

Abstract
Obinutuzumab is a glycoengineered tumor-targeting anti-CD20 mAb with a modified crystallizable fragment (Fc) domain designed to increase the affinity for the FcγRIIIA/CD16 receptor, which was recently approved for clinical use in CLL and follicular lymphoma. Here we extend our previous observation that, in human NK cells, the sustained CD16 ligation by obinutuzumab-opsonized targets leads to a markedly enhanced IFN-γ production upon a subsequent cytokine re-stimulation. The increased IFN-γ competence in response to IL-2 or IL-15 is attributable to post-transcriptional regulation, as it does not correlate with the upregulation of IFN-γ mRNA levels. Different from the reference molecule rituximab, we observe that the stimulation with obinutuzumab promotes the upregulation of microRNA (miR)-155 expression. A similar trend was also observed in NK cells from untreated CLL patients stimulated with obinutuzumab-opsonized autologous leukemia. miR-155 upregulation associates with reduced levels of SHIP-1 inositol phosphatase, which acts in constraining PI3K-dependent signals, by virtue of its ability to mediate phosphatidylinositol 3,4,5-trisphosphate (PIP3) de-phosphorylation. Downstream of PI3K, the phosphorylation status of mammalian target of rapamycin (mTOR) effector molecule, S6, results in amplified response to IL-2 or IL-15 stimulation in obinutuzumab-experienced cells. Importantly, NK cell treatment with the PI3K or mTOR inhibitors, idelalisib and rapamycin, respectively, prevents the enhanced cytokine responsiveness, thus, highlighting the relevance of the PI3K/mTOR axis in CD16-dependent priming. The enhanced IFN-γ competence may be envisaged to potentiate the immunoregulatory role of NK cells in a therapeutic setting.
Electronic supplementary material
The online version of this article (10.1007/s00262-020-02482-2) contains supplementary material, which is available to authorized users.
Keywords: Obinutuzumab, CD16, Natural killer cells, IFN-γ, PI3K/mTOR, miR-155
Introduction
NK cells doubly contribute to the therapeutic efficacy of tumor-targeting mAbs; besides the killing of tumor cells mediated by the engagement of the low affinity receptor for IgG, FcγRIIIA/CD16, activated NK cells secrete pro-inflammatory cytokines and chemokines which act in boosting the recruitment and activation of other immune effector cells and the development of long-lasting T cell immunity [2, 3]. In particular, NK-derived IFN-γ stands as a key immunoregulatory factor in the shaping of anti-tumor adaptive immune responses, by promoting the maturation of DC and the subsequent development of Th1 and CTL responses [4]. Indeed, the current understanding of anti-tumor responses driven by tumor-targeting mAbs introduced a novel ground by which NK cells may contribute to the development of a vaccine-like effect required for the long-term protection of mAb-treated patients [5–9].
By virtue of an accessible Ifng locus, NK cells represent a prompt source of IFN-γ. Such cytokine is transcribed constitutively at low levels in NK cells; its increased production in response to cytokines or after the engagement of activating receptors is tightly regulated at transcriptional and post-transcriptional levels [10–12]. In this context, microRNA (miR)-155 functions as a positive regulator of IFN-γ production stimulated by CD16 and cytokines [13] by directly targeting the hematopoietic cell-specific inositol 5-phosphatase, SHIP-1, which negatively regulates the PI3K pathway [14]. Downstream PI3K, the master metabolic regulator mammalian target of rapamycin (mTOR) promotes IFN-γ translation through the phosphorylation of the ribosomal protein S6 kinase (S6K) and the eukaryotic translation initiation factor 4E (eIF4E)-binding protein 1 (4E-BP1) [15–18].
To reach an enhanced clinical efficacy, new mAbs with increased affinity for CD16 have been generated. Among them, obinutuzumab, recently approved for clinical use [19–21], is a type II glycoengineered anti-CD20 mAb with a defucosylated crystallizable fragment (Fc) domain that binds to a CD20 epitope in a different space orientation with respect to the reference molecule rituximab [22, 23].
Our recent studies have revealed that the strength of CD16 ligation by tumor-targeting mAbs impacts on receptor signaling and functional properties [24–26].
Here, extending our previous observations [25], we demonstrate that following obinutuzumab pre-stimulation, NK cells undergo enhanced IFN-γ production in response to a subsequent re-stimulation with common γ chain (γc) cytokines IL-15 or IL-2, which correlates to the upregulation of miR-155 and to reduced SHIP-1 levels but not with the upregulation of IFN-γ mRNA levels; the increased IFN-γ competence depends on the PI3K/mTOR axis.
Such data add mechanistic insights into NK cell plasticity in therapeutic settings. Moreover, taking into account the current research efforts focused on the development of IL-2 and IL-15 cytokine variants with extended half-life and targeted action [27], our results suggest that obinutuzumab-based immunotherapy in combination with NK cell-activating cytokines may achieve a useful synergism for the development of long-lasting curative anti-tumor responses.
Materials and methods
Antibodies
The following anti-CD20 mAbs were used: the chimeric IgG1κ rituximab, the humanized IgG1κ obinutuzumab (GA101), and its non-glycoengineered parental molecule, GA101 wild type (WT), all kindly provided by Roche Innovation Center Zurich (Schlieren, Switzerland).
For functional assays, the following mAbs were used: anti-2B4 (clone:C1.7, #IM1607, Beckman Coulter Life Science), anti-NKp46 (clone: 9E2, #331902, Biolegend), anti-natural killer group 2 member D (NKG2D) (clone: 149810, #MAB139, R&D Systems), all mouse IgG1 isotype, and goat F(ab')2 fragment anti-mouse IgG (H + L) (#115-006-003, Jackson ImmunoResearch Laboratories). The following fluorochrome-conjugated mAbs were used for flow cytometric analysis: anti-CD25 APC (clone:M-A2511, #555434) and anti-CD215 PE (clone:JM7A4, #566589) were from BD Biosciences; the anti-pS6 ribosomal protein (S235/236) PE (clone: D57.2.2E, #5316S) was from Cell Signaling Technology. For immunoblot analysis, antibodies were obtained from the following sources: anti-SHIP-1 (clone:P1C1, #sc-8425) from Santa Cruz Biotechnology Inc); the anti-phospho-STAT4 (Tyr693) (clone:D2E4, #4134), anti-STAT4 (clone:C46B10, #2653), anti-Src homology 2 domain-containing leukocyte protein of 76 kDa (SLP-76) (#4958) and anti-Akt (#9272), all from Cell Signaling Technology.
Patients and healthy donors
PBMCs were obtained from anonymized healthy donors of Transfusion Center of Sapienza University (Rome, Italy) or CLL patients of Hematology Unit, S. Maria Goretti Hospital (Latina, Italy).
The diagnosis of CLL was based on criteria recommended by the International Workshop on Chronic Lymphocytic Leukemia (IWCLL); the stage of disease was assessed according to the Rai staging system [24]. In all specimens, the percentage of CD5+CD19+ CLL cells was more than 70%. From each patient, part of PBMC was used to obtain primary cultured NK cells (see below) and the rest was cryopreserved and stored at − 160 °C. The day before the experiment, thawed samples were rested overnight in complete medium. Only samples with viability more than 90% were used.
Cell system
Primary cultured human NK cells were obtained from healthy donors [28], or from CLL patients as previously described [24]. Briefly, PBMC were co-cultured for 10 days with irradiated (3000 rad) Epstein–Barr virus positive RPMI 8866 lymphoblastoid cell line in 10% FCS and 1% l-glutamine containing RPMI 1640 (all from Euroclone). Experiments were performed on NK cell cultures which were more than 80% pure (CD3−CD56+).
Anti-CD20-experienced NK cell preparation and purification
The human CD20+ lymphoblastoid Raji cell line or primary B-CLL cells were loaded with 10 μγ/ml of EZ-Link Sulfo-NHS-SS-Biotin (#21331, Thermo Fisher Scientific) for 30 min at room temperature [29], washed twice with PBS (Euroclone) and then opsonized for 20 min at room temperature with saturating doses of rituximab, obinutuzumab or obinutuzumab WT. Not opsonized and anti-CD20-opsonized targets were mixed at 1:2 ratio with primary cultured NK cells for 18 h, and then washed twice with cold 5 mM EDTA containing PBS. NK cells were purified by negative selection on biotin-binder Dynabeads (#11047), followed by anti-CD3-coated Dynabeads (#11151D) (both from Invitrogen, Life Technologies), according to the manufacturer’s protocols. Experienced NK populations were checked by flow cytometry to assess purity.
Evaluation of IFN-γ release
Purified experienced NK cells (5 × 105/ml) were resuspended in complete medium and left untreated or stimulated for 18 h at 37 °C with 500 U/ml of IL-2 (#200-02), or 10 ng/ml of IL-12 (#200-12), or 100 ng/ml of IL-15 (#200-15) (all from PeproTech). Where required, 10 μM idelalisib (CAL-101, GS1101; #S2226, Selleck Chemicals) or 20 nM rapamycin (#R0395, Merck) was added 2 h before cytokine treatment, and maintained until the end of stimulation. An equivalent volume of DMSO (#D5879, Merck), as vehicle, was added to control samples.
Stimulation of activating receptors was performed by plastic-immobilized goat F(abʹ)2 anti-mouse IgG (H + L) (1 μg/106) followed by anti-NKp46 (0.5 μg/106), anti-NKG2D (1 μg/106) or anti-2B4 (0.24 μg/106) mAbs, used alone or in combination. IFN-γ was quantified in the supernatants by commercial ELISA kit (#EHIFNG2, Thermo Fisher Scientific), according to the manufacturer’s instructions.
Stimulation and analysis of S6 phosphorylation
To determine the phosphorylation status of ribosomal protein S6, purified experienced NK cells were resuspended in complete medium, rested for 2 h at 37 °C and stimulated with 500 U/ml of IL-2 or 100 ng/ml of IL-15 at 37 °C for different lengths of time. Following stimulation, samples were washed, fixed and permeabilized with commercial kit (# 00-5523-00, eBioscience, Thermo Fisher Scientific), according to the manufacturer’s instructions, and stained with PE-conjugated anti-pS6 (S235/236) rabbit mAb. Samples were acquired on a FACSCanto II (BD Bioscience) and analyzed with FlowJo v.9.3.2 (TreeStar) software.
miR and mRNA analyses
Total RNA, including miRs, was extracted from highly purified experienced NK cells before and after cytokine stimulation using Total RNA Purification Kit (#3755, Norgen Biotek Corp). The concentration and purity of extracted RNA were determined spectrophotometrically using NanoDrop 2000 (Thermo Fisher Scientific). cDNA was generated by means of High-Capacity cDNA Reverse Transcription kit (#4374966) or TaqMan MicroRNA Reverse Transcription kit (#4366596), using RT primers specific for miR-155, miR-29a, RNU-44 and RNU-48. mRNA, and miR levels were determined by real-time quantitative PCR (RT-qPCR) using TaqMan gene expression or miR assays, according to the manufacturer’s instructions. All reagents were obtained from Applied Biosystem, Life Technologies (Thermo Fisher Scientific) and the list of all primers employed is reported below. Reactions were performed in 96-well plate format by StepOnePlus real-time PCR system and data obtained were analyzed by StepOne Software v2.3 (Applied Biosystem). Water (no template) was loaded as a negative control. mRNA and miR contents were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or RNU-44 and RNU-48 endogenous controls, respectively, and relative quantification was evaluated by the comparative cycling threshold (ΔΔCT) method. The fold change was calculated according to the formula 2−ΔΔCT, setting the control population to 1. The following TaqMan gene expression assays (#4331182) were used: Hs00989291_m1 IFNG, Hs00183290_m1 INPP5D, Hs00705164_s1 suppressor of cytokine signalling 1 (SOCS-1), Hs03929097_g1 GAPDH. The following TaqMan miRNA assays (#4427975) were used: hsa-miR-155 (#002623), hsa-miR-29a-5p (#002447), RNU-44 (#001094), RNU-48 (#001006) all conjugated with fluorochrome carboxyfluorescein (FAM).
Biochemical analysis
For assessing STAT4 phosphorylation, purified experienced NK populations were stimulated for 30 min at 37 °C with 500 U/ml of IL-2 or 100 ng/ml of IL-12, or left untreated, and lysed in radio-immunoprecipitation assay (RIPA) buffer (50 mM Tris–HCl, pH 7.5, 150 mM NaCl, 0.5% deoxycholate, 0.1% SDS, 1 mM EDTA, pH8, 1 mM ethylene glycol tetraacetic acid (EGTA), pH 8, 5 mM MgCl2, 5 mM NaF) supplemented with 1 mM phenylmethylsulfonyl fluoride, 1 mM Na3VO4, 1 μg/ml each of aprotinin and leupeptin inhibitors. To analyze SHIP-1, SLP-76 and Akt levels, whole cell lysates of purified experienced NK cells were obtained by incubating with 1% Triton X-100 lysis buffer (50 mM Tris–HCl, pH 7.5, 150 mM NaCl, 1 mM EGTA, pH 8, 1 mM MgCl2, 50 mM NaF) supplemented with protease and phosphatase inhibitors as above. Protein content was determined by Bradford Colorimetric Assay (#500-0006, Bio-Rad Laboratories) and equal amounts of proteins from each sample were separated by SDS-PAGE and transferred to nitrocellulose for immunoblot analysis. Quantification of specific bands was performed with ImageJ1.41o software (National Institutes of Health).
Statistical analysis
Wilcoxon matched-pairs signed rank or paired t tests were used to determine statistically significative differences (p value < 0.05) between two groups, as appropriate. Error bars represent the SEM. Analysis were performed using Prism v.6 (GraphPad Software).
Results
Pre-stimulation of CD16 with obinutuzumab-opsonized targets leads to an amplified IFN-γ production in response to IL-2 or IL-15, independently of transcriptional regulation
In line with our recent data demonstrating that the sustained CD16 pre-ligation in high-affinity conditions primes NK cells for IFN-γ production [25], we show here that primary cultured NK cells that were stimulated for 18 h with obinutuzumab-opsonized Raji cells (obinutuzumab-experienced NK cells) became able to secrete an increased amount of IFN-γ in response to high doses of IL-2 (500 U/ml) or IL-15 (100 ng/ml) and, to a lesser extent, of IL-12 (10 ng/ml), with respect to NK cells that were stimulated with rituximab-opsonized (rituximab-experienced NK cells) or not opsonized (control population) Raji cells, which behave comparably (Fig. 1a). The enhanced IFN-γ production was selectively observed in response to cytokine stimulation; indeed, we found a marked impairment of the ability to secrete IFN-γ in response to either ITAM-dependent (i.e., NKp46) or -independent (i.e., NKG2D and 2B4) receptors, alone or in combination, both in rituximab- and in obinutuzumab-experienced cells, with respect to control population (Fig. 1b).
Fig. 1.

Obinutuzumab-mediated CD16 pre-ligation induces an enhanced IFN-γ production in response to IL-2 or IL-15 stimulation. Primary cultured NK cells were immunomagnetically purified by negative selection upon 18 h of co-culture (2:1) with biotinylated rituximab (RTX-exp)-, obinutuzumab (GA101-exp)-opsonized or not opsonized Raji (Ctrl-exp). Experienced cells were left untreated (none) (a, b) or stimulated with (a) IL-2 (500 U/ml), IL-15 (100 ng/ml) or IL-12 (10 ng/ml) or with (b) the indicated plastic-immobilized mAbs. After 18 h, IFN-γ levels were measured in the collected supernatants. a Data are presented as mean ± SEM of n = 14 donors for IL-2 setting (***p = 0.0001), n = 11 donors for IL-15 setting (***p = 0.001, **p = 0.002) or n = 7 donors for IL-12 setting (*p = 0.0156) in five independent experiments. b Mean ± SEM from n = 3 donors are reported. c IL-2R α (CD25) or IL-15R α (CD215) expression was assessed by FACS analysis in purified experienced NK cells. Histogram overlays from one representative experiment are shown. Geometric MFI (gMFI) expressed as fold change with respect to Ctrl-exp population (set to 1) is reported. Data (mean ± SEM) from n = 3 donors are reported
To assess whether the increased IFN-γ production may be attributable to the modulation of specific cytokine receptor subunits, we evaluated IL-2 receptor (IL-2R) α-chain (CD25) and IL-15R α-chain (CD215) expression levels. While a robust upregulation of IL-2R α surface levels was observed in obinutuzumab- and, to a lesser extent, in rituximab-experienced cells, IL-15R α levels remained unaffected (Fig. 1c), thus indicating that the increased cytokine responsiveness does not completely rely on receptor affinity modulation.
To address the impact of cytokine stimulation on IFN-γ transcription, we analyzed IFN-γ mRNA levels. We observed that mRNA levels induced by IL-2 or IL-15 stimulation are comparable among all the experimental groups, indicating that the increased IFN-γ production in obinutuzumab-experienced cells is independent of transcriptional regulation. By contrast, IL-12 stimulation induces higher levels of IFN-γ transcript, in spite of comparable levels of IFN-γ release (Fig. 2a). In line with the necessary and sufficient role of STAT-4 phosphorylation for IL-12-induced IFN-γ transcription, we observed that IL-12 stimulation efficiently induces STAT-4 phosphorylation at comparable levels between all the experimental groups (Fig. 2b).
Fig. 2.
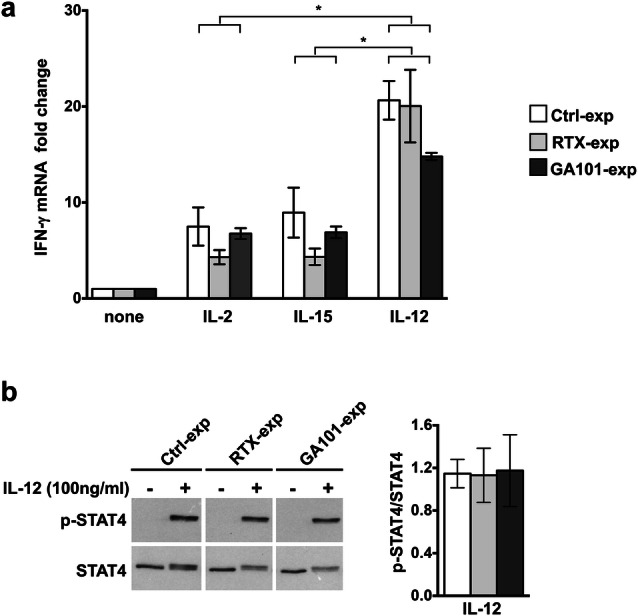
Cytokine-induced IFN-γ mRNA transcription in anti-CD20-experienced cells. a Purified experienced NK cells were obtained and stimulated with cytokines as in Fig. 1a. After 18 h, total RNA was extracted from untreated (none) or cytokine-activated NK cells and IFN-γ mRNA levels were measured by RT-qPCR. The fold change expression relative to untreated populations (set to 1) after normalizing with the GAPDH endogenous control is reported in the bar graph. Data (mean ± SEM) from n = 6 donors of three independent experiments are shown, *p = 0.0313. b Purified experienced NK cells were stimulated as indicated for 30 min. Western blot analysis of equal amounts of total lysates (4 × 105cells/point) were probed with anti-phospho-STAT4 (p-STAT4) followed by anti-STAT4 mAbs. All lanes were from the same experiment but were not contiguous. One representative experiment out of three performed is shown (left panels). Densitometric analysis of phosphorylated STAT4 (p-STAT4) normalized to total STAT4 in IL-12-stimulated samples is represented. Data (means ± SEM) from n = 3 donors (right panel)
Overall, these data demonstrate that in obinutuzumab-experienced cells the enhanced IFN-γ production in response to γc cytokine stimulation relies on post-transcriptional events.
miR-155 upregulation in obinutuzumab-experienced NK cells is associated with reduced SHIP-1 levels
We analyzed the levels of selected miRs previously involved in the regulation of IFN-γ production in NK cells [30, 31]. It is known that both miR-155 and miR-29a are poorly expressed in primary human NK cells and that their increased levels in activated cells act in the regulation of IFN-γ production [13, 32–34].
We quantified miR-155 and miR-29a by RT-qPCR by normalizing with RNU-44 endogenous control. Our data show that, upon 18 h of interaction with obinutuzumab-opsonized cells, miR-155 levels undergo significant upregulation, with respect to rituximab-stimulated cells or control population, which behaves comparably to unstimulated NK cells. Although at lower degree, but still significant with respect to control population, the upregulation of miR-155 was detected in rituximab-experienced cells. The observation that the fucosylated WT version of obinutuzumab, GA101WT, behaves similarly to rituximab, indicates that the strength of CD16 stimulation affects the levels of miR-155. On the contrary, no major modulation of miR-29a was observed (Fig. 3a).
Fig. 3.
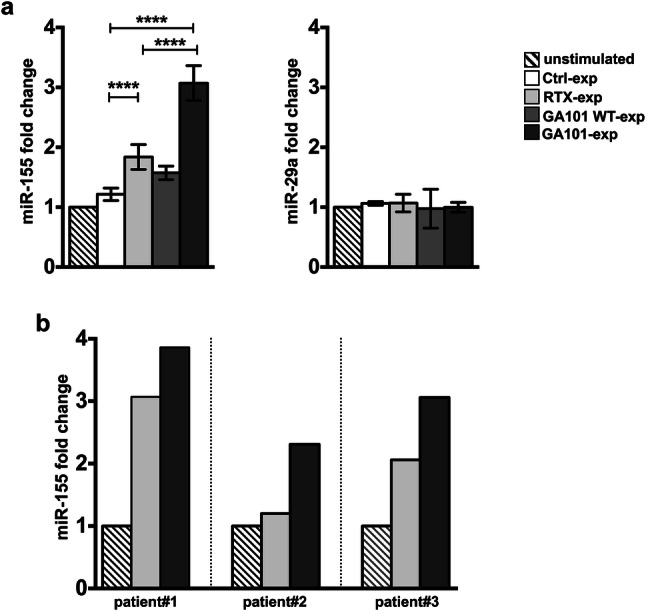
Obinutuzumab-mediated CD16 ligation upregulates miR-155 levels in NK cells from healthy donors and CLL patients. a Primary cultured NK cells were stimulated (2:1) for 18 h with biotinylated rituximab (RTX-exp)-, obinutuzumab WT (GA101 WT-exp)-, obinutuzumab (GA101-exp)-opsonized, not opsonized Raji (Ctrl-exp) or left unstimulated and immunomagnetically purified by negative selection, or b NK cells from three untreated CLL patients were stimulated with biotinylated rituximab (RTX-exp)-, obinutuzumab (GA101-exp)-opsonized autologous leukemia or left unstimulated and purified as in a. Relative miR-155 (a, b) or miR-29a (a) levels were measured by RT-qPCR. Bar graphs depict the fold change expression relative to the unstimulated population (set to 1) after normalizing with the RNU-44 endogenous control. a Data (mean ± SEM) from n = 17 donors from five independent experiments are shown. ****p < 0.0001
Remarkably, we found higher miR-155 levels in obinutuzumab- compared to rituximab-experienced NK cells also in primary NK cells derived from untreated CLL patients (Supplementary Table 1) stimulated for 18 h with anti-CD20-opsonized autologous leukemia (Fig. 3b). Similar results were obtained using RNU-48 as endogenous control (Supplementary Fig. 1).
Taking into account that SHIP-1 and SOCS-1 signaling intermediates, both relevant to IFN-γ production in NK cells, are known to be direct targets of miR-155 [13, 14, 32, 33], we assessed their expression levels in anti-CD20-experienced cells. We observed a significant reduction of SHIP-1 mRNA in obinutuzumab-, but not in rituximab- or GA101WT-experienced cells, with respect to control population (Fig. 4a, left). On the contrary, SOCS-1 mRNA levels were not modulated at significant levels (Fig. 4a, right).
Fig. 4.

SHIP-1 and SLP-76 downregulation in obinutuzumab-experienced NK cells. Primary cultured NK cells were stimulated (2:1) for 18 h with biotinylated rituximab (RTX-exp)-, obinutuzumab WT (GA101 WT-exp)-, obinutuzumab (GA101-exp)-opsonized or not opsonized Raji (Ctrl-exp) and immunomagnetically purified by negative selection. a Relative SHIP-1 or SOCS-1 mRNA levels were measured by RT-qPCR. The fold change expression relative to Ctrl-exp population (set to 1) after normalizing with the GAPDH endogenous control is reported in the bar graph. Data (mean ± SEM) from n = 15 donors of three independent experiments are shown. ****p < 0.0001, ***p = 0.0001. b Western blot analysis of equal amounts of total lysates. The same membrane was immunoblotted as indicated. One representative experiment is shown (left panels). The relative values of SHIP-1 or SLP-76 were obtained by normalizing to the levels of Akt and are expressed as fold change respect to control population (set to 1). Data (mean ± SEM, n = 8) from three independent experiments are depicted in bar graphs, **p = 0.0078, *p = 0.0156 (right panels)
Accordingly, we found a marked reduction of SHIP-1 at protein level of almost 65% in obinutuzumab-experienced cells, but not in rituximab- or GA101WT-stimulated samples, with respect to control population. A consensual downregulation of the signaling intermediate SLP-76, known to be a direct target of miR-155 too [32], was observed in obinutuzumab-experienced cells (Fig. 4b).
Collectively, these data demonstrate that the sustained CD16 ligation by means of obinutuzumab-opsonized targets induces the upregulation of miR-155 levels associated with the downregulation of both SHIP-1 and SLP-76.
Role of PI3K/mTOR pathway in the increased IFN-γ competence of obinutuzumab-experienced NK cells
The acute reduction of SHIP-1 in NK cells may lead to the amplification of PI3K-dependent signals, which is responsible for an increased IFN-γ production [35, 36].
To assess the functional role of PI3K in the priming of obinutuzumab-experienced cells, IL-2- or IL-15-dependent IFN-γ production was assessed in anti-CD20-experienced NK cells in the presence or absence of the p110δ PI3K inhibitor, idelalisib. As expected, the production of IFN-γ was reduced in the presence of inhibitor, although with a different sensitivity between populations; in fact, idelalisib completely counteracts the enhancement of IFN-γ production observed in obinutuzumab-experienced cells (Fig. 5a).
Fig. 5.

Amplified γc cytokine-dependent S6 phosphorylation and increased sensitivity to PI3K/mTOR inhibition in obinutuzumab-experienced NK cells. Primary cultured NK cells were stimulated (2:1) for 18 h with biotinylated rituximab (RTX-exp)-, obinutuzumab (GA101-exp)-opsonized (a–c) or not opsonized Raji (Ctrl-exp) (b) and immunomagnetically purified by negative selection. Experienced NK cells were treated with 10 μM idelalisib (a), 20 nM rapamycin (c) (white histograms) or with the same volume of DMSO as vehicle (grey histograms) for 2 h and then stimulated for additional 18 h with IL-2 (500 U/ml) or IL-15 (100 ng/ml), as indicated, in the presence of the inhibitors. Supernatants were then collected and assessed for IFN-γ levels. Data (mean ± SEM) from n = 8 donors from three independent experiments are depicted in bar graphs. ***p < 0.0008, **p < 0.0006, *p < 0.03. b Experienced NK cells were stimulated with IL-2 (500 U/ml) (top panels) or IL-15 (100 ng/ml) (bottom panels) for the indicated times. The phosphorylation levels of ribosomal protein S6 in S235/236 residues were evaluated by FACS analysis in fixed and permeabilized samples. Graphs depict for each time point the MFI value calculated as follow: (MFI of cytokine activated sample-MFI of untreated sample). Three representative donors and mean ± SEM from n = 6 donors are shown. *p = 0.031
Downstream of PI3K, mTOR acts in promoting mRNA translation [15]. We assessed ribosomal protein S6 phosphorylation in response to cytokine stimulation. The basal phosphorylation levels are comparable between all the experimental groups (Supplementary Fig. 2). In control population, the stimulation with IL-2 or IL-15 induces a time-dependent increase of S6 phosphorylation up to 60 min (Supplementary Fig. 3a). In obinutuzumab-experienced cells, in response to γc cytokines, we observed higher levels of S6 phosphorylation, particularly at later time points of stimulation, with respect to rituximab- or not opsonized target-experienced cells, which behave comparably (Fig. 5b).
The functional role of the mTOR/S6K pathway was evaluated by assessing IFN-γ production in the presence of the mTOR inhibitor, rapamycin. Rapamycin treatment inhibits, in a dose-dependent manner, cytokine-induced S6 phosphorylation (Supplementary Fig. 3b); by comparing the effect of rapamycin on anti-CD20-experienced populations, we observed a more pronounced inhibition of IFN-γ production in obinutuzumab- with respect to rituximab-experienced cells (Δ between untreated and treated cells is 1158 for GA101-exp vs 647 for RTX-exp for IL-2 setting; 1051 for GA101-exp vs 530 for RTX-exp for IL-15 setting) (Fig. 5c). Indeed, rapamycin prevents the enhanced response of obinutuzumab-experienced cells.
All together, these data indicate that the enhanced IFN-γ production in response to IL-2 or IL-15 stimulation relies on a potentiated PI3K/mTOR/S6K axis.
Discussion
We show here that the outcome of a sustained (18 h) interaction of NK cells with obinutuzumab-opsonized targets is an increased capability to produce IFN-γ in response to high doses of IL-2 or IL-15 which depends on a potentiated PI3K/mTOR pathway associated with miR-155 upregulation and SHIP-1 downregulation (Fig. 6) [1].
Fig. 6.

Obinutuzumab-experienced NK cells exhibit γc cytokine hyperresponsiveness. CD16-dependent miR-155 promotes SHIP-1 downregulation which in turn potentiates the PI3K/mTOR pathway responsible for enhanced IFN-γ translation
Recently, Pahl et al. demonstrated that CD16 aggregation by a tetravalent bispecific therapeutic antibody AFM13 (CD30/CD16A) followed by long-term re-stimulation with IL-2 or IL-15 results in a global NK cell hyperresponsiveness, attributable to increased cytokine receptor expression levels [37]. In our experimental conditions, we observed that the improved sensitivity of obinutuzumab-experienced cells to cytokine receptor stimulation coincided with increased CD25 expression levels, which, together with CD122 (IL-2Rβ) and CD132 (γc), assembles the trimeric high-affinity IL-2R [38], whereas IL-15R α chain, which by itself mediates the high-affinity binding [27, 39], remains unaffected upon CD16 stimulation. Such observations, along with the fully stimulatory doses of cytokines used, would exclude an increased receptor affinity as the unique mechanism responsible for the enhanced responsiveness.
The lack of correlation between IFN-γ protein and mRNA levels in γc cytokine-stimulated samples led us to hypothesize that post-transcriptional mechanisms may be responsible for the amplified response. In this context, our data confirm the existence of fundamental differences in the regulation of IFN-γ production induced by different cytokines [12, 40, 41]; in response to stimulation with IL-12, IFN-γ transcript increases almost 20-fold, whereas stimulation with IL-2 or IL-15 leads to a lower IFNG gene transcription, despite the efficient IFN-γ production. The stronger transcriptional effect of IL-12 is in line with a necessary role of STAT4 for maximal IL-12-mediated IFN-γ transcription [40, 41]. The increased IFN-γ competence is reminiscent of the recently described memory NK cell population which, in fact, exhibits enhanced IFN-γ production in response to CD16 stimulation [26]; however, memory NK cells display a defective responsiveness to IL-12 stimulation [42].
Several recent studies have demonstrated that IFN-γ production by NK cells is under the control of the metabolic sensor mTOR [16, 43, 44]. PI3K/Akt/mTOR pathway is an essential component of signaling via IL-15R and IL-2R [16, 45], with mTOR involved in translational activation through the phosphorylation of 4E-BP1 and S6K [15, 17, 18, 45].
In this context, our data evidence, in obinutuzumab-experienced cells, an amplified ribosomal protein S6 phosphorylation induced by γc cytokine stimulation along with a higher sensitivity to the mTOR inhibitor rapamycin, thus indicating that the increased IFN-γ competence depends on the mTOR pathway.
Based on these data, we reasoned that CD16 ligation in high-affinity conditions may lower the threshold for PI3K/mTOR pathway activation, thus leading to γc cytokine hyperresponsiveness.
SHIP-1 phosphatase acts in constraining PI3K/Akt-dependent signals [46] and negatively regulates IFN-γ production by monokines and CD16 stimulation in both human and mouse NK cells [35, 36]; its reduced expression levels in CD56bright with respect to CD56dim NK cells contribute to the increased IFN-γ producing potential of the former population [34]. SHIP-1, along with SOCS-1, are directly targeted and repressed by miR-155, which is involved in the regulation of IFN-γ production stimulated by cytokines in NK cells [13, 14, 32, 33].
On this line, we evidence that the upregulation of miR-155 transcript, in obinutuzumab-experienced cells, associates with SHIP-1 but not SOCS-1 downregulation; the observation that both rituximab and obinutuzumab WT stimulation induce still significant but lower levels of miR-155 indicates that the strength of CD16 ligation dictates the capability to promote miR-155 expression. On this issue, a recent paper highlighted a correlation between the strength of TCR stimulation and miR-155 expression levels that impacts IFN-γ production by anti-tumor CD8+ T cells [47].
SLP-76, a direct target of miR-155 in NK cells [32], is known to orchestrate the formation of a molecular platform critical in signal integration downstream activating receptors [2]. The reduction of SLP-76 levels, along with the degradation of FcεRIγ and CD3ξ adaptor chains and the Spleen-associated tyrosine kinase, Syk [25], may explain the profound impairment of IFN-γ production induced by the engagement of NKG2D and 2B4 activating receptors in anti-CD20-experienced cells. It should be noted that in a different experimental setting, based on shorter CD16 pre-ligation (90 min), we previously reported that the capability to produce IFN-γ in response to activating receptor stimulation was preserved [24]. In such conditions, no miR-155 upregulation and consequently no SLP-76 downmodulation have occurred (C. Capuano, unpublished observation).
The role of PI3K in the priming of obinutuzumab-experienced cells is strengthened by the observation that the inhibition of PI3K greatly counteracts γc cytokine hyperresponsiveness, while, as previously described, it has a more marginal effect on NK cell viability and cytotoxicity [48].
NK cell plasticity is being exploited by new NK cell-based intervention strategies against cancer [49–51]; by recapitulating the signaling pathway responsible for CD16-dependent priming, this study evidences new aspects of NK cell adaptation in a therapeutic setting. Taking into account that NK-derived IFN-γ is a key immunoregulatory factor in the shaping of anti-tumor adaptive immune responses, obinutuzumab-based therapy may be envisaged as a driver of a mAb-mediated vaccinal effect [5–9] that could significantly impact on long-term therapeutic efficacy. Suitable clinical trials focused on the prospective evaluation of the emergence of adaptive anti-tumor responses in obinutuzumab-treated patients are needed to highlight the actual translational impact.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgements
We are grateful to Prof. G. Cimino, Department of Translational and Precision Medicine, Cellular Biotechnology and Hematology, Hematology Unit (2U), S. Maria Goretti Hospital/ICOT, AUSL Latina, Italy, for CLL patients’ staging and recruitment. We also thank Dr. F.D. Batista (Ragon Institute of MGH, MIT and Harvard, Cambridge, MA, USA) for providing Raji cell line and P. Birarelli and B. Milana for their technical support.
Abbreviations
- 4E-BP
eukaryotic translation initiation factor 4E (eIF4E)-binding protein 1
- γc
common γ chain
- EGTA
ethylene glycol tetraacetic acid
- Fc
crystallizable fragment
- GAPDH
elyceraldehyde-3-phosphate dehydrogenase
- IL-2R
IL-2 receptor
- IL-15R
IL-15 receptor
- miR
microRNA
- mTOR
mammalian target of rapamycin
- NKG2D
Natural Killer group 2 member D
- PIP3
phosphatidylinositol 3,4,5-trisphosphate
- RT-qPCR
real-time quantitative PCR
- S6K
ribosomal protein S6 kinase
- SLP-76
src homology 2 domain-containing leukocyte protein of 76 kDa
- SOCS-1
suppressor of cytokine signaling 1
- WT
wild type
Author contributions
CC and CP designed the study, collected, analyzed and interpreted data and assisted to write the manuscript. RM and SB contributed to the experiments and to organize data. SM, GP, AS and CK contributed to data analysis and interpretation, and revised the manuscript. RG conceived and designed the study, analyzed and interpreted data and wrote the manuscript. All authors read and approved the final manuscript.
Funding
Cristina Capuano is supported by the Italian Ministry of University and Research (MIUR) (Grant number RBSI14022M). Simone Battella is a recipient of an Italian Association for Cancer Research (AIRC) fellowship. This work was supported by Roche Glycart AG, Schlieren, Switzerland.
Compliance with ethical standards
Conflict of interest
Christian Klein declares employment, patents and stock ownership with Roche Glycart AG, Schlieren, Switzerland. All other authors declare that they have no conflict of interests.
Ethical approval and ethical standards
All procedures involving human participants were in accordance with the ethical standards of the institutional research committee and with 1964 Declaration of Helsinki and its later amendments or comparable ethical standards. The study was approved by the Ethics Committee of Sapienza University of Rome (approval number 639/16 RIF/CE 4179).
Informed consent
All experiments were performed after obtaining written informed consent from patients and healthy donors. The informed consent was taken at the beginning of the study to use data and materials for research and publication of this study.
Cell line authentication
RPMI 8866 cell line was purchased by American Type Culture Collection, and Raji cell line was provided by Dr. F.D. Batista. Cells were authenticated (last testing May 2019) by morphology, growth and immunophenotypic characteristics, biologic behavior according to the provider recommendations, and routinely tested for mycoplasma contamination by EZ-PCR Mycoplasma test kit (#20–700-20, Biological Industries). All cell lines were kept in culture for less than two consecutive months.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Cristina Capuano and Chiara Pighi contributed equally to this work.
References
- 1.Pighi C, Capuano C, Maggio R et al (2019) CD16 aggregation in high affinity conditions by tumor-targeting mAb obinutuzumab promotes a PI3K/mTOR-dependent priming of Natural Killer cells for IFN-gamma production, associated to miR-155 upregulation. II Joint Meeting of the German Society for Immunology (DGfl) and the Italian Society of Immunology, Clinical Immunology and Allergology (SIICA). Eur J Immunol 49(1):271–272. 10.1002/eji.201970300 (Munich, September 10–13, 2019 [meeting abstract number P366]) [DOI]
- 2.Long EO, Kim HS, Liu D, Peterson ME, Rajagopalan S. Controlling natural killer cell responses: integration of signals for activation and inhibition. Annu Rev Immunol. 2013;31:227–258. doi: 10.1146/annurev-immunol-020711-075005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Morvan MG, Lanier LL. NK cells and cancer: you can teach innate cells new tricks. Nat Rev Cancer. 2016;16:7–19. doi: 10.1038/nrc.2015.5. [DOI] [PubMed] [Google Scholar]
- 4.Walzer T, Dalod M, Robbins SH, Zitvogel L, Vivier E. Natural-killer cells and dendritic cells: "l'union fait la force". Blood. 2005;106:2252–2258. doi: 10.1182/blood-2005-03-1154. [DOI] [PubMed] [Google Scholar]
- 5.Srivastava RM, Lee SC, Andrade Filho PA, et al. Cetuximab-activated natural killer and dendritic cells collaborate to trigger tumor antigen-specific T-cell immunity in head and neck cancer patients. Clin Cancer Res. 2013;19:1858–1872. doi: 10.1158/1078-0432.CCR-12-2426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Deligne C, Metidji A, Fridman WH, Teillaud JL. Anti-CD20 therapy induces a memory Th1 response through the IFN-γ/IL-12 axis and prevents protumor regulatory T-cell expansion in mice. Leukemia. 2015;29:947–957. doi: 10.1038/leu.2014.275. [DOI] [PubMed] [Google Scholar]
- 7.Battella S, Cox MC, Santoni A, Palmieri G. Natural killer (NK) cells and anti-tumor therapeutic mAb: unexplored interactions. J Leukoc Biol. 2016;99:87–96. doi: 10.1189/jlb.5VMR0415-141R. [DOI] [PubMed] [Google Scholar]
- 8.Cheadle EJ, Lipowska-Bhalla G, Dovedi SJ, et al. A TLR7 agonist enhances the antitumor efficacy of obinutuzumab in murine lymphoma models via NK cells and CD4 T cells. Leukemia. 2017;31:2278. doi: 10.1038/leu.2017.218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Muntasell A, Cabo M, Servitja S, et al. Interplay between natural killer cells and anti-HER2 antibodies: perspectives for breast cancer immunotherapy. Front Immunol. 2017;8:1544. doi: 10.3389/fimmu.2017.01544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hatton RD, Harrington LE, Luther RJ, et al. A distal conserved sequence element controls Ifng gene expression by T cells and NK cells. Immunity. 2006;25:717–729. doi: 10.1016/j.immuni.2006.09.007. [DOI] [PubMed] [Google Scholar]
- 11.Luetke-Eversloh M, Cicek BB, Siracusa F, et al. NK cells gain higher IFN-γ competence during terminal differentiation. Eur J Immunol. 2014;44:2074–2084. doi: 10.1002/eji.201344072. [DOI] [PubMed] [Google Scholar]
- 12.Mah AY, Cooper MA. Metabolic regulation of natural killer Cell IFN-γ production. Crit Rev Immunol. 2016;36:131–147. doi: 10.1615/CritRevImmunol.2016017387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Trotta R, Chen L, Ciarlariello D, et al. miR-155 regulates IFN-γ production in natural killer cells. Blood. 2012;119:3478–3485. doi: 10.1182/blood-2011-12-398099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.O'Connell RM, Chaudhuri AA, Rao DS, Baltimore D. Inositol phosphatase SHIP1 is a primary target of miR-155. Proc Natl Acad Sci USA. 2009;106:7113–7118. doi: 10.1073/pnas.0902636106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Laplante M, Sabatini DM. mTOR signaling in growth control and disease. Cell. 2012;149:274–293. doi: 10.1016/j.cell.2012.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Marçais A, Cherfils-Vicini J, Viant C, et al. The metabolic checkpoint kinase mTOR is essential for IL-15 signaling during the development and activation of NK cells. Nat Immunol. 2014;15:749–757. doi: 10.1038/ni.2936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ali AK, Nandagopal N, Lee SH. IL-15-PI3K-AKT-mTOR: a critical pathway in the life journey of natural killer cells. Front Immunol. 2015;6:355. doi: 10.3389/fimmu.2015.00355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yang M, Li D, Chang Z, Yang Z, Tian Z, Dong Z. PDK1 orchestrates early NK cell development through induction of E4BP4 expression and maintenance of IL-15 responsiveness. J Exp Med. 2015;212:253–265. doi: 10.1084/jem.20141703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Goede V, Fischer K, Busch R, et al. Obinutuzumab plus chlorambucil in patients with CLL and coexisting conditions. N Engl J Med. 2014;370:1101–1110. doi: 10.1056/NEJMoa1313984. [DOI] [PubMed] [Google Scholar]
- 20.Sehn LH, Chua N, Mayer J, et al. Obinutuzumab plus bendamustine versus bendamustine monotherapy in patients with rituximab-refractory indolent non-Hodgkin lymphoma (GADOLIN): a randomised, controlled, open-label, multicentre, phase 3 trial. Lancet Oncol. 2016;17:1081–1093. doi: 10.1016/S1470-2045(16)30097-3. [DOI] [PubMed] [Google Scholar]
- 21.Marcus R, Davies A, Ando K, et al. Obinutuzumab for the first-line treatment of follicular lymphoma. N Engl J Med. 2017;377:1331–1344. doi: 10.1056/NEJMoa1614598. [DOI] [PubMed] [Google Scholar]
- 22.Klein C, Lammens A, Schäfer W, et al. Epitope interactions of monoclonal antibodies targeting CD20 and their relationship to functional properties. MAbs. 2013;5:22–33. doi: 10.4161/mabs.22771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gagez AL, Cartron G. Obinutuzumab: a new class of anti-CD20 monoclonal antibody. Curr Opin Oncol. 2014;26:484–491. doi: 10.1097/CCO.0000000000000107. [DOI] [PubMed] [Google Scholar]
- 24.Capuano C, Romanelli M, Pighi C, et al. Anti-CD20 therapy acts via FcγRIIIA to diminish responsiveness of human natural killer cells. Cancer Res. 2015;75:4097–4108. doi: 10.1158/0008-5472.CAN-15-0781. [DOI] [PubMed] [Google Scholar]
- 25.Capuano C, Pighi C, Molfetta R, et al. Obinutuzumab-mediated high-affinity ligation of FcγRIIIA/CD16 primes NK cells for IFNγ production. Oncoimmunology. 2017;6:e1290037. doi: 10.1080/2162402X.2017.1290037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Capuano C, Battella S, Pighi C, et al. Tumor-targeting anti-CD20 antibodies mediate in vitro expansion of memory natural killer cells: impact of CD16 affinity ligation conditions and in vivo priming. Front Immunol. 2018;9:1031. doi: 10.3389/fimmu.2018.01031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Leonard WJ, Lin JX, O'Shea JJ. The γc family of cytokines: basic biology to therapeutic ramifications. Immunity. 2019;50:832–850. doi: 10.1016/j.immuni.2019.03.028. [DOI] [PubMed] [Google Scholar]
- 28.Capuano C, Paolini R, Molfetta R, Frati L, Santoni A, Galandrini R. PIP2-dependent regulation of Munc13-4 endocytic recycling: impact on the cytolytic secretory pathway. Blood. 2012;119:2252–2262. doi: 10.1182/blood-2010-12-324160. [DOI] [PubMed] [Google Scholar]
- 29.Molfetta R, Quatrini L, Capuano C, et al. c-Cbl regulates MICA- but not ULBP2-induced NKG2D down-modulation in human NK cells. Eur J Immunol. 2014;44:2761–2770. doi: 10.1002/eji.201444512. [DOI] [PubMed] [Google Scholar]
- 30.Beaulieu AM, Bezman NA, Lee JE, Matloubian M, Sun JC, Lanier LL. MicroRNA function in NK-cell biology. Immunol Rev. 2013;253:40–52. doi: 10.1111/imr.12045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sullivan RP, Leong JW, Fehniger TA. MicroRNA regulation of natural killer cells. Front Immunol. 2013;4:44. doi: 10.3389/fimmu.2013.00044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sullivan RP, Fogel LA, Leong JW, et al. MicroRNA-155 tunes both the threshold and extent of NK cell activation via targeting of multiple signaling pathways. J Immunol. 2013;191:5904–5913. doi: 10.4049/jimmunol.1301950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zawislak CL, Beaulieu AM, Loeb GB, et al. Stage-specific regulation of natural killer cell homeostasis and response against viral infection by microRNA-155. Proc Natl Acad Sci USA. 2013;110:6967–6972. doi: 10.1073/pnas.1304410110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Steiner DF, Thomas MF, Hu JK, et al. MicroRNA-29 regulates T-box transcription factors and interferon-γ production in helper T cells. Immunity. 2011;35:169–181. doi: 10.1016/j.immuni.2011.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Parihar R, Trotta R, Roda JM, et al. Src homology 2-containing inositol 5'-phosphatase 1 negatively regulates IFN-gamma production by natural killer cells stimulated with antibody-coated tumor cells and interleukin-12. Cancer Res. 2005;65:9099–9107. doi: 10.1158/0008-5472.CAN-04-4424. [DOI] [PubMed] [Google Scholar]
- 36.Trotta R, Parihar R, Yu J, et al. Differential expression of SHIP1 in CD56bright and CD56dim NK cells provides a molecular basis for distinct functional responses to monokine costimulation. Blood. 2005;105:3011–3018. doi: 10.1182/blood-2004-10-4072. [DOI] [PubMed] [Google Scholar]
- 37.Pahl JHW, Koch J, Götz JJ, et al. CD16A activation of NK cells promotes NK cell proliferation and memory-like cytotoxicity against cancer cells. Cancer Immunol Res. 2018;6:517–527. doi: 10.1158/2326-6066.CIR-17-0550. [DOI] [PubMed] [Google Scholar]
- 38.Waldmann TA. The biology of interleukin-2 and interleukin-15: implications for cancer therapy and vaccine design. Nat Rev Immunol. 2006;6:595–601. doi: 10.1038/nri1901. [DOI] [PubMed] [Google Scholar]
- 39.Ring AM, Lin JX, Feng D, et al. Mechanistic and structural insight into the functional dichotomy between IL-2 and IL-15. Nat Immunol. 2012;13:1187–1195. doi: 10.1038/ni.2449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Thierfelder WE, van Deursen JM, Yamamoto K, et al. Requirement for Stat4 in interleukin-12-mediated responses of natural killer and T cells. Nature. 1996;382:171–174. doi: 10.1038/382171a0. [DOI] [PubMed] [Google Scholar]
- 41.Parihar R, Dierksheide J, Hu Y, Carson WE. IL-12 enhances the natural killer cell cytokine response to Ab-coated tumor cells. J Clin Invest. 2002;110:983–992. doi: 10.1172/JCI15950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Schlums H, Cichocki F, Tesi B, et al. Cytomegalovirus infection drives adaptive epigenetic diversification of NK cells with altered signaling and effector function. Immunity. 2015;42:443–456. doi: 10.1016/j.immuni.2015.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Donnelly RP, Loftus RM, Keating SE, et al. mTORC1-dependent metabolic reprogramming is a prerequisite for NK cell effector function. J Immunol. 2014;193:4477–4484. doi: 10.4049/jimmunol.1401558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Keating SE, Zaiatz-Bittencourt V, Loftus RM, et al. Metabolic reprogramming supports IFN-γ production by CD56bright NK cells. J Immunol. 2016;196:2552–2560. doi: 10.4049/jimmunol.1501783. [DOI] [PubMed] [Google Scholar]
- 45.Nandagopal N, Ali AK, Komal AK, Lee SH. The critical role of IL-15-PI3K-mTOR pathway in natural killer cell effector functions. Front Immunol. 2014;5:187. doi: 10.3389/fimmu.2014.00187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Pauls SD, Marshall AJ. Regulation of immune cell signaling by SHIP1: a phosphatase, scaffold protein, and potential therapeutic target. Eur J Immunol. 2017;47:932–945. doi: 10.1002/eji.201646795. [DOI] [PubMed] [Google Scholar]
- 47.Martinez-Usatorre A, Sempere LF, Carmona SJ, et al. MicroRNA-155 expression is enhanced by T-cell receptor stimulation strength and correlates with improved tumor control in melanoma. Cancer Immunol Res. 2019;7:1013–1024. doi: 10.1158/2326-6066.CIR-18-0504. [DOI] [PubMed] [Google Scholar]
- 48.Herman SE, Gordon AL, Wagner AJ, et al. Phosphatidylinositol 3-kinase-δ inhibitor CAL-101 shows promising preclinical activity in chronic lymphocytic leukemia by antagonizing intrinsic and extrinsic cellular survival signals. Blood. 2010;116:2078–2088. doi: 10.1182/blood-2010-02-271171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Guillerey C, Huntington ND, Smyth MJ. Targeting natural killer cells in cancer immunotherapy. Nat Immunol. 2016;17:1025–1036. doi: 10.1038/ni.3518. [DOI] [PubMed] [Google Scholar]
- 50.Cooley S, Parham P, Miller JS. Strategies to activate NK cells to prevent relapse and induce remission following hematopoietic stem cell transplantation. Blood. 2018;131:1053–1062. doi: 10.1182/blood-2017-08-752170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Capuano C, Pighi C, Battella S, Santoni A, Palmieri G, Galandrini R. Memory NK cell features exploitable in anticancer immunotherapy. J Immunol Res. 2019;2019:8795673. doi: 10.1155/2019/8795673. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.