

Abstract
Although the recent SARS coronavirus (SARS-CoV) that appeared in 2002 has now been contained, the possibility of re-emergence of SARS-CoV remains. Due to the threat of re-emergence, the overall fatality rate of ∼10%, and the rapid dispersion of the virus via international travel, viable vaccine candidates providing protection from SARS are clearly needed. We developed an attenuated VSV recombinant (VSV-S) expressing the SARS coronavirus (SARS-CoV) spike (S) protein. In cells infected with this recombinant, S protein was synthesized, glycosylated at approximately 17 Asn residues, and transported via the Golgi to the cell surface. Mice vaccinated with VSV-S developed SARS-neutralizing antibody and were able to control a challenge with SARS-CoV performed at 1 month or 4 months after a single vaccination. We also demonstrated, by passive antibody transfer, that the antibody response induced by the vaccine was sufficient for controlling SARS-CoV infection. A VSV-vectored SARS vaccine could have significant advantages over other SARS vaccine candidates described to date.
Keywords: Intranasal, Neutralizing antibodies, Protective immunity, Spike
In the fall of 2002 an emerging human disease termed SARS (severe acute respiratory syndrome) drew global attention. The disease was characterized as an atypical pneumonia accompanied by high fever. According the World Health Organization, there were 8096 cases of SARS, 21% of which were in health care workers. Approximately 10% of the infected individuals died as a result of the illness, and the death rate exceeded 50% for those over 60 years old (http://www.who.int/csr/sars/en/WHOconsensus.pdf and Peiris et al., 2004). The causative agent was identified as a coronavirus (Drosten et al., 2003, Ksiazek et al., 2003), a positive-stranded enveloped RNA virus, and the 29.7-kb genome sequence was determined rapidly. Like other coronaviruses, the SARS coronavirus (SARS-CoV) has 6 major open reading frames, two encoding the polymerase, and four encoding the structural proteins spike (S), membrane, envelope, and nucleocapsid (Marra et al., 2003, Rota et al., 2003). The SARS S protein is a type I transmembrane glycoprotein. It is responsible for viral binding to host cellular receptors, identified as angiotensin converting enzyme 2 (Li et al., 2003) and CD209L (Jeffers et al., 2004), followed by viral fusion to host cells (Bosch et al., 2004, Liu et al., 2004, Yuan et al., 2004). Immunization with the S proteins of other coronaviruses (mouse hepatitis virus [MHV], transmissible gastroenteritis virus [TGEV], and infectious bronchitis virus [IBV]) generates protective immunity against these viruses in various animal models (De Diego et al., 1992, Ignjatovic and Galli, 1994, Wesseling et al., 1993). Expression of the S protein of SARS-CoV from either in vitro expression systems or from live viral vectors has recently been reported to generate protection against SARS-CoV infection in animal models (Bisht et al., 2004, Bukreyev et al., 2004, Chen et al., 2005, Gao et al., 2003, Yang et al., 2004).
The spread of SARS was curtailed by public health measures, and the outbreak was successfully contained in July 2003. Although several animals are susceptible to SARS-CoV infection, the source of the outbreak and host reservoir are uncertain (Guan et al., 2003). The possibility of a re-emergence still exists, and in such an event, a SARS vaccine would be valuable, especially to protect the most vulnerable groups, the elderly and health care workers.
In this study, an experimental vesicular stomatitis virus (VSV)-based SARS vaccine was developed and tested. VSV is a negative strand RNA virus with a non-segmented genome that encodes five structural proteins, nucleocapsid (N), phosphoprotein (P), matrix (M), glycoprotein (G), and an RNA-dependent RNA polymerase (L). VSV can elicit strong humoral and cellular immune responses in a variety of animals, and VSV-based vaccines have been shown to confer immunity in animal models of respiratory syncytial virus (Kahn et al., 2001), influenza virus (Roberts et al., 1998), SHIV/AIDS (Rose et al., 2001), measles virus (Schlereth et al., 2000), and human papilloma virus (Roberts et al., 2004). VSV has significant advantages over many other vaccine vectors. It has a relatively small RNA genome but can accommodate insertion of large foreign genes. It replicates exclusively in the cytoplasm via RNA intermediates only and does not undergo recombination. VSV seropositivity is also extremely low in the general population (Rose and Whitt, 2001). Recombinant VSV generated by the plasmid DNA system employed here have been shown to be less pathogenic (attenuated) when compared with naturally occurring strains of VSV (Roberts et al., 1998) and cause no disease symptoms in non-human primates (Rose et al., 2001) when given by intranasal, oral, or intramuscular routes. In the present study, we developed a recombinant, attenuated VSV expressing the SARS-CoV S protein and tested it as a SARS vaccine in a mouse model (Subbarao et al., 2004). In this model, SARS-CoV replicates to high titers in the respiratory tract, allowing protection from SARS infection to be assayed readily in vaccinated animals following SARS-CoV challenge.
Results
Construction and characterization of a VSV recombinant (VSV-S) expressing SARS-CoV S protein
To determine if the SARS-CoV S protein could be expressed from a VSV recombinant, we first generated a cDNA clone of the SARS S gene by reverse transcription and PCR amplification of RNA purified from SARS-CoV-infected cells. We next constructed a plasmid with the SARS-CoV S coding sequence inserted between the VSV G and VSV L genes (Fig. 1A). Appropriate VSV transcription start and stop sites flanked the S gene. We recovered a recombinant VSV (designated VSV-S) from this plasmid using published methods (Lawson et al., 1995). To determine if the recombinant virus expressed the SARS S protein, we infected BHK cells with the recovered virus or with wt VSV and metabolically labeled cells for 1 h with [35S]-methionine. Lysates of radiolabeled cells were then analyzed by SDS–PAGE. Since VSV shuts off host protein synthesis, wt VSV-infected cells express predominantly the five viral proteins L, G, N, P, and M. VSV-S-infected cells expressed the five VSV proteins (Fig. 1B), and also a protein of the expected size for S (calculated as ∼197 kDa assuming N-glycosylation at all 23 potential sites). To verify the identity of the S protein, S was immunoprecipitated from the cell lysates using a rabbit polyclonal antibody to the S cytoplasmic domain. The immunoprecipitated protein appeared as two bands (Fig. 1C, lane 1), potentially representing two different glycosylation states of S. To examine this possibility we treated the protein with PNGase F (Fig. 1C, lane 2), which cleaves all N-linked glycans. PNGase F-digestion yielded a single band of approximately 156 kDa, confirming that variable glycosylation states of the S protein were responsible for the observed size heterogeneity. Digestion of immunoprecipitated S protein with Endo H, an endoglycosidase that cleaves only unprocessed or partially processed glycans from proteins, yielded two bands (Fig. 1C, lane 3). The Endo-H-resistant band of original size accounted for ∼15% of total S protein. The lower band, representing protein which had not yet undergone processing of its carbohydrate side chains, accounted for ∼85% of total S protein. In a pulse-chase experiment with a 20-min pulse labeling, we observed that 50% of S expressed by VSV was Endo H resistant within 30 min after the chase was begun (data not shown). From these experiments, we concluded that the S protein is transported through the exocytic pathway at least as far as the Golgi apparatus where Endo H resistance is acquired.
Fig. 1.
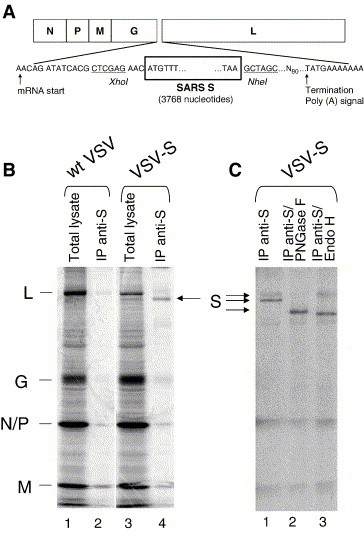
VSV recombinant expressing the SARS-CoV S protein. (A) The diagram illustrates the genome of the VSV-S recombinant expressing the S gene and shows the terminal coding and flanking VSV transcription start and stop signals. (B) BHK-21 cells were infected with wt VSV (lanes 1 and 2) or VSV-S (lanes 3 and 4), labeled with [35S]-methionine, and lysates were analyzed by SDS–PAGE without (lanes 1 and 3) or with immunoprecipitation with anti-S antibody (lanes 2 and 4). (C) Immunoprecipitated S was left undigested (lane 1), digested with PNGase F (lane 2), or Endo H (lane 3).
To determine if the S protein was transported from the Golgi to the cell surface, we examined VSV-S-infected cells using indirect immunofluorescence microscopy. BHK-21 cells were infected with VSV-S or wt VSV, fixed, and then incubated with serum from a person who had recovered from SARS-CoV infection. A secondary, Alexa Fluor 488-conjugated anti-human antibody was used for visualization with fluorescence microscopy. We found that the SARS S protein was clearly expressed on the cell surface as indicated by the strong fluorescent signal visible in VSV-S-but not wt VSV-infected cells (Fig. 2 ).
Fig. 2.
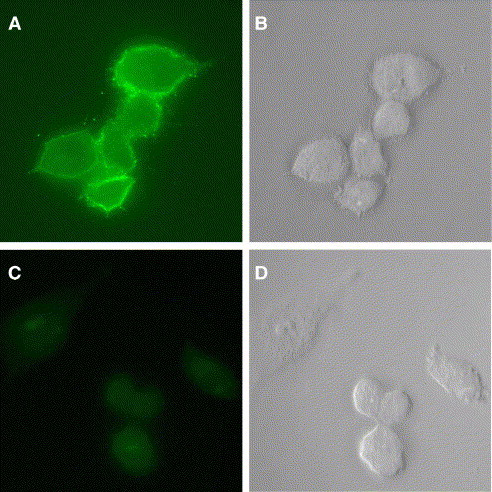
SARS-CoV S protein is expressed on the surface of VSV-S-infected cells. BHK-21 cells were infected with VSV-S (A and B) or wt VSV (C and D). Cells were fixed and stained for SARS S as described in Materials and methods. The fluorescence (left) and DIC (right) images were generated with a Nikon Microphot-FX microscope using a 60× objective.
Immunization with VSV-S elicits humoral immunity
Initially we immunized four groups of eight mice with either wt VSV, VSV-S, or SARS-CoV. Four weeks later, serum was collected from each mouse. To determine if the VSV and VSV-S inoculations were effective, we performed an assay to measure VSV-neutralizing antibodies. Pooled sera from mice infected with wt VSV or VSV-S had a VSV-neutralizing antibody titer of 1:2560, while mice infected with SARS-CoV did not have any detectable VSV-neutralizing antibody.
We then examined the individual serum samples for neutralizing antibodies to SARS-CoV (Table 1 ). Infection of mice with VSV-S generated a stronger neutralizing antibody response (average SARS-neutralizing titer of 1:32) to SARS-CoV than did infection with SARS-CoV (average SARS-neutralizing titer of 1:12). Sera from control mice immunized with wt VSV did not have detectable neutralizing antibody titers to SARS-CoV.
Table 1.
Neutralizing antibody titers to SARS-CoV
| Primary immunogen | VSV | VSV-S | SARS |
|---|---|---|---|
| Individual SARS-CoV-neutralizing antibody titer | <1:16 | 1:13 | <1:8 |
| <1:8 | 1:25 | 1:20 | |
| <1:8 | 1:20 | 1:10 | |
| <1:8 | 1:32 | 1:25 | |
| <1:4 | 1:81 | 1:11 | |
| <1:4 | 1:16 | 1:11 | |
| <1:8 | 1:40 | 1:8 | |
| <1:8 | |||
| Average | 1:32 | 1:12 |
Immunization with VSV-S protects against SARS-CoV infection
Mice inoculated intranasally with SARS-CoV do not show any clinical symptoms of infection; however, the virus replicates to high titers in the lungs and nasal turbinates (NTs). SARS-CoV replication in respiratory tissues peaks by 2 days post-infection and virus is cleared within a week (Subbarao et al., 2004). We examined SARS-CoV replication in vaccinated and control animals to evaluate the effectiveness of the VSV-S vaccine in protecting against SARS-CoV infection.
Three groups of four mice each were immunized with either wt VSV, VSV-S, or SARS-CoV. Four weeks later the mice were challenged with SARS-CoV, and 2 days after the challenge, the lungs and nasal turbinates were collected, and the corresponding viral titers were determined (Fig. 3A). Control mice, immunized with wt VSV, had high SARS viral titers in both lungs and nasal turbinates following SARS-CoV challenge. In contrast, mice immunized with VSV-S or SARS-CoV were completely protected and controlled the SARS-CoV challenge as indicated by SARS-CoV titers at or below the detection limits in both the lungs and nasal turbinates. These data indicate that vaccination with VSV-S is as effective as a primary infection with SARS-CoV in preventing subsequent SARS-CoV infections.
Fig. 3.
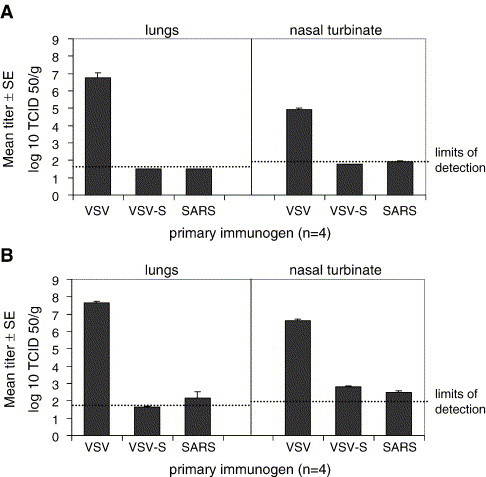
Protection from SARS-CoV infection in immunized mice. Balb/c mice were inoculated intranasally with wt VSV, VSV-S, or SARS-CoV. The mice were challenged 4 weeks later with 105 TCID50 SARS-CoV. Two days later, lungs and nasal turbinates were collected, and the viral titers were determined (A). Additional mice were challenged 4 months after immunization with wt VSV, VSV-SARS S, and SARS-CoV. Two days after the challenge, lungs and nasal turbinates were collected and viral titers were determined (B). Limits of detection were 1.5 log10TCID50/g tissue in 10% lung homogenates and 1.8 log10TCID50/g tissue in 5% nasal turbinate homogenates.
To determine if VSV-S vaccination could provide sustained protection from SARS-CoV infection, a similar experiment was carried out with a 4-month interval between immunizations and challenge (Fig. 3B). Two days after challenge, SARS-CoV titers in the lungs of VSV-S immunized mice were at the limit of detection (1 of 4 mice) or undetectable (3 of 4 mice), while the titer in the lungs of SARS-CoV immunized mice was slightly above the limit of detection (1 of 4 mice) or undetectable (3 of 4 mice). Viral titers in the nasal turbinates were slightly above the limit of detection for both VSV-S (3 of 4 mice)- and SARS-CoV (2 of 4 mice)-immunized mice. The mean SARS viral titers in nasal turbinates of both VSV-S- and SARS-CoV-inoculated mice were nearly 10,000-fold lower than the viral titers in nasal turbinates from wt VSV-immunized mice. These findings demonstrate that VSV-S vaccination provides sustained protection from SARS-CoV challenge and may provide protective immunity equal to or better than primary SARS-CoV infection.
Serum from mice immunized with VSV-S can confer passive protection
VSV infection can elicit strong humoral and cellular immune responses. To determine if antibody alone could provide protection against SARS-CoV infection, serum from wt VSV-, VSV-S-, or SARS-CoV-infected mice was administered intraperitoneally into naïve mice (Fig. 4 ). Mice were bled 24 h after administration of antisera and SARS-CoV-neutralizing antibody titers were measured. Mice were subsequently challenged with SARS-CoV, and 2 days later lungs were collected and SARS-CoV titers were determined. SARS-CoV-neutralizing antibody titers were detectable in the groups of mice that received VSV-S antisera and SARS-CoV antisera (diluted 1:10). Only these mice were able to protect against SARS-CoV infection upon challenge. Mice receiving wt VSV antisera or normal mouse sera did not have measurable neutralizing antibodies to SARS-CoV, nor were they protected from SARS-CoV infection upon challenge. These results demonstrate that VSV-S can elicit a strong antibody response that is sufficient for controlling SARS-CoV infection.
Fig. 4.
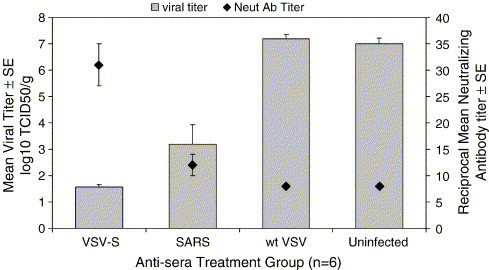
Inhibition of SARS-CoV replication by transfer of serum from immunized mice. Sera from uninfected mice or mice immunized with VSV-S, SARS-CoV (diluted 1:10 in PBS), or wt VSV was injected into naïve mice. Serum was then collected from mice to measure SARS-CoV-neutralizing antibody titer. (The limit of detection was a reciprocal dilution of 8.) The mice were then infected with SARS-CoV, and 2 days later the lungs were harvested to measure SARS-CoV titer. The limit of detection was 1.5 log10TCID50/g.
Discussion
In this study we used an attenuated VSV vector to express the SARS-CoV S protein gene. Because both SARS-CoV and VSV replicate in the cytoplasm of infected cells, we anticipated efficient expression of SARS-CoV S protein by a recombinant VSV without problems associated with nuclear transcription, including mRNA modification and mRNA export from the nucleus. The level of S protein expression obtained from the VSV-S recombinant was clearly sufficient to generate long-term protective immunity to SARS-CoV in this animal model.
The levels of SARS-CoV-neutralizing antibody titers following VSV-S immunization were equal to or better than those elicited following primary infection with SARS-CoV. Although passive transfer of immune sera demonstrated that antibody induced by either VSV-S or SARS-CoV inoculations is sufficient to prevent SARS-CoV infection, cellular immune responses may also contribute to protection. Similarly, it has been shown with other coronaviruses, such as mouse hepatitis virus type 3, that antibody-mediated immunity is sufficient for protection (Pope et al., 1996). Furthermore, the complete protection from SARS viral replication in the lower respiratory tissues of VSV-S-immunized mice observed when challenge was administered 4 months post-immunization suggests that prolonged protection may be provided from a single immunization with VSV-S. Ours is the first study to show such long-term protection after immunization with an experimental SARS-CoV vaccine. In previous studies on vaccination of mice with VSV expressing an influenza hemagglutinin, we found that protective influenza-neutralizing antibody titers were very long-lived, declining only 2- to 4-fold over the course of more than one year (Roberts and Rose, unpublished data).
Our VSV-based SARS vaccine could have significant advantages over previously described SARS vaccines (Bisht et al., 2004, Bukreyev et al., 2004, Chen et al., 2005, Gao et al., 2003, Yang et al., 2004). Pre-existing immunity to VSV is rare in the human population. Effectiveness of other vaccine candidates based on adenovirus or parainfluenza virus could be limited by a high prevalence of pre-existing immunity to the vectors in the human population. Vaccines based on modified vaccinia Ankara (MVA) could face the same problem because the parental vaccinia virus has been used worldwide as the smallpox vaccine, and there is extensive immunity to vaccinia, especially in the older population most susceptible to SARS (Crotty et al., 2003). DNA vaccines have been effective, especially in small animal models; however, efficacy in humans has yet to be demonstrated. VSV has the additional advantages of growing to very high titers in cell lines such as Vero that are approved for human vaccine production. VSV is also effective at very low doses (Roberts et al., 1998, Roberts et al., 1999) and is effective as an intranasal vaccine (Egan et al., 2004).
Inactivated SARS-CoV itself has also been shown to elicit an immune response (He et al., 2004, Qu et al., 2005, Takasuka et al., 2004, Tang et al., 2004, Xiong et al., 2004, Zhang et al., 2004); however, the production of such a vaccine would carry the inherent risks of exposure to SARS-CoV from handling large volumes of infectious material that could result in accidental infection and secondary spread. Additionally, there might be concerns of proper and complete inactivation of the virus.
The SARS-CoV S protein expressed by VSV was estimated from gel mobility to be approximately 200 kDa and was converted to about 156 kDa upon the removal of all N-linked glycans. Based on this calculation, we estimate that at least 17 of the 23 potential N-linked glycosylation sites are used (assuming 2.5 kDa per N-linked glycan). We also demonstrated that S expressed from the VSV recombinant moves to the Golgi apparatus where it acquires Endo H resistance. About 15% of S expressed during a 1-h labeling period contained Endo-H-resistant glycans. In a pulse-chase experiment with a 20-min pulse labeling, we observed that 50% of S expressed by VSV was Endo H resistant within 30 min after the chase was begun (data not shown). Finally, we readily observed S protein on the cell surface. Our results indicate that S protein moves efficiently through the exocytic pathway when it is expressed in the absence of other SARS virus proteins. Exposure of S protein on the cell surface of infected cells is likely to be important in the induction of a neutralizing antibody response.
Other coronavirus vaccines, including those for MHV, IBV, and TGEV, have been effective in controlling infection. In contrast, vaccine against feline infectious peritonitis virus using the S protein of that virus as an antigen enhanced disease in cats (Vennema et al., 1990). No such enhancement of infection was observed in the present mouse model in this and previous studies. However, since SARS-CoV does not cause a disease in mice, this model may not reveal such effects. Mild to moderate hepatic lesions have been observed in a single set of experiments where ferrets were vaccinated with MVA, MVA-SAR-S, or MVA-SARS-N and challenged with SARS-CoV (Czub et al., 2005, Weingartl et al., 2004). The significance of these lesions is unclear since they occur in MVA vaccinated, SARS-CoV-challenged animals, as well as unvaccinated (PBS controls) SARS-CoV challenged animals. Although two of three ferrets receiving MVA-SAR-S vaccine have slightly larger lesions following SARS-CoV challenge, it is difficult to ascertain whether hepatic disease is enhanced or just variable. Furthermore, SARS-CoV disease is primarily a respiratory disease, and the lesions in the liver may have little or no relevance to SARS-CoV disease. These observations may therefore be isolated to the use of this vector or this animal model or the specific combination. The potential for SARS-CoV-enhanced disease in previously vaccinated or SARS-exposed animals requires further studies in models that more closely mimic the human disease.
Materials and methods
Plasmid construction and recombinant VSV recovery
The SARS-CoV S gene was amplified from RNA of virus-infected cells (provided by Dr. William Bellini, CDC) by reverse transcription-PCR using Superscript One-Step RT-PCR for Long Templates (Invitrogen, Carlsbad, CA). The following primers were used in the RT-PCR: 5′-GATCGATCCTCGAGAACATGTTTATTTTCTTAATTATTTC-3′ and 5′-CGATCCCCCCGGGCTAGCTTATGTGTAATGTAATTTGACACCC. The PCR product was digested with XhoI and NheI and ligated into pVSVXN2 (Schnell et al., 1996), also digested with the same enzymes. The resulting clone, pVSV-SARS S, was sequenced. The S gene sequence matched the published sequence (GenBank accession no. AY278741) with the exception of a silent mutation at nucleotide 1173 which was G instead of A.
Recombinant virus was recovered as previously described (Lawson et al., 1995). Briefly, BHK-21 cells were infected with vTF7-3 (Fuerst et al., 1986) for 1 h and then transfected with pVSV-SARS S and support plasmids (pBS-N, pBS-M, pBS-G, and pBS-L) for 3 h. Two days after the transfection, the media were transferred onto fresh BHK-21 cells. The media were collected after 2 days. A stock of VSV-S from a single plaque was grown on BHK-21. Recombinant wild-type (wt) VSV (Lawson et al., 1995) was also grown on BHK-21.
SARS S antibody
In order to detect SARS-CoV S protein, antibodies directed to the c-terminal domain were produced. Rabbits were immunized and boosted with the peptide KFDEDDSEPVLKGVKLHYT coupled to KLH. Serum was collected and affinity purified (Pocono Rabbit Farm and Laboratory Inc., Canadensis, PA).
Metabolic labeling, immunoprecipitation, and SDS–PAGE
BHK-21 cells were infected with either VSV-SARS S or wt VSV at a multiplicity of infection (MOI) of 10. After 5 h, the cells were washed twice with methionine-free Dulbecco's modified Eagle's medium (DMEM) and incubated with DMEM containing 100 μCi of [35S]-methionine for 1 h. Cells were washed with phosphate-buffered saline (PBS) and then lysed with a detergent solution (1% Nonidet P-40, 0.4% deoxycholate, 50 mM Tris–HCl [pH 8], 62.5 mM EDTA). Protein samples were analyzed by SDS–10% PAGE.
SARS S was immunoprecipitated from lysate with the S-tail antiserum at a dilution of 1:100 at 4 °C overnight. Protein A-Sepharose (Zymed Laboratories Inc., San Francisco, CA) was added and allowed to incubate for 1 h at room temperature. The Sepharose was washed with HNTG buffer (50 mM HEPES, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 10% glycerol, 0.1% Triton X-100).
The immunoprecipitates were treated with Endoglycosidase (Endo) H and peptide N-Glycosidase (PNGase) F (New England Biolabs, Beverly, MA) according to manufacturer's instructions.
Immunofluorescence microscopy
BHK-21 cells seeded onto glass coverslips were infected with either VSV-S or wt VSV at an MOI of 10. 6 h after infection, the cells were washed with PBS and fixed with 3% paraformaldehyde. The cells were then washed with PBS–glycine (10 mM). Coverslips were incubated with a 1:200 dilution serum from a person who had recovered from SARS (provided by Dr. William Bellini, CDC). The coverslips were washed with PBS–glycine and incubated with a 1:500 dilution of Alexa Fluor 488 goat anti human IgG (Molecular Probes, Eugene, OR). Coverslips were washed and mounted on slides using glycerol containing 0.1 M n-propylgallate. Cells were observed with Nikon Microphot-FX epi-fluorescence microscope using a 60× objective.
Animal challenge protocol
Three groups of eight BABL/c mice were inoculated intranasally (i.n.) with either 1.6 × 104 pfu of wt VSV, 1.4 × 104 pfu of VSV-SARS S, or 105 TCID50 of SARS-CoV. Twenty-eight days after the immunization, serum was collected from all mice to measure neutralizing antibody titers. Four mice from each group were then challenged i.n. with 104 TCID50 SARS-CoV as previously described (Subbarao et al., 2004). Two days later, lungs and nasal turbinates of the mice were collected to determine viral titers (Subbarao et al., 2004). The remaining four mice were challenged 4 months post-immunization. Lungs and nasal turbinates were collected 2 days after the challenge, and viral titers were determined (Subbarao et al., 2004).
Neutralizing antibody titers
VSV-neutralizing antibody titers were measured by determining the highest dilution of serum that could prevent killing of a monolayer of BHK-21 cells by VSV (Rose et al., 2001). SARS-CoV-neutralizing antibody titers were determined by evaluating the serum dilution at which the cytopathic effects (CPE) of SARS-CoV were inhibited in half of the wells containing Vero E6 monolayer (Subbarao et al., 2004).
Passive protection study
Sera from mice immunized with wt VSV, VSV-S, and SARS-CoV were collected and heat inactivated. Two groups of six mice were injected intraperitoneally (i.p.) with 500 μl of wt VSV or VSV-S immune sera. Two additional groups of six mice were also injected i.p. with 500 μl of SARS immune sera diluted 1:10 in PBS or sera of naïve mice. Serum was collected from each mouse to determine neutralizing antibody titers against SARS-CoV (Subbarao et al., 2004). Mice were then inoculated i.n. with 105 TCID50 of SARS-CoV. Two days later, the lungs were collected from each mouse, and viral titer was determined as described above.
Acknowledgments
We thank Dr. William Bellini for providing RNA from SARS-CoV infected cells and for serum samples. This work was supported by NIH grant AI057158.
References
- Bisht H., Roberts A., Vogel L., Bukreyev A., Collins P.L., Murphy B.R., Subbarao K., Moss B. Severe acute respiratory syndrome coronavirus spike protein expressed by attenuated vaccinia virus protectively immunizes mice. Proc. Natl. Acad. Sci. U.S.A. 2004;101(17):6641–6646. doi: 10.1073/pnas.0401939101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bosch B.J., Martina B.E., Van Der Zee R., Lepault J., Haijema B.J., Versluis C., Heck A.J., De Groot R., Osterhaus A.D., Rottier P.J. Severe acute respiratory syndrome coronavirus (SARS-CoV) infection inhibition using spike protein heptad repeat-derived peptides. Proc. Natl. Acad. Sci. U.S.A. 2004;101(22):8455–8460. doi: 10.1073/pnas.0400576101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bukreyev A., Lamirande E.W., Buchholz U.J., Vogel L.N., Elkins W.R., St Claire M., Murphy B.R., Subbarao K., Collins P.L. Mucosal immunisation of African green monkeys (Cercopithecus aethiops) with an attenuated parainfluenza virus expressing the SARS coronavirus spike protein for the prevention of SARS. Lancet. 2004;363(9427):2122–2127. doi: 10.1016/S0140-6736(04)16501-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Z., Zhang L., Qin C., Ba L., Yi C.E., Zhang F., Wei Q., He T., Yu W., Yu J., Gao H., Tu X., Gettie A., Farzan M., Yuen K.Y., Ho D.D. Recombinant modified vaccinia virus Ankara expressing the spike glycoprotein of severe acute respiratory syndrome coronavirus induces protective neutralizing antibodies primarily targeting the receptor binding region. J. Virol. 2005;79(5):2678–2688. doi: 10.1128/JVI.79.5.2678-2688.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crotty S., Felgner P., Davies H., Glidewell J., Villarreal L., Ahmed R. Cutting edge: long-term B cell memory in humans after smallpox vaccination. J. Immunol. 2003;171(10):4969–4973. doi: 10.4049/jimmunol.171.10.4969. [DOI] [PubMed] [Google Scholar]
- Czub M., Weingartl H., Czub S., He R., Cao J. Evaluation of modified vaccinia virus Ankara based recombinant SARS vaccine in ferrets. Vaccine. 2005;23(17–18):2273–2279. doi: 10.1016/j.vaccine.2005.01.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Diego M., Laviada M.D., Enjuanes L., Escribano J.M. Epitope specificity of protective lactogenic immunity against swine transmissible gastroenteritis virus. J. Virol. 1992;66(11):6502–6508. doi: 10.1128/jvi.66.11.6502-6508.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drosten C., Gunther S., Preiser W., van der Werf S., Brodt H.R., Becker S., Rabenau H., Panning M., Kolesnikova L., Fouchier R.A., Berger A., Burguiere A.M., Cinatl J., Eickmann M., Escriou N., Grywna K., Kramme S., Manuguerra J.C., Muller S., Rickerts V., Sturmer M., Vieth S., Klenk H.D., Osterhaus A.D., Schmitz H., Doerr H.W. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 2003;348(20):1967–1976. doi: 10.1056/NEJMoa030747. [DOI] [PubMed] [Google Scholar]
- Egan M.A., Chong S.Y., Rose N.F., Megati S., Lopez K.J., Schadeck E.B., Johnson J.E., Masood A., Piacente P., Druilhet R.E., Barras P.W., Hasselschwert D.L., Reilly P., Mishkin E.M., Montefiori D.C., Lewis M.G., Clarke D.K., Hendry R.M., Marx P.A., Eldridge J.H., Udem S.A., Israel Z.R., Rose J.K. Immunogenicity of attenuated vesicular stomatitis virus vectors expressing HIV type 1 Env and SIV Gag proteins: comparison of intranasal and intramuscular vaccination routes. AIDS Res. Hum. Retroviruses. 2004;20(9):989–1004. doi: 10.1089/aid.2004.20.989. [DOI] [PubMed] [Google Scholar]
- Fuerst T.R., Niles E.G., Studier F.W., Moss B. Eukaryotic transient-expression system based on recombinant vaccinia virus that synthesizes bacteriophage T7 RNA polymerase. Proc. Natl. Acad. Sci. U.S.A. 1986;83(21):8122–8126. doi: 10.1073/pnas.83.21.8122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao W., Tamin A., Soloff A., D'Aiuto L., Nwanegbo E., Robbins P.D., Bellini W.J., Barratt-Boyes S., Gambotto A. Effects of a SARS-associated coronavirus vaccine in monkeys. Lancet. 2003;362(9399):1895–1896. doi: 10.1016/S0140-6736(03)14962-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guan Y., Zheng B.J., He Y.Q., Liu X.L., Zhuang Z.X., Cheung C.L., Luo S.W., Li P.H., Zhang L.J., Guan Y.J., Butt K.M., Wong K.L., Chan K.W., Lim W., Shortridge K.F., Yuen K.Y., Peiris J.S., Poon L.L. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science. 2003;302(5643):276–278. doi: 10.1126/science.1087139. [DOI] [PubMed] [Google Scholar]
- He Y., Zhou Y., Siddiqui P., Jiang S. Inactivated SARS-CoV vaccine elicits high titers of spike protein-specific antibodies that block receptor binding and virus entry. Biochem. Biophys. Res. Commun. 2004;325(2):445–452. doi: 10.1016/j.bbrc.2004.10.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ignjatovic J., Galli L. The S1 glycoprotein but not the N or M proteins of avian infectious bronchitis virus induces protection in vaccinated chickens. Arch. Virol. 1994;138(1–2):117–134. doi: 10.1007/BF01310043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeffers S.A., Tusell S.M., Gillim-Ross L., Hemmila E.M., Achenbach J.E., Babcock G.J., Thomas W.D., Jr., Thackray L.B., Young M.D., Mason R.J., Ambrosino D.M., Wentworth D.E., Demartini J.C., Holmes K.V. CD209L (L-SIGN) is a receptor for severe acute respiratory syndrome coronavirus. Proc. Natl. Acad. Sci. U.S.A. 2004;101(44):15748–15753. doi: 10.1073/pnas.0403812101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahn J.S., Roberts A., Weibel C., Buonocore L., Rose J.K. Replication-competent or attenuated, nonpropagating vesicular stomatitis viruses expressing respiratory syncytial virus (RSV) antigens protect mice against RSV challenge. J. Virol. 2001;75(22):11079–11087. doi: 10.1128/JVI.75.22.11079-11087.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ksiazek T.G., Erdman D., Goldsmith C.S., Zaki S.R., Peret T., Emery S., Tong S., Urbani C., Comer J.A., Lim W., Rollin P.E., Dowell S.F., Ling A.E., Humphrey C.D., Shieh W.J., Guarner J., Paddock C.D., Rota P., Fields B., DeRisi J., Yang J.Y., Cox N., Hughes J.M., LeDuc J.W., Bellini W.J., Anderson L.J. A novel coronavirus associated with severe acute respiratory syndrome. N. Engl. J. Med. 2003;348(20):1953–1966. doi: 10.1056/NEJMoa030781. [DOI] [PubMed] [Google Scholar]
- Lawson N.D., Stillman E.A., Whitt M.A., Rose J.K. Recombinant vesicular stomatitis viruses from DNA. Proc. Natl. Acad. Sci. U.S.A. 1995;92(10):4477–4481. doi: 10.1073/pnas.92.10.4477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li W., Moore M.J., Vasilieva N., Sui J., Wong S.K., Berne M.A., Somasundaran M., Sullivan J.L., Luzuriaga K., Greenough T.C., Choe H., Farzan M. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426(6965):450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu S., Xiao G., Chen Y., He Y., Niu J., Escalante C.R., Xiong H., Farmar J., Debnath A.K., Tien P., Jiang S. Interaction between heptad repeat 1 and 2 regions in spike protein of SARS-associated coronavirus: implications for virus fusogenic mechanism and identification of fusion inhibitors. Lancet. 2004;363(9413):938–947. doi: 10.1016/S0140-6736(04)15788-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marra M.A., Jones S.J., Astell C.R., Holt R.A., Brooks-Wilson A., Butterfield Y.S., Khattra J., Asano J.K., Barber S.A., Chan S.Y., Cloutier A., Coughlin S.M., Freeman D., Girn N., Griffith O.L., Leach S.R., Mayo M., McDonald H., Montgomery S.B., Pandoh P.K., Petrescu A.S., Robertson A.G., Schein J.E., Siddiqui A., Smailus D.E., Stott J.M., Yang G.S., Plummer F., Andonov A., Artsob H., Bastien N., Bernard K., Booth T.F., Bowness D., Czub M., Drebot M., Fernando L., Flick R., Garbutt M., Gray M., Grolla A., Jones S., Feldmann H., Meyers A., Kabani A., Li Y., Normand S., Stroher U., Tipples G.A., Tyler S., Vogrig R., Ward D., Watson B., Brunham R.C., Krajden M., Petric M., Skowronski D.M., Upton C., Roper R.L. The Genome sequence of the SARS-associated coronavirus. Science. 2003;300(5624):1399–1404. doi: 10.1126/science.1085953. [DOI] [PubMed] [Google Scholar]
- Peiris J.S., Guan Y., Yuen K.Y. Severe acute respiratory syndrome. Nat. Med. 2004;10(12 Suppl):S88–S97. doi: 10.1038/nm1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pope M., Chung S.W., Mosmann T., Leibowitz J.L., Gorczynski R.M., Levy G.A. Resistance of naive mice to murine hepatitis virus strain 3 requires development of a Th1, but not a Th2, response, whereas pre-existing antibody partially protects against primary infection. J. Immunol. 1996;156(9):3342–3349. [PubMed] [Google Scholar]
- Qu D., Zheng B., Yao X., Guan Y., Yuan Z.H., Zhong N.S., Lu L.W., Xie J.P., Wen Y.M. Intranasal immunization with inactivated SARS-CoV (SARS-associated coronavirus) induced local and serum antibodies in mice. Vaccine. 2005;23(7):924–931. doi: 10.1016/j.vaccine.2004.07.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts A., Kretzschmar E., Perkins A.S., Forman J., Price R., Buonocore L., Kawaoka Y., Rose J.K. Vaccination with a recombinant vesicular stomatitis virus expressing an influenza virus hemagglutinin provides complete protection from influenza virus challenge. J. Virol. 1998;72(6):4704–4711. doi: 10.1128/jvi.72.6.4704-4711.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts A., Buonocore L., Price R., Forman J., Rose J.K. Attenuated vesicular stomatitis viruses as vaccine vectors. J. Virol. 1999;73(5):3723–3732. doi: 10.1128/jvi.73.5.3723-3732.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts A., Reuter J.D., Wilson J.H., Baldwin S., Rose J.K. Complete protection from papillomavirus challenge after a single vaccination with a vesicular stomatitis virus vector expressing high levels of L1 protein. J. Virol. 2004;78(6):3196–3199. doi: 10.1128/JVI.78.6.3196-3199.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rose J.K., Whitt M.A. Rhabdoviridae: the viruses and their replication. In: Knipe D.M., Howley P.M., Griffin D.E., Lamb R.A., Martin M.A., Roizman B., Straus S.E., editors. Fields' Virology. 4th ed. Lippincott Williams and Wilkins; Philadelphia: 2001. [Google Scholar]
- Rose N.F., Marx P.A., Luckay A., Nixon D.F., Moretto W.J., Donahoe S.M., Montefiori D., Roberts A., Buonocore L., Rose J.K. An effective AIDS vaccine based on live attenuated vesicular stomatitis virus recombinants. Cell. 2001;106(5):539–549. doi: 10.1016/s0092-8674(01)00482-2. [DOI] [PubMed] [Google Scholar]
- Rota P.A., Oberste M.S., Monroe S.S., Nix W.A., Campagnoli R., Icenogle J.P., Penaranda S., Bankamp B., Maher K., Chen M.H., Tong S., Tamin A., Lowe L., Frace M., DeRisi J.L., Chen Q., Wang D., Erdman D.D., Peret T.C., Burns C., Ksiazek T.G., Rollin P.E., Sanchez A., Liffick S., Holloway B., Limor J., McCaustland K., Olsen-Rasmussen M., Fouchier R., Gunther S., Osterhaus A.D., Drosten C., Pallansch M.A., Anderson L.J., Bellini W.J. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science. 2003;300(5624):1394–1399. doi: 10.1126/science.1085952. [DOI] [PubMed] [Google Scholar]
- Schlereth B., Rose J.K., Buonocore L., ter Meulen V., Niewiesk S. Successful vaccine-induced seroconversion by single-dose immunization in the presence of measles virus-specific maternal antibodies. J. Virol. 2000;74(10):4652–4657. doi: 10.1128/jvi.74.10.4652-4657.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schnell M.J., Buonocore L., Whitt M.A., Rose J.K. The minimal conserved transcription stop-start signal promotes stable expression of a foreign gene in vesicular stomatitis virus. J. Virol. 1996;70(4):2318–2323. doi: 10.1128/jvi.70.4.2318-2323.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subbarao K., McAuliffe J., Vogel L., Fahle G., Fischer S., Tatti K., Packard M., Shieh W.J., Zaki S., Murphy B. Prior infection and passive transfer of neutralizing antibody prevent replication of severe acute respiratory syndrome coronavirus in the respiratory tract of mice. J. Virol. 2004;78(7):3572–3577. doi: 10.1128/JVI.78.7.3572-3577.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takasuka N., Fujii H., Takahashi Y., Kasai M., Morikawa S., Itamura S., Ishii K., Sakaguchi M., Ohnishi K., Ohshima M., Hashimoto S., Odagiri T., Tashiro M., Yoshikura H., Takemori T., Tsunetsugu-Yokota Y. A subcutaneously injected UV-inactivated SARS coronavirus vaccine elicits systemic humoral immunity in mice. Int. Immunol. 2004;16(10):1423–1430. doi: 10.1093/intimm/dxh143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang L., Zhu Q., Qin E., Yu M., Ding Z., Shi H., Cheng X., Wang C., Chang G., Fang F., Chang H., Li S., Zhang X., Chen X., Yu J., Wang J., Chen Z. Inactivated SARS-CoV vaccine prepared from whole virus induces a high level of neutralizing antibodies in BALB/c mice. DNA Cell Biol. 2004;23(6):391–394. doi: 10.1089/104454904323145272. [DOI] [PubMed] [Google Scholar]
- Vennema H., de Groot R.J., Harbour D.A., Dalderup M., Gruffydd-Jones T., Horzinek M.C., Spaan W.J. Early death after feline infectious peritonitis virus challenge due to recombinant vaccinia virus immunization. J. Virol. 1990;64(3):1407–1409. doi: 10.1128/jvi.64.3.1407-1409.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weingartl H., Czub M., Czub S., Neufeld J., Marszal P., Gren J., Smith G., Jones S., Proulx R., Deschambault Y., Grudeski E., Andonov A., He R., Li Y., Copps J., Grolla A., Dick D., Berry J., Ganske S., Manning L., Cao J. Immunization with modified vaccinia virus Ankara-based recombinant vaccine against severe acute respiratory syndrome is associated with enhanced hepatitis in ferrets. J. Virol. 2004;78(22):12672–12676. doi: 10.1128/JVI.78.22.12672-12676.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wesseling J.G., Godeke G.J., Schijns V.E., Prevec L., Graham F.L., Horzinek M.C., Rottier P.J. Mouse hepatitis virus spike and nucleocapsid proteins expressed by adenovirus vectors protect mice against a lethal infection. J. Gen. Virol. 1993;74(Pt. 10):2061–2069. doi: 10.1099/0022-1317-74-10-2061. [DOI] [PubMed] [Google Scholar]
- Xiong S., Wang Y.F., Zhang M.Y., Liu X.J., Zhang C.H., Liu S.S., Qian C.W., Li J.X., Lu J.H., Wan Z.Y., Zheng H.Y., Yan X.G., Meng M.J., Fan J.L. Immunogenicity of SARS inactivated vaccine in BALB/c mice. Immunol. Lett. 2004;95(2):139–143. doi: 10.1016/j.imlet.2004.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Z.Y., Kong W.P., Huang Y., Roberts A., Murphy B.R., Subbarao K., Nabel G.J. A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. Nature. 2004;428(6982):561–564. doi: 10.1038/nature02463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan K., Yi L., Chen J., Qu X., Qing T., Rao X., Jiang P., Hu J., Xiong Z., Nie Y., Shi X., Wang W., Ling C., Yin X., Fan K., Lai L., Ding M., Deng H. Suppression of SARS-CoV entry by peptides corresponding to heptad regions on spike glycoprotein. Biochem. Biophys. Res. Commun. 2004;319(3):746–752. doi: 10.1016/j.bbrc.2004.05.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang C.H., Guo Z.M., Zheng H.Y., Lu J.H., Wang Y.F., Yan X.G., Zhao Y., Du X.W., Zhang X., Fang L., Ling W.H., Qi S.Y., Yu X.B., Zhong N.S. Humoral immune responses in rabbits induced by an experimental inactivated severe acute respiratory syndrome coronavirus vaccine prepared from F69 strain. Chin. Med. J. (Engl) 2004;117(11):1625–1629. [PubMed] [Google Scholar]