

Abstract
The present study was an attempt to raise hen egg yolk Ig (IgY) simultaneously directed against Salmonella Enteritidis (SE) and Salmonella Typhimurium (ST) in the same egg yolk. The immunopotentiating effect of 2 different adjuvants—Freund’s adjuvants (FA) and immunostimulating complexes matrix (IM)—on antibody response was also evaluated. Bacterial outer membrane proteins (OMP) were selected as target antigens. The ISA Brown hens, specific-Salmonella spp.-free status, divided into 6 groups were intramuscularly injected with a mono-compound antigen preparation: SE-OMP (treatment SE-FA or SE-IM) or ST-OMP (treatment ST-FA or ST-IM), or a combined antigen preparation: ½ SE-OMP and ½ STOMP (treatment SEST-FA or SEST-IM). Titers of antibodies in yolk were evaluated biweekly with ELISA. There was no antigen × adjuvant interaction on antibody titers. Anti-SE IgY titers in hens that received treatment SEST-FA or SEST-IM were statistically similar (P > 0.05) as compared with those obtained from hens immunized with treatment SE-FA or SE-IM. Anti-ST IgY titers in hens immunized with treatment SEST-FA or SEST-IM were slightly lower than those of hens that received treatment ST-FA or ST-IM. The cross-reactivity of anti-SE IgY, induced by treatment SE-FA or SE-IM, with ST-OMP antigen and that of anti-ST IgY, induced by ST-FA or ST-IM, with SE-OMP antigen were arbitrarily assessed on d 43 and 155 by ELISA. The average cross-reactivity of anti-SE IgY with ST-OMP antigen was 71.7%. The average cross-reactivity of anti-ST IgY with SE-OMP antigen was 78.8%. In FA groups, antibody titers were found higher (P < 0.05) than those in IM groups. Furthermore, no extensive lesions or clinical abnormalities were detected in hens injected with FA. These findings showed the opportunity to raise IgY antibody against 2 Salmonella serovars in the same yolk and that FA was more efficient than IM in mediating antibody response.
Keywords: immunoglobulin Y, Salmonella Enteritidis, Salmonella Typhimurium, Freund’s adjuvant, immunostimulating complexes matrix
INTRODUCTION
The laying hen transfers large amount of antibody, referred to as IgY, to the egg yolk. This antibody gives immunity to its offspring (Klemperer, 1893). Using hen as an immunization host brings a number of advantages. An egg may contain 50 to 100 mg of IgY, of which 2 to 10% are specific antibodies (Schade and Hlinak, 1996). Moreover, for animal welfare, antibody production in hens is a refined method because the antibodies are harvested from the eggs. Therefore, no bleeding of the animal is necessary. The availability of a large amount of IgY from egg yolks makes it feasible to use this antibody in many applications such as passive immunization in human and veterinary medicine, and in diagnostics (Schade et al., 2005).
A pathogen specific antibody can be obtained in large quantities from eggs laid by hyper-immunized hens. Many investigators have reported the possibility of inducing substantial amounts of specific IgY antibody against a single antigen (for a review, see Schade et al., 2005). However, to our knowledge, the possibility of raising specific IgY simultaneously directed against various antigens has not been previously reported, particularly in the case of Salmonella spp.
The immunogenicity of an antigen is influenced by several factors, including the species or strain being immunized, antigen properties and dosage, the route of administration, and adjuvant (Kuby, 2007). Freund’s adjuvant (FA), which is the most effective and common adjuvant used for antibody production in laboratory animals, leads generally to severe inflammation at the injection site. This disadvantage is rather linked to its complete form (Freund’s complete adjuvant: FCA) than to the incomplete one (Freund’s incomplete adjuvant; Leenaars et al., 1994, 1998). In birds, FCA, which is usually used at the first immunization, does not seem to result in the same severe lesions as in mammals. Studies using FCA in the laying hen for antibodies production did not mention any pathological lesions associated with this adjuvant (Svendsen et al., 1996; Kapoor et al., 2000; Li et al., 2006). However, the majority of Animal Care and Use Committees encourage the use of alternative adjuvants rather than the FA. The immunostimulating complexes matrix (IM) is a relatively new adjuvant that has been shown to induce an efficient humoral immune response in animal models and human clinical trials (Pearse and Drane, 2005).
The aim of the present study was to conduct a comparative immunization assay to assess the possibility of inducing an efficient humoral immune response against 2 Salmonella serovars, Salmonella Enteritidis (SE) and Salmonella Typhimurium (ST), in the same egg yolk. These serovars were selected because they are the main cause of salmonellosis in humans. Bacterial outer membrane proteins (OMP) were selected as the target antigens. The efficacy of 2 different adjuvants FA and IM on antibody titer in chicken egg yolk was also analyzed.
MATERIALS AND METHODS
Bacteria and Growth Conditions
The SE and ST strains used in this study were obtained from the WCIB (Walloon Center of Industrial Biology, Gembloux, Belgium) collection. Salmonella cells were grown in Nutrient broth (231000, Difco Laboratories, Detroit, MI) at 37°C for 16 to 18 h with agitation (130 rpm). Following incubation, cells were harvested by centrifugation at 7,000 × g for 30 min at 4°C. Cells were then washed 1 time with sterile double-distilled water and 2 times with sterile 10 mM Tris-HCl buffer (pH: 7.8; T5941, Sigma Chemical Co., St. Louis, MO), and subsequently resuspended in Tris-HCl buffer containing 10 mM EDTA (Sigma Chemical Co.; Tris-HCl/EDTA buffer).
Preparation of Outer Membrane Proteins
The OMP from Salmonella were prepared as described by Nurminen (1985) with some modifications. The resupended bacteria were treated with lysozyme (L76R5, Sigma Chemical Co.; 0.5 mg/mL). After incubation for 1 h at 37°C, ribonuclease (R6513, Sigma Chemical Co.; 30 μg/mL) was added and the sample was allowed to stand for 10 min at room temperature. Then, 1 M MgCl2 solution (40 μL/mL) and deoxyribonuclease (D5025, Sigma Chemical Co.; 30 μg/mL) were added. After incubation for 5 min at room temperature, the mixture was centrifuged at 1,600 × g for 10 min at 4°C. The pellet, corresponding to bacterial envelopes, was washed twice with Tris-HCl/ EDTA buffer and once with Tris-HCl buffer containing 5 mM MgCl2 (Tris-HCl/MgCl2 buffer).
Lysozyme-EDTA-treated envelopes were suspended in 10 mM Tris-HCl (pH: 7.5) containing 10 mM EDTA and 2% Triton X-100 (T8787, Sigma Chemical Co.; TX-buffer) and incubated for 2 h at 37°C with trypsin (93615, Sigma Chemical Co.; 0.5 mg/mL). Then, a further 0.5 mg/mL of trypsin was added, and the mixture was incubated again for 2 h at 37°C. The resulting digested mixture was centrifuged at 10,000 × g for 10 min at 4°C, the supernatant was collected, and the pH was adjusted to 6.0 with acetic acid 10%. Two milliliters of 5 M NaCl solution was added to the supernatant, and the sample was leaved overnight at 42°C to precipitate the porin fraction. The precipitate was collected by centrifugation at 10,000 × g for 10 min at 4°C, dissolved in 10 mM Tris-HCl (pH: 7.5). The suspension was centrifuged at 100,000 × g for 60 min at 4°C. The precipitate was recovered and washed twice with the same buffer, then dissolved in 10 mM Tris-HCl (pH: 7.5) containing 0.03% thimerosal (T5125, Sigma Chemical Co.) as a preservative, and stored at −20°C.
The protein content of the OMP extracts was determined by BCA assay system (BCA1-1KT, Sigma Chemical Co.) according to the protocol of standard analysis of the manufacturer. Each sample was tested in duplicate. The purity of each OMP extract was analyzed by SDS-PAGE by the method of Laemmli (1972) in 5 to 15% gradient acrylamide gels.
The lipopolysaccharide content of the OMP extracts was assayed by the Limulus amebocyte lysate test (Kinetic-QCL Kinetic Chromogenic Assay, US50-650U, Cambrex, Verviers, Belgium).
Animals
Twenty-eight ISA Brown hens (Gallus domesticus, specific- Salmonella spp.-free status, Elevage avicole Saint Georges, Villemareuil, France), 21 wk old at the beginning of the experiment, were kept individually in pens in a standard animal room with a 16L:8D cycle. The temperature in the room was 20 ± 4°C. The hens were fed 125 g per day of a standard laying hen diet (All-mash pondeuse, SCAR, Herve, Belgium). Water was given ad libitum.
Immunization of Hens
Twenty-four hens were randomly allocated to 1 of 6 experimental treatments (Table 1) corresponding to 6 different immune preparations for the induction of Salmonella-specific antibodies in egg yolk. The inoculum (500 μL) was administered to the hens at d 1 (21 wk old) intramuscularly at 4 sites on the pectoral muscle. In the case of the FA, the dose of antigen used (100 μg) was chosen on the basis of the instructions relating to chicken immunization protocols using this adjuvant (Schade and Hlinak, 1996; Schade et al., 2005). In the case of the IM, the choice of the dose (10 μg) was essentially made on the basis of the recommendations of the manufacturer (Isconova AB, Uppsala SE-75183, Sweden). The hens of the control group (n = 4), received in the same way, a sterile saline solution (PBS).
Table 1.
Composition of the different immune preparations1
| Antigen | ||||||
|---|---|---|---|---|---|---|
| SE-OMP | ST-OMP | (½ SE-OMP + ½ ST-OMP) | ||||
| 1SE = Salmonella Enteritidis; ST = Salmonella Typhimurium; OMP = outer membrane protein; FA = Freund’s adjuvant; IM = immunostimulating complexes matrix. | ||||||
| 2FA = Freund’s adjuvant; Freund’s complete adjuvant was used for the first immunization, and Freund’s incomplete adjuvant was used for the booster immunizations. | ||||||
| Treatment2 | SE-FA3 | SE-IM | ST-FA | ST-IM | SE-ST-FA | SE-ST-IM |
| Dose of antigen (μg) | 100 | 10 | 100 | 10 | 50+50 | 5+5 |
Booster inoculations given in the same way and same amount were carried out at d 22 (24 wk old), 43 (27 wk old) and 134 (40 wk old), except in the case of FA groups where FCA (F5881, Sigma Chemical Co.), used in the first immunization, was replaced by Freund’s incomplete adjuvant (F5506, Sigma Chemical Co.).
In each group of hens (n = 4), the eggs were collected daily, labeled accordingly, and the rate of laying per group was recorded weekly from the start to the end of the experiment.
Every 2 wk, from d 1 till 155, 1 egg per hen was taken, labeled accordingly, and stored in a refrigerator at 4°C until immunoglobulins were isolated.
General appearance of the hens and possible clinical signs were monitored daily. Once a week hens were weighted and injection sites were palpated to detect signs of pain. The experimental protocol was approved by the Official Animal Care and Use Committee of the Gembloux Agricultural University (protocol N° 05/04 NPC) before the initiation of the research and followed the Belgian guidelines approved by the committee on care, handling, and sampling of the animals.
Isolation and Partial Purification of IgY
Eggs were individually broken, the egg white discarded, and the yolk rolled on filter paper (Whatman No. 1, 512-1002, VWR International Europe BVBA, Leuven 3001, Belgium) to remove residues of egg white. The yolks were then transferred into a funnel fitted with filter paper that was opened at the bottom of the funnel, the yolk membranes were ruptured, and yolks were collected in a measuring cylinder leaving the yolk membrane attached to the paper. The yolks were diluted (1:9, vol/vol) with acidified distilled (pH: 2.5 to 2.8) water and mixed. The pH of the yolk suspension was then adjusted to 5.0 with 0.1 N HCl. After the suspension was allowed to stand for a night at 4°C, the aggregate lipoproteins of the yolk were separated by centrifugation (10,000 × g for 30 min at 4°C). The supernatant (water-soluble fraction) was collected and subjected to salt precipitation. This precipitation was carried out at room temperature by adding solid ammonium sulphate 19% (wt/vol) to the water-soluble fraction under stirring. After incubation for 30 min at room temperature, the solution was centrifuged (10,000 × g for 30 min at 4°C). The pellet was dissolved at the original volume of the yolk in PBS containing 0.03% thimerosal (T5125, Sigma Chemical Co.) and stored at −20°C until analysis.
Specific Activity of IgY
Enzyme-linked immunosorbent assay was used to check titers of SE-specific IgY in eggs laid by the hens receiving treatments SE-FA, SE-IM, SEST-FA, and SEST-IM. Each well of the 96 well microtiter plates (Nunc International, Rochester, NY) was coated overnight at 4°C with 100 μL of 1 μg/mL of SE-OMP antigen in 0.05 M carbonate-bicarbonate buffer (pH: 9.5). The wells were washed with phosphate buffered saline containing 0.1% Tween 20 (P7949, Sigma Chemical Co.; PBS-T), and then blocked by incubation for 30 min at room temperature with 200 μL of 2% BSA (A7030, Sigma Chemical Co.) in PBS 0.01 M. After 3 washings with PBS-T, IgY samples diluted to 1:1,000 were added to the wells, and the plates were incubated for 60 min at room temperature. One hundred microliters of horseradish peroxidase conjugated goat antichicken IgG (E30-104, Bethyl Laboratories Inc., Montgomery, TX) at 10,000-fold dilution was added to the wells after other 3 washings. The plates were incubated at room temperature for 60 min and washed with PBS-T for 3 times. The peroxidase activity was revealed with tetramethylbenzidine (T-2885, Sigma Chemical Co.) in acetate buffer (pH: 5.5) containing 0.03% of H2O2. After incubation for 5 min at room temperature, the enzymatic reaction was stopped with 100 μL of 2 M H2SO4 and the optical density (OD) was read at 405 nm in ELISA reader (PR 5000, Lab systems Multiskan RC, Helsinki, Finland). The antibody titer was expressed as P/N values (Li et al., 2006), where P represents the net OD of IgY from immunized hens at 1:1,000 dilution and N, the OD of IgY from the control hens at the same dilution. The net OD of IgY from immunized hens was calculated as the OD values of IgY from immunized hens minus the OD of IgY from the control hens.
To determine ST-specific IgY titers in eggs laid by the hens receiving treatments ST-FA, ST-IM, SEST-FA, and SEST-IM, ELISA was performed as described above, except the microtiter plate was coated with 100 μL of 1 μg/ mL of ST-OMP antigen.
Cross-Reactivity of IgY
The cross-reactivity of IgY was determined by using ELISA method as described by Lee et al. (2002) and Sunwoo et al. (2002). The cross-reactivity of anti-SE IgY, induced with treatments SE-FA and SE-IM against ST-OMP antigen, was determined using ELISA. Wells of the microtiter plate were coated with 100 μL of 1 μg/mL of SE-OMP or ST-OMP antigens in 0.05 M carbonate-bicarbonate buffer (pH: 9.5). Anti-SE IgY samples at 1:1,000 dilution were added to react with coated antigens. The cross-reactivity of anti-SE IgY against ST-OMP antigen was determined by comparing anti-SE IgY activity against ST-OMP antigen with activity against SE-OMP antigen, which was considered as 100%.
The cross-reactivity of anti-ST IgY, in eggs laid by the hens receiving treatments ST-FA and ST-IM, against SE-OMP antigen, was assayed according to the same procedure. The cross-reactivity of anti-ST IgY against SE-OMP antigen was determined by comparing anti-ST IgY activity against SE-OMP antigen with activity against ST-OMP antigen, which was considered as 100%.
Statistical Analysis
The laying rate and the BW of the hens were analyzed according to a GLM of the GLM procedure of SAS 8.02 software (SAS Institute, 1999). Model sums of squares were separated into treatment and time. Differences between levels of a given effect were tested using an F-test and were declared significant at P < 0.05. Differences between means were compared using Dunnett’s test (Dagnelie, 1986).
Given the repeated measurement data structure, the statistical analysis of anti-SE IgY titers values defined as (P/N)SE and measured every 2 wk during the experiment period (from d 1 till 155) for treatments SE-FA, SE-IM, SEST-FA, and SEST-IM were conducted using a mixed linear model of the MIXED procedure of SAS 8.02 software (SAS Institute, 1999). The model fitted antigen, adjuvant, and time as fixed classified effects and all their interactions and hen as random effect. Different covariance structures were tested to model repeated records for every hen. The smallest value for the Akaike’s information criteria was used to select the most appropriate covariance structure. Differences between levels of a given fixed effect were tested using an F-test and were declared significant at P < 0.05. The same statistical analysis was also conducted on anti-ST IgY titers values defined as (P/N)ST, for treatments ST-FA, ST-IM, SEST-FA, and SEST-IM. The 2 statistical analyses were performed separately because they concerned 2 different types of antibody.
RESULTS AND DISCUSSION
Antigen Preparation
The purity of the 2 OMP antigens was determined by SDS-PAGE analysis. The lanes resolved the purified proteins into 2 (SE-OMP extract) or 3 (ST-OMP extract) intensely stained bands located between 34 and 36 kDa, which correspond to the monomeric molecular size of the major outer membrane proteins of Salmonella as reported by Nurminen (1985; Figure 1). The Limulus amebocyte lysate test showed the presence of lipopolysac-charide in SE-OMP and ST- OMP extracts at 29 and 24 pg/10 μg of OMP, respectively.
Figure 1.

The SDS-PAGE profile of Salmonella Enteritidis (SE) and Salmonella Typhimurium (ST)-outer membrane proteins (OMP) extracts. M: Molecular size standards. Lane 1: SE-OMP extract (7.5 μg of protein). Lane 2: ST-OMP extract (7.5 μg of protein). Lane 3: SE-OMP extract (15 μg of protein). Lane 4: ST-OMP extract (15 μg of protein).
Animal Welfare and Egg Laying Rate
All immune preparations used in this study were shown to be safe and well-tolerated. None of the hens used in this study developed visible granulomas at the immunization sites. Only FA groups developed a tiny localized tissue reaction (inflammation) at the injection sites, which disappeared after a week. This reaction was observed only when FCA was used. This is in correspondence with results of Svendsen et al. (1996) who did not observe granulomas in chicken after injection of 0.4 mL of FCA/antigen emulsion (1:1).
Furthermore, no animal showed any clinical signs of distress in response to the immunization procedure or to any of the immune preparations. As shown in Table 2, no significant difference was observed between means of laying rate of the hens receiving the 6 immune preparations as compared with that of the control hens, especially for FA groups. These results indicated that the use of the FA did not alter the laying capacity of the hens, even after the first immunization, which was conducted with FCA. Our results contradict the findings of Bollen and Hau (1996), who observed a significant decrease in egg-laying rate when hens were immunized with FCA.
Table 2.
Laying rate and BW of hens according to the treatments1
| Main effect | Laying rate (%) | BW (g) |
|---|---|---|
| Treatment | ||
| SE-FA | 94.9a | 1,946a |
| SE-IM | 95.9a | 1,940a |
| ST-FA | 95.1a | 2,041a |
| ST-IM | 95.8a | 2,073a |
| SEST-FA | 94.8a | 2,037a |
| SEST-IM | 94.3a | 1,979a |
| Control treatment | 95.7a | 1,999a |
| Source of variations | df | SEM | P | SEM | P |
|---|---|---|---|---|---|
| aLeast square means within each column without a common letter with the control treatment are significantly different (P < 0.05). | |||||
| 1Values represent the least square means of the entire observation period. Treatments: SE-OMP (treatment SE-FA or SE-IM) or ST-OMP (treatment ST-FA or ST-IM), or a combined antigen preparation: ½ SE-OMP and ½ ST-OMP (treatment SEST-FA or SEST-IM). SE = Salmonella Enteritidis; ST = Salmonella Typhimurium; OMP = outer membrane protein. FA = Freund’s adjuvant: Freund complete adjuvant for the first immunization and Freund incomplete adjuvant for the booster immunizations). IM = immunostimulating complexes matrix. | |||||
| Treatment | 6 | 0.79 | 0.0874 | 49 | 0.0923 |
| Time | 21 | 7.85 | <0.0001 | 269 | <0.0001 |
For body weights data, similar observation was made (Table 2). The immunization procedure and the 6 immune preparations did not affect this parameter.
Kinetic of Specific Activity of IgY
When the hens were immunized according to our procedure, SE or ST-specific IgY was detected only by the second week (Figure 2). The lag time of 2 wk can be explained by the time it takes for specific antibodies produced in hen serum to be transferred and accumulated in yolk as reported in other studies utilizing the laying hen for antibody production (Sunwoo et al., 1996; Kim and Patterson, 2003; Fu et al., 2006).
Figure 2.
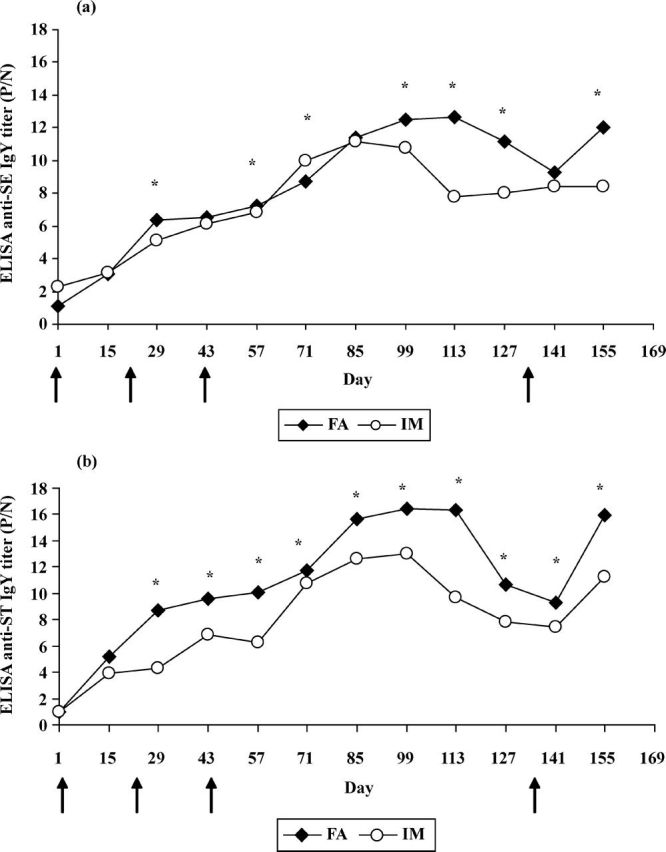
Kinetic of (a) anti-Salmonella Enteritidis (SE) IgY and (b) anti-Salmonella Typhimurium (ST) IgY in egg yolk laid by hens immunized with Freund’s adjuvants (FA) or immunostimulating complexes matrix (IM). Values are the mean of quadruple samples. *Within each day, IgY titer means differ significantly (P < 0.05). Arrows indicate the days of immunization. received treatment SE-IM at d 43 and 155 showed a cross-reactivity with ST-OMP antigen of 71.7 and 72.7%, respectively (Figure 3a).
From d 15 until 43, anti-Salmonella antibodies response climbed rapidly in the FA groups. In contrast, it rose more slowly in IM groups in spite the administration of a second injection at day 22. This may indicate that IM is not so potent than FA to produce rapidly high amounts of anti-Salmonella antibodies in the yolk.
After the third immunization (d 43), until d 57, the anti-Salmonella antibodies trended to increase more slowly than before. The effect of the third immunization is mainly marked after d 57.
Subsequently the antibody levels continued to increase and reached a peak approximately in d 99 or 113 in FA groups, and in d 85 or 99 in IM groups. After reaching peak, anti-Salmonella IgY titers decreased significantly (P < 0.05) in both adjuvant groups. However, this antibody decrease occurred more tardily in FA groups than in IM groups (Figure 2).
Thereafter, when a fourth immunization was conducted at the d 134, high levels of antibodies were recovered at d 155, especially for hens received treatments ST-FA, ST-IM, SEST-FA, and SEST-IM.
These findings indicated that 3 successive sensitizations at 3-wk intervals allow the induction of increasing antibodies titers practically lasting 4 mo. Subsequently, antibody titers fall. But when an additional booster was carried out, antibody titers start to increase again. Based on these findings, it would be undoubtedly preferable to carry out the fourth immunization when the maximum antibody level is reached to avoid the antibody titer decrease.
Antigen Effect
Independently of the adjuvant used, the mean of anti-SE antibody titer obtained from hens immunized with the combined vaccine preparation (P/NSE = 7.44 ± 0.15) was statistically similar (P > 0.05) as compared with that obtained from hens immunized with SE-OMP antigen given alone in a full-dose regimen (P/NSE = 7.50 ± 0.14; Table 3). The mean of anti-ST antibody titer raised against the combined vaccine preparation (P/NST = 9.16 ± 0.11) was statistically lower (P < 0.05) than that raised against ST-OMP antigen given alone in a full-dose regimen (P/ NST = 9.66 ± 0.11; Table 3). However, the relative difference between values is very slight and amounts to about 5%.
Table 3.
Antigen and adjuvant effects on the anti-Salmonella Enteritidis (SE) IgY and anti-Salmonella Typhimurium (ST) IgY titers1
| Main effect | Anti-SE IgY titer (P/N)SE | Anti-ST IgY titer (P/N)ST | |
|---|---|---|---|
| a,bLeast square means within each column for antigen or adjuvant without a common letter are significantly different (P < 0.05). | |||
| 1Values represent the least square means of the entire observation period. (P/N)SE = statistical analysis of anti-SE IgY titers values; (P/N)ST = statistical analysis of anti-ST IgY titers values. | |||
| 2SE-OMP = Salmonella Enteritidis outer membrane proteins extract. | |||
| 3ST-OMP = Salmonella Typhimurium outer membrane proteins extract. | |||
| 4FA = Freund’s adjuvant: Freund complete adjuvant for the first immunization and Freund incomplete adjuvant for the booster immunizations. | |||
| 5IM = immunostimulating complexes matrix. | |||
| Antigen | |||
| SE-OMP2 | 7.50a | — | |
| ST-OMP3 | — | 9.66a | |
| ½SE-OMP + ½ ST-OMP | 7.44a | 9.16b | |
| Adjuvant | |||
| FA4 | 8.22a | 10.91a | |
| IM5 | 6.73b | 7.91b | |
| Source of variations | df | Probability | |
| Antigen | 1 | 0.8843 | 0.0400 |
| Adjuvant | 1 | 0.0031 | <0.0001 |
| Antigen × Adjuvant | 1 | 0.5334 | 0.6213 |
| Sampling time | 11 | <0.0001 | <0.0001 |
| Antigen × time | 11 | 0.8045 | 0.1475 |
| Adjuvant × time | 11 | 0.0002 | <0.0001 |
| Antigen × adjuvant × time | 11 | 0.9648 | 0.5464 |
| Variance parameter estimate | |||
| Hen | 0.514 | 0.383 | |
| Residual | 2.803 | 1.035 | |
These data demonstrated that one-half the dose of each one of Salmonella spp.-OMP antigens might work equally or slightly lower as the full dose of this antigen when it is combined with a half-dose of its homologous. If the hens are to be used to produce antibodies in a large scale, the combined immune preparation has the advantage of reducing the number of hens to be immunized (only the half of animals will be needed) and the amounts of antigens to be used (a half-dose of each antigen vs. a full-dose), but especially, it offers the possibility to generate high levels of the 2 types of antibodies in the same egg. Following treatments SEST-FA and SEST-IM, the hens apparently produced high amounts of anti-SE and anti-ST antibodies in the same yolks because the antibodies could cross-react simultaneously with SE-OMP and STOMP antigens. To clarify this possibility, a cross-reactivity assay was conducted using ELISA.
The anti-SE IgY sampled in hens received SE-FA at d 43 and 155 cross-reacted 71.5 and 71.8% with ST-OMP antigen, respectively. Anti-SE IgY recovered from hens Anti-ST IgY recovered at d 43 and 155 from hens received treatment ST-FA showed a cross-reactivity with SE-OMP antigen of 76.9 and 79.5%, respectively. Anti-ST IgY sampled at d 43 and 155 in hens received treatment ST-IM cross-reacted 77.8 and 79.8% with SE-OMP antigen, respectively (Figure 3b).
Figure 3.
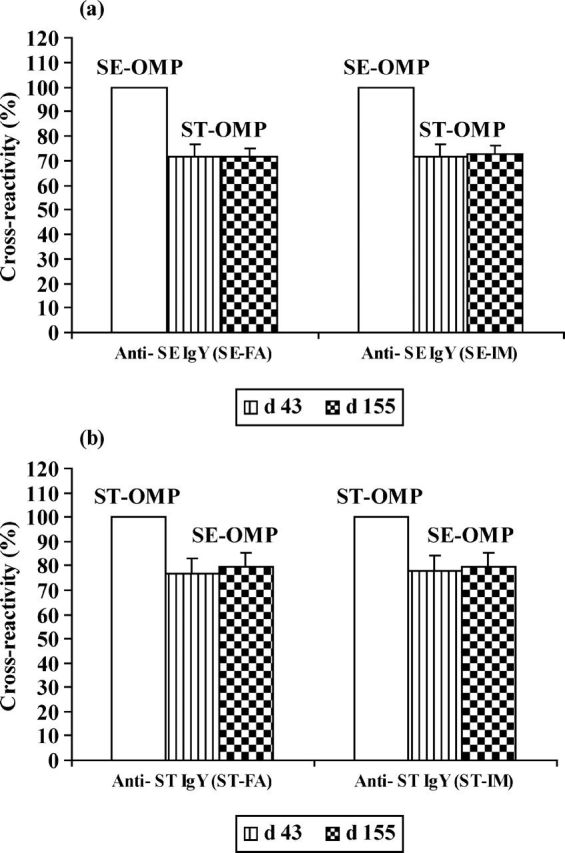
The cross-reactivity of (a) anti-Salmonella Enteritidis (SE) IgY with Salmonella Typhimurium (ST)-outer membrane proteins (OMP) antigen; (b) anti-ST IgY with SE-OMP antigen on d 43 and 155. Values are the mean of quadruple samples. Vertical bars indicate the standard deviation.
Regardless of the antibody type (anti-SE IgY or anti-ST IgY), there was no difference between cross-reactivity values measured for d 43 and those for d 155. This suggests that the cross-reactivity of an antibody with a de-fined antigen does not change over the time of sampling. Furthermore, for particular type of antibodies, results showed that the cross-reactivity values of antibodies induced with FA were similar to those of antibodies obtained with IM. This may indicate that the cross-reactivity of an antibody is independent of the type of the adjuvant used.
The high cross-reactivity of anti-Salmonella IgY observed between SE and ST can be explained by the fact that both Salmonella spp. share common epitopes on OMP (Kudrna et al., 1985). If the cross-reactivity has to be avoided, immunizing the hens with isolated proteins that are not shared by the 2 Salmonella serovars (Figure 1) could probably reduce it.
In this study, the observed values of cross-reactivity of anti-Salmonella IgY are higher than those shown in Lee et al. (2002), who have used SE and ST inactivated bacterial whole cell as antigens. It is thus likely that the degree of cross-reactivity of IgY directed against a purified antigen like OMP is greater than that of IgY raised against a crude antigen as bacterial whole cell.
Our results demonstrate that amount of anti-SE antibodies produced by the hens receiving treatment SEST-FA or SEST-IM were probably lesser than that produced by the hens receiving treatment SE-FA or SE-IM, respectively. The same conclusion may be drawn for anti-ST antibodies according to treatments SEST-FA or SEST-IM as compared with treatments ST-FA or ST-IM, respectively.
Independently of the adjuvant used, the overall average of anti-SE antibody titer developped in hens immunized according to treatments SE-FA and SE-IM (P/NSE = 7.50 ± 0.11) was lower than that of anti-ST antibody titer produced by the hens receiving treatments ST-FA and SE-IM (P/NST = 9.66 ± 0.11; Table 3). This finding suggests that SE-OMP antigen is weakly immunogenic as compared with ST-OMP antigen.
Adjuvant Effect
The choice of adjuvant had an effect on the antibody titer in the eggs. During the experiment period, FA resulted in titers significantly higher (P < 0.05) than IM. The same findings are observed for other animal species by Levi et al. (1993), Lockyer et al. (1993), and Sjölander et al. (1993). Furthermore, primary and secondary antibodies response magnitude induced by IM was significantly lower than that elicited by FA (P < 0.05). This may reflect the difference in the dose of antigen used for immunization with FA (100 μg) or IM (10 μg). But it has been reported that the necessary antigen dose required to mount a substantial antibody response can be considerably reduced (by 10- to 20-fold) when using immunostimulating complexes or immunostimulating complexes matrix (Coulter et al., 2003; Pearse and Drane, 2005). Because the side effects related to the use of the FA are small and of short duration, so for large-scale production of specific egg yolk antibody, it would be preferable to use FA because it induced higher antibody titers and costs much less than IM.
The adjuvant × time of sampling interaction was significant for anti-SE IgY and anti-ST IgY titers, which indicates that the effect of the adjuvant was closely related to the time of sampling (Figure 2).
In conclusions, our results demonstrate that antibodies simultaneously directed against SE and ST can be efficiently produced in the same egg yolk of hens immunized with SE-OMP and ST-OMP in a half-dose-mixture. This antibody mixture can be used as an additive in broiler chicken diets to fight both SE and ST, which are the predominant cause of salmonellosis in human often associated with poultry meat consumption. Nevertheless anti-SE IgY obtained with the combined antigen preparation showed lower activity than anti-ST IgY, which could be explained by the fact that SE-OMP antigen could be less immunogenic than ST-OMP. Furthermore, according to this immunization regimen, the amount of IgY properly specific to an antigen (SE-OMP or ST-OMP) is certainly lower than that produced with a full-dose of a single antigen because antibodies cross-react.
Moreover the results indicate that the antibodies response induced by IM is lower than that induced by FA. Our findings also show that FA did not cause serious side-effects in birds, especially when it is used in incomplete form at boosting immunizations.
As a final comment, our findings suggest that it is certainly desirable to carry out an additional immunization within the 3 mo following the first 3 immunizations to maintain a high level of antibodies in eggs.
Acknowledgments
This study was supported by the Gembloux Agricultural University (FUSAGx, Gembloux, Belgium), and the Funds for the Research in Industry and Agronomy (FRIA, Brussels, Belgium). The authors gratefully acknowledge Nicolas Gengler for his assistance in statistical analysis of the data and the Walloon Center of Industrial Biology (WCIB, Gembloux, Belgium) for its collaboration.
REFERENCES
- Bollen L. S., and J. Hau. 1996. Freund’s complete adjuvant has a negative impact on egg laying frequency in immunised chickens. In Vivo 13:107–108. [PubMed] [Google Scholar]
- Coulter A., R. Harris, R. Davis, D. Darne, J. Cox, D. Ryan, P. Sutton, S. Rockman, and M. Pearse. 2003. Intranasal vaccination with ISCOMATRIX adjuvanted influenza vaccine. Vaccine 21:946–949. [DOI] [PubMed] [Google Scholar]
- Dagnelie P. 1986. Théories et méthodes statistiques. Vol. 2. Presses Agron., Gembloux, Belgium.
- Fu C., H. Huang, X. Wang, Y. Liu, Z. Wang, S. Cui, H. Gao, Z. Li, J. Li, and X. Kong. 2006. Preparation and evaluation of anti-SARS coronavirus IgY from yolks of immunized SPF chickens. J. Virol. 133:112–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapoor P., M. M. Compton, and B. Howarth. 2000. Immunization of chickens with quail and turkey perivitelline membrane proteins: Production of antibodies and their effects on fertility. Poult. Sci. 79:245–256. [DOI] [PubMed] [Google Scholar]
- Kim W. K., and P. H. Patterson. 2003. Production of an egg yolk antibody specific to microbial uricase and its inhibitory effects on uricase activity. Poult. Sci. 82:1554–1558. [DOI] [PubMed] [Google Scholar]
- Klemperer F. 1893. Ueber natürlich Immunität und ihre verwerthung für die Immunisirungstherapie. Arch. Exp. Pathol. Pharmacol. 31:356–382. [Google Scholar]
- Kuby J. 2007. Antigens. Pages 87–186 in Immunology. 6th ed. J. Kuby, ed. W. H. Freeman and Co., New York, NY.
- Kudrna D. A., G. W. Teresa, J. M. Arnzen, and K. S. Beard. 1985. Immunoglobulin M and Immunoglobulin G responses in BALB/c mice to conjugated outer membrane extracts of four Salmonella serotypes. Infect. Immun. 49:598–608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laemmli U. K. 1972. Cleavage of structural proteins during the assembly of the head of bacteriolophage T4. Nature 227:680–685. [DOI] [PubMed] [Google Scholar]
- Lee E. N., H. H. Sunwoo, K. Menninen, and J. S. Sim. 2002. In vitro studies of chicken egg yolk antibody (IgY) against Salmonella enteritidis and Salmonella typhimurium.Poult. Sci. 81:632–641. [DOI] [PubMed] [Google Scholar]
- Leenaars P., C. F. Hendriksen, A. Angulo, M. Koedam, and E. Claassen. 1994. Evaluation of several adjuvants as alternatives to the use of Freund’s adjuvant in rabbits. Vet. Immunol. Immunopathol. 40:225–241. [DOI] [PubMed] [Google Scholar]
- Leenaars P., M. Koedam, P. Wester, V. Baumans, E. Claassen, and C. F. Hendriksen. 1998. Assessment of side effects induced by injection of different adjuvant/antigen combinations in rabbits and mice. Lab. Anim. 32:387–406. [DOI] [PubMed] [Google Scholar]
- Levi M., U. Ruden, D. Birx, L. Loomis, R. Redfield, K. Lovgren, L. Akerblom, E. Sandstrom, and B. Wahren. 1993. Effects of adjuvants and multiple antigen peptides on humoral and cellular immune responses to gp160 of HIV-1. J. Acquir. Immune Defic. Syndr. 6:855–864. [PubMed] [Google Scholar]
- Li X., J. Shuai, and W. Fang. 2006. Protection of Carassius auratus Gibelio against infection by Aeromonas hydrophila using specific immunoglobulins from hen egg yolk. J. Zheijiang Univ. Sci. B 7:922–928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lockyer M. J., H. Cooper, J. Tite, W. Rowan, and J. S. Crowe. 1993. Immunogenicity of a hybrid Plasmodium falciparum malaria antigen. Parasitology 106:451–457. [DOI] [PubMed] [Google Scholar]
- Nurminen M. 1985. Isolation of porin trimers. Pages 293–300 in Enterobacterial surface antigens: Methods for molecular characterisation. T. K. Korhonen, E. A. Dawes, and P. H. Mäkelä, ed. Elsevier Sci. Publ., Amsterdam, the Netherlands.
- Pearse M. J., and D. Drane. 2005. ISCOMATRIX adjuvant for antigen delivery. Adv. Drug Deliv. Rev. 57:465–474. [DOI] [PubMed] [Google Scholar]
- SAS Institute. 1999. SAS User’s Guide: Statistics, Version 8.02. SAS Inst. Inc., Cary. NC.
- Schade R., E. G. Calzado, R. Sarmiento, P. A. Chacana, J. Porankiewicz-Asplund, and H. R. Terzolo. 2005. Chicken egg yolk antibodies (IgY-technology): A review of progress in production and use in research and human and veterinary medicine. Altern. Lab. Anim. 33:129–154. [DOI] [PubMed] [Google Scholar]
- Schade R., and A. Hlinak. 1996. Egg Yolk Antibodies, State of the Art and Future Prospects. ALTEX 13:5–9. [PubMed] [Google Scholar]
- Sjölander A., S. Stahl, K. Lövgren, M. Hansson, L. Cavelier, A. Walles, H. Helmby, B. Wahlin, B. Morein, M. Uhlen, K. Berzins, P. Perlmann, and M. Wahlgren. 1993. Plasmodium falciparum: The immune response in rabbits to the clustered asparagine-rich protein (CARP) after immunization in Freund’s adjuvant or immunostimulating complexes (IS-COMs). Exp. Parasitol. 76:134–145. [DOI] [PubMed] [Google Scholar]
- Sunwoo H. H., E. N. Lee, K. Menninen, M. R. Suresh, and J. S. Sim. 2002. Growth inhibitory effect of chicken egg yolk antibody (IgY) on Escherichia coli O157:H7. J. Food Sci. 67:1486–1494. [Google Scholar]
- Sunwoo H. H., T. Nakano, W. T. Dixon, and J. S. Sim. 1996. Immune responses in chickens against lipopolysaccharide of Escherichia coli and Salmonella typhimurium.Poult. Sci. 75:342–345. [DOI] [PubMed] [Google Scholar]
- Svendsen L., A. Crowley, G. Stodulski, and J. Hau. 1996. Antibody production in rabbits and chickens immunized with human IgG. A comparison of titre and avidity development in rabbit serum, chicken serum and egg yolk using three different adjuvants. J. Immunol. Methods 191:113–120. [DOI] [PubMed] [Google Scholar]