

Abstract
Background
Brassica napus is an important oilseed crop that offers a considerable amount of biomass for global vegetable oil production. The establishment of an efficient genetic transformation system with a convenient transgenic-positive screening method is of great importance for gene functional analysis and molecular breeding. However, to our knowledge, there are few of the aforementioned systems available for efficient application in B. napus.
Results
Based on the well-established genetic transformation system in B. napus, five vectors carrying the red fluorescence protein encoding gene from Discosoma sp. (DsRed) were constructed and integrated into rapeseed via Agrobacterium-mediated hypocotyl transformation. An average of 59.1% tissues were marked with red fluorescence by the visual screening method in tissue culture medium, 96.1% of which, on average, were amplified with the objective genes from eight different rapeseed varieties. In addition, the final transgenic-positive efficiency of the rooted plantlets reached up to 90.7% from red fluorescence marked tissues, which was much higher than that in previous reports. Additionally, visual screening could be applicable to seedlings via integration of DsRed, including seed coats, roots, hypocotyls and cotyledons during seed germination. These results indicate that the highly efficient genetic transformation system combined with the transgenic-positive visual screening method helps to conveniently and efficiently obtain transgenic-positive rapeseed plantlets.
Conclusion
A rapid, convenient and highly efficient method was developed to obtain transgenic plants, which can help to obtain the largest proportion of transgene-positive regenerated plantlets, thereby avoiding a long period of plant regeneration. The results of this study will benefit gene functional studies especially in high-throughput molecular biology research.
Keywords: DsRed, Agrobacterium-mediated hypocotyl transformation, Visual screening, Brassica napus
Background
Brassica napus is an important oilseed crop, ranking as the second most important crop for oilseed production worldwide [1], and it was derived from the hybridization between B. rapa and B. oleracea [2]. To satisfy the increasing demand for oil, it is essential to improve various important economically and agriculturally traits via genetic engineering techniques, which are powerful tools for gene functional analysis and crop improvement [3]. Genetic transformation techniques have promoted the improvement of crop varieties by integration of novel genes to satisfy the needs for high-yield and good-quality crops, including characters for effective oil production [4], herbicide and disease-resistance [5, 6]. It has been successfully used to improve some major crops, such as soybean, maize and cotton [7–9]. New varieties of these aforementioned plants modified by transgenic methods are now planted widely in many countries, introducing great benefits for farmers [10]. However, to our knowledge, the genetic transformation technique in rapeseed is still relatively lower in transformation efficiency and inefficient in positive screening compared with other crops. Therefore, a suitable and favourable genetic transformation system accompanied by a highly efficient screening method is essential for rapeseed breeding and improvement.
To date, several genetic transformation methods have been reported and routinely applied in model plants and major crops, such as Arabidopsis thaliana, Nicotiana benthamiana, rice, wheat and maize [11–15]. In B. napus, various technologies, including PEG-mediated DNA uptake [16, 17], electroporation [18], particle bombardment [19], Agrobacterium-mediated transformation and microspore transfection [20, 21], have been used to obtain genetically modified plants. Among these technologies, Agrobacterium-mediated transformation is the most general, reliable and effective method [22–24]. Maheshwari et al. (2011) investigated the effect of hormonal combinations, donor plant age and explant types on the transgenic frequency and regeneration capacity of plants in four different rapeseed lines (nvigor 5020, Westar, Topas and its microspore derivative-Line 4079) [25]. They found that the transformation frequency was 54.2 and 53.7% in cultivars of Invigor 5020 and Westar, but 16.0 and 13.4% for Topas and Line 4079, respectively. However, due to the diverse genetic transformation ability caused by distinct genetic backgrounds within the cultivars [5], recalcitrance continued to persist in several cultivars which were not capable of being genetically transformed, especially in commercial varieties.
Obtaining highly efficient transgenic-positive regenerated plantlets is mainly dependent on the subsequent selection. Plant regeneration, however, is a time-consuming process that requires a long growth cycle; simultaneously, it often also results in undesired abortions and reduced fertility. Numerous methods have been frequently used for transgenic-positive plant screening, such as antibiotics, PCR, Southern blot analysis, GFP or GUS staining [14, 26]. These methods, however, are tedious, and they are usually performed with leaves, roots or siliques from the regenerated plantlets. Therefore, it is necessary to develop a convenient and efficient screening method to simplify and promote the process of plant transformation. Recently, red fluorescence protein from Discosoma sp. (DsRed), similar to Aequorea victoria green fluorescent protein (GFP) in the secondary structure, has been applied to identify the transgenic seeds by visual screening in plants [15, 27, 28]. Both the excitation and emission wavelength of DsRed (554 nm and 586 nm) are longer than that of GFP (480 nm and 505 nm), which could enable the transgenic-positive screening to be more sensitive [29, 30]. Although both DsRed and GFP do not affect the vegetative and reproductive growth of plants, GFP is more easily influenced by the intrinsic chlorophyll in plants [31, 32]. Stuitje et al. (2003) found that the sensitivity of DsRed was higher than that of GFP in Arabidopsis [33]. Thus, DsRed has been widely used to mark the transgenic pollen, leaf or seed, and even extraplastidic membranes of chloroplasts for visual selection or offspring separation [33–36]. Eckert et al. (2005) were the first to implement the expression of DsRed and another common reporter protein-GFP in Leptosphaeria spp. and Oculimacula spp. to observe the interactions between fungal pathogen species and rapeseed in vitro and in planta [37]. Zheng et al. (2019) investigated the potential of DsRed-labelled Verticillium longisporum dissemination by seeds of B. napus under greenhouse condition, and confirmed the systemic growth of the pathogen from roots to seeds [38]. However, until now, DsRed has not been widely used in tissue culture or in the process of seed development.
The objective of the present study was to construct a convenient and highly efficient method to obtain transgenic-positive lines, by combining a rapid transgenic-positive screening technology with a highly efficient Agrobacterium-mediated hypocotyl genetic transformation system in B. napus. To achieve this goal, highly efficient genetic transformation was conducted in eight rapeseed varieties and five expression vectors. In addition, DsRed was introduced to serve as a visual screening marker for screening transgenic-positive plants during tissue culture and seed development. The present study provides a good foundation for future studies focused on the efficient acquisition of high-throughput transgenic-positive lines.
Materials and methods
Plant materials and growth conditions
The allotetraploid rapeseed genotypes used included the spring varieties Jia 9709, semi-winter varieties Jia 2016, and six commercial winter varieties including Zhong Shuang 8, Zhong Shuang 11, Zhong You 821, 7633, B 351 and Shan 3B. The seeds of Jia 2016 and Jia 9709 were kindly provided by Prof. Chunyu Zhang from Huazhong Agriculture University (Wuhan, China). The seeds of Zhong Shuang 8, Zhong Shuang 11 and Zhong You 821 were developed and provided by the Institute of oil crops, Chinese Academy of Agricultural Sciences. The seeds of 7633, B 351 and Shan 3B were kindly provided by Prof. Dianrong Li from the Hybrid Rapeseed Center of Shaanxi Province. These rapeseed varieties were cultured in pots (6 × 6 × 9 cm) in a tissue culture room, and all the rooted plantlets were grown in pots (12 cm × 15 cm) in a greenhouse with 2000–2500 lux with 16 h of light and 8 h of dark at 25 ℃. During growth, water was supplied three times a week. Aphids were controlled with imidacloprid (Jiangsu Changqing Biotechnology Co., China) and sticky coloured cards (Chunhe, China).
Vector construction
For the BnaA07g17400D and BnaC05g34170D over-expression vectors, the CDS fragments were amplified and then purified with a QIA quick extraction kit (QIAGEN, America) and then linked to pCMABIA-1303 plasmid which was linearized by EcoR I digestion using in-fusion enzyme, as referenced in the In-Fusion® HD Cloning Kit User Manual. For the BnaA07g17400D knock-down vector, two regions (i.e., 364 nt-839 nt and 575 nt-986 nt) were designed. And for the BnaC05g34170D knock down vector, the region from 220 to 691 nt was designed. The sense and antisense strands of the three fragments were amplified with R1-S-F/R, R1-A-F/R, R2-S-F/R, R2-A-F/R, R3-S-F/R and R3-A-F/R (Additional file 4), respectively. After amplification and purification, they were cloned into p35S-1390 which was linearized with Sac I and SnaB I for the sense and antisense strand construction, respectively. Afterwards, the marker gene DsRed, amplified with the designed primers DsRed-F/R (Additional file 4), was purified and then cloned into these reconstructed vectors, digested with Hind III. These reconstructed vectors were transformed into Agrobacterium strain GV3101, and single positive clones were identified by PCR amplification and then stored at − 80 °C.
Agrobacterium tumefaciens-mediated transformation of hypocotyls in rapeseed
A.grobacterium tumefaciens-mediated genetic transformation was performed according to Zhou et al. (2002) with minor modifications [39]. A brief introduction is provided as follows.
First, seed germination. Clean the seed coat with sterilized water for approximately three times, and sterilized with 70% ethanol for 1 min, followed by 50% of 84 disinfectant (the active ingredient is approximately 2.5% NaClO in the ratio of mass-to-volume) for 3 min, and then rinsed 3–5 times with sterilized water. Finally, the seeds were transferred to 1/2 MS medium and germinated for approximately 7 days in a complete dark condition. Second, Agrobacterium preparation. Revive GV3101 harbouring the expression vector that stored at − 80 ℃ for approximately 30 min (28 ℃, 180 rpm) at the sixth day after the first step. Then, 50 μl of the revived Agrobacterium was inoculated into 20 ml LB liquid medium and cultured at 28 ℃ (180 rpm) for approximately 12 h. Generally, the OD600 reached approximately 0.4, and then 2 ml was obtained for centrifugation (3 min, 6000 rpm) to collect the pellet. The supernatant was removed and the pellet rinsed twice with liquid infection medium (LIM, 4.4 g/L MS, 30 g/L sucrose, 200 mM acetosyringone). The resuspended pellet was kept at 4 ℃ for further infection. Third, Agrobacterium infection and cocultivation. Hypocotyls were cut to approximately 1 cm into a sterilized plate containing 18 ml pre-added LIM, and then the aforementioned 2 ml of the resuspended pellet was added to the plate and allowed to infect for approximately 10 min. Next, place the infected hypocotyls in the cocultivation medium (4.4 g/L MS, 30 g/L sucrose, 18 g/L mannitol, 1 mg/L 2,4-D, 0.3 mg/L kinetin, 200 mM acetosyringone and 8 g/L agar) for two days in the dark, at 25 °C. Fourth, calli induction. Transfer all the hypocotyls to calli-inducing medium (CIM, 4.4 g/L MS, 30 g/L sucrose, 18 g/L mannitol, 1 mg/L 2,4-D, 0.3 mg/L kinetin, STS (0.1 M Na2S2O3: 0.1 M AgNO3 = 4:1), 300 mg/L timentin, 25 mg/L hygromycin and 8 g/L agar) for 20 days at 2000–2500 lux. Fifth, shoots induction. Transfer all the hypocotyls with embryogenic calli into a shoot-inducing medium (SIM, 4.4 g/L MS, 10 g/L glucose, 0.25 g/L xylose, 0.6 mg/L MES, 2.0 mg/L zeatin, 0.1 mg/L IAA, 3 mg/L AgNO3, 300 mg/L timentin, 25 mg/L hygromycin and 8 g/L agar) for shoots regeneration, and renewed the medium every 2–3 weeks until at least 3 leaves appeared. Sixth, roots induction and plantlet regeneration. Transfer the induced shoots into root-inducing medium (RIM, 4.4 g/L MS, 10 g/L sucrose, 1 mg/L IBA, 300 mg/L timentin and 8 g/L agar). Then, transplant the roots plantlets with 4–6 leaves into pots with nutritive soil and cultivated in a greenhouse. The semi-winter and winter plantlets should be vernalized in a cooler at 4 °C for approximately 4 weeks before transplanting.
Fluorescence observation and imaging
The red fluorescence in the calli and shoots on the medium during the preliminary stage were observed with a hand-held green fluorescent flashlight through a red filter in the dark, and the images were then captured with a D7100 camera (Nikon, Japan). The red fluorescence in roots, seed coats, hypocotyls, and cotyledons of seedlings were observed using a laser confocal fluorescence microscope FV1000 (Olympus, Japan). The images were processed and arranged with Photoshop software.
RNA extraction and real-time qPCR
To analyse the relative expression level of the DsRed and BnaA07g17400D gene, a set of roots, seed coats, hypocotyls, cotyledons and whole seedlings were collected one week after sowing the seeds on MS medium. Total RNA was extracted using the RNAprep Pure Plant Kit (polysaccharides & polyphenolics-rich) (Tiangen, China), according to the manufacturer’s protocol. Then, approximately 1 µL RNA samples was used to measure the RNA concentration with a NanoDrop 2000 spectrophotometer (Thermo Scientific, USA). The cDNA was synthesized with 1 μg RNA using a ReverTra Ace qPCR RT Master Mix kit with gDNA Remover (Toyobo, Japan) according to the manufacturer’s protocol, and it was stored at − 20 ℃.
The real-time PCR in a total volume of 20 µL were performed with SYBR Green Real-time PCR Master mix (Toyobo, Japan) on a Stepone Plus (ABI, America) according to the provided protocol. Three technical replicates were analysed. The relative expression of each gene was calculated using the ΔΔCt method. The primers used in RT-PCR are listed in Additional file 4.
DNA extraction, PCR-based identification and Southern blotting in transgenic rapeseed plants
For DNA extraction, calli were collected from the third renewed SIM, and young leaves were collected from greenhouse-grown T1 plantlets and from the field for T3 plants to the seedling stage, respectively. They were frozen in liquid nitrogen and stored in−80 °C. Genomic DNA was extracted with a NuClean PlantGen DNA kit (CWBIO, China) and kept at − 20 ℃.
To check the positive transgenic plants, presence of the BnaA07g17400D and BnaC05g34170D gene was performed by PCR amplification using the primer pair BnaA07g17400D-OE-F/R and BnaC05g34170D-OE-F/R for the E1 and E2 transgenic lines, respectively, while the presence of the DsRed gene was detected using the primer pair Red-F/R in the three RNAi lines. The primers are listed in Additional file 4.
Southern blotting analysis was performed in transgenic rapeseed plants according to the published methods with a minor modification [40]. Briefly, total genomic DNA was extracted using the standard CTAB method to obtain a large amount of DNA. Then, 30 μg DNA from each sample was digested with EcoR I, which flanked the DsRed gene in the E1, E2 and RNAi vector. The digested DNA samples were fractionated on a 0.8% agarose gel and transferred onto a nylon Hybond-N + membrane with a membrane transfer instrument. The DsRed PCR product was labelled with digoxigenin and used as a probe for hybridization with the digested DNA on the membrane. The hybridization and detection steps were performed according to the instructions supplied the DIG High Prime DNA Labelling and Detection Starter Kit II.
Results
High efficiency of Agrobacterium-mediated hypocotyl transformation in rapeseed
For Agrobacterium-mediated hypocotyl transformation in rapeseed, the over-expression vector pCMABIA-1303 and RNAi vector p35S-1390 were used for transformation (Fig. 1). Consequently, two over-expression vectors were constructed for BnaA07g17400D and BnaC05g34170D (hereafter called E1 and E2), respectively. Additionally, three RNAi vectors were also designed and constructed to knock down these two genes, i.e., two vectors were for BnaA07g17400D (hereafter called R1 and R2) while the third one was for BnaC05g34170D (hereafter called R3). In E1 and E2, the expression of BnaA07g17400D and BnaC05g34170D was controlled by the glycinin promoter, while in R1, R2 and R3, sense and antisense strands of RNAi fragments were guided by the CaMV35S promoter (Fig. 1). The marker gene-DsRed, was also under the control of the CaMV35S promoter (Fig. 1).
Fig. 1.

Vectors construction for gene over-expression and knock-down. a Diagram of the construction of the over-expression vector. The expression of BnaA07g17400D and BnaC05g34170D is driven by the glycinin promoter. The N-terminus of BnaA07g17400D and BnaC05g34170D is accompanied by a flag. b Diagram of the construction of the RNAi knock-down vector. The expression of sense strands and antisense strands is driven by the 2 × 35S promoter and a 2 × nuclear localization signal (NLS) at the N-terminus. DsRed in both vectors was also under the control of the CaMV35S promoter
Hypocotyls from eight different rapeseed varieties were used for Agrobacterium mediated transformation. The formation from calli to regenerated plantlets on a hypocotyl, i.e., calli formation, shoots formation, roots formation and plantlet regeneration, showed a good condition during the regeneration process (Additional file 1). It is known that the T-DNA integration ability into the plant genome is an important factor for plant transformation. Thus, the concentration of Agrobacterium was carefully controlled in this investigation. To simplify the final concentration adjustment, only the original OD600 needs to be examined for further experiments. Thus, 2 ml Agrobacterium solution at the original concentration was collectedat OD600 = 0.4 to suspended in 2 ml LIM and then diluted to 20 ml for infection. The original OD600 was sometimes higher than 0.4, and it might have reached up to 1.2 after inoculation. In this case, pipette approximately (0.4/OD600) * 2 ml Agrobacterium solution to collect the pellet. For example, pipette 1.5 ml at OD600 approximately 0.6 to collect the pellet, and finally suspended it in 2 ml LIM for further infection. Using this strategy, the final OD600 was approximately 0.04 for hypocotyl infection. In addition, 2,4-D was also used for cocultivation during calli induction to facilitate the integration of foreign DNA. Additionally, the ability of the transgenic calli to develop into a complete plant was also vital for plant regeneration. Thus, STS, which consisted of 0.1 M Na2S2O3 and 0.1 M AgNO3 at a 4:1 ratio, was added to the CIM to avoid tissue browning, while 3 mg/L AgNO3 was added to the SIM to promote shoot regeneration. Finally, 1 mg/L IBA was added to root formation medium for root generation (detailed methods in “Materials and methods” section). In short, eight different rapeseed varieties were separately transformed via the aforementioned Agrobacterium-mediated hypocotyl transformation in B. napus.
DsRed for rapid and convenient screening during tissue culture
Considering that the expression of DsRed occurred in the whole plant development process, it is presumed that the red fluorescence present in transgenic tissue could be observed through a red filter with a hand-held green fluorescent flashlight. Therefore, the red fluorescence emitted by DsRed might be utilized to screen transgenic-positive tissues in the primary stage of tissue culture. In this way, it was attempted to distinguish the transgenic-positive and transgenic-negative calli and shoots in tissue culture medium. As expected, strong red fluorescence was observed in transgenic-positive calli and shoots (Fig. 3a, b), while nearly no or weaker fluorescence could be observed in transgenic-negative ones (Fig. 3c, d). Moreover, red fluorescence emitted by putative transgenic-positive calli or shoots could also be clearly distinguished under a fluorescence stereomicroscope (Additional file 2). This strong contrast indicated the feasibility of using DsRed as a visual screening marker in tissue culture of B. napus. Thus, plenty of transformed rapeseed hypocotyls, containing calli and shoots, were observed with red fluorescence on CIM and SIM (Figs. 2b–e and 3e–i). Using this method, the transgenic-positive and transgenic-negative shoots could be clearly distinguished, demonstrating the feasibility of screening positive regenerative plantlets conveniently with a visual screening method. Furthermore, the calli, which could be observed with red fluorescence, were marked and counted under a hand-held green fluorescent flashlight. The calli with red fluorescence ranged from 50.8 to 71.4%, i.e., 56.9% (78/137) for E1, 50.6% (66/131) for E2, 66.9% (73/109) for R1, 50.8% (97/191) for R2 and 71.4% (167/234) for R3 (Table 1). These results showed that the average transformation efficiency was achieved to 59.1%, and they revealed that the visual screening method by red fluorescence provided an obvious advantage for rapid identification and convenient discrimination during the early stage of tissue culture.
Fig. 3.
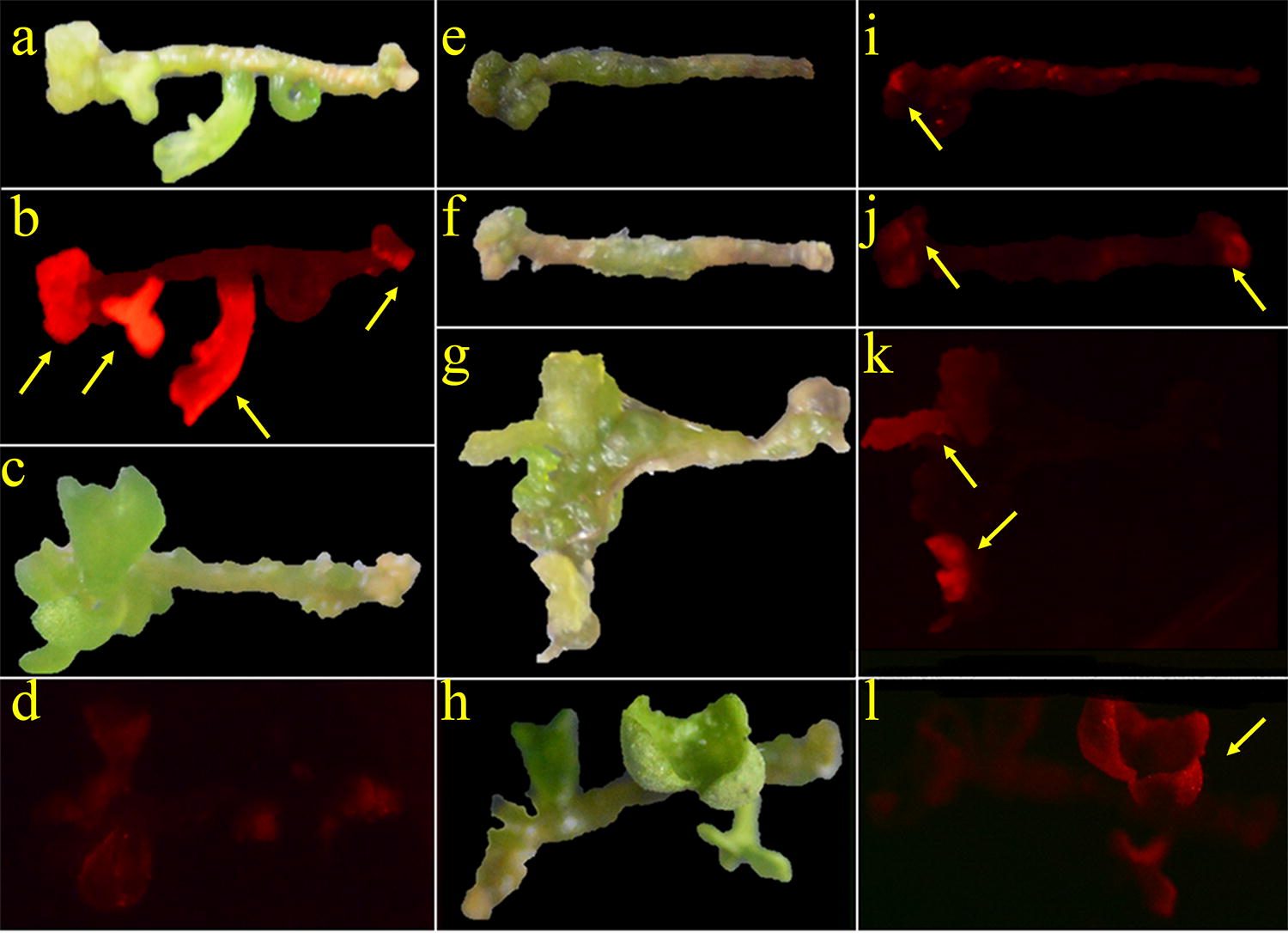
Efficient and convenient screening in the primary stage of tissue culture. a, b Examples of positive tissue with red fluorescence by visual screening. Strong red fluorescence could be observed in calli and shoots. c, d Examples of negative tissue with red fluorescence by visual screening. Nearly no or weak red fluorescence in calli and shoots could be observed. e–l Diagram of the convenient screening for calli and shoots at the primary culturing stage. Red fluorescence could be clearly observed in calli and shoots by visual screening with a hand-held green fluorescent flashlight. The yellow arrows indicate red fluorescence. a, c, e–h are in the white field, while b, d, i, g, k and l are in the excitatory light field
Fig. 2.

Overview of the efficient method for the high-efficiency acquisition of transgenic lines combined with a visual screening method compared with antibiotics screening in B. napus. a Highly efficient genetic transformation combined with a convenient visual screening marker DsRed via Agrobacterium-mediated hypocotyl transformation in B. napus. Hypocotyls were transformed with a vector accompanied by DsRed and hygromycin via Agrobacterium- mediated transformation. The light grey lines inside the oval represent hypocotyls from B. napus. b , c Calli formation and screening during the callus-induced stage with or without the visual screening method. The calli would survive under antibiotics screening with hygromycin in b, and they were further picked by the visual screening using a hand-held green fluorescent flashlight in c. The endpoints in the line represent the formed callus, and the red spots in the endpoints represented the red fluorescence observed in the calli. d ,e Shoots formation in shoot-induced medium with or without the visual screening method. The calli would survive under antibiotics screening with hygromycin in d, and they were further picked for visual screening using a hand-held green fluorescent flashlight in e. The red spots in the shoots represent the red fluorescence observed in the cotyledons. f, g highly efficient screening of the transgenic-positive rooted T0 plantlets and T1 transgenic-positive plants. Transgenic-positive rooted T0 plantlets and T1 transgenic-positive plants could be rapidly and high efficiently obtained when combined with the convenient visual screening method. h Visual screening in cotyledons, hypocotyls, seed coats and roots during seed germination. The seedlings could be screened with the visual marker DsRed using laser confocal fluorescence microscopy
Table 1.
Transformation efficiency of red fluorescence in different lines
| Target gene | Number of experiments | Number of hypocotyls inoculated | Hypocotyls with DsRed | Transformation efficiency (%) |
|---|---|---|---|---|
| BnaA07g17400D | 1 | 77 | 47 | 61.0 |
| 2 | 60 | 31 | 51.7 | |
| BnaC05g34170D | 1 | 57 | 30 | 52.6 |
| 2 | 74 | 36 | 48.6 | |
| R1 | 1 | 59 | 35 | 59.3 |
| 2 | 50 | 38 | 76.0 | |
| R2 | 1 | 92 | 47 | 51.1 |
| 2 | 99 | 50 | 50.5 | |
| R3 | 1 | 143 | 109 | 76.2 |
| 2 | 91 | 58 | 63.7 |
DsRed for efficient genetic transformation during tissue culture
To investigate whether the calli that emitted red fluorescence were integrated with the desired objective DNA fragments, the potential transgenic calli and shoots were randomly selected for identification by PCR amplification. Hypocotyls of three commercial rapeseed varieties (Zhong shuang 8, Zhong shuang 11, Zhong you 821), a spring rapeseed variety (Jia 9709) and a semi-winter variety (Jia 2016) were transformed with R2 vector. As expected, almost all the commercial cultivars carried the DsRed amplification products, i.e., 95.5% (21/22), 100% (22/22), 95.5% (21/22), 100% (22/22) and 100% (22/22) were identified with PCR products having a length of 426 bp (Additional file 3a). This high-efficiency genetic transformation system showed that it was a successful attempt in different rapeseed varieties with visual screening showing red fluorescence from the induced calli to shoots.
Additionally, to further identify whether this transformation system was also appropriate for other rapeseed varieties with different expression vectors, hypocotyls from another three commercial rapeseed varieties (7633, B 351 and Shan 3B), which were transformed with E1 vector, were also analysed by PCR detection. The results showed that 83.3% (15/18), 94.4% (17/18) and 100% (18/18) were amplified at the length of 722 bp for the BnaA07g17400D gene (Additional file 3b). Consequently, these results showed that the different commercial rapeseed varieties and different expression vectors used in this study were competent for transformation using the Agrobacterium-mediated hypocotyl transformation method (Fig. 2). Thus, an average of 96.1% of the tissues observed with red fluorescence could indeed be checked with the objective gene in eight different rapeseed varieties. These results verified that the visual screening method with DsRed could be used to shorten the screening period and accelerate the regeneration process to obtain high-efficiency positive transgenic plantlets.
High-efficiency for rooted transgenic-positive plantlets in rapeseed
To investigate the transgenic-positive efficiency via the visual screening method based on Agrobacterium-mediated transformation in rapeseed, plantlets transformed with E1, E2, R1, R2 and R3 vectors that were regenerated from hypocotyls of “Jia 2016” were collected, respectively. Approximately 196 independent rooted transgenic plantlets were obtained, and all these positive transgenic plants had a normal phenotype both in vegetative growth and reproductive growth; no sterile transgenic plants were found in the field.
Next, genomic DNA was extracted from putative transgenic rapeseed plantlets, and PCR amplification was performed to detect the integration of DsRed, BnaA07g17400D and BnaC05g34170D. As expected, most of the putative transgenic lines was presented positive bands on the agarose gel (Fig. 4). To be specific, 80% (8/10) and 90.7% (49/54) were detected for BnaA07g17400D and BnaC05g34170D amplification, respectively (Table 2). In addition, 88% (22/25), 81.4% (35/43) and 89.1% (57/64) were detected for R1, R2 and R3 by DsRed fragment amplification, respectively (Table 2). Hence, five different expression vectors, including over-expression and RNAi-mediated knock-down, could be efficiently transformed into rapeseed with the Agrobacterium-mediated hypocotyl transformation system.
Fig. 4.

Transgenic-positive plant identification in T0 plants. a Amplification of the BnaA07g17400D gene in E1 transformed lines. b Amplification of the BnaC05g34170D gene in E2 transformed lines. c–e Amplification of DsRed gene in R1, R2 and R3 transformed lines. The light bands indicated by the arrow indicate the size of the amplified genes noted on the left side. P, plasmid. WT, wild type plant. Marker, DL 100 bp ladder
Table 2.
Transformation efficiency using different expression vectors in J2016
| Total | Transgenic positive | Transformation efficiency (%) | |
|---|---|---|---|
| BnaA07g17400D | 10 | 8 | 80.0 |
| BnaC05g34170D | 54 | 49 | 90.7 |
| R1 | 25 | 22 | 88.0 |
| R2 | 43 | 35 | 81.4 |
| R3 | 64 | 57 | 89.1 |
DsRed for convenient and efficient screening during seed germination
Considering the thick tissue in B. napus, it is not easy to distinguish the positive transgenic seeds and seedlings with a hand-held green fluorescent flashlight after obtaining T1 seeds and regeneration plants. Red fluorescence, unlike green fluorescence, can be observed under laser confocal fluorescence microscopy without the interference of chlorophyll. To verify the screening feasibility during the primary stage of seed germination, experiments were performed using a confocal microscope. For each part, seed coats, roots, hypocotyls and cotyledons, were randomly selected for further analysis (Figs. 2h and 5a–e). It was revealed that the red fluorescence could be successfully observed in all the cells of different parts of the seedings (Fig. 5f–m).
Fig. 5.

Convenient screening during seed germination. a–m Observation of red fluorescence through the primary stage of seedling. The red fluorescence could be observed inthe cotyledon (b, f, j), hypocotyl (c, g, k), seed coat (d, h, l) and root (e, i, m) of rapeseed seedlings under a laser confocal fluorescence microscope FV1000, respectively. n RT-PCR identification of the relative gene expression in seedlings. The upper diagram represents the relative gene expression of DsRed in seedlings from different RNAi lines. The middle diagram represents the relative gene expression of BnaA07g17400D in seedlings from different RNAi lines. The lower diagram represents the relative gene expression of DsRed in roots, seed coats, hypocotyls and cotyledons from seedlings of different RNAi lines
Furthermore, RT-PCR analysis was performed to check the relative expression of the marker gene—DsRed and the target gene—BnaA07g17400D in the seedlings during seed germination (Fig. 5n). As expected, the exogenous marker gene—DsRed showed significant up-regulation in different transgenic lines, and achieved to even 1200 times higher levels compared with WT. Regarding the inferenced endogenous gene, the relative gene expression of BnaA07g17400D showed obvious down-regulation in the transgenic seedlings from different RNAi lines. Additionally, in the roots, seed coats, hypocotyls and cotyledons of seedlings from the aforementioned RNAi lines, the relative expressions of DsRed presented a comparable up-regulation in various organs, especially in cotyledons, which indicated that red fluorescence could be more easily detected in cotyledons. In brief, DsRed, as a marker gene, is reliable for positive screening without influencing the desired RNAi events.
Gene copy number identification of DsRed by southern blotting
To verify whether DsRed could be applied to identify the integrated copy number in regenerated plants, Southern blotting was used to detect the integration of DsRed cassettes in several transgenic plants. Three T3 plants for each line were randomly chosen for Southern blotting, and eight were confirmed to contain the insertion of the DsRed gene (Fig. 6). Southern blotting results revealed that the DsRed gene showed one—to—five copies in the transgenic rapeseed plants. More specifically, in the E1 lines, all three plants were inserted with three copies. In the R1 lines, one plant was inserted with a single copy and two plants with two copies. In the R2 lines, two plants were inserted with three copies, and the third plant lacked an insertion, which indicated that R2-20–10-1 was a marker-free transgenic plant (Fig. 6a). In the E2 lines, all three plants carried a single copy. In the R3 lines, two plants had five copies and one plant had three copies (Fig. 6b).
Fig. 6.

Southern blot analysis of DsRed genes for copy number identification in T3 transgenic rapeseed plants. a Southern blot analysis of BnaA07g17400D over-expression and knock down. b Southern blot analysis of BnaC05g34170D over-expression and knock down. WT, wild type; P-E1/E2, over-expression plasmid pCMABIA-1303; P-R1/R2/R3, knock down plasmid p35S-1390; M: λ DNA/Hind III- Plus DNA marker
It is worth noting that all E1 lines that stemmed from the same parent (E1-15–21) presented the same three T-DNA copies (Fig. 6a). Similarly, all the E2 lines from one T1 plant (E2-9) had one copy, and the two R2 lines from the same T2 plants (R2-20–6) also contained three T-DNA copies (Fig. 6a). These findings indicated that E1-15-21, R2-20-6 and E2-9 could be regarded as stable and homozygous transgenic plants. However, in R1 and R3 lines, the T-DNA copy number showed diversity in different plants), and the copy number of the third R2 plant (R2-20-10-1) also differed from the previous two plants because of the different parents (Fig. 6a). These results demonstrated that the T-DNA copy numbers were invariable between the same T2 lines (E1-15-21 and R2-20-6) but variable among the different T2 lines (R1-3-2, R1-3-19, R1-5-19 and R2-20-10), illustrating that stable transgenic plants could be obtained in T3 plant, even in tetraploid rapeseed plants.
Discussion
Over the last century, many attempts have been conducted in Agrobacterium-mediated transformation of rapeseed [5, 41–44]. However, few studies have been focused on the concentration of Agrobacterium. Previously, the concentration for Agrobacterium infection was controlled by adjusting the final OD600 [5]. Here, not the final OD600 adjustment but the calculation of the volume from the original Agrobacterium solution was performed, and different commercial rapeseed varieties, different expression vectors and different genes were transformed efficiently in this way. It is noteworthy that the OD600 was only detected once for the inoculated Agrobacterium, and the pipette volume for further experiments could be easily calculated by the aforementioned method. Previously, the final OD600 was adjusted to a fixed value in other studies, such as 0.05, 0.2 or 1, based on the original OD600 for explants infection [25, 26, 43]. However, the original OD600 usually ranged from 0.4 to 1.2 in this investigation, owning to the differences in the original inoculum, bacterial concentration, and bacterial activity. Considering the sensitivity of explant to the Agrobacterium, the final concentration was found to be easily controlled by the volume of the original Agrobacterium solution rather than an adjustment to avoid explants turning necrotic. Therefore, ensuring the appropriate bacterial activity of collected Agrobacterium was of great significance. Moreover, the timing of incubation infection and cocultivation stage should be carefully controlled, as excessive attachment can directly lead to explant necrosis or reduced regeneration [5, 45]. Usually, 10 min was used for hypocotyls infection, with maintenance for 36–48 h in the cocultivation medium in this investigation. Sometimes, the time for infection could be extended or shortened by 1–2 min appropriately and flexibly if the collected Agrobacterium or the hypocotyls were not in good condition. After 24 h of cocultivation in cocultivation medium, the hypocotyls should be tough and bright white, which can be transferred to CIM 24 h later. If the hypocotyls do not show the aforementioned features, it was suggested that they should be transferred to the CIM immediately or up to 12 h later to avoid explant necrosis. In this way, highly efficient transformation was obtained in eight rapeseed varieties. Previously, 2,4-D was reported to be added to the cocultivation medium to facilitate the integration of foreign DNA, owing to its ability to enable cell division at the incision [46]. Additionally, AgNO3 is known to promote the generation and elongation of shoot primordia, and increase the frequency of adventitious shoot differentiation [42]. Thus, an average of 85.8% transgenic-positive rooted plantlets is obtained with the addition of 1 mg/L 2,4-D to CIM and 3 mg/L AgNO3 to SIM. No obvious difference was observed in the transformation efficiency among different vectors, except for E1, which was utilized in our exploratory study for optimum experimental conditions. Therefore, the establishment of an efficient Agrobacterium-mediated rapeseed transformation system using hypocotyls will also enable genome editing of valuable genes controlling important traits.
Based on the well-established transformation system with high transformation efficiency, an effective transgenic-positive screening technology is of great importance for shortening the period of tissue culture and simplifying the transformation system in rapeseed (Fig. 2). DsRed, as a selectable marker gene for transgenic seed screening, is advantageous for separating transgenic and non-transgenic seeds in many plants, such as Arabidopsis, rice and Camelina sativa [15, 47, 48]. In Arabidopsis, DsRed has been used as a visual selection marker to promote transgenic seed screening [33, 49, 50]. However, considering the impediment of tissue thickness, DsRed has not been used in rapeseed for convenient screening to accelerate the process of transgenic-positive plantlet selection.
The red fluorescence emitted by DsRed marked calli and shoots could be easily observed through a red filter using a hand-held green fluorescent flashlight. Up to 61.2% of the calli were observed with red fluorescence, and almost all of these calli could be detected with the DsRed gene by PCR analysis. The red fluorescence in calli and rapeseed seedlings could also be observed under a fluorescence stereomicroscope and laser scanning confocal microscope, respectively. Moreover, it is worth noting that the fluorescence intensities varied from each other, potentially due to the rapeseed genotypes, i.e., homo- or hemizygosity of the transgene [50]. Additionally, DsRed, as a marker for Southern blotting analysis, was also successfully applied for estimating one to five copies in the transgenic offspring plants, as in other studies [41, 42]. These results indicated that the visual screening marker DsRed is a great success in accelerating the genetic transformation system in B. napus.
In addition, several methods provide alternatives for screening transgenic positive plants (Fig. 2 and Table 3). Antibiotic screening is frequently used in most transformation studies, such as kanamycin, hygromycin or herbicide [26, 51, 52]. However, the dosage of these substances might influence the regeneration rate and further break the balance of transgenic/non-transgenic seeds. PCR amplification is time and cost consuming in plant preparation and identification. Southern blot analysis, as a traditional method for identification of the transgenic copy number, is costly in terms of reagents, equipment, time and labour [40]. The GUS expression assay is a compromise in the matter of cost. However, it usually requires a long time for tissue decolouring. Moreover, the GUS expression assay is often performed following the expression of target gene in plant cells or tissues [53]. As for GFP, it is usually used for determining sub-cellular localization with a laser confocal fluorescence microscope, which should be associated with RFP or YFP due to its instability [54, 55]. Otherwise, it can also be detected with a fluorescence microscope and a green fluorescence pigment filter using tissue stained with fluorescein di-acetate (FDA), which induces a decreased fluorescence intensity [54, 56]. Nevertheless, the FDA assay has hardly used in plants to our limited knowledge. Compared with previous methods, the present study is the first to introduce the visual selectable marker gene DsRed to accelerate transgenic plantlets screening. Only a combination of light filter with hand-held fluorescent flashlight to easily distinguish the transgenic-positive and transgenic-negative plants during the primary stage of tissue culture and seed germination in rapeseed. Therefore, the application of DsRed, which served as a marker gene, will facilitate the screening of transgenic positive lines and the utilization of tissue culture or cell culture in plant biotechnology.
Table 3.
Comparisons of different screening methods in plants
| Methods | Special reagents | Equipments | Total time (D) /1000 samples | Cost/200 reaction | Labour | Injuries to plant |
|---|---|---|---|---|---|---|
| PCR amplification | DNA extarct kit | Thermal cycler | 2 | High | High | Leaf |
| Antibiotic/Herbicide selection | Autobiotics | – | ≥ 2 | Low | Moderate | Seed |
| Southern blot analysis | Southern blot hybridization kit | Hybridisation oven | ≥ 7 | High | High | Leaf |
| Gus expression assays | GUS dye | Microscopy | ≥ 2 | Moderate | Moderate | Leaf/Root |
| GFP | Fluorescein di-acetatea | Laser scanning confocal microscopy/green fluorescence pigment | ≥ 0.5 | Moderate | Moderate | Leaf/Root |
| Visual screening | No | Hand-held fluorescent flashlight, light filter | ≤ 0.5 | Zero | Low | No |
aIt has mostly been used in microorganisms
Conclusion
A convenient and highly efficient method was developed that combines an efficient genetic transformation system with a convenient and rapid transgenic-positive visual screening method in rapeseed. Using this method, potential transgenic-positive plants could be quickly and efficiently selected by a convenient visual screening method, which greatly accelerated the regeneration process. Additionally, a high transgenic positive efficiency could be easily achieved based on the optimized Agrobacterium-mediated hypocotyl transformation, which greatly increased the number of genetic transformed plants. The aforementioned system will be a good alternative for rapeseed genetic transformation and gene functional studies.
Supplementary information
Additional file 1. Formation of calli to a regenerated plantlet on a hypocotyl. a and b calli that were induced from hypocotyls. c Calli that were prepared for swollen to shoots. d and e Shoot formation and cotyledon development accompanied by the appearance of a growth point.
Additional file 2. Visual screening of transgenic-positive and/or -negative tissues under a fluorescence stereomicroscope. Red fluorescence could be observed under fluorescence stereomicroscope to facilitate visual screening in tissue culture medium. The image was captured in bright field a and its corresponding dark field b, respectively. The tissues indicated by the arrow represent transgenic-positive calli.
Additional file 3. PCR amplification for transgenic-positive identification in calli. a Amplification of DsRed from calli of Jia 9709, Jia 2016, Zhong shuang 8, Zhong shuang 11 and Zhong you 821, respectively. P, plasmid. WT, wild type plant. Marker, DL 100 bp ladder. b Amplification of BnaA07g17400D from calli in 7633, B 351 and Shan 3B, respectively. P, plasmid. N, wild type plant. Marker, DL 2000 bp.
Additional file 4. Primers used in this study.
Acknowledgements
Our thanks are extended to Prof. Chunyu Zhang from Huazhong Agriculture University and Prof. Dianrong Li from the Hybrid Rapeseed Center of Shaanxi Province for their contributions in offering the seeds of different rapeseed varieties.
Authors’ contributions
KZ participated in the entire experiment and wrote the manuscript. JH carried out the transformation and identification of the winter rapeseed varieties (7633, B 351 and Shan 3B) and modified the manuscript. LL, RX and LQ participated in the transformation and identification of the commercial rapeseed varieties (Zhong shuang 8, Zhong shuang 11, Zhong you 821). XL and WY participated in some of the tissue culture experiments. KC and YY provided helpful suggestions for the study. MMMK and AAS participated in the PCR detection. ML and CF designed, led and coordinated the overall study. KZ and JH contributed equally to this paper. All authors read and approved the final manuscript.
Funding
The work was supported by the National Natural Science Foundation of China (31671721) and the National key research and Development Program (2016YFD0101300).
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Kai Zhang and Jianjie He contributed equally to this work
Contributor Information
Chunhua Fu, Email: fuch2003@126.com.
Maoteng Li, Email: limaoteng426@hust.edu.cn.
Supplementary information
Supplementary information accompanies this paper at 10.1186/s13007-020-00585-6.
References
- 1.Gacek K, Bartkowiak-Broda I, Batley J. Genetic and molecular regulation of seed storage proteins (SSPs) to improve protein nutritional value of oilseed rape (Brassica napus L.) seeds. Front Plant Sci. 2018;9:890. doi: 10.3389/fpls.2018.00890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chalhoub B. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science. 2014;345:1255. doi: 10.1126/science.1253435. [DOI] [PubMed] [Google Scholar]
- 3.Tester M, Langridge P. Breeding technologies to increase crop production in a changing world. Science. 2010;327:818–822. doi: 10.1126/science.1183700. [DOI] [PubMed] [Google Scholar]
- 4.Zhai Y, Yu K, Cai S, Hu L, Amoo O, Xu L, et al. Targeted mutagenesis of BnTT8 homologs controls yellow seed coat development for effective oil production in Brassica napus L. Plant Biotechnol J. 2019 doi: 10.1111/pbi.13281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bhalla PL, Singh MB. Agrobacterium-mediated transformation of Brassica napus and Brassica oleracea. Nat Protoc. 2008;3:181–189. doi: 10.1038/nprot.2007.527. [DOI] [PubMed] [Google Scholar]
- 6.Li T, Liu B, Spalding MH, Weeks DP, Yang B. High-efficiency TALEN-based gene editing produces disease-resistant rice. Nat Biotechnol. 2012;30:390–392. doi: 10.1038/nbt.2199. [DOI] [PubMed] [Google Scholar]
- 7.Kuma KM, Lopes-Caitar VS, Romero CCT, Silva SMH, Kuwahara MK, Carvalho MCCG, et al. A high efficient protocol for soybean root transformation by Agrobacterium rhizogenes and most stable reference genes for RT-qPCR analysis. Plant Cell Rep. 2015;34:1987–2000. doi: 10.1007/s00299-015-1845-2. [DOI] [PubMed] [Google Scholar]
- 8.Huang SS, Gilbertson LA, Adams TH, Malloy KP, Reisenbigler EK, Birr DH, et al. Generation of marker-free transgenic maize by regular two-border Agrobacterium transformation vectors. Transgenic Res. 2004;13:451–461. doi: 10.1007/s11248-004-1453-3. [DOI] [PubMed] [Google Scholar]
- 9.Wang P, Zhang J, Sun L, Ma Y, Xu J, Liang S, et al. High efficient multi-sites genome editing in allotetraploid cotton (Gossypium hirsutum) using CRISPR/Cas9 system. Plant Biotechnol J. 2017;16:137–150. doi: 10.1111/pbi.12755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.James C. Global Status of Commercialized Biotech/GM Crops in 2018: biotech crops continue to help meet the challenges of increased population and climate change. ISAAA: Ithaca, NY. 2018. ISAAA Brief No. 54.
- 11.Cigan AM, Unger-Wallace E, Haug-Collet K. Transcriptional gene silencing as a tool for uncovering gene function in maize. Plant J. 2005;43:929–940. doi: 10.1111/j.1365-313X.2005.02492.x. [DOI] [PubMed] [Google Scholar]
- 12.Bashandy H, Jalkanen S, Teeri TH. Within leaf variation is the largest source of variation in agroinfiltration of Nicotiana benthamiana. Plant Methods. 2015;11:47. doi: 10.1186/s13007-015-0091-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Boccacci P, Mela A, Mina CP, Chitarra W, Perrone I, Gribaudo I, et al. Cultivar-specific gene modulation in Vitis vinifera: analysis of the promoters regulating the expression of WOX transcription factors. Sci Rep. 2017;7:45670. doi: 10.1038/srep45670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wang K, Liu H, Du L, Ye X. Generation of marker-free transgenic hexaploid wheat via an Agrobacterium-mediated co-transformation strategy in commercial Chinese wheat varieties. Plant Biotechnol J. 2017;15:614–623. doi: 10.1111/pbi.12660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chang ZY, Chen ZF, Wang N, Xie G, Lu JW, Yan W, et al. Construction of a male sterility system for hybrid rice breeding and seed production using a nuclear male sterility gene. P Natl Acad Sci USA. 2016;113:14145–14150. doi: 10.1073/pnas.1613792113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Golz C, Kohler F, Schieder O. Transfer of hygromycin resistance into Brassica napus using total DNA of a transgenic B nigra line. Plant Mol Biol. 1990;15:475–483. doi: 10.1007/BF00019164. [DOI] [PubMed] [Google Scholar]
- 17.Köhler F, Cardon G, Pöhlman M, Gill R, Schieder O. Enhancement of transformation rates in higher plants by low-dose irradiation: are DNA repair systems involved in the incorporation of exogenous DNA into the plant genome. Plant Mol Biol. 1989;12:189–199. doi: 10.1007/BF00020504. [DOI] [PubMed] [Google Scholar]
- 18.Bergman P, Glimelius K. Electroporation of rapeseed protoplasts - transient and stable transformation. Physiol Plantarum. 1993;88:604–611. doi: 10.1111/j.1399-3054.1993.tb01378.x. [DOI] [PubMed] [Google Scholar]
- 19.Chen JL, Beversdorf WD. A combined use of microprojectile bombardment and DNA imbibition enhances transformation frequency of canola (Brassica napus L.) Theor Appl Genet. 1994;88:187–192. doi: 10.1007/BF00225896. [DOI] [PubMed] [Google Scholar]
- 20.Palmer CE, Keller WA, Shivanna KR, Sawhney VK. Pollen embryos. Pollen Biotechnol Crop Prod Improv. 1997; 392
- 21.Jones-Villeneuve E, Huang B, Prudhomme I, Bird S, Kemble R, Hattori J, et al. Assessment of microinjection for introducing DNA into uninuclear microspores of rapeseed. Plant Cell Tiss Org. 1995;40:97–100. doi: 10.1007/BF00041124. [DOI] [Google Scholar]
- 22.Perez-Martin F, Yuste-Lisbona FJ, Pineda B, Angarita-Diaz MP, Garcia-Sogo B, Anton T, et al. A collection of enhancer trap insertional mutants for functional genomics in tomato. Plant Biotechnol J. 2017;15:1439–1452. doi: 10.1111/pbi.12728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sheikholeslam SN, Weeks DP. Acetosyringone promotes high efficiency transformation of Arabidopsis thaliana explants by Agrobacterium tumefaciens. Plant Mol Biol. 1987;8:291–298. doi: 10.1007/BF00021308. [DOI] [PubMed] [Google Scholar]
- 24.Cardoza V, Stewart CN. Increased Agrobacterium -mediated transformation and rooting efficiencies in canola (Brassica napus L.) from hypocotyl segment explants. Plant Cell Rep. 2003;21:599–604. doi: 10.1007/s00299-002-0560-y. [DOI] [PubMed] [Google Scholar]
- 25.Maheshwari P, Selvaraj G, Kovalchuk I. Optimization of Brassica napus (canola) explant regeneration for genetic transformation. New Biotechnol. 2011;29:144–155. doi: 10.1016/j.nbt.2011.06.014. [DOI] [PubMed] [Google Scholar]
- 26.Zhang Y, Singh MB, Swoboda I, Bhalla PL. Agrobacterium-mediated transformation and generation of male sterile lines of Australian canola. Aust J Agric Res. 2005;56:353. doi: 10.1071/AR04175. [DOI] [Google Scholar]
- 27.Matz MV, Fradkov AF, Labas YA, Savitsky AP, Zaraisky AG, Markelov ML, et al. Fluorescent proteins from nonbioluminescent Anthozoa species. Nat Biotechnol. 1999;17:969–973. doi: 10.1038/13657. [DOI] [PubMed] [Google Scholar]
- 28.Yarbrough D, Wachter RM, Kallio K, Matz MV, Remington SJ. Refined crystal structure of DsRed, a red fluorescent protein from coral, at 2.0-angstrom resolution. P Natl Acad Sci USA. 2001;98:462–467. doi: 10.1073/pnas.98.2.462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Baird GS, Zacharias DA, Tsien RY. Biochemistry, mutagenesis, and oligomerization of DsRed, a red fluorescent protein from coral. P Natl Acad Sci USA. 2000;97:11984–11989. doi: 10.1073/pnas.97.22.11984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Verkhusha VV, Chudakov DM, Gurskaya NG, Lukyanov S, Lukyanov KA. Common pathway for the red chromophore formation in fluorescent proteins and chromoproteins. Chem Biol. 2004;11:845–854. doi: 10.1016/j.chembiol.2004.04.007. [DOI] [PubMed] [Google Scholar]
- 31.Okwuonu IC, Achi OK, Egesi CN, Taylor NJ. Evaluation of red fluorescent protein (DsRed) as alternative visual marker of genetic transformation in cassava (Manihot esculenta Crantz) Vitro Cell Dev-Pl. 2015;51:571–579. doi: 10.1007/s11627-015-9718-5. [DOI] [Google Scholar]
- 32.Jach G, Binot E, Frings S, Luxa K, Schell J. Use of red fluorescent protein from Discosoma sp (dsRED) as a reporter for plant gene expression. Plant J. 2001;28:483–491. doi: 10.1046/j.1365-313X.2001.01153.x. [DOI] [PubMed] [Google Scholar]
- 33.Stuitje AR, Verbree EC, van der Linden KH, Mietkiewska EM, Nap JP, Kneppers TJA. Seed-expressed fluorescent proteins as versatile tools for easy (co)transformation and high-throughput functional genomics in Arabidopsis. Plant Biotechnol J. 2003;1:301–309. doi: 10.1046/j.1467-7652.2003.00028.x. [DOI] [PubMed] [Google Scholar]
- 34.Wada S, Ishida H, Izumi M, Yoshimoto K, Ohsumi Y, Mae T, et al. Autophagy plays a role in chloroplast degradation during senescence in individually darkened leaves. Plant Physiol. 2009;149:885–893. doi: 10.1104/pp.108.130013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wang Z, Xu C, Benning C. TGD4 involved in endoplasmic reticulum-to-chloroplast lipid trafficking is a phosphatidic acid binding protein. Plant J. 2012;70:614–623. doi: 10.1111/j.1365-313X.2012.04900.x. [DOI] [PubMed] [Google Scholar]
- 36.Yelina NE, Ziolkowski PA, Miller N, Zhao XH, Kelly KA, Munoz DF, et al. High-throughput analysis of meiotic crossover frequency and interference via flow cytometry of fluorescent pollen in Arabidopsis thaliana. Nat Protoc. 2013;8:2119–2134. doi: 10.1038/nprot.2013.131. [DOI] [PubMed] [Google Scholar]
- 37.Eckert M, Maguire K, Urban M, Foster S, Fitt B, Lucas J, et al. Agrobacterium tumefaciens-mediated transformation of Leptosphaeria spp. and Oculimacula spp. with the reef coral gene DsRed and the jellyfish gene gfp. Fems Microbiol Lett. 2005;253:67–74. doi: 10.1016/j.femsle.2005.09.041. [DOI] [PubMed] [Google Scholar]
- 38.Zheng X, Lopisso DT, Eseola AB, Koopmann B, von Tiedemann A. Potential for seed transmission of Verticillium longisporum in Oilseed Rape (Brassica napus) Plant Dis. 2019;8:129–130. doi: 10.1007/s00425-019-03154-x. [DOI] [PubMed] [Google Scholar]
- 39.Zhou YM, Wang H, Gilmer S, Whitwill S, Keller W, Fowke LC. Control of petal and pollen development by the plant cyclin-dependent kinase inhibitor ICK1 in transgenic Brassica plants. Planta. 2002;215:248–257. doi: 10.1007/s00425-002-0752-2. [DOI] [PubMed] [Google Scholar]
- 40.Stefano B, Patrizia B, Matteo C, Massimo G. Inverse PCR and quantitative PCR as alternative methods to southern blotting analysis to assess transgene copy number and characterize the integration site in transgenic woody plants. Biochem Genet. 2016;54:291–305. doi: 10.1007/s10528-016-9719-z. [DOI] [PubMed] [Google Scholar]
- 41.Radke SE, Andrews BM, Moloney MM, Crouch ML, Kridl JC, Knauf VC. Transformation of Brassica napus L. using Agrobacterium tumefaciens: developmentally regulated expression of a reintroduced napin gene. Theor Appl Genet. 1988;75:685–94. doi: 10.1007/BF00265588. [DOI] [Google Scholar]
- 42.Block M, De Brouwer D, Tenning P. Transformation of Brassica napus and Brassica oleracea using Agrobacterium tumefaciens and the expression of the bar and neo genes in the transgenic plants. Plant physiol. 1989;91:694–701. doi: 10.1104/pp.91.2.694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Damgaard O, Jensen LH, Rasmussen OS. Agrobacterium tumefaciens-mediated transformation of Brassica napus winter cultivars. Transgenic Res. 1997;6:279–288. doi: 10.1023/A:1018458628218. [DOI] [Google Scholar]
- 44.Liu XX, Lang SR, Su LQ, Liu X, Wang XF. Improved Agrobacterium-mediated transformation and high efficiency of root formation from hypocotyl meristem of spring Brassica napus 'Precocity' cultivar. Genet Mol Res. 2015;14:16840–16855. doi: 10.4238/2015.December.14.11. [DOI] [PubMed] [Google Scholar]
- 45.Bhalla PL, Smith N. Agrobacterium tumefaciens-mediated transformation of cauliflower. Brassica oleracea var botrytis Mol Breeding. 1998;4:531–541. doi: 10.1023/A:1009658614579. [DOI] [Google Scholar]
- 46.Moloney MM, Walker JM, Sharma KK. High efficiency transformation of Brassica napus using Agrobacterium vectors. Plant Cell Rep. 1989;8:238–242. doi: 10.1007/BF00778542. [DOI] [PubMed] [Google Scholar]
- 47.Morineau C, Bellec Y, Tellier F, Gissot L, Kelemen Z, Nogue F, et al. Selective gene dosage by CRISPR-Cas9 genome editing in hexaploid Camelina sativa. Plant Biotechnol J. 2017;15:729–739. doi: 10.1111/pbi.12671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chen K, Yin Y, Liu S, Guo Z, Zhang K, Liang Y, et al. Genome-wide identification and functional analysis of oleosin genes in Brassica napus L. BMC Plant Biol. 2019;19:294. doi: 10.1186/s12870-019-1891-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wu G, Rossidivito G, Hu TQ, Berlyand Y, Poethig RS. Traffic lines: new tools for genetic analysis in Arabidopsis thaliana. Genetics. 2015;200:35–45. doi: 10.1534/genetics.114.173435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Shimada TL, Shimada T, Hara-Nishimura I. A rapid and non-destructive screenable marker, FAST, for identifying transformed seeds of Arabidopsis thaliana. Plant J. 2010;61:519–528. doi: 10.1111/j.1365-313X.2009.04060.x. [DOI] [PubMed] [Google Scholar]
- 51.Baskar V, Gangadhar BH, Park SW, Nile SH. A simple and efficient Agrobacterium tumefaciens-mediated plant transformation of Brassica rapa ssp. pekinensis. 3 Biotech. 2016;6:88. doi: 10.1007/s13205-016-0402-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhang R, Liu JX, Chai ZZ, Chen S, Bai Y, Zong Y, et al. Generation of herbicide tolerance traits and a new selectable marker in wheat using base editing. Nat Plants. 2019;5:480–485. doi: 10.1038/s41477-019-0405-0. [DOI] [PubMed] [Google Scholar]
- 53.Kim HU, Li YB, Huang AHC. Ubiquitous and endoplasmic reticulum-located lysophosphatidyl acyltransferase, LPAT2, is essential for female but not male gametophyte development in Arabidopsis. Plant Cell. 2005;17:1073–1089. doi: 10.1105/tpc.104.030403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lentendu G, Hubschmann T, Muller S, Dunker S, Buscot F, Wilhelm C. Recovery of soil unicellular eukaryotes: An efficiency and activity analysis on the single cell level. J Microbiol Meth. 2013;95:463–469. doi: 10.1016/j.mimet.2013.05.006. [DOI] [PubMed] [Google Scholar]
- 55.Schornack S, Fuchs R, Huitema E, Rothbauer U, Lipka V, Kamoun S. Protein mislocalization in plant cells using a GFP-binding chromobody. Plant J. 2009;60:744–754. doi: 10.1111/j.1365-313X.2009.03982.x. [DOI] [PubMed] [Google Scholar]
- 56.Bechet Q, Laviale M, Arsapin N, Bonnefond H, Bernard O. Modeling the impact of high temperatures on microalgal viability and photosynthetic activity. Biotechnol Biofuels. 2017;10:136. doi: 10.1186/s13068-017-0823-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1. Formation of calli to a regenerated plantlet on a hypocotyl. a and b calli that were induced from hypocotyls. c Calli that were prepared for swollen to shoots. d and e Shoot formation and cotyledon development accompanied by the appearance of a growth point.
Additional file 2. Visual screening of transgenic-positive and/or -negative tissues under a fluorescence stereomicroscope. Red fluorescence could be observed under fluorescence stereomicroscope to facilitate visual screening in tissue culture medium. The image was captured in bright field a and its corresponding dark field b, respectively. The tissues indicated by the arrow represent transgenic-positive calli.
Additional file 3. PCR amplification for transgenic-positive identification in calli. a Amplification of DsRed from calli of Jia 9709, Jia 2016, Zhong shuang 8, Zhong shuang 11 and Zhong you 821, respectively. P, plasmid. WT, wild type plant. Marker, DL 100 bp ladder. b Amplification of BnaA07g17400D from calli in 7633, B 351 and Shan 3B, respectively. P, plasmid. N, wild type plant. Marker, DL 2000 bp.
Additional file 4. Primers used in this study.