

Abstract
The spectrum of infectious diseases has shifted in the past 50 years to include those caused by microbes that cause disease predominantly in immunocompromised individuals. This phenomenon has underscored the dependence of microbial virulence on the immune status of the host. The limited efficacy of the available antimicrobial armamentarium in immunocompromised individuals, combined with increasing resistance to these agents, has led to an urgent need for new therapies for infectious diseases. Immunomodulation represents a novel approach to antimicrobial therapy that depends on bolstering host immunity, rather than direct antimicrobial activity. Immunomodulators can be divided into those that are specific to pathogens (pathogen-specific) and those that are not specific to pathogens (non-specific). However, to date only a few immunomodulators have been evaluated for their efficacy as antimicrobial tools.
Introduction: immunomodulation in the context of the Damage-response framework
Immunomodulators are usually products of the immune system [1••]. As such, it is useful to consider immunomodulation approaches to infectious diseases in the context of microbial pathogenesis. In contrast to microbe-centric views, in which microbial pathogenesis and virulence are considered to reflect singular microbial functions, the Damage-response framework provides a flexible construct that accounts for the contribution of the host, as well as the microbe, to these entities [2]. The Damage-response framework considers host damage to be the common denominator in microbial pathogenesis. Based on this tenet, host damage can be plotted against the host immune response as a U-shaped curve, whereby the maximal host damage resulting from a given host–microbe interaction occurs both when the immune response is weak and when it is overly strong (Figure 1, Figure 2 ). The inherent flexibility afforded by this curve lies in its ability to account for the fact that certain microbes only cause disease in certain hosts, a phenomenon that cannot be explained by views of microbial pathogenesis that consider virulence to be a singular microbial trait [2].
Figure 1.
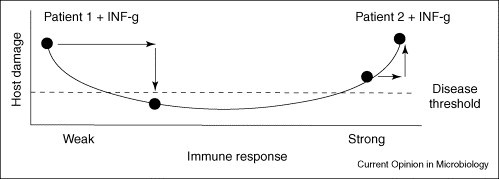
The possible effects of IFN–γ therapy in two patients with cryptococcosis in the context of the Damage-response framework. Patient 1 is an individual with AIDS-related cryptococcosis, where susceptibility to infection is associated with a profound defect in Th1-type immunity as a result of CD4 T-cell deficiency. In this patient, the administration of IFN–γ is pro-inflammatory and the increased inflammatory response might facilitate control of the infection, thus reducing damage and symptoms of disease. By contrast, Patient 2 is an individual with cryptococcal disease following immune reconstitution with HAART. In this patient, administration of IFN–γ might be detrimental, as cryptococcal disease is caused by an exuberant inflammatory response. Hence, the outcome of IFN–γ therapy depends on the immune status of the host.
Figure 2.
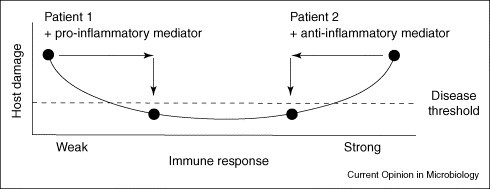
Illustration of the dichotomous requirements for immunomodulation in patients with different immune status. Patient 1 has an infectious disease that reflects the outcome of a weak immune response, such as HIV-associated histoplasmosis or aspergillosis following stem cell transplantation. In this individual enhancement of the inflammatory response with a pro-inflammatory immunomodulator could facilitate microbial clearance, thus reducing damage and symptoms of disease. By contrast, Patient 2 has an infectious disease that reflects the outcome of an overly exuberant immune response, such as mediastinal fibrosis from histoplasmosis or allergic aspergillosis. In this individual, an anti-inflammatory immunomodulator could dampen the host the inflammatory response, thus reducing damage and symptoms of disease. Notably, the immune response of this individual could have already resulted in microbial clearance. These patients illustrate that the kind of immunomodulator that would be beneficial is probably to be influenced by the immune status of the affected individual.
A logical corollary of the Damage-response framework is that infectious diseases only occur in susceptible hosts. Host immune mechanisms protect against infectious diseases by preventing or reducing the damage that can result from host–microbe interaction. The relationship between host immunity and microbial pathogenesis is clearly exemplified in immunocompromised hosts, by diseases that are caused by commensal microbes, such as Candida albicans and Staphylococcus epidermidis, and fungi, such as Cryptococcus neoformans and Pneumocystis jurevecci, and the success of immune reconstitution in preventing HIV-associated diseases caused by these microbes. The phenomenon of immune reconstitution disease that follows antiretroviral therapy with highly active anti-retroviral therapy, or HAART, in patients with AIDS (acquired immunodeficiency syndrome) illustrates how rebounding immunity can produce disease (discussed in [3]). Because the success of antimicrobial therapy is a function of its ability to ameliorate disease, and disease is a manifestation of host damage, the Damage-response framework provides a useful construct to consider approaches to treating infectious diseases that reduce host damage resulting from the host–microbe interaction.
The crisis in antimicrobial therapy, which has stemmed from antibiotic overuse, misuse and the limited number of new antimicrobial drugs on the near horizon is well documented [4••]. However, another area that limits the utility of antimicrobial drug-based therapy is that antimicrobial agents are frequently ineffective in individuals with impaired immunity, often despite being highly active in vitro or in individuals with intact immunity. This underscores the crucial relationship between host immunity and microbial virulence and provides a powerful rationale for approaches to antimicrobial therapy that regulate the immune response to reduce, ameliorate or prevent host damage.
Immunomodulators as antimicrobial tools
Approaches to immunomodulation can be divided into those that are specific to pathogens (pathogen-specific) and those that are not (non-specific). Pathogen-specific immunomodulators include antibody reagents and vaccines. With the exception of the rabies and varicella-zoster vaccines, currently licensed vaccines are administered to prevent acute infectious diseases rather than for therapy and are not discussed further here. Non-specific immunomodulators include cytokines, antimicrobial peptides, certain antimicrobial drugs and microbes such as probiotics. At present, clinical experience with non-specific immunomodulators as antimicrobial tools has been predominantly limited to cytokines.
Pathogen-specific immunomodulators: antibody-based agents
There are powerful historical precedents for the use of antibody-based therapies to treat infectious diseases (reviewed in [5]). The first era of antimicrobial therapy, early in the 20th century, was based on serum therapy with antibody preparations. Hence, the inaugural antimicrobial agents were immunomodulators [6]. First generation antibody reagents were abandoned because of their toxicity, which was a result of their impurity and derivation from non-human species, and the arrival of antimicrobial drugs that acted directly on the microbe. Nonetheless, there was evidence for synergism between antibiotics and serum therapies [7]. Serum therapy was validated in animal models before being administered to humans [7]; however, the mechanism by which it ameliorated infectious diseases or enhanced the efficacy of antimicrobial drugs was largely unknown. For most of the 20th century, the mechanisms of antibody action that were thought to influence antibody efficacy included their ability to neutralize, promote opsonization and phagocytosis or antibody-dependent cell-mediated cytotoxicity (ADCC) and/or to activate complement [8]. However, a significantly more robust menu of potential mechanisms of antibody action, which includes direct antimicrobial action, immunomodulation and generation of oxidative species, has emerged over the past decade [8]. For example, the efficacy of a pneumococcal capsular polysaccharide-specific antibody was associated with modulation of the cellular response to pneumococcus in the lungs of mice with pulmonary infection [9•]. This finding suggested that the well-documented ability of serum therapy to ameliorate fever and other clinical symptoms of pneumococcal pneumonia [10] could have reflected a downregulation of the host inflammatory response (or damage control). Perhaps the observation that pneumococcal pneumonia was only responsive to serum therapy with the homologous capsular polysaccharides-specific antiserum in the first three days of symptoms indicated that its capacity to mediate immunomodulation was limited to the early stages of disease. Antibodies to other microbes, including C. neoformans and Histoplasma capsulatum are able to modulate the cellular immune response to pulmonary infection (see [11•]). Studies of antibody action have shown that the older dichotomous view that antibody immunity was only effective against extracellular pathogens, whereas cellular immunity was responsible for immunity against intracellular pathogens has been deconstructed by evidence that antibody reagents can be effective against classical intracellular pathogens, such as Mycobacterium tuberculosis, C. neoformans, H. capsulatum and scores of viruses [11•].
Monoclonal antibody-based agents
Currently, there is only one antibody reagent licensed for use against an infectious disease in the United States — Palivizumab. Licensed in 1998, Palivizumab is a neutralizing, humanized monoclonal antibody (mAb) to protein F on respiratory syncytial virus (RSV) that reduced hospitalization for RSV in premature and other high-risk infants when given as prophylaxis [12]. Because the antiviral activity of Palivizumab was associated with a reduction in inflammatory mediator release in a murine model of RSV [13•], its mechanism of action probably involves immunomodulation. Despite ongoing controversies about the cost and target population of Palivizumab, its success in reducing the risk of RSV in high-risk infants promoted the development of second generation reagents and vaccine candidates [14].
Recently, Mycograb, a human recombinant antibody fragment was shown to significantly improve the response to amphotericin B in patients with invasive candidiasis [15••]. Patients who received Mycograb and amphotericin B showed a higher rate of complete overall response on day 10 of therapy, a significantly better mycological response and less Candida-attributable mortality than patients who received amphotericin B and a placebo. Mycograb was safe and well-tolerated; however, episodes of hypertension occurred more frequently in patients who received Mycograb than those who received a placebo. Mycograb is a recombinant antibody fragment lacking an Fc region, and is produced from a human anti-Hsp90 (heat-shock protein 90) cDNA library with an epitope that inhibits fungal Hsp90, NILKVIRKNIVKK [16]. The development of this antibody was driven by the observation that recovery from invasive candidiasis was associated with the appearance of antibodies to a 47 kDa determinant [17] that was found to be a fungal homolog of human Hsp90. Mycograb was tested in patients in comparison to standard therapy. As such, the question of whether or not its efficacy in vivo depends on synergy with antifungal drugs is unanswered. Nonetheless, the in vitro activity of Mycograb (with amphotericin B and other antifungal agents) against resistant Candida and other fungal species [18, 19] suggests it could hold promise as a broadly active antifungal agent.
The first mAb used to treat a fungal disease in humans was the mouse mAb 18B7, which binds to the cryptococcal capsular polysaccharide glucuronoxylomannan [20••, 21]. Extensive preclinical testing revealed that 18B7 augmented host defense mechanisms against C. neoformans, in vitro and in vivo (reviewed in [22]). In the clinical trial, administration of a single 1 mg kg−1 dose of 18B7 to HIV-infected patients treated for cryptococcal meningitis was well-tolerated and was associated with a reduction in serum glucuronoxylomannan levels [20••]. The tolerability and promising effect of this reagent in HIV-infected patients bolsters the prospect that immunotherapeutic interventions have the potential to augment host immune mechanisms in the treatment of infectious diseases in immunocompromised individuals.
Now, mAbs have been developed against a myriad of microbes responsible for emerging infectious diseases and/or those that cause disease in the setting of immune impairment. A human mAb to Bacillus anthracis toxin has recently successfully completed Phase I trials and stands as a potentially useful therapeutic in the event of an anthrax biological attack [23•]. Several human mAbs to the SARS (severe acute respiratory syndrome) coronavirus have been developed that might be useful if the disease reappears [24•]. Remarkably, these mAbs were developed to the point that clinical use was possible in less than five years. Human mAbs were highly effective against experimental shiga-toxin producing Escherichia coli in piglets [25]. Studies in experimental models have revealed that the efficacy of certain mAbs depends on intact cellular immunity (see [11•, 26]). As such, the use of mAbs in immunocompromised patients could depend on whether its efficacy requires the immune function (a subset or element of) that is lacking in the relevant patient(s). mAbs have the advantage of homogeneity and high specific activity. Although there is concern that mAbs could have limited usefulness for microbes that demonstrate high antigenic variation and mutability, combinations of mAbs have shown promise in overcoming this limitation [27••].
Polyclonal antibody-based agents
Another type of antibody-based therapy for infectious diseases consists of polyclonal immunoglobulin-based agents, including intravenous immunoglobulin (IVIG) and specific immune globulins (sIgs, sometimes called hyperimmune globulin). Treatment and prevention of rabies depends entirely on the combination of two immunomodulators: rabies vaccine and rabies immune globulin [28]. The sIgs are the mainstay of managing exposures to viral agents in susceptible individuals who are not candidates for live vaccines, such as pregnant women and patients with impaired immunity [29]. A polyclonal preparation derived from individuals with high serum antibody titers to staphylococcal fibrinogen-binding proteins, serine aspartate dipeptide repeat G and clumping factor A is under clinical development [30•]. An evaluation of this preparation in very low-weight infants revealed a trend toward fewer staphylococcal and candidal infections [31]. The ability of preformed antibodies to provide immediate defense against infectious diseases in susceptible individuals provides a potent justification for the use of antibodies, for example, in the setting of an act of bioterrorism or of epidemic diseases [32].
The use of IVIG in infectious diseases remains controversial. IVIG has also been shown to be useful for the treatment of Kawasaki Disease, cytomegalovirus pneumonitis in organ transplant recipients [33•] and parvovirus in patients with HIV infection [34]. IVIG is also useful in patients with toxic shock syndrome [35], West Nile virus infection [36] and sepsis [37], and is also invaluable in the management of patients with hypogammaglobulinemia who are at increased risk for infectious diseases, such as enteroviral meningitis [33•].
Non-specific immunomodulators: cytokines as antimicrobial tools
Cytokine-based therapies contrast with antibody reagents in that they are not pathogen-specific. The rationale for the use of cytokines as adjunctive immunomodulators for infectious diseases is based on the concept that replacement or augmentation of natural mediators of host defense should enhance the antimicrobial effect of host immune mechanisms and/or antimicrobial agents. Despite the logical basis for this concept, the potential antimicrobial power of these agents has been difficult to harness clinically. Currently, there are only a few examples of the use of adjunctive cytokines against infectious diseases. Notable exceptions are the use of recombinant α-interferons and nucleoside analogs for hepatitis B virus (HBV) and pegylated interferons and ribavirin for hepatitis C virus (HCV) [38]. The efficacy of interferons against HCV has been attributed to the induction of Th1 immunity [39]. A side effect of interferon-based therapies that limits their use in certain patients is depression [40•].
The effecacy of adjunctive interferon-gamma 1b (IFN–γ1b) with amphotericin B was studied in a Phase II, double-blind placebo-controlled trial for AIDS-associated cryptococcal meningitis [41••]. There was a trend towards mycological response and clinical improvement among interferon recipients, with 26% showing improvement, compared to 8% of placebo-controlled subjects. Although this difference did not reach statistical significance, the trend towards a beneficial effect of adjunctive interferon is encouraging, calling for further, larger scale studies and studies to identify those patients in whom adjunctive immunotherapy could be beneficial. The rationale for interferon therapy for cryptococcosis has a strong basis in preclinical studies in mice [42] and a human study showing an association between cerebrospinal fluid levels of IFN–γ and treatment in HIV-infected patients with cryptococcal meningitis [43•]. In light of the established benefit of interferon therapy for the prevention of bacterial diseases in patients with chronic granulomatous disease [44], adjunctive interferon could hold promise as an adjunctive agent for HIV-associated cryptococcal meningitis. However, the absence of surrogate markers that can predict the patients who would benefit from interferon therapy underscores the potential pitfalls in study-design and patient selection for clinical trials. This is particularly problematic for cryptococcal meningitis, a disease that can occur in the case of weak or reconstituted immunity in patients with HIV infection [3]. The failure to demonstrate the effect of a pro-inflammatory immunomodulator could reflect the induction of an excessive inflammatory response that promotes disease. Figure 1 provides a schematic interpretation of the use of IFN–γ therapy in the context of the Damage-response framework. Depending on the immunological status of the affected patient, adjunctive IFN–γ therapy could be beneficial or detrimental.
Preclinical data, demonstrating the importance of Th1 helper T-cell responses in protection against fungi, in experimental models has led to the proposal that adjunctive cytokines be used with antifungal agents for invasive fungal infections [45]. The rationale for the use of colony stimulating factors (CSFs), derived from granulocytes (G-CSF) or macrophages (GM-CSF), is in part based on their ability to alleviate neutropenia [45]. Data on the clinical use of adjunctive CSFs for fungal diseases is limited to small studies or case reports (see [45] for review). Nonetheless, administration of GM-CSF with antifungal agents in patients with invasive fungal infections was associated with decreased patient mortality or better response rates compared to a placebo [46] or antifungal therapy alone [47]. In view of the small number of affected patients and medical and ethical considerations in the design of randomized, double-blind placebo-controlled studies, it is possible that safe, well-tolerated candidate immunomodulators that are beneficial in preclinical studies will find their way into the antimicrobial armamentarium through compassionate use protocols and salvage therapy [48••].
Anti-inflammatory immunomodulators
The rationale for the use of adjunctive pro-inflammatory cytokines and certain antibodies for treating infectious diseases is to enhance the host response. However, a relatively underappreciated principle of microbial pathogenesis is that the damage resulting from host–microbe interaction can be the result of an overly exuberant host response [2]. Hence, there is a rationale for use of therapeutic interventions that dampen or reduce, as well as those that enhance or augment, the inflammatory response. It has been proposed that the beneficial effect of IVIG against inflammatory diseases involves engagement of the inhibitory Fc receptor, which downregulates the inflammatory response [49]. Probiotics, which are live bacteria derived from the human gastrointestinal tract, have been used as therapy for inflammatory bowel diseases, including antibiotic-associated diarrhea [50]. Probiotics remain outside the established antimicrobial armamentarium and are fraught with the potential for harm in immunocompromised hosts [51]. Nonetheless, it is logical to postulate that replacement and/or enhanced activity of the human microbiota could be beneficial for treatment of infectious diseases that reflect a failure of mucosal surfaces or of the innate mechanisms they bolster to protect against microbe-mediated damage (see [1••]).
Corticosteroids are important anti-inflammatory agents. Despite decades of controversy, corticosteroids have been validated as an important adjunct to antimicrobial therapy for bacterial meningitis [52•], HIV-associated Pneumocystis pneumonia [53] and tuberculosis meningitis [54•]. However, their mechanism of efficacy in these diseases might not be a direct anti-inflammatory effect [55]. Intriguingly, agents which have been proposed to have anti-inflammatory effects are macrolide antibiotics [56•]. The ability of macrolides to suppress the release of inflammatory mediators from phagocytes in vitro has been recognized for some time, but the clinical relevance of this phenomenon remains uncertain. Nonetheless, it has been hypothesized that the benefit of low-dose macrolide therapy in chronic pulmonary diseases, could be as a result of macrolide-induced reductions in levels of inflammatory cytokines, neutrophil recruitment and biofilm formation, which have been observed in animal models and/or in vitro [57]. In contrast to macrolides, amphotericin B [58] and penicillin [59] induce the release of inflammatory mediators in vitro through Toll-like receptor 2 (TLR2)-mediated stimulation. The inflammatory properties of these drugs raise the question of whether their immunomodulatory properties have an independent contribution to their therapeutic efficacy.
Conclusions: the future of adjunctive immunomodulators as antimicrobial tools
The future use of adjunctive immunomodulators for infectious diseases requires a better understanding of microbial pathogenesis and the relative need for immune activation versus immune modulation in the context of the immune response of the affected individual. In light of the fact that certain infectious diseases reflect an insufficient response, whereas others reflect an overly exuberant response, different types of interventions are likely to be required, depending on the immune status of the patient (Figure 2). The Damage-response framework can be a useful tool when considering the type of intervention that might be successful, but such predictions require experimental validation to be translated to the bedside and clinic.
References and recommended reading
Papers of particular interest, published within the annual period of review, have been highlighted as:
• of special interest
•• of outstanding interest
Acknowledgements
This work was supported by grants from the National Institutes of Health AI33580 (LP), AI44374 (LP), AI45459 (LP), AI033142 (AC), AI033774 (AC), and HL059842 (AC).
References
- 1••.Committee on New Directions in the Study of Antimicrobial Therapeutics: Immunomodulation: Treating infectious diseases in a microbial world: report of two workshops on novel antimicrobial therapies. Washington, DC: National Academies Press; 2006. [PubMed]; This 2006 National Academies report details the potential for modulating innate and acquired immunity and the microbiota to enhance the potential of immunomodulation for the treatment of infectious diseases.
- 2.Casadevall A., Pirofski L. The damage-response framework of microbial pathogenesis. Nat Rev Microbiol. 2003;1:17–24. doi: 10.1038/nrmicro732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Datta K., Pirofski L. Towards a vaccine for Cryptococcus neoformans: principles and caveats. FEM Yeast Res. 2006;6:536. doi: 10.1111/j.1567-1364.2006.00073.x. [DOI] [PubMed] [Google Scholar]
- 4••.Spellberg B., Powers J.H., Brass E.P., Miller L.G., Edwards J.E., Jr. Trends in antimicrobial drug development: implications for the future. Clin Infect Dis. 2004;38:1279–1286. doi: 10.1086/420937. [DOI] [PubMed] [Google Scholar]; This article details decline in new drug development, despite the need for new additions to the available antimicrobial armamentarium, and puts forth a powerful argument in support of discovery of novel antimicrobial agents.
- 5.Buchwald U.K., Pirofski L. Immune therapy for infectious diseases at the dawn of the 21(st) century: the past, present and future role of antibody therapy, therapeutic vaccination and biological response modifiers. Curr Pharm Des. 2003;9:945–968. doi: 10.2174/1381612033455189. [DOI] [PubMed] [Google Scholar]
- 6.Casadevall A. The third age of antimicrobial therapy. Clin Infect Dis. 2006;42:1414–1416. doi: 10.1086/503431. [DOI] [PubMed] [Google Scholar]
- 7.Casadevall A., Scharff M.D. Serum therapy revisited: animal models of infection and development of passive antibody therapy. Antimicrob Agents Chemother. 1994;38:1695–1702. doi: 10.1128/aac.38.8.1695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Casadevall A., Pirofski L.A. New concepts in antibody-mediated immunity. Infect Immun. 2004;72:6191–6196. doi: 10.1128/IAI.72.11.6191-6196.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9•.Burns T., Abadi M., Pirofski L. Modulation of the lung inflammatory response to serotype 8 pneumococcus infection by a human monoclonal IgM to serotype 8 capsular polysaccharide. Infect Immun. 2005;73:4530–4538. doi: 10.1128/IAI.73.8.4530-4538.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study demonstrates that the efficacy of a non-opsoinic human antibody to pneumococcal capsular polysaccharide is associated with modulation of the inflammatory response to pneumococcal infection in a mouse model of pneumococcal pneumonia.
- 10.Heffron R. Pneumonia. Harvard University Press; Cambridge: 1939. Serum treatment; pp. 805–880. [Google Scholar]
- 11•.Casadevall A., Pirofski L. A reappraisal of humoral immunity based on mechanisms of antibody-mediated protection against intracellular pathogens. Adv Immunol. 2006;91:1–44. doi: 10.1016/S0065-2776(06)91001-3. [DOI] [PubMed] [Google Scholar]; This article deconstructs the older view that antibody immunity is limited to extracellular organisms and details newer mechanisms of antibody action, including their ability to function as pro-inflammatory and anti-inflammatory immunomodulators as regulators of cellular immunity.
- 12.IMpact-RSV group Palivizumab, a humanized respiratory syncytial virus monoclonal antibody, reduces hospitalization from respiratory syncytial virus infection in high-risk infants. Pediatrics. 1998;102:531–537. [PubMed] [Google Scholar]
- 13•.Mejias A., Chavez-Bueno S., Rios A.M., Saavedra-Lozano J., Fonseca A.M., Hatfield J., Kapur P., Gomez A.M., Jafri H.S., Ramilo O. Anti-respiratory syncytial virus (RSV) neutralizing antibody decreases lung inflammation, airway obstruction, and airway hyperresponsiveness in a murine RSV model. Antimicrob Agents Chemother. 2004;48:1811–1822. doi: 10.1128/AAC.48.5.1811-1822.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study evaluated the effect of Palivizumab on virological and immunological parameters and lung inflammation in a mouse model of RSV pneumonia. The results showed that Palivizumab reduced the viral load in lung whether given before or up to 48 h after RSV infection, but that mAb administration before infection was associated with lower levels of inflammatory mediators and less lung damage. An important implication of this study is that the timing of mAb administration is a key determinant of its immunomodulatory activity.
- 14.Venkatesh M.P., Weisman L.E. Prevention and treatment of respiratory syncytial virus infection in infants: an update. Expert Rev Vaccines. 2006;5:261–268. doi: 10.1586/14760584.5.2.261. [DOI] [PubMed] [Google Scholar]
- 15••.Pachl J., Svoboda P., Jacobs F., Vandewoude K., van der H.B., Spronk P., Masterson G., Malbrain M., Aoun M., Garbino J. A randomized, blinded, multicenter trial of lipid-associated amphotericin B alone versus in combination with an antibody-based inhibitor of heat shock protein 90 in patients with invasive candidiasis. Clin Infect Dis. 2006;42:1404–1413. doi: 10.1086/503428. [DOI] [PubMed] [Google Scholar]; This study reports the result of a landmark trial that compared the outcome of patients with candidiasis treated with adjunctive Mycograb, a recombinant human antibody fragment, or placebo and amphotericin B. The result that Mycograb treatment was associated with a better mycological and clinical response suggests that Mycograb holds promise for treatment of candidiasis.
- 16.Matthews R., Burnie J. Antifungal antibodies: a new approach to the treatment of systemic candidiasis. Curr Opin Investig Drugs. 2001;2:472–476. [PubMed] [Google Scholar]
- 17.Matthews R.C., Burnie J.P., Tabaqchali S. Immunoblot analysis of the serological response in systemic candidosis. Lancet. 1984;2:1415–1418. doi: 10.1016/s0140-6736(84)91618-0. [DOI] [PubMed] [Google Scholar]
- 18.Matthews R.C., Rigg G., Hodgetts S., Carter T., Chapman C., Gregory C., Illidge C., Burnie J. Preclinical assessment of the efficacy of mycograb, a human recombinant antibody against fungal HSP90. Antimicrob Agents Chemother. 2003;47:2208–2216. doi: 10.1128/AAC.47.7.2208-2216.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nooney L., Matthews R.C., Burnie J.P. Evaluation of Mycograb(R), amphotericin B, caspofungin, and fluconazole in combination against Cryptococcus neoformans by checkerboard and time-kill methodologies. Diagn Microbiol Infect Dis. 2005;51:19–29. doi: 10.1016/j.diagmicrobio.2004.08.013. [DOI] [PubMed] [Google Scholar]
- 20••.Larsen R.A., Pappas P.G., Perfect J., Aberg J.A., Casadevall A., Cloud G.A., James R., FIller S., Dismukes W.E. Phase I evaluation of the safety and pharmacokinetics of murine-derived anticryptococcal antibody 18B7 in subjects with treated cryptococcal meningitis. Antimicrob Agents Chemother. 2005;49:952–958. doi: 10.1128/AAC.49.3.952-958.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study reports the results of a clinical trial of a mouse mAb to cryptococcal polysaccharide in patients with HIV-associated cryptococcal meningitis. The study is the first to evaluate antibody therapy for a fungal disease.
- 21.Casadevall A., Cleare W., Feldmesser M., Glatman-Freedman A., Kozel T.R., Lendvai N., Mukherjee J., Pirofski L., Rivera J., Rosas A.L. Characterization of a murine monoclonal antibody to C. neoformans polysaccharide which is a candidate for human therapeutic studies. Antimicrob Agents Chemother. 1998;42:1437–1446. doi: 10.1128/aac.42.6.1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Casadevall A., Pirofski L. Insights into mechanisms of antibody-mediated immunity from studies with Cryptococcus neoformans. Curr Mol Med. 2005;5:421–433. doi: 10.2174/1566524054022567. [DOI] [PubMed] [Google Scholar]
- 23•.Subramanian G.M., Cronin P.W., Poley G., Weinstein A., Stoughton S.M., Zhong J., Ou Y., Zmuda J.F., Osborn B.L., Freimuth W.W. A phase 1 study of PAmAb, a fully human monoclonal antibody against Bacillus anthracis protective antigen, in healthy volunteers. Clin Infect Dis. 2005;41:12–20. doi: 10.1086/430708. [DOI] [PubMed] [Google Scholar]; This study evaluated the safety profile and bioavailability of a human mAb to B. anthracis protective antigen, previously shown to be protective against inhalation anthrax in animal models, in human, underscoring that human mAb development is a viable approach to preparedness for agents of bioterror.
- 24•.Roberts A., Thomas W.D., Guarner J., Lamirande E.W., Babcock G.J., Greenough T.C., Vogel L., Hayes N., Sullivan J.L., Zaki S. Therapy with a severe acute respiratory syndrome-associated coronavirus-neutralizing human monoclonal antibody reduces disease severity and viral burden in golden Syrian hamsters. J Infect Dis. 2006;193:685–692. doi: 10.1086/500143. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study demonstrated the efficacy of a human mAb to SARS when given after the onset of infection. The efficacy of the mAb was associated with reduced pulmonary damage and inflammation, suggesting its mechanism of action involved immunomodulation.
- 25.Sheoran A.S., Chapman-Bonofiglio S., Harvey B.R., Mukherjee J., Georgiou G., Donohue-Rolfe A., Tzipori S. Human antibody against shiga toxin 2 administered to piglets after the onset of diarrhea due to Escherichia coli O157:H7 prevents fatal systemic complications. Infect Immun. 2005;73:4607–4613. doi: 10.1128/IAI.73.8.4607-4613.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Casadevall A., Pirofski L. Antibody mediated regulation of cellular immunity and the inflammatory response. Trends Immunol. 2003;24:474–478. doi: 10.1016/s1471-4906(03)00228-x. [DOI] [PubMed] [Google Scholar]
- 27••.Ter M.J., van den Brink E.N., Poon L.L., Marissen W.E., Leung C.S., Cox F., Cheung C.Y., Bakker A.Q., Bogaards J.A., van D.E. human monoclonal antibody combination against SARS coronavirus: synergy and coverage of escape mutants. PLoS. Med. 2006;3:e237. doi: 10.1371/journal.pmed.0030237. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study demonstrates that a combination of human mAbs prevented the development of escape mutants of the SARS coronavirus. The specificity of mAbs for a single determinant has been seen as a limitation of their use for diseases caused by mutable agents. This study shows that the use of combinations of mAbs can circumvent this problem, enhancing the feasibility of combination mAb therapy for diseases caused by mutable agents.
- 28.Anonymous Human rabies prevention–United States, 1999. recommendations of the advisory committee on immunization practices. MMWR Morb Mortal Wkly Rep. 1999;48:1–21. [PubMed] [Google Scholar]
- 29.Casadevall A., Pirofski L. The Damage-response framework of microbial pathogenesis. Nat Rev Microbiol. 2003;1:17–24. doi: 10.1038/nrmicro732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30•.Bloom B.T. INH-A21: a donor-selected Staphylococcal human immune globulin for the prevention of late-onset neonatal Staphylococcal infection. Expert Opin Investig Drugs. 2006;15:703–707. doi: 10.1517/13543784.15.6.703. [DOI] [PubMed] [Google Scholar]; This study evaluated the efficacy of hyperimmune globulin derived from individuals with high levels of antibodies to Staphylococcus aureus for prevention of staphylococcal sepsis in infants. The study is notable for showing promise against an increasingly common drug resistant nosocomial threat in a high-risk population.
- 31.Bloom B., Schelonka R., Kueser T., Walker W., Jung E., Kaufman D., Kesler K., Roberson D., Patti J., Hetherington S. Multicenter study to assess safety and efficacy of INH-A21, a donor-selected human staphylococcal immunoglobulin, for prevention of nosocomial infections in very low birth weight infants. Pediatr Infect Dis J. 2005;24:858–866. doi: 10.1097/01.inf.0000180504.66437.1f. [DOI] [PubMed] [Google Scholar]
- 32.Casadevall A. Antibodies for defense against biological attack. Nat Biotechnol. 2002;20:114. doi: 10.1038/nbt0202-114. [DOI] [PubMed] [Google Scholar]
- 33•.Orange J.S., Hossny E.M., Weiler C.R., Ballow M., Berger M., Bonilla F.A., Buckley R., Chinen J., El-Gamal Y., Mazer B.D. Use of intravenous immunoglobulin in human disease: a review of evidence by members of the Primary Immunodeficiency Committee of the American Academy of Allergy, Asthma and Immunology. J Allergy Clin Immunol. 2006;117:S525–S553. doi: 10.1016/j.jaci.2006.01.015. [DOI] [PubMed] [Google Scholar]; This review provides a rigorous, evidenced based analysis of the potential benefit of intravenous immunoglobulin for inflammatory and infectious diseases. The article summarizes important studies in an area that has been fraught with controversy and a paucity of universal recommendations.
- 34.Mouthon L., Guillevin L., Tellier Z. Intravenous immunoglobulins in autoimmune- or parvovirus B19-mediated pure red-cell aplasia. Autoimmun Rev. 2005;4:264–269. doi: 10.1016/j.autrev.2004.10.004. [DOI] [PubMed] [Google Scholar]
- 35.Kaul R., McGeer A., Norrby-Teglund A., Kotb M., Schwartz B., O’Rourke K., Talbot J., Low D.E. The Canadian Streptococcal Study Group: Intravenous immunoglobulin therapy for streptococcal toxic shock syndrome - a comparative observational study. Clin Infect Dis. 1999;28:800–807. doi: 10.1086/515199. [DOI] [PubMed] [Google Scholar]
- 36.Haley M., Retter A.S., Fowler D., Gea-Banacloche J., O’Grady N.P. The role for intravenous immunoglobulin in the treatment of West Nile virus encephalitis. Clin Infect Dis. 2003;37:e88–e90. doi: 10.1086/377172. [DOI] [PubMed] [Google Scholar]
- 37.Alejandria M.M., Lansang M.A., Dans L.F., Mantaring J.B. Intravenous immunoglobulin for treating sepsis and septic shock. Cochrane Database Syst Rev. 2002;2 doi: 10.1002/14651858.CD001090. CD001090. [DOI] [PubMed] [Google Scholar]
- 38.Forton D., Karayiannis P. Established and emerging therapies for the treatment of viral hepatitis. Dig Dis. 2006;24:160–173. doi: 10.1159/000090319. [DOI] [PubMed] [Google Scholar]
- 39.Trapero-Marugan M., Garcia-Buey L., Munoz C., Quintana N.E., Moreno-Monteagudo J.A., Borque M.J., Fernandez M.J., Salvanes F.R., Medina J., Moreno-Otero R. Sustained virological response to peginterferon plus ribavirin in chronic hepatitis C genotype 1 patients is associated with a persistent Th1 immune response. Aliment Pharmacol Ther. 2006;24:117–128. doi: 10.1111/j.1365-2036.2006.02954.x. [DOI] [PubMed] [Google Scholar]
- 40•.Asnis G.M., De La G.R. Interferon-induced depression in chronic hepatitis C: a review of its prevalence, risk factors, biology, and treatment approaches. J Clin Gastroenterol. 2006;40:322–335. doi: 10.1097/01.mcg.0000210099.36500.fe. [DOI] [PubMed] [Google Scholar]; This article describes the etiologies of depression in patients with hepatitis C, including the contribution of interferon therapy. This review serves as an important reminder that immunomodulation can have toxicity and that the therapy for infectious diseases, particularly those that are chronic, requires a multidisciplinary approach.
- 41••.Pappas P.G., Bustamante B., Ticona E., Hamill R.J., Johnson P.C., Reboli A., Aberg J., Hasbun R., Hsu H.H. Recombinant interferon- gamma 1b as adjunctive therapy for AIDS-related acute cryptococcal meningitis. J Infect Dis. 2004;189:2185–2191. doi: 10.1086/420829. [DOI] [PubMed] [Google Scholar]; This study compared the efficacy of adjunctive IFN–γ or placebo with amphotericin B in patients with HIV-associated cryptococcal meningitis. The trend towards a better outcome in interferon-treated patients demonstrated the feasibility of immunotherapy in immunocompromised patients and calls for further studies to define the patients who could benefit from adjunctive IFN–γ.
- 42.Lutz J.E., Clemons K.V., Stevens D.A. Enhancement of antifungal chemotherapy by interferon-gamma in experimental systemic cryptococcosis. J Antimicrob Chemother. 2000;46:437–442. doi: 10.1093/jac/46.3.437. [DOI] [PubMed] [Google Scholar]
- 43•.Siddiqui A.A., Brouwer A.E., Wuthiekanun V., Jaffar S., Shattock R., Irving D., Sheldon J., Chierakul W., Peacock S., Day N. IFN-gamma at the site of infection determines rate of clearance of infection in cryptococcal meningitis. J Immunol. 2005;174:1746–1750. doi: 10.4049/jimmunol.174.3.1746. [DOI] [PubMed] [Google Scholar]; This study evaluated the CSF response to C. neoformans infection and therapy in patients with HIV-associated cryptococcal meningitis. The results suggested that IFN–γ levels were positively correlated with the response to treatment. The significance of the study is that it provided a real time evaluation of the immune response in patients that was consistent with animal models and in vitro studies, demonstrating the crucial role of IFN–γ in host defense against C. neoformans.
- 44.A controlled trial of interferon gamma to prevent infection in chronic granulomatous disease. The International Chronic Granulomatous Disease Cooperative Study Group.N Engl J Med 1991, 324:509–516. [DOI] [PubMed]
- 45.Antachopoulos C., Roilides E. Cytokines and fungal infections. Br J Haematol. 2005;129:583–596. doi: 10.1111/j.1365-2141.2005.05498.x. [DOI] [PubMed] [Google Scholar]
- 46.Rowe J.M. Treatment of acute myeloid leukemia with cytokines: effect on duration of neutropenia and response to infections. Clin Infect Dis. 1998;26:1290–1294. doi: 10.1086/516360. [DOI] [PubMed] [Google Scholar]
- 47.Hazel D.L., Newland A.C., Kelsey S.M. Malignancy: granulocyte colony stimulating factor increases the efficacy of conventional amphotericin in the treatment of presumed deep-seated fungal infection in neutropenic patients following intensive chemotherapy or bone marrow transplantation for haematological malignancies. Hematology. 1999;4:305–311. doi: 10.1080/10245332.1999.11746453. [DOI] [PubMed] [Google Scholar]
- 48••.Dignani M.C., Rex J.H., Chan K.W., Dow G., Magalhaes-Silverman M., Maddox A., Walsh T., Anaissie E. Immunomodulation with interferon-gamma and colony-stimulating factors for refractory fungal infections in patients with leukemia. Cancer. 2005;104:199–204. doi: 10.1002/cncr.21142. [DOI] [PubMed] [Google Scholar]; This study showed the benefit of cytokines and CSF for invasive fungal disease in a small number of patients with leukemia. The promising results of this study support the use of immunomodulators as salvage therapy in desperately ill patients, although their benefit has not been established in large scale placebo-controlled trials.
- 49.Ott V.L., Fong D.C., Cambier J.C. Fc gamma RIIB as a potential molecular target for intravenous gamma globulin therapy. J Allergy Clin Immunol. 2001;108:S95–S98. doi: 10.1067/mai.2001.117822. [DOI] [PubMed] [Google Scholar]
- 50.O'Sullivan G.C., Kelly P., O’Halloran S., Collins C., Collins J.K., Dunne C., Shanahan F. Probiotics: an emerging therapy. Curr Pharm Des. 2005;11:3–10. doi: 10.2174/1381612053382368. [DOI] [PubMed] [Google Scholar]
- 51.Ezendam J., Van Loveren H. Probiotics: immunomodulation and evaluation of safety and efficacy. Nutr Rev. 2006;64:1–14. doi: 10.1111/j.1753-4887.2006.tb00168.x. [DOI] [PubMed] [Google Scholar]
- 52•.van de B.D., de G.J., McIntyre P., Prasad K. Steroids in adults with acute bacterial meningitis: a systematic review. Lancet Infect Dis. 2004;4:139–143. doi: 10.1016/S1473-3099(04)00937-5. [DOI] [PubMed] [Google Scholar]; This article reviews evidence to support the use of adjunctive corticosteroids in the treatment of bacterial meningitis and concludes that they should be used presumptively in the management of suspected community acquired bacterial meningitis.
- 53.Skerrett S.J., Park D.R. Anti-inflammatory treatment of acute and chronic pneumonia. Semin Respir Infect. 2001;16:76–84. doi: 10.1053/srin.2001.22731. [DOI] [PubMed] [Google Scholar]
- 54•.Thwaites G.E., Nguyen D.B., Nguyen H.D., Hoang T.Q., Do T.T., Nguyen T.C., Nguyen Q.H., Nguyen T.T., Nguyen N.H., Nguyen T.N. Dexamethasone for the treatment of tuberculous meningitis in adolescents and adults. N Engl J Med. 2004;351:1741–1751. doi: 10.1056/NEJMoa040573. [DOI] [PubMed] [Google Scholar]; This study compared the efficacy of adjunctive dexamethasone or placebo for tuberculous meningitis and demonstrated a survival benefit in individuals over the age of 14 years. The importance of this study is that it provided evidence that disproved the longstanding belief that corticosteroids aggravate mycobacterial disease. The result corroborates studies demonstrating the importance of reducing damage in the CNS in therapy for bacterial meningitis (see [52•]).
- 55.Simmons C.P., Thwaites G.E., Quyen N.T., Chau T.T., Mai P.P., Dung N.T., Stepniewska K., White N.J., Hien T.T., Farrar J. The clinical benefit of adjunctive dexamethasone in tuberculous meningitis is not associated with measurable attenuation of peripheral or local immune responses. J Immunol. 2005;175:579–590. doi: 10.4049/jimmunol.175.1.579. [DOI] [PubMed] [Google Scholar]
- 56•.Amsden G.W. Anti-inflammatory effects of macrolides–an underappreciated benefit in the treatment of community-acquired respiratory tract infections and chronic inflammatory pulmonary conditions? J Antimicrob Chemother. 2005;55:10–21. doi: 10.1093/jac/dkh519. [DOI] [PubMed] [Google Scholar]; This article reviews available evidence for beneficial immunomodulation, apart from antimicrobial activity, by macrolides in patients with chronic pulmonary disease.
- 57.Schultz M.J. Macrolide activities beyond their antimicrobial effects: macrolides in diffuse panbronchiolitis and cystic fibrosis. J Antimicrob Chemother. 2004;54:21–28. doi: 10.1093/jac/dkh309. [DOI] [PubMed] [Google Scholar]
- 58.Sau K., Mambula S.S., Latz E., Henneke P., Golenbock D.T., Levitz S.M. The antifungal drug amphotericin B promotes inflammatory cytokine release by a Toll-like receptor- and CD14-dependent mechanism. J Biol Chem. 2003;278:37561–37568. doi: 10.1074/jbc.M306137200. [DOI] [PubMed] [Google Scholar]
- 59.Moore L.J., Pridmore A.C., Dower S.K., Read R.C. Penicillin enhances the Toll-like receptor 2-mediated proinflammatory activity of Streptococcus pneumoniae. J Infect Dis. 2003;188:1040–1048. doi: 10.1086/378238. [DOI] [PubMed] [Google Scholar]