

Abstract
We previously demonstrated the presence of IgE directed to fetal calf serum (FCS) in the sera from dogs that developed allergic reactions after vaccination. In this study, by an immunoblot analysis, we investigated the IgE-reactive components of FCS using sera from 16 dogs that exhibited allergic reactions after vaccination. The immunoblot analysis revealed that several FCS proteins of approximately 25-, 50-, 66-, 75-, 120-, and 175-kDa strongly reacted with IgE in the sera from dogs that showed post-vaccination allergic reactions. The 66-kDa band was detected in the sera from 14 of the 16 dog serum samples analyzed in the immunoblot analysis for FCS, and it was speculated to be albumin based on its molecular weight; however, serum IgE reactivity to bovine serum albumin could be detected in only four of the 14 dog samples. These findings demonstrated that a variety of FCS components including albumin could function as allergens in dogs that developed allergic reactions after vaccination.
Keywords: Dog, Vaccine, Allergy, IgE, FCS
1. Introduction
Allergic reactions induced by vaccine injections are a problematic event in small animal practice; these reactions are regarded as one of the adverse events occurring after vaccination (Brooks, 1991, Gaskell et al., 2002). The clinical signs of possible allergic reactions after vaccination in dogs are classified into two groups. The first group includes cardiovascular and/or respiratory signs such as circulatory collapse, cyanosis, hypotension, hypothermia, and dyspnea, which constitute anaphylaxis, whereas the second group includes dermatological signs such as facial edema, pruritus, and erythema (Ohmori et al., 2005b). In addition, some dogs were reported to die from the severe anaphylaxis after vaccination (Ohmori et al., 2002, Ohmori et al., 2005b). Allergic reactions after vaccination, therefore, have been considered as an important issue that needs to be resolved in veterinary practice.
Our previous study showed that IgE-mediated type I hypersensitivity was involved in the development of immediate-type allergic reactions after vaccination in dogs; this was indicated by the presence of high levels of IgE directed to the injected vaccines in the sera from dogs that developed allergic reactions after the vaccination (Ohmori et al., 2005a). Furthermore, it was observed that in most of the dogs that exhibited immediate-type allergic reactions, serum IgE was directed to fetal calf serum (FCS) (Ohmori et al., 2005a). It was also revealed that a large amount of bovine serum albumin (BSA) was included in the commercially available vaccines for dogs, which could be derived from FCS and/or BSA that are used for the growth of virus strains and leptospira included in vaccines (Ohmori et al., 2005a). These results indicated that FCS was a possible component to induce allergic reactions after vaccination in dogs.
FCS contains a variety of proteins; however, the types of components that induce IgE-mediated type I hypersensitivity after vaccination in dogs have not been clarified. Therefore, in the present study, by using an immunoblot analysis, we investigated the FCS components that reacted with IgE in the sera from dogs that developed allergic reactions after vaccination.
2. Materials and methods
2.1. Dogs
Serum samples from 16 dogs that had developed allergic reactions including cardiovascular and/or respiratory signs (circulatory collapse) as well as dermatological signs (facial edema, pruritus, and/or erythema) after non-rabies vaccination were collected by the cooperation of small animal practitioners in Japan. In these dogs, serum IgE directed to the injected vaccines was shown to be present by the procedure previously reported by us (Ohmori et al., 2005a). As negative controls, serum samples from 50 healthy dogs that had not developed any adverse reactions after vaccination were collected.
2.2. Vaccines
The commercially available non-rabies vaccines for dogs in Japan were classified into the following four groups: monovalent live canine parvovirus vaccine (group 1); monovalent inactivated canine parvovirus or leptospira vaccine (group 2); combined live vaccines composed of canine parvovirus, canine distemper virus, canine adenovirus type 2, and/or canine parainfluenza virus (group 3); combined live (canine parvovirus, canine distemper virus, canine adenovirus type 2, canine parainfluenza virus, and/or canine coronavirus) and inactivated (canine coronavirus and/or leptospira) vaccines (group 4).
2.3. Fluorometric indirect ELISA for detection of serum IgE directed to the injected vaccines and FCS
Serum specific IgE directed to the injected vaccines and FCS was analyzed by a fluorometric ELISA system by using 1% cold water fish gelatin (Sigma–Aldrich) in PBS containing 0.05% Tween 20 as a blocking buffer and a mouse monoclonal anti-dog IgE antibody (DeBoer et al., 1993) as a secondary antibody according to the procedure reported by us previously (Sakaguchi et al., 1997, Sakaguchi et al., 2001, Ohmori et al., 2005a). Serum IgE level was defined as positive when the fluorescence unit (FU) was higher than the threshold FU, which was obtained from the samples of healthy control dogs. The threshold FU was calculated from the mean value of FU plus three times of the standard deviation (S.D.) obtained from the IgE levels directed to the injected vaccines in 50 control dogs.
2.4. SDS-PAGE and immunoblot analysis
Heat-inactivated FCS (at 56 °C, for 30 min; JRH Biosciences, Lenexa, KS) and purified bovine proteins including BSA, bovine IgG, transferrin and fibronectin (all purchased from Sigma–Aldrich) were suspended in an SDS-PAGE sample buffer (Laemmli sample buffer, Bio-Rad Laboratories, Hercules, CA) containing β-mercaptoethanol, and boiled at 100 °C for 5 min. The samples (FCS: 2 μg/lane and BSA, bovine IgG, transferrin and fibronectin: 1 μg/lane) were subjected to SDS-PAGE through a 5–20% gradient polyacrylamide gel (ATTO, Tokyo, Japan). The protein concentration of FCS was measured by the Bradford method standardized with BSA. After the electrophoresis, the gel was stained with the Coomassie brilliant blue G-250 staining solution (Bio-Rad Laboratories). In the immunoblot analysis, the gel was electronically transferred onto a polyvinylidene difluoride (PDVF) membrane (Amersham Biosciences, Piscataway, NJ). The membrane was then blocked with 1% cold water fish gelatin (Sigma–Aldrich) in PBS containing 0.1% Tween 20 (PBST) at 4 °C overnight. After washing with PBST, the membrane was cut into strips, and then incubated with diluted serum samples (1:10) in 1% gelatin PBST at room temperature (RT) for 3 h. After washing with PBST, the strips were incubated with an HRP-conjugated goat anti-dog IgE antibody (1:25,000) (Bethyl Laboratories, Montgomery, TX) in 1% gelatin PBST at RT for 1 h. After washing, the IgE-binding proteins on a membrane were visualized using Lumi-LightPLUS Western Blotting Substrate (Roche Applied Science, Indianapolis, IN) and captured using the ATTO Light Capture (ATTO).
3. Results and discussion
The clinical findings of the 16 dogs that developed allergic reactions after vaccination are shown in Table 1 . Of these 16 dogs, two (Nos. 1 and 2) showed cardiovascular and/or respiratory signs such as circulatory collapse, and the remaining 14 dogs (Nos. 3–16) manifested dermatological signs such as facial edema, pruritus, and erythema. Within 60 min after vaccination, seven dogs (Nos. 1–7) developed clinical signs; these were clinically classified as immediate-type allergic reactions. The remaining nine dogs (Nos. 8–16) exhibited clinical signs between 2 and 20 h after vaccination; these were clinically classified as nonimmediate-type allergic reactions based on a previous report (Ohmori et al., 2005b). The FU values of serum IgE directed to the injected vaccines in these 16 dogs ranged between 156 and 2952 FU (mean ± S.D.: 993.6 ± 717.2 FU), and these values were higher than the threshold FU obtained from 50 control dogs (0–96 FU, mean ± S.D.: 16.6 ± 27.9 FU, threshold: 100.3 FU). Moreover, in all 16 dogs, high levels of serum IgE directed to FCS were observed (1100–5385 FU, mean ± S.D.: 2613.3 ± 1228.6 FU).
Table 1.
Clinical findings and serum IgE reactivity to the injected vaccine and FCS in 16 dogs that developed allergic reactions after vaccination
| Dog number | Breed | Sex | Age | Number of previous vaccination | Onset time after vaccination | Clinical signs | Group of the injected vaccinea | Serum IgE reactivity (Fluorescence units) |
|
|---|---|---|---|---|---|---|---|---|---|
| Vaccine | FCS | ||||||||
| 1 | Shih tzu | F | 2 months | 0 | ≤5 min | Circulatory collapse | 1 and 3 | 639 | 2076 |
| 2 | Golden retriever | F | 5 years | ≥3 | ≤5 min | Circulatory collapse, vomiting, diarrhea | 4 | 382 | 4174 |
| 3 | Miniature dachshund | M | 3 months | Unknown | 30–60 min | Facial edema, pruritus | 4 | 1415 | 1366 |
| 4 | Pug | F | 1 years 3 months | ≥3 | 20 min | Facial edema | 4 | 493 | 2014 |
| 5 | Shetland sheepdog | M | Unknown | 1 | 60 min | Facial edema | 4 | 1517 | 4066 |
| 6 | Shih tzu | F | 2 years | ≥3 | 30 min | Facial edema | 4 | 156 | 2537 |
| 7 | Miniature dachshund | F | 2 years 6 months | ≥3 | 30 min | Facial edema, pruritus, vomiting | 4 | 415 | 1100 |
| 8 | Miniature dachshund | M | 5 years 5 months | ≥3 | 20 h | Facial edema | 4 | 766 | 3696 |
| 9 | Maltese | M | 2 years 4 months | ≥3 | 7 h | Facial edema, erythema, pruritus | 4 | 1041 | 2122 |
| 10 | Pug | M | 3 years 5 months | ≥3 | 7 h | Facial edema, pruritus, vomiting | 4 | 1429 | 3250 |
| 11 | Shetland sheepdog | M | 3 years 6 months | 2 | 2 h | Facial edema | 4 | 2952 | 5385 |
| 12 | Miniature dachshund | F | 1 years 2 months | ≥3 | 2 h | Facial edema | 4 | 389 | 1240 |
| 13 | Miniature dachshund | M | 2 years 3 months | ≥3 | 5 h | Facial edema | 3 | 1339 | 3412 |
| 14 | Yorkshire terrier | F | 5 months | 1 | 4–5 h | Facial edema, pruritus | 4 | 771 | 2625 |
| 15 | Miniature dachshund | F | 2 years | ≥3 | 2 h 30 min | Facial edema | 4 | 273 | 1446 |
| 16 | Beagle | F | 2 years 6 months | ≥3 | 10 h | Facial edema, vomiting | 4 | 1920 | 1303 |
Criteria for the grouping of vaccines are described in the materials and methods. F: female, M: male.
To investigate the allergenic component(s) of FCS, we carried out an immunoblot analysis using serum samples from the 16 dogs that had developed allergic reactions after vaccination. By SDS-PAGE, FCS was shown to contain several proteins including two major proteins that were observed at approximately 66- and 75-kDa (Fig. 1a). The 66-kDa protein was also identified as the most common IgE-binding protein in the immunoblot analysis of the serum of 14 of the 16 dogs (Dog Nos. 1–5 and 8–16) (Fig. 1b). In addition to the 66-kDa protein, several different types of FCS proteins, which strongly reacted with IgE in sera from the dogs, were detected at various molecular weights: 25-kDa (one dog: No.1), 50-kDa (one dog: No.1), 75-kDa (six dogs: Nos. 1, 6, 9, 10, 11, and 15), 120-kDa (three dogs: Nos. 6, 11, and 15) and 175-kDa (seven dogs: Nos. 6, 7, 11, 13, 14, 15, and 16) proteins (Fig. 1b). The sera from the two healthy control dogs did not react with any FCS proteins. These results suggest that the 66-kDa band obtained in the immunoblot analysis should contain the most common IgE-reactive component(s) of FCS in the dogs.
Fig. 1.
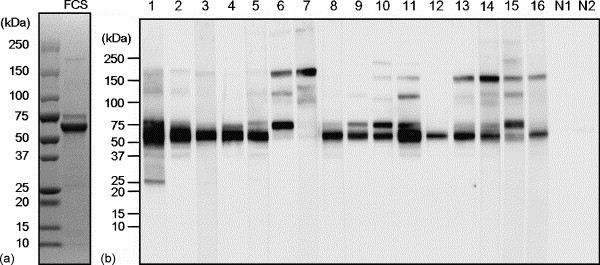
(a) SDS-PAGE analysis of FCS. An FCS sample (2 μg/lane) was subjected to SDS-PAGE through a 5–20% gradient polyacrylamide gel under reducing conditions, and the gel was then stained with Coomassie brilliant blue. Molecular weight size markers are indicated along the left side. (b) Immunoblot analysis of serum samples from dogs that developed post-vaccination allergic reactions to detect IgE directed to FCS components. Numbers 1–16 indicate the numbers of the dogs that exhibited allergic reactions after vaccination. N1 and N2 indicate the negative control dogs. Molecular weight size markers are indicated along the left side.
Based on the results of SDS-PAGE for FCS (Fig. 1a), the 66-kDa band that reacted with IgE in the immunoblot analysis (Fig. 1b) was suspected to be BSA, which is an FCS component. Therefore, we then examined IgE reactivity to purified BSA by using sera from the 14 dogs that were found to react with the 66-kDa band in the immunoblot analysis for FCS. Serum IgE directed to purified BSA was detected in four (Nos. 1–4) of the 14 dogs, but was neither detected in the remaining 10 dogs that showed post-vaccination allergic reactions nor in the two healthy control dogs (Fig. 2b). This finding indicated that the 66-kDa protein of FCS contained not only BSA but also other proteins that could be allergenic to the dogs. In addition, based on this result, it can be speculated that if we conducted an immunoblot inhibition assay for investigating the relationship between the 66-kDa protein of FCS and BSA in the four dogs, serum IgE directed to the 66-kDa protein of FCS would not be completely inhibited by BSA.
Fig. 2.
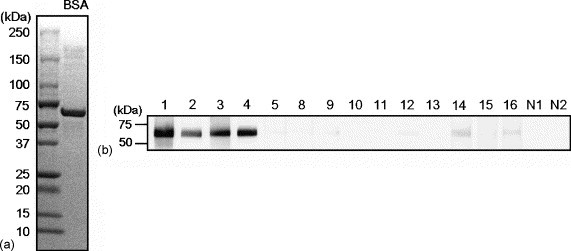
(a) SDS-PAGE analysis of BSA. A BSA sample (1 μg/lane) was subjected to SDS-PAGE through a 5–20% gradient polyacrylamide gel under reducing conditions, and the gel was then stained with Coomassie brilliant blue. Molecular weight size markers are indicated along the left side. (b) Immunoblot analysis of serum samples from dogs that developed post-vaccination allergic reactions to detect IgE directed to purified BSA. Numbers indicate the numbers of the 14 dogs in which IgE directed to the approximately 66-kDa protein detected in the immunoblot analysis of FCS was present. N1 and N2 indicate the negative control dogs. Molecular weight size markers are indicated along the left side.
Protein bands corresponding to molecular weights of approximately 25-, 50-, 66-, 75-, 120-, and 175-kDa were detected in the immunoblot analysis, which were performed to examine the IgE-binding FCS components; of these, the 25-, 50-, and 75-kDa bands were suspected to correspond to bovine IgG light chains, heavy chains, and transferrin, respectively, based on the molecular weights. In addition, it was reported that IgE directed to bovine fibronectin was shown in the sera form dogs after injection with a rabies vaccine (HogenEsch et al., 2002). Therefore, we further investigated serum IgE reactivity to the purified FCS components including bovine IgG, transferrin, and fibronectin by the immunoblot analysis. However, we did not detect any positive bands in these immunoblot analyses (data not shown). Since, IgE-reactive epitopes of these bovine proteins might have been destroyed by the denaturing step in the SDS-PAGE, further analysis using native antigen without denaturation is necessary to identify possible IgE-reactive FCS components that may be associated with allergic reactions after vaccination in dogs.
In the present study, in four of the 16 dogs, IgE directed to BSA was present in the sera. It was found that large amounts of BSA were included in all of the vaccines for dogs currently marketed in Japan (Ohmori et al., 2005a). Moreover, most of these vaccines are produced and marketed in the U.S. and Europe. It is highly plausible that the BSA in live vaccines is derived from FCS, which is routinely used in cell culture for growing virus strains that are included in vaccines for dogs, and the BSA in inactivated vaccines is derived from the culture media for leptospira. Therefore, it is conceivable that not only the FCS in live vaccines but also the BSA in inactivated leptospira vaccines can function as allergens that induce allergic reactions after vaccination in dogs.
Both serum IgE directed to the injected vaccines and FCS were detected not only in the sera of the seven dogs that showed allergic reactions within 60 min after vaccination but also in those of nine dogs that exhibited allergic reactions from 2–20 h after vaccination; the dogs of the latter group were clinically classified as nonimmediate-type allergic reactions. In humans, it was shown that children who developed allergic reactions several hours after vaccination did not have serum IgE directed to the injected vaccines (Kumagai et al., 1997). In these children, T-lymphocyte-mediated immune reactions were found to be involved in the development of nonimmeditate-type allergic reactions after vaccination (Kumagai et al., 1997). One of the possible explanations for the result obtained in this study is that the dog owners or the veterinarians might have missed the initial clinical signs of allergic reactions soon after vaccination, although some private veterinarians did report the onset of allergic reactions 2–20 h after vaccination. Alternatively, IgE might have also been involved in the development of nonimmediate-type allergic reactions after vaccination in the dogs. It was recently reported that IgE also contributed to the development of delayed-onset allergic inflammation in mice through the basophil-mediated immune response (Mukai et al., 2005). To clarify the immunological mechanisms underlying the development of nonimmediate-type allergic reactions after vaccination in dogs, further studies are needed.
In humans, allergen-free vaccines have successfully reduced the incidence of allergic reactions after vaccination (Ozaki et al., 2005). The present study clearly indicates that a variety of FCS components including BSA, but not a single component, can function as allergens in dogs that developed allergic reactions after vaccination. Therefore, together with our previous study, which showed that stabilizer proteins such as gelatin and casein also functioned as allergens in vaccines for dogs (Ohmori et al., 2005a), it can be concluded that FCS, BSA, and stabilizer proteins should be eliminated from the live and inactivated components of vaccines for dogs in order to manufacture safe vaccines with very low incidence of post-vaccination allergic reactions.
Acknowledgements
The authors would like to thank the private practitioners who provided the serum samples and Dr. Morimasa Yamanaka, Kyoto Biken Laboratories, Inc. for his assistance in collecting the serum samples. This work was supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology, Japan, and by Research Fellowships of the Japan Society for the Promotion of Science for Young Scientists.
References
- Brooks R. Adverse reactions to canine and feline vaccines. Aust. Vet. J. 1991;68:342–344. doi: 10.1111/j.1751-0813.1991.tb03096.x. [DOI] [PubMed] [Google Scholar]
- DeBoer D.J., Ewing K.M., Schultz K.T. Production and characterization of mouse monoclonal antibodies directed against canine IgE and IgG. Vet. Immunol. Immunopathol. 1993;37:183–199. doi: 10.1016/0165-2427(93)90193-8. [DOI] [PubMed] [Google Scholar]
- Gaskell R.M., Gettinby G., Graham S.J., Skilton D. Veterinary products committee working group report on feline and canine vaccination. Vet. Rec. 2002;150:126–134. [PubMed] [Google Scholar]
- HogenEsch H., Dunham A.D., Scott-Moncrieff C., Glickman L.T., DeBoer D.J. Effect of vaccination on serum concentrations of total and antigen-specific immunoglobulin E in dogs. Am. J. Vet. Res. 2002;63:611–616. doi: 10.2460/ajvr.2002.63.611. [DOI] [PubMed] [Google Scholar]
- Kumagai T., Yamanaka T., Wataya Y., Umetsu A., Kawamura N., Ikeda K., Furukawa H., Kimura K., Chiba S., Saito S., Sugawara N., Kurimoto F., Sakaguchi M., Inouye S. Gelatin-specific humoral and cellular immune responses in children with immediate- and nonimmediate-type reactions to live measles, mumps, rubella, and varicella vaccines. J. Allergy Clin. Immunol. 1997;100:130–134. doi: 10.1016/s0091-6749(97)70204-5. [DOI] [PubMed] [Google Scholar]
- Mukai K., Matsuoka K., Taya C., Suzuki H., Yokozeki H., Nishioka K., Hirokawa K., Etori M., Yamashita M., Kubota T., Minegishi Y., Yonekawa H., Karasuyama H. Basophils play a critical role in the development of IgE-mediated chronic allergic inflammation independently of T cells and mast cells. Immunity. 2005;23:191–202. doi: 10.1016/j.immuni.2005.06.011. [DOI] [PubMed] [Google Scholar]
- Ohmori K., Masuda K., Sakaguchi M., Kaburagi Y., Ohno K., Tsujimoto H. A retrospective study on adverse reactions to canine vaccines in Japan. J. Vet. Med. Sci. 2002;64:851–853. doi: 10.1292/jvms.64.851. [DOI] [PubMed] [Google Scholar]
- Ohmori K., Masuda K., Maeda S., Kaburagi Y., Kurata K., Ohno K., Deboer D.J., Tsujimoto H., Sakaguchi M. IgE reactivity to vaccine components in dogs that developed immediate-type allergic reactions after vaccination. Vet. Immunol. Immunopathol. 2005;104:249–256. doi: 10.1016/j.vetimm.2004.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohmori K., Sakaguchi M., Kaburagi Y., Maeda S., Masuda K., Ohno K., Tsujimoto H. Suspected allergic reactions after vaccination in 85 dogs in Japan. Vet. Rec. 2005;156:87–88. doi: 10.1136/vr.156.3.87. [DOI] [PubMed] [Google Scholar]
- Ozaki T., Nishimura N., Muto T., Sugata K., Kawabe S., Goto K., Koyama K., Fujita H., Takahashi Y., Akiyama M. Safety and immunogenicity of gelatin-free varicella vaccine in epidemiological and serological studies in Japan. Vaccine. 2005;23:1205–1208. doi: 10.1016/j.vaccine.2004.08.040. [DOI] [PubMed] [Google Scholar]
- Sakaguchi M., Nakano T., Tsujimoto H., Sasaki Y., DeBoer D.J., Inouye S. Fluorometric ELISA for allergen-specific dog IgE antibody. Allergol. Int. 1997:207–212. [Google Scholar]
- Sakaguchi M., Masuda K., Yasueda H., Saito S., DeBoer D.J., Tsujimoto H. IgE reactivity and cross-reactivity to Japanese cedar (Cryptomeria japonica) and cypress (Chamaecyparis obtusa) pollen allergens in dogs with atopic dermatitis. Vet. Immunol. Immunopathol. 2001;83:69–77. doi: 10.1016/s0165-2427(01)00362-2. [DOI] [PubMed] [Google Scholar]