitumor response in IFN-treated patients. In fact, IFNα can directly inhibit the proliferation of tumor cells both in vitro and in vivo, and can exert other direct effects on tumor cells (Figure 2 ) [61]. Next to a direct cytotoxic and cytostatic effect, IFNα also has wide stimulatory effects on other immune cells. As discussed above, monocytes can differentiate into killer DCs under the control of IFNα. In addition, recent studies have shown that IFNα also improves mDC survival and the capacity to store and process exogenous antigens, leading to enhanced cross-presentation and cross-priming of antigen-specific CD8+ T cell responses 62, 63. In vivo evidence was recently provided by two independent studies showing that, in mice, type I IFNs were critical for the induction of antitumor immune responses 62, 64. Furthermore, type I IFNs can regulate NK cell function, by enhancing the of NK cells to kill target cells and to produce IFNγ [65]. Also, type I IFNs promote the accumulation and/or survival of proliferating NK cells by the type I IFN and signal transducer and activator of transcription (STAT)1-dependent induction of IL-15 [65].
Figure 2.
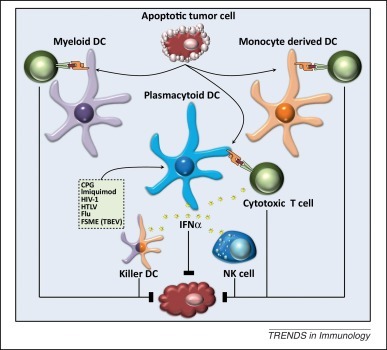
Indirect cytotoxicity of human killer DC subsets. Antigen-loaded killer DCs have the ability to activate antigen-specific cytotoxic T cells that in turn can lyse target cells. Whether killer DCs also have the ability to cross-present antigens obtained from killed target cells and thereafter cross-prime cytotoxic T cells is unclear. Furthermore, human pDCs produce large amounts of type I IFNs upon stimulation and, next to a direct cytotoxic effect, can exert indirect cytotoxic effects by: (i) activating NK cells; (ii) enhancing antigen cross-presentation and cross-priming of T cells; and (iii) generating of IFNα-induced killer MoDCs/mDCs. Abbreviations: CPG, CpG oligodeoxynucleotides; DC, dendritic cell; Flu, influenza; FSME, früh sommer meningo-encephalitis; HIV-1, human immunodeficiency virus 1; HTLV, human T lymphotropic virus; IFN, interferon; NK, natural killer; TBEV, tick-borne encephalitis virus.
These studies suggest that upon activation, pDCs can exert a wide variety of indirect cytotoxic antitumor effects (Figure 2). This notion is underscored by a study demonstrating that either pDC or IFNα depletion leads to a loss of the TRAIL-mediated tumor cell killing by CD14+ monocytes. This highlights a crucial role for pDC-derived IFNα in antitumor immunity [66]. The expression of TRAIL on a wide variety of immune cells is known to be regulated by type I IFNs 67, 68. Moreover, in some studies there has even been a direct link between pDC-derived type IFN, TRAIL expression, and target killing in the context of HIV 57, 69. Whether or not killer pDCs also acquire TRAIL expression under all other reported conditions in a paracrine or autocrine type-I-IFN-dependent manner remains to be determined.
Activation of tumor-infiltrating DCs to boost antitumor immunotherapy
The discovery of the ability of DCs to become tumor cell killers has generated new opportunities for future nonconventional immunotherapeutic strategies. It is generally accepted that macrophages and DCs outnumber NK cells and CTLs in tumor tissues, making these professional APCs ideal candidates to target and induce an antitumor response [10]. Human DCs infiltrate a vast range of tumors including skin cancer, ovarian carcinoma, and lung and colorectal cancer. However, this infiltration does not lead to a conclusive role in prognosis [48]. Infiltration by pDCs in breast cancer and ovarian carcinoma is correlated with an adverse prognostic outcome 70, 71, whereas infiltration of pDCs in lung cancer does not correlate with prognosis [72]. In particular, a tolerogenic state seems to be of importance because DCs can remain in an inactivated state due to the immunosuppressive environment of the tumor [48].
Nevertheless, some studies have shown tumor regression upon treatment with several TLR agonists, such as CpG or imiquimod. Stary et al. have shown that, upon treatment of basal cell carcinoma patients with imiquimod, mDCs and pDCs are recruited to the tumor site and express cytotoxic effector molecules [44]. In a mouse model of transplantable melanoma, treatment with imiquimod led to tumor clearance in a manner dependent on production of chemokine CC ligand (CCL)2 by mast cells. CCL2 induced the massive recruitment of pDCs into the skin, leading to a pDC-dependent reduced melanoma growth [73]. In another study, treatment of human basal cell carcinoma with imiquimod resulted in tumor regression by recruiting pDCs that specifically lysed the tumor cells expressing TRAIL receptor 1 in a TRAIL-dependent manner, and mDCs that expressed granzyme B [44]. The fact that pDCs only lysed cells expressing TRAILR1 indicates that these cells have the capacity to act as anticancer effector cells, thereby showing potential as targets for tumor clearance. These data suggest that tumor-infiltrating DCs can act as killer cells that are directly involved in tumor clearance. A recent study demonstrated that depletion of unactivated pDCs in an orthotopic mammary tumor model delayed tumor growth; however, in the same model, intratumoral administration of a TLR7 agonist led to pDC- and type-I-IFN-dependent tumor regression [74]. These findings point to a Janus-faced function of tumor infiltrating pDCs, and suggest that their function can be largely determined by context and activation state.
Indirect antitumoral activity of killer DCs via amplification of the adaptive immune response
In the past decade, clinical trials carried out by investigators worldwide have shown that vaccination with DCs loaded ex vivo with tumor peptides can induce tumor-specific immune responses in patients with advanced cancer [75]. However, the clinical results obtained so far have been rather disappointing, with only a minority of the treated patients showing long-lasting clinical responses. Many research efforts are currently being undertaken to improve the clinical efficacy of DC-based cancer immunotherapy protocols. Thus far, virtually all clinical trials were based on ex vivo generated DCs, either derived from monocytes or CD34+ progenitor cells. Only recently, the possibility has been explored to exploit scarce naturally circulating DCs such as pDCs to vaccinate end-stage melanoma patients, which showed promising results in terms of overall survival [2].
Several research groups have demonstrated that human killer DCs, apart from their direct tumoricidal activity, can present tumor antigen to T cells, providing a strong rationale for the use of killer DCs in DC-based vaccination protocols. Both IL-15 and IFNα differentiated CD56+ MoDCs were found to be efficient stimulators of antigen-specific T cell responses 26, 27. Human pDCs also have the capacity to present antigens and be potent stimulators of both CD4+ and CD8+ T cell responses 2, 50, 76. The high effector: target ratios that are required to detect killing activity argue in favor of a more predominant role for DCs in the acquisition and presentation of antigens rather than a role as effector cells capable of killing of tumor or virus-infected cells.
Concluding remarks
We have reviewed recent studies providing evidence for an effector role for DCs in cytolysis. We propose that DCs, and pDCs in particular, present a promising target for immunotherapy because they infiltrate tumor lesions and are, upon activation, capable of specifically kill tumor cells, either directly or via activation of other cytotoxic effector cells. However, many questions remain before killer DCs can be used in their full capacity in a clinical situation. How exactly do killer DCs recognize tumor/target cells, and in particular MHC-class-I-negative cells? What are the exact mechanisms that these DCs use, for example, exclusively TRAIL or other mechanisms? Furthermore, can human killer DC cross-present antigens derived from their killed target cells? Answering these questions may help to exploit these killer DCs for immunotherapy.
Acknowledgments
This work was supported by The Netherlands Organization for Scientific Research (NWO ZonMW) (Vidi grant 91776363 to IJMdV), the Netherlands Institute for Regenerative Medicine (NIRM, grant No. FES0908) and EU (ERC grant 269019). ES is a former postdoctoral research fellow of the Research Foundation Flanders (FWO Vlaanderen) and is supported by a training grant of the Belgian Hematological Society. SA is a former PhD fellow of the Research Foundation Flanders and currently holds an Emmanuel Van der Schueren Fellowship of the Flemish League against Cancer (Vlaamse Liga tegen Kanker). SA is also supported by a personal grant from the Belgian Foundation against Cancer and the Belgian public utility foundation VOCATIO. CGF was awarded with the NWO Spinoza award.
References
- 1.Banchereau J., Steinman R.M. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. doi: 10.1038/32588. [DOI] [PubMed] [Google Scholar]
- 2.Tel J. Natural human plasmacytoid dendritic cells induce antigen-specific T-cell responses in melanoma patients. Cancer Res. 2013;73:1063–1075. doi: 10.1158/0008-5472.CAN-12-2583. [DOI] [PubMed] [Google Scholar]
- 3.Shortman K. Dendritic cells and T lymphocytes: developmental and functional interactions. Ciba Found. Symp. 1997;204:130–138. doi: 10.1002/9780470515280.ch9. discussion 138–141. [DOI] [PubMed] [Google Scholar]
- 4.Chan C.W. Interferon-producing killer dendritic cells provide a link between innate and adaptive immunity. Nat. Med. 2006;12:207–213. doi: 10.1038/nm1352. [DOI] [PubMed] [Google Scholar]
- 5.Taieb J. A novel dendritic cell subset involved in tumor immunosurveillance. Nat. Med. 2006;12:214–219. doi: 10.1038/nm1356. [DOI] [PubMed] [Google Scholar]
- 6.Pillarisetty V.G. Natural killer dendritic cells have both antigen presenting and lytic function and in response to CpG produce IFN-gamma via autocrine IL-12. J. Immunol. 2005;174:2612–2618. doi: 10.4049/jimmunol.174.5.2612. [DOI] [PubMed] [Google Scholar]
- 7.Blasius A.L. Development and function of murine B220+CD11c+NK1.1+ cells identify them as a subset of NK cells. J. Exp. Med. 2007;204:2561–2568. doi: 10.1084/jem.20070991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vosshenrich C.A. CD11cloB220+ interferon-producing killer dendritic cells are activated natural killer cells. J. Exp. Med. 2007;204:2569–2578. doi: 10.1084/jem.20071451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Caminschi I. Putative IKDCs are functionally and developmentally similar to natural killer cells, but not to dendritic cells. J. Exp. Med. 2007;204:2579–2590. doi: 10.1084/jem.20071351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bonmort M. Killer dendritic cells: IKDC and the others. Curr. Opin. Immunol. 2008;20:558–565. doi: 10.1016/j.coi.2008.04.006. [DOI] [PubMed] [Google Scholar]
- 11.Larmonier N. Killer dendritic cells and their potential for cancer immunotherapy. Cancer Immunol. Immunother. 2010;59:1–11. doi: 10.1007/s00262-009-0736-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Griffith T.S. Monocyte-mediated tumoricidal activity via the tumor necrosis factor-related cytokine, TRAIL. J. Exp. Med. 1999;189:1343–1354. doi: 10.1084/jem.189.8.1343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Papewalis C. Increased numbers of tumor-lysing monocytes in cancer patients. Mol. Cell. Endocrinol. 2011;337:52–61. doi: 10.1016/j.mce.2011.01.020. [DOI] [PubMed] [Google Scholar]
- 14.Sconocchia G. Phenotype and function of a CD56+ peripheral blood monocyte. Leukemia. 2005;19:69–76. doi: 10.1038/sj.leu.2403550. [DOI] [PubMed] [Google Scholar]
- 15.Nakayama M. Involvement of TWEAK in interferon gamma-stimulated monocyte cytotoxicity. J. Exp. Med. 2000;192:1373–1380. doi: 10.1084/jem.192.9.1373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schmitz M. Tumoricidal potential of native blood dendritic cells: direct tumor cell killing and activation of NK cell-mediated cytotoxicity. J. Immunol. 2005;174:4127–4134. doi: 10.4049/jimmunol.174.7.4127. [DOI] [PubMed] [Google Scholar]
- 17.Schmitz M. Native human blood dendritic cells as potent effectors in antibody-dependent cellular cytotoxicity. Blood. 2002;100:1502–1504. [PubMed] [Google Scholar]
- 18.Vidalain P.O. Measles virus induces functional TRAIL production by human dendritic cells. J. Virol. 2000;74:556–559. doi: 10.1128/jvi.74.1.556-559.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fugier-Vivier I. Measles virus suppresses cell-mediated immunity by interfering with the survival and functions of dendritic and T cells. J. Exp. Med. 1997;186:813–823. doi: 10.1084/jem.186.6.813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Raftery M.J. Targeting the function of mature dendritic cells by human cytomegalovirus: a multilayered viral defense strategy. Immunity. 2001;15:997–1009. doi: 10.1016/s1074-7613(01)00239-4. [DOI] [PubMed] [Google Scholar]
- 21.Lichtner M. HIV type 1-infected dendritic cells induce apoptotic death in infected and uninfected primary CD4 T lymphocytes. AIDS Res. Hum. Retroviruses. 2004;20:175–182. doi: 10.1089/088922204773004897. [DOI] [PubMed] [Google Scholar]
- 22.Quaranta M.G. HIV-1 Nef equips dendritic cells to reduce survival and function of CD8+ T cells: a mechanism of immune evasion. FASEB J. 2004;18:1459–1461. doi: 10.1096/fj.04-1633fje. [DOI] [PubMed] [Google Scholar]
- 23.Majumder B. Dendritic cells infected with vpr-positive human immunodeficiency virus type 1 induce CD8+ T-cell apoptosis via upregulation of tumor necrosis factor alpha. J. Virol. 2007;81:7388–7399. doi: 10.1128/JVI.00893-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vidalain P.O. Cytotoxic activity of human dendritic cells is differentially regulated by double-stranded RNA and CD40 ligand. J. Immunol. 2001;167:3765–3772. doi: 10.4049/jimmunol.167.7.3765. [DOI] [PubMed] [Google Scholar]
- 25.Liu S. The involvement of TNF-alpha-related apoptosis-inducing ligand in the enhanced cytotoxicity of IFN-beta-stimulated human dendritic cells to tumor cells. J. Immunol. 2001;166:5407–5415. doi: 10.4049/jimmunol.166.9.5407. [DOI] [PubMed] [Google Scholar]
- 26.Papewalis C. IFN-alpha skews monocytes into CD56+-expressing dendritic cells with potent functional activities in vitro and in vivo. J. Immunol. 2008;180:1462–1470. doi: 10.4049/jimmunol.180.3.1462. [DOI] [PubMed] [Google Scholar]
- 27.Anguille S. Interleukin-15-induced CD56(+) myeloid dendritic cells combine potent tumor antigen presentation with direct tumoricidal potential. PLoS ONE. 2012;7:e51851. doi: 10.1371/journal.pone.0051851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Tomihara K. Gene transfer of CD40-ligand to dendritic cells stimulates interferon-gamma production to induce growth arrest and apoptosis of tumor cells. Gene Ther. 2008;15:203–213. doi: 10.1038/sj.gt.3303056. [DOI] [PubMed] [Google Scholar]
- 29.Lu G. Innate direct anticancer effector function of human immature dendritic cells. II. Role of TNF, lymphotoxin-alpha(1)beta(2), Fas ligand, and TNF-related apoptosis-inducing ligand. J. Immunol. 2002;68:1831–1839. doi: 10.4049/jimmunol.168.4.1831. [DOI] [PubMed] [Google Scholar]
- 30.Joo H.G. Human dendritic cells induce tumor-specific apoptosis by soluble factors. Int. J. Cancer. 2002;102:20–28. doi: 10.1002/ijc.10656. [DOI] [PubMed] [Google Scholar]
- 31.Yang R. Immature dendritic cells kill ovarian carcinoma cells by a FAS/FASL pathway, enabling them to sensitize tumor-specific CTLs. Int. J. Cancer. 2001;94:407–413. doi: 10.1002/ijc.1484. [DOI] [PubMed] [Google Scholar]
- 32.Zhao L., Tyrrell D.L. Myeloid dendritic cells can kill T cells during chronic hepatitis C virus infection. Virol. Immunol. 2013;26:25–39. doi: 10.1089/vim.2012.0058. [DOI] [PubMed] [Google Scholar]
- 33.Vanderheyde N. Tumoricidal activity of monocyte-derived dendritic cells: evidence for a caspase-8-dependent, Fas-associated death domain-independent mechanism. J. Immunol. 2001;167:3565–3569. doi: 10.4049/jimmunol.167.7.3565. [DOI] [PubMed] [Google Scholar]
- 34.Schiltz P.M. Human allogeneic and murine xenogeneic dendritic cells are cytotoxic to human tumor cells via two distinct pathways. Cancer Biother. Radiopharm. 2007;22:672–683. doi: 10.1089/cbr.2007.356. [DOI] [PubMed] [Google Scholar]
- 35.Vanderheyde N. Distinct mechanisms are involved in tumoristatic and tumoricidal activities of monocyte-derived dendritic cells. Immunol. Lett. 2004;91:99–101. doi: 10.1016/j.imlet.2003.11.011. [DOI] [PubMed] [Google Scholar]
- 36.Kim S.K. Enhanced anti-cancer activity of human dendritic cells sensitized with gamma-irradiation-induced apoptotic colon cancer cells. Cancer Lett. 2013;335:278–288. doi: 10.1016/j.canlet.2013.02.038. [DOI] [PubMed] [Google Scholar]
- 37.Sakhno L.V. Cytotoxic activity of dendritic cells as a possible mechanism of negative regulation of T lymphocytes in pulmonary tuberculosis. Clin. Dev. Immunol. 2012;2012:628635. doi: 10.1155/2012/628635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cottrez F. Priming of human CD4+ antigen-specific T cells to undergo apoptosis by HIV-infected monocytes. A two-step mechanism involving the gp120 molecule. J. Clin. Invest. 1997;99:257–266. doi: 10.1172/JCI119154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.MacDonald K.P. Characterization of human blood dendritic cell subsets. Blood. 2002;100:4512–4520. doi: 10.1182/blood-2001-11-0097. [DOI] [PubMed] [Google Scholar]
- 40.Fanger N.A. Human dendritic cells mediate cellular apoptosis via tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) J. Exp. Med. 1999;190:1155–1164. doi: 10.1084/jem.190.8.1155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Janjic B.M. Innate direct anticancer effector function of human immature dendritic cells. I. Involvement of an apoptosis-inducing pathway. J. Immunol. 2002;168:1823–1830. doi: 10.4049/jimmunol.168.4.1823. [DOI] [PubMed] [Google Scholar]
- 42.Ciesek S. Impaired TRAIL-dependent cytotoxicity of CD1c-positive dendritic cells in chronic hepatitis C virus infection. J. Viral Hepat. 2008;15:200–211. doi: 10.1111/j.1365-2893.2007.00930.x. [DOI] [PubMed] [Google Scholar]
- 43.Hemont C. Human blood mDC subsets exhibit distinct TLR repertoire and responsiveness. J. Leukoc. Biol. 2013;93:599–609. doi: 10.1189/jlb.0912452. [DOI] [PubMed] [Google Scholar]
- 44.Stary G. Tumoricidal activity of TLR7/8-activated inflammatory dendritic cells. J. Exp. Med. 2007;204:1441–1451. doi: 10.1084/jem.20070021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Manna P.P. IL-15 activated human peripheral blood dendritic cell kill allogeneic and xenogeneic endothelial cells via apoptosis. Cytokine. 2013;61:118–126. doi: 10.1016/j.cyto.2012.09.004. [DOI] [PubMed] [Google Scholar]
- 46.Dzionek A. BDCA-2, BDCA-3, and BDCA-4: three markers for distinct subsets of dendritic cells in human peripheral blood. J. Immunol. 2000;165:6037–6046. doi: 10.4049/jimmunol.165.11.6037. [DOI] [PubMed] [Google Scholar]
- 47.Cella M. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat. Med. 1999;5:919–923. doi: 10.1038/11360. [DOI] [PubMed] [Google Scholar]
- 48.Karthaus N. Deciphering the message broadcast by tumor-infiltrating dendritic cells. Am. J. Pathol. 2012;181:733–742. doi: 10.1016/j.ajpath.2012.05.012. [DOI] [PubMed] [Google Scholar]
- 49.Chaperot L. Virus or TLR agonists induce TRAIL-mediated cytotoxic activity of plasmacytoid dendritic cells. J. Immunol. 2006;176:248–255. doi: 10.4049/jimmunol.176.1.248. [DOI] [PubMed] [Google Scholar]
- 50.Tel J. Human plasmacytoid dendritic cells are equipped with antigen-presenting and tumoricidal capacities. Blood. 2012;120:3936–3944. doi: 10.1182/blood-2012-06-435941. [DOI] [PubMed] [Google Scholar]
- 51.Kalb M.L. TRAIL(+) human plasmacytoid dendritic cells kill tumor cells in vitro: mechanisms of imiquimod- and IFN-alpha-mediated antitumor reactivity. J. Immunol. 2012;188:1583–1591. doi: 10.4049/jimmunol.1102437. [DOI] [PubMed] [Google Scholar]
- 52.Matsui T. CD2 distinguishes two subsets of human plasmacytoid dendritic cells with distinct phenotype and functions. J. Immunol. 2009;182:6815–6823. doi: 10.4049/jimmunol.0802008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Bratke K. Functional expression of granzyme B in human plasmacytoid dendritic cells: a role in allergic inflammation. Clin. Exp. Allergy. 2010;40:1015–1024. doi: 10.1111/j.1365-2222.2010.03499.x. [DOI] [PubMed] [Google Scholar]
- 54.Jahrsdorfer B. Granzyme B produced by human plasmacytoid dendritic cells suppresses T-cell expansion. Blood. 2010;115:1156–1165. doi: 10.1182/blood-2009-07-235382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Colisson R. Free HTLV-1 induces TLR7-dependent innate immune response and TRAIL relocalization in killer plasmacytoid dendritic cells. Blood. 2010;115:2177–2185. doi: 10.1182/blood-2009-06-224741. [DOI] [PubMed] [Google Scholar]
- 56.Hardy A.W. HIV turns plasmacytoid dendritic cells (pDC) into TRAIL-expressing killer pDC and down-regulates HIV coreceptors by Toll-like receptor 7-induced IFN-alpha. Proc. Natl. Acad. Sci. U.S.A. 2007;104:17453–17458. doi: 10.1073/pnas.0707244104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Barblu L. Plasmacytoid dendritic cells (pDCs) from HIV controllers produce interferon-alpha and differentiate into functional killer pDCs under HIV activation. J. Infect. Dis. 2012;206:790–801. doi: 10.1093/infdis/jis384. [DOI] [PubMed] [Google Scholar]
- 58.Chehimi J. Inability of plasmacytoid dendritic cells to directly lyse HIV-infected autologous CD4+ T cells despite induction of tumor necrosis factor-related apoptosis-inducing ligand. J. Virol. 2010;84:2762–2773. doi: 10.1128/JVI.01350-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Vilcek J. Boosting p53 with interferon and viruses. Nat. Immunol. 2003;4:825–826. doi: 10.1038/ni0903-825. [DOI] [PubMed] [Google Scholar]
- 60.Arico E., Belardelli F. Interferon-alpha as antiviral and antitumor vaccine adjuvants: mechanisms of action and response signature. J. Interferon Cytokine Res. 2012;32:235–247. doi: 10.1089/jir.2011.0077. [DOI] [PubMed] [Google Scholar]
- 61.Anguille S. Interferon-alpha in acute myeloid leukemia: an old drug revisited. Leukemia. 2011;25:739–748. doi: 10.1038/leu.2010.324. [DOI] [PubMed] [Google Scholar]
- 62.Diamond M.S. Type I interferon is selectively required by dendritic cells for immune rejection of tumors. J. Exp. Med. 2011;208:1989–2003. doi: 10.1084/jem.20101158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Spadaro F. IFN-alpha enhances cross-presentation in human dendritic cells by modulating antigen survival, endocytic routing, and processing. Blood. 2012;119:1407–1417. doi: 10.1182/blood-2011-06-363564. [DOI] [PubMed] [Google Scholar]
- 64.Fuertes M.B. Host type I IFN signals are required for antitumor CD8+ T cell responses through CD8{alpha}+ dendritic cells. J. Exp. Med. 2011;208:2005–2016. doi: 10.1084/jem.20101159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Nguyen K.B. Coordinated and distinct roles for IFN-alpha beta, IL-12, and IL-15 regulation of NK cell responses to viral infection. J. Immunol. 2002;169:4279–4287. doi: 10.4049/jimmunol.169.8.4279. [DOI] [PubMed] [Google Scholar]
- 66.Kemp T.J. Plasmacytoid dendritic cell-derived IFN-alpha induces TNF-related apoptosis-inducing ligand/Apo-2L-mediated antitumor activity by human monocytes following CpG oligodeoxynucleotide stimulation. J. Immunol. 2003;171:212–218. doi: 10.4049/jimmunol.171.1.212. [DOI] [PubMed] [Google Scholar]
- 67.Kayagaki N. Type I interferons (IFNs) regulate tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) expression on human T cells: a novel mechanism for the antitumor effects of type I IFNs. J. Exp. Med. 1999;189:1451–1460. doi: 10.1084/jem.189.9.1451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Chawla-Sarkar M. Apoptosis and interferons: role of interferon-stimulated genes as mediators of apoptosis. Apoptosis. 2003;8:237–249. doi: 10.1023/a:1023668705040. [DOI] [PubMed] [Google Scholar]
- 69.Herbeuval J.P. Regulation of TNF-related apoptosis-inducing ligand on primary CD4+ T cells by HIV-1: role of type I IFN-producing plasmacytoid dendritic cells. Proc. Natl. Acad. Sci. U.S.A. 2005;102:13974–13979. doi: 10.1073/pnas.0505251102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Treilleux I. Dendritic cell infiltration and prognosis of early stage breast cancer. Clin. Cancer Res. 2004;10:7466–7474. doi: 10.1158/1078-0432.CCR-04-0684. [DOI] [PubMed] [Google Scholar]
- 71.Labidi-Galy S.I. Quantitative and functional alterations of plasmacytoid dendritic cells contribute to immune tolerance in ovarian cancer. Cancer Res. 2011;71:5423–5434. doi: 10.1158/0008-5472.CAN-11-0367. [DOI] [PubMed] [Google Scholar]
- 72.Tabarkiewicz J. CD1c+ and CD303+ dendritic cells in peripheral blood, lymph nodes and tumor tissue of patients with non-small cell lung cancer. Oncol. Rep. 2008;19:237–243. [PubMed] [Google Scholar]
- 73.Drobits B. Imiquimod clears tumors in mice independent of adaptive immunity by converting pDCs into tumor-killing effector cells. J. Clin. Invest. 2012;122:575–585. doi: 10.1172/JCI61034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Le Mercier I. Tumor promotion by intratumoral plasmacytoid dendritic cells is reversed by TLR7 ligand treatment. Cancer Res. 2013;73:4629–4640. doi: 10.1158/0008-5472.CAN-12-3058. [DOI] [PubMed] [Google Scholar]
- 75.Figdor C.G. Dendritic cell immunotherapy: mapping the way. Nat. Med. 2004;10:475–480. doi: 10.1038/nm1039. [DOI] [PubMed] [Google Scholar]
- 76.Tel J. Human plasmacytoid dendritic cells efficiently cross-present exogenous Ags to CD8+ T cells despite lower Ag uptake than myeloid dendritic cell subsets. Blood. 2013;121:459–467. doi: 10.1182/blood-2012-06-435644. [DOI] [PubMed] [Google Scholar]
- 77.Jahnisch H. TLR7/8 agonists trigger immunostimulatory properties of human 6-sulfo LacNAc dendritic cells. Cancer Lett. 2013;335:119–127. doi: 10.1016/j.canlet.2013.02.003. [DOI] [PubMed] [Google Scholar]
- 78.Chapoval A.I. In vitro growth inhibition of a broad spectrum of tumor cell lines by activated human dendritic cells. Blood. 2000;95:2346–2351. [PubMed] [Google Scholar]
- 79.Hubert P. Dendritic cells induce the death of human papillomavirus-transformed keratinocytes. FASEB J. 2001;15:2521–2523. doi: 10.1096/fj.00-0872fje. [DOI] [PubMed] [Google Scholar]
- 80.Shi J. Activated human umbilical cord blood dendritic cells kill tumor cells without damaging normal hematological progenitor cells. Cancer Sci. 2005;96:127–133. doi: 10.1111/j.1349-7006.2005.00017.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Korthals M. Monocyte derived dendritic cells generated by IFN-alpha acquire mature dendritic and natural killer cell properties as shown by gene expression analysis. J. Transl. Med. 2007;5:46. doi: 10.1186/1479-5876-5-46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Hill K.S. OK432-activated human dendritic cells kill tumor cells via CD40/CD40 ligand interactions. J. Immunol. 2008;181:3108–3115. doi: 10.4049/jimmunol.181.5.3108. [DOI] [PubMed] [Google Scholar]
- 83.Shibru D. Does the 3-gene diagnostic assay accurately distinguish benign from malignant thyroid neoplasms? Cancer. 2008;113:930–935. doi: 10.1002/cncr.23703. [DOI] [PubMed] [Google Scholar]
- 84.Law H.K. Toll-like receptors, chemokine receptors and death receptor ligands responses in SARS coronavirus infected human monocyte derived dendritic cells. BMC Immunol. 2009;10:35. doi: 10.1186/1471-2172-10-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Jung S.T. Aglycosylated IgG variants expressed in bacteria that selectively bind FcgammaRI potentiate tumor cell killing by monocyte-dendritic cells. Proc. Natl. Acad. Sci. U.S.A. 2010;107:604–609. doi: 10.1073/pnas.0908590107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Lakomy D. Cytotoxic dendritic cells generated from cancer patients. J. Immunol. 2011;187:2775–2782. doi: 10.4049/jimmunol.1004146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Koski G.K. A novel dendritic cell-based immunization approach for the induction of durable Th1-polarized anti-HER-2/neu responses in women with early breast cancer. J. Immunother. 2012;35:54–65. doi: 10.1097/CJI.0b013e318235f512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Manna P.P., Mohanakumar T. Human dendritic cell mediated cytotoxicity against breast carcinoma cells in vitro. J. Leukoc. Biol. 2002;72:312–320. [PubMed] [Google Scholar]
- 89.Stary G. Plasmacytoid dendritic cells express TRAIL and induce CD4+ T-cell apoptosis in HIV-1 viremic patients. Blood. 2009;114:3854–3863. doi: 10.1182/blood-2009-04-217927. [DOI] [PubMed] [Google Scholar]
- 90.Riboldi E. Engagement of BDCA-2 blocks TRAIL-mediated cytotoxic activity of plasmacytoid dendritic cells. Immunobiology. 2009;214:868–876. doi: 10.1016/j.imbio.2009.06.016. [DOI] [PubMed] [Google Scholar]