

Abstract
In this study, we tried to identify dengue virus-specific CD4+ T-cell epitopes, which can induce PBMC (peripheral blood mononuclear cells) isolated from DF convalescent patients (dengue virus type 1 infection) to secrete IFN-γ. PBMC of DF convalescent patients were stimulated in vitro with dengue virus-derived peptides, which were prepared based on the prediction of dengue virus-specific CD4+ T-cell epitopes by using RANKpep online software. Subsequently, the frequency of IFN-γ producing T cells and percentage of IFN-γ+ CD4+ T cells were measured by using ELISPOT assay and ICS assay (intracellular cytokine straining), respectively. The positive response of PBMC by ELISPOT showed that the numbers of SFC (spots forming cells) ranged from 50 to 310 SFC/1 × 106 PBMC. The positive response of PBMC by ICS assay showed that the percentage of IFN-γ+ CD4+ T cells ranged from 0.03 to 0.27%. As a result, C45-57 (KLVMAFIAFLRFL), E396-408 (SSIGKMFEATARG), NS323-35 (YRILQRGLLGRSQ), and NS3141-155 (NREGKIVGLYGNGVV) were identified as dengue virus-specific CD4+ T-cell epitopes.
Keywords: CD4+ T-cell epitope, ELISPOT, ICS
1. Introduction
Dengue virus (DEN) is a single-stranded positive RNA virus of the family Flaviviridae comprised of four distinct serotypes (DEN-1, 2, 3, 4) within the flavivirus genus. Infection with any of the four serotypes of dengue virus can result in an asymptomatic infection, a self-limited febrile disease (dengue fever, DF) or a severe, life-threatening illness (dengue hemorrhagic fever, DHF) (Halstead, 1988). DEN infections are a serious cause of morbidity and mortality in most tropical and subtropical areas of the world (Gubler, 1998, Gibbons and Vaughn, 2002). If untreated, the mortality rate of DHF is around 30% but can be reduced to 0.2–5% by fluid replacement.
Even though previous study suggested that antibody-dependent enhancement (ADE) take part in the pathogenesis of DHF, the pathogenesis of DHF cannot entirely be explained by antibody, however. Currently, many studies suggested that dengue-specific T-cell immune response are hypothesized to play an important role in the immunopathogenesis of DHF during a secondary DEN virus infection, and induction of immunopathology by T lymphocytes may occur by various mechanisms, including cell-mediated cytotoxicity and/or cytokine production (Miskovsky et al., 1994, Vergelli et al., 1997). In addition, other study showed that DHF is associated with high levels of T-cell cytokines such as IFN-γ and TNF-α, and evidence of marked T-cell activation (Libraty et al., 2002). Recently, immunopathogenesis in DHF has been proposed. The evidence is that both dengue-specific CD8+ and CD4+ lymphocytes have been implicated in pathogenesis and natural immunity to dengue viruses (Rothman and Ennis, 1999, Lei et al., 2001). How T cells contribute to this process is incompletely defined, however, since only a few studies have directly examined T-cell responses during acute disease and few DEN-specific T-cell epitopes have been identified (Green et al., 1999, Mongkolsapaya et al., 2003). Therefore, the establishment of a library of defined T-cell epitopes from dengue viruses will help to explore the role of T cells in immunity and disease pathogenesis.
In the present study, a combination of bioinformatics tools (epitope-prediction programs RANKpep) and in vitro assays (enzyme-linked immunospot assay (ELISPOT) and intracellular cytokine staining assay (ICS)) was used to screen and select antigen sequences as potential DEN-specific CD4+ T-cell epitopes, then the selected sequences are tested for biological function by their activation of T cells of DEN infected populations.
2. Materials and methods
2.1. Study populations
A total of 10 DF convalescent patients were enrolled in this study. They are early-convalescent-phase DEN infection cases (DEN-1) recruited from the eighth people's hospital of Guangzhou, China. We also included three flavivirus-naïve Guangdong residues. DEN-1 infection was confirmed by virus isolation in C6/36 cell culture and subsequent serotype identification with DEN serotype-specific RT-PCR during the viremia-phase (Lanciotti et al., 1992). In addition, DEN infection was also confirmed by using commercial capture-immunoglobulin M (IgM) and IgG enzyme-linked immunosorbent assay (ELISA) during the convalescent-phase as described previously (Kuno et al., 1985). After obtaining informed consent, peripheral blood samples were drawn for preparation of peripheral blood mononuclear cells (PBMC). Meanwhile, the ethics committees of the authors’ institutions approved this study.
2.2. PBMC separation
PBMC were isolated from the peripheral whole blood by Ficoll–Hypaque density gradient centrifugation (400 × g, 30 min, 24 °C) and used directly in ELISPOT assay and ICS assay.
2.3. Computational prediction of candidate CD4+ T-cell epitopes for dengue virus and synthesis of peptides
RANKpep online software (http://www.mif.dfci.harvard.edu/Tools/rankpep.html) was used for the prediction of DEN-specific CD4+ T-cell epitopes. The DEN strains used in the prediction process included DEN-1 strains, DEN-2 strains, DEN-3 strains and DEN-4 strains (Table 1 ).
Table 1.
The DEN strains used for the prediction of DEN-specific T-cell epitopes
| Serotypes | Strains | GenBank accession number |
|---|---|---|
| DEN-1 | A88 | AB074761 |
| 45AZ5 | NC_001477 | |
| GZ/80 | AF350498 | |
| Singapore 8114/93 | AY762084 | |
| 71/GZ02 | EF025110 | |
| Mochizuki | AB074760 | |
| DEN-2 | New Guinea-C strain | M29095 |
| ZS01/01 | EF051521 | |
| ThD2_0038_74 | DQ181806 | |
| ThD2_0498_84 | DQ181804 | |
| ThD2_0078_01 | DQ181797 | |
| Taiwan-1008DHF | AY776328 | |
| DR59/01 | AB122022 | |
| Jamaica/N.1409 | M20558 | |
| FJ11/99 | AF359579 | |
| DEN-3 | H87 | NC_001475 |
| 99TW628 | DQ675533 | |
| Sleman/78 | AY648961 | |
| Singapore 8120/95 | AY766104 | |
| BR74886/02 | AY679147 | |
| Feb-80 | AF317645 | |
| DEN-4 | H241 | AY947539 |
| Jun-30 | AF326573 | |
| Singapore 8976/95 | AY762085 | |
| ThD4_0087_77 | AY618991 | |
| Taiwan-2K0713 | AY776330 | |
The following criteria were used to select the candidate DEN-specific CD4+ T-cell epitopes: 1, having a higher ranking scores on RANKpep prediction; 2, predicted epitopes should be highly conserved in these selected DEN strains. Four higher-ranking candidate CD4+ T-cell epitopes (C45-57, E396-408, NS323-35 and NS3141-155) were synthesized using 9-fluorenylmethoxy carbonyl-based solid-phase chemistry on a multiple peptide synthesizer (purity >95% pure; HuaChen Company, Xi’an, China) and the coincidence of the actual molecular mass with the theoretical molecular mass was confirmed by mass spectrography analysis. Peptides were dissolved in phosphate-buffered saline (PBS) and diluted to 2 mg/ml in PBS, and stored at −80 °C for subsequent ELISPOT assay and ICS assay. The sequences and other information of these peptides are detailed in Table 2 .
Table 2.
Candidate DEN-specific CD4+ T-cell epitopes predicted by RANKpep
| Peptide no. | Proteins | Position | Sequences | Restricting HLA-II allelesa | Serotypesb |
|---|---|---|---|---|---|
| C45-57 | C | 45–57 | KLVMAFIAFLRFL | DP4, DPw4, DQ1, DQ5, DQ7 | Den-1, Den-3 |
| E396-408 | E | 396–408 | SSIGKMFEATARG | DP9, DQ8, DQ1, DQ7 | Den-1, Den-3 |
| NS323-35 | NS3 | 23–35 | YRILQRGLLGRSQ | DR1, DQ1, DQ6 | Den-1 |
| NS3141-155 | NS3 | 141–155 | NREGKIVGLYGNGVV | DR15 | Den-1 |
Predicted by using RANKpep online software.
Those sequences of peptides are conserved among those serotypes.
2.4. Interferon (IFN)-γ ELISPOT assay
Assay was performed using Human IFN-γ ELISPOT kit (R&D systems Minneapolis, MN), following the protocol provided by the manufacturer. Briefly, 96-well polyvinylidene difluoride-backed plates, precoated with anti-IFN-γ monoclonal antibody, were blocked with 200 μl of sterile RPMI-1640 medium (containing 2 mM l-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin and 10% heat-inactivated fetal calf serum) for 20 min at room temperature. A total of 1 × 105 PBMC were added in 100 μl RPMI-1640 medium per well, and individual peptides were subsequently added to a final concentration of 10 μg/ml. Unstimulated and phytohemagglutinin (PHA, 10 μg/ml) stimulated cells served as negative and positive controls, respectively. In addition, SARS-CoV peptide (Severe Acute Respiratory Syndrome-coronary virus, SHVPLATSRTLA) used as irrelevant peptide control. After an overnight incubation at 37 °C and 5% CO2 in air, plates were washed with PBS containing 0.05% Tween-20. Next, the biotinylated anti-IFN-γ antibody was added. After an overnight incubation at 4 °C, plates were washed, and then incubated for 2 h at room temperature with the diluted streptavidin-alkaline phosphatase. After four final washes, plates were developed with 100 μl/well of alkaline phosphatase substrate 5-bromo-4-chloro-3-indolylphosphate and nitroblue tetrazolium substrates (BCIP/NBT) Chromogen for 1 h at room temperature (protecting from light). Color development was monitored and stopped by washing with water. After dried overnight at room temperature, SFC (spots forming cells) per well were counted using an ELISPOT reader (Beijing SageCreation Science Co., Ltd., China). The deep-brown-colored spots with decreasing density radiating from the center were counted and expressed as responding cells in 1 × 106 PBMC.
The assay was considered valid only if positive control wells showed >100 spots/1 × 106 PBMC and if negative control wells showed <10 spots/1 × 106 PBMC. A positive response to a peptide was defined as having ≥50 spots/1 × 106 PBMC after subtraction of background as described previously (Simmons et al., 2005).
2.5. Intracellular cytokine staining and flow cytometry
PBMC were resuspended in RPMI-1640 medium and used directly in IFN-γ ICS assay as previously described (Mothe et al., 2002). Briefly, each test contained 0.7 × 106 PBMC and was incubated with either peptides (SARS-CoV peptide as irrelevant peptide control) at 10 μg/ml or not stimulated (no peptide control) for 6 h at 37 °C and 5% CO2. Meanwhile, Brefeldin A (BFA, 10 μg/ml, Biolegend Inc. CA) was added. Cells were washed using PBS and then stained using PE-labelled anti-CD3, APC-labelled anti-CD4 and PE-CY5-labelled anti-CD8 antibodies (Biolegend Inc. CA) for 40 min at 4 °C. Subsequently, cells were fixed in 4% paraformaldehyde, permeabilized with 0.1% saponin, and then stained using fluorescein isothiocyanate (FITC)-labelled anti-IFN-γ antibody (Biolegend Inc. CA). Data acquisition was performed on a FACS Calibur flow cytometer (BD bioscience) using CellQuest software. The number of events collected for each sample varied between 100,000 and 300,000, depending on the donors. CD4+/CD8+ cells were gated and analyzed for cytokine expression. Peptide-specific cells were defined as IFN-γ-producing CD4+/CD8+ T cells.
3. Results
3.1. Characteristics of study subjects
Between October 2006 and January 2007, we recruited 10 convalescent patients having dengue virus type 1 infection. The mean age was 37 years (ranging from 18 to 64 years) and the mean time (from discharge to drawing blood sample) was 58 days (ranging from 14 to 120 days). The characteristics of the study population are detailed in Table 3 .
Table 3.
Characteristics of the DF convalescent patients
| Donors | Age (years) | Sex (F/M) | Clinical diagnosis | Diagnostic methodsa | Timeb (days) |
|---|---|---|---|---|---|
| Donor 1 | 18 | M | DF | ELISA, RT-PCR | 20 |
| Donor 2 | 37 | M | DF | ELISA, RT-PCR | 14 |
| Donor 3 | 64 | F | DF | ELISA, RT-PCR | 18 |
| Donor 4 | 30 | F | DF | ELISA, RT-PCR | 20 |
| Donor 5 | 43 | F | DF | ELISA, RT-PCR | 60 |
| Donor 6 | 24 | M | DF | ELISA, RT-PCR | 120 |
| Donor 7 | 34 | F | DF | ELISA, RT-PCR | 90 |
| Donor 8 | 48 | F | DF | ELISA, RT-PCR | 60 |
| Donor 9 | 44 | M | DF | ELISA, RT-PCR | 120 |
| Donor 10 | 28 | M | DF | ELISA, RT-PCR | 60 |
RT-PCR was used for DEN infection during febrile phase while ELISA was used during convalescent phase.
Indicating the time from being discharged to drawing blood samples.
3.2. IFN-γ secreting cell frequencies in PBMC stimulated with peptides
The positive control assay showed that the numbers of SFC ranged from 600 to 1320 SFC/1 × 106 PBMC while the negative control (unstimulated PBMC) showed that the numbers of SFC was 0. In 10 DF convalescent patients, we identified one or more individual peptides that reproducibly induced an IFN-γ response in the ELISPOT assay that exceeded 50 SFU/1 × 106 PBMC. In total, T cells from 10 DF convalescent patients recognized four different peptides. The magnitude of responses to individual peptides ranged from 50 to 310 SFU/million PBMC. The breadth of the response in individual patients ranged from 1 to 4 peptides. In addition, the mean numbers of spots obtained for DEN-1-seronegative individuals were 0 for C45-57, NS323-35 and irrelevant peptide, and 10 for E396-408 and NS3141-155. In total, these four peptides (C45-57, E396-408, NS323-35 and NS3141-155) are DEN-specific T-cell epitopes. A summary of the individual peptides that reproducibly induced responses in IFN-γ ELISPOT assay is provided in Fig. 1 .
Fig. 1.
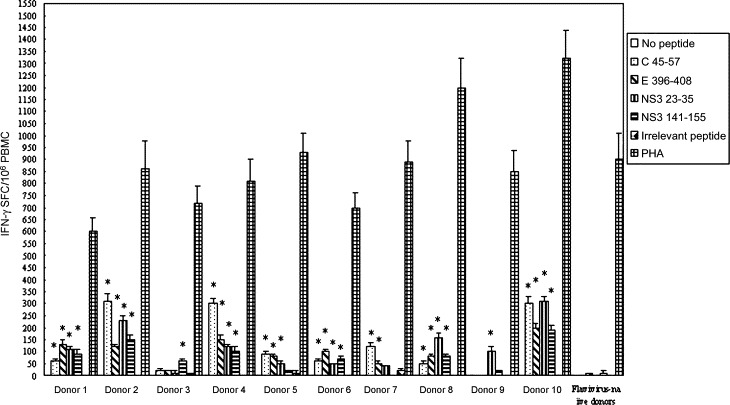
Enumeration of IFN-γ SFC in a 24 h ELISPOT assay. The numbers of SFC per 106 PBMC is shown as the mean ± S.D. of duplicate cells. *Indicating the positive response to a peptide.
3.3. Percentages of IFN-γ+ CD4+ T cells after peptides stimulation
The levels of circulating CD4+ T cells of 10 patients ranged from 10.89 to 21.46% while the levels of circulating CD8+ T cells ranged from 6.66 to 21.03%. The CD4/CD8 ratios were above one (varying from 1.01 to 1.89) (Table 4 ). For any subject, there were no IFN-γ+ CD4+ T cells in no peptide control and irrelevant peptide control. We identified individual epitopes that producibly evoked IFN-γ+ CD4+ T cells in the ICS assay that exceeded 0.01% (IFN-γ+ cells less than or equal to 0.01% of CD4+ T cells were considered as negative). The positive response of PBMC showed that the percentages of IFN-γ+ CD4+ T cells ranged from 0.03 to 0.27% (see Fig. 2 ) while the percentages of IFN-γ+ CD8+ T cells were 0 (data not shown). In addition, no responses to these peptides were seen in PBMC of the flavivirus-naïve donor (data not shown). As a result, C45-57, E396-408, NS323-35 and NS3141-155 are DEN-specific CD4+ T-cell epitopes (see Fig. 2).
Table 4.
The percentages of lymphocyte subsets of the DF convalescent patients
| Donors | CD3+CD4+T cells (%) | CD3+CD8+T cells (%) | CD4/CD8 |
|---|---|---|---|
| Donor 1 | 21.31 | 19.72 | 1.08 |
| Donor 2 | 13.65 | 21.03 | 1.13 |
| Donor 3 | 18.79 | 15.62 | 1.2 |
| Donor 4 | 13.07 | 10.4 | 1.27 |
| Donor 5 | 20.84 | 19.03 | 1.1 |
| Donor 6 | 10.89 | 10.75 | 1.01 |
| Donor 7 | 21.46 | 16.38 | 1.31 |
| Donor 8 | 19.82 | 14.25 | 1.39 |
| Donor 9 | 17.11 | 13.06 | 1.31 |
| Donor 10 | 12.57 | 6.66 | 1.89 |
Fig. 2.
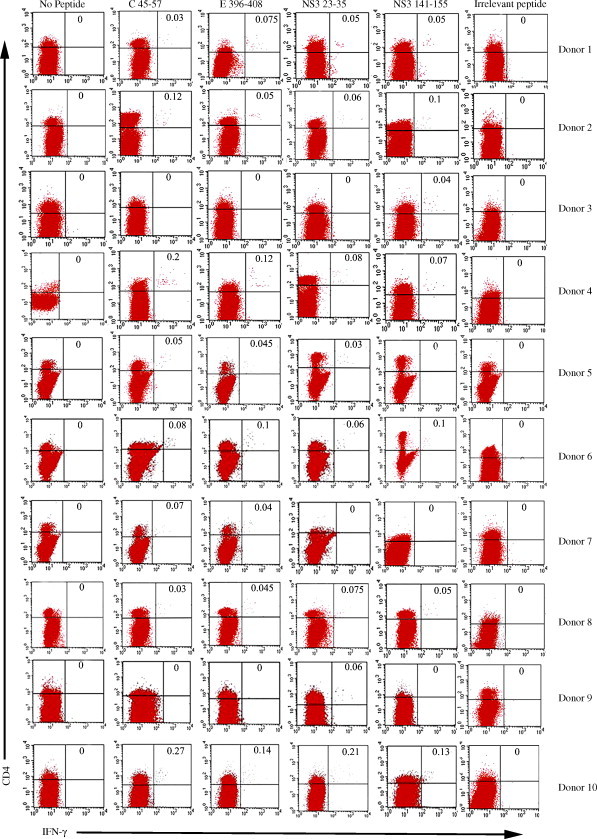
Frequencies of IFN-γ+ CD4+ T cells in PBMC of subjects. Numbers on the upper right hand corner of each panel indicate IFN-γ+ CD4+ T cells as percentage of CD4+ T cell. Data acquisition was performed on a FACS Calibur flow cytometer (BD bioscience) using CellQuest software. The number of events collected for each sample varied between 100,000 and 300,000, depending on the donors. CD4+/CD8+cells were gated and analyzed for cytokine expression.
4. Discussion
Initially, the viral T-cell epitopes were identified either by sequencing of eluted peptides bound to MHC molecules (Major Histocompability Complex) or by screening panels of overlapping peptides derived from the viral protein. However, these two methods have their shortcomings. The former is cumbersome and needs sophisticated equipments (such as mass spectrometer or peptide sequencer). The latter needs synthesis of a series of the overlapping peptides and massive experiments to identify which peptide is epitope, making it an extensive and time-consuming process (Drabner et al., 2002). At present, a novel strategies that utilize computer-based T-cell epitopes prediction methods combined with in vitro/in vivo studies was established to predict potential T-cell epitopes and are becoming increasingly popular as the amino acid sequences of pathogenic proteins are available in database (Huang et al., 2004, Turcanová and Höllsberg, 2004).
Epitope-prediction program RANKpep was developed by the “molecular immunology foundation” by P.A. Reche et al., which uses position specific scoring matrics (PSSM), cleavage model and predicts MHC binding peptide from an input protein based on their similarity to a set of peptides known to bind to a given MHC molecule (Reche et al., 2004). Among epitopes-prediction programs, RANKpep prediction algorithm has higher sensitivity and specificity and represents a quick and effective way to identify T-cell epitopes (Drabner et al., 2002, Gomez-Nunez et al., 2006, Reche et al., 2002, Reche et al., 2004). Recently, ELISPOT assay and ICS assay were extensively used to measure the frequencies of memory T cells. Compared with other in vitro methods, ELISPOT assay and ICS assay own the higher sensitivity and the latter have the advantage of defining the phenotype of the cytokine secreting cells (Waldrop et al., 1997, Butz and Bevan, 1998, Pala et al., 2000). Therefore, the combination of ELISPOT assay and ICS assay is necessary to identify T-cell epitopes.
Up to date, studies identified some DEN-specific CD4+ T-cell epitopes, which are mainly located in NS3 protein or C protein (Kurane et al., 1995, Kurane et al., 1998, Gagnon et al., 1996, Mangada et al., 2004). However, most of these epitopes are only conserved in DEN-4 or DEN-3 or DEN-2, 3, 4 and the epitope which is highly conserved in DEN-1 had not been identified. Furthermore, the low sensitive methods/technologies (the limiting dilution method and cytotoxic assay) limited general laboratories to identify T-cell epitopes. In this study, we measured a higher frequencies of DEN peptide-specific IFN-γ responding CD4+ T cells by using ELISPOT assay and ICS assay and then identified four DEN-specific CD4+ T-cell epitopes. More importantly, among the 10 DF convalescent patients, as for each peptide, the breadth of the positive response ranged from 7 to 8 subjects. This suggested that these epitopes could be the immunodominant epitopes and restricted by the high-frequency HLA alleles of human population. Of course, future experiments would be performed to determine the HLA restriction of these epitopes. Among these four epitopes, C45-57 and NS3141-155 contains the sequence that was previously characterized as DEN-specific CD4+ T-cell epitopes (C47-55 and NS3146-154, respectively) (Kurane et al., 1995, Gagnon et al., 1996). Therefore, our results indicated that C47-55 and NS3146-154 could be the minimal sequence recognized by MHC-II molecule. Interestingly, NS3146-154 was previously identified as HLA-DR15-restricted epitope (Kurane et al., 1995) while the putative HLA allele of NS3141-155 by RANKpep is HLA-DR15. In addition, C47-55 was HLA-DPw4-restricted epitope (Gagnon et al., 1996) while the restricting HLA alleles of C45-57 by RANKpep include DPw4. Therefore, these information suggested that the restricting HLA alleles of C45-57 and NS3141-155 could be closely related with DPw4 and DR15, respectively. In this study, E396-408 and NS323-35 are two novel identified CD4+ T-cell epitopes. Since E protein is the important structural protein in DEN infection and immunity, more researchers aimed to identified T-cell epitopes located in this protein and defined some T-cell epitopes (Rothman et al., 1996, Simmons et al., 2005, Bashyam et al., 2006). However, to our knowledge, E396-408 is the first CD4+ T-cell epitopes identified in human population. Importantly, E396-408 is only highly conserved in DEN-1 strains. Therefore, the identification of E396-408 would help to explore the role of T-cell immune response induced by E protein in DEN infection or immunity. Previous study shown that NS3 protein appears to be the most immunogenic antigen for cellular responses to DEN and there exist many T-cell epitopes in NS3200-324 (Mathew et al., 1996). Consequently, most research focused on identifying T-cell epitopes located in this region (Kurane et al., 1995, Kurane et al., 1998). In this study, the identification of NS323-35 suggested that the other region of NS3 could also contribute to T-cell immune response.
In conclusion, this study indicated a good correlation between experimentally observed and computationally predicted peptides. Therefore, the combination of bioinformatics tools and in vitro assays is a better strategy for the identification of T-cell epitopes.
Acknowledgement
The work was supported by grants from the team Project of Nature Science Foundation of Guangdong Province, China (5200638).
References
- Bashyam H.S., Green S., Rothman A.L. Dengue virus-reactive CD8+T cells display quantitative and qualitative differences in their response to variant epitopes of heterologous viral serotypes. J. Immunol. 2006;176:2817–2824. doi: 10.4049/jimmunol.176.5.2817. [DOI] [PubMed] [Google Scholar]
- Butz E.A., Bevan M.J. Massive expansion of antigen-specific CD8+T cells during an acute virus infection. Immunity. 1998;8:167–175. doi: 10.1016/s1074-7613(00)80469-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drabner B., Reineke U., Schneider-Mergener J., Humphreys R.E., Hartmann S., Lucius R. Identification of T helper cell-recognized epitopes in the chitinase of the filarial nematode Onchocerca volvulus. Vaccine. 2002;20:3685–3694. doi: 10.1016/s0264-410x(02)00395-x. [DOI] [PubMed] [Google Scholar]
- Gagnon S.J., Zeng W., Kurane I., Ennis F.A. Identification of two epitopes on the dengue 4 virus capsid protein recognized by a serotype-specific and a panel of serotype-cross-reactive human CD4+cytotoxic T-lymphocyte clones. J. Virol. 1996;70:141–147. doi: 10.1128/jvi.70.1.141-147.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibbons R.V., Vaughn D.W. Dengue: an escalating problem. BMJ. 2002;324:1563–1566. doi: 10.1136/bmj.324.7353.1563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez-Nunez M., Pinilla-Ibarz J., Dao T., May R.J., Pao M., Jaggi J.S., Scheinberg D.A. Peptide binding motif predictive algorithms correspond with experimental binding of leukemia vaccine candidate peptides to HLA-A*0201 molecules. Leuk. Res. 2006;30:1293–1298. doi: 10.1016/j.leukres.2006.02.010. [DOI] [PubMed] [Google Scholar]
- Green S., Pichyangkul S., Vaughn D.W., Kalayanarooj S., Nimmannitya S., Nisalak A., Kurane I., Rothman A.L., Ennis F.A. Early CD69 expression on peripheral blood lymphocytes from children with dengue hemorrhagic fever. J. Infect. Dis. 1999;180:1429–1435. doi: 10.1086/315072. [DOI] [PubMed] [Google Scholar]
- Gubler D.J. Dengue and dengue hemorrhagic fever. Clin. Microbiol. Rev. 1998;11:480–496. doi: 10.1128/cmr.11.3.480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halstead S.B. Pathogenesis of dengue: challenges to molecular biology. Science. 1988;239:476–481. doi: 10.1126/science.3277268. [DOI] [PubMed] [Google Scholar]
- Huang Y.H., Tao M.H., Hu C.P., Syu W.J., Wu J.C. Identification of novel HLA-A*0201-restricted CD8+T-cell epitopes on hepatitis delta virus. J. Gen. Virol. 2004;85:3089–3098. doi: 10.1099/vir.0.80183-0. [DOI] [PubMed] [Google Scholar]
- Kuno G., Gubler D.J., Santiago de Weil N.S. Antigen capture ELISA for the identification of dengue viruses. J. Virol. Methods. 1985;12:93–103. doi: 10.1016/0166-0934(85)90011-4. [DOI] [PubMed] [Google Scholar]
- Kurane I., Okamoto Y., Dai L.C., Zeng L.L., Brinton M.A., Ennis F.A. Flavivirus-cross-reactive HLA-DR15-restricted epitope on NS3 recognized by human CD4+ CD8− cytotoxic T lymphocyte clones. J. Gen. Virol. 1995;76:2243–2249. doi: 10.1099/0022-1317-76-9-2243. [DOI] [PubMed] [Google Scholar]
- Kurane I., Zeng L., Brinton M.A., Ennis F.A. Definition of an epitope on NS3 recognized by human CD4+ cytotoxic T lymphocyte clones cross-reactive for dengue virus types 2, 3, and 4. Virology. 1998;240:169–174. doi: 10.1006/viro.1997.8925. [DOI] [PubMed] [Google Scholar]
- Lanciotti R.S., Calisher C.H., Gubler D.J., Chang G.J., Vorndam A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992;30:545–551. doi: 10.1128/jcm.30.3.545-551.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lei H.Y., Yeh T.M., Liu H.S., Lin Y.S., Chen S.H., Liu C.C. Immunopathogenesis of dengue virus infection. J. Biomed. Sci. 2001;8:377–388. doi: 10.1007/BF02255946. [DOI] [PubMed] [Google Scholar]
- Libraty D.H., Endy T.P., Houng H.S., Green S., Kalayanarooj S., Suntayakorn S., Chansiriwongs W., Vaughn D.W., Nisalak A., Ennis F.A., Rothman A.L. Differing influences of viral burden and immune activation on disease severity in secondary dengue 3 virus infections. J. Infect. Dis. 2002;185:1213–1221. doi: 10.1086/340365. [DOI] [PubMed] [Google Scholar]
- Mangada M.M., Ennis F.A., Rothman A.L. Quantitation of dengue virus specific CD4+ T cells by intracellular cytokine staining. J. Immunol. Methods. 2004;284:89–97. doi: 10.1016/j.jim.2003.10.003. [DOI] [PubMed] [Google Scholar]
- Mathew A., Kurane I., Rothman A.L., Zeng L.L., Brinton M.A., Ennis F.A. Dominant recognition by human CD8+cytotoxic T lymphocytes of dengue virus nonstructural proteins NS3 and NS1.2a. J. Clin. Invest. 1996;98:1684–1691. doi: 10.1172/JCI118964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miskovsky E.P., Liu A.Y., Pavlat W., Viveen R., Stanhope P.E., Finzi D., Fox W.M., 3rd., Hruban R.H., Podack E.R., Siliciano R.F. Studies of the mechanism of cytolysis by HIV-1-specific CD4+ human CTL clones induced by candidate AIDS vaccines. J. Immunol. 1994;153:2787–2799. [PubMed] [Google Scholar]
- Mongkolsapaya J., Dejnirattisai W., Xu X.N., Vasanawathana S., Tangthawornchaikul N., Chairunsri A., Sawasdivorn S., Duangchinda T., Dong T., Rowland-Jones S., Yenchitsomanus P.T., McMichael A., Malasit P., Screaton G. Original antigenic sin and apoptosis in the pathogenesis of dengue hemorrhagic fever. Nat. Med. 2003;9:921–927. doi: 10.1038/nm887. [DOI] [PubMed] [Google Scholar]
- Mothe B.R., Sidney J., Dzuris J.L., Liebl M.E., Fuenger S., Watkins D.I., Sette A. Characterization of the peptide-binding specificity of Mamu-B*17 and identification of Mamu-B*17-restricted epitopes derived from simian immunodeficiency virus proteins. J. Immunol. 2002;169:210–219. doi: 10.4049/jimmunol.169.1.210. [DOI] [PubMed] [Google Scholar]
- Pala P., Hussell T., Openshaw P.J. Flow cytometric measurement of intracellular cytokines. J. Immunol. Methods. 2000;243:107–124. doi: 10.1016/s0022-1759(00)00230-1. [DOI] [PubMed] [Google Scholar]
- Reche P.A., Glutting J.P., Reinherz E.L. Prediction of MHC class I binding peptides using profile motifs. Hum. Immunol. 2002;63:701–709. doi: 10.1016/s0198-8859(02)00432-9. [DOI] [PubMed] [Google Scholar]
- Reche P.A., Glutting J.P., Zhang H., Reinherz E.L. Enhancement to the RANKPEP resource for the prediction of peptide binding to MHC molecules using profiles. Immunogenetics. 2004;56:405–419. doi: 10.1007/s00251-004-0709-7. [DOI] [PubMed] [Google Scholar]
- Rothman A.L., Kurane I., Ennis F.A. Multiple specificities in the murine CD4+and CD8+T-cell response to dengue virus. J. Virol. 1996;70:6540–6546. doi: 10.1128/jvi.70.10.6540-6546.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rothman A.L., Ennis F.A. Immunopathogenesis of dengue hemorrhagic fever. Virology. 1999;257:1–6. doi: 10.1006/viro.1999.9656. [DOI] [PubMed] [Google Scholar]
- Simmons C.P., Dong T., Chau N.V., Dung N.T., Chau T.N., Thao le T.T., Hien T.T., Rowland-Jones S., Farrar J. Early T-cell responses to dengue virus epitopes in Vietnamese adults with secondary dengue virus infections. J. Virol. 2005;79:5665–5675. doi: 10.1128/JVI.79.9.5665-5675.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turcanová V., Höllsberg P. Sustained CD8+T-cell immune response to a novel immunodominant HLA-B*0702-associated epitope derived from an Epstein-Barr virus helicase-primase-associated protein. J. Med. Virol. 2004;72:635–645. doi: 10.1002/jmv.20023. [DOI] [PubMed] [Google Scholar]
- Vergelli M., Hemmer B., Muraro P.A., Tranquill L., Biddison W.E., Sarin A., McFarland H.F., Martin R. Human autoreactive CD4+ T cell clones use perforin- or Fas/Fas ligand-mediated pathways for target cell lysis. J. Immunol. 1997;158:2756–2761. [PubMed] [Google Scholar]
- Waldrop S.L., Pitcher C.J., Peterson D.M., Maino V.C., Picker L.J. Determination of antigen-specific memory/effector CD4+T cell frequencies by flow cytometry: evidence for a novel, antigen-specific homeostatic mechanism in HIV-associated immunodeficiency. J. Clin. Invest. 1997;99:1739–1750. doi: 10.1172/JCI119338. [DOI] [PMC free article] [PubMed] [Google Scholar]