

Abstract
The recent outbreak of severe acute respiratory syndrome (SARS) provided an opportunity to study the antibody response of infected individuals to the causative virus, SARS coronavirus. We examined serum samples obtained from 46 patients with SARS, 40 patients with non-SARS pneumonia, and 38 healthy individuals, by use of Western blotting (WB), enzyme-linked immunoassay (ELISA), and immunofluorescence assay, using both native and bacterially produced antigens of the virus. We found a highly restricted, immunoglobulin G-dominated antibody response in patients with SARS, directed most frequently (89% by ELISA) and predominantly at the nucleocapsid. Almost all of the subjects without SARS had no antinucleocapsid antibodies. The spike protein was the next most frequently targeted, but only 63% of the patients (by ELISA) responded. Other targets of the response identified by use of WB included antigens of 80 and 60 kDa. Several nonstructural proteins cloned were not antigenic, and the culture-derived nucleocapsid appeared to be specifically degraded.
Severe acute respiratory syndrome (SARS) is a new infectious disease in humans caused by a novel coronavirus (CoV), SARS CoV [1–4]. This RNA virus is quite distinct from other CoVs known to infect humans or animals, on the basis of the structure of its 29, 751-bpgenome [5, 6]. However, like all CoVs, it has genes for polymerase and the structural proteins spike (S), envelope (E), membrane (M), and nucleocapsid (N) (figure 1A). In addition, it has genes for at least another 18 proteins, mostly nonstructural proteins (NSPs) and some putative proteins [7]. Presumably, as with other CoVs, SARS CoV infects cells through the S protein, which binds to specific cell receptors, such as aminopeptidase N [8]. After this initial attachment, the viral E protein fuses with the plasma membrane of the host cell, and a cascade of intracellular events follows, including interaction between theMand N proteins. This interaction is important for packaging of the progeny virions [9].
Figure 1.
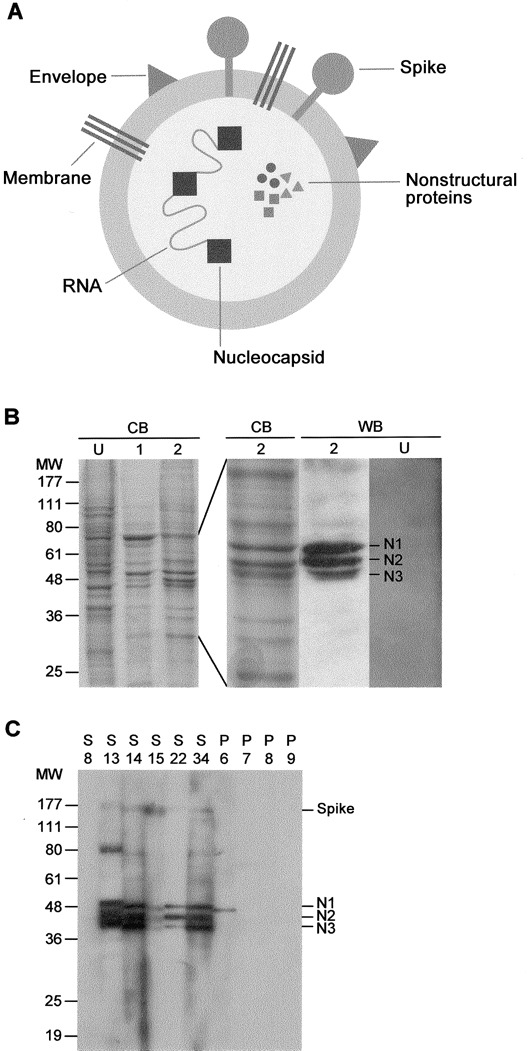
Identification of viral antigens that are reactive with severe acute respiratory syndrome (SARS) serum. A, Drawing of the SARS coronavirus structure showing the main antigens. B, Crude viral antigens obtained from culture cells were separated on gel and stained with Coomassie blue (CB). U, uninfected control cells. Lanes 1 and 2, Different antigen preparations showing slight variation in protein intensities at 36–48 kDa. Shown are results of Western blotting (WB) of preparation no. 2 or preparation U reacted with a serum sample from a patient with SARS showing the highly reactive antigens—nucleocapsid (N) 1, N2, and N3—in the former. MW, molecular weight markers (in kDa). C, WB resultsof 6 patients with SARS (S) and 4 patients with non-SARS pneumonia(P), showing strong reactivities of the N1–N3 antigens and lesser reactivities of the spike and the 80- and 60-kDa proteins or the partial reactivity (N1) or total lack of reactivity.
After infection, patients with SARS develop antibodies to the virus, and 190% of them experience seroconversion within 20 days [10]. However, little else is known about the antibody responses in these patients. The conventional antibody test, which detects antibodies to antigens present in virus-infected cells by use of immunofluorescence assay (IFA) [3], does not reveal which viral antigens are targeted by the immune response. Knowing the viral targets is important for several reasons. First, this allows the development of cell-free antibody tests that are less cumbersome and subjective than IFA and that can be used for mass screening in times of epidemics. Second, this enhances our understanding of the immunopathology of the disease, and moreover, helps in design of a vaccine. Both the humoral and cellular arms of the adaptive immune response are presumed to be important in controlling the infection. As with other CoVs, antibodies can function by blocking the adsorption or entry of the virus to the target cell [11–13] or by interfering with viral transcription [14]. In the present study, we identify the viral antigens to which our patients with SARS had responded.
Subjects, Materials, And Methods
Patients and control subjects. We diagnosed SARS according to the World Health Organization criteria [15], on the basis ofthe following symptoms: acute onset of fever (138°C) accompanied by frequent chills or rigor, dyspnea, headache, myalgia, hypoxemia, and radiological evidence of pneumonia. In addition, the patients with SARS had anti-SARS CoV IgG antibodies detected by use of an IFA [3], which showed seroconversion or a 4-fold increase in titer. In this assay, which uses virus-infected monkey kidney (Vero) cells and which is performed routinely in our laboratory, a serum titer of ⩽1:40 was considered to be positive. All 46 patients (15 males and 31 females; age range, 8–68 years; mean±SD age, 35.9±13.0 years) had contact with patients with confirmed SARS within 10 days of the onset of their symptoms and were admitted to the Prince of Wales Hospital, Hong Kong, during the SARS outbreak that started in early March 2003. Two of the patients died, 5 required intensive carebut eventually recovered, and the rest had a mild clinical course. Because methods of detection were not available early during the outbreak, the SARS CoV was cultured from only 1 patient, and viral RNA was detected by use of reverse-transcription polymerasechain reaction (PCR) [16] from the respiratory or stool samples from 4 patients. The serum samples used in the present study were obtained 7–35 days (mean±SD, 17.1±6.4 days) after the onset of fever.
Control subjects included 40 inpatients (21 males and 19 females; age range, 8–84 years; mean age, 43.3 years) who had presented at the Prince of Wales Hospital with clinical features of atypical pneumonia and were treated accordingly at the hospital in 2000 before the emergence of SARS and 38 healthy individuals (blood donors and students). Institutional approval for both human and animal ethics was obtained for the study, and the institutional guidelines were followed.
Viral culture and crude native antigens. Vero cells (ATCCCRL-1586) were grown in Dulbecco's modified Eagle medium containing 5% fetal calf serum at 37°C in a 5% CO2 humidified incubator. The CUHK-W1 SARS CoV strain (GenBank accession no. AY278554) used was isolated from a patient in our hospital. Virus stocks were prepared by infecting the Vero cells and harvestingthe culture supernatant 20–48 h after infection. To prepare the crude viral antigens, a subconfluent monolayer of Vero cells was infected with the virus stock. At 16–48 h after infection,the cells were harvested and resuspended in lysis buffer (10 mmol/L HEPES [pH 7.0], 40 mmol/L KCl, 3 mmol/L MgCl2, 5% glycerol, 0.2% nonidet-P40, 1 mmol/L 1, 4-dithiothreitol, 1mmol/L phenylmethylsulfonyl fluoride [PMSF], and 1×protease inhibitor cocktail [Sigma]). After incubation for 1 h on ice, the supernatant obtained from the cell lysate by centrifugation was heated for 30 min at 55°C, to in activate any live virus present,and was stored at −70°C until use. Control cell lysate was similarly prepared from uninfected Vero cells.
Recombinant viral antigens. Total RNA extracted from SARS CoV-infected Vero cells by use of a QIAmp Viral RNA Kit (Qiagen) was reverse-transcribed with random hexamers (Applied Biosystems), and the cDNA obtained was used to generate the various gene segments. The following forward (F) primers (containing a BamH1 site, underlined) and reverse (R) primers (containing an EcoR1 site, underlined) were used in PCRs described elsewhere [17] for the corresponding gene segments (sizes given): rSa (N-terminal spike, 957 bp; F, 5′-CGTGGATCCAATTCTACTAATGTTGTTATA-3′; R, 5′-CGATGAATTCCATTATAATTACCAGTTGAAGT-3′), rSb (median spike, 1365 bp; F, 5′-CGTGGATCCACTTCAACTGGTAATTATAAT-3′; R, 5′ -CGATGAATTCCTGCCATTTGCATAGCAAAAGG-3′), rSc (C-terminal spike, 948 bp; F, 5′-CGTGGATCCCCTTTTGCTATGCAAATGGCA-3′; R, 5′ -CGATGAATTCCCCATTTAATATATTGCTCATA-3′), rNa (N-terminal N, 660 bp;F,5′-CGTGGATCCATGTCTGATAATGGACCCCAA-3′; R, 5′ -CGATGAATTCCGAGGGCAGTTTCACCACCTCC-3′), rNSP12a (N-terminal, 522 bp; F, 5′-CGTGGATCCAGTTTAGAAAATGTGGCTTAT-3′; R, 5′ -CGATGAATTCCTGTTTTTACTGATTCTCCAAT-3′), rNSP12b (C-terminal, 37 bp; F, 5′-CGTGGATCCATTGGAGAATCAGTAAAAACA-3′; R, 5′ -CGATGAATTCCTTGTAGTTTTGGGTAGAAGGT-3′), rNSP13 (whole, 894 bp; F, 5′-CGTGGATCCGCAAGTCAAGCGTGGCAACCA-3′; R,5′ -CGATGAATTCCGTTGTTAACAAGAATATCACT-3′), and rNSP9 (N-terminal, 720 bp; F, 5′-CGTGGATCCGAGGAAGGCAATTTATTAGAC-3′; R, 5′-CGATGAATTCCGTTAATACAATTGGGATGGAT-3′).
The PCR conditions used were 3 min at 94°C, 34 cycles of 1 min at 94°C, 1 min at 55°C, 1 min at 72°C, and 15 min at 72°C. The gel- and affinity-purified fragments were cloned into thebacterial expression vector pGEX-2T (Amersham Bioscience), which is fused to the bacterial glutathione S-transferase (GST) gene. The ligated vector was transfected to Escherichia coli BL21.
Recombinant antigens were recovered from selected transformants induced with isopropyl-²-d-thiogalactopyranoside. In brief, the cell pellet was resuspended in ice-cold lysis buffer (25 mmol/L Tris-HCl, 100 mmol/L NaCl, 0.1% Triton X-100,1 mmol/L PMSF, and 1×protease inhibitor cocktail [pH 8.0; Sigma]) and was sonicated (30 mm). The recombinant antigen was recovered from the supernatant of the lysate by use of affinity chromatography using glutathione-coupled agarose (Amersham) and, for elution, 25 mmol/L Tris-HCl buffer (pH 8.0) containing 20 mmol/L reduced-form glutathione, 100 mmol/L NaCl, 0.1% Triton X-100, and 5 mmol/L DDT. The eluted antigen was examined on SDS-PAGE gels stained with Coomassie blue (expected size and good purity were observed in all cases; data not shown).
SDS-PAGE and Western blotting (WB). In SDS-PAGE, the antigen preparation was heated for 5 min at 100°C in loadingbuffer (0.25 mol/L Tris-HCl [pH 6.8], 20% 2-mercaptoethanol, 40% glycerol, 8% SDS, and 0.01% bromophenol blue)and electrophoresed (150 V for 80 min at room temperature [RT]) on 10% polyacrylamide. Antigens of interest (pN2, pN3, and pS) were located in the unstained gel by use of a parallelgel stained with Coomassie blue and were recovered with aneluter (Harvard Bioscience) at 40 V overnight in 25 mmol/LTris buffer (pH 8.3) containing 192 mmol/L glycine, 20% methanol, and 0.5% SDS. In other experiments, the whole gel was electroblotted onto a 0.22-µm polyvinylidene fluoride membrane (Bio-Rad) and was used for protein sequencing (Protein Facility, Iowa State University) or for WB. In the latter assay, the membrane was blocked with skim milk and incubated (for 1 hat RT) with the unknown human or mouse serum (diluted 1:200 in 2 mL of PBS containing 5% skim milk) [8]. After washing, the blot was incubated (for 1 h at RT) with peroxidase-labeled goat anti-human IgG or anti-mouse Ig (all classes) (BD Biosciences) and later with the chemiluminescence substrate ECL (Amersham). The assay was developed by exposure to hyperfilm ² max (Amersham). In inhibition WB, the unknown serum (10 µL) was preincubated with the inhibiting antigen (600 µg/mL) in 50 µL of PBS containing 5% skim milk overnight at 4°C.
ELISA. The native viral antigens, either crude or purified (pN2, pN3, and pS; 1:200 stock dilution), or the recombinant antigens (rNa, rSa, rSb, rSc, rNSP12a, rNSP12b, rNSP13, and rNSP9; 1 µg/mL) were coated onto 96-well Immunon-2 plates(Dynex) in bicarbonate buffer (pH 9.6) overnight at 4°C, and the assay was performed as described elsewhere [17]. In brief, 100 µL of the unknown human serum diluted 1:50 in PBS (containing 1.3% bovine serum albumin, 0.25% casein, and 0.05% Tween-20) were added to the wells and incubated for 30 min at RT. The plates were washed and incubated (for 15min at RT) with horse radish peroxidase-labeled goat anti-human Ig (IgG, IgM, or IgA specific) (BD Biosciences). Afterwashing, substrate (3,3′,5,5′-tetramethylbenzidine) was added, and reaction was allowed for 15 min at RT. The results were read at 450 nm in a Dynex MRX II reader.
Mouse immunization. BALB/c mice (3 mice/group) were injected intraperitoneally with the affinity-purified, alum-precipitated antigen (200 µg/mouse) in complete Freund's adjuvantand received booster injections with 50 µg of the antigen in incomplete Freund's adjuvant 2 weeks later. Blood was obtained from the retro-orbital plexus 4 days after administration of the booster injection.
Statistics. The relationships of the (rNa) ELISA with other immunoassays or with sampling times were examined by use of regression analysis (GraphPad Prism 3; GraphPad Software).
Results
We first examined the antibody response of the patients with SARS by use of WB. Figure 1B shows the gel-separated crude extract of the culture-grown virus. Several virus-specific antigens are discernible by comparing the control (uninfected) extract with the viral extract, particularly ones at 48, 46, and 44 kDa molecular weight. The antigenicity of these proteins, labeled N1, N2, and N3, respectively, is shown by immunoblotting with serum samples obtained from patients with SARS (figure 1B). The relative abundance of N1–N3 varied among batches of the extracts, but N3, which is actually a doublet, was consistently lowest in quantity.
The WB results from representative patients with SARS and patients with non-SARS pneumonia are shown in figure 1C. These results were based on the IgG response; the IgM reactions were either weak or absent. The N1–N3 antigens were the most reactive antigens found in the patients with SARS but were absent in patients with non-SARS pneumonia. These 3 antigens always appeared as a triplet. One of the non-SARS serum samples reacted weakly with an antigen at the position of N1. Less reactive antigens found at 150, 80, and 60 kDa were not seenin the non-SARS serum samples. The molecular size of N1 suggests that it is the nucleocapsid, but N2 and N3 could notbe identified in this way. Protein sequencing of these antigenswas not successful, but it confirmed the 150-kDa “S” antigen (SDLDR) to be the S protein.
On the basis of WB results for a total of 43 patients with SARS, 23 patients with non-SARS pneumonia, and 16 healthy subjects, most (79%) of the patients with SARS produced antibodies tothe N1–N3 antigens, but only 40% produced antibodies to theS protein (figure 2). In addition, 35%, 26%, 19%, and 5% of the patients with SARS produced antibodies to the 80-, 60-, 32-, and 24-kDa antigens, respectively (data not shown). None ofthe control subjects produced antibodies to any of these antigens, although 13% of the serum samples from patients withn on-SARS pneumonia showed weak reactivity with the (presumably) N1 antigen (see below). The SARS serum samples were positive for viral antibodies when examined by use of an IFA using infected cells, although some (17%) had low titers (<1:80) only (figure 2). None of the serum samples from the 2 control cohorts examined were positive by either IFA or WB.
Figure 2.
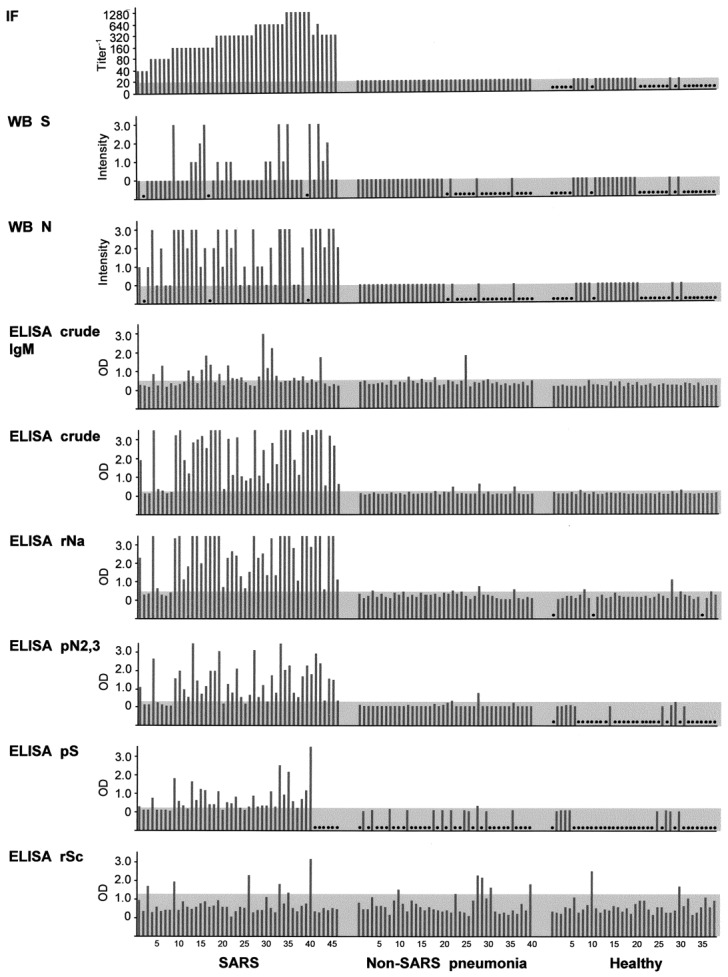
Comparison of the efficiency of various detection assays for severe acute respiratory syndrome (SARS). Individual serum samples from each group (46 patients with SARS, 40 patients with non-SARS pneumonia, and 38 healthy individuals) were examined by use of immunofluore scenceassay (IFA), Western blotting (WB), or ELISA. Antigens used included native spike (S), native nucleocapsid (N), crude antigen, crude viral extract, recombinant nucleocapsid (rNa), gel-purified native N2 and N3 antigens (pN2, 3), gel-purified native spike (pS), and recombinant spike (rSc). Except where marked “IgM”, all tests are based on detection of IgG. In each test, the cut off for positivity is shown by the shaded bar; for ELISA, this is based on the mean+1 SD value of the combined cohorts of patients with non-SARS pneumonia and healthy individuals. In WB, the intensity of the reaction was arbitrarily scored by eye (3, strongest). Serum samples that were not examined because of a lack of antigen or serum or because the results were not readable are shown by a dot.
Using a bigger study group (46 patients with SARS, 40 patients with non-SARS pneumonia, and 38 healthy individuals), we examined the antibody responses further by use of ELISA. First, when the crude viral extract was used as antigen, 91% of the patients with SARS were positive for antibodies, compared with <6% of the combined control subjects (figure 2). All 4 patients with SARS who were negative for antibodies had low IFA titers (<1:80). All serum samples were not reactive with the control antigen, GST (data not shown). The results are based on the IgG response. The IgM response, in contrast, was less robust and less frequent (43%) among the patients with SARS (figure 2) and less discriminatory between the SARS and non-SARS cohorts. Second, we made a recombinant antigen of the N-terminal half of the N protein (rNa), and, when it was used in an IgG ELISA, we found results almost identical to those found with the crude viral extract (89% sensitivity and 94%–95% specificity), including 4 negative cases in common (figure 2). The IgM responses were similarly low and infrequent among the patients with SARS (data not shown).
We investigated the possibility that the 20% of serum samples that were negative or weakly positive in the ELISA but were IFA positive might have antibodies directed to antigens other than the nucleocapsid or those present in the crude extract. We thus examined the recombinant antigens made from several NSPs of the virus. However, although NSP12 (both subunits) and NSP9 were found to be nonantigenic, NSP13 showed only weak reactivities with some (11%) of the serum samples from patients with SARS (data not shown).
Native S antigen that was purified and used in an ELISA detected antibody responses in 63% of the patients with SARS (figure 2), slightly more than those detected by use of WB. However, most of the responses were weak. Using recombinantS antigens, we found that only the C-terminal end (rSc) of the protein was antigenic and that only a small number (13%) of serum samples from patients with SARS were reactive, similar to results for serum samples from patients with non-SARS pneumonia (figure 2).
We purified the native N2 and N3 antigens together from the gel-separated crude extract and used these in an ELISA. The results obtained were very similar to those of the rNa ELISA, particularly with respect to the negative or positive cases (figure 2). This suggests that N2 or N3 or both might be antigenically related to rNa. To investigate this possibility, we performed WB experiments using rNa as inhibitor. rNa was indeed found to reduce the reactivity of not only N1, but also of N2 and N3 (figure 3A). In fact, inhibition was greatest with N3 and least with N1. Of interest, there appeared to be a fourth, minor N fragment (N4) with a molecular weight of 32 kDa (figure 3A) that was seen with some serum samples (e.g., S13; figure 1). Similar results were obtained when the purified N2 and N3 antigens were used as the inhibitor in the WB analysis (figure 3A). In contrast, when the S antigen, rSc, was used as inhibitor, reactivity at the 150-kDa region (“S”), but not that of N1–N4 , was affected (i.e., abolished) (figure 3A).
Figure 3.
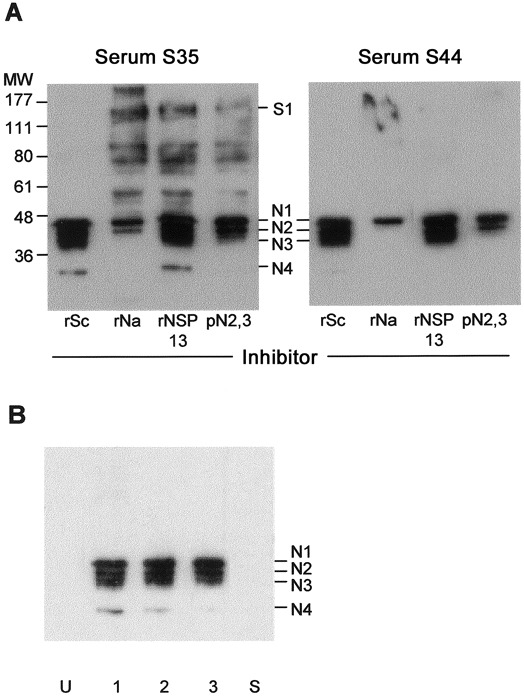
Demonstration that nucleocapsid (N) 1-N4 are all N antigens. A, Western blotting (WB) of 2 serum samples from patients with severe acute respiratory syndrome (SARS) (nos. S35 and S44) against the crude viral extract in the presence of various antigens used as inhibitor. B, WB of serum samples obtained from 3 mice immunized with recombinant N (rNa)-GST (lanes 1, 2, and 3) or from a representative mouse immunized with recombinant spike (rSa)-GST or recombinant spike (rSc)-GST (lane S), against the crude viral extract. MW, molecular weight (in kDa); pN2,3, gel-purified native N2 and N3 antigens; rNSP, recombinant nonstructural protein; U, unimmunized mouse.
We proved further that N1, N2, N3, and N4 are all N antigens. Mouse serum made against the recombinant antigens rNa, rSa, and rSc (rSb was not available at the time) were used in WB against the crude viral extract. All 3 mice immunized with rNa produced antibodies that reacted specifically with N1,N2, N3, and N4, but not with other antigens (figure 3B). In contrast, neither of the S antigens produced any antibodies against the crude extract (figure 3B).
Discussion
The N antigen of SARS CoV stands out as the most important diagnostic antigen of the virus. The majority of our patients who developed SARS (89% by ELISA) produced antibodies to this antigen. When present, these antibodies are also the most abundant of the antibodies made to the virus. It is not clear why the N antigen is so immunogenic, but we have found thateven a bacterially produced fragment of it induced good production of antibodies in mice. The N antigen also has been found to be highly immunogenic in the elk CoV [18] and in the infectious bronchitis virus [19].
We have found the N antigen to be the predominant antigenin the crude viral extract. This is shown by the good correlation between the rNa and crude antigen ELISAs (figures 2 and 4A). In contrast, there was only weak correlation between the rNa ELISA and the IFA performed on these serum samples (figure 4B). This result was not due to the fact that we used a single dilution measurement in the ELISA, rather than titrating the serum (figure 4C). It is possible that some antigens cannot be extracted from the cell or become degraded during the extraction, but this was not the case with NSP12 or NSP9. It is noteworthy that the serum samples from the 3 patients with non-SARS pneumonia, which reacted with the (presumably) N1 component by WB, were negative by the rNa ELISA. This result suggests that the reactive epitope is located elsewhere in the N antigen or in a nonviral (Vero) antigen.
Figure 4.

Performance of the recombinant nucleocapsid (rNa)—ELISA using serum samples from patients with severe acute respiratory syndrome (SARS). A, Comparison with the crude ELISA by regression analysis. B, Comparison with immunofluorescence assay (IFA). C, Two examples where both of the serum samples have similar ELISA but vastly different IFA titers. D, Relationship between stage of disease and test sensitivity.
As expected, detection of the anti-N antigen antibodies in patients with SARS by use of the rNa ELISA improved with increasing intervals between the time samples were obtained and the onset of fever (figure 4D). Thus, in 3 of the 5 negative cases, the serum was obtained within 10 days after the onset of fever. On the other hand, 4 other serum samples obtained during the same period were found to be positive. Although this means that more than half of the early SARS cases (and all of the late [⩾21 days] cases) could be detected, it is important to note that we used serum samples that were IFA selected.
Of note with regard to the anti-N antibody response in the patients with SARS is the predominance of IgG antibodies over IgM antibodies, even early (second week) during the course of the disease. This result implies that there was strong T helper cell involvement. An exaggerated T cell response probably also accounts for the pneumonia-associated immunopathology seen in patients with SARS [10]. The N antigen may be important here. It may indeed be both a potent B cell and a potent T cell immunogen; that is, an important candidate for a vaccine. Although antibodies to the nucleocapsid are nonneutralizing, whether such antibodies can be protective in other ways, as observed for mouse hepatitis virus [20] and rotavirus [21], needs to be addressed formally. The latter phenomenon maybe due to the ability of the IgA antibodies to enter the infected cell by the IgA-transcytosis pathway, thereby interfering with viral transcription [14].
We found evidence that the N antigen is degraded in cell cultures of the virus. The degradation is specific, since the 3 major N antigens, N1–N3, were found in every preparation ofthe crude extract that we used. A minor antigen, N4, was also sometimes seen. N1 is presumably the full-length protein, and the others are fragments of N1 that lack varying lengths of the C-terminal end. Because of the high specificity seen, we do not consider the degradation to be an artifact of the antigen-extraction procedure. Rather, we suspect that N1 is cleaved by caspases in the Vero cells as the cells undergo apoptosis, similar to what Eleouet et al. [22] observed for the N antigen from the transmissible gastroenteritis CoV.
After the N antigen, the S antigen is the one most often targeted by the immune response of the patients with SARS. However, <63% (by ELISA) of the patients responded, and their responses were generally weak. It is possible, on the other hand, that the responses were underestimated because the S antigen contained in the crude extract used for WB or ELISA was degraded or under represented and the recombinant antigen used for ELISA that was produced in bacteria lacked the necessary glycosylation normally associated with the native antigen (see: http://www.cbs.dtu.dk).
Other antigens of the virus targeted by the immune response are presumably minor. Examples are the 80- and 60-kDa antigens. The latter could be NSP13, recently postulated to be an mRNA cap-1 methyltransferase [23]. The 24-kDa antigen to which some patients responded could be the M protein. Both this and the E protein of the virus (the latter was too small [8.5 kDa] to assess in our WB gels) are presumed to be important in protection.
Acknowledgment
We thank Peggy Fung for excellent secretarial help.
Footnotes
Presented in part: Closed-door meeting of the SARS Laboratory Network of the World Health Organization, Geneva, Switzerland, 22 October 2003.
Financial support: Research Grants Council of Hong Kong (grant CUHK 4527/03M).
References
- 1.Poutanen SM, Low DE, Henry B, et al. Identification of severe acute respiratory syndrome in Canada. N Engl J Med. 2003;348:1995–2005. doi: 10.1056/NEJMoa030634. [DOI] [PubMed] [Google Scholar]
- 2.Peiris JSM, Lai ST, Poon LLM, et al. Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet. 2003;361:1319–25. doi: 10.1016/S0140-6736(03)13077-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ksiazek TG, Erdman D, Goldsmith CS, et al. A novel coronavirus associated with severe acute respiratory syndrome. N Engl J Med. 2003;348:1953–66. doi: 10.1056/NEJMoa030781. [DOI] [PubMed] [Google Scholar]
- 4.Drosten C, Gunther S, Preiser W, et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N Engl J Med. 2003;348:1967–76. doi: 10.1056/NEJMoa030747. [DOI] [PubMed] [Google Scholar]
- 5.Marra MA, Jones SJ, Astell CR, et al. The genome sequence of the SARS-associated coronavirus. Science. 2003;300:1399–404. doi: 10.1126/science.1085953. [DOI] [PubMed] [Google Scholar]
- 6.Rota PA, Oberste MS, Monroe SS, et al. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science. 2003;300:1394–9. doi: 10.1126/science.1085952. [DOI] [PubMed] [Google Scholar]
- 7.Ruan YJ, Wei CL, Ee LA, et al. Comparative full-length genome sequence analysis of 14 SARS coronavirus isolates and common mutations associated with putative origins of infection. Lancet. 2003;361:1779–85. doi: 10.1016/S0140-6736(03)13414-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bonavia A, Zelus BD, Wentworth DE, Talbot PJ, Holmes KV. Identification of a receptor-binding domain of the spike glycoprotein of human coronavirus HCoV-229E. J Virol. 2003;77:2530–8. doi: 10.1128/JVI.77.4.2530-2538.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Narayanan K, Maeda A, Maeda J, Makino S. Characterization of the coronavirus M protein and nucleocapsid interaction in infected cells. J Virol. 2000;74:8127–34. doi: 10.1128/jvi.74.17.8127-8134.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Peiris JS, Chu CM, Cheng VC, et al. Clinical progression and viral load in a community outbreak of coronavirus-associated SARS pneumonia: a prospective study. Lancet. 2003;361:1767–72. doi: 10.1016/S0140-6736(03)13412-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Torres JM, Sanchez C, Sune C, et al. Induction of antibodies protecting against transmissible gastroenteritis coronavirus (TGEV) by recombinant adenovirus expressing TGEV spike protein. Virology. 1995;213:503–16. doi: 10.1006/viro.1995.0023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lamarre A, Talbot PJ. Protection from lethal coronavirus infection by immunoglobulin fragments. J Immunol. 1995;154:3975–84. [PubMed] [Google Scholar]
- 13.Castilla J, Sola I, Enjuanes L. Interference of coronavirus infection by expression of immunoglobulin G (IgG) or IgA virus-neutralizing antibodies. J Virol. 1997;71:5251–8. doi: 10.1128/jvi.71.7.5251-5258.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Feng N, Lawton JA, Gilbert J, et al. Inhibition of rotavirus replication by a non-neutralizing, rotavirus VP6-specific IgA mAb. J Clin Invest. 2002;109:1203–13. doi: 10.1172/JCI14397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.World Health Organization. Case definitions for surveillance of severe acute respiratory syndrome (SARS) Available at: http://www.who.int/csr/sars/casedefinition/en/print.html. Accessed 1 May 2003. [Google Scholar]
- 16.Chan PKS, Tam JS, Lam CW, et al. Human metapneumovirus detection in patients with severe acute respiratory syndrome. Emerg Infect Dis. 2003;9:1058–63. doi: 10.3201/eid0909.030304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Leung DTM, Yam NWC, Chui YL, Wong KC, Lim PL. A human and a mouse anti-idiotypic antibody specific for human T14+ anti-DNA antibodies reconstructed by phage display. Gene. 2000;255:373–80. doi: 10.1016/s0378-1119(00)00312-7. [DOI] [PubMed] [Google Scholar]
- 18.Daginakatte GC, Chard Bergstrom C, Andrews GA, Kapil S. Production, characterization, and uses of monoclonal antibodies against recombinant nucleoprotein of elk coronavirus. Clin Diagn Lab Immunol. 1999;6:341–4. doi: 10.1128/cdli.6.3.341-344.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ndifuna A, Waters AK, Zhou M, Collisson EW. Recombinant nucleocapsid protein is potentially an inexpensive, effective serodiagnostic reagent for infectious bronchitis virus. J Virol Methods. 1998;70:37–44. doi: 10.1016/S0166-0934(97)00170-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nakanaga K, Yamanouchi K, Fujiwara K. Protective effect of monoclonal antibodies on lethal mouse hepatitis virus infection in mice. J Virol. 1986;59:168, 71. doi: 10.1128/jvi.59.1.168-171.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Burns JW, Siadat Pajouh M, Krishnaney AA, Greenberg HB. Protective effect of rotavirus VP6-specific IgA monoclonal antibodies that lack neutralizing activity. Science. 1996;272:104–7. doi: 10.1126/science.272.5258.104. [DOI] [PubMed] [Google Scholar]
- 22.Eleouet JF, Slee EA, Saurini F, et al. The viral nucleocapsid protein of transmissible gastroenteritis coronavirus (TGEV) is cleaved by caspase- 6 and -7 during TGEV-induced apoptosis. J Virol. 2000;74:3975–83. doi: 10.1128/jvi.74.9.3975-3983.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.von Grotthuss M, Wyrwicz LS, Rychlewski L. mRNA cap-1 methyltransferase in the SARS genome. Cell. 2003;113:701–2. doi: 10.1016/S0092-8674(03)00424-0. [DOI] [PMC free article] [PubMed] [Google Scholar]