

Abstract
Veterinary vaccines currently available in Europe and in other parts of the world are developed by the veterinary pharmaceutical industry. The development of a vaccine for veterinary use is an economic endeavour that takes many years. There are many obstacles along the path to the successful development and launch of a vaccine. The industrial development of a vaccine for veterinary use usually starts after the proof of concept that is based on robust academic research. A vaccine can only be made available to the veterinary community once marketing authorisation has been granted by the veterinary authorities.
This review gives a brief description of the regulatory requirements which have to be fulfilled before a vaccine can be admitted to the market. Vaccines have to be produced in a quality controlled environment to guarantee delivery of a product of consistent quality with well defined animal and consumer safety and efficacy characteristics. The regulatory and manufacturing legislative framework in which the development takes place is described, as well as the trend in developments in production systems. Recent developments in bacterial, viral and parasite vaccine research and development are also addressed and the development of novel adjuvants that use the expanding knowledge of immunology and disease pathology are described.
Keywords: Vaccine, Industry, Regulatory, Good manufacturing practice (GMP)
Introduction
The industrial development of a veterinary vaccine starts once the proof of concept has been established. Proof of concept is based on robust academic exploration of fundamental science. The scientific literature has reported many academic proofs of various novel vaccine concepts in recent years. Industrial vaccine development programs (see Table 1 for its phases) starts once this proof of concept has been demonstrated and should eventually lead to market authorisation (MA), i.e. the permission to place a vaccine on the market, which is granted by the appropriate veterinary authorities.
Table 1.
Research an development phases for a vaccinea
| Documentary | Feasibility | Pre-development | Development | Registration | Commercial |
|---|---|---|---|---|---|
| Product profile | Antigen selection | Prototype batch | Pilot batches | Registration dossier compilation and submission | Sales and marketing |
| Formulation | Formulation | Licensing | Pharmacovigilance | ||
| Experimental batch | Production process | Production process validation | |||
| Control tests | Control test validation | ||||
| Proof of concept | Dose–effect (safety and efficacy) | Safety studies, Efficacy studies | |||
| Pre-stability studies | Stability studies | ||||
| Field trials | |||||
|
Table with minor modifications adapted from Soulebot et al. (1997).
In contrast to academic proof of concept demonstrations, the industrial development of a vaccine should be seen in an economic context. Veterinary vaccines are produced and marketed by pharmaceutical companies which are in competition with one another. For veterinary vaccines there are two main target markets that can be recognised each with its own dynamics. These markets are (1) the agricultural or production animal sector and (2) companion animals.
In recent decades the general trend has been that the agricultural sector has become more industrial and price sensitive. In contrast, the public perception of companion animals has changed leading to increased demand for medicinal care. In general, vaccines for production animals are produced in larger quantities at low cost, whereas vaccines for companion animals are produced in lower quantities and sold at higher prices. Moreover, it should be realised that it is difficult for profit-making enterprises to justify the development of vaccines for diseases of low incidence or vaccines for species that are kept in low numbers (Schetters, 1995).
Apart from operational costs there is the expense of complying with the legal and regulatory requirements for MA and manufacturing. Regulatory requirements are under continuous review and vaccines which could be registered 20 years ago may not be registrable today, owing to changes in the regulatory framework. Furthermore, regulatory requirements are sometimes difficult to fulfil for a particular vaccine concept. Vaccines are produced on an industrial scale in government certified manufacturing facilities. The Good Manufacturing Practice (GMP) requirements, which have been incorporated into law, describe in detail the manufacturing standard. Since the introduction of veterinary medicinal product licensing in Europe some 25 years ago, the economic environment, the increasing legal and regulatory pressure and the changed agricultural scenery have lead to a marked consolidation among the veterinary vaccine manufacturers.
In this review, we first outline the regulatory and manufacturing legal context in the EU. Second we discuss developments in production methods, and third we describe concepts of viral, bacterial and parasitic vaccines and immune potentiators.
Regulatory and manufacturing legislation
Vaccines for animals, like all other medicinal products have to be licensed by the relevant authorities who have to ensure that the medicine is of adequate quality and purity, that it is safe and that it works in the target species as claimed for the indication for which it is intended (Jones et al., 2007; Schetters and Gravendyck, 2006).
Licensing of veterinary vaccines was formally introduced in the EU in 1981 (CD 81/851/EEC, 1981, CD 81/852/EEC, 1981, CD 90/677/EEC, 1990, CD 92/18/EEC, 1991, Eudralex, XXX, Eudralex, XXX 1 Ph. Eur, 2005). All the information and test results required by the guidelines (CD 81/851/EEC, 1981, CD 81/852/EEC, 1981, CD 90/677/EEC, 1990, CD 92/18/EEC, 1991, Eudralex, XXX, Eudralex, XXX, Ph. Eur, 2005), which demonstrate quality, purity, safety and efficacy are compiled in a dossier. The MA is based on this dossier after review and further questioning.
The gathering of all the required information and test results on the vaccine (vaccine development) is usually organised in structured and well phased manner. The various development phases that can be distinguished are summarised in Table 1 and a flow chart of the key stages of the development is given in Fig. 1 . Regulations may of course change during the course of the development and may have an impact on development time and cost.
Fig. 1.
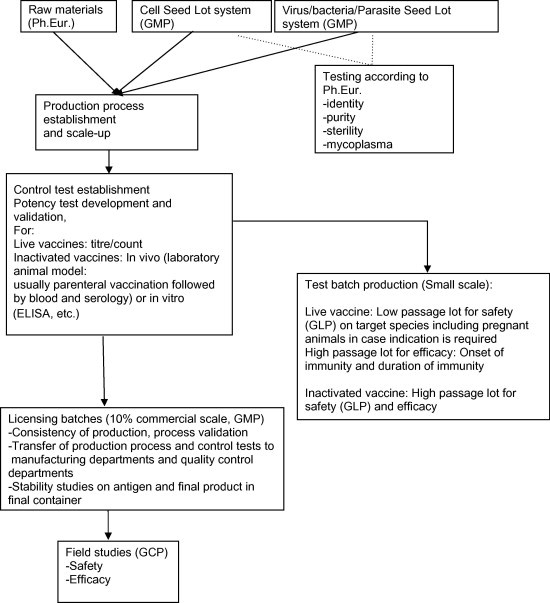
Flow diagram for vaccine development.
Before marketing the vaccine, a manufacturing license and a GMP certificate of each facility involved in the manufacturing of the vaccine are required, in addition to a MA license. Here we describe briefly quality, safety, efficacy and GMP (manufacturing) requirements for veterinary vaccines.
Quality
To ensure consistent quality, the production method and technology underlying a vaccine must operate in compliance with the principles of current GMP, which is monitored by a Quality Assurance system (CD 91/412/EEC, 1991, Eudralex, XXX, Eudralex, XXX). Vaccine production under GMP is required by law (CD 91/412/EEC, 1991, Eudralex, XXX, Eudralex, XXX).
The quality of starting materials used for production is assured by testing each batch purchased or is made to ensure that it meets acceptance limits. Additional requirements exist for starting materials of biological origin, which are commonly used for vaccine production (e.g. ensuring exclusion of extraneous agents). Seed stocks of the vaccine strain and production cell-lines are laid down in a seed lot system, making passages from the Master Seed to establish a bank of Working Seed from which all production batches are produced. The initial Master Seed must have a known history of its isolation and previous passages to minimise the risk of transmission of transmissible spongiform encephalopathies (TSEs). In addition, the purity (absence of extraneous agents such as bacteria, fungi, mycoplasma and viruses) and identity of vaccine antigen needs to be demonstrated.
To ensure that each batch of a commercial vaccine is equivalent in quality, the manufacturer must register all relevant in process tests as well as tests to be performed on the finished product, giving limits of acceptance that must be met before the batch can be released for sale. In addition, the manufacturer must prove that the quality of the vaccine when produced on a (semi) commercial scale, is guaranteed until the end of its shelf life based on at least three batches of vaccine in the final container.
Safety
Safety is of paramount importance. The determination of safety is fundamentally a firmness of purpose that the benefits of the product outweigh any potential risks, not only to the target species being vaccinated, but also to the administrator of the vaccine, the environment, the food derived from treated animals from which food is derived, and the consumer.
Experimental data obtained with batches with the highest potency or titre (see below) must be generated in specially designed experiments carried out under the Good Laboratory Practice (GLP) regimen (CD 92/18/EEC, 1991, OECD, 2002 2). In addition to the safety of a single dose, the safety of an overdose and repeated doses of the vaccine must be shown (such as injection site reaction, clinical signs). Safety studies have to be designed in accordance with the recommended schedules and using susceptible target species.
There are additional special requirements for live vaccines, such as (genetic) stability (i.e. the vaccine should not revert to virulence during consecutive passages) and the risks of genetic recombination and/or genomic re-assortment should be minimal. Also studies on the immunological functions or reproductive capacity may have to be carried out depending on the nature of the vaccine, adjuvant, or disease. Additional requirements have to be met for vaccines containing genetically modified organisms (GMO).
Efficacy and potency
A product must be able to do what is claimed on the label (e.g. reduce virus shedding, limit typical clinical signs, disease) Preferably, these data are obtained from field trials performed under Good Clinical Practice-Veterinary (VICH-GL9) 3 conditions and in laboratory studies in which, if possible, validated experimental challenge models are used.
Onset and duration of immunity, for instance a few days (onset) to 1 year (duration) followed by a yearly vaccination to sustain the level of immunity, must be supported by experimental data. These data must show that 1 year after the primary vaccination, animals are still satisfactorily protected and also that animals that receive a single booster vaccination 1 year after initial vaccination are still protected 1 year later.
Confirmation of the protective effect of a specific antigen is usually established by vaccination-challenge experiments. This protective effect is correlated to alternative laboratory animal or in vitro tests which are carried out on each batch of finished vaccine. The pass level for batch release is set at the minimum level that was shown to be efficacious in the target animal (correlate of protection). If the correlate of protection is unknown, it may take years before an accurate potency test can be developed. Moreover the manufacturer must provide data that guarantee the immunising capability and thereby the protective effect of a product over its entire shelf life.
Label claim
In the EU, each MA is granted with an approved summary of product characteristics (SPC) describing the pharmaceutical form of the product, the (categories of) target animal, contra-indications, recommended vaccination schedule and route of administration. Moreover, the SPC contains recommendations for use such as withdrawal period, use with other products and advice for minimisation of the risk of concurrent infections during the vaccination period, as these could interfere with the induction of the active immune response.
Commercial production under GMP
Since licensing of veterinary vaccines was formally introduced in the EU in 1981 it has been a requirement that both the active ingredient (antigen) and the finished product (vaccine) must be manufactured according to GMP (CD 91/412/EEC, 1991, CD 92/18/EEC, 1991, Eudralex, XXX, Eudralex, XXX). Apart from validated methods and tests, a preventive maintenance system should be in-place for all critical equipment and buildings. Responsibilities of key personnel and key departments have to be documented in policy documents. Trend analysis of different parameters (Utilities, Environmental Monitoring) and In Process Controls (IPC) should provide useful information about process performance and quality. In general, all tests and processes must be validated and reported including buildings and equipment. Moreover, every one of these quality related systems, procedures and policies have to be written down in a Site Master File.
Even during commercial production it can be necessary to make changes to the processes or to the tests that have been developed. With a full operational GMP quality system it is still possible to optimise the process and to make significant improvement changes. A Change Control system ensures that all changes are formally requested, documented and approved. The impact of these changes on the marketing authorisation and the validation status of the process need to be determined and documented in detail as this may have regulatory consequences.
Moreover, a vaccine producer must employ at least one Qualified Person who is, without prejudice to his relationship with the holder of the manufacturing authorisation, personally responsible for release of vaccine onto the market (CD 81/851/EEC, 1981, CD 91/412/EEC, 1991, CD 2001/83/EC, 2001). Regular internal and external audits by competent authorities aim at surveying the quality control procedures in-place described here.
Production systems
The increasing regulatory, GMP and economic burden imposed on vaccine manufacturers necessitate the development of reliable, large-scale antigen production methods, such as the use of cell culture substrates. In recent years, the quest for highly productive cell culture processes has been focused on the front-end to continuous cell-line development rather than the use of primary cells such as chicken embryo fibroblasts (CEF). The improvement in protein expression systems, high throughput screening methods and cell characterisation techniques have enabled the development of more productive and stable cell-lines used for antigen production. Final product quality and consumer safety is already considered at cell-line development, i.e. at the very early stage of a vaccine development project. The development of and transition to animal-component-free culture media for cells, the identification of suitable animal-free replacements for certain components such as bovine serum, and, ultimately, the development of fully chemically defined culture media, are examples of this consideration.
With an increasing need for higher antigen yields in combination with shorter product development and production process times, sophisticated production process development methods are required in every stage of the process from early development to scale-up to large volumes. The current generation of miniature bioreactors, with working volumes of a few millilitres, modelling large-scale cell culture processes, provide the capability to examine different conditions within a single experiment. This offers the opportunity to understand the process at the cellular level and enables better and more predictable extrapolation to large-scale bioreactor processes. The establishment of analytical technologies enabling in-line monitoring of antigen production has the advantage of studying important immunogenic antigen properties such as glycosylation or cell metabolites influencing overall growth of cells and consequently antigen and contributes to the consistency, reliability and scalability of entire processes.
Early effort to produce proteins expressed by bacteria, yeast, and mammalian cells in commercial quantities required costly re-usable hardware. This involved an extensive cleaning and re-sterilisation processes and the validation of these processes as required by the rules set-out in GMP regulations. This longstanding approach placed heavy demands on standard operating procedures (SOPs), and usually resulted in high costs and extensive personnel training.
The most significant costs involved in manufacturing are facility time (that can represent up to 55% of the total manufacturing costs of a plant) and validation (that can account for between 10% and 20% of a plant cost). These costs can be reduced through increasing facility throughput in terms of number of campaigns or runs per year, increasing the antigen yield per run and installing equipment that allows plants to operate both as multi-purpose and multi-product production facilities.
An option that is adopted more and more by biotech manufactures in an effort to reduce costs is moving to disposable manufacturing. “Use once, throw away” technology is used more and more by many manufacturers in various stages of production processes. It results in improvement in throughput by reducing the amount of downtime between campaigns, and it also allows the development of a multi-purpose plant design because of the flexibility in use of disposable systems in existing clean rooms. Furthermore, the disposable individual components or whole systems can eliminate the long lead times in the initial manufacturing and installation of stainless steel equipment. The design of a disposable system is primarily dictated by the application and usually custom made.
Key factors to take into consideration during the design phase of such an application may be the volume to be processed, chemical compatibility with the product, the number of production runs per year, processing conditions (as pressure, temperature, flow rates, mixing times), sensitivity of the product to extractable materials, which are present in all plastics, and validation support from the vendors. Currently, the most widely used components include containers, filtration, connections, clamps, bioreactors and tubing. Nowadays many of the compounds in a biotechnology production suite can be supplied both in hard piped stainless steel or disposable systems. If components are already supplied pre-assembled and sterilised, significant reductions will be made in contamination risks, validation efforts and labour time. Pre-assembled disposable components dramatically increase the manufacturer’s dependence on its supplies and may jeopardise sustainable delivery of vaccine product (Table 2 ).
Table 2.
Comparison of disposable versus stainless steel production equipment
| Advantages | Disadvantages | |
|---|---|---|
| Stainless steel | Robust | Less flexible |
| Low operating cost | High installation cost | |
| Limited supplier dependency | Cleaning and steaming required | |
| Accepted material quality standard | Validations (e.g. cleaning) | |
| All types of equipment available (USP and DSP) | ||
| No extractables | ||
| Disposable | Flexible | Extractables |
| Low start-up cost | Supplier dependency | |
| Low down time between cycles | Higher raw material cost | |
| Limited validation required | Not all types of equipment available (DSP and probes) | |
| TSE compliance | ||
| No accepted material quality standard |
As already indicated, disposable systems do contain extractables (components of the disposable material leaking into the product stream), which may interfere with antigen production or product safety. Manufacturers of disposable systems undertake extractable testing using model solvents as part of their own validation protocols. However, system users must generate their own validation data to demonstrate that extractables do not adversely affect the product being stored or processed within or by the disposable. This is done with a worst-case exposure where the conditions are based on known characteristics of the product and the model solvent covering the entire disposable assembly. Moreover, plastics are not, by definition, animal component-free and disposables also have to comply with TSE guidelines (Table 2).
The use of disposable technology still requires hardware and control panels that need initial validation and maintenance. Some elements of current manufacturing processes, such as centrifugation, large-scale chromatography and large-scale tangential flow filtration systems are not yet available as disposable options. In-line or real time monitoring of production processes is still in a development stage and disposable sensor technology is also not yet available. This means cleaning in-place or discarding expensive re-usable sensors, or avoiding sensors all together, which makes the processes uncontrollable (Table 2).
Recent improvements in chromatography and TFF equipment, aseptic fluid transfer devices, as well as improvements on plastic film strength and resistance will lead eventually to the replacement of traditional stainless steel GMP manufacturing components with single-use disposable formats. Thinking even further ahead, the development of a totally disposable manufacturing system can be imagined. When such disposable factories may become commonplace will depend largely on new disposable product developments and evolution of regulatory and GMP guidelines. The industry is adapting to these developments as shown by the introduction of short-term disposable mixing and monitoring systems.
Bacterial vaccines
Bacterial vaccines are mainly inactivated whole cell cultures (or bacterins), inactivated culture supernatants (or toxoids), crude extracts of the cell surface or attenuated live preparations. Advances in our knowledge and biological technologies give the opportunity to develop improved vaccines.
The inactivated crude preparations had the advantage that knowledge of the molecular basis of pathogenesis was not a prerequisite to vaccine development since there was a good chance they contained the antigens necessary to stimulate protective immunity. The disadvantages were the greater potential for antigenic competition and/or diversion so that immune responses to protective antigens (that were nonetheless present in the preparations) failed, and that they usually contained lipopolysaccharide, techoic acid or other cell wall constituents. These components led to the heavy stimulation of the innate immune systems so that undesirable adverse effects were likely. Nonetheless, the traditional approaches remain a perfectly acceptable and an acceptable solution for certain diseases (Andre-Fontaine et al., 2003).
The early live vaccines had the advantages of the inactivated antigens combined with the potential for in vivo dependent gene expression to provide a more protective immune stimulus (Feberwee et al., 2001). The non-specific or unstable genetic alterations inherent in these vaccines could lead to suboptimal growth for adequate immune stimulus (i.e. over attenuated) or insufficient attenuation to prevent adverse reactions and reversions to virulence (i.e. under attenuated and in a reversible fashion).
Later, vaccine development took advantage of new studies of the molecular basis of bacterial pathogenesis. Initially these depended on physicochemical separations of bacterial components and assessment of the interaction of these with isolated host tissues, cells and biochemistry. In addition, the effects of immunisations with purified preparations on the outcome of experimental challenges, and of the epidemiological distribution of the efficacious components amongst clinical and non-clinical isolates (Moon and Bunn, 1993), were taken into account. These studies heralded the introduction of the first so-called subunit vaccines.
Later still the introduction of molecular cloning enabled the genetic isolation of characterised protective immunogens and in particular the construction of host/vector combinations with enhanced expression of the recombinant antigen to provide higher yields of less reactive vaccine components more cheaply (Petersen et al., 1991). Commercially available vaccines developed by these methods were first introduced only in comparatively recent times.
The arrival of whole bacterial genome sequencing and bioinformatic analyses (prediction of structure, function and cellular location from sequence) and the development of micro-array analysis of whole genome expression are bringing about a revolution in the design and development of bacterial vaccines. This is particularly the case for those diseases where the traditional approaches have not been efficacious (Scarselli et al., 2005). In simple terms, with all of the genes hypothetically identified for a bacterial pathogen it is possible to construct an array of sites on a solid sheet each containing bound synthetic DNA complementary to all of the genes in the genome. The messenger RNA (mRNA) extracted from the bacteria living in different environments is used to prepare probes in strengths proportionate to the amounts of each mRNA species present at the time of extraction. By detecting differences in the strength of probe hybridisation to the gene array it is possible to see which gene’s levels of expression have changed relative to the different environments.
Genes upregulated in vivo, or in cultures designed to mimic in vivo environments, that bioinformatic analyses indicate are likely to be surface expressed or secreted during critical stages of host colonisation or pathogenesis (and hence accessible to interference from host immunity) enable the identification of attractive candidate vaccine antigens without necessarily knowing their function. It is then possible to delete the gene(s) in question to confirm their role in pathogenesis in challenge models and also to produce recombinant purified antigens to use in vaccination trials. Provided the antigens are reasonably conserved amongst clinical isolates this strategy can provide a powerful route to efficacious vaccine development where crude preparations in the past have not achieved a desirable level of efficacy or safety. This technology now makes it possible to create live vaccines incapable of reversion to virulence, with minimal interference in the expression of important protective immunogens whilst inactivating powerful immune diverting immunogens that may interfere with the strength of efficacious responses.
DNA vaccination is a relatively recent consideration where genes encoding protective immunogens are inserted into non-replicating elements able to be taken up by host cells and initially express the foreign antigen intracellularly and then presented on the cell surface in association with MHC antigens to T cells. This offers the advantages (1) that the Th1/Th2 balance of immune responses can be modulated to achieve more effective immunity and (2) that DNA is highly stable at room temperature and easily quality assurance (QA) controlled during production (Jechlinger, 2006). Unfortunately, for many projects the payload of DNA required has been prohibitively expensive and new technologies are needed along the lines of adjuvants that enhance responses to antigens.
One of the ironies of the molecular genomic approach is that protective subunit antigens and DNA vaccines are often poor immunogens because they are not associated with other bacterial components present in crude preparations that non-specifically enhance immune responses, that is to say with inherent adjuvant properties. Consequently, not only are new adjuvant strategies needed to make recombinant subunit or DNA vaccines work, but bacteria may themselves provide the solution. Most intensively investigated as adjuvants are cholera and Escherichia coli heat labile ADP-ribosylating enterotoxins (Lycke, 2004), the zonula occludens toxin of Vibrio cholerae (De Magistris, 2006), Mycobacterium tuberculosis heat shock protein 70 (Bulut et al., 2005), Salmonella Typhimurium FljB (Simon and Samuel, 2007), synthetic analogs of bacterial lipoproteins (Ghielmetti et al., 2005), synthetic oligodeoxynucleotides containing unmethylated CpG dinucleotides (McCluskie and Krieg, 2006, Gomis et al., 2007) and monophosphoryl lipid A (Jiang et al., 2007).
Veterinary bacterial vaccine development can still call upon perfectly valid traditional techniques but we have only just started the era where bacterial behaviour from the gene to genome will allow us to predict what we need to do for successful control of disease. It is only a matter of time before efficacious vaccination could be a reality for almost any bacterial disease for which there is a market.
Viral vaccines
Alpha herpesviruses
Viruses belonging to this subfamily cause significant diseases in horses, pigs, cattle and poultry. Genomes of several members in the herpesvirus subfamily have been sequenced and molecular virology has played a significant role in the identification of virulence-associated, virus encoded genes and their functions (Wittmann and Rziha, 1989, Mettenleiter, 1991, Kimman et al., 1992). This has led to the development of effective and safe, conventional as well as biotechnological marker vaccines (Kit et al., 1987, Marchioli et al., 1987, Quint et al., 1987, Pensaert et al., 1992, Van Oirschot, 2000).
The deletion mutants’ biotech approach has been particularly successful for pseudorabies virus (PRV) and bovine herpesvirus 1 (BHV-1). In contrast, for equine herpesvirus-1 and -4 (EHV-1; EHV-4), deletion of non-essential virus glycoprotein and enzyme genes with the aim of deriving attenuated live virus vaccine candidates, similar to PRV and BHV-1, have been fruitless (Patel and Heldens, 2005). There is, however, a clear need for improved EHV-1 vaccines, particularly to protect against EHV-1 induced abortion and paresis and also to reduce the incidence and therefore the transmission of EHV-1 and EHV-4 by unweaned passively immune foals (Patel and Heldens, 2005). In this regard, a highly promising experimental EHV-1 vaccine was a temperature-sensitive (ts) strain cloned from a classically mutagenised stock of an abortigenic EHV-1 isolate (Patel et al., 2003a, Patel et al., 2003b, Patel et al., 2004).
Marek’s disease virus (MDV) occurs worldwide and is the cause of significant loss in chickens. MDV has been developed as a vector for Newcastle disease virus glycoprotein recombinant vaccine. Effective commercial Marek’s disease virus vaccines for poultry contain cell-associated, low passage variants of the original Rispens isolate. Quests to generate efficacious and safe cell-free MDV vaccines, using conventional or biotech approaches have been unsuccessful so far despite the veterinary medicinal and logistic need for such vaccine.
Circoviruses
Porcine circovirus-2 (PCV-2) is a cause of multi-organ disease and significant economic loss in domestic pigs (Allan and Ellis, 2000, Krakowa et al., 2002). Immunoprophylaxis against PCV-2 infection of domestic pigs is with conventional and biotech vaccines, such as a baculovirus expressed PCV-ORF2 protein. Additionally, there have been many biotech experimental PCV-2 vaccines investigated. Examples of such vaccines are PCV-2 ORF 2 gene expressed in PRV vector, an apathogenic PCV-1/PCV-2 chimera vaccine, and DNA plasmid vaccines (Fenaux et al., 2004, Kamstrup et al., 2004, Chunmei et al., 2005).
Flaviviruses
Routine prophylactic vaccination against classical swine fever virus (CSFV) in the EU ceased in 1990 (Westergaard, 2000). Outside the EU, conventional vaccines as well as biotech CSFV-E2 glycoprotein subunit marker vaccine are widely used (Lin and Lee, 1981, Van Oirschot, 2000, De Smit et al., 2000). Several biotech approaches have been investigated for CSFV vaccines and include baculovirus expressed E2 glycoprotein (Ahrens et al., 2000), chimeric bovine virus diarrhoea virus (BVDV)–CSFV vaccine (van Gennip et al., 2000) and recombinant PRV expressing CSFV-E2 glycoprotein that was protective for diseases due to PRV and CSFV (Van Zijl et al., 1991). Some of these biotech vaccines are experimental but they allow serological discrimination of vaccinated and infected animals, allowing the use of CSFV vaccination, where previously it has been banned.
Application of biotech approaches for protective BVDV vaccines include glycoprotein E2 expression and production in baculovirus and defective BVDV replicons. No biotech BVDV vaccines are currently available in the EU but there is a choice of live and killed conventional vaccines (Straub, 2002). However, only three killed vaccines claim to afford protection against BVDV abortion (Brownlie et al., 1996, Patel et al., 2002, Salt et al., 2004).
Over the last decade West Nile virus (WNV), which causes significant disease in horses and other animal species including man, has become a potent threat. Currently biotech vaccines marketed for use in horses are a canarypox virus vectored live vaccine and a chimerical WNV glycoprotein and yellow fever virus backbone vaccine (Monath, 2001).
Influenza and paramyxoviruses
Influenza A viruses cause greater problems in birds than mammals. In particular, some strains of H5 and H7 subtypes are highly pathogenic causing high mortality and pose a zoonotic risk to man along with some H9 subtypes. In the EU, vaccination against avian influenza is discouraged, but the emphasis is changing (Capua and Alexander, 2006). The likely approach to develop vaccines would be, first, the cloning and site directed mutagenesis to turn the HA-gene into a non-pathogenic form, and, second, the production of so-called high growth re-assortants producing considerable amounts of the new HA protein, which is, among others, the protective antigen in influenza virus. The latter can be achieved by reverse genetics and transfection techniques (Wood and Robertson, 2006). This has been the approach for the currently circulating and potentially pandemic H5N1 avian influenza virus, which has caused more than 250 human deaths so far.
Other biotech approaches, such as the cloning of the changed HA-gene into other viral vectors such as Newcastle disease virus (NDV) can be carried out as well. Reverse genetics has been used to alter the cleavage site of the NDV-F protein, but so far conventional live and killed vaccines dominate the markets. Non-replicating vector avipoxvirus approaches, such as canarypox virus expressing canine distemper virus HF glycoprotein gene have been on the market in the USA since 1997.
Modern biotech vaccine approaches do not always give better vaccines, however. For avian influenza H5 for instance, it has been shown in the field that classically grown, inactivated vaccines formulated with potent adjuvants can offer cross-protection against related pathogenic strains (Swayne et al., 2006).
Iridoviruses and orbiviruses
African swine fever (ASF) is a highly contagious fatal disease of pigs caused by an iridovirus (ASFV). An effective vaccine is clearly needed and there are indications that this is possible (Mettraux et al., 1992). Commercial development of the approach has however never been undertaken for economic reasons or has been unsuccessful for reasons such as manufacturing difficulties.
African horse sickness (AHS) is a highly fatal, insect transmitted disease of equidae caused by African horse sickness virus (AHSV), an orbivirus. Currently, conventional mouse brain or tissue culture grown vaccines are in use but future vaccines may use AHSV VP2 protein as a subunit biotech vaccine (Ranz et al., 1992). As is the case for ASFV, commercial development of the approach has never been undertaken for economic reasons or has been unsuccessful owing to manufacturing difficulties.
For bluetongue virus (BTV), whilst there are egg-adapted polyvalent vaccines to prevalent serotypes in use, biotech approaches have been investigated which hold much promise. Thus BTV-like and virus core-like structures have been constructed using major (VP3, VP7) and minor (VP1, VP4, VP5) capsid proteins. The VP proteins were produced in baculovirus multiple expression vectors. These virus-like single and double shelled particles emulsified in Freund’s incomplete adjuvant or Montanide ISA-50 adjuvants were highly immunogenic and protective for naı¨ve sheep (Roy, 1992). The approach holds promise since baculovirus expression vectors for different BTV serotypes could be prepared in advance and stored in a bank on similar lines to foot and mouth disease virus (FMDV; Westergaard, 2000) and brought out when an outbreak occurs.
Rabies virus
Conventional rabies virus vaccines, mostly killed, for immunising various mammalian species are in common use. Rabies virus is one of the examples where biotech approaches have or are likely to make good progress. The latter is well exemplified by the attempts to reduce the incidence of wildlife rabies, mostly in the red fox (Vulpes vulpes) in mainland Europe. A replicating recombinant vaccinia virus expressing rabies virus glycoprotein has been highly effective in reducing the incidence of wildlife rabies in mainland Europe (Kihm et al., 1992, Pastoret and Brochier, 1996, Westergaard, 2000).
Retroviruses and viruses without vaccines
Monovalent and subunit feline leukaemia virus (FeLV) glycoprotein expressed in E. coli is marketed in the EU. Immunogenicity of an experimental vaccinia virus recombinant that expresses bovine leukaemia virus envelope protein (gp51) has given promising results in bovine calves and rabbits (Valikhov et al., 1997).
It is important and relevant to point out that there are animal diseases for which there are no vaccines at present. The diseases affect domesticated and wild ruminants and deer for instance. Other wildlife diseases, currently unknown or undetected, may be transmitted to domesticated animals and possibly to man. Recent notable examples of animal viruses crossing into man are of retroviruses like HIV, SARS coronavirus, West Nile virus and avian influenza H5N1 subtype virus.
The gamma herpesviruses named alcephaline herpes virus-1 (AHV-1) and ovine herpesvirus-2 (OHV-2) cause fatal excessive lymphoid proliferation in secondary dead-end secondary ruminant hosts, namely cattle and deer species (Nettleton et al., 1988, Reid and Buxton, 1989, Plowright, 1990, Hussey et al., 2000). AHV-1 and OHV-2 are innocuous in reservoir hosts, wildebeest and sheep, respectively. Two retroviruses, one belonging to subfamily lentivirus (visna-maedi virus and caprine arthritis/encephalitis virus) and one to the subfamily oncovirus (sheep pulmonary adenomatosis virus), cause significant diseases in sheep, for which no vaccines are available. Similarly, it is not possible at present to control some other veterinary retroviruses such as bovine leukaemia, avian type C oncovirus and reticuloendotheliosis virus.
Two new fatal zoonotic infections due to related paramyxoviruses, named Hendra and Nipah viruses, affect horses and pigs, respectively, and also man. The reservoir for both viruses are petropid bats found in Australia and the Far East (Haplin et al., 2000). Currently no vaccines are available for these viruses.
Parasite vaccines
Classically, the most important parasitic diseases in humans and animals are treated and/or controlled by using chemotherapeutics (Cornelissen and Schetters, 1996). A series of developments (including drug resistance) has given impetus to the research into parasite vaccines. As a result, a number of vaccines against parasitic diseases are now commercially available. It is envisioned that more parasite vaccines will come to market and will aid in the control of parasitic diseases (Vercruysse et al., 2004).
In many parasitic diseases, the host develops some level of immunity once the infection is cured. This indicates that the parasite has sufficient immunogenic potential that could be exploited as a vaccine. Research aims at the development of a vaccine and/or vaccination protocol that induces protective immunity while limiting the induction of pathology. Different approaches can be recognized varying from the use of live vaccines, attenuated live vaccines, killed vaccines and subunit vaccines.
Live vaccines (complete life-cycles)
Transient infections
The most obvious examples of such a vaccine are the live vaccines against coccidiosis in chickens (Williams, 2002). As this type of infection is transient (the parasite “passes” through the chicken) the infection is self-limiting and no chemotherapeutic treatment is necessary to cure the infection. To limit the induction of pathology it is necessary that a defined low-dose is given to the animals, and that this infection is initiated simultaneously in all chickens from a flock.
Selection for less virulent strains
The virulence of parasite strains derived from a single isolate can be variable. For example, using Babesia bovis isolates, passage through splenectomised animals can select for strains of reduced virulence. Such parasite strains are being used to vaccinate cattle in Africa and Australia. The infection that develops is less virulent and the animals develop immunity against subsequent challenge infection (De Waal and Combrink, 2006). These vaccines are distributed by government institutions.
Similarly, strains with reduced virulence can be selected from Eimeria isolates. Some commercially available coccidiosis vaccines for broilers contain strains that are selected after repeated passage through chickens. These so-called precocious strains require less time to develop into oocysts, and the numbers of progeny are reduced compared to the wild-type parent population (Williams, 1994).
Temperature-sensitive strains
Some parasite strains have been selected that differ from the wild-type strains in that they cause a self-limiting infection. One such example is the temperature-sensitive strain of Toxoplasma gondii. This strain, which resulted from chemically induced attenuation, can be propagated successfully in vitro at relatively low temperatures, but will not propagate successfully at the body temperature of the target animal. As a result the infection will self-cure (Lindsay et al., 1993). This vaccine has not been commercialised.
Infection treatment
In case the parasite has a tendency to survive in the host for longer periods of time, chemotherapeutic cure of the infection is also required. An example of this approach is the live vaccine against Theileria parva infection. The vaccine is based on isolates of virulent T. parva strains which are used to infect cattle that are simultaneously treated with a long-acting tetracycline preparation to control the infection (Boulter and Hall, 1999). This method is still being used in Africa, and the vaccine is produced by Centre for Ticks and Tick borne Diseases (CTTBD) in Malawi.
Live vaccines (incomplete life-cycles)
Many parasite species have complicated life-cycles characterised by distinct life-cycle stages, sometimes involving more than one host. In some cases the early life-cycle stages are sufficiently immunogenic to induce protective immunity; selection for parasite strains with truncated life-cycles is another strategy to develop vaccines. A good example is the T. gondii S48 strain. This strain has lost the capacity to develop from the tachyzoite into the bradyzoite stage, and thus does not form tissue cysts. The tachyzoites induce a transient infection in the host, while triggering protective immune reactions (Buxton, 1993).
Irradiation of parasites has also been used as a mechanism to truncate the life-cycle. The live vaccine against lungworm infection in cattle contains L3 larvae of Dictyocaulus viviparus that do not develop further than the L4 stage. Vaccinated cattle are immune to challenge with L3 larvae (Urquhart, 1985).
Theoretically, virulent parasite strains could be genetically modified to reduce their virulence. For instance, parasite strains could be genetically manipulated such that during the production of the vaccine, parasites would be fully virulent and when administered to the host would cause a self-limiting infection. This principle has been investigated using Toxoplasma gondii in which a tetracycline-dependent regulatory element was cloned in front of an essential gene. The parasite may be propagated during the vaccine production phase in the presence of tetracycline. Once injected into the target animal, the parasite will not be able to continue propagation in the absence of tetracycline, as this will lead to blocking of the expression of the essential gene (Van Poppel et al., 2006).
Killed vaccines
Whole organisms
If no live vaccine strains are available, or the use of live vaccines is undesirable, one may want to inactivate the parasites prior to the formulation of a vaccine. Such preparations by themselves do not induce protective immunity and an appropriate adjuvant and formulation must be developed. Examples of such vaccines are the vaccine against abortion in cattle due to Neospora caninum infection (Schetters, 2004) and a vaccine against giardiasis in dogs (Olson et al., 2000).
Subunit vaccines
A more detailed analysis of the immune response acquired after natural infection or vaccine induced immunity, can lead to the discovery of critical antigenic components of an organism that can be used in a vaccine. Again such preparations require an adjuvant for the induction of protective immunity. The vaccine against babesiosis of dogs due to Babesia canis infection is one such example (Schetters, 2005). It contains soluble antigens secreted/excreted from the parasite. Research has shown that during B. canis infection in dogs these antigens are released and cause disease. Vaccination helps animals to quickly produce neutralising antibodies against these antigens. The adjuvant appears critical: saponin works, whereas oil-based adjuvants have shown little or no efficacy (Schetters, 2005).
In some cases the antigens are produced using recombinant DNA technology. The best example is a vaccine against Taenia ovis in sheep, which is based on recombinantly produced parasite antigens that induce antibodies that block the attachment of oncospheres to the gut epithelium (Harrison et al., 1999). Saponin adjuvant was shown to be most efficacious. Another example is the vaccine against the cattle tick Boophilus microplus (Willadsen, 2004). The vaccine contains recombinantly produced gut wall antigens of the tick. Upon vaccination of cattle, high levels of antibodies to the gut wall of ticks are produced. During feeding of the tick on the vaccinated animal these antibodies are ingested and destroy the gut epithelium of the tick thus killing the parasite.
Vaccine immunopotentiators or adjuvants
In recent times, immunopotentiators received abundant attention in the media as critical adjuvants in novel human vaccines. Examples include the prophylactic vaccines against human papilloma virus (HPV), novel pandemic influenza virus, as well as experimental allergy and tumour vaccines. Indeed, vaccine adjuvants, also referred to as major platform technologies, are recognised to make the difference between competing vaccines based on identical antigens. In addition, it is recognised that vaccines designed for certain diseases require a matching combination of selected antigen(s) together with a critical immunopotentiator that selectively drives the essential immune pathway with minimal adverse reactions.
The increased awareness of immunopotentiator importance is prominent in particular among human vaccinologists. For veterinary vaccine designers, the application and importance of adjuvants has always been evident. Traditional veterinary vaccines, consisting of relatively crude extracts of microbial cultures, are inherently more immunogenic relative to purified subunits or peptide antigens, which are preferred in human vaccines. Obviously, the constitution of the antigen of interest may contribute to immunogenicity and overall level of immune responsiveness.
Another trend in recent years is an increasing scientific knowledge of mechanistic activities of many immunopotentiators, especially as a result of research focused on innate immunity receptors. These insights enable more rational adjuvant and vaccine design, which, ideally, is based on predictable immunophenotypes following vaccination. Schijns, 2000, Schijns, 2001, Schijns, 2002, Schijns, 2003, Schijns and Tangeras, 2005, Schijns, 2006, Schijns et al., in press have highlighted recent developments in immunopotentiators.
Mechanistically, vaccine adjuvants can be classified in two major groups: (1) vaccine delivery systems that facilitate the timing, dosing and geography of the antigen, – also called facilitators of signal 1, and (2) signal 2 facilitators, which directly activate certain (innate) immune cells (Degen et al., 2003). Vaccine adjuvants also influence the quality of the immune response, since they may preferably instruct for the development of T helper (Th)1, Th2, Th3, Th17, or regulatory T cells (T reg), a stimulus originally referred to as signal 3 (Kapsenberg et al., 1999).
Immunopotentiators make the vaccine work, that is they evoke and potentiate antigen-specific T and B cell responses to both poorly immunogenic subunit vaccines and crude antigen preparations, respectively. They largely determine the immunophenotype of the response to the antigen in the vaccine. For example, vaccine adjuvants are able to influence and accelerate the onset of immunity, which may become urgent during emergency vaccination, or at early age (post-hatch or after birth) when the offspring is naı¨ve to microbial attack.
Vaccine adjuvants may also increase the overall magnitude of the antigen-specific response in order to reach a minimal level of protective antibody concentration or effector T cell population. In addition, adjuvants can prolong the duration of vaccine effector immune responses, allowing for fewer or no booster immunisations, which becomes critical during mass vaccinations. Moreover, certain vaccine adjuvants are able to positively affect the quality of the immunophenotype, for example induction of cell-mediated T cell immunity, considered necessary for control of many types of intracellular pathogens. Antigen dose (cost) sparing is enabled by certain adjuvants, which is relevant in case of expensive or cumbersome antigen production systems.
Importantly, immunopotentiators may cause transient unwanted adverse reactions, either at the injection site or systemically. The level of acceptance depends on the relative benefit of the vaccine and the medical need to prevent or treat the disease of interest. Unfortunately, there is no single adjuvant for all needs. Instead, there are many different choices of potential immunopotentiators. Rational selection of vaccine adjuvants during vaccine design is hampered by limited knowledge of the immunophenotype evoked by most classical vaccine adjuvants.
Selection of the best immunopotentiator has long been a matter of trial-and-error and serendipity. Fortunately, in recent times mechanistic studies based on new insights of especially innate immunity receptors have allowed for more systematic and scientific investigations on immune induction in general and immunopotentiation in particular. This not only holds for mice and humans (the best studied immune systems) but increasingly for species of veterinary relevance. This new knowledge will allow for the development of new and improved immunopotentiator-based prophylactic vaccines as well as novel therapeutic vaccines.
Discussion
Vaccines play an important role in the control of animal diseases, particularly in the food producing industries. However, this is currently largely achieved by conventionally developed and produced vaccines. Progress on veterinary vaccine development in industry is largely influenced by boundaries set by regulatory, GMP and economic constraints, as outlined above. For obvious reasons regulatory requirements are strict, and will be further strengthened, to ensure delivery of safe and efficacious vaccines. GMP and GLP quality systems were developed to achieve these ends. From the proof of concept of a new vaccine, these QA systems are implemented at all stages of vaccine development. In early phases only seed material is produced under a GMP regimen whereas in the final stages, a fully operational GMP system is applied.
The burden placed on vaccine manufacturers caused them to develop reliable and flexible large-scale production methods. The development of more cell culture systems, animal-component-free culture media, multi-purpose facilities and disposable manufacturing materials are clear examples of this trend. Against this background we have dealt with the current trends in industry, namely the use of biotechnology in vaccine development and production. Examples of the success of biotech and classical vaccine approaches have been given for bacterial, viral and parasitic vaccines.
With respect to virus diseases, a separate review dealing in more detail with epidemiology, pathogenesis and vaccines available or in development has been written (Patel and Heldens, accepted for publication). This will give a more complete and comprehensive overview of vaccine options in development and marketing. As indicated, the number of biotechnology-based vaccines presently on the market or in late stage development against bacterial, viral or parasitic pathogens is rather limited, but it is expected that products based on biotechnology/bio-engineering will increase in the future.
Knowledge of the working mechanism of adjuvants is increasing, based on progress in understanding animal diseases, immunology and adjuvant working mechanisms. This allows more rational vaccine design. Notwithstanding the fact that the majority of current veterinary vaccines are derived and produced by conventional attenuation and/or inactivation processes in tissue culture, genetic engineering techniques have been widely used and will be used even more in the future but they have yet to fulfil the promise of improved vaccines. It is important to realise that there is no perfect vaccine and there may never be since pathogens are constantly mutating in order to survive and to evade their host’s defences.
Basic academic research to increase our understanding of pathogen behaviour must be an ongoing activity so that we can improve our ability to protect the health and welfare of both humans and animals. To accomplish this we must continue and improve our vaccines that can be made in an industrial manner.
Acknowledgements
The secretarial support from Hanny van Lare for typing (sometimes hand-written) manuscripts and for editing (hand-written) suggestions for improvement to the paper is acknowledged.
Footnotes
References
- Ahrens U., Kaden V., Drexler C., Visser N. Efficacy of the classical swine fever (CSF) marker vaccine Porcilis pesti in pregnant sows. Veterinary Microbiology. 2000;77:83–97. doi: 10.1016/s0378-1135(00)00265-0. [DOI] [PubMed] [Google Scholar]
- Allan G.M., Ellis J.A. Porcine circoviruses: a review. Journal of Veterinary Diagnostic Investigation. 2000;12:3–14. doi: 10.1177/104063870001200102. [DOI] [PubMed] [Google Scholar]
- Andre-Fontaine G., Branger C., Gray A.W., Klaasen H.L. Comparison of the efficacy of three commercial bacterins in preventing canine leptospirosis. Veterinary Record. 2003;153:165–169. doi: 10.1136/vr.153.6.165. [DOI] [PubMed] [Google Scholar]
- Boulter N., Hall R. Immunity and vaccine development in the bovine theileriosis. Advances in Parasitology. 1999;44:41–97. doi: 10.1016/s0065-308x(08)60230-4. [DOI] [PubMed] [Google Scholar]
- Brownlie J., Clarke M.C., Hooper L.B., Bell G.D. Protection of the bovine foetus from BVDV infection by means of a new inactivated vaccine. Veterinary Record. 1996;137:582–586. doi: 10.1136/vr.137.3.58. [DOI] [PubMed] [Google Scholar]
- Bulut Y., Michelsen K.S., Hayrapetian L., Naiki Y., Spallek R., Singh M., Arditi M. Mycobacterium tuberculosis heat shock proteins use diverse Toll-like receptor pathways to activate pro-inflammatory signals. Journal of Biological Chemistry. 2005;280:20961–20967. doi: 10.1074/jbc.M411379200. [DOI] [PubMed] [Google Scholar]
- Buxton D. Toxoplasmosis: the first commercial vaccine. Parasitology Today. 1993;9:335–337. doi: 10.1016/0169-4758(93)90236-9. [DOI] [PubMed] [Google Scholar]
- Capua I., Alexander D.J. The challenge of avian influenza to the veterinary community. Avian Pathology. 2006;35:189–205. doi: 10.1080/03079450600717174. [DOI] [PubMed] [Google Scholar]
- CD 2001/83/EC, 2001. Directive 2001/83/EC of the European Parliament and of the Council of 6 November 2001 on the Community code relating to medicinal products for human use (O.J. No. L311 of 28.11.2001).
- CD 81/851/EEC, 1981. Directive 81/851/EEC of 28 September 1981 on the approximation of the laws of the member states relating to veterinary medicinal products (O.J. No. L317 of 6.11.81).
- CD 81/852/EEC, 1981. Council Directive 81/852/EEC of 28 September 1981 on the approximation of the laws of the Member States relating to analytical, pharmacotoxicological and clinical standards and protocols in respect of the testing of veterinary medicinal products (O.J. No. L317 of 6.11.81).
- CD 90/677/EEC, 1990. Council Directive 90/677/EEC of 13 December 1990 extending the scope of Directive 81/851/EEC on the approximation of the laws of the Member States relating to veterinary medicinal products and laying down additional provisions for immunological veterinary medicinal products (O.J. No. L373 of 31.12.90).
- CD 91/412/EEC, 1991. Directive 91/412/EC of the European Parliament and of the Council of 23 July 1991 laying down the principles and guidelines of Good Manufacturing Practice for veterinary medicinal products (O.J. No. L228 of 17.08.91).
- CD 92/18/EEC, 1991. Commission directive 92/18/EC of 20 March 1992 modifying de Annex to Council Directive 81/852/EEC on the approximation of the laws of Member States relating to analytical, pharmacotoxicological and clinical standards and protocols in respect of the testing of veterinary medicinal products (O.J. No. L97/1 of 10.4.92).
- Chunmei J., Fan H., Tan Y., Liu Z., Xi X., Cao S., Wu B., Chen H. Immunogenicity of a recombinant pseudorabies virus expressing ORF1-ORF2 fusion protein of circovirus type 2. Veterinary Microbiology. 2005;109:179–190. doi: 10.1016/j.vetmic.2005.06.001. [DOI] [PubMed] [Google Scholar]
- Cornelissen A.W.C.A., Schetters Th.P.M. Vaccines against protozoal diseases of veterinary importance. FEMS Immunology and Medical Microbiology. 1996;15:61–72. doi: 10.1111/j.1574-695X.1996.tb00055.x. [DOI] [PubMed] [Google Scholar]
- Degen W.G., Jansen T., Schijns V.E. Vaccine adjuvant technology: from mechanistic concepts to applications. Expert Review Vaccines. 2003;2(2):327–335. doi: 10.1586/14760584.2.2.327. [DOI] [PubMed] [Google Scholar]
- De Magistris M.T. Zonula occludens toxin as a new promising adjuvant for mucosal vaccines. Vaccine. 2006;24(Suppl. 2) doi: 10.1016/j.vaccine.2005.01.123. S2-60-1. [DOI] [PubMed] [Google Scholar]
- De Smit, A.J., Van de Wetering, G., Colijn, E.C., Hulst, M., Kramps, J.A., Van den Blink, A., Moormann, R.J.M., 2000. The detection of antibodies against the ERNS envelope protein of classical swine fever virus. In: Brocchi, E., Lavaza, A. (Eds.), fifth International Congress of Veterinary Virology, Istituto Zooprofillatico Sperimentale della Lombardia e dell Emilia Romagna, Brescia Fondazione Iniziative Zooprofilattiche e Zootechniche, Brescia, Italy, pp. 414–415.
- De Waal D.T., Combrink M.P. Live vaccines against babesiosis. Veterinary Parasitology. 2006;138(1–2):88–96. doi: 10.1016/j.vetpar.2006.01.042. [DOI] [PubMed] [Google Scholar]
- Eudralex. The Rules governing Medicinal Products in the European Union, vol. IV, Good Manufacturing Practice for medicinal products (EC Catalog number CO-71-91-760-EN-C).
- Eudralex. The Rules governing Medicinal Products in the European Union, vol. VII, Guidelines for the testing of Veterinary Medicinal Products (EC Catalog number CO-86-94-383-EN-C).
- Feberwee A., de Vries T.S., Hartman E.G., de Wit J.J., Elbers A.R., de Jong W.A. Vaccination against Salmonella enteritidis in Dutch commercial layer flocks with a vaccine based on a live Salmonella gallinarum 9R strain: evaluation of efficacy, safety, and performance of serologic Salmonella tests. Avian Diseases. 2001;45:83–91. [PubMed] [Google Scholar]
- Fenaux M., Opriessnig T., Halbur P.G., Elvinger F., Meng X.J. A chimeric porcine circovirus (PCV) with the immunogenic capsid gene of the pathogenic PCV type 2 (PCV2) cloned into the genomic backbone of the nonpathogenic PCV1 induces protective immunity against PCV2 infection in pigs. Journal of Virology. 2004;78:6297–6303. doi: 10.1128/JVI.78.12.6297-6303.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghielmetti M., Reschner A., Zwicker M., Padovan E. Synthetic bacterial lipopeptide analogs: structural requirements for adjuvanticity. Immunobiology. 2005;210:211–215. doi: 10.1016/j.imbio.2005.05.015. [DOI] [PubMed] [Google Scholar]
- Gomis S., Babiuk L., Allan B., Willson P., Waters E., Hecker R., Potter A. Protection of chickens against a lethal challenge of Eschericia coli by a vaccine containing CpG oligodeoxynucleotides as an adjuvant. Avian Diseases. 2007;51:78–83. doi: 10.1637/0005-2086(2007)051[0078:POCAAL]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Haplin K., Young P.L., Field H.E., MacKenzie J.S. Isolation of Hendra virus from petropid bats: a natural reservoir of Hendra virus. Journal of General Virology. 2000;81:1927–1932. doi: 10.1099/0022-1317-81-8-1927. [DOI] [PubMed] [Google Scholar]
- Harrison G.B., Shakes T.R., Robinson C.M., Lawrence S.B., Heath D.D., Dempster R.P., Lightowlers M.W., Rickard M.D. Duration of immunity, efficacy and safety in sheep of a recombinant Taenia ovis vaccine formulated with saponin or selected adjuvants. Veterinary Immunology and Immunopathology. 1999;20:161–172. doi: 10.1016/s0165-2427(99)00039-2. [DOI] [PubMed] [Google Scholar]
- Hussey, D., Mueller-Doblies, U., Stauber, N., Hauzer, B., Braun, U., Ackermann, M., 2000. In: Brocchi, E., Lavaza, A. (Eds.), fifth International Congress of Veterinary Virology, Istituto Zooprofillatico Sperimentale della Lombardia e dell Emilia Romagna, Brescia Fondazione Iniziative Zooprofilattiche e Zootechniche, Brescia, Italy, pp. 105–106.
- Jechlinger W. Optimization and delivery of plasmid DNA for vaccination. Expert Reviews in Vaccines. 2006;5:803–825. doi: 10.1586/14760584.5.6.803. [DOI] [PubMed] [Google Scholar]
- Jiang Z.H., Budzynski W.A., Qiu D., Yalamati D., Koganty R.R. Monophosphoryl lipid A analogues with varying 3-O-substitution: synthesis and potent adjuvant activity. Carbohydrate Research. 2007;342:784–796. doi: 10.1016/j.carres.2007.01.012. [DOI] [PubMed] [Google Scholar]
- Jones P.G.H., Cowan G., Gravendyck M., Nagata T., Robinson S., Waits M. Regulatory requirements for vaccine authorization. Revue Scientifique Et Technique De L Office International Des Epizooties. 2007;26(2):379–392. [PubMed] [Google Scholar]
- Kamstrup S., Barfoed A.M., Frimann T.H., Ladekjaer-Mikkelsen A.S., Botner A. Immunisation against PCV2 structural protein by DNA vaccination of mice. Vaccine. 2004;22:1358–1361. doi: 10.1016/j.vaccine.2004.01.032. [DOI] [PubMed] [Google Scholar]
- Kapsenberg M.L., Hilkens C.M., Wierenga E.A., Kalinski P. The paradigm of type 1 and type 2 antigen-presenting cells. Implications for atopic allergy. Clinical and Experimental Allergy. 1999;29(Suppl. 2):33–36. [PubMed] [Google Scholar]
- Kihm U., Flamand A., Pastoret P.-P., Peterhans E. Round table on epidemiology and control of fox rabies. Veterinary Microbiology. 1992;33:297–301. doi: 10.1016/0378-1135(92)90057-z. [DOI] [PubMed] [Google Scholar]
- Kimman T.G., Pol J.M.A., De Wind N., Oie-Lie N., Berns A.J.M., Gielkens A.L.J. Role of different gene in the virulence and pathogenesis of Aujeszky’s disease virus. Veterinary Microbiology. 1992;33:45–52. doi: 10.1016/0378-1135(92)90034-q. [DOI] [PubMed] [Google Scholar]
- Kit S., Sheppard M., Ichimura H., Kit M. Second-generation pseudorabies virus vaccine with deletions in thymidine kinase and glycoprotein genes. American Journal of Veterinary Research. 1987;48:780–793. [PubMed] [Google Scholar]
- Krakowa S., Ellis J.A., McNeilly F., Gilpin D., Meehan B., McCullough K., Allan G. Immunologic features of porcine circovirus type 2 infection. Viral Immunology. 2002;15:567–582. doi: 10.1089/088282402320914511. [DOI] [PubMed] [Google Scholar]
- Lin, T.C., Lee, R.C.T., 1981. An overall report on the development of a highly safe and potent lapinized hog cholera virus strain for hog cholera control in Taiwan. NSC special Publication no. 5.
- Lindsay D.S., Blagburn B.L., Dubey J.P. Safety and results of challenge of weaned pigs given a temperature-sensitive mutant of Toxoplasma gondii. Journal of Parasitology. 1993;79:71–76. [PubMed] [Google Scholar]
- Lycke N. ADP-ribosylating bacterial enzymes for the targeted control of mucosal tolerance and immunity. Annals of the New York Academy of Sciences. 2004;1029:193–208. doi: 10.1196/annals.1309.036. [DOI] [PubMed] [Google Scholar]
- Marchioli C.C., Yancey R.J., Wardley R.C., Thomsen D.R., Post L.E. A vaccine strain of pseudorabies virus with deletions in the thymidine kinase and glycoprotein X genes. American Journal of Veterinary Research. 1987;48:1577–1583. [PubMed] [Google Scholar]
- McCluskie M.J., Krieg A.M. Enhancement of infectious disease vaccines through TLR9-dependent recognition of CpG DNA. Current Topics in Microbiological Immunology. 2006;311:155–178. doi: 10.1007/3-540-32636-7_6. [DOI] [PubMed] [Google Scholar]
- Mettenleiter T.C. Molecular biology of pseudorabies (Aujeszky’s disease) virus. Comparative Immunology and Microbiology of Infectious Diseases. 1991;14:151–163. doi: 10.1016/0147-9571(91)90128-z. [DOI] [PubMed] [Google Scholar]
- Mettraux C., Ackermann M., Tratschin J.-D., Kihm U. Approaches to the identification of non-essential genes of African swine fever virus. Veterinary Microbiology. 1992;33:101–115. doi: 10.1016/0378-1135(92)90039-v. [DOI] [PubMed] [Google Scholar]
- Monath T.P. Prospects for the development of a vaccine against the West Nile virus. Annals of the New York Academy of Sciences. 2001;951:1–12. doi: 10.1111/j.1749-6632.2001.tb02680.x. [DOI] [PubMed] [Google Scholar]
- Moon H.W., Bunn T.O. Vaccines for preventing enterotoxigenic Escherichia coli infections in farm animals. Vaccine. 1993;11:113–200. doi: 10.1016/0264-410X(93)90020-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nettleton P.F., Thiry E., Reid H., Pastoret P.P. Herpes virus infections in cervidae. Reviews of Scientific Office of International Epizootiology. 1988;7:977–988. doi: 10.20506/rst.7.4.384. [DOI] [PubMed] [Google Scholar]
- OECD, 2002. OECD series on principles of good laboratory practice and compliance monitoring, numbers 1–13. Organisation for Economic Co-operation and Development.
- Olson M.E., Ceri H., Morck D.W. Giardia vaccination. Trends in Parasitology. 2000;16:213–217. doi: 10.1016/s0169-4758(99)01623-3. [DOI] [PubMed] [Google Scholar]
- Pastoret P.-P., Brochier B. The development and use of a vaccinia-rabies recombinant oral vaccine for the control of wildlife rabies; a link between Jenner and Pasteur. Epidemiology and Infection. 1996;116:235–240. doi: 10.1017/s0950268800052535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel J.R., Heldens J. Equine herpes viruses 1 (EHV-1) and 4 (EHV-4) – epidemiology, disease and immunoprophylaxis: a brief review. The Veterinary Journal. 2005;170:14–23. doi: 10.1016/j.tvjl.2004.04.018. [DOI] [PubMed] [Google Scholar]
- Patel, J.R., Heldens, J.G.M., accepted for publication. Immunoprophylaxis against important virus diseases of farm animals: a review. Veterinary Journal. [DOI] [PubMed]
- Patel J.R., Shilleto R.W., Williams J., Alexander D.C.S. Prevention of transplacental infection of bovine foetus by bovine viral diarrhea virus through vaccination. Archives of Virology. 2002;147:2453–2463. doi: 10.1007/s00705-002-0878-3. [DOI] [PubMed] [Google Scholar]
- Patel J.R., Bateman H., Williams J., Didlick S. Derivation and characterisation of a live equid herpes virus 1 (EHV-1) vaccine to protect against and respiratory disease due to EHV-1. Veterinary Microbiology. 2003;91:23–39. doi: 10.1016/s0378-1135(02)00259-6. [DOI] [PubMed] [Google Scholar]
- Patel J.R., Földi J., Bateman H., Williams J., Didlick S., Stark R. Equid herpes virus (EHV-1) live vaccine strain C147: efficacy against respiratory diseases following EHV types 1 and 4 challenges. Veterinary Microbiology. 2003;92:1–17. doi: 10.1016/s0378-1135(02)00358-9. [DOI] [PubMed] [Google Scholar]
- Patel J.R., Didlick S., Bateman H. Efficacy of a live equine herpes virus – 1 (EHV-1) strain C147 vaccine in foals with maternally derived antibody. Protection against EHV-1 infection. Equine Veterinary Journal. 2004;36:447–451. doi: 10.2746/0425164044868332. [DOI] [PubMed] [Google Scholar]
- Pensaert M., Gielkens A.L.J., Lomniczi B., Kimman T.G., Vannier P., Eloit M. Round table on control of Aujeszky’s disease and vaccine development based on molecular biology. Veterinary Microbiology. 1992;33:53–67. doi: 10.1016/0378-1135(92)90035-r. [DOI] [PubMed] [Google Scholar]
- Petersen S.K., Foged N.T., Bording A., Nielsen J.P., Riemann H.K., Frandsen P.L. Recombinant derivatives of Pasteurella multocida toxin: candidates for a vaccine against progressive atrophic rhinitis. Infection and Immunity. 1991;59:1387–1393. doi: 10.1128/iai.59.4.1387-1393.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ph. Eur . fourth ed. The European Department for the Quality of Medicines; Strasbourg: 2005. European Pharmacopoeia. [Google Scholar]
- Plowright W. Malignant catarrhal fever virus. In: Dinter Z., Morein B., editors. Virus Infection of Ruminants. Elsevier; Amsterdam: 1990. pp. 123–150. [Google Scholar]
- Quint W.G.V., Gielkens A.L.J., Van Oirschot J.T., Berns A.J.M., Cuypers H.T. Construction and characterisation of deletion mutants of pseudorabies virus: a new generation of “live” vaccines. Journal of General Virology. 1987;68:523–534. doi: 10.1099/0022-1317-68-2-523. [DOI] [PubMed] [Google Scholar]
- Ranz A.I., Miguet J.G., Anaya C., Venteo A., Cortés E., Vela C., Sanz A. Diagnostic methods for African horse sickness virus using monoclonal antibodies to structural and non-structural proteins. Veterinary Microbiology. 1992;33:143–153. doi: 10.1016/0378-1135(92)90042-r. [DOI] [PubMed] [Google Scholar]
- Reid H.W., Buxton D. Malignant catarrhal fever and gamma herpesviridae of Bovidae. In: Witteman G., editor. Herpes Virus Diseases of Cattle, Horses and Pigs. Kluwer Academic Publishers; Dordrecht: 1989. pp. 116–162. [Google Scholar]
- Roy P. From genes to complex structures of bluetongue virus and their efficacy as vaccines. Veterinary Microbiology. 1992;33:155–168. doi: 10.1016/0378-1135(92)90043-s. [DOI] [PubMed] [Google Scholar]
- Salt J.S., Anthonis A.F.G., Peters A.R., Brune A., Jahnecke S., Traeder W., Harmeyer S.S. Breite kreuzneutralisation von europäische BVDV Typ-1 und Typ-2 Stämmen und signifikante Verbesserung der Fertilität nach Testinfektionen. Tierärztliche Praxis. 2004;32:191–195. [Google Scholar]
- Scarselli M., Giuliani M.M., Adu-Bobie J., Pizza M., Rappuoli R. The impact of genomics on vaccine design. Trends in Biotechnology. 2005;23:84–91. doi: 10.1016/j.tibtech.2004.12.008. [DOI] [PubMed] [Google Scholar]
- Schetters T. Vaccine development from a commercial point of view. Veterinary Parasitology. 1995;57:267–275. doi: 10.1016/0304-4017(94)03125-g. [DOI] [PubMed] [Google Scholar]
- Schetters Th.P.M. Intervet symposium bovine neosporosis. Veterinary Parasitology. 2004;125:137–146. doi: 10.1016/j.vetpar.2004.05.010. [DOI] [PubMed] [Google Scholar]
- Schetters Th.P.M. Vaccination against canine babesiosis. Trends in Parasitology. 2005;21:179–184. doi: 10.1016/j.pt.2005.02.006. [DOI] [PubMed] [Google Scholar]
- Schetters T.P., Gravendyck M. Regulations and procedures in parasite vaccine development. Parasitology. 2006;133(Suppl.):S189–S195. doi: 10.1017/S0031182006001879. [DOI] [PubMed] [Google Scholar]
- Schijns V.E.C.J. Immunological concepts of vaccine adjuvant activity. Current Opinion in Immunology. 2000;12:456–463. doi: 10.1016/s0952-7915(00)00120-5. [DOI] [PubMed] [Google Scholar]
- Schijns V.E.C.J. Induction and direction of immune responses by vaccine adjuvants. Critical Reviews in Immunology. 2001;21:75–85. [PubMed] [Google Scholar]
- Schijns V.E.C.J. Antigen delivery systems and immunostimulation. Veterinary Immunology and Immunopathology. 2002;87:195–198. doi: 10.1016/s0165-2427(02)00072-7. [DOI] [PubMed] [Google Scholar]
- Schijns V.E.C.J. Mechanisms of vaccine adjuvant activity: initiation and regulation of immune responses by vaccine adjuvants. Vaccine. 2003;21:829–831. doi: 10.1016/s0264-410x(02)00527-3. [DOI] [PubMed] [Google Scholar]
- Schijns V.E.C.J. Unravelling “the immunologist’s dirty little secret”. In: Schijns Virgil E.J.C., O’Hagan Derek., editors. Immunopotentiators in Modern Vaccines. Elsevier/Academic Press; Amsterdam: 2006. [Google Scholar]
- Schijns V.E.C.J., Tangeras A. Vaccine adjuvant technology: from theoretical mechanisms to practical approaches. Developments in Biologicals (Basel) 2005;121:127–134. [PubMed] [Google Scholar]
- Schijns, V.E.C.J., Jagdev Sharma, J., Tarpey, I., in press. Practical aspects of vaccination. In: Davidson, Schat, Kaspers (Eds.), Avian Immunology (in press).
- Simon R., Samuel C.E. Activation of NF-kappaB-dependent gene expression by Salmonella flagellins FliC and FljB. Biochemical and Biophysical Research Communications. 2007;355:280–285. doi: 10.1016/j.bbrc.2007.01.148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soulebot J.P., Folkers C., Taylor J., Pastoret P.P. Properties of vaccines. In: Pastoret P.P., Blancou J., Vannier P., Verschueren C., editors. Veterinary Vaccinology. Elsevier; Amsterdam: 1997. pp. 159–166. [Google Scholar]
- Straub, O.C., 2002. Advantages and disadvantages of common vaccination programs. In: Kaske, M., Scholz, H., Höltershinken, M. (Eds.). Recent Developments and Perspectives in Bovine Medicine: Keynote Lectures of the XXII World Buiatrics Congress, Klinik für Rinderkrankheiten, Hannover, pp. 86–96.
- Swayne D.E., Lee C.W., Spackman E. Inactivated North American and European H5N2 avain influenza vaccines protect chickens from Asian H5N1 high pathogenicity influenza virus. Avian Pathology. 2006;35(2):141–146. doi: 10.1080/03079450600597956. [DOI] [PubMed] [Google Scholar]
- Urquhart G.M. Field experience with the bovine lungworm vaccine. Developments in Biological Standardization. 1985;62:109–112. [PubMed] [Google Scholar]
- Valikhov, A., Ivanova, L., Vassin, A., Zakharova, N., Makarova, L., Altenstein, A., 1997. Immunogenicity of vaccinia virus (VV) recombinants, expressing bovine leukemia virus (BLV) env gene, in rabbits and calves. In: Fourth International Congress of Veterinary Virology, Edinburgh, Scotland, UK, p. 200.
- van Gennip H.G., van Rijn P.A., Widjojoatmodjo M.N., de Smit A.J., Moormann R.J. Chimeric classical swine fever viruses containing envelope protein E(RNS) or E2 of bovine viral diarrhoea virus protect pigs against challenge with CSFV and induce a distinguishable antibody response. Vaccine. 2000;19:447–459. doi: 10.1016/s0264-410x(00)00198-5. [DOI] [PubMed] [Google Scholar]
- Van Oirschot, J.T., 2000. Present and Future of veterinary viral vaccinology. In: Brocchi, E., Lavaza, A. (Eds.). Fifth International Congress of Veterinary Virology, Istituto Zooprofillatico Sperimentale della Lombardia e dell Emilia Romagna, Brescia Fondazione Iniziative Zooprofilattiche e Zootechniche, Brescia, Italy, pp. 18–27.
- Van Poppel N.F., Welagen J., Duisters R.F., Vermeulen A.N., Schaap D. Tight control of transcription in Toxoplasma gondii using an alternative tet repressor. International Journal of Parasitology. 2006;36:443–452. doi: 10.1016/j.ijpara.2006.01.005. [DOI] [PubMed] [Google Scholar]
- Van Zijl N., Wensvoort G., De Kluijver E.P., Hulst M.M., Van der Gulden H., Gielkens A.L.J., Berns A., Moormann R.J.N. Live attenuated pseudorabies virus expressing envelope glycoprotein E2 of hog cholera virus protects swine against both pseudorabies and hog cholera. Journal of Virology. 1991;65:2761–2765. doi: 10.1128/jvi.65.5.2761-2765.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vercruysse J., Knox D.P., Schetters Th.P.M., Willadsen P. Veterinary parasitic vaccines: pitfalls and future directions. Trends in Parasitology. 2004;20:488–492. doi: 10.1016/j.pt.2004.07.009. [DOI] [PubMed] [Google Scholar]
- VICH-GL9, 2000. Good Clinical Practice. International Cooperation on Harmonisation of Technical Requirements for Registration of Veterinary Medicinal Products http://vich.eudra.org/. [DOI] [PubMed]
- Westergaard, J.M., Control strategies of major viral infectious diseases of live stock in Europe, ten years of experience, in: Brocchi, E., Lavazza, A. (Eds.), Fifth International Congress of Veterinary Virology, Istituto Zooprofillatico Sperimentale della Lombardia e dell Emilia Romagna, Brescia Fondazione Iniziative Zooprofilattiche e Zootechniche, Brescia, Italy, 2000, pp. 3–12.
- Willadsen P. Anti-tick vaccines. Parasitology. 2004;129(Suppl.):S367–S387. doi: 10.1017/s0031182003004657. [DOI] [PubMed] [Google Scholar]
- Williams R.B. Safety of the attenuated anticoccidial vaccine Paracox in broiler chickens isolated from extraneous coccidial infection. Veterinary Research Communications. 1994;18:189–198. doi: 10.1007/BF01839268. [DOI] [PubMed] [Google Scholar]
- Williams R.B. Anticoccidial vaccines for broiler chickens: pathways to success. Avian Pathology. 2002;31:317–353. doi: 10.1080/03079450220148988. (Erratum in Avian Pathology 32, 429) [DOI] [PubMed] [Google Scholar]
- Wittmann G., Rziha H.J. Aujeszky’s disease (pseudo rabies) in pigs. In: Wittmann A., editor. Herpes Virus Disease of Cattle, Horses and Pigs. Kluwer Academic Publishers; Boston: 1989. pp. 230–325. [Google Scholar]
- Wood J.M., Robertson J.S. Reference viruses for seasonal and pandemic influenza vaccine preparation. Influenza and Other Respiratory Viruses. 2006;1:4–9. doi: 10.1111/j.1750-2659.2006.00003.x. [DOI] [PMC free article] [PubMed] [Google Scholar]