

Highlights
-
•
Middle East Respiratory Syndrome (MERS) is a zoonotic viral disease transmissible from dromedaries to human beings.
-
•
MERS coronavirus (MERS-CoV) infection in dromedaries is usually asymptomatic but mild respiratory signs may occur.
-
•
No MERS-CoV or antibodies against the virus have been detected in camelids other than dromedaries.
-
•
Dromedaries acquire MERS-CoV at less than 1 year of age and 100% of adult dromedaries are seropositive for the virus.
-
•
MERS-CoV in dromedaries is detected by virus isolation, quantitative reverse transcriptase-PCR or antigen detection.
Keywords: Middle East respiratory syndrome (MERS), Coronavirus, Dromedary, Diagnosis
Abstract
Middle East Respiratory Syndrome (MERS) is a zoonotic viral disease that can be transmitted from dromedaries to human beings. More than 1500 cases of MERS have been reported in human beings to date. Although MERS has been associated with 30% case fatality in human beings, MERS coronavirus (MERS-CoV) infection in dromedaries is usually asymptomatic. In rare cases, dromedaries may develop mild respiratory signs. No MERS-CoV or antibodies against the virus have been detected in camelids other than dromedaries. MERS-CoV is mainly acquired in dromedaries when they are less than 1 year of age, and the proportion of seropositivity increases with age to a seroprevalence of 100% in adult dromedaries. Laboratory diagnosis of MERS-CoV infection in dromedaries can be achieved through virus isolation using Vero cells, RNA detection by real-time quantitative reverse transcriptase-PCR and antigen detection using respiratory specimens or serum. Rapid nucleocapsid antigen detection using a lateral flow platform allows efficient screening of dromedaries carrying MERS-CoV. In addition to MERS-CoV, which is a lineage C virus in the Betacoronavirus (betaCoV) genus, a lineage B betaCoV and a virus in the Alphacoronavirus (alphaCoV) genus have been detected in dromedaries. Dromedary CoV UAE-HKU23 is closely related to human CoV OC43, whereas the alphaCoV has not been detected in human beings to date.
Introduction
Since the emergence of Middle East respiratory syndrome (MERS) in 2012, more than 1500 human cases have been reported, with a case fatality of ~30% (World Health Organization (WHO), 2016). The causative agent of MERS has been confirmed to be a novel coronavirus (CoV), named MERS-CoV (van Boheemen et al, 2012, Zaki et al, 2012). CoVs are positive-sense single-stranded RNA viruses with genome size of ~30 kb. They have the characteristic crown-shaped appearance on electron microscopic examination. CoVs are classified into four genera, Alphacoronavirus, Betacoronavirus (with four lineages), Gammacoronavirus and Deltacoronavirus (Woo et al., 2012). MERS-CoV belongs to lineage C of Betacoronavirus (Fig. 1 ) (van Boheemen et al., 2012).
Fig. 1.
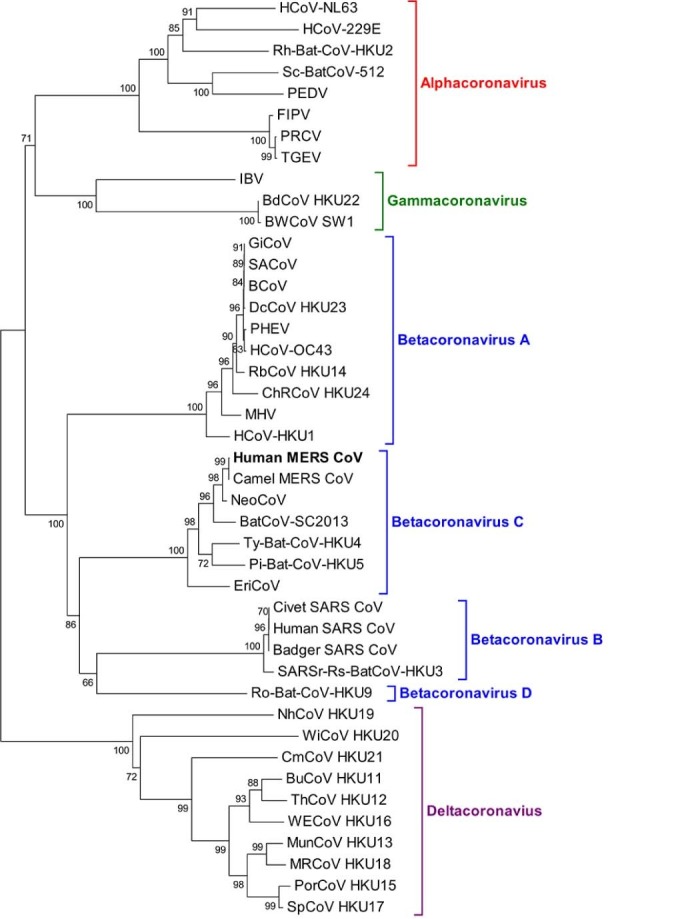
Phylogenetic analysis of RNA-dependent-RNA-polymerase (RdRp) of Middle East respiratory syndrome (MERS) coronavirus (MERS-CoV) and other coronaviruses. The tree was constructed by neighbour-joining method using maximum composite likelihood substitution model with bootstrap values calculated from 1000 trees. Virus list and GenBank accession numbers as follow: HCoV-NL63, human CoV NL63 (NC_005831); HCoV-229E (NC_002645); RhBatCoV HKU2, rhinolophus bat CoV HKU2 (EF203064); Sc-BatCoV-512, scotophilus bat CoV 512 (NC_009657); PEDV, porcine epidemic diarrhoea virus (NC_003436); FIPV, feline infectious peritonitis virus (AY994055); PRCV, porcine respiratory CoV (DQ811787); TGEV, transmissible gastroenteritis virus (NC_002306); IBV, infectious bronchitis virus (NC_001451); BdCoV HKU22, bottlenose dolphin CoV HKU22 (KF793826); BWCoV-SW1, Beluga whale CoV SW1 (NC_010646); GiCoV, giraffe CoV (EF424622); SACoV, sable antelope CoV (EF424621); BCoV, bovine CoV (NC_003045); DcCoV HKU23, dromedary camel CoV HKU23 (KF906251); PHEV, porcine haemagglutinating encephalomyelitis virus (NC_007732); HCoV-OC43, human CoV OC43 (NC_005147); RbCoV HKU14, rabbit CoV HKU14 (JN874559); ChRCoV HKU24, China Rattus CoV HKU24 (KM349742); MHV, murine hepatitis virus (NC_001846); HCoV-HKU1, human CoV HKU1 (NC_006577); human MERS-CoV, human Middle East respiratory syndrome CoV (JX869059); Camel MER-CoV, Camel Middle East respiratory syndrome CoV (KT751244); NeoCoV, Neoromicia CoV (KC869678); BatCoV-SC2013, Bat coronavirus SC2013 (KJ473821); Ty-BatCoV HKU4, tylonycteris bat CoV HKU4 (NC_009019); Pi-BatCoV HKU5, pipistrellus bat CoV HKU5 (NC_009020); EriCoV, ErinaceusCoV (KC545383); Civet SARS CoV, SARS-related palm civet CoV (AY304488); human SARS-CoV, severe acute respiratory syndrome-associated human CoV (NC_004718); badger SARS-CoV, SARS-related Chinese ferret badger CoV (AY545919); SARSr-Rs-BatCoV-HKU3, SARS-related rhinolophus bat CoV HKU3 (DQ022305); Ro-BatCoV HKU9, rousettus bat CoV HKU9 (NC_009021); NhCoV HKU19, night-heron CoV HKU19 (NC_016994); WiCoV HKU20, wigeon CoV HKU20 (NC_016995); CmCoV HKU21, common-moorhen CoV HKU21 (NC_016996); BuCoV HKU11, bulbul CoV HKU11 (FJ376619); ThCoV HKU12, thrush CoV HKU12 (FJ376621); WECoV HKU16, white-eye CoV HKU16 (NC_016991); MunCoV HKU13, munia CoV HKU13 (FJ376622); MRCoV HKU18, magpie-robin CoV HKU18 (NC_016993); PorCoV HKU15, porcine CoV HKU15 (NC_016990); SpCoV HKU17, sparrow CoV HKU17 (NC_016992).
Subsequent investigations have shown that dromedary or one humped camels (Camelus dromedarius) are so far the only reservoir of MERS-CoV (Alagaili et al, 2014, Chu et al, 2014, Wernery et al, 2015a, Wernery et al, 2015b, Sabir et al, 2016). Adult dromedaries have almost 100% seropositivity against MERS-CoV while the virus is found mainly in dromedary calves (Alagaili et al, 2014, Wernery et al, 2015b). Camel-to-human transmission of MERS-CoV and subsequent human-to-human transmission result in MERS in human beings, many of whom develop severe lower respiratory tract infections, with renal failure in some cases (Arabi et al, 2014, Saad et al, 2014). In contrast to human infection, MERS-CoV causes no or mild disease in dromedaries. In this article, our current understanding of MERS-CoV infection in dromedaries is reviewed and the presence of other CoVs in dromedaries is also discussed.
MERS-CoV infection in dromedaries
More than 20 MERS-CoV isolates have been recovered from nasal swabs from young dromedaries from different farms in Dubai over the last 2 years at the Central Veterinary Research Laboratory. These animals died from diseases unrelated to MERS, such as selenium deficiency, cryptosporidiosis, salmonellosis, Escherichia coli septicaemia and Clostridium perfringens A enterotoxaemia. None had nasal discharge and it is believed that the virus isolation was incidental to the death of the dromedaries. Histopathological investigations did not show any lesions consistent with virus infection.
However, experimental infections of dromedaries with MERS-CoV in the USA and Spain predominantly induced respiratory tract disease with no or mild clinical respiratory signs (Adney et al, 2014, Haagmans et al, 2016). The experimentally infected animals demonstrated moderate rhinitis, with nasal discharge, tracheitis and bronchitis, but no involvement of the alveolar tissue. The dromedaries were infected with high doses of MERS-CoV intranasally, i.e. 1 × 107 50% tissue culture infectious doses (TCID50) in the study of Adney et al. (2014) and 5 × 106 TCID50 in the study of Haagmans et al. (2016). Experimental MERS-CoV infections were also performed in alpacas, with similar results (Adney et al., 2016). Experimentally infected alpacas transmitted the virus to two of three contact animals. Experimentally infected animals were protected against reinfection 70 days later and those infected by contact were only partially protected.
Laboratory diagnosis of MERS-CoV infection in dromedaries
Detection of MERS-CoV in dromedaries is performed to understand the epidemiology and evolutionary dynamics of the virus and to reduce the risk of human transmission. MERS-CoV was first isolated from a human patient suffering from fatal lower respiratory tract infection and acute renal failure in Saudi Arabia in 2012 (van Boheemen et al, 2012, Zaki et al, 2012). Since then, the virus has been isolated from both human beings and dromedaries. Although Vero cells are usually the cell line used in clinical laboratories, the virus can be propagated in a variety of human and non-human cell lines, which may have implications for its tissue tropism and the high fatality associated with MERS (Chan et al, 2013, Eckerle et al, 2014, Zhou et al, 2015).
MERS-CoV has been isolated from nasal swabs of young dromedaries (Wernery, 2014, Wernery et al, 2015a, Bin et al, 2016), but has not been recovered from adult dromedaries. Over the last 3 years, several hundred samples have been tested at the Central Veterinary Research Laboratory, Dubai, for the presence of MERS-CoV at post-mortem examination from serologically positive MERS-CoV adult dromedaries greater than 4 years of age. Most of the dromedaries came from farms where MERS-CoV was isolated from the nasal cavity of young animals. However, no MERS-CoV has been recovered from any samples from adult dromedaries, including nasal swabs, tonsils, lungs, intestinal lymph nodes, mammary lymph nodes and milk.
Although isolating the virus will be useful for further research, such as antiviral susceptibility testing, diagnosis of MERS-CoV infection in dromedaries is mainly achieved through nucleic acid detection. Among all the nucleic acid detection technologies, the most widely used for MERS-CoV diagnosis is real-time quantitative reverse transcriptase (RT)-PCR (RT-qPCR), using a number of possible targets, including the RNA-dependent RNA polymerase, the region upstream to the envelope gene, or the nucleocapsid gene (Corman et al, 2012a, Corman et al, 2012b). RT-qPCR has high sensitivity and specificity for diagnosis of MERS-CoV infection.
Rapid diagnosis of MERS-CoV infections in dromedaries can also be achieved by direct antigen detection assays. In 2015, we published a monoclonal antibody-based ELISA for the detection of the nucleocapsid antigen of MERS-CoV in respiratory samples, with high sensitivity and specificity (Chen et al., 2015). Recently, we have used a lateral flow immunoassay platform for rapid detection of nucleocapsid antigen in respiratory samples of dromedaries; this has a sensitivity of ~80% and a specificity of 100% when compared to RT-qPCR (Chen et al., 2016). This technology has enabled the on-site screening of a large number of dromedaries in a short period of time. It also avoids the use of expensive equipment and does not require expertise in molecular diagnostics.
Seroepidemiology
The first evidence of dromedaries being the reservoir of MERS-CoV came from serological studies. High levels of MERS-CoV antibodies have been observed in dromedaries in the Middle East and Africa (Reusken et al, 2013, Corman et al, 2014a, Hemida et al, 2014, Meyer et al, 2014, Muller et al, 2014, Mackay, Arden, 2015). Serological follow-up of dromedary dams and their calves has showed a typical pattern of juvenile infection. Maternal antibodies against MERS-CoV in dromedary calves generally disappear between 4 and 8 months of age, permitting infection to occur during the seronegative period (Alagaili et al, 2014, Bin et al, 2016); young infected dromedaries then develop antibodies that persist for a long time. However, in a few cases, MERS-CoV has been isolated at the age of 20 days or even at younger, indicating that maternal antibodies are not necessarily protective. So far, the specific source of infection for young dromedaries is not known, although it is likely to be from other dromedaries. Extensive investigations in other animal species, including rodents, ticks, horses and small ruminants, have not demonstrated other reservoirs of infection to date.
Prevention and control
Protective experimental immunisations in dromedaries have already started using a modified vaccinia virus Ankara (MVA) vaccine expressing the MERS-CoV spike protein (Haagmans et al., 2016). Preliminary data showed a significant reduction in excretion of infectious virus and viral RNA in small numbers of vaccinated and challenged dromedaries compared to controls. Protection is correlated with the presence of serum neutralising antibodies against MERS-CoV. MVA-specific antibodies that cross-neutralise camelpox virus are another very important advantage of this vaccine, since outbreaks of camelpox still occur in dromedaries (Higgins et al., 1992). Another approach would be to add a MERS-CoV component to the already existing attenuated camelpox vaccine Ducapox. Since Ducapox has been used in the Middle East for many years, the acceptance of such a vaccine can be anticipated (Wernery et al., 2014). However, it is important for the success of a vaccine to adhere strictly to the exact time of vaccination, since the window of disappearance of maternal antibodies and appearance of antibodies as a result of infection is narrow.
MERS as a zoonotic disease
MERS is a zoonotic viral disease that can be transmitted from dromedaries to human beings. Several groups have performed seroepidemiological studies in different camel species in various countries and continents. So far, no antibodies to MERS-CoV have been detected in South American camelids or in Bactrian camels (Chan et al, 2015, Mackay, Arden, 2015). It seems that infection of dromedaries with MERS-CoV is limited to Africa and the Middle East (Perera et al, 2013, Reusken et al, 2013, Corman et al, 2014a, Hemida et al, 2014, Meyer et al, 2014, Muller et al, 2014). Over 90% of all dromedaries investigated from this region possess antibodies to the virus, some of them at least for more than 30 years (Muller et al., 2014). MERS-CoV infection in dromedaries is not a new entity and only very few young animals (<1%) may develop nasal discharge without other clinical signs. This picture changes when dromedaries are experimentally infected through the intranasal route (Haagmans et al., 2016).
In human beings, close contact with these young dromedaries may result in MERS-CoV infection, sometimes with fatal consequences. Although the mode of human-to-human transmission of MERS-CoV is not fully understood, the virus has caused major nosocomial outbreaks at hospitals in Saudi Arabia (Assiri et al, 2013, Azhar et al, 2014) and South Korea (Oboho et al., 2015). In South Korea, environmental contamination has been demonstrated, with MERS-CoV detected in patients' rooms, medical devices and air ventilating units, leading to closure of some hospitals (Cowling et al., 2015). It is worthwhile mentioning that no human MERS-CoV cases have been seen on camel dairy farms in the United Arab Emirates, even though the virus has been isolated regularly from dromedary calves.
Transmission from dromedaries to human beings is presumably uncommon, because the seroprevalence in human beings is low, even in people in frequent contact with dromedaries (Hemida et al., 2015). There may be several reasons for the low frequency of transmission from camels to human beings. Only young dromedaries with no or low maternal antibodies to MERS-CoV are susceptible to infection and the virus is shed only for 8 days (Wernery et al., 2015b). These young dromedaries are reared with their mothers for a year and have no or very little contact with human beings. Additionally, less than 1% of infected calves exhibit nasal discharge and therefore the quantity of virus excreted may be low. Another reason for the low transmission rate is likely to be the lack expression of the MERS-CoV receptor-dipeptidyl peptidase 4 (DPP4) in the human upper respiratory tract (Widagdo et al., 2016). Care must be taken when camel calves have to be treated. In these cases, caretakers and veterinarians should wear protective gear to avoid any transmission from camel calves to human beings.
In contrast to the monophylectic origin of SARS-CoV, MERS-CoV strains from human beings are polyphylectic as a result of multiple camel-to-human transmission events (Arabi et al, 2014, Ling et al, 2015). This implies that MERS-CoV has been present in dromedaries for many decades. It also provides an explanation why MERS has now persisted for more than 3 years, unlike SARS which disappeared rapidly after civets were removed by closure of animal markets in Southern China. MERS-CoV is likely to continue to be a threat to human beings unless vaccines are available.
Coronaviruses other than MERS-CoV in camelids
Coronaviruses are widespread and infect a broad range of species. An alphaCoV has been isolated from New World camelids (NWCs, also known as South American camelids) with respiratory disease (Jin et al., 2007). A betaCoV (bovine CoV-like CoV with no genome sequenced) was isolated from NWCs and dromedaries with diarrhoea (Crossley et al, 2010, Crossley et al, 2012). Transmission of the betaCoV occurs directly through close contact and aerosols, and the incubation period in crias is short (24–48 h). Disease in NWCs occurs in association with infection with rotavirus, E. coli, and Cryptosporidium and Salmonella spp. (Wernery et al., 2014). CoVs have been linked to several outbreaks of diarrhoea affecting NWCs of all ages on farms in north west USA and South America. Serological surveys and virus identification have also been carried out. CoVs were also detected in dromedary faecal samples and antibodies have been detected in dromedary sera in East Africa and on the Arabian Peninsula (Reusken et al, 2013, Meyer et al, 2014, Muller et al, 2014). However, there are no reports on CoV in Bactrian camels (Chan et al., 2015).
In 2014, we discovered a novel CoV named dromedary camel CoV UAE-HKU23 (DcCoV UAE-HKU23); the complete genome of this virus was sequenced directly from dromedary faecal samples and its phylogenetic relationship with other CoVs was determined (Table 1 ) (Wernery et al., 2014). We speculate that the bovine CoV-like CoV, with limited sequence information, described previously (Crossley et al, 2010, Crossley et al, 2012), is probably DcCoV UAE-HKU23. Phylogenetic analysis shows that CoV UAE-HKU23 is closely related to other CoVs of the Betacoronavirus 1 species in betaCoV lineage A, of which bovine CoV and human CoV OC43 are members (Woo et al., 2014d). Recently, we successfully isolated DcCoV UAE-HKU23 using the human rectal tumour HRT-18G cell line and confirmed that it is different from the betaCoV in alpacas using bioinformatics approaches (Woo et al., 2016b). Although both MERS-CoV and DcCoV UAE-HKU23 are beta CoVs, there is minimal cross antigenicity between them, as shown by various serological tests (Woo et al., 2014d). Last year, another alphaCoV that has been found in alpacas was also discovered in nasal samples of dromedaries (Table 1). The camelid alphaCoV, together with MERS-CoV and DcCoV UAE-HKU23, were observed to be co-circulating in camels in Saudi Arabia (Sabir et al., 2016).
Table 1.
Coronaviruses found in dromedaries.
| Viruses | Genera | Hosts | Geographical locations |
|---|---|---|---|
| Alpha CoV | Alphacoronavirus | Alpacas, llamas, dromedaries | USA, Saudi Arabia |
| DcCoV UAE HKU23 | Betacoronavirus (lineage A) | Dromedaries | UAE, Saudi Arabia |
| MERS-CoV | Betacoronavirus (lineage C) | Human beings, dromedaries | Saudi Arabia, Yemen, Oman, United Arab Emirates, Egypt, Jordan, Iran, Turkey, Greece, Italy, Germany, France, England, USA, Korea, China, Thailand, Malaysia |
Discussion
In the last 15 years, we have witnessed unprecedented changes in the CoV field. Two novel human CoVs (human CoV NL63 and human CoV HKU1) have been discovered, along with more than 40 novel animal CoVs (Lau et al, 2005, Lau et al, 2010, Lau et al, 2012, Lau et al, 2015, Woo et al, 2005, Woo et al, 2009, Woo et al, 2012, Woo et al, 2014a, van der Hoek et al, 2006, Chu et al, 2008, Mihindukulasuriya et al, 2008, Bermingham et al, 2012, Corman et al, 2014b). These include three novel lineages (B, C and D) of betaCoVs and a novel CoV genus (Deltacoronavirus) in the Coronavirus family. In addition, two large human CoV epidemics have affected numerous countries globally. The number of publications on CoVs in PubMed search has doubled in the last 10 years and a model of CoV evolution has been built. Due to the high recombination rate in CoVs and its unique mechanism of replication, it is expected that more and more ‘new’ CoVs will be generated. Some of these CoVs may have the capability to jump from one species to another and some of these interspecies transmission events may result in further major epidemics in human beings and animals.
In addition to CoVs, the MERS epidemic has also boosted interest in the discovery of other novel viruses in dromedaries. Since 2013, novel viruses of at least five different families (Astroviridae, Circoviridae, Hepeviridae, Picobirnaviridae and Picornaviridae) have been discovered in dromedaries (Woo et al, 2014b, Woo et al, 2014c, Woo et al, 2015, Woo et al, 2016a). Furthermore, West Nile virus has also been isolated from dromedaries (Joseph et al., 2016). Notably, shortly after the discovery of a novel genotype of hepatitis E virus in dromedaries in Dubai, the same virus was found in a human liver transplant recipient from the Middle East with chronic hepatitis E virus infection. The patient had the habit of regular consumption of camel meat and milk, suggesting that the virus may be transmitted from dromedaries to human beings (Lee et al., 2016). Both the transmission of MERS-CoV and hepatitis E virus from camels to human beings have shown that some viruses from dromedaries can be transmitted to human beings.
Conclusions
MERS-CoV is a zoonotic virus that can be transmitted from dromedaries to human beings, in which it causes severe respiratory disease with a high case fatality. Dromedaries develop no or only mild respiratory signs. In addition to MERS-CoV, a lineage B betaCoV and a virus in the Alphacoronavirus (alphaCoV) genus have been detected in dromedaries. More ‘new’ CoVs are likely to be generated due to the high recombination rate in this group of viruses. More intensive surveillance of CoVs in camels should be performed to improve understanding of these viruses in this unique group of animals, which have been closely associated with human beings for thousands of years.
Conflict of interest statement
None of the authors of this paper has a financial or personal relationship with other people or organisations that could inappropriately influence or bias the content of the paper.
Acknowledgements
This work is partly supported by the Theme-based Research Scheme (project number T11/707/15), University Grants Committee, Health and Medical Research Fund, University Development Fund and Strategic Research Theme Fund, University of Hong Kong. We thank Emily Y. M. Wong for her assistance in preparing the manuscript.
References
- Adney D.R., van Doremalen N., Brown V.R., Bushmaker T., Scott D., de Wit E., Bowen R.A., Munster V.J. Replication and shedding of MERS-CoV in upper respiratory tract of inoculated dromedary camels. Emerging Infectious Diseases. 2014;20:1999–2005. doi: 10.3201/eid2012.141280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adney D.R., Bielefeldt-Ohmann H., Hartwig A.E., Bowen R.A. Infection, replication, and transmission of Middle East respiratory syndrome coronavirus in alpacas. Emerging Infectious Diseases. 2016;22:1031–1037. doi: 10.3201/eid2206.160192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alagaili A.N., Briese T., Mishra N., Kapoor V., Sameroff S.C., Burbelo P.D., de Wit E., Munster V.J., Hensley L.E., Zalmout I.S. Middle East respiratory syndrome coronavirus infection in dromedary camels in Saudi Arabia. mBio. 2014;5:e00884-14. doi: 10.1128/mBio.00884-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arabi Y.M., Arifi A.A., Balkhy H.H., Najm H., Aldawood A.S., Ghabashi A., Hawa H., Alothman A., Khaldi A., Al Raiy B. Clinical course and outcomes of critically ill patients with Middle East respiratory syndrome coronavirus infection. Annals of Internal Medicine. 2014;160:389–397. doi: 10.7326/M13-2486. [DOI] [PubMed] [Google Scholar]
- Assiri A., McGeer A., Perl T.M., Price C.S., Al Rabeeah A.A., Cummings D.A., Alabdullatif Z.N., Assad M., Almulhim A., Makhdoom H. Hospital outbreak of Middle East respiratory syndrome coronavirus. New England Journal of Medicine. 2013;369:407–416. doi: 10.1056/NEJMoa1306742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azhar E.I., El-Kafrawy S.A., Farraj S.A., Hassan A.M., Al-Saeed M.S., Hashem A.M., Madani T.A. Evidence for camel-to-human transmission of MERS coronavirus. New England Journal of Medicine. 2014;370:2499–2505. doi: 10.1056/NEJMoa1401505. [DOI] [PubMed] [Google Scholar]
- Bermingham A., Chand M.A., Brown C.S., Aarons E., Tong C., Langrish C., Hoschler K., Brown K., Galiano M., Myers R. Severe respiratory illness caused by a novel coronavirus, in a patient transferred to the United Kingdom from the Middle East, September 2012. Euro Surveillance: Bulletin Européen sur les Maladies Transmissibles – European Communicable Disease Bulletin. 2012;17:20290. [PubMed] [Google Scholar]
- Bin S.Y., Heo J.Y., Song M.S., Lee J., Kim E.H., Park S.J., Kwon H.I., Kim S.M., Kim Y.I., Si Y.J. Environmental contamination and viral shedding in MERS patients during MERS-CoV outbreak in South Korea. Clinical Infectious Diseases. 2016;62:755–760. doi: 10.1093/cid/civ1020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan J.F., Chan K.H., Choi G.K., To K.K., Tse H., Cai J.P., Yeung M.L., Cheng V.C., Chen H., Che X.Y. Differential cell line susceptibility to the emerging novel human betacoronavirus 2c EMC/2012: Implications for disease pathogenesis and clinical manifestation. The Journal of Infectious Diseases. 2013;207:1743–1752. doi: 10.1093/infdis/jit123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan S.M., Damdinjav B., Perera R.A., Chu D.K., Khishgee B., Enkhbold B., Poon L.L., Peiris M. Absence of MERS-coronavirus in Bactrian camels, Southern Mongolia, November 2014. Emerging Infectious Diseases. 2015;21:1269–1271. doi: 10.3201/eid2107.150178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y., Chan K.H., Kang Y., Chen H., Luk H.K., Poon R.W., Chan J.F., Yuen K.Y., Xia N., Lau S.K. A sensitive and specific antigen detection assay for Middle East respiratory syndrome coronavirus. Emerging Microbes & Infections. 2015;4:e26. doi: 10.1038/emi.2015.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y., Chan K.H., Hong C., Kang Y., Ge S., Chen H., Wong E.Y., Joseph S., Patteril N.G., Wernery U. A highly specific rapid antigen detection assay for on-site diagnosis of MERS. The Journal of Infection. 2016;73:82–84. doi: 10.1016/j.jinf.2016.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu D.K., Peiris J.S., Chen H., Guan Y., Poon L.L. Genomic characterizations of bat coronaviruses (1A, 1B and HKU8) and evidence for co-infections in Miniopterus bats. The Journal of General Virology. 2008;89:1282–1287. doi: 10.1099/vir.0.83605-0. [DOI] [PubMed] [Google Scholar]
- Chu D.K., Poon L.L., Gomaa M.M., Shehata M.M., Perera R.A., Abu Zeid D., El Rifay A.S., Siu L.Y., Guan Y., Webby R.J. MERS coronaviruses in dromedary camels, Egypt. Emerging Infectious Diseases. 2014;20:1049–1053. doi: 10.3201/eid2006.140299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corman V.M., Eckerle I., Bleicker T., Zaki A., Landt O., Eschbach-Bludau M., van Boheemen S., Gopal R., Ballhause M., Bestebroer T.M. Detection of a novel human coronavirus by real-time reverse-transcription polymerase chain reaction. Euro Surveillance. 2012;17:20285. doi: 10.2807/ese.17.39.20285-en. [DOI] [PubMed] [Google Scholar]
- Corman V.M., Müller M.A., Costabel U., Timm J., Binger T., Meyer B., Kreher P., Lattwein E., Eschbach-Bludau M., Nitsche A. Assays for laboratory confirmation of novel human coronavirus (hCoV-EMC) infections. Euro Surveillance. 2012;17:20334. doi: 10.2807/ese.17.49.20334-en. [DOI] [PubMed] [Google Scholar]
- Corman V.M., Jores J., Meyer B., Younan M., Liljander A., Said M.Y., Gluecks I., Lattwein E., Bosch B.J., Drexler J.F. Antibodies against MERS coronavirus in dromedary camels, Kenya, 1992–2013. Emerging Infectious Diseases. 2014;20:1319–1322. doi: 10.3201/eid2008.140596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corman V.M., Kallies R., Philipps H., Gopner G., Muller M.A., Eckerle I., Brunink S., Drosten C., Drexler J.F. Characterization of a novel betacoronavirus related to Middle East respiratory syndrome coronavirus in European hedgehogs. Journal of Virology. 2014;88:717–724. doi: 10.1128/JVI.01600-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cowling B.J., Park M., Fang V.J., Wu P., Leung G.M., Wu J.T. Preliminary epidemiological assessment of MERS-CoV outbreak in South Korea, May to. Euro Surveillance. 2015;20:7–13. doi: 10.2807/1560-7917.es2015.20.25.21163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crossley B.M., Barr B.C., Magdesian K.G., Ing M., Mora D., Jensen D., Loretti A.P., McConnell T., Mock R. Identification of a novel coronavirus possibly associated with acute respiratory syndrome in alpacas (Vicugna pacos) in California, 2007. Journal of Veterinary Diagnostic Investigation. 2010;22:94–97. doi: 10.1177/104063871002200118. [DOI] [PubMed] [Google Scholar]
- Crossley B.M., Mock R.E., Callison S.A., Hietala S.K. Identification and characterization of a novel alpaca respiratory coronavirus most closely related to the human coronavirus 229E. Viruses. 2012;4:3689–3700. doi: 10.3390/v4123689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eckerle I., Corman V.M., Muller M.A., Lenk M., Ulrich R.G., Drosten C. Replicative capacity of MERS coronavirus in livestock cell lines. Emerging Infectious Diseases. 2014;20:276–279. doi: 10.3201/eid2002.131182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haagmans B.L., van den Brand J.M., Raj V.S., Volz A., Wohlsein P., Smits S.L., Schipper D., Bestebroer T.M., Okba N., Fux R. An orthopoxvirus-based vaccine reduces virus excretion after MERS-CoV infection in dromedary camels. Science. 2016;351:77–81. doi: 10.1126/science.aad1283. [DOI] [PubMed] [Google Scholar]
- Hemida M.G., Perera R.A., Al Jassim R.A., Kayali G., Siu L.Y., Wang P., Chu K.W., Perlman S., Ali M.A., Alnaeem A. Seroepidemiology of Middle East respiratory syndrome (MERS) coronavirus in Saudi Arabia (1993) and Australia (2014) and characterisation of assay specificity. Euro Surveillance. 2014;19:20828. doi: 10.2807/1560-7917.es2014.19.23.20828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hemida M.G., Al-Naeem A., Perera R.A., Chin A.W., Poon L.L., Peiris M. Lack of middle East respiratory syndrome coronavirus transmission from infected camels. Emerging Infectious Diseases. 2015;21:699–701. doi: 10.3201/eid2104.141949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higgins A.J., Silvey R.E., Abdelghafir A.E., Kitching R.P. The epidemiology and control of an outbreak of camelpox in Bahrain. In: Allen W.R., Higgins A.J., Mayhew I.G., Snow D.H., Wade J.F., editors. Proceedings of the 1st International Camel Conference, Dubai, United Arab Emirates, 2–6 February 1992. R&W Publications; Newmarket, UK: 1992. pp. 100–104. [Google Scholar]
- Jin L., Cebra C.K., Baker R.J., Mattson D.E., Cohen S.A., Alvarado D.E., Rohrmann G.F. Analysis of the genome sequence of an alpaca coronavirus. Virology. 2007;365:198–203. doi: 10.1016/j.virol.2007.03.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joseph S., Wernery U., Teng J.L., Wernery R., Huang Y., Patteril N.A., Chan K.H., Elizabeth S.K., Fan R.Y., Lau S.K. First isolation of West Nile virus from a dromedary camel. Emerging Microbes & Infections. 2016;5:e53. doi: 10.1038/emi.2016.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau S.K., Woo P.C., Li K.S., Huang Y., Tsoi H.W., Wong B.H., Wong S.S., Leung S.Y., Chan K.H., Yuen K.Y. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proceedings of the National Academy of Sciences of the United States of America. 2005;102:14040–14045. doi: 10.1073/pnas.0506735102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau S.K., Poon R.W., Wong B.H., Wang M., Huang Y., Xu H., Guo R., Li K.S., Gao K., Chan K.H. Coexistence of different genotypes in the same bat and serological characterization of Rousettus bat coronavirus HKU9 belonging to a novel Betacoronavirus subgroup. Journal of Virology. 2010;84:11385–11394. doi: 10.1128/JVI.01121-10. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Lau S.K., Woo P.C., Yip C.C., Fan R.Y., Huang Y., Wang M., Guo R., Lam C.S., Tsang A.K., Lai K.K. Isolation and characterization of a novel Betacoronavirus subgroup A coronavirus, rabbit coronavirus HKU14, from domestic rabbits. Journal of Virology. 2012;86:5481–5496. doi: 10.1128/JVI.06927-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau S.K., Woo P.C., Li K.S., Tsang A.K., Fan R.Y., Luk H.K., Cai J.P., Chan K.H., Zheng B.J., Wang M. Discovery of a novel coronavirus, China Rattus coronavirus HKU24, from Norway rats supports the murine origin of Betacoronavirus 1 and has implications for the ancestor of Betacoronavirus lineage A. Journal of Virology. 2015;89:3076–3092. doi: 10.1128/JVI.02420-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee G.H., Tan B.H., Chi-Yuan Teo E., Lim S.G., Dan Y.Y., Wee A., Aw P.P., Zhu Y., Hibberd M.L., Tan C.K. Chronic infection with camelid hepatitis E virus in a liver transplant recipient who regularly consumes camel meat and milk. Gastroenterology. 2016;150:355–357. doi: 10.1053/j.gastro.2015.10.048. [DOI] [PubMed] [Google Scholar]
- Ling Y., Qu R., Luo Y. Clinical analysis of the first patient with imported Middle East respiratory syndrome in China. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue. 2015;27:630–634. doi: 10.3760/cma.j.issn.2095-4352.2015.08.002. in Chinese. [DOI] [PubMed] [Google Scholar]
- Mackay I.M., Arden K.E. MERS coronavirus: Diagnostics, epidemiology and transmission. Virology Journal. 2015;12:222. doi: 10.1186/s12985-015-0439-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer B., Muller M.A., Corman V.M., Reusken C.B., Ritz D., Godeke G.J., Lattwein E., Kallies S., Siemens A., van Beek J. Antibodies against MERS coronavirus in dromedary camels, United Arab Emirates, 2003 and 2013. Emerging Infectious Diseases. 2014;20:552–559. doi: 10.3201/eid2004.131746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mihindukulasuriya K.A., Wu G., St Leger J., Nordhausen R.W., Wang D. Identification of a novel coronavirus from a beluga whale by using a panviral microarray. Journal of Virology. 2008;82:5084–5088. doi: 10.1128/JVI.02722-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muller M.A., Corman V.M., Jores J., Meyer B., Younan M., Liljander A., Bosch B.J., Lattwein E., Hilali M., Musa B.E. MERS coronavirus neutralizing antibodies in camels, Eastern Africa, 1983–1997. Emerging Infectious Diseases. 2014;20:2093–2095. doi: 10.3201/eid2012.141026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oboho I.K., Tomczyk S.M., Al-Asmari A.M., Banjar A.A., Al-Mugti H., Aloraini M.S., Alkhaldi K.Z., Almohammadi E.L., Alraddadi B.M., Gerber S.I. 2014 MERS-CoV outbreak in Jeddah – A link to health care facilities. New England Journal of Medicine. 2015;372:846–854. doi: 10.1056/NEJMoa1408636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perera R.A., Wang P., Gomaa M.R., El-Shesheny R., Kandeil A., Bagato O., Siu L.Y., Shehata M.M., Kayed A.S., Moatasim Y. Seroepidemiology for MERS coronavirus using microneutralisation and pseudoparticle virus neutralisation assays reveal a high prevalence of antibody in dromedary camels in Egypt, June 2013. Euro Surveillance : Bulletin Européen sur les Maladies Transmissibles = European Communicable Disease Bulletin. 2013;18:20574. doi: 10.2807/1560-7917.es2013.18.36.20574. [DOI] [PubMed] [Google Scholar]
- Reusken C.B., Haagmans B.L., Muller M.A., Gutierrez C., Godeke G.J., Meyer B., Muth D., Raj V.S., Smits-De Vries L., Corman V.M. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: A comparative serological study. The Lancet. Infectious Diseases. 2013;13:859–866. doi: 10.1016/S1473-3099(13)70164-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saad M., Omrani A.S., Baig K., Bahloul A., Elzein F., Matin M.A., Selim M.A., Al Mutairi M., Al Nakhli D., Al Aidaroos A.Y. Clinical aspects and outcomes of 70 patients with Middle East respiratory syndrome coronavirus infection: A single-center experience in Saudi Arabia. International Journal of Infectious Diseases. 2014;29:301–306. doi: 10.1016/j.ijid.2014.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sabir J.S., Lam T.T., Ahmed M.M., Li L., Shen Y., Abo-Aba S.E., Qureshi M.I., Abu-Zeid M., Zhang Y., Khiyami M.A. Co-circulation of three camel coronavirus species and recombination of MERS-CoVs in Saudi Arabia. Science. 2016;351:81–84. doi: 10.1126/science.aac8608. [DOI] [PubMed] [Google Scholar]
- van Boheemen S., de Graaf M., Lauber C., Bestebroer T.M., Raj V.S., Zaki A.M., Osterhaus A.D.M.E., Haagmans B.L., Gorbalenya A.E., Snijder E.J. Genomic characterization of a newly discovered coronavirus associated with acute respiratory distress syndrome in humans. mBio. 2012;3:e00473-12. doi: 10.1128/mBio.00473-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Hoek L., Pyrc K., Berkhout B. Human coronavirus NL63, a new respiratory virus. FEMS Microbiology Reviews. 2006;30:760–773. doi: 10.1111/j.1574-6976.2006.00032.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wernery U. Some epidemiological studies on MERS Coronavirus in dromedaries in the United Arab Emirates. Journal of Camel Practice and Research. 2014;21:1–4. [Google Scholar]
- Wernery U., Kinne J., Schuster R.F. World Organization for Animal Health/Office International des Épizooties (OIE); Paris, France: 2014. Camelid Infectious Diseases; pp. 283–295. [Google Scholar]
- Wernery U., Corman V.M., Wong E.Y., Tsang A.K., Muth D., Lau S.K., Khazanehdari K., Zirkel F., Ali M., Nagy P. Acute Middle East respiratory syndrome coronavirus infection in livestock dromedaries, Dubai, 2014. Emerging Infectious Diseases. 2015;21:1019–1022. doi: 10.3201/eid2106.150038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wernery U., Rasoul I.E., Wong E.Y., Joseph M., Chen Y., Jose S., Tsang A.K., Patteril N.A., Chen H., Elizabeth S.K. A phylogenetically distinct Middle East respiratory syndrome coronavirus detected in a dromedary calf from a closed dairy herd in Dubai with rising seroprevalence with age. Emerging Microbes & Infections. 2015;4:e74. doi: 10.1038/emi.2015.74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Widagdo W., Raj V.S., Schipper D., Kolijn K., van Leenders G.J., Bosch B.J., Bensaid A., Segalés J., Baumgärtner W., Osterhaus A.D.M.E. Differential expression of the Middle East respiratory syndrome coronavirus receptor in the upper respiratory tracts of humans and dromedary camels. Journal of Virology. 2016;90:4838–4842. doi: 10.1128/JVI.02994-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C., Lau S.K., Chu C.M., Chan K.H., Tsoi H.W., Huang Y., Wong B.H., Poon R.W., Cai J.J., Luk W.K. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. Journal of Virology. 2005;79:884–895. doi: 10.1128/JVI.79.2.884-895.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C., Lau S.K., Lam C.S., Lai K.K., Huang Y., Lee P., Luk G.S., Dyrting K.C., Chan K.H., Yuen K.Y. Comparative analysis of complete genome sequences of three avian coronaviruses reveals a novel group 3c coronavirus. Journal of Virology. 2009;83:908–917. doi: 10.1128/JVI.01977-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C., Lau S.K., Lam C.S., Lau C.C., Tsang A.K., Lau J.H., Bai R., Teng J.L., Tsang C.C., Wang M. Discovery of seven novel Mammalian and avian coronaviruses in the genus Deltacoronavirus supports bat coronaviruses as the gene source of Alphacoronavirus and Betacoronavirus and avian coronaviruses as the gene source of Gammacoronavirus and Deltacoronavirus. Journal of Virology. 2012;86:3995–4008. doi: 10.1128/JVI.06540-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C., Lau S.K., Lam C.S., Tsang A.K., Hui S.W., Fan R.Y., Martelli P., Yuen K.Y. Discovery of a novel bottlenose dolphin coronavirus reveals a distinct species of marine mammal coronavirus in Gammacoronavirus. Journal of Virology. 2014;88:1318–1331. doi: 10.1128/JVI.02351-13. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Woo P.C., Lau S.K., Teng J.L., Tsang A.K., Joseph M., Wong E.Y., Tang Y., Sivakumar S., Bai R., Wernery R. Metagenomic analysis of viromes of dromedary camel fecal samples reveals large number and high diversity of circoviruses and picobirnaviruses. Virology. 2014;471–473:117–125. doi: 10.1016/j.virol.2014.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C., Lau S.K., Teng J.L., Tsang A.K., Joseph M., Wong E.Y., Tang Y., Sivakumar S., Xie J., Bai R. New hepatitis E virus genotype in camels, the Middle East. Emerging Infectious Diseases. 2014;20:1044–1048. doi: 10.3201/eid2006.140140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C., Lau S.K., Wernery U., Wong E.Y., Tsang A.K., Johnson B., Yip C.C., Lau C.C., Sivakumar S., Cai J.P. Novel betacoronavirus in dromedaries of the Middle East, 2013. Emerging Infectious Diseases. 2014;20:560–572. doi: 10.3201/eid2004.131769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C., Lau S.K., Teng J.L., Tsang A.K., Joseph S., Xie J., Jose S., Fan R.Y., Wernery U., Yuen K.Y. A novel astrovirus from dromedaries in the Middle East. The Journal of General Virology. 2015;96:2697–2707. doi: 10.1099/jgv.0.000233. [DOI] [PubMed] [Google Scholar]
- Woo P.C., Lau S.K., Choi G.K., Huang Y., Wernery R., Joseph S., Wong E.Y., Elizabeth S.K., Patteril N.A., Li T. Equine rhinitis B viruses in horse fecal samples from the Middle East. Virology Journal. 2016;13:94. doi: 10.1186/s12985-016-0547-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C., Lau S.K., Fan R.Y., Lau C.C., Wong E.Y., Joseph S., Tsang A.K., Wernery R., Yip C.C., Tsang C.C. Isolation and characterization of dromedary camel coronavirus UAE-HKU23 from dromedaries of the Middle East: Minimal serological cross-reactivity between MERS coronavirus and dromedary camel coronavirus UAE-HKU23. International Journal of Molecular Sciences. 2016;17:E691. doi: 10.3390/ijms17050691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- World Health Organization (WHO) Middle East respiratory syndrome coronavirus (MERS-CoV) – Saudi Arabia. 2016. http://www.who.int/csr/don/16-may-2016-mers-saudi-arabia/en/ (accessed 16 June 2016)
- Zaki A.M., van Boheemen S., Bestebroer T.M., Osterhaus A.D.M.E., Fouchier R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. New England Journal of Medicine. 2012;367:1814–1820. doi: 10.1056/NEJMoa1211721. [DOI] [PubMed] [Google Scholar]
- Zhou J., Chu H., Chan J.F., Yuen K.Y. Middle East respiratory syndrome coronavirus infection: Virus-host cell interactions and implications on pathogenesis. Virology Journal. 2015;12:218. doi: 10.1186/s12985-015-0446-6. [DOI] [PMC free article] [PubMed] [Google Scholar]