

Highlights
-
•
There is an urgent need for antivirals against Chikungunya virus (CHIKV), a medically significant virus.
-
•
Novel compounds with viral targets offer a new direction for anti-CHIKV research.
-
•
More research into CHIKV biology is required to identify new viral targets.
Abstract
Chikungunya virus (CHIKV) has re-emerged as a significant public health threat since the 2005 chikungunya fever epidemic in La Réunion. Driven by the medical importance of this virus, as well as the lack of approved antivirals, research into the field of CHIKV antivirals has recently intensified. Potential therapeutics that have been reported to show anti-CHIKV activity in vitro range from known broad-spectrum antivirals like chloroquine to novel strategies involving RNA silencing technology. Although most of the earlier efforts focused on compounds that target host components, some recent studies have reported viral targets such as nonstructural proteins. This article examines the reported in vitro and in vivo efficacies, as well as the therapeutic potential of these antiviral compounds.
Introduction
Chikungunya virus (CHIKV) is a mosquito-transmitted alphavirus belonging to the family Togaviridae [1]. CHIKV is the causative agent of chikungunya fever, a disease characterized by myalgia, polyarthralgia, fever, nausea, headaches and maculopapular rash 1, 2. Owing to mechanisms that are poorly understood, recurrent and persistent myalgia and arthralgia have been reported to last for years after the infection clears in some patients. A recent study with a macaque model suggested that the chronic phase could be caused by inflammatory responses toward persistent CHIKV in certain tissues, rather than an autoimmune-mediated response, as was initially believed 3, 4.
CHIKV was first isolated in the Makondé plateau in Tanzania during the earliest recorded epidemic in 1953 [5]. In 2005, the re-emergence of CHIKV in several Indian Ocean islands caused outbreaks of unprecedented magnitude. In the French island of La Réunion alone, one-third its 785,000 inhabitants were infected with CHIKV, resulting in more than 250 fatalities 1, 6. Apart from the newfound pathogenicity, chikungunya fever cases reported during the La Réunion epidemic were also associated with complications including lymphopenia, lethal hepatitis, dermatologic lesions and encephalitis 7, 8. Cases of fetal transmission were also recorded, leading to miscarriages and neonatal encephalopathy [9]. Apart from the classical transmission vector, Aedes aegypti, an additional mosquito vector, Aedes albopictus, was noted during this epidemic. This was accompanied by a mutation of alanine to valine in the CHIKV E1 glycoprotein (A226V), which mediates viral fusion 1, 6. A. albopictus, although native to Southeast Asia, is today established in almost all southern temperate regions [10]. To date, CHIKV has infected millions of people in more than 40 countries, including India, Malaysia, Indonesia, Thailand, Singapore, USA, Italy, Cambodia and Yemen 11, 12, 13, 14.
Despite the medical threat posed by CHIKV, there is currently no approved antiviral treatment or vaccine for CHIKV infection. Treatments are usually symptomatic, with administration of non-steroidal anti-inflammatory drugs or corticosteroids to control arthralgia and myalgia 15, 16. Given the efficiency of the mosquito vectors in establishing CHIKV infection in many geographical regions, there is an urgent need for the development of safe and effective antivirals against CHIKV to control symptoms and minimize complications in future epidemics. With the re-emergence of CHIKV in recent years, numerous efforts have been made toward the development of effective antivirals targeting CHIKV. This review highlights recent developments in the field of antiviral therapy for CHIKV infections. A summary of compounds that have exhibited anti-CHIKV activities is listed in Table 1 . The concentrations of compounds that produced 50% of the total anti-CHIKV effect (EC50), as well as the concentrations that caused cytotoxicity of 50% of the cells (CC50), are also displayed in Table 1.
Table 1.
Compounds with inhibitory activities against CHIKV
| Compound | Structure | EC50(μM; unless stated otherwise) | CC50(μM; unless stated otherwise) | Refs |
|---|---|---|---|---|
| Inhibitors of viral entry | ||||
| Chloroquine |  |
7.0a 10.0b 17.2c |
∼260d | [31] |
| Arbidol |  |
12.2 | 376 | [36] |
| Chlorpromazine |  |
39.4 | 67.3 | [39] |
| Perphenazine | 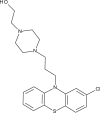 |
48.1 | 155 | [39] |
| Ethopropazine | 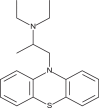 |
61.5 | 166.9 | [39] |
| Thiethylperazine |  |
63.8 | 83.1 | [39] |
| Thioridazine | 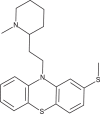 |
71.5 | 179.4 | [39] |
| Methdilazine |
 |
84.5 |
63.8 |
[39] |
| Inhibitors of viral protein translation | ||||
| siRNA (against CHIKV nsP3 and E1) | N/A | N/R | N/R | [40] |
| shRNA (against CHIKV nsP1 and E1) | N/A | N/R | N/R | [41] |
| Harringtonine |  |
0.24 | >10 | [43] |
| Homoharringtonine |
 |
N/R |
>10 |
[43] |
| Inhibitors of viral replicase | ||||
| Apigenin |  |
70.8 | >200 | [39] |
| Chrysin |  |
126.6 | >200 | [39] |
| Naringenin |  |
118.4 | 94.1 | [39] |
| Silybin |  |
92.3 | >200 | [39] |
| Prothipendyl |  |
97.3 | >200 | [39] |
| Trigocherrin A |  |
1.5 | 35 | [47] |
| Trigocherrin B |  |
2.6 | 93 | [47] |
| Trigocherrin F |  |
3.0 | 23.1 | [47] |
| Trigocherriolide A | 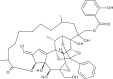 |
1.9 | 4.6 | [47] |
| Trigocherriolide B | 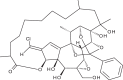 |
2.5 | 5.3 | [47] |
| Trigocherriolide C |  |
3.9 | 10.5 | [47] |
| ID1452-2 |  |
31 | >31 | [51] |
| Compound 1 |
 |
5 |
72 |
[52] |
| Inhibitors of viral genome replication | ||||
| Ribavirin | 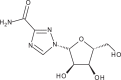 |
341.1 | 30.7 mM | [57] |
| 6-Azauridine |  |
0.82 | 208 | [57] |
| Mycophenolic acid |
 |
0.1 (MOI 0.01) 0.21 (MOI 0.1) 1.8 (MOI 1.0) 3.21 (MOI 10) 7.81 (MOI 100) |
30 |
[59] |
| Modulators of host immune response | ||||
| IFN-α2a | N/A | 11.1 IU/ml | >10,000 IU/ml | [57] |
| IFN-α2b | N/A | 9.7 IU/ml | >10,000 IU/ml | [57] |
| Polyinosinic acid:polycytidylic acid |
N/A |
N/R |
N/R |
[73] |
| Inhibitor of viral glycoprotein maturation | ||||
| Decanoyl-RVKR-chloromethyl ketone |
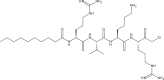 |
∼50d |
N/R |
[24] |
| Inhibitors with unknown targets | ||||
| Trigowiin A | 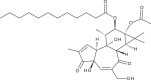 |
43.5 | >100 | [76] |
| Prostratin | 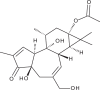 |
2.6 | 79 | [76] |
| 12-O-Tetradecanoylphorbol 13-acetate (TPA) |  |
0.0029 | 5.7 | [76] |
| 4α-12-O-Tetradecanoylphorbol 13-acetate (4α-TPA) |  |
2.8 | 5.3 | [76] |
CHIKV, Chikungunya virus; EC50, 50% effective concentration; CC50, 50% cytotoxic concentration; N/A, not applicable; N/R, not reported.
Values for pre-treatment with compound.
Values for co-treatment with compound.
Values for post-treatment with compound.
Estimated value.
General virology
The CHIKV replication cycle (Fig. 1 ) offers a good starting point for the identification of potential targets during the development of antiviral compounds. Although knowledge of CHIKV biology is lacking in many areas, mechanisms of viral attachment, fusion and entry can be supposed from studies done with related alphaviruses, including Sindbis virus (SINV) and Semliki Forest virus (SFV). CHIKV is an enveloped virus with a single-stranded, RNA genome of positive polarity. The CHIKV genome is approximately 11 kb long and consists of two open reading frames: a 7 kb frame that encodes the viral nonstructural proteins (nsP1, nsP2, nsP3 and nsP4) and a 4 kb frame that encodes the viral structural proteins, including the capsid (C) and envelope (E1, E2, E3 and 6k) proteins [17]. The envelope proteins E1 and E2 form glycoprotein spikes on the viral particle surface.
Figure 1.

CHIKV replication cycle. Upon receptor binding, the virus particle is endocytosed. Within the endosome, the pH lowers, triggering the fusion of the viral envelope and endosomal membrane. This releases the nucleocapsid into the cytoplasm which disassembles, liberating the viral genome. Once the genome is in the cytoplasm, viral nonstructural proteins (nsP1–4) are translated (using the host's translation machinery) and complex to form the viral replicase. The viral replicase then synthesizes a negative-sense RNA strand from the positive-sense genome template. The negative-sense RNA serves as a template for the generation of two things: the full-length positive-sense genome (which becomes packaged into the new viral particles) and the subgenomic (26S) RNA, which enables the expression of the structural polyprotein (C-E3-E2-6K-E1). Further processing cleaves the structural polyprotein into the individual structural proteins, including the capsid, which assembles together with the genome to form the nucleocapsid. As the nucleocapsid buds out through the plasma membrane it acquires a portion of the host plasma membrane with embedded viral glycoproteins that will form the envelope of the viral particle. Abbreviations: nsP, non-structural protein; ER, endoplasmic reticulum; pE2, precursor of E2.
Initial binding to host-cell receptors is facilitated by the E2 glycoprotein. Upon receptor binding, the virus particle is endocytosed. Although studies in an A. albopictus cell line have suggested that endocytosis of CHIKV is clathrin-mediated [18], studies in mammalian cells have indicated otherwise [19]. Apart from being clathrin-independent, endocytosis into mammalian cells has been found to be dependent on Eps15 and Rab5 proteins [19]. Acidification of the endosome results in the E1-mediated fusion of the viral envelope and endomosal membrane, followed by uncoating of the viral genome [20]. Translation of viral RNA by the host cell translational machinery produces the viral nonstructural proteins, which complex to form the CHIKV replicase [21]. The viral replicase generates more copies of the viral genome, as well as the subgenomic (26S) RNA [22]. The subgenomic RNA encodes the structural polyprotein, which is processed into C, pE2, 6k and E1 [23]. During export to the plasma membrane, the pE2 precursor is cleaved by furin to yield the mature E2 and E3 proteins [24]. Finally, viral components are assembled into nucleocapsids that bud out of infected cells, acquiring a lipid envelope with embedded E1 and E2 glycoproteins [25].
Inhibitors of viral entry
Inhibition of CHIKV entry presents an attractive therapeutic strategy because the damage caused by virulence factors during intracellular viral replication can be minimized. In addition, inhibitors of entry might target extracellular components, for instance host-cell receptors, which are more accessible, thereby reducing the required dosages and limiting cytotoxicity [26].
The search for specific entry inhibitors, however, has been impeded by the poor understanding of CHIKV-specific receptors on human cellular targets. In fact, only a single putative CHIKV receptor, prohibitin, has been identified to date [27]. The identification of more CHIKV-specific receptors, especially those implicated in the in vivo pathogenesis of CHIKV infection, is crucial to drive future research into CHIKV entry inhibitors.
Chloroquine
Chloroquine, an antimalarial drug, has shown antiviral activity against several viruses, including HIV [28], severe acute respiratory syndrome coronavirus [29] and alphaviruses [30]. A dose-dependent inhibition of CHIKV replication in Vero cells was observed upon treatment with chloroquine before, during and after infection [31]. Further investigation into the mechanism of chloroquine action suggested that chloroquine disrupted endosome-mediated CHIKV internalization, possibly through the prevention of endosomal acidification. The authors also noted that the EC50 values of the different treatment groups (7 μM for pre-treatment, 10 μM for co-treatment and 17 μM for post-treatment) were similar to the plasma concentrations of chloroquine during treatment of acute malaria, indicating potential for use against CHIKV infection in vivo.
Despite this, a double-blind clinical trial conducted with 54 CHIKV patients during the La Réunion epidemic failed to show significant improvement of symptoms or decrease in the duration of viremia upon administration of chloroquine over 5 days (day 1: 600 mg; days 2 and 3: 300 mg twice daily; days 4 and 5: 300 mg) [32]. Conversely, at day 200, patients in the chloroquine treatment group complained more frequently of chronic arthralgia (P < 0.01) than those from the placebo group. By contrast, Brighton [33] reported a significant improvement in chronic arthritis symptoms upon chloroquine phosphate treatment (250 mg/day for 20 weeks) of ten patients who had been suffering from chronic arthralgia for at least 5 years following CHIKV infection.
It must be noted that in vitro studies [31] were performed at a multiplicity of infection (MOI) of 0.1, which might not reflect the true viremia during the acute phase of CHIKV infection. Indeed, viral load has been observed to reach 1010 copies of viral genome per ml of serum during acute infection [32]. Both clinical trials were conducted with a patient cohort that was too small for conclusive data on the effectiveness of chloroquine to be obtained. In addition, the difference in treatment lengths in both chloroquine trials could have had an effect on patient outcomes. The discrepancy in data from chloroquine treatment has also been observed with HIV-1-infected patients. Savarino et al. [34] observed that the decrease in viral load of HIV-1 patients was not as significant in a study where a lower dose of chloroquine was used. The authors noted that it was important to use the appropriate dosage during treatment regimens. Therefore, before dismissing chloroquine as a potential antiviral, more studies need to be performed to determine whether changes in treatment regimens would have an impact on viral load and symptoms.
Arbidol
Arbidol is a broad-spectrum antiviral that has been approved in Russia and China for the treatment and prophylaxis of influenza and other respiratory infections 35, 36. Arbidol was shown to exhibit potent inhibitory effects against CHIKV infection in MRC-5 cells (EC50 = 12.2 μM, CC50 = 376 μM) [36]. An arbidol-resistant mutant strain isolated in the same study displayed a mutation of glycine to arginine (G407R) in domain A of the CHIKV E2 glycoprotein, postulated to be involved in binding to host receptors [37]. Taken together with data from hemagglutination assays, the study suggested that, as observed with influenza viruses, arbidol inhibits viral entry by preventing adsorption of CHIKV to target cells.
Having been in clinical use for more than 15 years in Russia, arbidol has been shown to be well-tolerated with minimal side effects. In addition, no arbidol-resistant viruses have been isolated to date [38], suggesting that it might be an attractive option as an antiviral.
10H-phenothiazines
An antiviral screen conducted with a temperature-sensitive SFV mutant identified entry inhibitors that were also reported to be effective against a recombinant strain of CHIKV carrying a luciferase gene [39]. These comprised six compounds containing a 10H-phenothiazine core (Table 1), including chlorpromazine, perphenazine, ethopropazine, thiethylperazine, thioridazine and methdilazine (EC50 = 39.4–84.5 μM). Phenothiazines are clinically approved antipsychotics, and their ability to transverse the blood–brain barrier could be useful in CHIKV cases with neurologic complications, similar to those documented during the La Réunion outbreak [39]. The selected phenothiazines are expected to inhibit CHIKV entry, as they do with SFV, but studies identifying their molecular targets remain to be conducted.
Inhibitors of viral protein translation
RNA interference
Like most RNA viruses, translation of the CHIKV genome is carried out by the host cell translation machinery. RNA interference (RNAi) offers a promising approach toward inhibiting viral protein expression by selectively targeting the viral genome template. Small interfering RNA (siRNA) sequences targeting CHIKV nsP3 and E1 genes were reported to reduce CHIKV titers and genome copy numbers significantly (P < 0.05) by about 1 log10 unit at 24 hours post-infection (hpi) [40]. The inhibition was however found to be transient and diminished by 72 hpi, possibly owing to the rapidly replicating nature of alphaviruses [40] and susceptibility of siRNAs to intracellular degradation [41].
More recently, plasmid-based small hairpin RNA (shRNA) was evaluated as a potential CHIKV antiviral [41]. Stable cell clones expressing shRNA against CHIKV E1 and nsP1 displayed significant (P < 0.05) and sustained inhibition of CHIKV infection and E2 protein production for at least 3 days post-infection. Infectious titers decreased progressively from day 1 post-infection to day 3 post-infection, where CHIKV replication was completely abolished. Inoculation of 60 μg of E1 shRNA into a suckling mouse model 24 hours before infection with 106 plaque-forming units (PFU) of CHIKV resulted in complete survival of treated groups for 15 days post-infection [41]. By contrast, control groups inoculated with scrambled E1 shRNA displayed complete mortality within 10 days post-infection. Target specificity was confirmed when stable E1 shRNA clones did not show decrease in viral titers upon infection with SINV and dengue virus.
The approval of Vitravene® (fomivirsen) by the FDA in 1998 for treatment of retinitis following cytomegalovirus infection suggests that RNAi technology has the potential to be developed for therapeutic applications [42]. The efficacy of siRNA in vitro, and shRNA in vivo, suggests that RNAi could be useful in combating CHIKV infection as well. Despite its transient action, siRNA treatment could be useful in lowering viral titers to aid the host immune response in clearing infection [40]. shRNA efficacy in vivo shows promising efficacy as a prophylaxis, but the potential for use post-infection must also be explored.
Harringtonine and homoharringtonine
Harringtonine, a cephalotaxine alkaloid derived from Cephalotaxus harringtonia, displayed potent inhibition of CHIKV infection (EC50 = 0.24 μM) with minimal cytotoxicity [43]. Further investigations revealed that harringtonine inhibited production of nsP3 and E2 proteins, positive- and negative-sense CHIKV RNA, and caused a reduction in infectious CHIKV titers. Harringtonine has been found to inhibit the elongation phase of eukaryotic translation by competing with incoming aminoacyl tRNAs for binding to the A site in the 60S ribosome subunit [44]. The authors therefore postulated that the antiviral action of harringtonine could also depend on its inhibition of the eukaryotic translation machinery, directly interrupting nonstructural and structural viral protein synthesis and indirectly affecting all downstream products. Homoharringtonine, an analog of harringtonine with an additional methyl group, was also observed to possess anti-CHIKV activity. Interestingly, harringtonine and homoharringtonine displayed greater potency with a CHIKV strain carrying the E1 A226V mutation as compared with a strain carrying the wild type E1. However, a causal link between the A226V mutation and difference in drug potency was not established, because both isolates used in the study contained other genetic differences.
Homoharringtonine (omacetaxine mepesuccinate) was recently approved by the FDA for the treatment of chronic myeloid leukemia. Although harringtonine and homoharringtonine have been reported to possess antileukemia properties, homoharringtonine has been observed to be more potent owing to its ability to persist at its target sites for a longer duration [45]. As such, extensive in vivo studies in a suitable murine model are required to determine whether harringtonine or homoharringtonine is more suitable as a CHIKV therapeutic.
Inhibitors of viral replicase
5,7-Dihydroxyflavones and prothipendyl
Although the initial steps of the CHIKV replication cycle employ host factors, the replication of the viral genome represents a step that is driven mainly by viral factors. Pohjala et al. [39] developed a screening assay using BHK-CHIKV-NCT, a stable cell line expressing a CHIKV replicon containing only nonstructural proteins with enhanced green fluorescent protein and Renilla luciferase as reporters. Four natural compounds containing a 5,7-dihydroxyflavone structure were identified as CHIKV inhibitors. These were compounds with known anticancer properties [46] like apigenin, chrysin, naringenin and silybin (EC50 = 70.8–126.6 μM). Prothipendyl, a pharmaceutical phenothiazine, was also identified with an EC50 of 97.3 μM.
Given that alphavirus replicases have been found to complex with host proteins as well [21], it is necessary to verify the targets of the selected hit compounds in downstream studies. The importance of the 5,7-dihydroxyflavone structure can also be evaluated further in studies of mechanism and SAR analysis.
Daphnane diterpenoids
Efforts to identify novel antivirals have also been driven by the isolation of new compounds from natural sources. Allard et al. [47] performed a bioassay-guided purification on extracts of the bark of Trigonostemon cherrieri to screen for inhibitors of the RNA-dependent RNA polymerase portion of the dengue virus NS5 protein. From this screen, the authors isolated a new series of highly oxygenated daphnane diterpenoid orthoesters carrying an uncommon chlorinated moiety. Trigocherrins A, B and F and trigocherriolides A, B and C suppressed CHIKV-induced cytopathic effect (CPE) (EC50 = 1.5–3.9 μM, CC50 = 4.6–93 μM). Among these, the selectivity indices (SI: CC50/EC50) of trigocherrins (SI = 8–36) were significantly higher than those of trigocherriolides (SI = 2–3), suggesting that trigocherrins are more potent inhibitors of CHIKV infection. Given the targeted approach used in the screening assay, there is a possibility that these compounds inhibit the CHIKV replicase. However, the precise targets and mechanism of action of these diterpenoids have yet to be investigated in a CHIKV model.
Inhibitors of CHIKV nsP2
CHIKV nsP2 carries a protease domain involved in processing viral polyproteins, an RNA triphosphatase involved in viral RNA capping and a nucleotide triphosphatase, which supports the RNA helicase function of nsP2 [48]. CHIKV nsP2 has also been found to inhibit host antiviral response by suppressing transcription and type I/II interferon-stimulated JAK/STAT signaling 49, 50. A mutation (P718G) in the nsP2 region has been found to inhibit CHIKV replication, affirming nsP2 as an attractive target for CHIKV antivirals [51].
Lucas-Hourani et al. [51] designed a trans-reporter assay to screen for inhibitors of CHIKV nsP2. From the screen, a natural compound derivative, ID1452-2 (EC50 = 31 μM), was validated as an inhibitor of CHIKV infection in vitro using a recombinant CHIKV strain carrying a luciferase insert. Several compounds with a similar taxoid oxetan ring structure were found to be only minimally active in the trans-reporter assay, suggesting that more work needs to be done to determine the SAR of ID1452-2.
Bassetto et al. [52] developed a homology model of CHIKV nsP2 based on the crystal structure of the nsP2 of Venezuelan equine encephalitis virus (VEEV), a related alphavirus. From their virtual screen of about five million compounds, the authors identified a lead compound (Compound 1) that suppressed CHIKV-induced CPE (EC50 = 5 μM, CC50 = 72 μM). Compound 1 was predicted to bind to the central portion of the nsP2 protease active site. SAR analysis conducted with structural analogs suggested that the cyclopropylic moiety and hydrazone group of Compound 1 are important for its anti-CHIKV activity.
Inhibitors of viral genome replication
Ribavirin
Ribavirin is a synthetic purine nucleoside analog with broad-spectrum antiviral activities [53]. Ribavirin has been licensed by the FDA for the treatment of respiratory syncytial virus [54], and in combination with pegylated interferon (IFN)-α for treatment of acute and relapsing hepatitis C virus (HCV) 55, 56.
Ribavirin displayed potent inhibition of CHIKV-induced CPE (EC50 = 341.1 μM, CC50 = 30.7 mM) as well as a significant (P < 0.05) dose-dependent reduction of CHIKV titer [57]. In concordance with previous data, ribavirin also displayed a synergistic antiviral effect with IFN-α. A study conducted with patients experiencing arthritis and lower limb pains for at least 2 weeks after CHIKV infection found that administration of ribavirin (200 mg, twice a day for 7 days) reduced soft tissue swelling and joint pains [58]. Six out of the ten patients treated were able to walk freely upon ribavirin treatment and seven patients did not require analgesics after 4 weeks. Despite the promising data, the patient cohort used was too small for conclusive observations on the efficacy of ribavirin. However, the observed in vitro and in vivo anti-CHIKV activity of ribavirin does warrant further study into its potential antiviral use.
Despite its well-documented antiviral effects, the mechanism of action of ribavirin remains controversial and is likely to vary for different viruses [53]. A structural analog of guanosine, ribavirin is known to be a competitive inhibitor of inosine monophosphate dehydrogenase (IMPDH), an enzyme involved in the de novo synthesis of guanine nucleotides 53, 59. Studies with SINV have shown that ribavirin treatment depletes intracellular guanosine triphosphate (GTP) pools, and that resistance to ribavirin maps to nsP1, a guanylyltransferase that is involved in capping the viral RNA [60]. It is therefore possible that ribavirin results in an increase in degradation of intracellular viral RNA, and halts cap-dependent viral protein translation. However, mechanistic studies are still required to confirm and elucidate the mode of action for ribavirin against CHIKV.
6-Azauridine
6-Azauridine inhibited CHIKV replication and CPE at noncytotoxic concentrations (EC50 = 0.82 μM, CC50 = 208 μM) [57]. Like ribavirin, 6-azauridine is a nucleoside analog that has demonstrated broad-spectrum antiviral activity [61]. However, antiviral activity has been difficult to replicate in vivo [62]. A uridine analog, 6-azauridine is an inhibitor of orotidine monophosphate decarboxylase, which is involved in de novo synthesis of pyrimidines [63]. Clinically, 6-azauridine triacetate has been used for treatment of severe psoriasis without severe adverse effects. 6-Azauridine can be administered at high doses (200 mg/kg), reaching plasma concentrations that are higher than levels required for alphavirus inhibition, suggesting possible therapeutic use [57]. Apart from mechanistic studies, the efficacy of 6-azauridine requires careful evaluation in suitable in vivo models of CHIKV infection.
Mycophenolic acid
Mycophenolic acid (MPA) is a noncompetitive inhibitor of IMPDH, and has been used clinically as an immunosuppressant to prevent the rejection of transplant organs [64]. MPA appears to inhibit CHIKV replication in a mechanism similar to that of ribavirin. In vitro, MPA inhibited CHIKV-induced CPE across different MOIs in a dose-dependent manner (EC50 = 0.1 μM at MOI 0.01, and 7.81 μM at MOI 100, CC50 = 30 μM) [59]. A dose-dependent reduction in CHIKV genome copy number and infectious titer was also noted. The mode of action of MPA was traced to depletion of GTP pools and subsequent inhibition of CHIKV-induced apoptosis, resulting in decreased CPE. Addition of exogenous guanosine monophosphate (GMP) abolished MPA-induced inhibition of CHIKV infection. MPA-resistant SINV strains have displayed mutations in nsP1, resulting in a guanylyl methyltransferase with increased affinity for GTP, and it is possible that a similar mechanism could apply for CHIKV [60]. In addition, the authors noted that MPA treatment caused a reduction in infectivity of CHIKV from 3.33% to 0.148%, suggesting that an additional mechanism of MPA action could rely on the increase in mutation rates during viral genome replication owing to the lack of intracellular GTP [59].
Modulators of host immune response
CHIKV infection clears rapidly within 4–7 days, suggesting a crucial role for the innate immune system in the acute phase of the infection [65]. As with most viruses, type I IFNs play a central role in CHIKV infection clearance. The production of type I IFNs is believed to be triggered by detection of viral RNA by Toll-like receptors (TLRs) 3, 7 and 8, as well as Rig-like receptors during CHIKV infection [65]. Type I IFNs activate the transcription of interferon-stimulated genes (ISGs), which encode proteins involved in the host antiviral defense 66, 67. Therefore, activation of the innate immune response may contribute to a quicker and more efficient resolution of viral infection.
IFN-α
IFN-α treatment has been reported to protect mice against infections with VEEV [68], SFV [69] and nerurovirulent SINV [70]. IFN-α2a (EC50 = 11.1 IU/ml, CC50 > 10,000 IU/ml) and IFN-α2b (EC50 = 9.7 IU/ml, CC50 > 10,000 IU/ml) caused dose-dependent inhibition of CPE and CHIKV titers [57]. As was observed in the harringtonine study [43], Bordi et al. [71] noted that a CHIKV strain carrying the E1 A226V mutation was more sensitive to IFN-α treatment than a strain carrying the wild-type genotype. Protection against CHIKV-induced CPE vanished upon treatment of 0.5–1.5 IU/ml of IFN-α for isolates with the E1 A226V mutation, as opposed to 14 IU/ml for the wild-type isolate. However, the authors clarified that the genetic backgrounds of both isolates were unknown, and that additional genetic differences could have contributed to the observations. Nevertheless, it is worthwhile to ascertain the effect of the A226V mutation on antivirals, given that isolates carrying this mutation became more prevalent as the epidemic progressed in La Réunion. Evidence for the in vitro antiviral effects of IFN-α, as well as the synergistic effects of IFN-α and ribavirin, warrants further exploration in suitable in vivo models.
Polyinosinic acid:polycytidylic acid
Polyinosinic acid:polycytidylic acid [poly(I:C)] is a synthetic analog of double-stranded RNA (dsRNA) that interacts with TLR3, causing potent induction of IFN [72]. Treatment with 4 μg/ml of poly(I:C) before CHIKV infection at various MOI (0.01, 1 or 5) suppressed CPE for up to 72 hpi [73]. Under the same treatment conditions, CHIKV infectious titers were reduced by about 2–3 log10 units at 24 and 48 hpi and 1–2 log10 units at 72 hpi for all MOIs tested. Downstream studies indicated that poly(I:C) treatment induced secretion of IFN-β, which peaked at 4 hours post-treatment, as opposed to CHIKV-induced secretion of IFN-β, which peaked at 24 hpi. TLR3 expression was also upregulated during poly(I:C) treatment as well as CHIKV infection. A significant upregulation at the mRNA level was observed for IFN-α, IFN-β and interferon-inducible antiviral factors including 2′,5′-oligoadenylate synthetase (OAS) and myxovirus resistance protein (MxA) upon poly(I:C) treatment in uninfected cells. It is probable that the antiviral effects of poly(I:C) are dependent on its immunomodulatory effects. Poly(I:C) has also been used as an adjuvant in vaccine research, and this is another possible application for this TLR3 analog 74, 75.
Inhibitor of viral glycoprotein maturation
Decanoyl-RVKR-chloromethyl ketone
In the final stages of the CHIKV replication cycle, the viral precursor protein E3E2 is cleaved by furin into mature E3 and E2 [24]. Treatment of CHIKV-infected cells with decanoyl-RVKR-chloromethyl ketone (dec-RVKR-cmk), a furin inhibitor, was found to suppress the level of CHIKV infection as determined by immunofluorescence detection of alphavirus nucleocapsid. In particular, the authors observed that inhibition correlated with a decrease in the number of antigen-positive cells per CHIKV-induced focus, instead of a depletion in the number of foci. This implied that dec-RVKR-cmk impairs the spread of CHIKV to neighboring cells. Treatment of dec-RVKR-cmk before, during and after CHIKV infection also revealed significant inhibition of viral titer, indicating that dec-RVKR-cmk could inhibit early steps of the viral replication cycle as well. However, mechanistic studies investigating this have not been performed.
Inhibitors with unknown targets
Tigliane diterpenoids
Bourjout et al. isolated a novel tigliane diterpenoid, trigowiin A, during a chemical study of Trigonostemon howii. Although trigowiin A (EC50 = 43.5 μM, CC50 > 100 μM) displayed only moderate anti-CHIKV activities, several well-known tigliane diterpenoids with a similar structure displayed potent inhibition of CHIKV-induced CPE [76]. These included prostratin (EC50 = 2.6 μM, CC50 = 79 μM), 12-O-tetradecanoylphorbol 13-acetate (TPA) (EC50 = 0.0029 μM, CC50 = 5.7 μM) and 4α-TPA (EC50 = 2.8 μM, CC50 = 5.3 μM). In particular, TPA was the most potent compound of the set, with an SI of 1965. Based on preliminary SAR data, the authors proposed that antiviral activity against CHIKV requires a basic phorbol carbon skeleton esterified at position 13, with a free hydroxyl group at position 12.
Despite its attractive selectivity index, the tumor-promoting properties of TPA could make it unsuitable as an antiviral agent 76, 77. In addition, TPA has been reported to inhibit HIV-induced syncytia formation via a protein kinase C (PKC)-dependent pathway while enhancing HIV replication 77, 78. This indicates that the antiviral activities of compounds displaying CPE reduction should always be confirmed with assays determining infectious viral titers. This is especially so for TPA and its structural analogs, given the data from HIV studies.
Concluding remarks and future directions
The magnitude of the chikungunya fever epidemic in La Réunion in 2005 emphasized the importance of the search for appropriate CHIKV antivirals. Most of the antivirals discussed in this review are only in the preliminary stages of research and this can be attributed partly to the gaps in current understanding of the biology of alphaviruses. Among the potential therapeutics discussed are well-known broad-spectrum antivirals like chloroquine, ribavirin and IFN-α, which have displayed some efficacy in vivo. Having been clinically approved for other conditions, existing information on pharmacologic parameters of these compounds might aid more-extensive in vivo studies on their anti-CHIKV properties, expediting the drug development process. The recent discovery of novel antivirals targeting CHIKV components like nsP2 (ID1452-2 and Compound 1), replicase complex (5,7-dihydroxyflavones) and viral genome translation (siRNA and shRNA) have shown promise in vitro, and further studies on their in vivo efficacies are warranted, especially because compounds with viral targets are expected to display lower toxicity owing to their selectivity. Nevertheless, compounds with host targets like harringtonine (ribosome inhibitor) and dec-RVKR-cmk (furin inhibitor) remain relevant to the antiviral discovery process, as shown by the FDA approval of host-targeting compounds like ribavirin.
There is a lot that remains unknown with regards to CHIKV biology and pathogenesis in vivo. To date, the processes contributing to CHIKV-induced encephalitis and chronic arthritis are poorly understood, posing a hindrance to antiviral drug discovery. Even at the molecular level, there are gaps in the understanding of CHIKV replication, including host-cell receptors as well as the events prior to viral budding. Indeed, research into basic biology and in vivo pathogenesis of CHIKV are crucial for the identification of novel viral targets as well as antivirals.
Acknowledgement
The authors would like to thank Ms Lim Pei Jin for her valuable contributions to this paper.
References
- 1.Sourisseau M. Characterization of reemerging Chikungunya virus. PLoS Pathog. 2007;3:e89. doi: 10.1371/journal.ppat.0030089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Maek A.N.W., Silachamroon U. Presence of autoimmune antibody in Chikungunya infection. Case Rep. Med. 2009;2009:840183. doi: 10.1155/2009/840183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Labadie K. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J. Clin. Invest. 2010;120:894–906. doi: 10.1172/JCI40104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dupuis-Maguiraga L. Chikungunya disease: infection-associated markers from the acute to the chronic phase of arbovirus-induced arthralgia. PLoS Negl. Trop. Dis. 2012;6:e1446. doi: 10.1371/journal.pntd.0001446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Robinson M.C. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952–53. I. Clinical features. Trans. R. Soc. Trop. Med. Hyg. 1955;49:28–32. doi: 10.1016/0035-9203(55)90080-8. [DOI] [PubMed] [Google Scholar]
- 6.Schuffenecker I. Genome microevolution of Chikungunya viruses causing the Indian Ocean outbreak. PLoS Med. 2006;3:e263. doi: 10.1371/journal.pmed.0030263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Robin S. Neurologic manifestations of pediatric Chikungunya infection. J. Child. Neurol. 2008;23:1028–1035. doi: 10.1177/0883073808314151. [DOI] [PubMed] [Google Scholar]
- 8.Mavalankar D. Increased mortality rate associated with Chikungunya epidemic, Ahmedabad, India. Emerg. Infect. Dis. 2008;14:412–415. doi: 10.3201/eid1403.070720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lenglet Y. Chikungunya infection in pregnancy: evidence for intrauterine infection in pregnant women and vertical transmission in the parturient, Survey of the Réunion Island outbreak. J. Gynecol. Obstet. Biol. Reprod. (Paris) 2006;35:578–583. doi: 10.1016/s0368-2315(06)76447-x. [DOI] [PubMed] [Google Scholar]
- 10.Hawley W.A. Aedes albopictus in North America: probable introduction in used tires from northern Asia. Science. 1987;236:1114–1116. doi: 10.1126/science.3576225. [DOI] [PubMed] [Google Scholar]
- 11.Hapuarachchi H.C. Re-emergence of Chikungunya virus in South-east Asia: virological evidence from Sri Lanka and Singapore. J. Gen. Virol. 2010;91:1067–1076. doi: 10.1099/vir.0.015743-0. [DOI] [PubMed] [Google Scholar]
- 12.Hochedez P. Cases of Chikungunya fever imported from the islands of the South West Indian Ocean to Paris, France. Euro. Surveill. 2007;12 [Epub] [Google Scholar]
- 13.Lim P.L. Chikungunya in Singapore: imported cases among travelers visiting friends and relatives. J. Travel Med. 2009;16:289–291. doi: 10.1111/j.1708-8305.2009.00313.x. [DOI] [PubMed] [Google Scholar]
- 14.Pfeffer M., Loscher T. Cases of Chikungunya imported into Europe. Euro. Surveill. 2006;11:E060316.2. [Epub] [PubMed] [Google Scholar]
- 15.Brehin A.C. The large form of human 2′,5′-oligoadenylate synthetase (OAS3) exerts antiviral effect against Chikungunya virus. Virology. 2009;384:216–222. doi: 10.1016/j.virol.2008.10.021. [DOI] [PubMed] [Google Scholar]
- 16.Queyriaux B. Clinical burden of Chikungunya virus infection. Lancet Infect. Dis. 2008;8:2–3. doi: 10.1016/S1473-3099(07)70294-3. [DOI] [PubMed] [Google Scholar]
- 17.Sreekumar E. Genetic characterization of 2006–2008 isolates of Chikungunya virus from Kerala, South India, by whole genome sequence analysis. Virus Genes. 2010;40:14–27. doi: 10.1007/s11262-009-0411-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lee R.C. Mosquito cellular factors and functions in mediating the infectious entry of Chikungunya virus. PLoS Negl. Trop. Dis. 2013;7:e2050. doi: 10.1371/journal.pntd.0002050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bernard E. Endocytosis of Chikungunya virus into mammalian cells: role of clathrin and early endosomal compartments. PLoS ONE. 2010;5:e11479. doi: 10.1371/journal.pone.0011479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Helenius A. On the entry of Semliki forest virus into BHK-21 cells. J. Cell Biol. 1980;84:404–420. doi: 10.1083/jcb.84.2.404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Barton D.J. Solubilization and immunoprecipitation of alphavirus replication complexes. J. Virol. 1991;65:1496–1506. doi: 10.1128/jvi.65.3.1496-1506.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gifford G.E., Heller E. Effect of actinomycin D on interferon production by ‘active’ and ‘inactive’ Chikungunya virus in chick cells. Nature. 1963;200:50–51. doi: 10.1038/200050a0. [DOI] [PubMed] [Google Scholar]
- 23.Strauss J.H., Strauss E.G. The alphaviruses: gene expression, replication, and evolution. Microbiol. Rev. 1994;58:491–562. doi: 10.1128/mr.58.3.491-562.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ozden S. Inhibition of Chikungunya virus infection in cultured human muscle cells by furin inhibitors: impairment of the maturation of the E2 surface glycoprotein. J. Biol. Chem. 2008;283:21899–21908. doi: 10.1074/jbc.M802444200. [DOI] [PubMed] [Google Scholar]
- 25.Ekstrom M. Membrane protein lateral interactions control Semliki Forest virus budding. EMBO J. 1994;13:1058–1064. doi: 10.1002/j.1460-2075.1994.tb06354.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Teissier E. Targeting cell entry of enveloped viruses as an antiviral strategy. Molecules. 2011;16:221–250. doi: 10.3390/molecules16010221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wintachai P. Identification of prohibitin as a Chikungunya virus receptor protein. J. Med. Virol. 2012;84:1757–1770. doi: 10.1002/jmv.23403. [DOI] [PubMed] [Google Scholar]
- 28.Savarino A. Anti-HIV effects of chloroquine: inhibition of viral particle glycosylation and synergism with protease inhibitors. J. Acquir. Immune Defic. Syndr. 2004;35:223–232. doi: 10.1097/00126334-200403010-00002. [DOI] [PubMed] [Google Scholar]
- 29.Keyaerts E. In vitro inhibition of severe acute respiratory syndrome coronavirus by chloroquine. Biochem. Biophys. Res. Commun. 2004;323:264–268. doi: 10.1016/j.bbrc.2004.08.085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Helenius A. Inhibition of Semliki forest virus penetration by lysosomotropic weak bases. J. Gen. Virol. 1982;58:47–61. doi: 10.1099/0022-1317-58-1-47. [DOI] [PubMed] [Google Scholar]
- 31.Khan M. Assessment of in vitro prophylactic and therapeutic efficacy of chloroquine against Chikungunya virus in vero cells. J. Med. Virol. 2010;82:817–824. doi: 10.1002/jmv.21663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.de Lamballerie X. Antiviral treatment of Chikungunya virus infection. Infect. Disord. Drug Targets. 2009;9:101–104. doi: 10.2174/187152609787847712. [DOI] [PubMed] [Google Scholar]
- 33.Brighton S.W. Chloroquine phosphate treatment of chronic Chikungunya arthritis. An open pilot study. S. Afr. Med. J. 1984;66:217–218. [PubMed] [Google Scholar]
- 34.Savarino A. New insights into the antiviral effects of chloroquine. Lancet Infect. Dis. 2006;6:67–69. doi: 10.1016/S1473-3099(06)70361-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Boriskin Y.S. Arbidol: a broad-spectrum antiviral that inhibits acute and chronic HCV infection. Virol. J. 2006;3:56. doi: 10.1186/1743-422X-3-56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Delogu I. In vitro antiviral activity of arbidol against Chikungunya virus and characteristics of a selected resistant mutant. Antiviral. Res. 2011;90:99–107. doi: 10.1016/j.antiviral.2011.03.182. [DOI] [PubMed] [Google Scholar]
- 37.Voss J.E. Glycoprotein organization of Chikungunya virus particles revealed by X-ray crystallography. Nature. 2010;468:709–712. doi: 10.1038/nature09555. [DOI] [PubMed] [Google Scholar]
- 38.Liu M.Y. Pharmacokinetic properties and bioequivalence of two formulations of arbidol: an open-label, single-dose, randomized-sequence, two-period crossover study in healthy Chinese male volunteers. Clin. Ther. 2009;31:784–792. doi: 10.1016/j.clinthera.2009.04.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pohjala L. Inhibitors of alphavirus entry and replication identified with a stable Chikungunya replicon cell line and virus-based assays. PLoS ONE. 2011;6:e28923. doi: 10.1371/journal.pone.0028923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Dash P.K. RNA interference mediated inhibition of Chikungunya virus replication in mammalian cells. Biochem. Biophys. Res. Commun. 2008;376:718–722. doi: 10.1016/j.bbrc.2008.09.040. [DOI] [PubMed] [Google Scholar]
- 41.Lam S. Expression of plasmid-based shRNA against the E1 and nsP1 genes effectively silenced Chikungunya virus replication. PLoS ONE. 2012;7:e46396. doi: 10.1371/journal.pone.0046396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Karagiannis T.C., El-Osta A. RNA interference and potential therapeutic applications of short interfering RNAs. Cancer Gene Ther. 2005;12:787–795. doi: 10.1038/sj.cgt.7700857. [DOI] [PubMed] [Google Scholar]
- 43.Kaur P. Inhibition of Chikungunya virus replication by harringtonine, a novel antiviral that suppresses viral protein expression. Antimicrob. Agents Chemother. 2013;57:155–167. doi: 10.1128/AAC.01467-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Fresno M. Inhibition of translation in eukaryotic systems by harringtonine. Eur. J. Biochem. 1977;72:323–330. doi: 10.1111/j.1432-1033.1977.tb11256.x. [DOI] [PubMed] [Google Scholar]
- 45.Jiang T.L. Comparative in vitro antitumor activity of homoharringtonine and harringtonine against clonogenic human tumor cells. Invest. New Drugs. 1983;1:21–25. doi: 10.1007/BF00180188. [DOI] [PubMed] [Google Scholar]
- 46.Weng C.J., Yen G.C. Flavonoids, a ubiquitous dietary phenolic subclass, exert extensive in vitro anti-invasive and in vivo anti-metastatic activities. Cancer Metast. Rev. 2012;31:323–351. doi: 10.1007/s10555-012-9347-y. [DOI] [PubMed] [Google Scholar]
- 47.Allard P.M. Antiviral chlorinated daphnane diterpenoid orthoesters from the bark and wood of Trigonostemon cherrieri. Phytochemistry. 2012;84:160–168. doi: 10.1016/j.phytochem.2012.07.023. [DOI] [PubMed] [Google Scholar]
- 48.Bourai M. Mapping of Chikungunya virus interactions with host proteins identified nsP2 as a highly connected viral component. J. Virol. 2012;86:3121–3134. doi: 10.1128/JVI.06390-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Fros J.J. Chikungunya virus nonstructural protein 2 inhibits type I/II interferon-stimulated JAK-STAT signaling. J. Virol. 2010;84:10877–10887. doi: 10.1128/JVI.00949-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Akhrymuk I. Evasion of the innate immune response: the Old World alphavirus nsP2 protein induces rapid degradation of Rpb1, a catalytic subunit of RNA polymerase II. J. Virol. 2012;86:7180–7191. doi: 10.1128/JVI.00541-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lucas-Hourani M. A phenotypic assay to identify Chikungunya virus inhibitors targeting the nonstructural protein nsP2. J. Biomol. Screen. 2013;18:172–179. doi: 10.1177/1087057112460091. [DOI] [PubMed] [Google Scholar]
- 52.Bassetto M. Computer-aided identification, design and synthesis of a novel series of compounds with selective antiviral activity against Chikungunya virus. Antiviral Res. 2013;98:12–18. doi: 10.1016/j.antiviral.2013.01.002. [DOI] [PubMed] [Google Scholar]
- 53.Graci J.D., Cameron C.E. Mechanisms of action of ribavirin against distinct viruses. Rev. Med. Virol. 2006;16:37–48. doi: 10.1002/rmv.483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Cooper A.C. Management and prevention strategies for respiratory syncytial virus (RSV) bronchiolitis in infants and young children: a review of evidence-based practice interventions. Pediatr. Nurs. 2003;29:452–456. [PubMed] [Google Scholar]
- 55.Poynard T. Randomised trial of interferon alpha2b plus ribavirin for 48 weeks or for 24 weeks versus interferon alpha2b plus placebo for 48 weeks for treatment of chronic infection with hepatitis C virus, International Hepatitis Interventional Therapy Group (IHIT) Lancet. 1998;352:1426–1432. doi: 10.1016/s0140-6736(98)07124-4. [DOI] [PubMed] [Google Scholar]
- 56.Mangia A. Individualized treatment duration for hepatitis C genotype 1 patients: a randomized controlled trial. Hepatology. 2008;47:43–50. doi: 10.1002/hep.22061. [DOI] [PubMed] [Google Scholar]
- 57.Briolant S. In vitro inhibition of Chikungunya and Semliki Forest viruses replication by antiviral compounds: synergistic effect of interferon-α and ribavirin combination. Antiviral Res. 2004;61:111–117. doi: 10.1016/j.antiviral.2003.09.005. [DOI] [PubMed] [Google Scholar]
- 58.Ravichandran R., Manian M. Ribavirin therapy for Chikungunya arthritis. J. Infect. Dev. Ctries. 2008;2:140–142. [PubMed] [Google Scholar]
- 59.Khan M. Cellular IMPDH enzyme activity is a potential target for the inhibition of Chikungunya virus replication and virus induced apoptosis in cultured mammalian cells. Antiviral Res. 2011;89:1–8. doi: 10.1016/j.antiviral.2010.10.009. [DOI] [PubMed] [Google Scholar]
- 60.Scheidel L.M., Stollar V. Mutations that confer resistance to mycophenolic acid and ribavirin on Sindbis virus map to the nonstructural protein nsP1. Virology. 1991;181:490–499. doi: 10.1016/0042-6822(91)90881-b. [DOI] [PubMed] [Google Scholar]
- 61.Rada B., Dragun M. Antiviral action and selectivity of 6-azauridine. Ann. N. Y. Acad. Sci. 1977;284:410–417. doi: 10.1111/j.1749-6632.1977.tb21977.x. [DOI] [PubMed] [Google Scholar]
- 62.Klein R.J. Herpes simplex virus skin infection in hairless mice: treatment with antiviral compounds. Antimicrob. Agents Chemother. 1974;5:318–322. doi: 10.1128/aac.5.3.318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Cinatl J., Jr. Development of antiviral therapy for severe acute respiratory syndrome. Antiviral Res. 2005;66:81–97. doi: 10.1016/j.antiviral.2005.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lipsky J.J. Mycophenolate mofetil. Lancet. 1996;348:1357–1359. doi: 10.1016/S0140-6736(96)10310-X. [DOI] [PubMed] [Google Scholar]
- 65.Schwartz O., Albert M.L. Biology and pathogenesis of Chikungunya virus. Nat. Rev. Microbiol. 2010;8:491–500. doi: 10.1038/nrmicro2368. [DOI] [PubMed] [Google Scholar]
- 66.Chow A. Persistent arthralgia induced by Chikungunya virus infection is associated with interleukin-6 and granulocyte macrophage colony-stimulating factor. J. Infect. Dis. 2011;203:149–157. doi: 10.1093/infdis/jiq042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.de Veer M.J. Functional classification of interferon-stimulated genes identified using microarrays. J. Leukoc. Biol. 2001;69:912–920. [PubMed] [Google Scholar]
- 68.Grieder F.B., Vogel S.N. Role of interferon and interferon regulatory factors in early protection against Venezuelan equine encephalitis virus infection. Virology. 1999;257:106–118. doi: 10.1006/viro.1999.9662. [DOI] [PubMed] [Google Scholar]
- 69.Pinto A.J. Comparative therapeutic efficacy of recombinant interferons-alpha, -beta, and -gamma against alphatogavirus, bunyavirus, flavivirus, and herpesvirus infections. J. Interferon Res. 1990;10:293–298. doi: 10.1089/jir.1990.10.293. [DOI] [PubMed] [Google Scholar]
- 70.Ryman K.D. Alpha/beta interferon protects adult mice from fatal Sindbis virus infection and is an important determinant of cell and tissue tropism. J. Virol. 2000;74:3366–3378. doi: 10.1128/jvi.74.7.3366-3378.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bordi L. Chikungunya virus isolates with/without A226V mutation show different sensitivity to IFN-a, but similar replication kinetics in non human primate cells. New Microbiol. 2011;34:87–91. [PubMed] [Google Scholar]
- 72.Field A.K. Inducers of interferon and host resistance, II. Multistranded synthetic polynucleotide complexes. Proc. Natl. Acad. Sci. U. S. A. 1967;58:1004–1010. doi: 10.1073/pnas.58.3.1004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Li Y.G. Poly (I:C), an agonist of Toll-like receptor-3, inhibits replication of the Chikungunya virus in BEAS-2B cells. Virol. J. 2012;9:114. doi: 10.1186/1743-422X-9-114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ichinohe T. Synthetic double-stranded RNA poly(I:C) combined with mucosal vaccine protects against influenza virus infection. J. Virol. 2005;79:2910–2919. doi: 10.1128/JVI.79.5.2910-2919.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Ichinohe T. PolyI:polyC12U adjuvant-combined intranasal vaccine protects mice against highly pathogenic H5N1 influenza virus variants. Vaccine. 2009;27:6276–6279. doi: 10.1016/j.vaccine.2009.04.074. [DOI] [PubMed] [Google Scholar]
- 76.Bourjot M. Prostratin and 12-O-tetradecanoylphorbol 13-acetate are potent and selective inhibitors of Chikungunya virus replication. J. Nat. Prod. 2012;75:2183–2187. doi: 10.1021/np300637t. [DOI] [PubMed] [Google Scholar]
- 77.Chowdhury I.H. The phorbol ester TPA strongly inhibits HIV-1-induced syncytia formation but enhances virus production: possible involvement of protein kinase C pathway. Virology. 1990;176:126–132. doi: 10.1016/0042-6822(90)90237-l. [DOI] [PubMed] [Google Scholar]
- 78.Nabel G., Baltimore D. An inducible transcription factor activates expression of human immunodeficiency virus in T cells. Nature. 1987;326:711–713. doi: 10.1038/326711a0. [DOI] [PubMed] [Google Scholar]