

Highlights
► Natural compounds inhibiting Porcine reproductive and respiratory syndrome virus replicative cycle. ► Ion transport is important for Porcine reproductive and respiratory syndrome virus replicative cycle. ► Sub micro molar concentration inhibitors of Porcine reproductive and respiratory syndrome virus.
Keywords: PRRSV, Natural product compounds, Viral inhibitor
Abstract
Porcine reproductive and respiratory syndrome virus (PRRSV) is an economically important pathogenic virus in the swine production. Current vaccines against PRRSV do not induce sterile immunity and the virus evolves at a rapid rate with frequent appearances of new strains. In this study, we screened a library of 502 highly purified natural product compounds to identify specific inhibitors of PRRSV replication cycle. Our observations showed that many of the inhibitory compounds identified have activity on the cellular ion transport mechanisms. We identified for the first time, four compounds which inhibit the PRRSV replication cycle at micro molar concentration or less, namely, 12-deoxyphorbol 13-phenylacetate 20-acetate, ouabain, bufalin and valinomycin. Further, we have identified 15 other compounds which can inhibit the PRRSV replication at the concentration of 8 μM. This study provides a basis for further development of pharmacological agents to inhibit PRRSV replication.
Porcine reproductive and respiratory syndrome virus (PRRSV) is an important pathogen of Porcine species which causes an estimated annual loss of more than 600 million dollars to the swine production industry in the United States alone (http://nationalhogfarmer.com August 17, 2011). The emergence of PRRSV as a pathogen was first observed in the 1980s and is currently a global epizootic (Murtaugh and Genzow, 2011). The virus belongs to the order Nidovirales, family Arteriviridae and is characterized by positive strand RNA genome of approximately 15 kb (kilobases) and is enveloped. The PRRSV viral genomic RNA is capped and polyadenylated. The virus has been classified into two major strains, the Type I (European) and Type II (North American) (Shi et al., 2010). The structural proteins of the virus are encoded from the distal third of the viral genome and are transcribed as sub-genomic mRNAs. The non-structural proteins take up the proximal two thirds of the viral genome and are synthesized as a shorter and a longer polyprotein, which are then processed into smaller proteins consisting of a variety of enzymes, such as proteases, RNA polymerases and RNA helicases. The non-structural proteins are shown to suppress the induction of host type I interferon genes and other cytokines (Beura et al., 2010, Patel et al., 2010). The PRRSV is estimated to have a high rate of evolution of 3.29 × 10−3 substitutions per nucleotide site per year and new strains frequently appear in the field (Song et al., 2010).
The currently available inactivated and modified live vaccines are inadequate to eradicate the virus and do not provide complete immunity from infection (Murtaugh and Genzow, 2011). Modified live vaccine PRRSV strains are also characterized by the shedding of the virus from vaccinated animals, which often revert to virulent strains (Murtaugh and Genzow, 2011, Storgaard et al., 1999). Since the global PRRSV epizootic in pigs is not completely amenable to immunological intervention, a pharmacological intervention might be a valuable alternative or additional control measure. For PRRSV, past studies in cell culture models have revealed only few compounds displaying antiviral activity (see Table 1 ). In general, natural compounds constitute a major source of effective pharmacological agents including, artemisinin, anti tumor alkaloids, salicylates, opiates, etc. (Mishra and Tiwari, 2011). Therefore, we evaluated a commercially available library (Screen-Well™ Natural Product Library, Enzo life sciences) of 502 purified natural compounds for their activity against PRRSV infection and replication. To this end, we established a high-throughput screening method.
Table 1.
A list of compounds in the published literature known to inhibit PRRSV replication and IC50 values for those estimated. NP – not published.
| Compound | IC50 | Reference |
|---|---|---|
| Macrolide antibiotic | ||
| Bafilomycin | ∼0.2 μM | Kreutz and Ackermann (1996) |
| Cellular ion transport targeting agents | ||
| Verapamil | NP | Kreutz and Ackermann (1996) |
| Amantadine | <0.2 μM | |
| Pyrithione | NP | te Velthuis et al. (2010) |
| Endosomal acidification blockers | ||
| Ammonium chloride | <0.5 mM | |
| Chloroquinone | >6 μM | |
| Monensin | NP | |
| Plant derived lectins | ||
| Indigowoad root polysaccharide | NP | van der Meer et al. (2007) |
| Hippeastrum hybrid agglutinin | >2 μM | |
| Galanthus nivalis agglutinin | >2 μM | |
| Cymbidium sp. agglutinin | NP | |
| Urtica dioica agglutinin | 4.8 ± 3.2 μM | |
| Non-plant derived lectins | ||
| Pradimicin-A | >120 μM | van der Meer et al. (2007) |
| Cyanovirin-N | NP | |
| Chelator | ||
| N-acetylpenicillamine | NP | Jiang et al. (2010) |
An immunofluorescence assay (IFA) based high-throughput screening platform in a 384 well format to detect viral replication was established utilizing MARC-145 cells permissive to PRRSV infection. The viral proteins were detected using a highly specific monoclonal antibody against the nucleocapsid protein (ORF7) of the PRRSV (Clone 5H2; in-house produced). After empirical optimization, the assay was performed by seeding 1000 cells per well, infected at a multiplicity of infection (MOI) of 10 at the time of seeding. The antiviral effect was evaluated at 48 h post-infection by fixing the cells with 4% paraformaldehyde. We included mock-infected cells and PRRSV-infected cells with additional 1 μM of mycophenolic acid (MPA), a de novo purine synthesis inhibitor (Sievers et al., 1997), as negative and positive controls, respectively, for virus inhibition of replication. For the immunofluorescence assay, the primary mouse monoclonal antibody from hybridoma supernatant was used undiluted and the secondary donkey anti-mouse immunoglobulins antibody labeled with Alexa Fluor 546 (stock 2 μg/μL) was used at a dilution of 1:500 in PBS–Tween (0.1%). The primary and secondary antibodies were stocked in bulk quantities and aliquots of these stocks were used consistently throughout the studies. Following the antibody staining, the cell nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI). The assay results were machine read with Cellomics® scanner, measuring the intensities of DAPI (indicating the nuclei of cells) and Alexa Fluor 543 (indicating the virus infection of cells). The percentage of infected cells was determined from this assay. The assay showed a Z-score of 0.53, when the PRRSV infected and infected cells with 1 μM MPA were compared, which validates the assay as highly reliable. Utilizing the assay, the compounds in the natural product library were screened for their ability to inhibit viral entry or/and replication by adding 10 μM the compound in DMSO (final concentration of 0.1%) to the cell plus virus mixture at the time of seeding. Mock-infected cells, infected cells, infected cells with 1 μM MPA and infected cells with 0.1% DMSO were used as controls. The compounds which caused a decrease of the DAPI intensity of treated cells to less than 70% of that of infected cells with only 0.1% DMSO were considered toxic to the cells and excluded from further analysis. The screening was repeated twice and we identified 19 compounds which inhibited the virus replication to less than 50% at a 10 μM concentration, as observed by IFA signal intensity, without adversely affecting the host cells (Table 2 ).
Table 2.
The list of the 19 compounds found to inhibit the PRRSV replication at 10 μM concentration to less than 50% as observed by IFA signal intensity without decreasing of the DAPI intensity of treated cells to less than 70%. Con – control inhibitor MPA.
| No. | Compound | Percent viral signal compared to 0.1% DMSO treated infected cells | Percent DAPI signal (nuclei) compared to 0.1% DMSO treated infected cells | ||||
|---|---|---|---|---|---|---|---|
| Concentration (μM) |
Concentration (μM) |
||||||
| 10 | 1 | 0.1 | 10 | 1 | 0.1 | ||
| 1 | Antimycin A1 | 23 | – | – | 81 | – | – |
| 2 | Bufalinb | 0 | 2 | 17 | 78 | 94 | 96 |
| 3 | Catechin hydrate (+)- | 18 | 92 | 96 | 91 | 95 | 96 |
| 4 | Cepharanthineb | 21 | 86 | 88 | 89 | 95 | 95 |
| 5 | Cinobufagina | 34 | – | – | 86 | – | – |
| 6 | Coumermycin A1 | 19 | 74 | 81 | 82 | 94 | 95 |
| 7 | Cryptotanshinone | 16 | 85 | 88 | 86 | 96 | 96 |
| 8 | Cycloheximide | 7 | – | – | 72 | – | – |
| 9 | Cyclopiazonic acidb | 14 | 78 | 86 | 83 | 94 | 96 |
| 10 | 12-Deoxyphorbol 13-phenylacetate 20-acetate | 0 | 4 | 36 | 89 | 94 | 95 |
| 11 | Dicoumarol | 36 | – | – | 89 | – | – |
| 12 | Digitoxina | 12 | – | – | 84 | – | – |
| 13 | Gitoxigenina | 11 | – | – | 85 | – | – |
| 14 | Harmaline | 41 | – | – | 86 | – | – |
| 15 | Hydrastine, d-β- | 24 | 94 | 98 | 79 | 97 | 97 |
| 16 | Hypocrellin B | 31 | – | – | 73 | – | – |
| 17 | Ouabain (−)-a | 0 | 0 | 2 | 81 | 93 | 96 |
| 18 | Strophanthidin acetatea | 27 | – | – | 77 | – | – |
| 19 | Valinomycin | 0 | 0 | 1 | 83 | 95 | 96 |
| Con | Mycophenolic acid | 0 | 12 | 76 | 38 | 96 | 96 |
Cardiac glycosides.
Compounds known to act on ion channels.
The compound library used for screening comprises of several classes of natural compounds; glycosides, terpenoids, coumarins, isoflavones, peptolides, alkaloids, flavones, macrolides, etc. The compounds belonging to the glycosides (known for their action on cardiac system) and cardio-active steroids were found in higher frequency to inhibit the viral replication in the assay. Of the identified 19 inhibitors, five were the glycosides and two were cardio-active steroids. These compounds share a similar mode of action on the ion channels in cells. Whether these compounds act on the cellular mechanisms and indirectly affect the virus or act directly on a viral component or mechanism is to be determined. The most likely viral target for these compounds could be the viral porin on the PRRSV envelope formed by the multimerization of the small envelope protein (ORF2b) (Lee and Yoo, 2006). Amantadine, known to block the M2 ion channel of the Influenza A, also blocks the PRRSV ORF2b porin (Lee and Yoo, 2006). Other compounds that inhibited the PRRSV included fungal metabolites and flavones. Subsequently, we selected ten compounds (Table 3 ), considering presence of known related pharmacological compounds and their biological effects. Compounds such as Cycloheximide, Dicoumarol and Harmaline, were not selected from the panel of 19 due to known undesirable biological activity profile. The toxicity effect of the selected ten compounds on MARC-145 cells was analyzed using alamar blue test (Invitrogen) by following the manufacturer’s instructions. The alamar blue fluorescence of the infected cells treated with the compounds was not less than 70% of the untreated infected cells and correlated with the DAPI fluorescence intensities estimated previously (data not shown). The inhibitory effect of these compounds were evaluated using the signal intensity in IFA assay, as described earlier, at 0.1, 1 and 8 μM concentrations of each compound (Table 2), by adding the compounds to the virus and cells at the time of seeding. The treatments were performed in triplicates and the experiment was performed twice.
Table 3.
The list of 10 compounds selected for secondary assays, to estimate their efficacy to inhibit PRRSV at doses of 0.1, 1 and 8 μM concentration.
| No. | Compound | Known pharmacological action |
|---|---|---|
| 1 | Bufalin | Cardiac glycoside with antiviral activity (Cui et al., 2010). |
| 2 | Catechin hydrate, (+)- | Flavonoid, anti-oxidant, cyclo-oxygenase inhibitor, anti inflammatory (Kuzuhara et al., 2009). |
| 3 | Cepharanthine | An alkaloid with antiviral activity against SARS CoV; anti-inflammatory, inhibits lipooxygenase (Baba et al., 2001, Zhang et al., 2005). |
| 4 | Coumermycin A1 | Novobiocin related antibiotic with antiviral activity (Palù et al., 1986). |
| 5 | Cryptotanshinone | A herb derived tanshinone that blocks Signal transducer and activator of transcription 3 (STAT3) dimerization and inhibits cytokine production by immune cells (Kang et al., 2000, Shin et al., 2009). |
| 6 | Cyclopiazonic acid | A fungal metabolite that acts on endoplasmic reticulum resident Ca2+ ATPase and mediates activation of NFkB, a key component of immune cell signaling (Xia et al., 2006). |
| 7 | 12-Deoxyphorbol 13-phenylacetate 20-acetate | A phorbol ester that activates PKCβ, and mimics IFN mediated signaling (Saraiva et al., 2004). |
| 8 | Hydrastine, d-β- | An alkaloid anti-inflammatory agent; inhibits cytochrome P450 mediated oxidation (Raner et al., 2007). |
| 9 | Ouabain (−)- | Cardiac glycoside with antiviral activity (Su et al., 2008). |
| 10 | Valinomycin | A dodeca-depsipeptide antibiotic, anti inflammatory, lipo-oxygenase inhibitor. Known to be a potent antiviral agent against Severe Acute Respiratory Syndrome Corona virus (SARS CoV) (De Clercq, 2006, Wu et al., 2004). |
All the ten compounds inhibited the virus replication at 8 μM concentration. Only four of the compounds were inhibitory at 1 μM or sub-micro molar concentrations; 12-deoxyphorbol 13-phenylacetate 20-acetate (dPPA), ouabain, valinomycin and bufalin. The representative images of infection are shown in Fig. 1 . Interestingly, the dPPA is known to activate the PKCβ and stimulate interferon signaling. Ouabain and bufalin are known to act on the cardiac system by affecting the ion channels. Further, bufalin perturbs signaling mediated by STAT3. Valinomycin, a dodecadepsipeptide antibiotic, is a potassium ionophore. Previous studies have shown that valinomycin has a potent antiviral activity against severe acute respiratory syndrome corona virus (SARS CoV), another important Nidovirus (Wu et al., 2004). We estimated the 50% inhibitory concentrations (IC50) for the inhibition of viral replication of the four compounds as described earlier (Wu et al., 2004). The IC50 values derived by curve fitting of mean of percent infected cells in triplicates observed by IFA at 8, 1, 0.1, 0.01, 0.001 and 0.0001 μM concentrations of the inhibitors, from three independent experiments. The IC50 were well below 1 μM (Table 4 ) for all the four compounds.
Fig. 1.


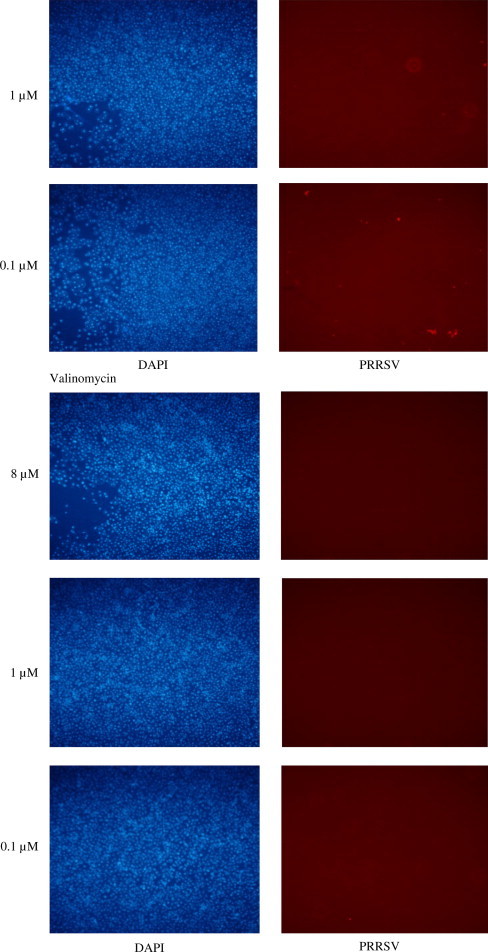
12-Deoxyphorbol 13 phenylacetate 20 acetate, ouabain, valinomycin and bufalin exhibited inhibition of the PRRSV replication at micro-molar or sub-micro-molar concentrations. Viral replication in the presence of 8, 1, 0.1 μM of the above compounds was examined by IFA. Nuclei – DAPI staining (blue); PRRSV protein ORF7 staining (red). MPA – mycophenolic acid as a positive control inhibitor. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Table 4.
The estimated IC50 of the four compounds with the highest inhibitory activity.
| Compound | IC50 (±standard deviation) |
|---|---|
| Bufalin | 52 nM (±3.8) |
| 12-Deoxyphorbol 13-phenylacetate 20-acetate | 118 nM (±8.4) |
| Ouabain (−)- | 41 nM (±3.2) |
| Valinomycin | 24 nM (+4.6) |
We next evaluated if these four compounds acted on the binding and entry steps of the virus or during the replication process. MARC-145 cells were incubated with each of the compounds (1 μM) for 1 h prior to the addition of the virus at an MOI of 10 and subsequently the cells were incubated at 37 °C for 2 h. After which, the cells were washed thrice with phosphate buffered saline (PBS) and the cells were incubated in fresh media and fixed after 48 h. The results showed that identified four compounds do not block the binding and entry of the virus, but act during the subsequent replication process. Next, the cells were infected with the virus at an MOI of 10 and incubated with compounds (1 μM) from 0, 2 or 8 h post infection (hpi) and fixed at 48 hpi. The IFA signal quantification results (Table 5 ) showed that virus replication was almost completely inhibited by all the four compounds when added 2 hpi. However, when added at 8 hpi, dPPA decreased the viral signal only by 30% while the other three compounds almost completely inhibited the signal. The results show that dPPA inhibits the virus replication by acting on the early event in virus–cell interaction. The dPPA is known to mimic the cellular interferon signaling (Saraiva et al., 2004), and possibly acts by augmenting the interferon mediated virus suppression. The other three compounds are known act on ion channels and could have a different mode of action. These compounds which are inhibitory even when added in the later stages of infection could have a better value in the in vivo inhibition of the virus. Further detailed analysis needs to be performed to identify their exact mode of action. Next the inhibitory activities of the compounds were tested in a rapid cytopathic effect (CPE) inhibition assay adapted from Cotarelo et al., 1999. The MARC145 cells were infected with PRRS virus at an MOI of 10 with or without the above four compounds (at 8, 1 and 0.1 μM concentration) and examined for CPE at 72 h post infection. The untreated infected cells showed extensive CPE at 72 hpi. At 8 and 1 μM concentrations, all of the four compounds completely inhibited observable CPE at 72 hpi. However, at 0.1 μM concentration dPPA showed mild CPE. The CPE inhibition assay showed that compounds were effective in inhibiting the virus induced cellular pathology.
Table 5.
Effect of time of addition after infection on the inhibition of virus replication of the compounds at 1 μM. hpi – hours post infection.
| Compound | Percent viral signal compared to 0.1% DMSO treated infected cells when compounds were added at indicated hpi |
||
|---|---|---|---|
| 0 | 2 | 8 | |
| Bufalin | 0 | 3 | 7 |
| 12-Deoxyphorbol 13-phenylacetate 20-acetate | 0 | 2 | 68 |
| Ouabain (−)- | 0 | 0 | 4 |
| Valinomycin | 0 | 1 | 5 |
| Mycophenolic acid | 0 | 3 | 16 |
Apart from these four compounds, the other compounds from the original 19 identified to inhibit PRRSV at 10 μM concentration or the analogs of these compounds could have a better profile of activity in other cell culture models for PRRSV replication or in in vivo studies. However, as PRRSV infection could not be successfully modeled in a laboratory animal, the in vivo studies will have to be performed in pigs or piglets. Considering the rapid evolution rate of PRRSV (Murtaugh and Genzow, 2011, Song et al., 2010), the eradication of the virus cannot be achieved only by immunological intervention, which is the trend observed in the field so far. Therefore, a therapeutic intervention with pharmacological agents inhibiting virus replication could represent a valuable alternative or additional tool against PRRSV. For the latter, a combined immunological and pharmacological intervention could decrease the risk of the emergence of resistant strains as well. In conclusion, we here identified several natural compounds displaying potent antiviral activity, providing a basis for further research for pharmacological agents against PRRSV.
Contributor Information
Justin Jang-Hann Chu, Email: miccjh@nus.edu.sg.
Jimmy Kwang, Email: kwang@tll.org.sg.
References
- Baba M., Okamoto M., Kashiwaba N., Ono M. Anti-HIV-1 activity and structure–activity relationship of cepharanoline derivatives in chronically infected cells. Antivir. Chem. Chemother. 2001;12:307–312. doi: 10.1177/095632020101200506. [DOI] [PubMed] [Google Scholar]
- Beura L.K., Sarkar S.N., Kwon B., Subramaniam S., Jones C., Pattnaik A.K., Osorio F.A. Porcine reproductive and respiratory syndrome virus nonstructural protein 1beta modulates host innate immune response by antagonizing IRF3 activation. J. Virol. 2010;84:1574–1584. doi: 10.1128/JVI.01326-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cotarelo M., Catalán P., Sánchez-Carrillo C., Menasalvas A., Cercenado E., Tenorio A., Bouza E. Cytopathic effect inhibition assay for determining the in-vitro susceptibility of herpes simplex virus to antiviral agents. J. Antimicrob. Chemother. 1999;44:705–708. doi: 10.1093/jac/44.5.705. [DOI] [PubMed] [Google Scholar]
- Cui X., Inagaki Y., Xu H., Wang D., Qi F., Kokudo N., Fang D., Tang W. Anti-hepatitis B virus activities of cinobufacini and its active components bufalin and cinobufagin in HepG2.2.15 cells. Biol. Pharm. Bull. 2010;33:1728–1732. doi: 10.1248/bpb.33.1728. [DOI] [PubMed] [Google Scholar]
- De Clercq E. Potential antivirals and antiviral strategies against SARS coronavirus infections. Expert. Rev. Anti. Infect. Ther. 2006;4:291–302. doi: 10.1586/14787210.4.2.291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang Y., Fang L., Luo R., Xiao S., Chen H. N-Acetylpenicillamine inhibits the replication of porcine reproductive and respiratory syndrome virus in vitro. Vet. Res. Commun. 2010;34:607–617. doi: 10.1007/s11259-010-9435-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang B.Y., Chung S.W., Kim S.H., Ryu S.Y., Kim T.S. Inhibition of interleukin-12 and interferon-gamma production in immune cells by tanshinones from Salvia miltiorrhiza. Immunopharmacology. 2000;49:355–361. doi: 10.1016/s0162-3109(00)00256-3. [DOI] [PubMed] [Google Scholar]
- Kreutz L.C., Ackermann M.R. Porcine reproductive and respiratory syndrome virus enters cells through a low pH-dependent endocytic pathway. Virus Res. 1996;42:137–147. doi: 10.1016/0168-1702(96)01313-5. [DOI] [PubMed] [Google Scholar]
- Kuzuhara T., Iwai Y., Takahashi H., Hatakeyama D., Echigo N. Green tea catechins inhibit the endonuclease activity of influenza A virus RNA polymerase. PLoS Curr. 2009;1:RRN1052. doi: 10.1371/currents.RRN1052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee C., Yoo D. The small envelope protein of porcine reproductive and respiratory syndrome virus possesses ion channel protein-like properties. Virology. 2006;10:30–43. doi: 10.1016/j.virol.2006.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murtaugh M.P., Genzow M. Immunological solutions for treatment and prevention of porcine reproductive and respiratory syndrome (PRRS) Vaccine. 2011;29:8192–8204. doi: 10.1016/j.vaccine.2011.09.013. [DOI] [PubMed] [Google Scholar]
- Mishra B.B., Tiwari V.K. Natural products: an evolving role in future drug discovery. Eur. J. Med. Chem. 2011;46:4769–4807. doi: 10.1016/j.ejmech.2011.07.057. [DOI] [PubMed] [Google Scholar]
- Palù G., Meloni G.A., von Berger J., Masotti L. On the complex nature of the antiviral activity of coumermycin A1: its interference with the replication of herpes simplex virus type 1. Antiviral Res. 1986;6:19–32. doi: 10.1016/0166-3542(86)90036-7. [DOI] [PubMed] [Google Scholar]
- Patel D., Nan Y., Shen M., Ritthipichai K., Zhu X., Zhang Y.J. Porcine reproductive and respiratory syndrome virus inhibits type I interferon signaling by blocking STAT1/STAT2 nuclear translocation. J Virol. 2010;84:11045–11055. doi: 10.1128/JVI.00655-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raner G.M., Cornelious S., Moulick K., Wang Y., Mortenson A., Cech N.B. Effects of herbal products and their constituents on human cytochrome P450(2E1) activity. Food Chem. Toxicol. 2007;45:2359–2365. doi: 10.1016/j.fct.2007.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saraiva L., Fresco P., Pinto E., Gonçalves J. Characterization of phorbol esters activity on individual mammalian protein kinase C isoforms, using the yeast phenotypic assay. Eur. J. Pharmacol. 2004;491:101–110. doi: 10.1016/j.ejphar.2004.03.035. [DOI] [PubMed] [Google Scholar]
- Shi M., Lam T.T., Hon C.C., Hui R.K., Faaberg K.S., Wennblom T., Murtaugh M.P., Stadejek T., Leung F.C. Molecular epidemiology of PRRSV: a phylogenetic perspective. Virus Res. 2010;154:7–17. doi: 10.1016/j.virusres.2010.08.014. [DOI] [PubMed] [Google Scholar]
- Shin D.S., Kim H.N., Shin K.D., Yoon Y.J., Kim S.J., Han D.C., Kwon B.M. Cryptotanshinone inhibits constitutive signal transducer and activator of transcription 3 function through blocking the dimerization in DU145 prostate cancer cells. Cancer Res. 2009;69:193–202. doi: 10.1158/0008-5472.CAN-08-2575. [DOI] [PubMed] [Google Scholar]
- Sievers T.M., Rossi S.J., Ghobrial R.M., Arriola E., Nishimura P., Kawano M., Holt C.D. Mycophenolate mofetil. Pharmacotherapy. 1997;17:1178–1197. [PubMed] [Google Scholar]
- Song J., Shen D., Cui J., Zhao B. Accelerated evolution of PRRSV during recent outbreaks in China. Virus Genes. 2010;41:241–245. doi: 10.1007/s11262-010-0507-2. [DOI] [PubMed] [Google Scholar]
- Storgaard T., Oleksiewicz M., Bøtner A A. Examination of the selective pressures on a live PRRS vaccine virus. Arch. Virol. 1999;144:2389–2401. doi: 10.1007/s007050050652. [DOI] [PubMed] [Google Scholar]
- Su C.T., Hsu J.T., Hsieh H.P., Lin P.H., Chen T.C., Kao C.L., Lee C.N., Chang S.Y. Anti-HSV activity of digitoxin and its possible mechanisms. Antiviral Res. 2008;79:62–70. doi: 10.1016/j.antiviral.2008.01.156. [DOI] [PubMed] [Google Scholar]
- te Velthuis A.J., van den Worm S.H., Sims A.C., Baric R.S., Snijder E.J., van Hemert M.J. Zn(2+) inhibits coronavirus and arterivirus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathog. 2010;6:e1001176. doi: 10.1371/journal.ppat.1001176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Meer F.J., de Haan C.A., Schuurman N.M., Haijema B.J., Peumans W.J., Van Damme E.J., Delputte P.L., Balzarini J., Egberink H.F. Antiviral activity of carbohydrate-binding agents against Nidovirales in cell culture. Antiviral Res. 2007;76:21–29. doi: 10.1016/j.antiviral.2007.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu C.Y., Jan J.T., Ma S.H., Kuo C.J., Juan H.F., Cheng Y.S., Hsu H.H., Huang H.C., Wu D., Brik A., Liang F.S., Liu R.S., Fang J.M., Chen S.T., Liang P.H., Wong C.H. Small molecules targeting severe acute respiratory syndrome human coronavirus. Proc. Natl. Acad. Sci. USA. 2004;6:10012–10017. doi: 10.1073/pnas.0403596101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xia W., Shen Y., Xie H., Zheng S. Involvement of endoplasmic reticulum in hepatitis B virus replication. Virus Res. 2006;121:116–121. doi: 10.1016/j.virusres.2006.01.020. [DOI] [PubMed] [Google Scholar]
- Zhang C.H., Wang Y.F., Liu X.J., Lu J.H., Qian C.W., Wan Z.Y., Yan X.G., Zheng H.Y., Zhang M.Y., Xiong S., Li J.X., Qi S.Y. Antiviral activity of cepharanthine against severe acute respiratory syndrome coronavirus in vitro. Chin. Med. J. (Engl.) 2005;20:493–496. [PubMed] [Google Scholar]