

Abstract
Interleukin-34 (IL-34) is a novel cytokine that plays an important role in innate immunity and inflammatory processes by binding to the colony-stimulating factor-1 receptor (CSF-1R). However, information on the function of IL-34 in fish remains limited. In the present study, we identified an IL-34 homolog from mudskippers (Boleophthalmus pectinirostris). In silico analysis showed that the mudskipper IL-34 (BpIL-34) was similar to other known IL-34 variants in sequence and structure and was most closely related to an orange-spotted grouper (Epinephelus coioides) homolog. BpIL-34 transcripts were constitutively expressed in various tissues, with the highest level of expression found in the brain. Edwardsiella tarda infection significantly up-regulated the mRNA expression of BpIL-34 in the mudskipper tissues. The recombinant mature BpIL-34 peptide (rBpIL-34) was purified and used to produce anti-rBpIL-34 IgG. Western blot analysis combined with PNGase F digestion revealed that native BpIL-34 in monocytes/macrophages (MOs/MФs) was N-glycosylated. In vitro, rBpIL-34 treatment enhanced the phagocytotic and bactericidal activity of mudskipper MOs/MФs, as well as the mRNA expression of pro-inflammatory cytokines like tumor necrosis factor α (BpTNF-α) and BpIL-1β in these cells. Furthermore, the knockdown of mudskipper CSF-1R1 (BpCSF-1R1), but not mudskipper BpCSF-1R2, significantly inhibited the rBpIL-34-mediated enhanced effect on MO/MФ function. In conclusion, our results indicate that mudskipper BpIL-34 modulates the functions of MOs/MФs via BpCSF-1R1.
Keywords: Interleukin-34, Mudskipper, Monocyte/macrophage function, Edwardsiella tarda, Colony-stimulating factor-1 receptor
INTRODUCTION
Cytokines comprise a large group of proteins, peptides, and glycoproteins that are secreted by specific cells of the immune system to mediate and regulate immunity, inflammation, and hematopoiesis (Wang & Secombes, 2013; Zou & Secombes, 2016). There are many types of cytokines including chemokines, interferons, interleukins, lymphokines, and tumor necrosis factor. In recent years, considerable progress has been made in the cloning, sequencing, and functional analysis of fish cytokine genes (Ogryzko et al., 2014; Secombes & Zou, 2017; Wang & Secombes, 2013). Our increasing knowledge on the regulation of fish immune responses by cytokines and their potential use as candidate drugs for disease control in aquaculture makes fish cytokine biology an attractive and rapidly expanding field (Secombes, 2016).
In a brilliant study that aimed to understand the system of secreted proteins and receptors involved in cell–cell signaling, interleukin 34 (IL-34), a secreted protein with high functional selectivity that stimulates monocyte survival, was identified as the second ligand of the colony-stimulating factor-1 receptor (CSF-1R) (Lin et al., 2008). The IL-34 structure is comprised of two β strands, four short helices, and four long helices, with an antiparallel four-helix core homologous to that of CSF-1 (Ma et al., 2012). CSF-1 is a cytokine that controls the production, differentiation, and function of macrophages, and CSF-1R mediates most, if not all, of the biological effects of CSF-1 (Liu et al., 2012; Nakamichi et al., 2013). IL-34 binds to CSF-1R with higher affinity than that of CSF-1 and induces the enhanced phosphorylation of CSF-1R and stronger activation of signaling pathways (Chihara et al., 2010). In addition, IL-34, but not CSF-1, binds to two other receptors, namely PTP-ζ and syndecan-1 (Baghdadi et al., 2018). PTP-ζ is a chondroitin sulfate proteoglycan, with expression restricted to the brain (neural progenitors and glial cells) and kidney (Masteller & Wong, 2014). IL-34 binds to the extracellular domains of PTP-ζ, resulting in the activation of several signaling pathways that regulate cell proliferation, motility, and clonogenicity (Baghdadi et al., 2018). The binding of IL-34 to syndecan-1 modulates the activation of CSF-1R and appears to be involved in the regulation of myeloid cell migration (Vasek et al., 2016). IL-34 and CSF-1 are equivalent in their ability to induce macrophage differentiation but exhibit different polarization potentials (Baghdadi et al., 2018). In contrast to CSF-1, IL-34 has a restricted pattern of expression (Nandi et al., 2012) and seems to be required for the generation of a restricted set of tissue-specific macrophages, including microglia and Langerhans cells, two types of phagocytes present in the brain and skin, respectively (Greter et al., 2012; Nakamichi et al., 2013; Wang et al., 2012). Over the past decade, accumulating evidence has shown that IL-34 exerts potent immunomodulatory effects under many pathological states such as infections, inflammatory diseases, autoimmune diseases, cancer, transplant rejection, and neurological diseases (Baghdadi et al., 2018; Grayfer & Robert, 2014, 2015; Kim & Turka, 2015; Masteller & Wong, 2014).
In fish, single copies of the IL-34 gene have been found in the genome of rainbow trout (Oncorhynchus mykiss), zebrafish (Danio rerio), fugu (Takifugu rubripes), orange-spotted grouper (Epinephelus coioides), large yellow croaker (Larimichthys crocea), and grass carp (Ctenopharyngodon idella) (Mo et al., 2015; Wang et al., 2013, 2018; Xue et al., 2019). To date, however, information on the expression of IL-34 in fish is still limited. Analysis of gene expression in tissues showed that IL-34 is constitutively expressed in rainbow trout, large yellow croaker, and grass carp, with highest expression in the spleen (Wang et al., 2013, 2018; Xue et al., 2019); however, in orange-spotted grouper, the highest level of expression is reported in the brain (Mo et al., 2015). Furthermore, IL-34 expression is tightly associated with pathogen stimulation. For example, studies on rainbow trout and grouper have demonstrated that parasitic infection can lead to elevated IL-34 transcript levels in tissues, especially in infected sites (Mo et al., 2015; Wang et al., 2013). Stimulation by viral or bacterial pathogens has also been shown to up-regulate the expression of IL-34 in large yellow croaker (Wang et al., 2018), golden pompano (Trachinotus ovatus) (Wu et al., 2019), and grass carp (Xue et al., 2019). However, the functions and underlying mechanisms of IL-34 in fish immune responses remain unclear. Recent study showed that recombinant grass carp IL-34 produced in bacteria and HEK293T cells has a stimulatory effect on the expression of IL-1β, IL-6, and IL-8 but inhibits the expression of IL-10 and TGF-β in primary head kidney macrophages (Xue et al., 2019), suggesting that IL-34 is a pro-inflammatory cytokine. Another study on zebrafish showed that ectopically expressed IL-34 can induce macrophage migration to the liver in vivo (Jiang et al., 2019). As putative receptors for IL-34, CSF-1Rs have already been identified as specific markers of macrophages in some teleost fish species, such as gilthead seabream (Sparus aurata) (Roca et al., 2006), goldfish (Carassius auratus) (Katzenback & Belosevic, 2012), ayu (Plecoglossus altivelis) (Chen et al., 2016b), and grass carp (Chen et al., 2015). Unlike mammals, teleost fish commonly possess two CSF-1R genes (CSF-1R1 and CSF-1R2), which are widely expressed in tissues, including the head kidney and spleen (Chen et al., 2015, 2016b; Dan et al., 2013; Honda et al., 2005; Katzenback & Belosevic, 2012; Mulero et al., 2008; Williams et al., 2002). A soluble form of CSF-1R has also been described in goldfish, which is important in the regulation of leucocyte-related immune responses (Rieger et al., 2013, 2014, 2015). However, the relationship between IL-34 and CSF-1Rs in fish has not been investigated to date.
Mudskipper fish (Boleophthalmus pectinirostris) are usually found in soft mudflat estuaries and coastal waters after ebb tides. Their behavioral, physiological, and morphological characteristics have become specialized and adapted to an amphibious lifestyle (You et al., 2018). The complete genomic sequences of mudskippers show many immune genes for adaptations to their complex habitats (You et al., 2014). Edwardsiella tarda is a gram-negative, facultative anaerobic bacterium that infects multifarious hosts, including fish, amphibians, and humans (Slaven et al., 2001; Xu & Zhang, 2014). Bacterial infections cause large aquacultural losses in Asia, especially in China and Japan (Xu & Zhang, 2014). Our previous report showed that intraperitoneal infection with E. tarda is lethal to mudskippers (Chen et al., 2016a). Given the importance of IL-34 in anti-bacterial innate immune responses, studies on the biological function and mechanism of IL-34 in mudskippers against E. tarda infection are crucial. In this study, we identified an IL-34 gene (BpIL-34) in mudskippers based on genomic sequences and investigated mRNA expression in the tissues of healthy and E. tarda-infected fish. Moreover, we prepared the recombinant mature peptide of BpIL-34 (rBpIL-34) and determined its effect on monocyte/macrophage (MO/MФ) functions, including phagocytosis, bactericidal activity, and cytokine mRNA expression. Furthermore, two mudskipper CSF-1R genes (BpCSF-1R1 and BpCSF-1R2) were identified, with the effect of BpCSF-1R1 and BpCSF-1R2 knockdown on BpIL-34-activated MO/MФ functions also investigated. These findings should provide new insights into IL-34 as a drug candidate for controlling biological diseases in fish.
MATERIALS AND METHODS
Fish maintenance
Healthy mudskippers without visible pathological symptoms, weighing 35–40 g each, were purchased from a commercial farm in Ningbo city, China. The fish were kept at a salinity of 10 and temperature of 24–26 °C in a filtered-water recirculating system. The fish were allowed two weeks to acclimatize to laboratory conditions. In addition, they were randomly selected for polymerase chain reaction (PCR) determination of E. tarda in blood and liver with E. tarda-specific primers gyrBF (5′–TGGCGACACCGAGCAGA–3′) and gyrBR (5′–ACAAACGCCTTAATCCCACC–3′) and showed to be E. tarda-free (Guan et al., 2017). All experiments were performed according to the Experimental Animal Management Law of China and approved by the Animal Ethics Committee of Ningbo University.
Molecular characterization of BpIL-34 cDNA
The cDNA sequence of BpIL-34 was retrieved from our transcriptome data of mudskipper by BLAST search (http://blast.ncbi.nlm.nih.gov/Blast.cgi) and verified in mudskipper genomic sequences (You et al., 2014). The molecular weight and isoelectric point of putative BpIL-34 were calculated using the ExPASy Compute pI/MW tool ( https://www.expasy.org/tools). SignalP 5.0 ( http://www.cbs.dtu.dk/services/SignalP/) was used to predict the sequence of the signal peptide. SMART ( http://smart.embl-heidelberg.de/) was used to predict the domain architecture of the putative protein. Potential N-glycosylation sites were predicted using the NetNGlyc1.0 Server ( http://www.cbs.dtu.dk/services/NetNGlyc/). Multiple sequence alignments were performed and analyzed using ClustalW ( http://clustalw.ddbj.nig.ac.jp/). Phylogenetic tree analysis was conducted with MEGA v5.0 (Tamura et al., 2011). The related IL-34 and CSF-1 sequences are listed in Table 1.
1. IL-34 and CSF-1 sequences used for multiple sequence alignment and phylogenetic tree analysis.
| GenBank accession No. | Species | Gene | |
| Latin name | English name | ||
| XM_020935293 | Boleophthalmus pectinirostris | Mudskipper | IL-34 |
| NM_001128701 | Danio rerio | Zebrafish | IL-34 |
| KM350155 | Epinephelus coioides | Orange-spotted grouper | IL-34 |
| NM_001257301 | Oncorhynchus mykiss | Rainbow trout | IL-34 |
| XM_030427145 | Sparus aurata | Gilthead seabream | IL-34 |
| NM_001305607 | Takifugu rubripes | Tiger puffer | IL-34 |
| XM_020104586 | Paralichthys olivaceus | Japanese flounder | IL-34 |
| XM_019360775 | Oreochromis niloticus | Nile tilapia | IL-34 |
| XM_010743578 | Larimichthys crocea | Large yellow croaker | IL-34 |
| XM_014124241 | Salmo salar | Atlantic salmon | IL-34 |
| XM_017465109 | Ictalurus punctatus | Channel catfish | IL-34 |
| XM_019087026 | Cyprinus carpio | Common carp | IL-34 |
| XM_011476357 | Oryzias latipes | Japanese rice fish | IL-34 |
| XM_027161223 | Tachysurus fulvidraco | Yellow catfish | IL-34 |
| XM_013138989 | Esox lucius | Northern pike | IL-34 |
| XM_026287434 | Carassius auratus | Goldfish | IL-34 |
| XM_007249177 | Astyanax mexicanus | Mexican tetra | IL-34 |
| MK297321 | Ctenopharyngodon idella | Grass carp | IL-34 |
| XM_020456685 | Oncorhynchus kisutch | Coho salmon | IL-34 |
| XM_024802984 | Maylandia zebra | Zebra mbuna | IL-34 |
| NM_152456 | Homo sapiens | Human | IL-34 |
| NM_001135100 | Mus musculus | Mouse | IL-34 |
| NM_001025766 | Rattus norvegicus | Rat | IL-34 |
| NM_001285975 | Sus scrofa | Pig | IL-34 |
| XM_022419217 | Canis lupus familiaris | Dog | IL-34 |
| XM_023637306 | Equus caballus | Horse | IL-34 |
| XM_018260639 | Xenopus laevis | African clawed frog | IL-34 |
| XM_003641892 | Gallus gallus | Chicken | IL-34 |
| XM_019535589 | Crocodylus porosus | Crocodile | IL-34 |
| NM_001100324 | Bos taurus | Cattle | IL-34 |
| NM_001114480 | Danio rerio | Zebrafish | CSF-1 |
| NM_001080076 | Danio rerio | Zebrafish | CSF-1-2 |
| NM_001124394 | Oncorhynchus mykiss | Rainbow trout | CSF-1 |
| NM_001160476 | Oncorhynchus mykiss | Rainbow trout | CSF-1-2 |
| KM350156 | Epinephelus coioides | Orange-spotted grouper | CSF-1-2 |
| NM_001280600 | Xenopus laevis | African clawed frog | CSF-1 |
| XM_017000369 | Homo sapiens | Human | CSF-1 |
| XM_024984551 | Bos taurus | Cattle | CSF-1 |
| XM_008761428 | Rattus norvegicus | Rat | CSF-1 |
| NM_001113530 | Mus musculus | Mouse | CSF-1 |
| NM_001193295 | Gallus gallus | Chicken | CSF-1 |
In vivo bacterial challenge and tissue collection
In vivo E. tarda challenge was performed on the mudskippers as described previously (Guan et al., 2017). The E. tarda strain MCCC 235 (purchased from the Marine Culture Collection of China) was cultured in Tryptic Soy Broth (TSB) medium at 28 °C with shaking and collected in the logarithmic growth phase. The fish were then intraperitoneally injected with 1.0×104 colony forming units (CFUs) of live E. tarda per fish, with the control group treated with the same volume of phosphate-buffered saline (PBS). At 4, 8, 12, and 24 h post-injection (hpi), the liver, spleen, kidney, and brain were collected. The tissues of healthy fish without treatment, including the kidney, muscle, liver, skin, spleen, gill, brain, and intestine, were also collected. Prior to tissue dissection, fish were anesthetized with 0.03% (v/v) ethylene glycol monophenyl ether and sacrificed.
Quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from fish tissues and MOs/MΦs using RNAiso reagent (TaKaRa, Dalian, China). After treatment with DNase I (TaKaRa, Dalian, China), first-strand cDNA was synthesized using AMV reverse transcriptase (TaKaRa, Dalian, China). qRT-PCR was performed on an ABI StepOne Real-Time PCR System (Applied Biosystems, Foster City, USA) using SYBR premix Ex Taq II (TaKaRa, Dalian, China) as described previously (Chen et al., 2019). The primers used are listed in Table 2. The reaction mixture was incubated for 5 min at 94 °C, and then subjected to 40 amplification cycles of 30 s at 94 °C, 30 s at 60 °C, and 30 s at 72 °C, followed by melting curve analysis for 30 s at 94 °C, 30 s at 72 °C, and 30 s at 94 °C. Relative gene expression of BpIL-34 in healthy or E. tarda-infected samples was calculated using the 2−ΔCT and 2−ΔΔCT methods, respectively, and the data were normalized to Bp18S rRNA levels. Each PCR run was performed with four samples and repeated three times.
2. Oligonucleotide primers used in the qRT-PCR analysis of mudskipper genes.
| Gene | GenBank accession No. | Primer | Nucleotide sequence (5′–3′) |
| BpIL-34 | XM_020935293 | BpIL-34RF | GCAGGAGCTTCCAGAGTCAG |
| BpIL-34RR | CCTCCAATGGGACCTGTCAC | ||
| BpIL-1β | KX492895 | BpIL-1βF | ACGAGTGGTGAATGTGGTCA |
| BpIL-1βR | GAACTGAGGTTGTGCTGCAA | ||
| BpTNF-α | KX492896 | BpTNF-αF | GGACAACAACGAGATCGTGA |
| BpTNF-αR | GTTCCACCGTGTGACTGATG | ||
| BpIL-6 | XM_020932674 | BpIL-6F | GCAGCACGTCAGAAGATGAGA |
| BpIL-6R | TCTCTGAGAAACTCGTGCAGC | ||
| BpTGF-β | XM_020928521 | BpTGF-βF | TCAAAGGACACTTGCACAGC |
| BpTGF-βR | CAGGGCCAAGATCTGTGAAT | ||
| BpIL-10 | XM_020936977 | BpIL-10F | GTGGAGGGGTTCCCTCTAAG |
| BpIL-10R | GTGCGGAGGTAAAAGCTCAG | ||
| Bp18S rRNA | KX492897 | Bp18SF | GGCCGTTCTTAGTTGGTGGA |
| Bp18SR | CCCGGACATCTAAGGGCATC | ||
| BpCSF-1R1 | XM_020921963 | BpCSF-1R1F | GGCCCACGTGTAAGGAGAAT |
| BpCSF-1R1R | TACTCCTCCCTCTGCACCTC | ||
| BpCSF-1R2 | XM_020941064 | BpCSF-1R2F | GTCTTGAGGTTGGACTCGGG |
| BpCSF-1R2R | CTCACACCTGTCGGTGAGTC |
Prokaryotic expression and purification of recombinant mature BpIL-34 (rBpIL-34)
The primers BpIL-34pF (5′-CG GAATTCGCCCCCACTCCTTCGAGC-3′, underline indicates an introduced EcoR I site) and BpIL-34pR (5′-CCG CTCGAGTCAGCTTTTTGTGTTCACATTCT-3′, underline indicates an introduced Xho I site) were designed to amplify the sequence encoding the mature BpIL-34 (mBpIL-34) peptide. After digestion with EcoR I and Xho I, the amplicon was cloned into the pET-28a vector and the constructed plasmid (pET28a-mBpIL-34) was subsequently transformed into Escherichia coli BL21 (DE3). Here, rBpIL-34 was overexpressed by the induction of isopropyl-β-D-thiogalactopyranoside (IPTG) and subsequently purified using a Ni-NTA column (TaKaRa, Dalian, China) according to the manufacturer’s instructions. Endotoxin in the recombinant proteins was detected using the Limulus amebocyte lysate test and was found to be less than 0.1 EU/mg after toxin removal with an endotoxin-removal column (Pierce, Rockford, USA).
Antibody preparation and Western blot assays
Antibody production was performed as reported previously (Chen et al., 2019). Briefly, purified rBpIL-34 was used as an immunogen to produce antiserum in Institute of Cancer Research (ICR) mice. Protein A agarose beads (Invitrogen, Shanghai, China) were used to precipitate IgG (anti-rBpIL-34 IgG) from the antisera. The IgG from saline-injected ICR mice (isotype IgG) was also purified.
Western blotting was performed to detect native rBpIL-34 in mudskipper MOs/MФs using anti-rBpIL-34 IgG as per previous research (Chen et al., 2019). Briefly, protein samples were resolved by SDS-PAGE and transferred to a polyvinylidene fluoride membrane. Anti-rBpIL-34 IgG was used as the primary antibody at a 1:1 000 dilution, followed by horseradish peroxidase (HRP)-labeled goat anti-mouse IgG (1:5 000) as the secondary antibody. Proteins were visualized using an enhanced chemiluminescence (ECL) kit from Amersham (GE Healthcare, Pittsburgh, USA). To determine whether native BpIL-34 is N-glycosylated, denatured proteins of mudskipper MOs/MФs were treated with PNGase F (New England Biolabs, Beverly, USA) at 37 °C overnight and analyzed by Western blotting.
Primary culture of mudskipper kidney-derived MOs/MФs
Mudskipper kidney-derived MOs/MФs were isolated and cultured as described previously (Ding et al., 2019). Briefly, fish kidney leukocyte-enriched fractions were obtained using Ficoll-Hypaque PREMIUM (1.077 g/mL; GE Healthcare, USA). After washing twice with RPMI1640, the MOs/MФs were cultured in 35 mm dishes at a concentration of 2×107 cells/mL. Cells (2 mL) were then incubated for at least 12 h at 24 °C with 5% CO2. Non-adherent cells were washed off, and adherent cells were incubated in complete medium (RPMI 1640, 5% mudskipper serum, 5% fetal bovine serum (FBS), 100 U/mL penicillin, 100 μg/mL streptomycin) at 24 °C with 5% CO2. The purity of the isolated mudskipper MOs/MФs was greater than 95%, as measured by Wright-Giemsa staining.
RNA interference (RNAi)
Target gene siRNAs, BpCSF-1R1 siRNA (BpCSF-1R1si, 5′-GCCAACGCCUCAAUCACAUTT-3′) and BpCSF-1R2 siRNA (BpCSF-1R2si, 5′-GCAGCAAUGAGAAUCACUUTT-3′), as well as mismatched control siRNA (MsiRNA, 5′-UUCUCCGAACGUGUCACGUTT-3′), were designed and synthesized by Invitrogen. The transfection of cells with siRNA was performed using Lipofectamine RNAiMAX (Invitrogen) according to the manufacturer’s protocols. Briefly, 5 μL of lipofectamine RNAiMAX in 250 μL of Opti-MEM (Invitrogen) was mixed with 100 pmoL siRNA in 250 μL of Opti-MEM. The mixture was then incubated for 20 min at room temperature before adding it to MOs/MФs at a final siRNA concentration of 40 nmol/L. Media were changed to complete media after 5.5-h incubation, and cells were cultured for another 48 h before collection for expression analysis. qRT-PCR confirmed the knockdown of BpCSF-1R1 and BpCSF-1R2 mRNA. The primers used are listed in Table 2.
Phagocytosis assay
Mudskipper MO/MФ phagocytosis assays using E. tarda were performed as described previously (Chen et al., 2016a). Briefly, E. tarda were collected in the logarithmic growth phase, labeled with fluorescein isothiocyanate (FITC) (Sigma, Saint Louis, USA), and designated FITC-E. tarda. The MOs/MФs were cultured in 6-well plates at a concentration of 2×106 cells/mL and each well contained a 2-mL cell suspension. The MOs/MФs were treated with 10.0 μg/mL rBpIL-34 or the same volume of PBS for 12 h. The heat-killed FITC-E. tarda were then added at a multiplicity of infection (MOI) of 10 and incubated for another 30 min. After washing extensively with sterile PBS to remove extracellular particles from the cells, Trypan Blue (0.4%) was used to quench fluorescence outside the cell. The engulfed bacteria were examined by flow cytometry using a Gallios Flow Cytometer (Beckman Coulter, Miami, USA). The relative mean fluorescence intensity (MFI) of bacteria engulfed by the cells was analyzed using FlowJo software. The MFIs of the PBS- and rBpIL-34-treated groups were expressed as fold-change relative to that without bacteria, and the MFI of the PBS-treated group was assigned a value of 100. Another set of assays were conducted with the addition of the siRNA reagents. After transfection with BpCSF-1R1si, BpCSF-1R2si, or MsiRNA for 48 h, MOs/MФs were treated with 10.0 μg/mL rBpIL-34 for 12 h. The remaining steps were performed as described above. The MFIs of the rBpIL-34- and siRNA-treated groups were expressed as fold-change relative to that without bacteria, and the MFI of the rBpIL-34-treated group was assigned a value of 100.
Bacterial killing assay
A bacterial killing assay was performed as described previously (Chen et al., 2018). Briefly, 4×106 MOs/MФs were treated with 10.0 μg/mL rBpIL-34 or the same volume of PBS for 12 h. Live FITC-E. tarda were added at a MOI of 10 and incubated for another 30 min. Bacterial uptake by MOs/MΦs was allowed to occur for 30 min at 24 °C in an atmosphere with 5% CO2. Non-internalized E. tarda were removed by extensive washing with sterile PBS. One set of samples (uptake group) was collected for RNA extraction, whereas the other set of samples (kill group) was further incubated for 1.5 h to facilitate bacterial killing before cell lysis. Cells were subjected to classic RNA isolation and subsequent qRT-PCR assays for E. tarda using the primers gyrBF and gyrBR (Guan et al., 2017). The Ct values, based on standard curves generated previously, were used to calculate the total CFUs/mL in all samples. Bacterial survival was determined by dividing the number of CFUs in the kill group by that in the uptake group. Four independent experiments were carried out. In addition, another set of assays were conducted with the addition of siRNA reagents. After transfection with BpCSF-1R1si, BpCSF-1R2si, or MsiRNA for 48 h, MOs/MФs were treated with 10.0 μg/mL rBpIL-34 for 12 h. The remaining steps were performed as described above.
Cytokine mRNA expression analysis in MOs/MФs
After transfection with BpCSF-1R1si, BpCSF-1R2si, and MsiRNA, MOs/MФs were treated with 10.0 μg/mL rBpIL-34 for 12 h; MOs/MФs without any treatment were used as a blank control. qRT-PCR was carried out as described in Section 2.4 with target gene-specific primers (Table 2). The mRNA expression levels of BpTNF-α, BpIL-1β, BpIL-6, BpTGF-β, and BpIL-10 were normalized to those of Bp18S rRNA. Four independent experiments were performed.
Statistical analysis
All data are described as means±standard error of the mean (SEM). Data were analyzed by one-way analysis of variance (ANOVA) or Student’s t-test using SPSS v13.0 (SPSS Inc, Chicago, IL, USA). A P-value of 0.05 was considered statistically significant.
RESULTS
Sequence identification of BpIL-34
The cDNA sequence of BpIL-34 was obtained from the DDBJ/EMBL/GenBank databases under accession No. XM_020935293. The sequence was 1 169 nucleotides (nt) in length and possessed an open reading frame (ORF) of 642 nt, which was predicted to encode a 213-amino acid (aa) polypeptide with a calculated MW of 25.0 kDa and an isoelectric point (pI) of 8.57. Sequence analyses revealed that BpIL-34 possessed a 24-aa N-terminal signal peptide, and the mature peptide was comprised of 189 aa, with a calculated MW of 22.4 kDa. Multiple alignment showed that BpIL-34 contained five highly conserved cysteine residues (Cys32, Cys177, Cys182, Cys187, and Cys197), four of which formed two disulfide bonds (Cys32-Cys187 and Cys182-Cys197) (Figure 1). Two potential N-glycosylation sites (Asn39 and Asn71) were found in BpIL-34 (Figure 1). BpIL-34 was also found to contain a conserved motif with multiple cationic amino acid residues (RKx [R/K] K) at aa position 203–207, similar to that in other fish homologs (Liu et al., 2012) (Figure 1).
1. Multiple alignment of amino acid sequences of BpIL-34 and related fish IL-34 sequences.

Threshold for shading was >60%; similar residues are marked with a gray shadow, identical residues with a black shadow, and alignment gaps with “-”. BpIL-34: Mudskipper IL-34; DrIL-34: Zebrafish IL-34; OmIL-34: Rainbow trout IL-34; SaIL-34: Gilthead seabream IL-34; TrIL-34: Tiger puffer IL-34; SsIL-34: Atlantic salmon IL-34; IpIL-34: Channel catfish IL-34; CcIL-34: Common carp IL-34; OlIL-34: Japanese rice fish IL-34; EcIL-34: Orange-spotted grouper IL-34. Predicted cleavage site for signal peptide is marked as “↓”. Five conserved cysteine residues are marked as “*”. Two cysteine residues joined by solid line represent disulfide bond. Possible N-linked glycosylation sites of BpIL-34 are indicated by “▼”. RKx [R/K] K motif is double underlined. GenBank accession Nos. of sequences used are presented in Table 1.
Sequence comparisons showed that BpIL-34 shared the highest aa identity (78.4%) with the orange-spotted grouper IL-34. A phylogenetic tree was constructed based on the complete aa sequences of IL-34, with the CSF-1 sequences serving as an outgroup (Figure 2). Results showed that fish IL-34 grouped together to form a cluster distinct from the mammalian IL-34 cluster (Figure 2). The fish and mammalian IL-34 clusters formed a clade that was well separated from the CSF-1 clade (Figure 2).
2. Phylogenetic tree analysis of complete amino acid sequences of IL-34 and CSF-1 using neighbor-joining method.

CSF-1 sequences as an outgroup. Percentage of bootstrap values is shown next to branches based on 1 000 bootstrap replications (shown only when >60%). Scale bar represents number of substitutions per base position. Site of mudskipper IL-34 is indicated by “▲”. GenBank accession Nos. of sequences used are listed in Table 1.
Tissue BpIL-34 mRNA expression in response to E. tarda infection
The relative mRNA expression of BpIL-34 in selected tissues was detected by qRT-PCR. In healthy fish, BpIL-34 transcripts were detected in all tested tissues, with the highest level observed in the brain, followed by the gill and intestine (Figure 3A). Upon E. tarda infection, BpIL-34 mRNA expression rapidly increased in the four selected tissues (brain, liver, spleen, and kidney) compared to that in the control group, and the highest level of up-regulation (45.7-fold) was found in the kidney at 12 hpi (Figure 3B–E).
3. qRT-PCR analysis of BpIL-34 mRNA expression in tissues of healthy (A) and Edwardsiella tarda-infected mudskippers (B–E) .

A: BpIL-34 mRNA expression relative to that of Bp18S rRNA was calculated using 2−ΔCT method. Values denoted by different letters are significantly different when compared by ANOVA (P<0.05);n=4. B–E: BpIL-34 mRNA expression relative to that of Bp18S rRNA was calculated using 2−ΔΔCT method. Tissues were collected at 4, 8, 12, and 24 h after bacterial infection. Data are expressed as means±SEM; n=4, *: P<0.05.
Prokaryotic expression and purification of rBpIL-34 and antibody preparation
After induction by IPTG, recombinant rBpIL-34 was overexpressed in E. coli BL21 (DE3). The size of rBpIL-34 obtained based on SDS-PAGE analysis was ~26 kDa, similar to the calculated value (22.4 kDa for the mBpIL-34 plus 3.83 kDa for 6×His-tag; Figure 4A). rBpIL-34 was purified by affinity chromatography using an Ni-NTA column, with purity not less than 96% (data not shown). Purified rBpIL-34 was then used to immunize mice, and anti-rBPIL-34 IgG was purified from antisera. As determined by Western blot analysis, the MW of native BpIL-34 in mudskipper MOs/MΦs was approximately 32 kDa, which was converted to a 25-kDa band after PNGase F digestion (Figure 4B).
4. Prokaryotic expression and Western blot analysis of BpIL-34.
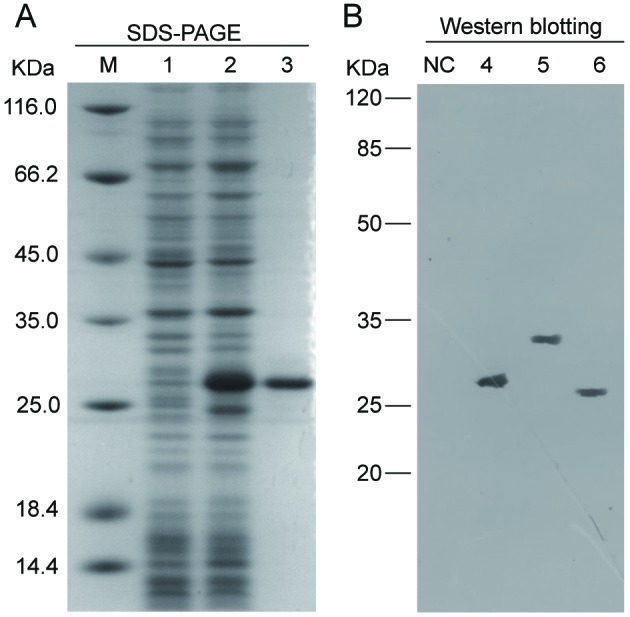
A: SDS-PAGE analysis of prokaryotically expressed rBpIL-34. Lane M: Protein marker; 1, 2: Crude protein extracts from BL21 (DE3) transformed with pET-28a-BpIL-34 plasmid before and after IPTG induction, respectively; 3: Purified rBpIL-34. B: Western blot analysis of BpIL-34 using anti-rBpIL-34 IgG. NC: Negative control; 4: Purified rBpIL-34; 5: Proteins extracted from mudskipper MOs/MФs; 6: Proteins extracted from mudskipper MOs/MФs with PNGase F digestion.
Effect of rBpIL-34 on phagocytotic and bacterial killing activity of MOs/MФs
MOs/MФs play a pivotal role in immune responses in fish against invasion by pathogens, and phagocytosis and intracellular bacterial killing are the main biological functions of MOs/MФs (Lu & Chen, 2019). Consequently, we investigated the effects of rBpIL-34 on the functions of mudskipper MOs/MФs. Phagocytosis of FITC-E. tarda by rBpIL-34-treated MOs/MΦs was significantly higher (up to 3.18-fold) than that in the PBS-treated group (Figure 5A). In addition, qRT-PCR measurement of intracellular E. tarda CFUs in mudskipper MOs/MΦs showed that the bacterial survival rate in the rBpIL-34-treated group (36.12%±1.54%) was much lower than that in the PBS-treated group (75.36%±3.97%; Figure 5B).
5. Effects of rBpIL-34 on phagocytosis and bacterial killing of Edwardsiella tarda by mudskipper MO/MΦs .

A: Effect of rBpIL-34 on phagocytosis of E. tarda by MOs/MΦs. Mudskipper MOs/MΦs were pre-treated with PBS or rBpIL-34 before adding FITC-E. tarda (MOI=10). After an additional 30 min incubation, phagocytosis of FITC-E. tarda was determined by flow cytometry. MFI is presented as fold-change over the value for PBS-treated group, which was assigned a unit of 100. B: Effect of rBpIL-34 on bacterial killing of E. tarda by MOs/MΦs. MOs/MΦs were infected with live E. tarda after treatment with PBS or rBpIL-34, and viability of E. tarda (MOI=10) was determined by qRT-PCR. Killing of E. tarda by mudskipper MOs/MФs was measured using CFU assay based on a standard curve. Data are expressed as means±SEM; n=4, *: P<0.05.
Effect of rBpIL-34 on cytokine mRNA expression in MOs/MФs
We also investigated the effects of rBpIL-34 on mRNA expression of typical inflammatory cytokines (pro-inflammatory cytokines: BpTNF-α, BpIL-1β, and BpIL-6; anti-inflammatory cytokines: BpTGF-β and BpIL-10) in mudskipper MOs/MΦs. qRT-PCR showed that the mRNA expression levels of BpTNF-α, BpIL-1β, and BpIL-6 were up-regulated 7.36-, 3.01-, and 1.42-fold, respectively, in MOs/MΦs treated with rBpIL-34 compared to those in the control group (Figure 6). No significant change was found in the mRNA expression of BpTGF-β between the rBpIL-34-treated and control groups; however, BpIL-10 mRNA expression was down-regulated 0.67-fold in the rBpIL-34-treated group compared to that in the control group (Figure 6).
6. Effects of rBpIL-34 on mRNA expression of BpTNF-α (A), BpIL-1β (B), BpIL-6 (C), BpTGF-β (D), and BpIL-10 (E) in mudskipper MOs/MΦs .

MOs/MΦs were treated with rBpIL-34 for 12 h. PBS-treated group was used as the control. mRNA expression levels of selected cytokines were normalized to those of Bp18S rRNA. Data are expressed as means±SEM; n=4, *: P<0.05.
Effect of BpCSF-1R1 and BpCSF-1R2 knockdown on rBpIL-34-enhanced phagocytotic and bacterial killing activity of MOs/MΦs
As CSF-1R is reportedly the receptor of IL-34 in mammals (Ségaliny et al., 2015; Wang et al., 2012) and mudskippers have two CSF-1R genes, we further determined whether BpCSF-1R1 or BpCSF-1R2 knockdown would influence the rBpIL-34-enhanced phagocytotic and bacterial killing activity of mudskipper MOs/MΦs. Firstly, we used RNAi to knock down the expression of BpCSF-1R1 and BpCSF-1R2 in MOs/MФs. When MOs/MФs were transfected with BpCSF-1R1si, the mRNA expression of BpCSF-1R1 decreased to 29.17%±6.31% of control levels at 48 h, whereas BpCSF-1R2 expression showed no significant change (Figure 7B, C). Similarly, when MOs/MФs were transfected with BpCSF-1R2si, the mRNA expression of BpCSF-1R2 decreased to 21.66%±1.11% of control levels at 48 h, whereas BpCSF-1R1 expression showed no significant change (Figure 7B, C). These results suggest that the target genes BpCSF-1R1 and BpCSF-1R2 were effectively and specifically knocked down. The transfection of MsiRNA had no obvious effect on either BpCSF-1R1 or BpCSF-1R2 expression (Figure 7B, C).
7. RNAi-mediated gene knockdown of BpCSF-1R1 and BpCSF-1R2 in mudskipper MOs/MФs .

A: qRT-PCR analysis of mRNA expression of BpCSF-1R1 and BpCSF-1R2 in primary MOs/MФs. B: qRT-PCR analysis of BpCSF-1R1 transcripts in BpCSF-1R1si- or BpCSF-1R2si-transfected MOs/MФs. C: qRT-PCR analysis of BpCSF-1R2 transcripts in BpCSF-1R1si- or BpCSF-1R2si-transfected MOs/MФs. MsiRNA-transfected MOs/MФs group was used as the control. mRNA levels of BpCSF-1R1 and BpCSF-1R2 were normalized to those of Bp18S rRNA. Data are expressed as means±SEM; n=4, *: P<0.05.
We then used BpCSF-1R1si and BpCSF-1R2si to explore whether BpCSF-1R1 and BpCSF-1R2 mediated the enhancing effect of rBPIL-34 on the phagocytotic and bacterial killing activity of MOs/MФs. After BpCSF-1R1si transfection, MOs/MΦs treated with rBpIL-34 showed a significant decrease in phagocytosis of E. tarda (0.16-fold) compared to that in the control group (normal MO/MΦs treated with rBpIL-34), whereas MsiRNA or BpCSF-1R2si transfection resulted in no obvious changes (Figure 8A). On the other hand, the bacterial survival rate in BpCSF-1R1-knockdown MOs/MΦs after rBPIL-34 treatment was significantly increased (2.15-fold) compared to that in the control group (normal MO/MΦ treated with rBpIL-34), with MsiRNA or BpCSF-1R2si transfection resulting in no obvious changes (Figure 8B).
8. Effects of BpCSF-1R1 and BpCSF-1R2 knockdown on rBpIL-34-enhanced phagocytosis and bacterial killing of Edwardsiella tarda by mudskipper MOs/MΦs .

A: Effect of BpCSF-1R1 or BpCSF-1R2 knockdown on rBpIL-34-enhanced phagocytosis of E. tarda by mudskipper MOs/MΦs. After transfection with BpCSF-1R1si, BpCSF-1R2si, or MsiRNA (control) for 48 h, mudskipper MOs/MФs were treated with rBpIL-34 for 12 h. Normal MOs/MΦs treated with rBpIL-34 (rBpIL-34 group) were used as controls. Thereafter, FITC-E. tarda were added at a MOI of 10 and incubated for an additional 30 min. Phagocytosis of FITC-E. tarda was determined by flow cytometry. Mean fluorescence intensity (MFI) is presented as a fold-change over value for rBpIL-34 group, which was assigned a value of 100. B: Effect of BpCSF-1R1 and BpCSF-1R2 knockdown on rBpIL-34-enhanced bacterial killing of E. tarda by mudskipper MOs/MΦs. After transfection with BpCSF-1R1si, BpCSF-1R2si, or MsiRNA for 48 h, mudskipper MOs/MФs were treated with rBpIL-34 for 12 h. Normal MOs/MΦs treated with rBpIL-34 (rBpIL-34 group) were used as controls. Live E. tarda were added at a MOI of 10 and incubated for an additional 30 min. Killing of E. tarda by mudskipper MOs/MФs was measured using a CFU assay based on standard curve. Data are expressed as means±SEM; n=4, *: P<0.05.
Effect of BpCSF-1R1 and BpCSF-1R2 knockdown on rBpIL-34-altered cytokine mRNA expression in MOs/MФs
qRT-PCR analysis showed that BpCSF-1R1 knockdown decreased the mRNA expression of BpTNF-α, BpIL-1β, and BpIL-6 in mudskipper MOs/MΦs treated with rBpIL-34 by approximately 0.31-, 0.36-, and 0.49-fold, respectively, compared to levels in the MsiRNA-treated control group (Figure 9), whereas BpTGF-β and BpIL-10 mRNA expression was not obviously changed (Figure 9). BpCSF-1R2 knockdown had no noticeable effect on the mRNA levels of all selected cytokines in MOs/MФs compared to those in the control group (Figure 9).
9. Effects of BpCSF-1R1 and BpCSF-1R2 knockdown on rBpIL-34-altered mRNA expression of selected cytokines in MOs/MФs .

Mudskipper MOs/MФs were pre-treated with BpCSF-1R1si, BpCSF-1R2si, or MsiRNA (control) and further incubated with rBpIL-34 for 12 h. MsiRNA-treated group was used as the control. mRNA levels of BpTNF-α (A), BpIL-1β (B), BpIL-6 (C), BpTGF-β (D), and BpIL-10 (E) were normalized to those of Bp18S rRNA. Data are expressed as means±SEM; n=4, *: P<0.05.
DISCUSSION
IL-34 is a recently discovered cytokine identified as the second ligand of CSF-1R (Lin et al., 2008). Over the past decade, accumulating evidence has shown that IL-34 exerts potent immunomodulatory effects in many physiological and pathological states. However, studies on the function of IL-34 in teleosts are limited. In this study, we identified an IL-34 homolog sequence from mudskippers. Sequence analysis revealed that BpIL-34 had structural similarities to other fish IL-34 homologs. Phylogenetic tree analysis showed that BpIL-34 belonged to the IL-34 clade and was most closely related to an orange-spotted grouper homolog. It has been reported that the C-terminal region of IL-34 is heavily glycosylated via O-linked glycans in humans (Felix et al., 2013), but this region is not found in fish IL-34 homologs (Xue et al., 2019). In contrast, two to six potential N-glycosylation sites are present in each fish IL-34 homolog (Wang et al., 2013). Our results showed that the native BpIL-34 in cells was N-glycosylated. Glycosylation plays an important role in functions such as intracellular transport, ligand binding, structural stability, and signal transduction (Huang et al., 2015). Recent study has shown that this post-translational glycosylation does not appear to influence the functions of recombinant grass carp IL-34 as the effects of both bacteria-derived rCiIL-34 and HEK293T cell-derived rCiIL-34 on fish MOs/MΦs are similar (Xue et al., 2019). Signatures of positive selection are also found in mammalian IL-34 (Neves et al., 2014), which might be explained by the multitude of biological processes involving IL-34.
Gene expression patterns commonly reflect functions. IL-34 expression is mainly detected in the skin and central nervous system in mice (Wang et al., 2012). In fish, like rainbow trout, large yellow croaker, and grass carp, IL-34 is constitutively expressed in various tissues, with highest expression found in the spleen (Wang et al., 2013, 2018; Xue et al., 2019), or the brain in the case of orange-spotted grouper (Mo et al., 2015). IL-34 expression is tightly associated with pathogen stimulation. Most studies have shown that pathogen infection increases IL-34 transcript levels in tissues, especially at the infected sites (Mo et al., 2015; Wang et al., 2013, 2018; Wu et al., 2019; Xue et al., 2019). However, a significant decrease in the expression of IL-34 in hepatitis B virus (HBV) patients has also been identified (Cheng et al., 2017). Changes in IL-34 expression, either increase or decrease, are involved in disease pathogenesis, and are correlated with progression, severity, and chronicity (Baghdadi et al., 2018). In this study, we determined that BpIL-34 transcripts were constitutively expressed in all tested tissues, with the highest level found in the brain. This result is in accordance with that reported in the orange-spotted grouper (Mo et al., 2015) and in mammals (Wang et al., 2012). IL-34 is a tissue-restricted ligand of CSF-1R and is required for the development of Langerhans cells and microglia in mammals (Wang et al., 2012); moreover, it can protect blood-brain barrier integrity by restoring expression levels of tight junction proteins (Jin et al., 2014). Fish IL-34 may also exhibit such functions. Upon E. tarda infection, the BpIL-34 mRNA level was significantly up-regulated in the liver, spleen, kidney, and brain, in accordance with that reported in fish previously (Mo et al., 2015; Wang et al., 2013; Wu et al., 2019; Xue et al., 2019), suggesting that BpIL-34 may be involved in mudskipper immune responses against E. tarda.
MO/MΦ plays key roles in the fish protection system, and phagocytosis, the killing of invading bacteria, and production of cytokines are their main effector functions (Lu & Chen, 2019; Zou & Secombes, 2016). IL-34 was first identified as a potent activator of MO and MΦ via CSF-1R based on functional screening of the extracellular proteome (Lin et al., 2008). In the African clawed frog (Xenopus laevis), recombinant IL-34-derived MΦs not only display higher bactericidal activity than control cells but also exhibit potent antiviral activity against FV3 ranavirus, which is dependent on reactive oxygen production (Grayfer & Robert, 2014, 2015). Intriguingly, recombinant CSF-1-treated rather than recombinant IL-34-treated MΦs are more phagocytic for both tadpoles and adult frogs (Grayfer & Robert, 2014, 2015). IL-34 also activates microglia (tissue-resident MΦs of the central nervous system) to rescue neurons by enhancing the phagocytosis of toxicants or damaged debris (Suzumura, 2013; Wang & Colonna, 2014). In this study, we showed that rBpIL-34-derived mudskipper MOs/MΦs displayed increased phagocytic and bactericidal activities compared to those of the PBS-treated control group, suggesting that rBpIL-34 could activate these cells and possibly drive their differentiation. Cytokines are signaling proteins that regulate a wide range of biological functions, and macrophages are a major source of many cytokines. Studies have shown that IL-34 can alter cytokine expression in vertebrate species. For example, IL-34 treatment can induce the up-regulation of IL-6, CXCL10, IL-8, and CCL2 in human whole blood (Eda et al., 2010) and increased TNF-α expression in lamina propria mononuclear cells (Franzè et al., 2015). Furthermore, the addition of IL-34 to primary lung fibroblasts can significantly promote IL-6 and IL-8 expression in a dose- and time-dependent manner (Zhou et al., 2018). Treatment with chicken IL-34 increases the expression of Th1 and Th17 cytokines in chicken cell lines (Truong et al., 2018). In grass carp, in vitro IL-34 treatment up-regulates the expression of IL-1β, IL-6, and IL-8 but inhibits the expression of IL-10 and TGF-β1 in MΦs (Xue et al., 2019). Here, we determined that rBpIL-34-derived MOs/MΦs expressed higher levels of BpTNF-α, BpIL-1β, and BpIL-6 and lower levels of BpIL-10 compared to that in the PBS-treated control group, coinciding well with previous reports (Eda et al., 2010; Franzè et al., 2015; Truong et al., 2018; Xue et al., 2019; Zhou et al., 2018). These results suggest that IL-34 promotes the differentiation of mudskipper MO/MΦ into the pro-inflammatory phenotype.
CSF-1R is critical for the proliferation, survival, and differentiation of MФs, as its knockdown results in the marked depletion of MФs in most tissues (Dai et al, 2002; Droin & Solary, 2010). In addition, CSF-1R signaling controls development of the MФ lineage under steady conditions and during certain inflammatory reactions (Lenzo et al, 2012). CSF-1R is a major receptor of IL-34 in mammals (Baghdadi et al., 2018; Jeannin et al., 2018; Peyraud et al., 2017). For example, in human whole blood, the IL-34-enhanced mRNA expression levels of IL-6 and chemokines such as MCP-1 are inhibited by GW2580, a CSF-1R kinase inhibitor (Eda et al., 2010). In this study, we determined that BpCSF-1R1, but not BpCSF-1R2, mediated the enhancing effects of rBpIL-34 on MO/MΦ phagocytosis, bactericidal activity, and pro-inflammatory cytokine mRNA expression. Interestingly, although rBpIL-34 inhibited the mRNA expression of the anti-inflammatory cytokine BpIL-10 in MOs/MΦs, knockdown of BpCSF-1R1 and BpCSF-1R2 had no significant effect on BpIL-10 mRNA expression in mudskipper MOs/MΦs compared to that in the MsiRNA-treated control group, suggesting that the inhibitory effect of rBPIL-34 on BpIL-10 mRNA expression was independent of BpCSF-1R1 and BpCSF-1R2. To our knowledge, this is the first report to show that CSF-1R1 mediates the functions of IL-34 in fish MOs/MΦs.
In summary, we identified an IL-34 homolog from mudskippers. BpIL-34 mRNA expression was up-regulated in mudskipper tissue upon E. tarda infection. In vitro, rBpIL-34 treatment not only enhanced the phagocytotic and bactericidal activity of mudskipper MOs/MФs but also elevated the mRNA levels of pro-inflammatory cytokines. Results of RNAi suggested that BpCSF-1R1, but not BpCSF-1R2, mediated the effects of rBpIL-34. However, further investigations are needed to determine the underlying intracellular signaling pathways associated with IL-34.
COMPETING INTERESTS
The authors declare that they have no competing interests.
AUTHORS' CONTRIBUTIONS
H.Y.S., Q.J.Z, and J.C. conceived and designed the experiments. H.Y.S., Y.Z., and Q.J.Z performed the experiments. H.Y.S., Q.J.Z, and J.C. analyzed the data and wrote the manuscript. M.Y.L. and J.C. reviewed and corrected the manuscript. All authors read and approved the final version of the manuscript.
Funding Statement
The study was supported by the National Natural Science Foundation of China (31972821; 31772876), the Program of Zhejiang Provincial Natural Science Foundation of China (LZ18C190001), Science and Technology Department of Zhejiang Province (LGN18C180002), Natural Science Foundation of Ningbo City (2018A610342), and the K.C. Wong Magna Fund in Ningbo University
Contributor Information
Qian-Jin Zhou, Email: mumu2325@163.com.
Jiong Chen, Email: jchen1975@163.com.
References
- 1.Baghdadi M, Umeyama Y, Hama N, Kobayashi T, Han N, Wada H, Seino KI Interleukin-34, a comprehensive review. Journal of Leukocyte Biology. 2018;104(5):931–951. doi: 10.1002/JLB.MR1117-457R. [DOI] [PubMed] [Google Scholar]
- 2.Chen J, Chen Q, Lu XJ, Chen J The protection effect of LEAP-2 on the mudskipper (Boleophthalmus pectinirostris) against Edwardsiella tarda infection is associated with its immunomodulatory activity on monocytes/macrophages . Fish & Shellfish Immunology. 2016a;59:66–76. doi: 10.1016/j.fsi.2016.10.028. [DOI] [PubMed] [Google Scholar]
- 3.Chen J, Nie L, Chen J Mudskipper (Boleophthalmus pectinirostris) hepcidin-1 and hepcidin-2 present different gene expression profile and antibacterial activity and possess distinct protective effect against Edwardsiella tarda infection . Probiotics and Antimicrobial Proteins. 2018;10(2):176–185. doi: 10.1007/s12602-017-9352-0. [DOI] [PubMed] [Google Scholar]
- 4.Chen K, Shi YH, Chen J, Li MY A soluble FcγR homolog inhibits IgM antibody production in ayu spleen cells. Zoological Research. 2019;40(5):404–415. doi: 10.24272/j.issn.2095-8137.2019.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chen Q, Lu XJ, Chen J Identification and functional characterization of the CSF1R gene from grass carp Ctenopharyngodon idellus and its use as a marker of monocytes/macrophages . Fish & Shellfish Immunology. 2015;45(2):386–398. doi: 10.1016/j.fsi.2015.04.029. [DOI] [PubMed] [Google Scholar]
- 6.Chen Q, Lu XJ, Li MY, Chen J Molecular cloning, pathologically-correlated expression and functional characterization of the colonystimulating factor 1 receptor (CSF-1R) gene from a teleost, Plecoglossus altivelis . Zoological Research. 2016b;37(2):96–102. doi: 10.13918/j.issn.2095-8137.2016.2.96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cheng ST, Tang H, Ren JH, Chen X, Huang AL, Chen J Interleukin-34 inhibits hepatitis B virus replication in vitro and in vivo . PLoS One. 2017;12(6):e0179605. doi: 10.1371/journal.pone.0179605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chihara T, Suzu S, Hassan R, Chutiwitoonchai N, Hiyoshi M, Motoyoshi K, Kimura F, Okada S IL-34 and M-CSF share the receptor Fms but are not identical in biological activity and signal activation. Cell Death & Differentiation. 2010;17(12):1917–1927. doi: 10.1038/cdd.2010.60. [DOI] [PubMed] [Google Scholar]
- 9.Dai XM, Ryan GR, Hapel AJ, Dominguez MG, Russell RG, Kapp S, Sylvestre V, Stanley ER Targeted disruption of the mouse colony-stimulating factor 1 receptor gene results in osteopetrosis, mononuclear phagocyte deficiency, increased primitive progenitor cell frequencies, and reproductive defects. Blood. 2002;99(1):111–120. doi: 10.1182/blood.V99.1.111. [DOI] [PubMed] [Google Scholar]
- 10.Dan XM, Zhong ZP, Li YW, Luo XC, Li AX Cloning and expression analysis of grouper (Epinephelus coioides) M-CSFR gene post Cryptocaryon irritans infection and distribution of M-CSFR+ cells . Fish & Shellfish Immunology. 2013;35(2):240–248. doi: 10.1016/j.fsi.2013.04.019. [DOI] [PubMed] [Google Scholar]
- 11.Ding FF, Li CH, Chen J Molecular characterization of the NK-lysin in a teleost fish, Boleophthalmus pectinirostris: Antimicrobial activity and immunomodulatory activity on monocytes/macrophages . Fish & Shellfish Immunology. 2019;92:256–264. doi: 10.1016/j.fsi.2019.06.021. [DOI] [PubMed] [Google Scholar]
- 12.Droin N, Solary E Editorial: CSF1R, CSF-1, and IL-34, a "ménage à trois" conserved across vertebrates. Journal of Leukocyte Biology. 2010;87(5):745–747. doi: 10.1189/jlb.1209780. [DOI] [PubMed] [Google Scholar]
- 13.Eda H, Zhang J, Keith RH, Michener M, Beidler DR, Monahan JB Macrophage-colony stimulating factor and interleukin-34 induce chemokines in human whole blood. Cytokine. 2010;52(3):215–220. doi: 10.1016/j.cyto.2010.08.005. [DOI] [PubMed] [Google Scholar]
- 14.Felix J, Elegheert J, Gutsche I, Shkumatov AV, Wen YR, Bracke N, Pannecoucke E, Vandenberghe I, Devreese B, Svergun DI, Pauwels E, Vergauwen B, Savvides SN Human IL-34 and CSF-1 establish structurally similar extracellular assemblies with their common hematopoietic receptor. Structure. 2013;21(4):528–539. doi: 10.1016/j.str.2013.01.018. [DOI] [PubMed] [Google Scholar]
- 15.Franzè E, Monteleone I, Cupi ML, Mancia P, Caprioli F, Marafini I, Colantoni A, Ortenzi A, Laudisi F, Sica G, Sileri P, Pallone F, Monteleone G Interleukin-34 sustains inflammatory pathways in the gut. Clinical Science. 2015;129(3):271–280. doi: 10.1042/CS20150132. [DOI] [PubMed] [Google Scholar]
- 16.Grayfer L, Robert J Divergent antiviral roles of amphibian (Xenopus laevis) macrophages elicited by colony-stimulating factor-1 and interleukin-34 . Journal of Leukocyte Biology. 2014;96(6):1143–1153. doi: 10.1189/jlb.4A0614-295R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Grayfer L, Robert J Distinct functional roles of amphibian (Xenopus laevis) colony-stimulating factor-1- and interleukin-34-derived macrophages . Journal of Leukocyte Biology. 2015;98(4):641–649. doi: 10.1189/jlb.4AB0315-117RR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Greter M, Lelios I, Pelczar P, Hoeffel G, Price J, Leboeuf M, Kündig TM, Frei K, Ginhoux F, Merad M, Becher B Stroma-derived interleukin-34 controls the development and maintenance of langerhans cells and the maintenance of microglia. Immunity. 2012;37(6):1050–1060. doi: 10.1016/j.immuni.2012.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Guan F, Lu XJ, Li CH, Chen J Molecular characterization of mudskipper (Boleophthalmus pectinirostris) hypoxia-inducible factor-1α (HIF-1α) and analysis of its function in monocytes/macrophages . PLoS One. 2017;12(5):e0177960. doi: 10.1371/journal.pone.0177960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Honda T, Nishizawa T, Uenobe M, Kohchi C, Kuroda A, Ototake M, Nakanishi T, Yokomizo Y, Takahashi Y, Inagawa H, Soma GI Molecular cloning and expression analysis of a macrophage-colony stimulating factor receptor-like gene from rainbow trout, Oncorhynchus mykiss . Molecular Immunology. 2005;42(1):1–8. doi: 10.1016/j.molimm.2004.07.002. [DOI] [PubMed] [Google Scholar]
- 21.Huang ZL, Chen S, Liu JC, Xiao J, Yan J, Feng H IFNa of black carp is an antiviral cytokine modified with N-linked glycosylation. Fish & Shellfish Immunology. 2015;46(2):477–485. doi: 10.1016/j.fsi.2015.07.020. [DOI] [PubMed] [Google Scholar]
- 22.Jeannin P, Paolini L, Adam C, Delneste Y The roles of CSFs on the functional polarization of tumor-associated macrophages. The FEBS Journal. 2018;285(4):680–699. doi: 10.1111/febs.14343. [DOI] [PubMed] [Google Scholar]
- 23.Jiang YY, Chen JH, Yen K, Xu J Ectopically expressed IL-34 can efficiently induce macrophage migration to the liver in zebrafish. Zebrafish. 2019;16(2):165–170. doi: 10.1089/zeb.2018.1685. [DOI] [PubMed] [Google Scholar]
- 24.Jin SJ, Sonobe Y, Kawanokuchi J, Horiuchi H, Cheng Y, Wang Y, Mizuno T, Takeuchi H, Suzumura A Interleukin-34 restores blood-brain barrier integrity by upregulating tight junction proteins in endothelial cells. PLoS One. 2014;9(12):e115981. doi: 10.1371/journal.pone.0115981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Katzenback BA, Belosevic M Colony-stimulating factor-1 receptor protein expression is a specific marker for goldfish (Carassius auratus L.) macrophage progenitors and their differentiated cell types . Fish & Shellfish Immunology. 2012;32(3):434–445. doi: 10.1016/j.fsi.2011.12.003. [DOI] [PubMed] [Google Scholar]
- 26.Kim JI, Turka LA Transplant tolerance: a new role for IL-34. Journal of Clinical Investigation. 2015;125(10):3751–3753. doi: 10.1172/JCI84010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lenzo JC, Turner AL, Cook AD, Vlahos R, Anderson GP, Reynolds EC, Hamilton JA Control of macrophage lineage populations by CSF-1 receptor and GM-CSF in homeostasis and inflammation. Immunology & Cell Biology. 2012;90(4):429–440. doi: 10.1038/icb.2011.58. [DOI] [PubMed] [Google Scholar]
- 28.Lin HS, Lee E, Hestir K, Leo C, Huang MM, Bosch E, Halenbeck R, Wu G, Zhou A, Behrens D, Hollenbaugh D, Linnemann T, Qin MM, Wong J, Chu KT, Doberstein SK, Williams LT Discovery of a cytokine and its receptor by functional screening of the extracellular proteome. Science. 2008;320(5877):807–811. doi: 10.1126/science.1154370. [DOI] [PubMed] [Google Scholar]
- 29.Liu H, Leo C, Chen XY, Wong BR, Williams LT, Lin HS, He XL The mechanism of shared but distinct CSF-1R signaling by the non-homologous cytokines IL-34 and CSF-1. Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 2012;1824(7):938–945. doi: 10.1016/j.bbapap.2012.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lu XJ, Chen J Specific function and modulation of teleost monocytes/macrophages: polarization and phagocytosis. Zoological Research. 2019;40(3):146–150. doi: 10.24272/j.issn.2095-8137.2019.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ma XL, Lin WY, Chen YM, Stawicki S, Mukhyala K, Wu Y, Martin F, Bazan JF, Starovasnik MA Structural basis for the dual recognition of helical cytokines IL-34 and CSF-1 by CSF-1R. Structure. 2012;20(4):676–687. doi: 10.1016/j.str.2012.02.010. [DOI] [PubMed] [Google Scholar]
- 32.Masteller EL, Wong BR Targeting IL-34 in chronic inflammation. Drug Discovery Today. 2014;19(8):1212–1216. doi: 10.1016/j.drudis.2014.05.016. [DOI] [PubMed] [Google Scholar]
- 33.Mo ZQ, Li YW, Zhou L, Li AX, Luo XC, Dan XM Grouper (Epinephelus coioides) IL-34/MCSF2 and MCSFR1/MCSFR2 were involved in mononuclear phagocytes activation against Cryptocaryon irritans infection . Fish & Shellfish Immunology. 2015;43(1):142–149. doi: 10.1016/j.fsi.2014.12.023. [DOI] [PubMed] [Google Scholar]
- 34.Mulero I, Pilar Sepulcre M, Roca FJ, Meseguer J, García-Ayala A, Mulero V Characterization of macrophages from the bony fish gilthead seabream using an antibody against the macrophage colony-stimulating factor receptor. Developmental & Comparative Immunology. 2008;32(10):1151–1159. doi: 10.1016/j.dci.2008.03.005. [DOI] [PubMed] [Google Scholar]
- 35.Nakamichi Y, Udagawa N, Takahashi N IL-34 and CSF-1: similarities and differences. Journal of Bone and Mineral Metabolism. 2013;31(5):486–495. doi: 10.1007/s00774-013-0476-3. [DOI] [PubMed] [Google Scholar]
- 36.Nandi S, Gokhan S, Dai XM, Wei SW, Enikolopov G, Lin HS, Mehler MF, Stanley ER The CSF-1 receptor ligands IL-34 and CSF-1 exhibit distinct developmental brain expression patterns and regulate neural progenitor cell maintenance and maturation. Developmental Biology. 2012;367(2):100–113. doi: 10.1016/j.ydbio.2012.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Neves F, Abrantes J, Steinke JW, Esteves PJ Maximum-likelihood approaches reveal signatures of positive selection in IL genes in mammals. Innate Immunity. 2014;20(2):184–191. doi: 10.1177/1753425913486687. [DOI] [PubMed] [Google Scholar]
- 38.Ogryzko NV, Renshaw SA, Wilson HL The IL-1 family in fish: swimming through the muddy waters of inflammasome evolution. Developmental & Comparative Immunology. 2014;46(1):53–62. doi: 10.1016/j.dci.2014.03.008. [DOI] [PubMed] [Google Scholar]
- 39.Peyraud F, Cousin S, Italiano A CSF-1R inhibitor development: current clinical status. Current Oncology Reports. 2017;19(11):70. doi: 10.1007/s11912-017-0634-1. [DOI] [PubMed] [Google Scholar]
- 40.Rieger AM, Konowalchuk JD, Havixbeck JJ, Robbins JS, Smith MK, Lund JM, Barreda DR A soluble form of the CSF-1 receptor contributes to the inhibition of inflammation in a teleost fish. Developmental & Comparative Immunology. 2013;39(4):438–446. doi: 10.1016/j.dci.2012.12.001. [DOI] [PubMed] [Google Scholar]
- 41.Rieger AM, Hanington PC, Belosevic M, Barreda DR Control of CSF-1 induced inflammation in teleost fish by a soluble form of the CSF-1 receptor. Fish & Shellfish Immunology. 2014;41(1):45–51. doi: 10.1016/j.fsi.2014.03.035. [DOI] [PubMed] [Google Scholar]
- 42.Rieger AM, Havixbeck JJ, Belosevic M, Barreda DR Teleost soluble CSF-1R modulates cytokine profiles at an inflammatory site, and inhibits neutrophil chemotaxis, phagocytosis, and bacterial killing. Developmental & Comparative Immunology. 2015;49(2):259–266. doi: 10.1016/j.dci.2014.12.003. [DOI] [PubMed] [Google Scholar]
- 43.Roca FJ, Sepulcre MP, López-Castejón G, Meseguer J, Mulero V The colony-stimulating factor-1 receptor is a specific marker of macrophages from the bony fish gilthead seabream. Molecular Immunology. 2006;43(9):1418–1423. doi: 10.1016/j.molimm.2005.07.028. [DOI] [PubMed] [Google Scholar]
- 44.Secombes CJ What's new in fish cytokine research? Fish & Shellfish Immunology. 2016;53:1–3. doi: 10.1016/j.fsi.2016.03.035. [DOI] [PubMed] [Google Scholar]
- 45.Secombes CJ, Zou J Evolution of interferons and interferon receptors. Frontiers in Immunology. 2017;8:209. doi: 10.3389/fimmu.2017.00209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ségaliny AI, Brion R, Brulin B, Maillasson M, Charrier C, Téletchéa S, Heymann D IL-34 and M-CSF form a novel heteromeric cytokine and regulate the M-CSF receptor activation and localization. Cytokine. 2015;76(2):170–181. doi: 10.1016/j.cyto.2015.05.029. [DOI] [PubMed] [Google Scholar]
- 47.Slaven EM, Lopez FA, Hart SM, Sanders CV Myonecrosis caused by Edwardsiella tarda: a case report and case series of extraintestinal E. tarda infections . Clinical Infectious Diseases. 2001;32(10):1430–1433. doi: 10.1086/320152. [DOI] [PubMed] [Google Scholar]
- 48.Suzumura A Neuron-microglia interaction in neuroinflammation. Current Protein & Peptide Science. 2013;14(1):16–20. doi: 10.2174/1389203711314010004. [DOI] [PubMed] [Google Scholar]
- 49.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution. 2011;28(10):2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Truong AD, Hong Y, Lee J, Lee K, Kil DY, Lillehoj HS, Hong YH Interleukin-34 regulates Th1 and Th17 cytokine production by activating multiple signaling pathways through CSF-1R in chicken cell lines. International Journal of Molecular Sciences. 2018;19(6):1665. doi: 10.3390/ijms19061665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Vasek MJ, Garber C, Dorsey D, Durrant DM, Bollman B, Soung A, Yu JS, Perez-Torres C, Frouin A, Wilton DK, Funk K, DeMasters BK, Jiang XP, Bowen JR, Mennerick S, Robinson JK, Garbow JR, Tyler KL, Suthar MS, Schmidt RE, Stevens B, Klein RS A complement-microglial axis drives synapse loss during virus-induced memory impairment. Nature. 2016;534(7608):538–543. doi: 10.1038/nature18283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wang LP, Jiang LH, Wu CW, Lou B Molecular characterization and expression analysis of large yellow croaker (Larimichthys crocea) interleukin-12A, 16 and 34 after poly I: C and Vibrio anguillarum challenge . Fish & Shellfish Immunology. 2018;74:84–93. doi: 10.1016/j.fsi.2017.12.041. [DOI] [PubMed] [Google Scholar]
- 53.Wang TH, Kono T, Monte MM, Kuse H, Costa MM, Korenaga H, Maehr T, Husain M, Sakai M, Secombes CJ Identification of IL-34 in teleost fish: differential expression of rainbow trout IL-34, MCSF1 and MCSF2, ligands of the MCSF receptor. Molecular Immunology. 2013;53(4):398–409. doi: 10.1016/j.molimm.2012.09.008. [DOI] [PubMed] [Google Scholar]
- 54.Wang TH, Secombes CJ The cytokine networks of adaptive immunity in fish. Fish & Shellfish Immunology. 2013;35(6):1703–1718. doi: 10.1016/j.fsi.2013.08.030. [DOI] [PubMed] [Google Scholar]
- 55.Wang YM, Szretter KJ, Vermi W, Gilfillan S, Rossini C, Cella M, Barrow AD, Diamond MS, Colonna M IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia. Nature Immunology. 2012;13(8):753–760. doi: 10.1038/ni.2360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wang YM, Colonna M Interkeukin-34, a cytokine crucial for the differentiation and maintenance of tissue resident macrophages and Langerhans cells. European Journal of Immunology. 2014;44(6):1575–1581. doi: 10.1002/eji.201344365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Williams H, Brenner S, Venkatesh B Identification and analysis of additional copies of the platelet-derived growth factor receptor and colony stimulating factor 1 receptor genes in fugu. Gene. 2002;295(2):255–264. doi: 10.1016/S0378-1119(02)00736-9. [DOI] [PubMed] [Google Scholar]
- 58.Wu Y, Zhou YC, Cao ZJ, Sun Y, Chen Y, Xiang YJ, Wang L, Zhang SN, Guo WL Comparative analysis of the expression patterns of IL-1β, IL-11, and IL-34 in golden pompano (Trachinotus ovatus) following different pathogens challenge . Fish & Shellfish Immunology. 2019;93:863–870. doi: 10.1016/j.fsi.2019.08.018. [DOI] [PubMed] [Google Scholar]
- 59.Xu TT, Zhang XH Edwardsiella tarda: an intriguing problem in aquaculture . Aquaculture. 2014;431:129–135. doi: 10.1016/j.aquaculture.2013.12.001. [DOI] [Google Scholar]
- 60.Xue YJ, Jiang XY, Gao JD, Li X, Xu JW, Wang JY, Gao Q, Zou J Functional characterisation of interleukin 34 in grass carp Ctenopharyngodon idella . Fish & Shellfish Immunology. 2019;92:91–100. doi: 10.1016/j.fsi.2019.05.059. [DOI] [PubMed] [Google Scholar]
- 61.You XX, Bian C, Zan QJ, Xu X, Liu X, Chen JM, Wang JT, Qiu Y, Li WJ, Zhang XH, Sun Y, Chen SX, Hong WS, Li YX, Cheng SF, Fan GY, Shi CC, Liang J, Tom Tang Y, Yang CY, Ruan ZQ, Bai J, Peng C, Mu Q, Lu J, Fan MJ, Yang S, Huang ZY, Jiang XT, Fang XD, Zhang GJ, Zhang Y, Polgar G, Yu H, Li J, Liu ZJ, Zhang GQ, Ravi V, Coon SL, Wang J, Yang HM, Venkatesh B, Wang J, Shi Q Mudskipper genomes provide insights into the terrestrial adaptation of amphibious fishes. Nature Communications. 2014;5(1):5594. doi: 10.1038/ncomms6594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.You XX, Chen JM, Bian C, Yi YH, Ruan ZQ, Li J, Zhang XH, Yu H, Xu JM, Shi Q Transcriptomic evidence of adaptive tolerance to high environmental ammonia in mudskippers. Genomics. 2018;110(6):404–413. doi: 10.1016/j.ygeno.2018.09.001. [DOI] [PubMed] [Google Scholar]
- 63.Zhou J, Sun XY, Zhang J, Yang Y, Chen DP, Cao J IL-34 regulates IL-6 and IL-8 production in human lung fibroblasts via MAPK, PI3K-Akt, JAK and NF-κB signaling pathways . International Immunopharmacology. 2018;61:119–125. doi: 10.1016/j.intimp.2018.05.023. [DOI] [PubMed] [Google Scholar]
- 64.Zou J, Secombes CJ The function of fish cytokines. Biology. 2016;5(2):23. doi: 10.3390/biology5020023. [DOI] [PMC free article] [PubMed] [Google Scholar]