

Highlights
-
•
Chikungunya fever is caused by a mosquito-borne alphavirus originating in East Africa.
-
•
During the past 7 years, the disease has spread to islands of the Indian Ocean, Asia and Europe.
-
•
Its spread has been facilitated by a mutation favouring replication in the mosquito Ae. albopictus.
-
•
No vaccines or antiviral drugs are available to prevent or treat chikungunya fever.
-
•
This paper provides an extensive review of the virus and disease, including Supplementary Tables.
Keywords: Chikungunya virus, Chikungunya fever, Arbovirus, Alphavirus, Antiviral therapy
Abstract
Chikungunya virus (CHIKV) is the aetiological agent of the mosquito-borne disease chikungunya fever, a debilitating arthritic disease that, during the past 7 years, has caused immeasurable morbidity and some mortality in humans, including newborn babies, following its emergence and dispersal out of Africa to the Indian Ocean islands and Asia. Since the first reports of its existence in Africa in the 1950s, more than 1500 scientific publications on the different aspects of the disease and its causative agent have been produced. Analysis of these publications shows that, following a number of studies in the 1960s and 1970s, and in the absence of autochthonous cases in developed countries, the interest of the scientific community remained low. However, in 2005 chikungunya fever unexpectedly re-emerged in the form of devastating epidemics in and around the Indian Ocean. These outbreaks were associated with mutations in the viral genome that facilitated the replication of the virus in Aedes albopictus mosquitoes. Since then, nearly 1000 publications on chikungunya fever have been referenced in the PubMed database. This article provides a comprehensive review of chikungunya fever and CHIKV, including clinical data, epidemiological reports, therapeutic aspects and data relating to animal models for in vivo laboratory studies. It includes Supplementary Tables of all WHO outbreak bulletins, ProMED Mail alerts, viral sequences available on GenBank, and PubMed reports of clinical cases and seroprevalence studies.
1. Introduction
Chikungunya virus (CHIKV) is an arthropod-borne virus that is transmitted by Aedes (Ae.) mosquitoes. It was first isolated in 1952 in the Makonde Plateau of the southern province of Tanzania (former Tanganyika). The virus transmission cycle requires infection of female mosquitoes via a viraemic bloodmeal taken from a susceptible vertebrate host and, following a suitable extrinsic incubation period, transmission to another vertebrate host during subsequent feeding (Solignat et al., 2009). After an incubation period, most patients suffer from polyarthralgia and myalgia, with a significant impact on their quality of life. Chikungunya fever is characterised by a very high viraemic load and concomitant abnormalities such as pronounced lymphopenia and moderate thrombocytopenia. The rate of asymptomatic cases is lower, and the percentage of infected patients requiring medical attention is higher, than in most other common arboviral infections. After the acute stage, some patients experienced relapse, persistent arthralgia or musculoskeletal pains. Increase of age is the most obvious risk factor associated with severe disease or persistent symptoms in adults, whilst in paediatric populations, newborns have a higher risk of severe disease.
Since the first reports of chikungunya fever in Africa in the early 1950s, more than 1500 scientific publications on different aspects of the disease and its causative agent have been produced. Analysis of these publications shows that, following a number of studies in the 1960s and 1970s, and in the absence of autochthonous cases in developed countries, the interest of the scientific community remained low (Fig. 1 ). However, in 2005 chikungunya fever unexpectedly re-emerged in the form of devastating epidemics in and around the Indian Ocean. These outbreaks were associated with mutations in the viral genome that facilitated the replication of the virus in Aedes albopictus mosquitoes. Since then, nearly 1000 publications on chikungunya fever have been referenced in the PubMed database. The reader is referred to Supplementary Tables 1–6 for lists of all WHO outbreak bulletins, ProMED Mail alerts, viral sequences available on GenBank, and PubMed reports of clinical cases and seroprevalence studies.
Fig. 1.
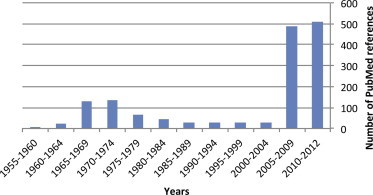
Publications related to outbreaks of chikungunya fever in the PubMed database. Articles published between 1950 and September, 2012 were identified using the MeSH term “chikungunya,” and are reported by 5-year periods.
Two distinct transmission cycles have been described for CHIKV: a sylvatic cycle in Africa and an urban human–mosquito–human virus transmission cycle seen in Asia, the Indian Ocean, Africa and more recently, in Europe. The two major vectors of the disease currently identified are Ae. aegypti and since 2006, Ae. albopictus. The important role of Ae. albopictus in recent outbreaks is due to adaptive mutations of the viral genome, in particular the A226V mutation in the E1 glycoprotein, that increase viral replication in this specific vector. Based on the partial E1 structural glycoprotein or complete genomic sequences, three phylogroups of CHIKV (West-African, Asian, and East-Central-South-African) which apparently circulate in regions that display different ecological backgrounds have been identified.
No licenced vaccine against chikungunya is commercially available, but several strategies are under study. In sections below we review several drugs which have shown antiviral activity against CHIKV or activity against the inflammatory symptoms associated with CHIKV infection. Treatment of standard presentations of chikungunya fever currently relies on paracetamol/acetaminophen and non-steroidal anti-inflammatory drugs. Chloroquine is not recommended at the acute phase of the disease. Ribavirin has been used in some severe presentations but very limited information is available to confirm its efficacy. Recent investigations may lead to the identification of new antiviral candidates with a clearly defined mechanism of viral inhibition in cell-based systems and significant activity in animal models. Therapeutic protocols for severe cases may also be established based on specific immunoglobulins or molecules that can interfere with some aspects of the inflammatory response associated with CHIKV infection. For chronic rheumatic manifestation and inflammatory polyarthritis lasting more than 2–3 months, disease-modifying anti-rheumatic drugs such as methotrexate are recommended. As described below, studies in animal models suggest that inflammation, macrophage tissue tropism and local viral persistence are involved in the establishment of chronic disease.
2. Chikungunya virus in brief
2.1. Classification
CHIKV belongs to the alphavirus genus of the family Togaviridae (Enserink, 2006, King et al., 2012, Powers and Logue, 2007). CHIKV was first isolated from the serum of a febrile patient during an outbreak that occurred in the southern province of Tanzania (Tanganyika, Makonde Plateau) in 1952–1953 (Robinson, 1955, Ross, 1956). The name “Chikungunya” in the Bantu language of the Makonde people refers to the stooped posture due to the frequent and debilitating joint pain induced during chikungunya fever (Enserink, 2006). There is substantial evidence that CHIKV is transmitted and dispersed by mosquito vectors, in particular by Aedes species (spp.) (Gilotra and Bhattacharya, 1968, Gilotra and Shah, 1967, McIntosh et al., 1963, Paterson and McIntosh, 1964, Sarkar, 1967, Shah et al., 1964, Weinbren et al., 1958).
CHIKV is a member virus of the Semliki Forest (Eurasia) virus antigenic complex, together with a number of other alphaviruses that are found in Africa (O’nyong–nyong virus (ONNV)), in South America (e.g., Mayaro virus) and in the Australia/Oceania region (Ross River virus (RRV)) that cause acute arthropathy in humans (Powers et al., 2000, Vanlandingham et al., 2006).
2.2. Virus structure and genomic organisation
The virion has an icosahedral capsid enclosed by a lipid envelope and a diameter of 60–70 nm. It is sensitive to desiccation and to temperatures >58 °C. The genome is a single-stranded, positive sense, RNA molecule of approximately 12 kb in length (Khan et al., 2002) (Fig. 2 ). The genomic organisation is arranged as with others alphaviruses: 5′-nsP1–nsP2–nsP3–nsP4-junction region-C–E3–E2–6k–E1-poly (A)-3′ with two open reading frames (ORFs). The 5′ end of the genome has a 7-methylguanosine cap and there is a polyadenylation signal at the 3′ end. The 5′ ORF is translated from genomic RNA and encodes four non-structural proteins (nsP1, 2, 3 and 4) (Jose et al., 2009). The 3′ ORF is translated from a subgenomic 26S RNA and encodes a polyprotein that is processed as the capsid protein (C), two surface envelope glycoproteins (E1 and E2) and two small peptides designated E3 and 6k (King et al., 2012, Simizu et al., 1984, Voss et al., 2010).
Fig. 2.
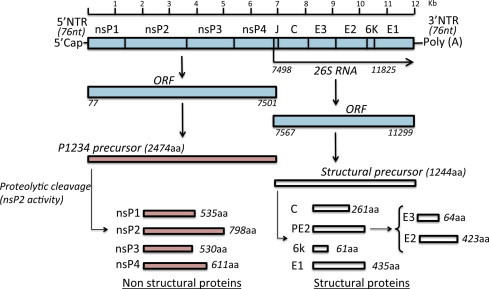
Organisation of the CHIKV genome and gene products. The genomic organisation is arranged as with other alphaviruses, with two open reading frames (ORFs), 5′ cap structures and a 3′ poly(A) tail. The 5′ and 3′ proximal sequences of the CHIKV genome include nontranslated regions (NTR). The junction region (J) is also noncoding. The 5′ ORF is translated from genomic RNA and encodes four nonstructural proteins (nsP1, 2, 3 and 4). The 3′ ORF is translated from a subgenomic 26S RNA and encodes the capsid protein (C), two surface envelope glycoproteins (E1 and E2) and two small peptides designated E3 and 6k. The different non-structural proteins (nsP1–nsP4) and structural proteins (C, Capsid; E1, E2, E3, envelope; 6k) are generated after proteolytic cleavage of polyprotein precursors (adapted from Solignat et al., 2009, with authors’ permission).
The glycoproteins E1 and E2 are embedded in a heterodimeric form in the viral envelope and are responsible for virus attachment and membrane fusion. Virus fusion with the cell membrane is mediated by the E1 glycoprotein, a class II fusion protein, in a process dependent on low-pH. Acidic conditions induce a conformational change in the virus envelope proteins, dissociation of the E2–E1 heterodimers and formation of E1 homotrimers. The E1 trimer is inserted into the target membrane via its hydrophobic fusion peptide and refolds to form a hairpin-like structure. Cholesterol is required for both cell membrane fusion and budding during alphavirus infection (Solignat et al., 2009). A large number of more recent studies have been dedicated to the structural characterisation of the envelope proteins of alphaviruses and to structural modifications that occur during fusion (Gibbons et al., 2004, Li et al., 2010, Liu and Kielian, 2009, Roussel et al., 2006, Voss et al., 2010, Zhang et al., 2011).
2.3. Replication cycle
The CHIKV replication cycle is essentially similar to the replication cycle of other alphaviruses (Solignat et al., 2009). The non-structural proteins (nsP1–4) and their cleavage intermediates are involved in RNA replication. The five structural proteins (C, E3, E2, 6k, E1) and their cleavage intermediates are required for viral encapsidation and budding (Fros et al., 2010, Leung et al., 2011). Replication occurs in the cytoplasm, both in vertebrate and insect cells, in close association with the Golgi apparatus.
Virus enters cells at the plasma membrane, mostly by endocytosis, via a pH-dependent mechanism which culminates in fusion pore formation and release of the nucleocapsid into the cytosol (Fig. 3 ). It begins with attachment (E2 is primarily responsible for interactions with cellular receptors) and fusion of virus particles in the membrane of the host cell. The fusion peptide is located at the tip of the E1 molecule in domain II, close to amino acid 226. However, a CHIKV resistant mutant selected and adapted to growth in the presence of high concentrations of Arbidol (a molecule that typically blocks fusion (Boriskin et al., 2008, Leneva et al., 2009)) included a single amino acid substitution (G407R) localised in the E2 envelope protein (Delogu et al., 2011), suggesting that close cooperation between E1 and E2 proteins is necessary during the fusion process.
Fig. 3.
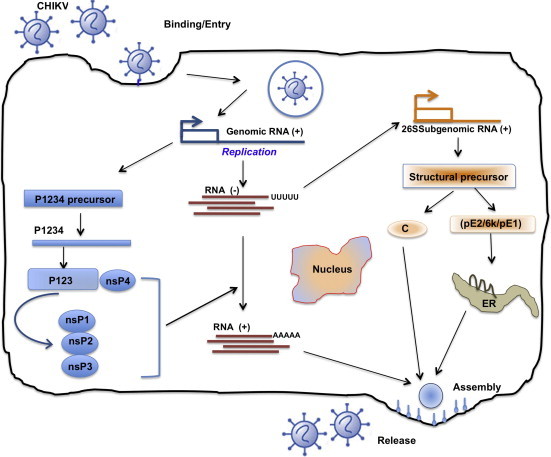
Life cycle of CHIKV in infected cells. Virus enters cells at the plasma membrane, mostly by endocytosis, via a pH-dependent mechanism, which culminates in fusion pore formation and release of the nucleocapsid into the cytosol. It begins with attachment (E2 is primarily responsible for interactions with cellular receptors) and fusion of virus particles with the host cell membrane. The fusion peptide is located at the tip of the E1 molecule in domain II, close to amino acid 226. Following virus entry, two rounds of translation occur. Positive-sense genomic RNA acts directly as mRNA and is partially translated (5’ end) to produce non-structural proteins (nsP’s). These proteins are responsible for replication and formation of a complementary negative strand, the template for further positive-strand synthesis. Subgenomic mRNA (26S) replication occurs through the synthesis of full-length negative intermediate RNA, which is regulated by nsP4 and p123 precursor in early infection, and later by mature nsPs. Translation of the 26S sub-genomic RNA results in production of 5 structural proteins (C, E3, E2, 6k, E1), which are required for viral encapsidation and budding. Assembly occurs at the cell surface, and the envelope is acquired as the virus buds from the cell. Release and maturation occur almost simultaneously.
From Thiboutot et al. (2010), with permission.
3. Clinical syndrome
3.1. Incubation period
The incubation period of chikungunya fever has never been investigated in great detail, but a range of 1–12 days is often cited (Burt et al., 2012, Singh and Unni, 2011). In 1977, an outbreak occurred amongst a group of high-school children from Pretoria after a visit to the northern Transvaal bushveld; two days after their arrival, the first campers were taken ill. Overall, the incubation period during this episode was estimated to vary from 2 to 10 days (Fourie and Morrison, 1979).
Amongst cases imported into metropolitan France during the 2006 Indian Ocean outbreak, 43 patients had a defined onset of disease after their departure from the endemic area. The mean duration of incubation following arrival (which represented the minimal duration of incubation) was 1.6 days (95% CI: 1.1–2.1), with a range of 0–8 days and a median of 1 day (data from Institut de Veille Sanitaire – InVS – 2006, personal communication: Harold Noël). These data are globally in accordance with the medical literature and support the belief that the incubation period is generally short (2–10 days).
3.2. Mild cases and asymptomatic infections
The proportion of individuals infected by CHIKV who develop clinical symptoms requiring medical attention is higher than in most other arboviral infections (Chastel, 2011). Following outbreaks of chikungunya fever, several recent seroprevalence studies were performed indicating different rates of asymptomatic cases ranging from 3.8% to 27.7%. After the outbreak in Reunion island, 162/967 (16.7%) cases reported no symptoms including 46/967 (4.8%) who ‘did not know’ (Gerardin et al., 2008b). Several months after the outbreak in Italy, 6/33 (18.2%) cases with positive serology declared no symptoms (Moro et al., 2010). Similarly, after the outbreak in Mayotte, 122/440 (27.7%) cases with positive serology declared no clinical signs that could be compatible with a history of chikungunya fever (Sissoko et al., 2010).
Two years after the chikungunya fever outbreak in the state of Kerala in India, a seroprevalence study found that amongst the 260 patients with positive serology, only 10 (3.8%) declared no symptoms compatible with a history of chikungunya fever (Kumar et al., 2011). In a study amongst French military policemen after the chikungunya outbreak in Reunion island, only 3.2% of cases did not report symptoms (Queyriaux et al., 2008). Finally in a case-control study in Thailand, 9% of laboratory-proven CHIKV infection cases were considered as asymptomatic and the viral loads observed in the symptomatic individuals were not significantly different from those observed in the viremic asymptomatic individuals (Appassakij et al., 2012).
3.3. Clinical features of chikungunya fever
Since the Indian Ocean outbreak in 2005–2006, the information available for the scientific community relating specifically to the clinical characteristics of patients infected by CHIKV has significantly increased. Two stages of the disease are now described: acute illness and the late stage of illness, with persistent arthropathy.
3.3.1. Acute illness
Several recent and prospective studies are currently available to describe accurately the acute stage of illness (Bandyopadhyay and Ghosh, 2010, Borgherini et al., 2007, Chow et al., 2011, Nkoghe et al., 2012, Queyriaux et al., 2008, Rezza et al., 2007, Riyaz et al., 2010, Simon et al., 2007, Staikowsky et al., 2009, Taubitz et al., 2007, Thiberville et al., 2013, Win et al., 2010).
Symptomatic patients generally report an abrupt onset of disease characterised by high fever, polyarthralgia, backache, headache, and fatigue. Fever and characteristic clinical symptoms appear within 4–7 days.
Poly-arthralgia is reported in 87–98% of cases and represents the most characteristic symptom. Joint pain is mostly polyarticular, bilateral, symmetrical and occurs mainly in peripheral joints (wrists, ankles, and phalanges) and some large joints (shoulders, elbows and knees). Joint swelling is less frequent: 25–42% of cases. Pains in the ligaments (pubalgia, sternocleidomastoid, occipital insertions and talalgia), temporomandibular or sternocostoclavicular joints and tenosynovitis, have also been described. Myalgia was observed in 46–59% of cases in recent prospective studies, whilst retrospective studies reported a higher prevalence (93%). Myalgia has been observed predominantly in the arms, thighs and calves without myositis.
Cutaneous manifestations were reported in 40–50% of cases and characterised by a macular or maculopapular rash involving mainly the extremities, trunk and face. The skin lesions are transient and mostly occur a few (2–5) days after the onset of the disease. Generalised pruritus was reported in one-fourth of cases. A large variety of skin and mucous membrane lesions has also been reported during the acute stage of the disease: hypermelanosis, hyperpigmentation, photosensitivity, exfoliative dermatitis, vesicles, bullae, vasculitic lesions, erythema nodosum like lesions, exacerbation of pre-existing dermatoses such as psoriasis and mucosal ulceration.
Digestive symptoms such as diarrhoea, vomiting, nausea or abdominal pain occurs in 15–47% of cases during the acute stage of the disease. As a consequence, chikungunya fever has a major impact on quality of life during the acute stage of illness. Incapacitation or limitation of normal activity occurs in more than 60% of cases whilst tiredness is considered as significant or very significant in 47% of cases. Psychological impact was observed and some patients were depressed or demoralised.
More recently, a study described the classical and daily evolution (until day 14 after onset) of the main symptoms of a cohort of outpatients infected by CHIKV during the Reunion island outbreak. Two stages were identified: the ‘viral stage’ (days 1–4), associated with rapid decrease of viraemia, followed by rapid improvement of clinical presentation and the ‘convalescent stage’ (days 5–14) that was associated with no detectable viraemia and a slow clinical improvement (Thiberville et al., 2013).
3.3.2. Viraemia and changes in clinical laboratory tests
The acute stage of chikungunya fever is characterised by a very high viraemic load (107 pfu/ml on average) with a median duration of viraemia of 6 days (range 3–10 days) and concomitant abnormalities such as pronounced lymphopenia and/or moderate thrombocytopenia. In a prospective series of cases during the Reunion island outbreak in 2006, 79% of patients had a lymphopenia (<1000 cells/mm3), which was severe (<500 cells/mm3) in 39% of cases. Lymphopenia was more pronounced in patients with higher viraemia. Moderate thrombocytopenia (<150,000 cells/mm3 and >100,000 cells/mm3) was recorded in 40–50% of patients. Other less common biological abnormalities have been observed such as leukopenia, elevated liver enzymes, anemia, elevated creatinine, elevated creatinine kinase and hypocalcemia.
3.3.3. Differential diagnosis
Differentiating chikungunya fever from other possible etiologies is important for physicians, notably to ensure good management of outpatients. In a retrospective serological study in Mayotte Island after the outbreak of 2006, Sissoko et al. (2010) found that fever together with polyarthralgia provided an 84% sensitivity (Se) with a 74% positive predictive value (PPV) and an 83% negative predictive value (NPV). In a prospective study of patients with suspected chikungunya fever during a recent outbreak in Gabon, fever and arthralgia had a Se of 73% with a PPV and NPV of 79% and 44% respectively (Nkoghe et al., 2012). In another study, Staikowsky et al. (2009) reported that CHIKV positive patients had an increased frequency of skin rashes, arthralgia on the feet and wrist, asthenia, a higher temperature and a lower frequency of digestive symptoms and pruritus than CHIKV negative patients.
In a retrospective study aiming to compare dengue and chikungunya fever on presentation, chikungunya was independently associated with arthralgia and rash, whilst dengue was associated with myalgia, raised aspartate transaminase, and leucopaenia (Mohd Zim et al., 2013). Biological parameters had also been associated with CHIKV-positive patients, notably lymphopenia, elevated liver enzymes, anemia, and elevated creatinine (Nkoghe et al., 2012, Staikowsky et al., 2009). More recently, clinical and clinicobiological diagnostic scores based on a cohort of outpatients during the Reunion island outbreak have been proposed, using very simple criteria (arthralgia on hands and wrists, minor or absent myalgia that can be combined (clinico-biological score) or not (clinical score) with the presence of lymphopenia (<1G/L)) (Thiberville et al., 2013).
3.3.4. Late stage of illness and persistent arthropathy
After the acute stage described above, some patients experience relapse or persistent symptoms. The clinical description and the risk factors of the late stage have recently gained increased interest as evidence by several studies (Borgherini et al., 2008, Bouquillard and Combe, 2009, Couturier et al., 2012, de Andrade et al., 2010, Gerardin et al., 2011, Hoarau et al., 2010a, Manimunda et al., 2010, Marimoutou et al., 2012, Moro et al., 2012, Narsimulu and Prabhu, 2011, Nkoghe et al., 2012, Ribera et al., 2012, Simon et al., 2007, Sissoko et al., 2009, Thiberville et al., 2013, Win et al., 2010).
Amongst the symptoms declared during the late stage, arthralgia or musculoskeletal pains were the most frequent long-lasting signs. Following the chikungunya fever outbreak in Reunion island, several studies have prospectively followed-up patients who presented with chronic rheumatic manifestations. Rheumatoid arthritis (RA) according to the American College of Rheumatology (ACR) criteria, spondylarthopathy and other non-classified rheumatism were diagnosed and associated with the chikungunya fever. Others symptoms were less frequently reported such as fever, fatigue, headaches, neuropathic pain syndromes, cerebral disorders, sensorineural impairments, dysesthesia and/or paraesthesia, carpal, tarsal, or cubital tunnel syndromes, digestive disorders, skin involvement, rash, alopecia, pruritus, bilateral Raynaud phenomenon or erythermalgia, joint stiffness, bursitis, tenosynovitis and synovitis with or without effusions. As a consequence, the quality of life and psychological health could be impaired at the late stage. Except for some patients with a diagnosis of RA, standard laboratory findings including inflammatory markers are remarkably within normal limits during this stage.
The proportion of chikungunya patients who fully recovered, partially improved or had persistent symptoms vary between studies and according to the time of the study assessment from the onset of the disease. In all cases, the frequency of persons presenting with symptoms of chikungunya fever decreased with increasing time of onset. In a cross-sectional study after the outbreak on Reunion island, 16% of patients declared that they had suffered for less than one month, 31% between one and three months and 53% still suffered, on average, 128 days after the onset of the disease.
In an outpatient cohort during the Reunion island outbreak, approximately 20% of patients declared incomplete recovery or persistent arthralgia 300 days after the onset of the disease. Twelve months after the outbreak in Italy, 66% and 60.8% of cases still reported at least one of the most frequent symptoms (arthralgia, myalgia and asthenia) or only arthralgia respectively. In another study, 15 months after the outbreak of Mayotte 2006, 36% of patients reported that they had permanent symptoms and 21% reported experiencing at least one episode of recurrence. In contrast, during the outbreak of Gabon 2010, 83% of patients recovered fully by 30 days after the onset of the disease. In West India after the 2006 epidemic, 12% and 5% suffered from musculoskeletal pains and arthritis at 1 and 2 years after the disease onset respectively.
3.3.5. Risk factors for persistent arthropathy
Dupuis-Maguiraga et al. (2012) have reviewed the host and virological factors recorded during the late stage of clinical disease. Increased age was associated with a longer duration of illness and the presence of joint pain at the late stages of infection. In contrast age was not a risk factor for chronic disease during the outbreak in Singapore, possibly due to a younger population being studied. Pre-existing joint pain or osteoarthritis comorbidity are also associated with an increased risk of persistent symptoms. Female gender was associated with a longer duration of illness and persistent symptoms, but only after univariate analysis.
Amongst the biological markers of the acute stage, viral load has been associated with persistent symptoms during the Reunion island outbreak whereas no relationship between viral load and late stage was found during the outbreak in Singapore. A strong early Immunoglobulin G3 (IgG3) response triggered by a high viral load was also proposed to protect against chronic long-term effects of CHIKV infection. Recently, two studies have observed a positive association between high titres of CHIKV specific IgG antibodies and long lasting arthralgias (Gerardin et al., 2013, Moro et al., 2012). In one study, peak creatinine levels were significantly lower in patients with persistent arthralgia (Win et al., 2010).
3.4. Atypical cases
Since the 1960–1970s CHIKV has been known to affect the central nervous system. During the recent Indian Ocean outbreak, neurological complications were reported in less than 25% of cases. Seizures were most often reported in patients who had a past medical history of epilepsy and/or in those with a history of heavy alcohol consumption. Other neurological complications reported are encephalopathy, encephalitis, Guillain–Barre, encephalomyeloradiculitis or subarachnoid cerebellar haemorrhages (Borgherini et al., 2007, Das et al., 2010, Staikowsky et al., 2009).
Haemorrhagic signs are rare (1–7% of patients) and minor, such as bleeding of the nose and gums (Borgherini et al., 2007, Win et al., 2010). In most cases, they are not associated with clotting abnormalities or major thrombocytopenia (Nkoghe et al., 2012, Staikowsky et al., 2009).
A variety of other clinical symptoms have been reported during the acute stage of chikungunya fever, such as conjunctivitis, neuroretinitis, iridocyclitis, myocarditis, pericarditis, pneumonia, dry cough, lymphadenopathy, nephritis, hepatitis and pancreatitis (Borgherini et al., 2007, Economopoulou et al., 2009, Mahendradas et al., 2008, Mirabel et al., 2007, Nair et al., 2012, Rajapakse et al., 2010, Renault et al., 2007, Rezza et al., 2007, Rose et al., 2011, Simon et al., 2007, Simon et al., 2008, Staikowsky et al., 2009, Win et al., 2010). Overall, atypical cases were estimated at 0.3% of all symptomatic cases during the 2006 Reunion island outbreak (Economopoulou et al., 2009). Amongst these atypical cases, 36% were considered to be severe, 14% were admitted to an intensive care unit and 10% died (Economopoulou et al., 2009). The reported causes of death were heart failure, multiple organ failure syndrome, toxic hepatitis, encephalitis, bullous dermatosis, respiratory failure, renal failure, pneumonia, acute myocardial infarction, cerebrovascular disease, hypothyroidism or septicaemia.
During the Reunion island outbreak, an excessive death-rate was observed and mortality associated with chikungunya fever was reported to have had a case-fatality ratio of about 1 in 1000 (Josseran et al., 2006). A similar case-fatality ratio was observed following the outbreak in Port Blair, capital city of the Union Territory of Andaman and Nicobar Islands, India, in 2006 (Manimunda et al., 2011). However, the comparison between expected and observed mortality in Reunion island, 2006, identified a high-mortality rate between January and May and a low-mortality rate during the rest of the year (Renault et al., 2012), suggesting the existence of a ‘harvesting’ phenomenon and implying that the global excess of death might have been overestimated when derived only from the epidemic period.
3.5. Chikungunya fever in children
In children, clinical manifestations of chikungunya fever appear to be quite specific and although rheumatological manifestations are less frequent, they remain a group at high risk of atypical or severe manifestations. The main clinical characteristics in infants are the high prevalence of dermatological manifestations (hyperpigmentation, generalised erythema, maculopapular rash and vesiculobullous lesions) and neurological complications (encephalitis, seizures, meningeal syndrome or acute encephalopathy). Other clinical features are also described such as digestive disorder (loose stools), peripheral cyanosis and minor haemorrhagic manifestations (Robin et al., 2008, Robin et al., 2010, Sebastian et al., 2009, Valamparampil et al., 2009).
3.6. Chikungunya fever in pregnant women
Although chikungunya fever apparently has no observable teratogenic effects during pregnancy (Fritel et al., 2010), vertical transmission was reported for the first time, during the 2006 Reunion island outbreak and was observed exclusively in near-term deliveries in the context of intrapartum viremia, with a rate of 49%. Caesarean section had no protective effect on transmission. Severe illness was observed in 53% of newborns and mainly consisted of encephalopathy with persistent disabilities in 44% of them (Gerardin et al., 2008a). The others complications included seizures, haemorrhagic syndrome, haemodynamic disorders, cardiologic complication (myocardial hypertrophy, ventricular dysfunction, pericarditis, coronary artery dilatation), necrotizing enterocolitis or dermatologic manifestation (Nair, 2008, Ramful et al., 2007, Rao et al., 2008).
3.7. Risk factors for severe disease
Risk factors for severe acute disease have been thoroughly investigated (Borgherini et al., 2007, Chow et al., 2011, Economopoulou et al., 2009, Gerardin et al., 2008a, Gerardin et al., 2008c, Kam et al., 2012c, Kee et al., 2010, Kumar et al., 2010, Lohachanakul et al., 2012, Ng et al., 2009b, Nkoghe et al., 2012, Paquet et al., 2006, Renault et al., 2007, Sissoko et al., 2008, Staikowsky et al., 2009). Several studies reported that, in adults, the incidence of atypical cases, severe cases, hospitalisation and the mortality rate increased with age. In pediatric populations, newborns had a high risk of severe disease. Although comorbidity and increase of age are often linked, comorbidity or underlying respiratory diseases, the use of NSAIDs prior to hospitalisation, hypertension and underlying cardiac disorders have been associated with hospitalisation or disease severity. Alcohol abuse was also associated with increased mortality.
Female gender has not been clearly associated with more severe illness but, during the Indian Ocean outbreaks, women were over-represented based on reported cases, whereas cross-sectional studies found similar seroprevalence values for both genders, suggesting a variability of symptomatic disease depending on gender. In contrast, in a single study, males and blood Rhesus-positive individuals were found to be more susceptible to infection by CHIKV. Case reports of immunocompromised patients have described atypical and severe disease but no studies have compared immunocompromised or immunodefficient patients. The association between viral load and acute severe illness is controversial. Indeed, several studies reported higher viral load in hospitalised patients or patients with severe illness whilst others studies did not reveal significant association between the viral load and the clinical presentation. Other biological abnormalities or immunological markers have been proposed to be associated with disease severity such as elevated liver enzyme, creatinine, C reactive protein (CRP), hypocalcemia, elevated IL-1β or IL-6, decreases in RANTES or early CHIKV-specific IgG3.
3.8. Supportive therapy
No studies have precisely evaluated the efficacy of the various symptomatic treatments usually employed during the acute or chronic stage of chikungunya fever. Nevertheless, in previously published reviews, paracetamol/acetaminophen and non-steroidal anti-inflammatory drugs (NSAIDs) have been recommended. Aspirin should be avoided because of the risk of bleeding. Systemic corticosteroids should also be avoided because of the strong rebound effect when the treatment is stopped (Ali Ou Alla and Combe, 2011, Burt et al., 2012, Simon et al., 2011, Singh and Unni, 2011).
For severe chronic arthritis, disease-modifying anti-rheumatic drugs (DMARDs), including methotrexate, hydroxychloroquine or sulphasalazine have been proposed (Bouquillard and Combe, 2009). Although anti-tumour necrosis factor-α (anti-TNF) has been associated with an exacerbated disease in mouse models at the early stage of illness (Zaid et al., 2011), patients with a diagnosis of post-chikungunya rheumatoid arthritis have been successfully treated with anti-TNF antibodies (Bouquillard and Combe, 2009).
Ribera et al. (2012) recently proposed a therapeutic protocol for patients with chronic rheumatic manifestation; for patients without a diagnosis of polyarthritis symptomatic treatment such as analgesic ladder 1 (paracetamol, NSAIDs) or 2 (codeine, tramadol) with kinesitherapia, muscle relaxant drugs, acupuncture and phytotherapia; for patients with a defined inflammatory polyarthritis during more than 2–3 months, DMARDS (methotrexate) are proposed. For patients who do not respond well to the usual analgesics at the chronic stage, neuropathic syndromes should be more carefully evaluated to assess whether or not specific treatments such as antiepileptics and antidepressants could be used to alleviate suffering (de Andrade et al., 2010).
4. Epidemiology
4.1. Arthropod vectors
The main CHIKV mosquito vectors are Ae. aegypti (Stegomya aegypti) and Ae. albopictus (Stegomya albopicta). Until recently, Ae. aegypti was considered the primary vector for CHIKV transmission but in 2006, Ae. albopictus was surprisingly identified as a second major vector of the virus both in places where Ae. aegypti is considered to be rare (e.g., in Reunion island) and also in places where both mosquito species are prevelant (e.g., in Madagascar, India and Gabon). This emergence of Ae. albopictus as a major vector of CHIKV was largely attributable to a single mutation in the E1 protein of CHIKV which facilitated enhanced virus uptake, replication and transmission by the vector (Tsetsarkin et al., 2011b).
Ae. albopictus has recently shown a remarkable capacity to adapt to peri-domestic environments, enabling it to displace Ae. aegypti in some places and to become a significant vector of CHIKV and DENV (Knudsen, 1995). In Africa, a wide range of other mosquitoes has also been incriminated in the transmission of CHIKV, such as other Ae. spp. (Ae. furcifer, Ae. taylori, Ae. vittatus, Ae. fulgens, Ae. luteocephalus, Ae. dalzieli, Ae. vigilax, Ae. camptorhyntites, Ae. africanus) and also Culex annulirostris and Mansonia uniformis (Lam et al., 2001, Yadav et al., 2003).
4.2. Vertebrate reservoirs and transmission cycle
Human beings serve as reservoir hosts for the virus during epidemic periods whereas during inter-epidemic periods, several other reservoir hosts have been incriminated such as monkeys, rodents and birds (Inoue et al., 2003, Wolfe et al., 2001). The significance of non-human primates as reservoir hosts is discussed in the section devoted to pathogenesis.
Two distinct transmission cycles have been described for CHIKV: a sylvatic cycle in Africa (Wolfe et al., 2001), the general pattern of which resembles the yellow fever virus sylvatic cycle (i.e., involving virus transmission between forest-or savannah-associated mosquitoes and non-human primates (and possibly rodents) with occasional spillover of the virus into nearby human populations living in or close to the sylvatic environment). Under these latter circumstances the virus may then encounter mosquitoes primarily associated with urban environments (i.e., domestic or peridomestic mosquitoes) thus initiating an urban human–mosquito–human virus transmission cycle such as those seen in Asia, the Indian Ocean, Africa and more recently, Europe (Rezza et al., 2007). In general, when Ae. aegypti is the predominant vector, these urban outbreaks or epidemics resemble the virus transmission cycle of dengue virus in the urban environment.
In Africa, CHIKV is maintained in a sylvatic cycle involving wild non-human primates and a variety of forest-dwelling mosquito species (Powers and Logue, 2007), such as Ae. africanus in Uganda and Bangui, Ae. cordellieri in South Africa, Ae. furcifer–taylori in South Africa and Senegal (Diallo et al., 1999) and Ae. luteocephalus and Ae. dalzieli in Senegal (Diallo et al., 1999, Jupp and McIntosh, 1990, McIntosh et al., 1963). In rural regions of Africa, the outbreaks tend to affect small populations and appear to be heavily dependent on the sylvatic mosquito population densities that increase during periods of heavy rainfall (Diallo et al., 1999, Thonnon et al., 1999). However, during recent outbreaks in Cameroon, Congo and Gabon, CHIKV was vectored by the Asian mosquito Ae. albopictus which has gradually widened its geographic range, out of Asia into Africa, Europe and the Americas (Benedict et al., 2007). Currently, the principle factors (if indeed there are any) responsible for the maintenance of this Ae. albopictus-adapted virus life cycle in the sylvatic environment have not been defined.
In Asia, CHIKV is maintained in human-mosquito-human urban cycles involving both Ae. aegypti and Ae. albopictus. Whether or not these transmission cycles are exclusive for each mosquito species is not yet known and has not yet been fully investigated. For many years, CHIKV persistence was thought to depend primarily upon continuous introductions of the virus into immunologically naïve populations (Powers and Logue, 2007). However, the evidence for long lasting persistence of the Asian genotype (see infra) and several recent studies suggest that sylvatic cycles could also play a part in the transmission of CHIKV in Asia (Powers et al., 2000, Apandi et al., 2009).
4.3. Infection in humans
CHIKV is endemic and epidemic in Africa, Asia and since 2005, the Indian Ocean (Kariuki Njenga et al., 2008, Powers and Logue, 2007). Cases of chikungunya fever, retrieved from the literature and from different public health alert systems, are shown in Fig. 4 A and B. The following paragraphs summarise the information relating to virus circulation in original endemic areas (i.e., Africa and Asia), to the recent epidemiological expansion and to the possible introduction of the virus in non-endemic areas by infected travellers (Fig. 5 ). Supplementary Tables 1–6 list all WHO outbreak bulletins, ProMED Mail alerts, viral sequences available on GenBank, and PubMed reports of clinical cases and seroprevalence studies.
Fig. 4.
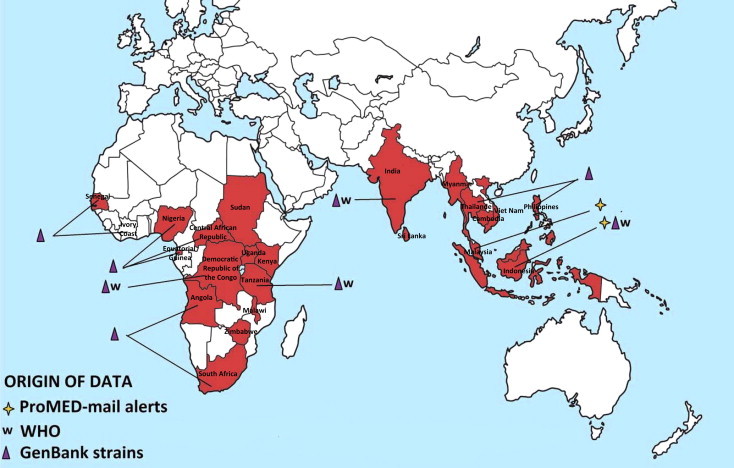
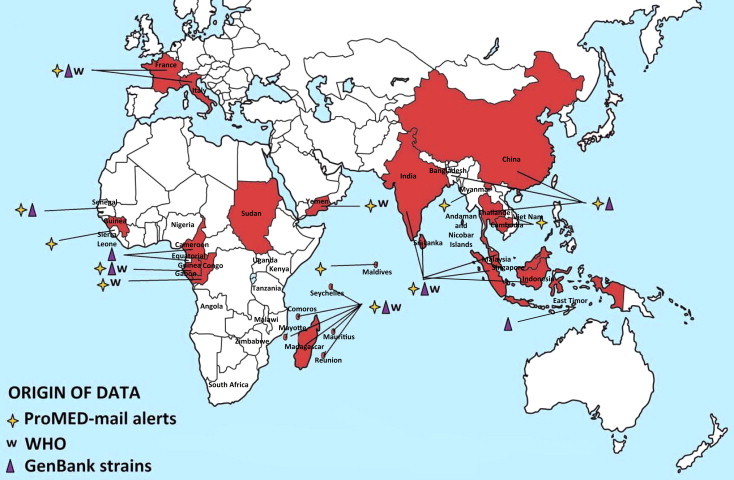
Geographic distribution of CHIKV cases. Autochthonous cases before and after the emergence of CHIKV in the Indian Ocean (2005) are reported in Fig. 4A and B, respectively. Cases were collected from GenBank, PubMed, WHO weekly epidemiological records and the ProMED Mail alert databases. More details and complete tables are provided in the Supplementary Tables.
Fig. 5.
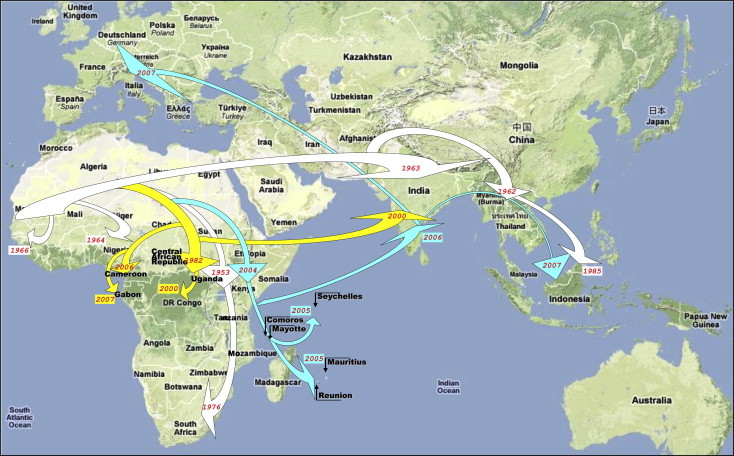
Dispersal pattern of CHIKV from Africa to the Indian Ocean and Europe during the past 20–50 years. Viral evolution and spread are represented according to recent phylogenetic studies. Different evolutionary lineages are identified using arrows with specific colours. This figure was reproduced with permission (de Lamballerie et al., 2008).
4.3.1. Early description of chikungunya
The first human documented epidemic caused by CHIKV was recognised in East Africa and Austral Africa (Tanzania, Uganda and Zimbabwe) in 1952 and 1953 (Mason and Haddow, 1957, Weinbren, 1958, Weinbren et al., 1958). In Asia, the first epidemic was documented in 1958 in Bangkok (Aikat et al., 1964, Volk et al., 2010), followed by a number of outbreaks documented in the Philippines, Cambodia, Vietnam, Laos, Myanmar, Malaysia and Indonesia (Halstead et al., 1969a, Halstead et al., 1969b).
Between 1954 and 1990, the virus was implicated as the cause of epidemics in:
-
•
The Philippines, Thailand, Myanmar (Khai Ming et al., 1974),
-
•
Singapore, Sri Lanka (Hermon, 1967, Mendis, 1967, Vesenjak-Hirjan et al., 1969),
-
•
Cambodia (Chastel, 1964), India, Malaysia, Vietnam (Vu Qui and Nguyen-Thi, 1967),
-
•
Taiwan, Indonesia (Porter et al., 2004) and
-
•
Pakistan (Darwish et al., 1983).
Other ancient possible cases of chikungunya fever in Asia have been reported (Carey, 1971, Ng and Hapuarachchi, 2010) that include a widespread epidemic of self-limited febrile illness in Africa, the Caribbean, West Indies and India in the 1820s, with a large proportion of the population of Calcutta (current Kolkatta, India) affected, and subsequent cases in Calcutta in 1853 and 1871, possibly originating from Zanzibar and then spreading to Aden (Red sea), Calcutta, Burma (Myanmar) and Java. Of note, whilst this epidemic pattern would locate the most ancient reported cases of chikungunya fever in Asia, it is fully compatible with an African origin of the outbreaks.
4.3.2. CHIKV epidemics in Africa and Asia
4.3.2.1. In Africa
Since 1960, cases of chikungunya fever have been reported in several African countries (see Supplementary Table 6):
-
•
South Africa (McIntosh et al., 1963, Paterson and McIntosh, 1964, Powers and Logue, 2007)
-
•
Angola (Filipe and Pinto, 1973, Pinto and Filipe, 1973)
-
•
Niger and Nigeria (Moore et al., 1974, Powers and Logue, 2007, Tomori et al., 1975)
-
•
Central African Republic (Pastorino et al., 2004, Saluzzo et al., 1980)
-
•
Democratic Republic of Congo (Pastorino et al., 2004),
-
•
Ivory Coast, Malawi and Sudan (Watts et al., 1994).
-
•
Senegal (Diallo et al., 1999, Powers and Logue, 2007, Roche and Robin, 1967, Thonnon et al., 1999)
-
•
Zimbabwe (Powers and Logue, 2007)
-
•
Uganda (Powers and Logue, 2007)
- •
The Democratic Republic of Congo declared an epidemic in 1999–2000 (Muyembe-Tamfum et al., 2003). During the period 2004 through 2010, epidemics were identified sequentially in Kenya, Senegal, Sudan (2005), the Cameroon (2006) and Gabon (2007 and 2010). In 2011, an epidemic was reported for the first time in the Republic of Congo (Kelvin, 2011).
Paradoxically, our knowledge of the natural history and environmental cycle of the virus in Africa remains fragmentary. Whilst infection of nonhuman primates (which may act as reservoir or amplifying hosts) has been demonstrated (see section below on pathogenesis), and the virus identified in a variety of sylvatic mosquitoes (see previous sections) the determinants or emergence remain poorly understood. In both Africa and Asia, outbreaks were reported to be unpredictable, with an interval of 7–20 years between individual epidemics (Powers and Logue, 2007) and an interval of 4–5 decades between major epidemics. Whilst the precise factors underlying this pattern are unknown, it is probable that, in addition to a variety of ecological and viral genetic factors and given the long lasting immune protection acquired after chikungunya infection, the immune status of the populations plays a central role in the periodicity of recurrence. This is in coherence with the observation that recurrent outbreaks prefentially target the youngest age-groups (i.e., non immunised during the previous epidemics) (Robert et al., 1996).
4.3.2.2. In Asia
More recent epidemic re-emergence was documented in 2001–2003 in Java (Indonesia), after a 20-year period of its absence (Laras et al., 2005)(see next section). The first ever case of CHIKV in the Hong Kong Special Administrative Region was reported on 24 March 2006 (corresponding to an imported case from Mauritius (ProMED-mail accession number 20060402.0989)). In 2010, an outbreak in Guangdong province (China) of chikungunya fever was reported (Wu et al., 2012). Interestingly, a detailed clinical report by David Bylon in the city of Batavia (current Jakarta, Indonesia) in the year 1779 is highly evocative of chikungunya (see Ng and Hapuarachchi, 2010) and may have been for a long time mistakenly reported as a documentation of dengue fever (Carey, 1971, Halstead, 2009).
Our understanding of the origin of the Asian genotype of CHIKV remains fragmentary. It is probable that the first documented case reported in Bangkok in 1958 reflected more ancient circulation of CHIKV in the region, thus explaining the extent of genetic divergence between the East-Central-South-African (ECSA) and Asian genotypes. According to Ng et al. (Ng and Hapuarachchi, 2010) the availability of historical clinical records suggestive of chikungunya fever outbreaks in Asia since 1779 suggests the independent evolution of an African ancestor of CHIKV in Asia for several centuries, resulting in the Asian genotype. However, this is difficult to accommodate with the most recent estimates of CHIKV evolutionary rates since the most recent ancestor of the Asian and ECSA genotypes is placed at the beginning of the 20th century (Cherian et al., 2009, Volk et al., 2010). Alternately if the report of chikungunya fever in Jakarta in 1779 is exact, it may implicate a limited importation of CHIKV from Africa, the installation of the Asian genotype occurring later (possibly at the end of the 19th century).
Globally, the chikungunya epidemiological situation in Asia remains poorly documented. First, the respective roles of the endemic Asian genotype and of the other genotypes (including the viruses responsible for the Indian Ocean outbreak) have been poorly investigated, despite evidence for co-circulation in some areas (e.g., India, Indonesia… see Fig. 6 ). As noted (Ng and Hapuarachchi, 2010), whilst the ECSA genotype has been taking centre stage in the recent Asian outbreaks, the Asian lineage has not faded into oblivion, with recent cases reported in Malaysia (Kumarasamy et al., 2006), Singapore (Ng and Hapuarachchi, 2010) and Taiwan (Huang et al., 2009) (see also Fig. 6). The long-standing persistence of the Asian genotype, and the recent isolation of an Asian genotype strain from wild macaque monkeys in Malaysia (Apandi et al., 2009) are evocative of a sylvatic maintenance which remains to be further documented (Powers et al., 2000). Secondly, seroepidemiological studies are required to understand in more depth, the level of exposure of the populations and thus to propose a cartography of risk.
Fig. 6.
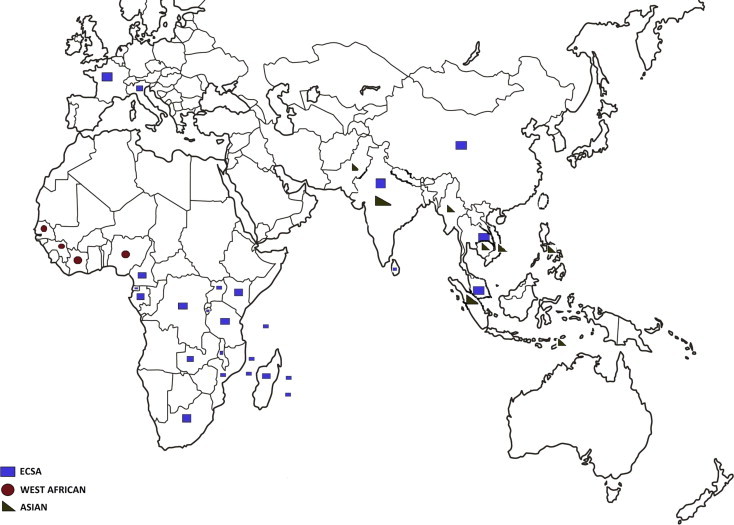
Geographic distribution of CHIKV isolates, according to genotype, as obtained from GenBank and PubMed publications.
4.3.3. Epidemics in the Indian Ocean, India and Southeast Asia
In May 2004, an outbreak of chikungunya fever affected the population on the Kenyan coast. It was first identified on Lamu island where the epidemic peak was reached in July (Renault et al., 2012). It subsequently appeared on the Comoros Islands in December 2004 (Paquet et al., 2006, Schuffenecker et al., 2006) where the epidemic peak was recorded at the end of March 2005, mostly affecting Grande Comore (Ngazidja) island. The epidemic then encompassed the inhabited islands in the entire Southwest Indian Ocean (Mayotte, Reunion Island, Mauritius, Seychelles and Madagascar) (Fig. 5).
Importantly, this outbreak started and spread in areas where the most prevalent vector is Ae. aegypti (i.e., Kenya, Comoros, Seychelles, some areas of Madagascar). Transmission by this mosquito was extremely efficient since a cross-sectional seroprevalence study, performed 9 weeks after the epidemic peak on a representative sample of the Lamu island (Kenya) population, revealed that 75% of the population presented with anti-chikungunya antibodies (Renault et al., 2012, Sergon et al., 2008). Similarly, a cross-sectional seroprevalence study performed on a representative sample of Grande Comore island in March 2005 reported a seroprevalence of 63%, with a considerable social impact (more than one half of infected individuals had to take work leave) (Renault et al., 2012, Sergon et al., 2007).
Little information is available regarding the situation in Mauritius (Renault et al., 2012), where the most prevalent vector is Ae. albopictus. The first detected epidemic wave occurred in March–June 2005, with a second peak in March 2006 (Laras et al., 2005). No case has been reported since August 2006. In the absence of seroprevalence data, the number of cases officially declared (less than 1% of the population) is presumably considerably underestimated.
In the Seychelles archipelago, the first suspected case was reported in July 2005, with a first limited (11 cases) peak in September 2005. The second peak occurred in February 2006 and (probably reflecting the limited impact of the first waves) a third peak occurred in May–June 2007. The total attack rate was estimated to be 12% of the population. Since the end of December 2007, a few suspected cases have been occasionally notified. No retrospective seroprevalence study was conducted.
In Madagascar, epidemiological data are scarce, with initial cases mostly established in travellers returning from the island and clinical epidemiological studies made difficult by the co-circulation of dengue during the same period (Renault et al., 2012). The impact of the 2006 chikungunya outbreak was probably important, and chikungunya fever is suspected to be now endemically present in the island since several cases imported from Madagascar have been identified in Reunion island between 2009 and 2010. It is probable that an outbreak affected the South-east of Madagascar in 2010, which was considered over at the end of March 2010. No seroprevalence data are available.
In Reunion and Mayotte islands, which are French overseas territories, the 2006 chikungunya outbreak was analysed in detail (Charrel et al., 2007, Pialoux et al., 2007, Renault et al., 2012). On Reunion island, where Ae. albopictus is the only epidemiologically relevant vector, the first peak of the outbreak was observed in May 2005 (450 cases). The incidence then decreased and became stable at around 100 cases per week during the austral winter (Renault et al., 2012). A second peak was reached in January–February 2006 (with up to 47,000 cases in a single week in a population of less than 800,000 inhabitants). The last indigenous case was reported in December 2006.
A retrospective seroepidemiological study showed that approximately 40% of the population had been infected, demonstrating the massive impact of the epidemics caused by this dispersing virus (Gerardin et al., 2008b). This is in agreement with prospective surveillance data which reported 266,000 symptomatic infections (attack rate: 34%) and 22 severe presentations requiring assistance for at least one vital function (Renault et al., 2012), of whom 65 died (29%); 25 severe presentations in patients under 15 years of age were detected, of whom 2 died (8%). Forty four cases of mother-to-infant transmission were reported (Economopoulou et al., 2009).
In 2009, a new indigenous cluster was observed in the West of the island (5 cases, (D’Ortenzio et al., 2009)) and in 2010 a second cluster was detected in the same region in March (Saint-Paul, more than 150 cases, first case detected on March 17). Epidemiological investigations established that the probability of unnoticeable virus circulation on the island was unlikely and that the most probable origin of the outbreak was imported cases from Madagascar associated with high vector density. During the outbreak, the house index and Breteau index collected around chikungunya fever cases was much higher than the level of epidemic risk defined by WHO and comparable with levels observed during the 2005–2006 outbreak (D’Ortenzio et al., 2011, Vilain et al., 2012). This result demonstrates that a global level of immunity around 40% was not sufficient to prevent autochthonous transmission of chikungunya fever on Reunion island.
In Mayotte, 63 cases were detected in February–June 2005. A second peak was observed in 2006, with an attack rate estimated at 4% (Renault et al., 2012). No case has been notified since April 2006. In Sri Lanka and the Maldives, the outbreaks appeared in November and December 2006 respectively (WHO, 2007, Yoosuf et al., 2009), followed by explosive epidemics, involving millions of people in at least 13 Indian states, (Kalantri et al., 2006, Lahariya and Pradhan, 2006) where the virus had apparently been virtually absent for the previous 32 years (it is not widely reported that cities in India had experienced massive outbreaks of chikungunya fever during the 1960s and early 1970s). The recent epidemics then spread to Pakistan, Malaysia, Singapore and Thailand (Pulmanausahakul et al., 2011).
4.3.4. Chikungunya fever in Europe and the Americas
4.3.4.1. Imported cases
Imported cases have been reported in France (Cordel et al., 2006, Grandadam et al., 2011, Krastinova et al., 2006), Germany (Frank et al., 2011), Switzerland (Bodenmann and Genton, 2006) and Norway (WHO, 2006) and, based on public health reports, in other countries such as the United Kingdom, Belgium, Spain and the Czech Republic (see Supplementary Tables). Surprisingly, more than 1000 imported cases were reported in 2006 in Western Europe. An attempt at creating risk stratified surveillance zones in Europe indicated that France and Italy are likely to be at greater risk due to the number of visitors they receive from chikungunya active regions, principally viremic visitors from India (Tilston et al., 2009).
Between 2006 and 2011, imported cases were also reported in North America, Canada, French Guyana, Brazil, Guadeloupe, Martinique, New Caledonia and Australia (Bonilauri et al., 2008, Depoortere et al., 2008, Parola et al., 2006). Despite the abundance of Ae. mosquitoes in most of these areas, there was no evidence for significant autochthonous transmission of CHIKV. This may be in part due to the active surveillance of these imported cases, but may also indicate that the spread of the virus in the population was hampered by undetermined entomological or ecological reasons. Nevertheless, a recent study suggests that the vectorial competence of local Ae. albopictus is as efficient as the typical vector Ae. aegypti for CHIKV and DENV (Vega-Rua et al., 2013).
4.3.4.2. Autochthonous cases
Until 2007, CHIKV had never been known to circulate in Europe (MMWR 2006; 55: 1040–1042). In 2007, an outbreak of autochthonous chikungunya fever cases occurred in Italy. This outbreak was initiated by an individual returning to Europe from a visit to India (Angelini et al., 2007). The virus responsible for this outbreak was disseminated in the Emilia Romagna region of northern Italy by Ae. albopictus mosquitoes. More recently, in September 2010, an autochthonous transmission of CHIKV was recorded in southeastern France with two confirmed cases (Gould et al., 2010, Grandadam et al., 2011).
In conclusion, the epidemiological impact of recent chikungunya fever outbreaks has proved to be considerable in regions where the epidemic has spread. High seroconversion rates have been observed (commonly around 35% and up to 75%) and an enormous social and economic burden has been inflicted on affected communities (de Andrade et al., 2010, Enserink, 2007, Krishnamoorthy et al., 2009, Queyriaux et al., 2008). Moreover, despite the limited proportion of cases, outbreaks can have a significant impact on the health services due to the high proportion of symptomatic cases.
4.3.5. Nosocomial transmission of CHIKV
Since chikungunya fever is associated with extremely high and early viremia, the risk of blood-borne transmission of the virus has a high likelihood and interruption of blood supply for transfusion has been reported in affected countries to minimise the risk of transmission via infected blood products (Brouard et al., 2008, Liumbruno et al., 2008, Ng et al., 2009a).
Although direct person-to-person transmission has not been reported, nosocomial transmission most probably can occur following needlestick injury, i.e. when the skin is accidentally punctured by a used (contaminated) needle. However, in the context of an outbreak, the nosocomial nature of transmission is difficult to establish. Interestingly, the only published case of documented nosocomial infection was reported in France, in a ‘cold’ epidemiological context associated with an imported case from Reunion island. It highlighted the risk of transmission for health care workers (Parola et al., 2006). During the CHIKV outbreak in Reunion island, corneas taken from asymptomatic donors proved to be infected with CHIKV and transmission of the virus occurred via this ocular route (Couderc et al., 2012).
5. Phylogenetic analysis
The first focused phylogenetic analysis of CHIKV identified three phylogroups based on the partial E1 viral structural glycoprotein sequence. The CHIKV strains represented in these different geographic lineages apparently circulate in regions that display different ecological backgrounds. The three genotypes are West-African, Asian, and East-Central-South-African (ECSA) (Powers et al., 2000, Schuffenecker et al., 2006).
The geographical distribution of the different genotypes has been described elsewhere (De Lamballerie et al., 2008, Volk et al., 2010) and is summarised in Fig. 6. It is generally believed that the geographical origin of the virus is Africa, and this is supported by several observations (Ng and Hapuarachchi, 2010):
-
1.
the most genetically divergent group of viruses is the West-African genotype;
-
2.
the ECSA and Asian genotypes are paraphyletic in phylogenetic reconstructions (Cherian et al., 2009, Powers et al., 2000, Volk et al., 2010);
-
3.
a complex sylvatic cycle has been known and reported for a longer period of time in Africa than in other regions and a larger number of vertebrate hosts and invertebrate vectors has been identified in Africa, i.e., propitious conditions for creating the observed viral genetic diversity;
-
4.
more ancestral divergence times are estimated for West-African and ECSA genotypes compared with the Asian genotype (Cherian et al., 2009, Volk et al., 2010).
On the basis of recent in-depth phylogenetic analyses of available CHIKV isolates (Fig. 7 ), the evolutionary timescale of CHIKV was estimated to be during the past 300 years (Cherian et al., 2009, Volk et al., 2010). This supports a scenario in which sylvatic African CHIKV emerged as a human pathogen, probably during the 18th century, whereas all the currently identified epidemic genotypes (West-African, Asian, and ECSA) are estimated to have emerged during the 20th century.
Fig. 7.
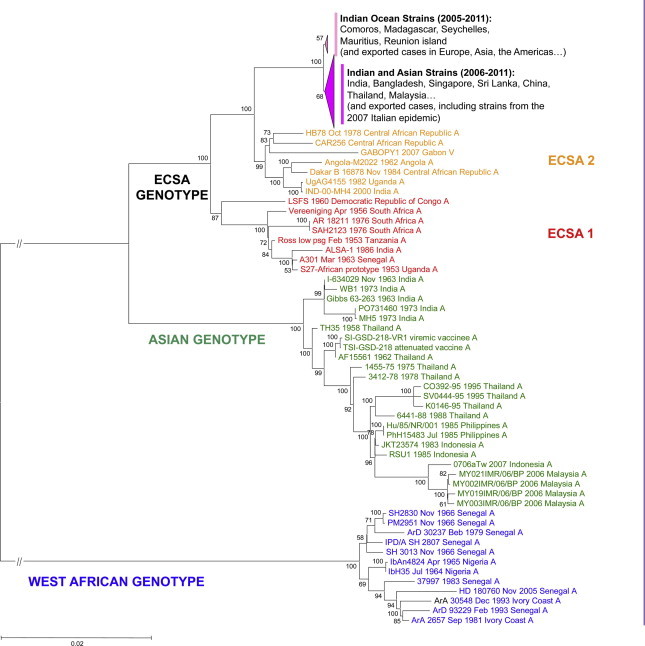
Phylogenetic reconstruction of CHIKV evolution. Phylogenetic trees were produced using Clustal W alignments of complete or nearly complete Chikungunya virus coding nucleotide sequences. Phylogenetic trees were produced using the Neighbour Joining method implemented in MEGA version 5. Bootstrap resampling values are indicated at the main branches. The major evolutionary groups are indicated. Colours identify the different lineages (East-, Central- South-African (ECSA), Asian and West African).
The most important recent event is the spread of the ECSA lineages in the Indian Ocean and Asia. The strains form a monophyletic group within the ECSA lineages, with one branch including mostly Indian strains (and strains subsequently exported in various Asian, European and American countries) and the other strains identified in the South West Indian ocean islands (and strains subsequently exported). The progenitor of the viruses implicated was found to have existed 3 (Lo Presti et al., 2012, Volk et al., 2010) to 9 (Cherian et al., 2009) years before the 2004–2005 and subsequent epidemics arose.
During 2004–2005 chikungunya fever was transmitted to humans via Ae. aegypti (Arankalle et al., 2007) in Kenya, Comoros, Seychelles and some regions of Madagascar (de Lamballerie et al., 2008) but during the outbreak, a genetic change occurred at position 226 of the gene for the membrane fusion glycoprotein E1 with the substitution of a valine residue for an alanine (E1: A226V) (Schuffenecker et al., 2006). There is strong epidemiological and experimental evidence that this is an adaptive mutation that increases the viral replication in Ae. albopictus (Tsetsarkin et al., 2007). This mutation was reported to have occured independently in different geographical locations and in different genetic backgrounds within the ECSA genotype (see the case of Gabonese strains in (de Lamballerie et al., 2008)). It represents an exellent example of convergent evolution (in the amino acid position 226) by different strains of CHIKV (de Lamballerie et al., 2008). However, it must be noted that the original, non-mutated Ae. aegypti-adapted strain was extremely efficiently transmitted in the regions where this mosquito was prevalent, with seroprevalences reaching more than 60% in Grande Comore and Lamu ismands (see previous sections). Accordingly, it has been proposed that the emergence of the epidemic virus was linked with an initial adaptation to urban domestic Ae. aegypti mosquitoes (Ng and Hapuarachchi, 2010), a postulation further supported by compelling evidence of the urban transmission of the ECSA genotype of CHIKV in central Africa during the past decade (Pages et al., 2009).
The A226V adaptative mutation to Ae. albopictus increased the transmission by this mosquito (Tsetsarkin et al., 2009, Tsetsarkin et al., 2007) but marginally compromised the viral fitness in Ae. aegypti (Tsetsarkin et al., 2007, Vazeille et al., 2007). Despite its apparent high frequency, the A226V adaptative evolution did not occur in all CHIKV genetic backgrounds (it was limited to strains belonging to the ECSA genotype) and a complex pattern of other mutations may have been implicated in CHIKV adaptation to a specific Ae. spp. vector (Tsetsarkin et al., 2011a, Tsetsarkin et al., 2009). In particular, the E2-I211T substitution is suspected to have provided the background rendering possible the A226V mutation. It was reported to occur independently on three occasions (South Africa, 1976; Gabon, 2007; Kenya, 2004). However, whilst this mutation is present in the Asian genotype, the A226V mutation has never been observed in this genotype, and experimental mutagenesis failed to create efficient Ae. albopictus-adapted Asian genotype viruses that included this mutation.
Tsetsarkin et al. (2011a) demonstrated that lineage-specific epistatic interactions between substitutions at amino acid positions 226 and 98 of the E1 envelope glycoprotein have restricted the ability of endemic Asian CHIKV strains to adapt to Ae. albopictus. The same authors (Tsetsarkin and Weaver, 2011) identified an additional substitution, E2-L210Q which caused a significant increase in the ability of CHIKV to develop a disseminated infection in Ae. albopictus, but had no effect on CHIKV fitness in Ae. aegypti, or in vertebrate cell lines.
6. Pathogenesis of chikungunya fever
6.1. Target cells
Several recent publications have studied cell susceptibility to CHIKV replication. A panel of immortalised primary human cells was analysed and it was demonstrated that human epithelial and endothelial cells, primary fibroblasts and, to a lesser extent, monocyte-derived macrophages were susceptible to CHIKV infection whereas no replication was identified in lymphoid and monocytoid cell lines, primary lymphocytes and monocytes, or monocyte-derived dendritic cells (Sourisseau et al., 2007a). CHIKV has also been isolated in a tremendous variety of continuous or diploid cell lines, including mammalian cells (Vero, BHK21, HEK-213T, MRC5, BGM, HeLa etc.), amphibian cells (XTC) and mosquito cells (C6/36, Ae, A20 etc.).
In animal models, CHIKV was detected in the cytoplasm of numerous mononuclear cells in the spleen and lymph nodes. Moreover, the sinusoidal endothelium of the liver and macrophages were identified as the main cellular reservoirs during the late stage of CHIKV infection in a macaque model (Labadie et al., 2010). In mouse models, fibroblasts constituted the main target cell of CHIKV in joint, muscle and dermal tissues, whilst in skeletal muscle cells CHIKV was rarely present in satellite cells. No infected leukocytes were detectable in the blood. In the central nervous system choroid plexus epithelial cells and ependymocytes were infected in contrast to microglial cells, astrocytes and microvascular endothelial cells. CHIKV was not found in placental tissue. In addition, the human syncytiotrophoblastic cell line BeWo was found to be refractory to infection (Couderc et al., 2008, Couderc and Lecuit, 2009).
In humans, viral growth in muscle satellite cells was detected whereas myotubes were essentially refractory to infection (Ozden et al., 2007). In biopsies of skeletal muscles, joints and skin, CHIKV antigens were also detected but appeared to be confined to fibroblasts of the joint capsule of skeletal muscle fascia and of the dermis (Couderc et al., 2008, Couderc and Lecuit, 2009). CHIKV was also found (Hoarau et al., 2010a) in perivascular synovial macrophages in one chronic patient 18 months post infection (pi).
6.2. Animal models
6.2.1. Mice
Historically, the mouse was the model of choice to follow susceptibility to arbovirus infection, and was thus more often used for the study of the virus cycle than as a disease model. This is particularly clear for Sindbis virus (SINV), which induces arthritis in humans but was largely used as a model to study virus-induced encephalitis in mice. This highlights the main difficulty associated with the use of mouse models in assessing if they accurately mimic disease in humans both in term of specificity and severity. A recent review was dedicated to this purpose (Teo et al., 2012).
Numerous studies were limited because the majority of adult wild-type (wt) mouse strains were asymptomatic following standard virus inoculation (intraperitoneal, intradermal, intravenous or subcutaneous (sc)). Thus, mainly neonatal mice or interferon (IFN)-α/β receptor knockout mice were used. These models and their derivatives (IFN-α pathway defective mice) (Rudd et al., 2012)) have provided information about the innate immune response against alphaviruses and have been used for drug testing, since mortality rate and organ viral load can be used as convenient indicators of antiviral activity. (Couderc et al., 2009). The use of various mutants of IFN-α pathway defective mice enabled the mechanisms of the innate immune response specific to CHIKV to be deciphered (Couderc et al., 2008, Gardner et al., 2012, Schilte et al., 2012, Schilte et al., 2010, Werneke et al., 2011) implicating roles for Myd88, ISG15, IRF-3 and IRF-7 or viperine (Teng et al., 2012). Recently, a relative role of autophagy and the pathway in the host response against CHIKV was also assessed using Atg16L1HM mice (Joubert et al., 2012).
More recently, A. Suhrbier and collaborators provided a model of chikungunya rheumatic disease in 6-week-old C57BL/6J mice by shifting the site of injection to the dorsal side of the footpad (Gardner et al., 2010). This subcutaneous (sc) injection route provided histological evidence of acute and persistence arthritis, tenosynovitis and myositis at the infected footpad, similar to that observed in RRV infected mice (Rulli et al., 2009, Rulli et al., 2011). The acute phase was characterised by one-week viral replication peaking at 106 CCID50/ml and clear swelling and oedema of the inoculated foot (Gardner et al., 2010, Morrison et al., 2011). Tissue viral replication was detectable from day 1 to 21 in muscle, spleen lymph node and liver. In addition, in this adult mouse model, it was shown that mice initially infected with RRV are partially protected from superinfection with the CHIKV LR-2006 OPY-1 strain inoculated 4 weeks later. In mice, young age remained a factor of severity of the disease as is also observed in human babies but mechanisms of neurological disease are not perfectly compatible with these observations. Following intranasal infection of BALB/C or C57BL/6J mice, direct tissue necrosis is observed in the olfactory lobe but no direct infection of neurones was found (Teo et al., 2012). Finally in a recent study using IRF3/7(−/−) mice, it was demonstrated that inadequate IFN-α/β responses following virus infection can be sufficient to induce haemorrhagic fever and shock, a finding with possible implications for understanding severe CHIKV disease (Rudd et al., 2012).
Altogether, there is no current mouse model able to depict the long-lasting chronic arthralgic features found in up to 60% of the CHIKV-infected patient (Schilte et al., 2012 submitted) even if gene profiling of CHIKV arthritis in a mouse model reveals significant overlap with rheumatoid arthritis (Nakaya et al., 2012). However, mouse models are convenient and remain of specific interest for the in vivo screening of drugs targeting the acute phase of infection.
6.2.2. Nonhuman primates
NHP have been used extensively for studies of virus-induced pathology and evaluation of drugs and vaccines in a number of diseases (Vierboom et al., 2007, Walsh et al., 1996). Very early in the history of CHIKV studies it was found that NHP are susceptible to infection and are probably part of the natural reservoir in Africa and Asia (Inoue et al., 2003, Marchette et al., 1978, McCrae et al., 1971, Peiris et al., 1993). In field epidemiological studies in Central Africa (Central African Republic, Gabon) Cercopithecus and baboon monkeys have been used as sentinels to assess the presence of arboviruses in the wild (Saluzzo et al., 1980, Saluzzo et al., 1981, Saluzzo et al., 1982). African green monkeys and macaques were also used as sentinels for estimation of vector efficacy of different mosquito species (Jupp et al., 1981, Levitt et al., 1986, Turell and Beaman, 1992, Turell and Malinoski, 1992). These studies led to the identification of the major relevance of Ae. spp. in epidemic outbreaks.
The first experimental CHIKV infection was achieved in rhesus (Macaca mulata) and bonnet macaques (M radiata) in 1967, but only M mulata showed episodes of fever within a few days of virus inoculation (Binn et al., 1967, Paul and Singh, 1968). The viral replication pattern in macaques was very similar to that reported in humans in 1965 with the same Asian strains, with plasma viremia found to be positive in macaques from days 1 to 6 and viral titres ranging from 3 to 5.3 log10 pfu/ml (Binn et al., 1967, Paul and Singh, 1968).
In these early experiments, little was recorded concerning the clinical signs associated with infection perhaps due to technical limitations. Amazingly, Paul and Sing in the 1968 study noted that: “the rate of infection in Ae. albopictus was consistently higher than that of Ae. aegypti when both species of mosquitoes were fed simultaneously on the same (viremic) macaque”. Despite this premonitory observation, Ae. albopictus was regarded as a minor potential vector of CHIKV, until the 2005–2006 Indian Ocean outbreak and its spread to India and South-east Asia. Subsequently, wild macaque susceptibility to infection was reported following epidemiological studies (Inoue et al., 2003, Peiris et al., 1993).
In the early 1990s, studies of CHIKV infection in NHP focused on macaque to mosquito transmission efficacy (Turell and Beaman, 1992, Turell and Malinoski, 1992). Recently, Roques and collaborators have infected cynomolgus macaques (Macaca fascicularis) using Reunion island strains of CHIKV and, by following biomarkers of CHIKV, found that replication was detectable during the first week of infection with a level of replication similar to human infection (Labadie et al., 2010). Performing in vivo titrations they demonstrated that as little as 10 pfu given IV, could produce infection in macaques, with viremic levels up to 108 pfu/ml being detected. At day 4 postinfection (pi) CHIKV is detected in the cerebrospinal fluid of all tested macaques but clinical neurological disease is detected only in macaques receiving the highest infectious doses. Interestingly, the acute infection seemed to be tightly controlled by poorly-characterised antiviral mechanisms given that the viral titre was reduced to basal levels at day 10 postinfection as described in humans or mice (Ziegler et al., 2008). These viral replication profiles were also recorded in rhesus macaques (Akahata et al., 2010, Chen et al., 2010).
In humans, early leukopenia was observed (Akahata et al., 2010, Borgherini et al., 2009, Borgherini et al., 2007, Labadie et al., 2010) together with markers of antiviral response (IFN-α/β) inflammation and cell immune activation (Higgs and Ziegler, 2010, Labadie et al., 2010). Whilst virus has been found in cerebrospinal fluid samples during the acute phase of infection, viral presence in the central nervous system is not clearly related to neuronal disease. It is more likely associated with non-specific inflammation that in rare cases might be associated with encephalopathy in animals infected with a very large amount of virus, as was probably the case for neonatal patients (Gerardin, 2010, Gerardin et al., 2008a, Labadie et al., 2010).
Recent studies in cynomolgus macaques demonstrated that CHIKV persists in target tissues after its clearance from the blood, as demonstrated by viral RNA detection using in situ hybridization assays (Labadie et al., 2010). At day 7 or 9, CHIKV is detectable in nearly every organ or compartment tested. Joints, secondary lymphoid organs and, to a lesser extent, muscles are affected and virus genetic material as well as isolation of replicative CHIKV is obtained up to 3 months pi. CHIKV replicates in several cell types during the acute phase (Higgs and Ziegler, 2010, Labadie et al., 2010) but is thereafter mainly detectable in macrophages, by immunohistochemical analysis (double staining). Infected monocytes and macrophages could be detected in the blood 6 h after infection (Roques et al., 2011) and in most tissues on the following day (in situ hybridization, immunohistochemistry, RT-PCR and virus isolation). Significant macrophage infiltration was also detected by histological analysis throughout the study and long after viral clearance from blood (Labadie et al., 2010).
This monocyte-macrophage tropism is consistent with the infection of human monocytes in vitro. These infected monocytes generate new viruses, which can be detected using HEK293 cells and titration assays, albeit at low levels (Her et al., 2010). Similarly, CHIKV can infect primary macrophages in vitro (Rinaldo et al., 1975, Sourisseau et al., 2007b) resulting in the production of highly variable amounts of virus, from 103 to 106 pfu/ml, regardless of the species being used for the studies (human, macaque, mouse) (Gardner et al., 2010, Labadie et al., 2010, Sourisseau et al., 2007b) (Fig. 8 ). These results are consistent with human studies, which reported that macrophages are susceptible to CHIKV infection both in vivo and in vitro (Hoarau et al., 2010b, Krejbich-Trotot et al., 2011). The alleviation of chikungunya-associated arthritis and myositis by treatment, with Bindarit, a MCP-1/CCL-2 inhibitor, in mice (Rulli et al., 2011) also strongly suggests that monocytes/macrophages, the main targets in MCP-1/CCL-2 tissue tropism, are central to muscle and joint disease (see below on the treatment section).
Fig. 8.
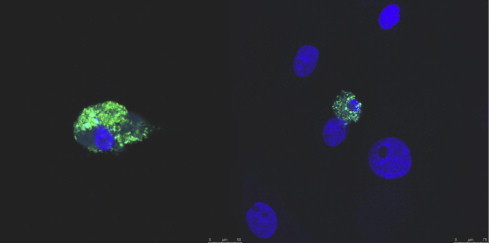
Accumulation of CHIKV proteins in macaque monocyte-derived macrophages 21 days post-infection (Labadie et al., 2010). Monocyte-derived macrophages were separated from monocytes by adhesion for 7 days, then infected with 0.5 MOI of CHIKV expressing nsp3-ZsGreen protein. Macrophages growing on glass slides were fixed and conterstained with DAPI. Pictures were acquired using a Confocal SPE Leica microscope, (×40).
There is currently no animal model fully reproducing the chronic rheumatoid syndrome of chikungunya fever. Indeed, the disease picture in mice is mainly driven by destruction of tissues with huge cell infiltration (Gardner et al., 2010, Morrison et al., 2011, Rulli et al., 2011) and despite virus persistence, severe joint damage was not observed in macaques (Chen et al., 2010, Labadie et al., 2010). Nevertheless, both models suggest that inflammation, macrophage tissue tropism, and local viral persistence are involved in the establishment of chronic disease.
7. Vaccines
No vaccine against chikungunya is currently available. The first attempts to develop an inactivated vaccine were reported at the end of the 1960s and involved detergent treatment of virus (White et al., 1972). Unfortunately, these formalin-killed vaccines which showed promise appeared to be moderately immunogenic and limited to short time effects in a clinical trial (Harrison et al., 1971). As live attenuated vaccines are often more effective and, particularly for arboviruses, live vaccines are better at inducing rapid protection against homologous and heterologous challenge, the US Army developed a live, attenuated vaccine (TSI-GSD-218), based on an Asian 1970s strain, by serial passage in MRC-5 cells resulting in a virus (CHIK 181/Clone 25), which carried significant genetic changes (Hoke et al., 2012, Levitt et al., 1986). This showed promise in protecting NHP against challenge and was later tested in clinical trials showing good immunogenicity (Edelman et al., 2000, McClain et al., 1998). However, several adverse events were associated with TSI-GSD-218, the most severe being arthralgia in 8% of vaccinees.
Because of loss of information about the vaccine production, TSI-GSD-218 is no-longer considered suitable for clinical use, but was used in proof of concept studies to evaluate the potential mechanisms of protection and resistance to the virus induced disease. Thus (Partidos et al. (2011) probed the attenuation and protective efficacy of this attenuated vaccine strain in mice with compromised IFN signalling. They showed that infection of AG129 mice (defective in IFN-α/β and IFN-γ receptor signalling) with CHIK 181/Clone 25 virus resulted in rapid mortality within 3–4 days. In contrast, all infected A129 mice (defective in IFN-α/β receptor signalling) survived with temporary morbidity characterised by ruffled appearance and body weight loss. A129 heterozygote mice that retain partial IFN-α/β receptor signalling activity remained healthy. Infection of A129 mice with CHIK 181/Clone 25 virus induced significant levels of IFN-γ and IL-12 whilst the inflammatory cytokines, IFN-α and IL-6 remained low. A single administration of the CHIK 181/Clone 25 vaccine strain provided both short-term and long-term protection (38 days and 247 days post-prime, respectively) against challenge with wt CHIKV-La Reunion (CHIKV-LR). This protection was at least partially mediated by antibodies since passively transferred immune serum protected both A129 and AG129 mice from wt CHIKV-LR and CHIK 181/Clone 25 virus challenge.
These data were in accordance with others studies showing that IFN-α/β deficiency greatly exacerbates arthritogenic disease in mice infected with wt CHIKV (Couderc et al., 2008, Gardner et al., 2010), but not with the cell culture-adapted live-attenuated CHIK 181/Clone 25 vaccine candidate (Gorchakov et al., 2012). The Weaver group showed that attenuation of CHIK 181/Clone 25 vaccine strain is determined by two amino acid substitutions in the E2 envelope glycoprotein tested in two different murine strains: infant CD1 (5–6 days) and adult A129 mice (both defective in IFN pathway) (Gorchakov et al., 2012). Overall, these data highlight the importance of IFNs in controlling the CHIK 181/Clone 25 vaccine strain, and demonstrate the ability of this vaccine to elicit neutralising antibody responses that confer short- and long-term protection against wt CHIKV-LR challenge.
7.1. Single recombinant antigens or Vero cell-adapted, formalin-inactivated vaccine
A vaccine based on recombinant envelope proteins of an Indian strain of CHIKV, adjuvanted with Alum, FCA or Montanide, elicited a balanced Th1/Th2 response with virus-neutralising antibodies in BALB/C mice, but the authors did not further challenge these mice (Khan et al., 2012). Kumar et al. (2012) evaluated recombinant E2 protein-based and whole-virus inactivated candidate vaccines, given intramuscularly, against CHIKV in BALB/c mice. They tested 4 adjuvants: Alum, Mw, CadB (rE2p), Alum/Mw (formalin-inactivated CHIKV) and Alum (BPL-inactivated CHIKV). Humoral immunity was assessed by ELISA and in vitro neutralisation test, using homologous and heterologous (Asian genotype) strains of CHIKV.
Two cohorts of vaccinated mice were challenged separately via the intranasal route with homologous virus 2 and 20 weeks after the second dose. Anti-CHIKV-antibody titres were dose dependent for all the immunogenic formulations. BPL-inactivated vaccines led to the highest ELISA/neutralising antibody (nAb) titres, whilst alum was the most effective adjuvant. In this adult BALB/c mouse/intra-nasal infection model (6–8 weeks), complete protection was observed following immunisation with the alum-adjuvanted rE2p, and both the inactivated vaccines, as no virus was detected in the tissues (muscle, brain, spleen) and blood (real time PCR) after challenge 2 weeks or 20 weeks-post-the second vaccine dose. However, with rE2p–CadB, very low viremia was recorded on the second day-post-challenge. Globally, a majority of vaccination trials in mice, even with the very simple inactivation system of whole CHIKV particles inactivated with aziridine (Brown, 2001), gave effective protection with as low as 0.01 μg of vaccine in C57BL6/adult mice (12 weeks of age) with suppression of viremia and leg oedema.
7.2. Structural components of CHIKV in non-infectious virus-like particles (VLPs)
VLPs representing CHIKV envelope protein were produced in 293A renal epithelial cells and 293T human kidney cell lines infected by lentivirus vectors containing the subgenomic RNA sequence (C–E3–E2–6k–E1) (Akahata et al., 2010). The authors showed that, using VLP plus Ribi adjuvant, an antibody response 20 times higher than when using a DNA vaccine with the same sequence in Balb/C mice could be detected. Finally they reported protection in both mice (Ifnar −/−) and non-human primates challenged with the LR-2006 OPY-1 strain of CHIKV. However, the amount of produced VLPs remained very low in this system: from 0.1 to 20 mg/l within the 293T supernatant medium. The method was improved recently when the authors showed that a specific domain of the CHIKV E2 protein (amino-acid 233N–234K–252Q) regulated particle formation in human cells (Akahata and Nabel, 2012) and thus production of functional VLPs might be enhanced up to 200 fold.
A group from Wageningen University developed production of VLPs using chimeric baculovirus with the salmonid alphavirus (SAV) in insect cells and obtained functional processing and secretion (Metz et al., 2011). They recently developed such a process to obtain CHIKV VLPs within chimeric baculovirus. The first vaccination assays and virus challenge experiments were performed using the A Suhrbier mouse model system (Metz and Pijlman, 2011, Pijlman, 2012).
7.3. Vaccination with chimeric alphaviruses
Chimeric alphaviruses have been developed containing Venezuelan equine encephalitis virus (VEEV) or equine encephalitis virus (EEV) nonstructural protein coding sequences and CHIKV structural protein coding sequence with a programmed, attenuated, cell-type-restricted phenotype because of the lack of Old World alphavirus nsP2 or New World alphavirus capsid involved in innate cell response shutdown (Kim et al., 2011). To make these viruses incapable of transmission by mosquito vectors and to differentially regulate the expression of viral structural proteins, their replication was made dependent on the internal ribosome entry sites (IRES), derived from the encephalomyocarditis virus IRES (Plante et al., 2011).
The design of the genomes was complemented by selection procedures, which adapted viruses to replication in tissue culture and produced variants which (i) demonstrated different levels of replication and production of the individual structural proteins, (ii) efficiently induced the antiviral response in infected cells, (iii) could not replicate in mosquito cells (Darwin et al., 2011), and (iv) efficiently replicated in Vero cells. Finally, this vaccine candidate was tested in highly sensitive A129 mice (IFN-α/β R−/−) and was found not deleterious in contrast to CHIK 181/Clone 25 vaccine strain but efficiently induced a protective immune response (Wang et al., 2011b) and was able to provide cross-protection for alphaviruses of the Semliki Forest antigenic complex (Partidos et al., 2012).
Using the same procedure and CHIKV vaccine candidate, Partidos et al. induced a cross-protective immunity in adult AG129 mice (IFN-α/β R−/−; IFNγ R−/−) against ONNV 9 weeks post vaccine (Partidos et al., 2012). The authors also showed that transplacental passive anti-CHIKV antibody transfer to 3 week old A129 offspring from the vaccinated dam was protective against a ONNV intra-dermal challenge.
Using a completely different strategy, GenPhar developed a recombinant adenovirus CadVax-CHIKV containing codon optimised sequence containing the CHIKV structural genes (C–E3–E2–6k–E1). This vaccine was tested using the A Suhrbier mouse model (Wang et al., 2011a). At 5 weeks post-vaccination, the reciprocal of the neutralisation titre (RNT) was close to 5 × 103 which was similar to the titre observed after infection with the wild type CHIKV (100% protection against cytopathic effect of 200 CCID50 of virus). The mice were challenged 45 days after vaccination and were shown to be fully protected against both foot swelling and viral replication (undetectable compared to 5 × 106 CCID50 in control group).
Other recombinant viruses tested in mice include chimeric vesiculo/alphavirus, in which the entire CHIKV envelope polyprotein was expressed (E3–E2–6k–E1) (Chattopadhyay et al., 2013). These VSV-G-CHIKV chimeras were attenuated in tissue culture, propagated to high titre without VSV-G complementation and shown to protect adult mice C57BL/6 challenged 34 days postvaccination by the sc route of inoculation in the left rear foot-pad (Morrison model). Efficiency of vaccination was assessed for neutralising antibodies using the plaque reduction neutralisation test PRNT80 160 to >640 and IFN-γ ELISPOT assay against E1 and E2 peptides (400–600 ± 200 SFC/106 cells). However, in this study the viral replication and disease induced by the CHIKV in non-vaccinated mice was not very high (2.85 ± 0.47 log10 pfu/ml at day 2), i.e., 2–3 orders of magnitude below the value obtained in two others studies (Gardner et al., 2010, Teng et al., 2012), perhaps because of a shorter delay between vaccination and challenge.
7.4. Electroporated DNA
A plasmid containing codon and RNA-structure-optimised E3–E2–E1 protein sequences without the 6k peptide but including the furin cleavage sites as well as a Kozak-IgE leader sequence at the 5′ end of the genome, was generated by GeneArt (Regensburg, Germany) and vaccination was performed by 3 subsequent electroporations into quadriceps from 8-week-old female BALB/c mice (Mallilankaraman et al., 2011). One week after the third electroporation CHIKV neutralising antibodies were significantly enhanced with a RNT median of 320 (which produced 100% protection against cytopathic effect using a challenge dose of 100 CCID50 of virus) and a high CHIKV specific cellular response was demonstrated as measured by IFN-γ ELISPOT assay against E1 and E2 peptide pools (1613 ± 170 SFC/106 cells). This construction thus induced an improved immune response, when compared with the previous C–E1–E2 contruction from the same group (Muthumani et al., 2008). At this time mice were challenged by intranasal exposure with 107 pfu (25 μl) of the CHIKV-PC-08 strain (ECSA genotype).
In summary, the mice were protected from the high morbidity normally associated with intranasal infection (i.e., spongiform lesion apoptotic bodies, microglial nodules in the external granular layer of the cerebral cortex; severe myocardial and liver degeneration/necrosis) which required euthanasia of control mice at day 6–12 pi. Despite marked weight loss during the first 3 days, vaccinated mice returned to normal values thereafter. However, in the vaccinated animals sacrified at day 5 pi, some microglial oedema was seen in the brains suggesting that immunisation decreased CHIKV-mediated pathology but did not fully protect the brain in the intranasal inoculation model. In contrast, the disease in mice vaccinated with DNA expressing only the CHIKV capsid induced a morbidity picture identical to the control non-immunised mice. Finally Mallilankaraman et al., 2011 tested the immune response induced following this vaccination strategy in four rhesus macaques (5 immunizations). Two weeks later, they detected variable neutralising antibody responses (RNT 80–1200) and specific T cell responses (200–500 SFC/106 PBMCs), but they did not challenge the animals. Thus, all candidate vaccines except one (TSI-GSD-218) have so far only been tested in mice, and the studies have mainly focused on neutralising antibodies and not on cellular responses. From the literature, it is unclear whether any of these vaccines have proceeded into human clinical trials.
In addition, recent studies have shown that (1) the recognised protective immune response against CHIKV consists predominantly of IgG3 antibodies and (2) the early neutralising IgG response to CHIKV in infected patients targets a dominant linear epitope on the E2 glycoprotein (Kam et al., 2012a, Kam et al., 2012b). However, another study reported that CHIKV is able to escape the impact of neutralising antibodies by direct cell-to-cell transmission (Lee et al., 2011). This observation implies that an effective vaccine cannot be assessed purely by its ability to induce detectable neutralising antibodies.
In conclusion, whilst it is evident that an effective vaccine against CHIKV infection is clearly needed, inactivated vaccines are unlikely to provide strong and long-lasting immunity, and they will almost inevitably require multiple doses, thus reducing their cost effectiveness. DNA vaccines, thus far, have proved less effective in humans than in other mammals. Live, attenuated alphaviruses are being used as the genetic backbone for vaccines under development by Chiron, Alphavax and others and this approach is the most likely to lead to an effective vaccine with long-lasting immunity for CHIKV. However, it remains to be determined whether the inevitable high development costs of a safe and effective live, attenuated chimaeric vaccine against CHIKV will prove to be a restraining influence.
7.5. Attenuation of CHIKV by large-scale codon re-encoding
This novel method of attenuating viruses (Burns et al., 2006, Coleman et al., 2008, Mueller et al., 2010, Mueller et al., 2006, Song et al., 2012) was recently applied to the LR2006 CHIKV strain (ECSA genotype): the nucleic acid composition of large coding regions was modified, without modifying the encoded proteins, by introducing a large number of synonymous mutations (Nougairede et al., 2013). Using reverse genetic methods, synonymous mutations were randomly introduced in three regions of around 1.4 kb that encode structural (nsP1 and nsP4) and non-structural (E1–E2) proteins. Up to 882 synonymous mutations were introduced representing, 7.5% of the complete genome of the virus.
The replicative fitness of the resulting re-encoded viruses was studied in cellulo using primate and mosquito cells. Introducing a large number of presumably slightly deleterious synonymous mutations significantly decreased the replicative fitness of CHIKV in both primate and arthropod cells and this diminution correlated directly with the degree of re-encoding giving the possibility to modulate precisely the degree of fitness loss. The use of several re-encoded regions located throughout the viral genome reduces the likelihood of complete phenotypic reversion due to recombination between wild-type and re-encoded CHIKVs. In addition, the stability of these re-encoded viruses was assessed by serial passaging of these viruses 50 times either in primate or insect cells or in each cell line alternately. The analysis of these passaged viruses showed that it exhibited a stable phenotype, and that the response to codon re-encoding was largely compensatory in nature, with little reversion of mutations.
Studies in animal models are obviously needed to evaluate the potential of these re-encoded CHIKVs for producing vaccine candidates. The important message is that generating an attenuated strain of CHIKV by large-scale re-encoding represents a potentially important route to vaccine development, because this method does not modify amino acid sequences and therefore potentially alleviates the likelihood of novel phenotypic properties, and also provides the advantages of all the live attenuated vaccines, including reduced costs and single dose induction of long-term immunity.
8. Antiviral therapy
Several drugs are known to be effective against CHIKV when tested in vitro, but no recognised antiviral treatment is currently available.
8.1. Experimental therapies targeting steps in viral replication
8.1.1. Antibody-based therapies
Passive immunotherapy has been used for over a century in the treatment of viral infectious disease. In a recent study, monkeys were immunised with a vaccine based on CHIKV VLPs. The immunoglobulin G (IgG) extracted from the sera of these monkeys was administered passively to mice, which were then infected with CHIKV. The immune IgG protected the mice from infection by CHIKV (Akahata et al., 2010). Human immunoglobulins purified from plasma of CHIKV convalescent patients also exhibited a high in vitro neutralising activity and a powerful prophylactic and therapeutic efficacy against CHIKV infection in mice (Couderc et al., 2009).
More recently, human recombinant monoclonal antibodies exhibited strong and specific neutralising activity in vitro when tested against a wide variety of CHIKV strains and were able to significantly delay the CHIKV-driven lethality in AGR129 mice (IFN-α/β/γ R−/−/− and RAG-2–) (Fric et al., 2013, Warter et al., 2011). Passive immunotherapy may constitute an efficacious prevention strategy and treatment for individuals exposed to CHIKV who are at risk of severe infection, such as neonates born to viremic mothers and adults with chronic underlying medical conditions (Couderc et al., 2009).
8.1.2. Interferon
The IFN system is capable of inhibiting virus infections in the absence of adaptive immunity. Treatment of cells with IFN-α/β upregulates the expression of several hundred genes which, in combination, specify the antiviral state. No single gene is pivotal and, for any given virus, a subset of genes is probably required to limit viral replication. Several of the upregulated genes encode enzymes that have been studied intensively, such as dsRNA-dependent protein kinase R (PKR), 2′,5′-oligoadenylate synthetase (OAS) and MxA (Randall and Goodbourn, 2008). The most important current application of IFN therapy as an antiviral treatment is for the chronic phase of viral infections, such as HCV and HBV (Heim, 2012). Nevertheless, IFN therapy has also been proposed for acute viral infection such as SARS or influenza (Wang and Fish, 2012).
IFN is known to inhibit the growth of other viruses within the genus Alphavirus (Lukaszewski and Brooks, 2000, Pinto et al., 1990). In the case of SINV, it was found that the balance between type I IFN induction and the ability of the virus to develop further rounds of infection is determined in the first few hours of virus replication, when only low numbers of cells and infectious virus are involved (Frolov et al., 2012). In another study, an emerging alphavirus in salmonid aquaculture called Salmonid alphavirus-3 (SAV-3), was sensitive to the preinfection antiviral state induced by IFN-α, whilst pi antiviral responses or pi treatment with IFN-α was not able to limit viral replication (Xu et al., 2010). This is consistent with recent studies in mice, in which it was found that IFN-α treatment prevented arthritis due to CHIKV only if given before infection. Since such a situation would not exist in most cases of human infection, IFN-α is not likely to prove useful for therapy (Gardner et al., 2010).
8.1.3. Small-molecule drugs
8.1.3.1. Ribavirin
Ribavirin, the nucleoside analogue 1-β-D-ribofuranosyl-1,2,4-triazole-3-carboxamide, exhibits antiviral activity against a variety of RNA viruses in cell culture through at least three distinct mechanisms: inhibition of the cellular protein inosine monophosphate dehydrogenase (IMPDH), immunomodulatory effects, and incorporation as a mutagenic nucleoside by the viral RNA polymerase. All three activities may play an antiviral role in vivo (Crotty et al., 2002).
Ribavirin is known to inhibit in cellulo a large range of RNA viruses and notably viruses in the genus Alphavirus, such as Semliki forest virus (SFV) and CHIKV (Ho et al., 2010, Smee et al., 1988). In animal models, ribavirin is effective against a more limited set of viruses, mainly RNA viruses. In humans, ribavirin is currently used in combination with interferon-α to treat hepatitis C virus (HCV) infections and it has been used as monotherapy for Lassa fever and severe respiratory syncytial virus (RSV) infections (Crotty et al., 2002).
Some in vivo antiviral activity of ribavirin during chikungunya fever has been reported recently in a non-randomised human cohort, but no statistical analysis was available due to the small number of participants (Ravichandran and Manian, 2008). The combination of interferon and ribavirin has been found to exert a synergistic antiviral effect against two alphaviruses (CHIKV and SFV) in vitro (Briolant et al., 2004).
8.1.3.2. Chloroquine
Chloroquine and its hydroxy-analogue, hydroxychloroquine are weak bases that are known to affect acid vesicles leading to dysfunction of several enzymes. Some viruses enter their target cells by endocytosis in the lysosomal compartment, where the low pH, along with the action of enzymes, disrupts the viral particle, thus liberating the infectious nucleic acid and, in several cases, enzymes necessary for viral replication. For some enveloped viruses, post-translational modification of the envelope glycoproteins also occurs within the endoplasmic and trans-Golgi network vesicles, and involves proteases and glycosyl-transferases, some of which require a low pH. Thus it was shown that chloroquine/hydroxychloroquine can impair the replication of several viruses by interacting with the endosome-mediated viral entry or the late stages of replication of enveloped viruses (Savarino et al., 2003).
The potential for in cellulo application of chloroquine as an antiviral treatment was first reported more than 40 years ago (Inglot, 1969), but in the case of alphaviruses, and notably SFV, it was also reported that chloroquine enhanced virus replication in mice (Maheshwari et al., 1991). In vitro inhibition of CHIKV replication by chloroquine has been reported (Delogu and de Lamballerie, 2011) and should be distinguished from the anti-inflammatory effect previously reported in patients at the chronic stage of infection (Brighton, 1984). In 2006, a double-blind, placebo-controlled randomised trial evaluating the efficacy of chloroquine against chikungunya fever did not identify a significant difference between the chloroquine and placebo groups, in terms of either the mean duration of febrile arthralgia or the decrease of viremia from day 1 to 3. Moreover, at day 200 postinfection, patients treated with chloroquine declared more frequently that they still suffered from arthralgia than patients who had received a placebo (De Lamballerie et al., 2008).
In conclusion, whilst in vitro inhibition of CHIKV replication is strongly established, the narrow therapeutic index and the absence of obvious biological or clinical improvement during a clinical trial do not argue in favour of a curative use for chloroquine in noncomplicated cases of chikungunya fever (Delogu and de Lamballerie, 2011). This suggests that previous similar favourable results obtained in cellulo only with chloroquine for highly pathogenic viruses, such as the SARS-CoV (Keyaerts et al., 2004), should be considered carefully.
8.1.3.3. Arbidol
The antiviral drug Arbidol (ARB) (1-methyl-2-phenyl-thiomethyl-3-carbotoxy-4-dimetylaminomethyl-5-hydroxy-6-bromoindolehydrochloride monohydrate) was developed by the Centre for Drug Chemistry, Moscow for use against respiratory viral infections (Delogu et al., 2011). ARB has been reported to be clinically active against influenza A and B viruses, respiratory syncytial virus (RSV) and other respiratory viruses and more recently against HCV (Brooks et al., 2012). In addition to possible immune-modulatory effects, the antiviral activity of ARB could in part be due to its membranotropism. Recent data showed that ARB incorporates into cellular membranes leading to perturbed membrane structures and inhibition of virus mediated fusion. Indeed, it has been shown that ARB was an entry inhibitor of influenza virus infection by stabilizing the influenza HA and preventing the endosomal membrane fusion. Similarly, ARB inhibited the HCV glycoprotein conformational changes needed for the membrane fusion process. A recent study found that ARB was also effective against CHIKV in vitro and it was concluded that ARB interferes with the early stages of CHIKV infection (virus attachment or entry) by targeting the cellular membrane (Delogu et al., 2011).
8.1.3.4. Furin inhibitors
Alphavirus envelope glycoproteins are initially produced first as precursors (E3E2 or p62) and during virion maturation further cleaved at short multibasic motifs. Amongst cellular proteases, the basic amino acid-specific furin or furin-like proprotein convertases (PCs) have been shown to be involved in such a calcium-dependent processing, resulting in the cleavage of surface glycoproteins (Ozden et al., 2008). Since it was demonstrated that uncleaved glycoprotein precursors induce defects in viral production for other alphaviruses such as SINV and SFV, a recent study tested the hypothesis that an irreversible furin-inhibiting peptide (decanoyl-RVKR-chloromethyl ketone (dec-RVKR-cmk)) could inhibit CHIKV infection (Ozden et al., 2008). Like other alphaviruses, the envelope glycoproteins of CHIKV are initially produced as precursors that are further cleaved by furin-like proprotein convertases (PCs) (Voss et al., 2010). dec-RVKR-cmk induced in cellulo stronger inhibition of viral infection than chloroquine when added immediately following infection. A combination of both drugs induced an additive effect, which was further enhanced when the drugs were added to the cells before the virus (Ozden et al., 2008).
8.1.3.5. Structure-based-specific inhibitors
Crystal structures of the replicative proteins of CHIKV and several other alphaviruses have been reported (Lampio et al., 2000, Malet et al., 2009, Russo et al., 2006, Voss et al., 2010) and provide new opportunities to develop structure-based, specific inhibitors. Using these principles, Singh et al. (2012) identified four lead compounds in a large library of compounds that potentially could inhibit the nsP2 protease of CHIKV, based on a homology model of the crystal structure of the nsP2 protein of VEEV. More recently, Lucas-Hourani et al. (2012) identified, amongst 3040 molecules, one natural compound (ID1452-2) that inhibits CHIKV replication in vitro by interfering with nsP2 activity.
8.1.3.6. Other inhibitors of virus replication
A series of complex polycyclic molecules isolated from a rare native plant of New Caledonia (Trigonostemon cherrieri), Trigocherrins 1,2 and 6 and Trigocherriolides 7–9, have recently been tested in cellular assays against CHIKV, SINV and SFV. Comparisons of selectivity indices led to the conclusion that currently Trigocherrins are the most potent identified inhibitors of CHIKV replication. The mechanism of inhibition has not yet been determined (Allard et al., 2012a, Allard et al., 2012b). More recently, another species of the same genus (Trigonostemon howii) collected in central Vietnam has also proved to be a potent inhibitor of CHIKV (Bourjot et al., 2012a).
Similarly, products extracted from the leaves of the Madagascan plant Anacolosa pervilleana, demonstrated moderate activity in vitro against CHIKV replication (Bourjot et al., 2012b, Bourjot et al., 2012c). More recently, Kaur et al. (2013) identified amongst a highly purified natural product compound library, a cephalotaxine alkaloid called harringtonine that displayed potent inhibition of CHIKV infection affecting CHIKV RNA production as well as viral protein expression with minimal cytotoxicity.
8.1.4. Antisense oligonucleotides and siRNA
Peptide-conjugated phosphorodiamidate morpholino oligomers (PPMO) are nuclease resistant and water-soluble single-stranded-DNA-analogues that can enter cells readily and act as steric-blocking antisense agents through stable duplex formation with complementary RNA (Stein, 2008). When applied in cellulo, PPMO decreased viral titres by several orders of magnitude, and reduced viral replication and/or increased survival of mice experimentally infected with poliovirus, coxsackievirus B3, dengue virus, West Nile virus, respiratory syncytial virus, Ebola virus, influenza A virus and the alphavirus VEEV (Paessler et al., 2008, Stein, 2008).
Small interfering RNA (siRNA) is an ancient, highly conserved and natural mechanism that is induced by double-stranded RNA. It defends cellular genomes against infection by viruses and transposons. SiRNA specifically degrades RNA in the cytoplasm of eukaryotic cells (Hannon, 2002). It has been successfully utilised to inhibit replication of many RNA and DNA viruses (influenza, human immunodeficiency virus, Japanese encephalitis virus, West Nile virus, foot and mouth disease virus, hepatitis B virus, and herpes simplex virus) and notably alphaviruses such as VEEV and SFV (Dash et al., 2008).
In 2008, Dash et al. (2008) designed two siRNAs that bind to conserved regions of the CHIKV genome (nsP3 and E1). When added to cells up to 72 h postinfection, these siRNAs produced significant viral inhibition, reducing both the infectious virus titre and the total viral load. In another study, siRNA was used to inhibit the autophagy machinery in CHIKV-infected cells (siRNA against the transcript of Beclin 1, an autophagic protein). This procedure substantially decreased CHIKV replication (Krejbich-Trotot et al., 2011). More recently, plasmid-based small hairpin RNA (shRNA) against CHIKV E1 genes has provided significant inhibition of infectious virus production in vitro. Strong and sustained anti-CHIKV protection was also conferred in suckling mice pretreated with shRNA E1 (Lam et al., 2012).
8.2. Experimental therapies targeting host responses to infection
8.2.1. Sphingosine kinase 1 activity
In a recent review, Carr et al. summarised the information available regarding sphingosine kinase 1 (SphK1) activity against virus infections (Carr et al., 2012). SphK1 has been proposed as a potential therapeutic agent for the treatment of cancer, immunological and inflammatory disorders. SphK1 catalyses the phosphorylated form of sphingosine to generate sphingosine-1-phosphate (S1P). The SphK1/S1P axis has well described roles in cell signalling, viz., the cell death/survival decision, the production of a pro-inflammatory response, immunomodulation and control of vascular integrity. The authors argued that the clinical picture of chikungunya fever is similar to the painful joint syndrome of rheumatoid arthritis (RA). Accordingly, SphK1/S1P could play a significant role in infections by RNA viruses. Indeed, data suggest that raising S1P systematically, or reducing SphK1 activity and S1P locally in joints, does appear to have beneficial effects in clinical cases of RA. Moreover S1P lyase inhibitors are under investigation for RA
8.2.2. Inhibitor of monocyte chemotactic protein (MCP)
The mechanisms by which arthritogenic alphaviruses such as CHIKV cause disease are poorly understood, but the inflammatory response to infection contributes to viral elimination from the blood and clinical recovery at both the acute stage and the chronic form of the disease (Dupuis-Maguiraga et al., 2012). It has also been suggested that macrophages play a critical role in the inflammatory response (Dupuis-Maguiraga et al., 2012).
An inhibitor of monocyte chemotactic protein (MCP) synthesis, called Bindarit, was recently shown to be a potent anti-inflammatory drug, which does not cause systemic immunosuppression or affect arachidonic acid metabolism (Rulli et al., 2009). Clinical trials have confirmed the positive safety of Bindarit and its oral availability (Rulli et al., 2009). In two recent studies, Bindarit treatment improved the articular symptoms of CHIKV and RRV disease in mice and reduced tissue destruction, without affecting the viral load in the tissues (Rulli et al., 2009, Rulli et al., 2011).
In summary, no antiviral drug is currently available for the treatment of standard presentations of chikungunya fever. Chloroquine, which has been used in the past, should not be utilised at the acute phase of the disease, based on data collected during a prospective clinical trial in Reunion Island. In severe presentations, ribavirin may be proposed, but very limited information is available and the precise indications of the treatment and the therapeutic protocol (e.g., doses and duration) have not been established.
However, on a more positive note, current investigations may lead to the identification of new antiviral candidates, as an increasing number of molecules are being associated with a clearly defined mechanism of viral inhibition in cell-based systems, and show significant activity in animal models. Therapeutic protocols for severe cases of chikungunya fever may also be established, based on specific immunoglobulins or molecules that can interfere with some aspects of the inflammatory response associated with CHIKV infection.
Acknowledgments
The authors are grateful to Dr Christian Devaux for providing access to figures from Solignat et al. (2009), Dr Jean-Claude Desenclos and Dr Harold Noël (Institut de Veille Sanitaire, France) for providing unpublished data about French imported cases and Pr Fabrice Simon for helpful discussion. LDM is a post-doc financed by the ICRES Project of the European Commission.
Footnotes
Supplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/j.antiviral.2013.06.009.
Appendix A. Supplementary data
Epidemiological record and outbreak bulletin of chikungunya disease from the World Health Organisation (WHO). The weekly epidemiological records are available every Friday on the website: http://www.who.int/wer/en/. The outbreak bulletins of the WHO Regional Office for Africa are available on the website http://www.afro.who.int. We present here all epidemiological records and outbreak bulletins that include the keyword “chikungunya”.
ProMED-Mail alerts of chikungunya disease. Alerts are available on the website: http://www.promedmail.org. ProMED (Program for Monitoring Emerging Diseases) is an internet-based reporting system dedicated to rapid global dissemination of information on infectious disease outbreaks. Sources of information include media reports, official reports, online summaries, local observations, and others. A team of expert moderators screens, reviews and investigates reports before posting to the network. ProMED-mail currently reaches over 60,000 subscribers in at least 185 countries. This table summarises ProMED-mail alerts relating to chikungunya disease.
CHIKV sequences available from the NIH GenBank genetic sequence database. GenBank is a comprehensive database that contains publicly available nucleotide sequences for more than 380,000 organisms named at the genus level or lower, obtained primarily through submissions from individual laboratories and batch submissions from large-scale sequencing projects. GenBank is built and distributed by the National centre for Biotechnology Information, a division of the National Library of Medicine, located on the campus of the US National Institutes of Health in Bethesda, MD, USA. We summarise here the GenBank sequences of CHIKV available on the website: http://www.ncbi.nlm.nih.gov/nuccore and with nucleotide size equal or higher than 200 bp.
PubMed references of chikungunya cases. PubMed is a free database accessing primarily the MEDLINE database of references and abstracts on life sciences and biomedical topics. This database is maintained by the United States National Library of Medicine at the National Institutes of Health in Bethesda, MD, USA. We summarise here the main publications that report outbreaks or imported cases of chikungunya disease referenced on the website: http://www.ncbi.nlm.nih.gov/pubmed.
PubMed references of seroprevalence studies of chikungunya virus. The table summarises studies that report serologic surveys (IgG) within human populations and referenced on NCBI, available on the website: http://www.ncbi.nlm.nih.gov/pubmed.
Summary of the Pubmed references, WHO publications, GenBank sequences and ProMED alerts for chikungunya virus. We summarise here (i) PubMed references [excluding imported cases], (ii) WHO publications [excluding imported cases], (iii) GenBank sequences and (iv) ProMED-mail alerts, relevant to chikungunya virus, described in Supplemental Tables 1–5.
References
- Aikat B.K., Konar N.R., Banerjee G. Haemorrhagic Fever in Calcutta Area. Indian J. Med. Res. 1964;52:660–675. [PubMed] [Google Scholar]
- Akahata W., Nabel G.J. A specific domain of the chikungunya virus e2 protein regulates particle formation in human cells: implications for alphavirus vaccine design. J. Virol. 2012;86:8879–8883. doi: 10.1128/JVI.00370-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akahata W., Yang Z.Y., Andersen H., Sun S., Holdaway H.A., Kong W.P., Lewis M.G., Higgs S., Rossmann M.G., Rao S., Nabel G.J. A virus-like particle vaccine for epidemic Chikungunya virus protects nonhuman primates against infection. Nat. Med. 2010;16:334–338. doi: 10.1038/nm.2105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ali Ou Alla S., Combe B. Arthritis after infection with Chikungunya virus. Best Pract. Res. Clin. Rheumatol. 2011;25:337–346. doi: 10.1016/j.berh.2011.03.005. [DOI] [PubMed] [Google Scholar]
- Allard P.M., Leyssen P., Martin M.T., Bourjot M., Dumontet V., Eydoux C., Guillemot J.C., Canard B., Poullain C., Gueritte F., Litaudon M. Antiviral chlorinated daphnane diterpenoid orthoesters from the bark and wood of Trigonostemon cherrieri. Phytochemistry. 2012;84:160–168. doi: 10.1016/j.phytochem.2012.07.023. [DOI] [PubMed] [Google Scholar]
- Allard P.M., Martin M.T., Dau M.E., Leyssen P., Gueritte F., Litaudon M. Trigocherrin A, the first natural chlorinated daphnane diterpene orthoester from Trigonostemon cherrieri. Org. Lett. 2012;14:342–345. doi: 10.1021/ol2030907. [DOI] [PubMed] [Google Scholar]
- Angelini R., Finarelli A.C., Angelini P., Po C., Petropulacos K., Macini P., Fiorentini C., Fortuna C., Venturi G., Romi R., Majori G., Nicoletti L., Rezza G., Cassone A. An outbreak of chikungunya fever in the province of Ravenna, Italy. Eur. Surveill. 2007;12:E070906.1. doi: 10.2807/esw.12.36.03260-en. [DOI] [PubMed] [Google Scholar]
- Apandi Y., Nazni W.A., Noor Azleen Z.A., Vythilingam I., Noorazian M.Y., Azahari A.H., Zainah S., Lee H.L. The first isolation of chikungunya virus from non-human primates in Malaysia. J. Gen. Mol. Virol. 2009;1:035–039. [Google Scholar]
- Appassakij H., Khuntikij P., Kemapunmanus M., Wutthanarungsan R., Silpapojakul K. Viremic profiles in asymptomatic and symptomatic chikungunya fever: a blood transfusion threat? Transfusion. 2012 doi: 10.1111/j.1537-2995.2012.03960.x. [DOI] [PubMed] [Google Scholar]
- Arankalle V.A., Shrivastava S., Cherian S., Gunjikar R.S., Walimbe A.M., Jadhav S.M., Sudeep A.B., Mishra A.C. Genetic divergence of Chikungunya viruses in India (1963–2006) with special reference to the 2005–2006 explosive epidemic. J. Gen. Virol. 2007;88:1967–1976. doi: 10.1099/vir.0.82714-0. [DOI] [PubMed] [Google Scholar]
- Bandyopadhyay D., Ghosh S.K. Mucocutaneous manifestations of Chikungunya fever. Indian J. Dermatol. 2010;55:64–67. doi: 10.4103/0019-5154.60356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benedict M.Q., Levine R.S., Hawley W.A., Lounibos L.P. Spread of the tiger: global risk of invasion by the mosquito Aedes albopictus. Vector Borne Zoonotic Dis. 2007;7:76–85. doi: 10.1089/vbz.2006.0562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Binn L.N., Harrison V.R., Randall R. Patterns of viremia and antibody observed in rhesus monkeys inoculated with chikungunya and other serologically related group A arboviruses. Am. J. Trop. Med. Hyg. 1967;16:782–785. [PubMed] [Google Scholar]
- Bodenmann P., Genton B. Chikungunya: an epidemic in real time. Lancet. 2006;368:258. doi: 10.1016/S0140-6736(06)69046-6. [DOI] [PubMed] [Google Scholar]
- Bonilauri P., Bellini R., Calzolari M., Angelini R., Venturi L., Fallacara F., Cordioli P., Angelini P., Venturelli C., Merialdi G., Dottori M. Chikungunya virus in Aedes albopictus, Italy. Emerg. Infect. Dis. 2008;14:852–854. doi: 10.3201/eid1405.071144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borgherini G., Poubeau P., Jossaume A., Gouix A., Cotte L., Michault A., Arvin-Berod C., Paganin F. Persistent arthralgia associated with chikungunya virus: a study of 88 adult patients on reunion island. Clin. Infect. Dis. 2008;47:469–475. doi: 10.1086/590003. [DOI] [PubMed] [Google Scholar]
- Borgherini G., Poubeau P., Paganin F. Chikungunya epidemic in Reunion Island. Epidemiol. Infect. 2009;137:542–543. doi: 10.1017/S0950268809002209. author reply 543. [DOI] [PubMed] [Google Scholar]
- Borgherini G., Poubeau P., Staikowsky F., Lory M., Le Moullec N., Becquart J.P., Wengling C., Michault A., Paganin F. Outbreak of chikungunya on Reunion Island: early clinical and laboratory features in 157 adult patients. Clin. Infect. Dis. 2007;44:1401–1407. doi: 10.1086/517537. [DOI] [PubMed] [Google Scholar]
- Boriskin Y.S., Leneva I.A., Pecheur E.I., Polyak S.J. Arbidol: a broad-spectrum antiviral compound that blocks viral fusion. Curr. Med. Chem. 2008;15:997–1005. doi: 10.2174/092986708784049658. [DOI] [PubMed] [Google Scholar]
- Bouquillard E., Combe B. A report of 21 cases of rheumatoid arthritis following Chikungunya fever. A mean follow-up of two years. Joint Bone Spine. 2009;76:654–657. doi: 10.1016/j.jbspin.2009.08.005. [DOI] [PubMed] [Google Scholar]
- Bourjot M., Delang L., Nguyen V.H., Neyts J., Gueritte F., Leyssen P., Litaudon M. Prostratin and 12-o-tetradecanoylphorbol 13-acetate are potent and selective inhibitors of chikungunya virus replication. J. Nat. Prod. 2012;75:2183–2187. doi: 10.1021/np300637t. [DOI] [PubMed] [Google Scholar]
- Bourjot M., Leyssen P., Eydoux C., Guillemot J.C., Canard B., Rasoanaivo P., Gueritte F., Litaudon M. Chemical constituents of Anacolosa pervilleana and their antiviral activities. Fitoterapia. 2012;83:1076–1080. doi: 10.1016/j.fitote.2012.05.004. [DOI] [PubMed] [Google Scholar]
- Bourjot M., Leyssen P., Eydoux C., Guillemot J.C., Canard B., Rasoanaivo P., Gueritte F., Litaudon M. Flacourtosides A–F, Phenolic Glycosides Isolated from Flacourtia ramontchi. J. Nat. Prod. 2012;75:752–758. doi: 10.1021/np300059n. [DOI] [PubMed] [Google Scholar]
- Brighton S.W. Chloroquine phosphate treatment of chronic Chikungunya arthritis. An open pilot study. S. Afr. Med. J. 1984;66:217–218. [PubMed] [Google Scholar]
- Briolant S., Garin D., Scaramozzino N., Jouan A., Crance J.M. In vitro inhibition of Chikungunya and Semliki Forest viruses replication by antiviral compounds: synergistic effect of interferon-alpha and ribavirin combination. Antiviral Res. 2004;61:111–117. doi: 10.1016/j.antiviral.2003.09.005. [DOI] [PubMed] [Google Scholar]
- Brooks M.J., Burtseva E.I., Ellery P.J., Marsh G.A., Lew A.M., Slepushkin A.N., Crowe S.M., Tannock G.A. Antiviral activity of arbidol, a broad-spectrum drug for use against respiratory viruses, varies according to test conditions. J. Med. Virol. 2012;84:170–181. doi: 10.1002/jmv.22234. [DOI] [PubMed] [Google Scholar]
- Brouard C., Bernillon P., Quatresous I., Pillonel J., Assal A., De Valk H., Desenclos J.C. Estimated risk of Chikungunya viremic blood donation during an epidemic on Reunion Island in the Indian Ocean, 2005 to 2007. Transfusion. 2008;48:1333–1341. doi: 10.1111/j.1537-2995.2008.01646.x. [DOI] [PubMed] [Google Scholar]
- Brown F. Inactivation of viruses by aziridines. Vaccine. 2001;20:322–327. doi: 10.1016/s0264-410x(01)00342-5. [DOI] [PubMed] [Google Scholar]
- Burns C.C., Shaw J., Campagnoli R., Jorba J., Vincent A., Quay J., Kew O. Modulation of poliovirus replicative fitness in HeLa cells by deoptimization of synonymous codon usage in the capsid region. J. Virol. 2006;80:3259–3272. doi: 10.1128/JVI.80.7.3259-3272.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt F.J., Rolph M.S., Rulli N.E., Mahalingam S., Heise M.T. Chikungunya: a re-emerging virus. Lancet. 2012;379:662–671. doi: 10.1016/S0140-6736(11)60281-X. [DOI] [PubMed] [Google Scholar]
- Carey D.E. Chikungunya and dengue: a case of mistaken identity? J. Hist. Med. Allied Sci. 1971;26:243–262. doi: 10.1093/jhmas/xxvi.3.243. [DOI] [PubMed] [Google Scholar]
- Carr J.M., Mahalingam S., Bonder C.S., Pitson S.M. Sphingosine kinase 1 in viral infections. Rev. Med. Virol. 2012;12:73–84. doi: 10.1002/rmv.1718. [DOI] [PubMed] [Google Scholar]
- Charrel R.N., de Lamballerie X., Raoult D. Chikungunya outbreaks – the globalization of vectorborne diseases. N. Engl. J. Med. 2007;356:769–771. doi: 10.1056/NEJMp078013. [DOI] [PubMed] [Google Scholar]
- Chastel C. Human Infections in Cambodia by the Chikungunya Virus or a Closely Related Agent. 3. Epidemiology. Bull. Soc. Pathol. Exot. Filiales. 1964;57:65–82. [PubMed] [Google Scholar]
- Chastel C. Asymptomatic infections in man: a Trojan horse for the introduction and spread of mosquito-borne arboviruses in non-endemic areas? Bull. Soc. Pathol. Exot. 2011;104:213–219. doi: 10.1007/s13149-011-0165-1. [DOI] [PubMed] [Google Scholar]
- Chattopadhyay A., Wang E., Seymour R., Weaver S.C., Rose J.K. A Chimeric Vesiculo/Alphavirus is an effective Alphavirus Vaccine. J. Virol. 2013;87:395–402. doi: 10.1128/JVI.01860-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C.I., Clark D.C., Pesavento P., Lerche N.W., Luciw P.A., Reisen W.K., Brault A.C. Comparative pathogenesis of epidemic and enzootic Chikungunya viruses in a pregnant Rhesus macaque model. Am. J. Trop. Med. Hyg. 2010;83:1249–1258. doi: 10.4269/ajtmh.2010.10-0290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cherian S.S., Walimbe A.M., Jadhav S.M., Gandhe S.S., Hundekar S.L., Mishra A.C., Arankalle V.A. Evolutionary rates and timescale comparison of Chikungunya viruses inferred from the whole genome/E1 gene with special reference to the 2005–07 outbreak in the Indian subcontinent. Infect. Genet. Evol. 2009;9:16–23. doi: 10.1016/j.meegid.2008.09.004. [DOI] [PubMed] [Google Scholar]
- Chow A., Her Z., Ong E.K., Chen J.M., Dimatatac F., Kwek D.J., Barkham T., Yang H., Renia L., Leo Y.S., Ng L.F. Persistent arthralgia induced by Chikungunya virus infection is associated with interleukin-6 and granulocyte macrophage colony-stimulating factor. J. Infect. Dis. 2011;203:149–157. doi: 10.1093/infdis/jiq042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coleman J.R., Papamichail D., Skiena S., Futcher B., Wimmer E., Mueller S. Virus attenuation by genome-scale changes in codon pair bias. Science. 2008;320:1784–1787. doi: 10.1126/science.1155761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cordel H., Quatresous I., Paquet C., Couturier E. Imported cases of chikungunya in metropolitan France, April 2005–February 2006. Euro Surveill. 2006;11:E060420.3. doi: 10.2807/esw.11.16.02944-en. [DOI] [PubMed] [Google Scholar]
- Couderc T., Chretien F., Schilte C., Disson O., Brigitte M., Guivel-Benhassine F., Touret Y., Barau G., Cayet N., Schuffenecker I., Despres P., Arenzana-Seisdedos F., Michault A., Albert M.L., Lecuit M. A mouse model for Chikungunya: young age and inefficient type-I interferon signaling are risk factors for severe disease. PLoS Pathog. 2008;4:e29. doi: 10.1371/journal.ppat.0040029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Couderc T., Gangneux N., Chretien F., Caro V., Le Luong T., Ducloux B., Tolou H., Lecuit M., Grandadam M. Chikungunya virus infection of corneal grafts. J. Infect. Dis. 2012;206:851–859. doi: 10.1093/infdis/jis296. [DOI] [PubMed] [Google Scholar]
- Couderc T., Khandoudi N., Grandadam M., Visse C., Gangneux N., Bagot S., Prost J.F., Lecuit M. Prophylaxis and therapy for Chikungunya virus infection. J. Infect. Dis. 2009;200:516–523. doi: 10.1086/600381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Couderc T., Lecuit M. Focus on Chikungunya pathophysiology in human and animal models. Microbes Infect. 2009;11:1197–1205. doi: 10.1016/j.micinf.2009.09.002. [DOI] [PubMed] [Google Scholar]
- Couturier E., Guillemin F., Mura M., Leon L., Virion J.M., Letort M.J., De Valk H., Simon F., Vaillant V. Impaired quality of life after chikungunya virus infection: a 2-year follow-up study. Rheumatology (Oxford, England) 2012;51:1315–1322. doi: 10.1093/rheumatology/kes015. [DOI] [PubMed] [Google Scholar]
- Crotty S., Cameron C., Andino R. Ribavirin’s antiviral mechanism of action: lethal mutagenesis? J. Mol. Med. (Berl.) 2002;80:86–95. doi: 10.1007/s00109-001-0308-0. [DOI] [PubMed] [Google Scholar]
- D’Ortenzio E., Grandadam M., Balleydier E., Dehecq J.S., Jaffar-Bandjee M.C., Michault A., Andriamandimby S.F., Reynes J.M., Filleul L. Sporadic cases of chikungunya, Reunion Island, August 2009. Euro Surveill. 2009;14 [PubMed] [Google Scholar]
- D’Ortenzio E., Grandadam M., Balleydier E., Jaffar-Bandjee M.C., Michault A., Brottet E., Baville M., Filleul L. A226V strains of Chikungunya virus, Reunion Island, 2010. Emerg. Infect. Dis. 2011;17:309–311. doi: 10.3201/eid1702.101056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darwin J.R., Kenney J.L., Weaver S.C. Transmission potential of two chimeric Chikungunya vaccine candidates in the urban mosquito vectors, Aedes aegypti and Ae. albopictus. Am. J. Trop. Med. Hyg. 2011;84:1012–1015. doi: 10.4269/ajtmh.2011.11-0049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darwish M.A., Hoogstraal H., Roberts T.J., Ahmed I.P., Omar F. A sero-epidemiological survey for certain arboviruses (Togaviridae) in Pakistan. Trans. R. Soc. Trop. Med. Hyg. 1983;77:442–445. doi: 10.1016/0035-9203(83)90106-2. [DOI] [PubMed] [Google Scholar]
- Das T., Jaffar-Bandjee M.C., Hoarau J.J., Krejbich Trotot P., Denizot M., Lee-Pat-Yuen G., Sahoo R., Guiraud P., Ramful D., Robin S., Alessandri J.L., Gauzere B.A., Gasque P. Chikungunya fever: CNS infection and pathologies of a re-emerging arbovirus. Prog. Neurobiol. 2010;91:121–129. doi: 10.1016/j.pneurobio.2009.12.006. [DOI] [PubMed] [Google Scholar]
- Dash P.K., Tiwari M., Santhosh S.R., Parida M., Lakshmana Rao P.V. RNA interference mediated inhibition of Chikungunya virus replication in mammalian cells. Biochem. Biophys. Res. Commun. 2008;376:718–722. doi: 10.1016/j.bbrc.2008.09.040. [DOI] [PubMed] [Google Scholar]
- de Andrade D.C., Jean S., Clavelou P., Dallel R., Bouhassira D. Chronic pain associated with the Chikungunya Fever: long lasting burden of an acute illness. BMC Infect. Dis. 2010;10:31. doi: 10.1186/1471-2334-10-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Lamballerie X., Boisson V., Reynier J.C., Enault S., Charrel R.N., Flahault A., Roques P., Le Grand R. On chikungunya acute infection and chloroquine treatment. Vector Borne Zoonotic Dis. 2008;8:837–839. doi: 10.1089/vbz.2008.0049. [DOI] [PubMed] [Google Scholar]
- de Lamballerie X., Leroy E., Charrel R.N., Ttsetsarkin K., Higgs S., Gould E.A. Chikungunya virus adapts to tiger mosquito via evolutionary convergence: a sign of things to come? Virol. J. 2008;5:33. doi: 10.1186/1743-422X-5-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delogu I., de Lamballerie X. Chikungunya disease and chloroquine treatment. J. Med. Virol. 2011;83:1058–1059. doi: 10.1002/jmv.22019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delogu I., Pastorino B., Baronti C., Nougairede A., Bonnet E., de Lamballerie X. In vitro antiviral activity of arbidol against Chikungunya virus and characteristics of a selected resistant mutant. Antiviral Res. 2011;90:99–107. doi: 10.1016/j.antiviral.2011.03.182. [DOI] [PubMed] [Google Scholar]
- Depoortere E., Salmaso S., Pompa M.G., Guglielmetti P., Coulombier D. Chikungunya in Europe. Lancet. 2008;371:723. doi: 10.1016/S0140-6736(08)60336-0. [DOI] [PubMed] [Google Scholar]
- Diallo M., Thonnon J., Traore-Lamizana M., Fontenille D. Vectors of Chikungunya virus in Senegal: current data and transmission cycles. Am. J. Trop. Med. Hyg. 1999;60:281–286. doi: 10.4269/ajtmh.1999.60.281. [DOI] [PubMed] [Google Scholar]
- Dupuis-Maguiraga L., Noret M., Brun S., Le Grand R., Gras G., Roques P. Chikungunya disease: infection-associated markers from the acute to the chronic phase of arbovirus-induced arthralgia. PLoS Negl. Trop. Dis. 2012;6:e1446. doi: 10.1371/journal.pntd.0001446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Economopoulou A., Dominguez M., Helynck B., Sissoko D., Wichmann O., Quenel P., Germonneau P., Quatresous I. Atypical Chikungunya virus infections: clinical manifestations, mortality and risk factors for severe disease during the 2005–2006 outbreak on Reunion. Epidemiol. Infect. 2009;137:534–541. doi: 10.1017/S0950268808001167. [DOI] [PubMed] [Google Scholar]
- Edelman R., Tacket C.O., Wasserman S.S., Bodison S.A., Perry J.G., Mangiafico J.A. Phase II safety and immunogenicity study of live chikungunya virus vaccine TSI-GSD-218. Am. J. Trop. Med. Hyg. 2000;62:681–685. doi: 10.4269/ajtmh.2000.62.681. [DOI] [PubMed] [Google Scholar]
- Enserink M. Infectious diseases. Massive outbreak draws fresh attention to little-known virus. Science. 2006;311:1085. doi: 10.1126/science.311.5764.1085a. [DOI] [PubMed] [Google Scholar]
- Enserink M. Infectious diseases. Chikungunya: no longer a third world disease. Science. 2007;318:1860–1861. doi: 10.1126/science.318.5858.1860. [DOI] [PubMed] [Google Scholar]
- Filipe A.F., Pinto M.R. Arbovirus studies in Luanda, Angola. 2. Virological and serological studies during an outbreak of dengue-like disease caused by the Chikungunya virus. Bull. World Health Organ. 1973;49:37–40. [PMC free article] [PubMed] [Google Scholar]
- Fourie E.D., Morrison J.G. Rheumatoid arthritic syndrome after chikungunya fever. S. Afr. Med. J. 1979;56:130–132. [PubMed] [Google Scholar]
- Frank C., Schoneberg I., Stark K. Trends in imported chikungunya virus infections in Germany, 2006–2009. Vector Borne Zoonotic Dis. 2011;11:631–636. doi: 10.1089/vbz.2010.0269. [DOI] [PubMed] [Google Scholar]
- Fric J., Bertin-Maghit S., Wang C.I., Nardin A., Warter L. Use of human monoclonal antibodies to treat chikungunya virus infection. J. Infect. Dis. 2013;207:319–322. doi: 10.1093/infdis/jis674. [DOI] [PubMed] [Google Scholar]
- Fritel X., Rollot O., Gerardin P., Gauzere B.A., Bideault J., Lagarde L., Dhuime B., Orvain E., Cuillier F., Ramful D., Samperiz S., Jaffar-Bandjee M.C., Michault A., Cotte L., Kaminski M., Fourmaintraux A. Chikungunya virus infection during pregnancy, Reunion, France, 2006. Emerg. Infect. Dis. 2010;16:418–425. doi: 10.3201/eid1604.091403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frolov I., Akhrymuk M., Akhrymuk I., Atasheva S., Frolova E.I. Early events in alphavirus replication determine the outcome of infection. J. Virol. 2012;86:5055–5066. doi: 10.1128/JVI.07223-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fros J.J., Liu W.J., Prow N.A., Geertsema C., Ligtenberg M., Vanlandingham D.L., Schnettler E., Vlak J.M., Suhrbier A., Khromykh A.A., Pijlman G.P. Chikungunya virus nonstructural protein 2 inhibits type I/II interferon-stimulated JAK-STAT signaling. J. Virol. 2010;84:10877–10887. doi: 10.1128/JVI.00949-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner C.L., Burke C.W., Higgs S.T., Klimstra W.B., Ryman K.D. Interferon-alpha/beta deficiency greatly exacerbates arthritogenic disease in mice infected with wild-type chikungunya virus but not with the cell culture-adapted live-attenuated 181/25 vaccine candidate. Virology. 2012;425:103–112. doi: 10.1016/j.virol.2011.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner J., Anraku I., Le T.T., Larcher T., Major L., Roques P., Schroder W.A., Higgs S., Suhrbier A. Chikungunya virus arthritis in adult wild-type mice. J. Virol. 2010;84:8021–8032. doi: 10.1128/JVI.02603-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerardin P. Paediatric features of Dengue and Chikungunya fevers. Arch. Pediatr. 2010;17:86–90. doi: 10.1016/j.arcped.2009.09.001. [DOI] [PubMed] [Google Scholar]
- Gerardin P., Barau G., Michault A., Bintner M., Randrianaivo H., Choker G., Lenglet Y., Touret Y., Bouveret A., Grivard P., Le Roux K., Blanc S., Schuffenecker I., Couderc T., Arenzana-Seisdedos F., Lecuit M., Robillard P.Y. Multidisciplinary prospective study of mother-to-child chikungunya virus infections on the island of La Reunion. PLoS Med. 2008;5:e60. doi: 10.1371/journal.pmed.0050060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerardin P., Fianu A., Malvy D., Mussard C., Boussaid K., Rollot O., Michault A., Gauzere B.A., Breart G., Favier F. Perceived morbidity and community burden after a Chikungunya outbreak: the TELECHIK survey, a population-based cohort study. BMC Med. 2011;9:5. doi: 10.1186/1741-7015-9-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerardin P., Fianu A., Michault A., Mussard C., Boussaid K., Rollot O., Grivard P., Kassab S., Bouquillard E., Borgherini G., Gauzere B.A., Malvy D., Breart G., Favier F. Predictors of Chikungunya rheumatism: a prognostic survey ancillary to the TELECHIK cohort study. Arthritis Res. Ther. 2013;15:R9. doi: 10.1186/ar4137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerardin P., Guernier V., Perrau J., Fianu A., Le Roux K., Grivard P., Michault A., de Lamballerie X., Flahault A., Favier F. Estimating Chikungunya prevalence in La Reunion Island outbreak by serosurveys: two methods for two critical times of the epidemic. BMC Infect. Dis. 2008;8:99. doi: 10.1186/1471-2334-8-99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerardin P., Perrau J., Fianu A., Favier F. Institut de Veille Sanitaire; 2008. Determinants of Chikungunya Virus Infection in the Reunion island: Results of the SEROCHIK Seroprevalence Survey in the Population, August–October 2006, BEH du 21 Octobre 2008 / n 38–39-40. [Google Scholar]
- Gibbons D.L., Vaney M.C., Roussel A., Vigouroux A., Reilly B., Lepault J., Kielian M., Rey F.A. Conformational change and protein-protein interactions of the fusion protein of Semliki Forest virus. Nature. 2004;427:320–325. doi: 10.1038/nature02239. [DOI] [PubMed] [Google Scholar]
- Gilotra S.K., Bhattacharya N.C. Mosquito vectors of dengue–chikungunya viruses in a rural area near Calcutta. Bull. Calcutta Sch. Trop. Med. 1968;16:41–42. [PubMed] [Google Scholar]
- Gilotra S.K., Shah K.V. Laboratory studies on transmission of Chikungunya virus by mosquitoes. Am. J. Epidemiol. 1967;86:379–385. doi: 10.1093/oxfordjournals.aje.a120748. [DOI] [PubMed] [Google Scholar]
- Gorchakov R., Wang E., Leal G., Forrester N.L., Plante K., Rossi S.L., Partidos C.D., Adams A.P., Seymour R.L., Weger J., Borland E.M., Sherman M.B., Powers A.M., Osorio J.E., Weaver S.C. Attenuation of Chikungunya virus vaccine strain 181/clone 25 is determined by two amino acid substitutions in the E2 envelope glycoprotein. J. Virol. 2012;86:6084–6096. doi: 10.1128/JVI.06449-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould E.A., Gallian P., De Lamballerie X., Charrel R.N. First cases of autochthonous dengue fever and chikungunya fever in France: from bad dream to reality! Clin. Microbiol. Infect. 2010;16:1702–1704. doi: 10.1111/j.1469-0691.2010.03386.x. [DOI] [PubMed] [Google Scholar]
- Grandadam M., Caro V., Plumet S., Thiberge J.M., Souares Y., Failloux A.B., Tolou H.J., Budelot M., Cosserat D., Leparc-Goffart I., Despres P. Chikungunya virus, southeastern France. Emerg. Infect. Dis. 2011;17:910–913. doi: 10.3201/eid1705.101873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halstead, S.B. 2009. Chikungunya. In: Feigin, R.D., Cherry, J.D., Demmler-Harrison, G.J., Kaplan, S.L. (Eds), Feigin and Cherry’s Textbook of Pediatric Infectious Diseases, Saunders, sixth ed., pp. 2314–2319.
- Halstead S.B., Nimmannitya S., Margiotta M.R. Dengue d chikungunya virus infection in man in Thailand, 1962–1964. II. Observations on disease in outpatients. Am. J. Trop. Med. Hyg. 1969;18:972–983. doi: 10.4269/ajtmh.1969.18.972. [DOI] [PubMed] [Google Scholar]
- Halstead S.B., Scanlon J.E., Umpaivit P., Udomsakdi S. Dengue and chikungunya virus infection in man in Thailand, 1962–1964. IV. Epidemiologic studies in the Bangkok metropolitan area. Am. J. Trop. Med. Hyg. 1969;18:997–1021. doi: 10.4269/ajtmh.1969.18.997. [DOI] [PubMed] [Google Scholar]
- Hannon G.J. RNA interference. Nature. 2002;418:244–251. doi: 10.1038/418244a. [DOI] [PubMed] [Google Scholar]
- Harrison V.R., Eckels K.H., Bartelloni P.J., Hampton C. Production and evaluation of a formalin-killed Chikungunya vaccine. J. Immunol. 1971;107:643–647. [PubMed] [Google Scholar]
- Heim M.H. Interferons and hepatitis C virus. Swiss Med. Wkly. 2012;142:w13586. doi: 10.4414/smw.2012.13586. [DOI] [PubMed] [Google Scholar]
- Her Z., Malleret B., Chan M., Ong E.K., Wong S.C., Kwek D.J., Tolou H., Lin R.T., Tambyah P.A., Renia L., Ng L.F. Active infection of human blood monocytes by Chikungunya virus triggers an innate immune response. J. Immunol. 2010;184:5903–5913. doi: 10.4049/jimmunol.0904181. [DOI] [PubMed] [Google Scholar]
- Hermon Y.E. Virological investigations of Arbovirus infections in Ceylon, with special reference to the recent Chikungunya fever epidemic. Ceylon Med. J. 1967;12:81–92. [PubMed] [Google Scholar]
- Higgs S., Ziegler S.A. A nonhuman primate model of chikungunya disease. J. Clin. Invest. 2010;120:657–660. doi: 10.1172/JCI42392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho P.S., Ng M.M., Chu J.J. Establishment of one-step SYBR green-based real time-PCR assay for rapid detection and quantification of chikungunya virus infection. Virol. J. 2010;7:13. doi: 10.1186/1743-422X-7-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoarau J.J., Jaffar Bandjee M.C., Krejbich Trotot P., Das T., Li-Pat-Yuen G., Dassa B., Denizot M., Guichard E., Ribera A., Henni T., Tallet F., Moiton M.P., Gauzere B.A., Bruniquet S., Jaffar Bandjee Z., Morbidelli P., Martigny G., Jolivet M., Gay F., Grandadam M., Tolou H., Vieillard V., Debre P., Autran B., Gasque P. Persistent chronic inflammation and infection by Chikungunya arthritogenic alphavirus in spite of a robust host immune response. J. Immunol. 2010;184:5914–5927. doi: 10.4049/jimmunol.0900255. [DOI] [PubMed] [Google Scholar]
- Hoarau J.J., Jaffar Bandjee M.C., Trotot P.K., Das T., Li-Pat-Yuen G., Dassa B., Denizot M., Guichard E., Ribera A., Henni T., Tallet F., Moiton M.P., Gauzere B.A., Bruniquet S., Jaffar Bandjee Z., Morbidelli P., Martigny G., Jolivet M., Gay F., Grandadam M., Tolou H., Vieillard V., Debre P., Autran B., Gasque P. Persistent chronic inflammation and infection by Chikungunya arthritogenic alphavirus in spite of a robust host immune response. J. Immunol. 2010;184:5914–5927. doi: 10.4049/jimmunol.0900255. [DOI] [PubMed] [Google Scholar]
- Hoke C.H., Jr., Pace-Templeton J., Pittman P., Malinoski F.J., Gibbs P., Ulderich T., Mathers M., Fogtman B., Glass P., Vaughn D.W. US Military contributions to the global response to pandemic chikungunya. Vaccine. 2012;30:6713–6720. doi: 10.1016/j.vaccine.2012.08.025. [DOI] [PubMed] [Google Scholar]
- Huang J.H., Yang C.F., Su C.L., Chang S.F., Cheng C.H., Yu S.K., Lin C.C., Shu P.Y. Imported chikungunya virus strains, Taiwan, 2006–2009. Emerg. Infect. Dis. 2009;15:1854–1856. doi: 10.3201/eid1511.090398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inglot A.D. Comparison of the antiviral activity in vitro of some non-steroidal anti-inflammatory drugs. J. Gen. Virol. 1969;4:203–214. doi: 10.1099/0022-1317-4-2-203. [DOI] [PubMed] [Google Scholar]
- Inoue S., Morita K., Matias R.R., Tuplano J.V., Resuello R.R., Candelario J.R., Cruz D.J., Mapua C.A., Hasebe F., Igarashi A., Natividad F.F. Distribution of three arbovirus antibodies among monkeys (Macaca fascicularis) in the Philippines. J. Med. Primatol. 2003;32:89–94. doi: 10.1034/j.1600-0684.2003.00015.x. [DOI] [PubMed] [Google Scholar]
- Jose J., Snyder J.E., Kuhn R.J. A structural and functional perspective of alphavirus replication and assembly. Future Microbiol. 2009;4:837–856. doi: 10.2217/fmb.09.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Josseran L., Paquet C., Zehgnoun A., Caillere N., Le Tertre A., Solet J.L., Ledrans M. Chikungunya disease outbreak, Reunion Island. Emerg. Infect. Dis. 2006;12:1994–1995. doi: 10.3201/eid1212.060710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joubert P.E., Werneke S.W., de la Calle C., Guivel-Benhassine F., Giodini A., Peduto L., Levine B., Schwartz O., Lenschow D.J., Albert M.L. Chikungunya virus-induced autophagy delays caspase-dependent cell death. J. Exp. Med. 2012;209:1029–1047. doi: 10.1084/jem.20110996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jupp P.G., McIntosh B.M. Aedes furcifer and other mosquitoes as vectors of chikungunya virus at Mica, northeastern Transvaal, South Africa. J. Am. Mosq. Control Assoc. 1990;6:415–420. [PubMed] [Google Scholar]
- Jupp P.G., McIntosh B.M., Dos Santos I., DeMoor P. Laboratory vector studies on six mosquito and one tick species with chikungunya virus. Trans. R. Soc. Trop. Med. Hyg. 1981;75:15–19. doi: 10.1016/0035-9203(81)90005-5. [DOI] [PubMed] [Google Scholar]
- Kalantri S.P., Joshi R., Riley L.W. Chikungunya epidemic: an Indian perspective. Natl. Med. J. India. 2006;19:315–322. [PubMed] [Google Scholar]
- Kam Y.W., Lee W.W., Simarmata D., Harjanto S., Teng T.S., Tolou H., Chow A., Lin R.T., Leo Y.S., Renia L., Ng L.F. Longitudinal analysis of the human antibody response to Chikungunya virus infection: implications for serodiagnosis and vaccine development. J. Virol. 2012;86:13005–13015. doi: 10.1128/JVI.01780-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kam Y.W., Lum F.M., Teo T.H., Lee W.W., Simarmata D., Harjanto S., Chua C.L., Chan Y.F., Wee J.K., Chow A., Lin R.T., Leo Y.S., Le Grand R., Sam I.C., Tong J.C., Roques P., Wiesmuller K.H., Renia L., Rotzschke O., Ng L.F. Early neutralizing IgG response to Chikungunya virus in infected patients targets a dominant linear epitope on the E2 glycoprotein. EMBO Mol. Med. 2012;4:330–343. doi: 10.1002/emmm.201200213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kam Y.W., Simarmata D., Chow A., Her Z., Teng T.S., Ong E.K., Renia L., Leo Y.S., Ng L.F. Early appearance of neutralizing immunoglobulin G3 antibodies is associated with chikungunya virus clearance and long-term clinical protection. J. Infect. Dis. 2012;205:1147–1154. doi: 10.1093/infdis/jis033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kariuki Njenga M., Nderitu L., Ledermann J.P., Ndirangu A., Logue C.H., Kelly C.H., Sang R., Sergon K., Breiman R., Powers A.M. Tracking epidemic Chikungunya virus into the Indian Ocean from East Africa. J. Gen. Virol. 2008;89:2754–2760. doi: 10.1099/vir.0.2008/005413-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaur P., Thiruchelvan M., Lee R.C., Chen H., Chen K.C., Ng M.L., Chu J.J. Inhibition of chikungunya virus replication by harringtonine, a novel antiviral that suppresses viral protein expression. Antimicrob. Agents Chemother. 2013;57:155–167. doi: 10.1128/AAC.01467-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kee A.C., Yang S., Tambyah P. Atypical chikungunya virus infections in immunocompromised patients. Emerg. Infect. Dis. 2010;16:1038–1040. doi: 10.3201/eid1606.091115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelvin A.A. Outbreak of Chikungunya in the Republic of Congo and the global picture. J. Infect. Dev. Ctries. 2011;5:441–444. [PubMed] [Google Scholar]
- Keyaerts E., Vijgen L., Maes P., Neyts J., Van Ranst M. In vitro inhibition of severe acute respiratory syndrome coronavirus by chloroquine. Biochem. Biophys. Res. Commun. 2004;323:264–268. doi: 10.1016/j.bbrc.2004.08.085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khai Ming C., Thain S., Thaung U., Tin U., Myint K.S., Swe T., Halstead S.B., Diwan A.R. Clinical and laboratory studies on haemorrhagic fever in Burma, 1970–72. Bull. World Health Organ. 1974;51:227–235. [PMC free article] [PubMed] [Google Scholar]
- Khan A.H., Morita K., Parquet Md Mdel C., Hasebe F., Parquet.Md. E.G., Igarashi A. Complete nucleotide sequence of chikungunya virus and evidence for an internal polyadenylation site. J. Gen. Virol. 2002;83:3075–3084. doi: 10.1099/0022-1317-83-12-3075. [DOI] [PubMed] [Google Scholar]
- Khan M., Dhanwani R., Rao P.V., Parida M. Subunit vaccine formulations based on recombinant envelope proteins of Chikungunya virus elicit balanced Th1/Th2 response and virus-neutralizing antibodies in mice. Virus Res. 2012;167:236–246. doi: 10.1016/j.virusres.2012.05.004. [DOI] [PubMed] [Google Scholar]
- Kim D.Y., Atasheva S., Foy N.J., Wang E., Frolova E.I., Weaver S., Frolov I. Design of chimeric alphaviruses with a programmed, attenuated, cell type-restricted phenotype. J. Virol. 2011;85:4363–4376. doi: 10.1128/JVI.00065-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King A.M.Q., Adams M.J., Carstens E.B., Lefkowitz E.J., San Diego . Elsevier Academic Press; 2012. Virus taxonomy: classification and nomenclature of viruses: Ninth Report of the International Committee on Taxonomy of Viruses. [Google Scholar]
- Knudsen A.B. Global distribution and continuing spread of Aedes albopictus. Parassitologia. 1995;37:91–97. [PubMed] [Google Scholar]
- Krastinova E., Quatresous I., Tarantola A. Imported cases of chikungunya in metropolitan France: update to June 2006. Euro Surveill. 2006;11:E060824.1. doi: 10.2807/esw.11.34.03030-en. [DOI] [PubMed] [Google Scholar]
- Krejbich-Trotot P., Gay B., Li-Pat-Yuen G., Hoarau J.J., Jaffar-Bandjee M.C., Briant L., Gasque P., Denizot M. Chikungunya triggers an autophagic process which promotes viral replication. Virol. J. 2011;8:432. doi: 10.1186/1743-422X-8-432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krishnamoorthy K., Harichandrakumar K.T., Krishna Kumari A., Das L.K. Burden of chikungunya in India: estimates of disability adjusted life years (DALY) lost in 2006 epidemic. J. Vector Borne Dis. 2009;46:26–35. [PubMed] [Google Scholar]
- Kumar M., Sudeep A.B., Arankalle V.A. Evaluation of recombinant E2 protein-based and whole-virus inactivated candidate vaccines against chikungunya virus. Vaccine. 2012;30:6142–6149. doi: 10.1016/j.vaccine.2012.07.072. [DOI] [PubMed] [Google Scholar]
- Kumar N.C., Nadimpalli M., Vardhan V.R., Gopal S.D. Association of ABO blood groups with Chikungunya virus. Virol. J. 2010;7:140. doi: 10.1186/1743-422X-7-140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar N.P., Suresh A., Vanamail P., Sabesan S., Krishnamoorthy K.G., Mathew J., Jose V.T., Jambulingam P. Chikungunya virus outbreak in Kerala, India, 2007: a seroprevalence study. Mem. Inst. Oswaldo Cruz. 2011;106:912–916. doi: 10.1590/s0074-02762011000800003. [DOI] [PubMed] [Google Scholar]
- Kumarasamy V., Prathapa S., Zuridah H., Chem Y.K., Norizah I., Chua K.B. Re-emergence of Chikungunya virus in Malaysia. Med. J. Malaysia. 2006;61:221–225. [PubMed] [Google Scholar]
- Labadie K., Larcher T., Joubert C., Mannioui A., Delache B., Brochard P., Guigand L., Dubreil L., Lebon P., Verrier B., de Lamballerie X., Suhrbier A., Cherel Y., Le Grand R., Roques P. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J. Clin. Invest. 2010;120:894–906. doi: 10.1172/JCI40104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lahariya C., Pradhan S.K. Emergence of chikungunya virus in Indian subcontinent after 32 years: A review. J. Vector Borne Dis. 2006;43:151–160. [PubMed] [Google Scholar]
- Lam S., Chen K.C., Ng M.M., Chu J.J. Expression of plasmid-based shRNA against the E1 and nsP1 genes effectively silenced Chikungunya virus replication. PLoS One. 2012;7:e46396. doi: 10.1371/journal.pone.0046396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lam S.K., Chua K.B., Hooi P.S., Rahimah M.A., Kumari S., Tharmaratnam M., Chuah S.K., Smith D.W., Sampson I.A. Chikungunya infection – an emerging disease in Malaysia. Southeast Asian J. Trop. Med. Public Health. 2001;32:447–451. [PubMed] [Google Scholar]
- Lampio A., Kilpelainen I., Pesonen S., Karhi K., Auvinen P., Somerharju P., Kaariainen L. Membrane binding mechanism of an RNA virus-capping enzyme. J. Biol. Chem. 2000;275:37853–37859. doi: 10.1074/jbc.M004865200. [DOI] [PubMed] [Google Scholar]
- Laras K., Sukri N.C., Larasati R.P., Bangs M.J., Kosim R., Djauzi, Wandra T., Master J., Kosasih H., Hartati S., Beckett C., Sedyaningsih E.R., Beecham H.R., 3rd, Corwin A.L., Djauzi Wandra. Tracking the re-emergence of epidemic chikungunya virus in Indonesia. Trans. R. Soc. Trop. Med. Hyg. 2005;99:128–141. doi: 10.1016/j.trstmh.2004.03.013. [DOI] [PubMed] [Google Scholar]
- Lee C.Y., Kam Y.W., Fric J., Malleret B., Koh E.G., Prakash C., Huang W., Lee W.W., Lin C., Lin R.T., Renia L., Wang C.I., Ng L.F., Warter L. Chikungunya virus neutralization antigens and direct cell-to-cell transmission are revealed by human antibody-escape mutants. PLoS Pathog. 2011;7:e1002390. doi: 10.1371/journal.ppat.1002390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leneva I.A., Russell R.J., Boriskin Y.S., Hay A.J. Characteristics of arbidol-resistant mutants of influenza virus: implications for the mechanism of anti-influenza action of arbidol. Antiviral Res. 2009;81:132–140. doi: 10.1016/j.antiviral.2008.10.009. [DOI] [PubMed] [Google Scholar]
- Leung J.Y., Ng M.M., Chu J.J. Replication of alphaviruses: a review on the entry process of alphaviruses into cells. Adv. Virol. 2011;2011:249640. doi: 10.1155/2011/249640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levitt N.H., Ramsburg H.H., Hasty S.E., Repik P.M., Cole F.E., Jr., Lupton H.W. Development of an attenuated strain of chikungunya virus for use in vaccine production. Vaccine. 1986;4:157–162. doi: 10.1016/0264-410x(86)90003-4. [DOI] [PubMed] [Google Scholar]
- Li L., Jose J., Xiang Y., Kuhn R.J., Rossmann M.G. Structural changes of envelope proteins during alphavirus fusion. Nature. 2010;468:705–708. doi: 10.1038/nature09546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C.Y., Kielian M. E1 mutants identify a critical region in the trimer interface of the Semliki forest virus fusion protein. J. Virol. 2009;83:11298–11306. doi: 10.1128/JVI.01147-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liumbruno G.M., Calteri D., Petropulacos K., Mattivi A., Po C., Macini P., Tomasini I., Zucchelli P., Silvestri A.R., Sambri V., Pupella S., Catalano L., Piccinini V., Calizzani G., Grazzini G. The Chikungunya epidemic in Italy and its repercussion on the blood system. Blood Transfus. 2008;6:199–210. doi: 10.2450/2008.0016-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lo Presti A., Ciccozzi M., Cella E., Lai A., Simonetti F.R., Galli M., Zehender G., Rezza G. Origin, evolution, and phylogeography of recent epidemic CHIKV strains. Infect. Genet. Evol. 2012;12:392–398. doi: 10.1016/j.meegid.2011.12.015. [DOI] [PubMed] [Google Scholar]
- Lohachanakul J., Phuklia W., Thannagith M., Thonsakulprasert T., Ubol S. High concentrations of circulating interleukin-6 and monocyte chemotactic protein-1 with low concentrations of interleukin-8 were associated with severe chikungunya fever during the 2009–2010 outbreak in Thailand. Microbiol. Immunol. 2012;56:134–138. doi: 10.1111/j.1348-0421.2011.00417.x. [DOI] [PubMed] [Google Scholar]
- Lucas-Hourani M., Lupan A., Despres P., Thoret S., Pamlard O., Dubois J., Guillou C., Tangy F., Vidalain P.O., Munier-Lehmann H. A phenotypic assay to identify chikungunya virus inhibitors targeting the nonstructural protein nsP2. J. Biomol. Screen. 2012;18:172–179. doi: 10.1177/1087057112460091. [DOI] [PubMed] [Google Scholar]
- Lukaszewski R.A., Brooks T.J. Pegylated alpha interferon is an effective treatment for virulent venezuelan equine encephalitis virus and has profound effects on the host immune response to infection. J. Virol. 2000;74:5006–5015. doi: 10.1128/jvi.74.11.5006-5015.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahendradas P., Ranganna S.K., Shetty R., Balu R., Narayana K.M., Babu R.B., Shetty B.K. Ocular manifestations associated with chikungunya. Ophthalmology. 2008;115:287–291. doi: 10.1016/j.ophtha.2007.03.085. [DOI] [PubMed] [Google Scholar]
- Maheshwari R.K., Srikantan V., Bhartiya D. Chloroquine enhances replication of Semliki Forest virus and encephalomyocarditis virus in mice. J. Virol. 1991;65:992–995. doi: 10.1128/jvi.65.2.992-995.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malet H., Coutard B., Jamal S., Dutartre H., Papageorgiou N., Neuvonen M., Ahola T., Forrester N., Gould E.A., Lafitte D., Ferron F., Lescar J., Gorbalenya A.E., de Lamballerie X., Canard B. The crystal structures of Chikungunya and Venezuelan equine encephalitis virus nsP3 macro domains define a conserved adenosine binding pocket. J. Virol. 2009;83:6534–6545. doi: 10.1128/JVI.00189-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mallilankaraman K., Shedlock D.J., Bao H., Kawalekar O.U., Fagone P., Ramanathan A.A., Ferraro B., Stabenow J., Vijayachari P., Sundaram S.G., Muruganandam N., Sarangan G., Srikanth P., Khan A.S., Lewis M.G., Kim J.J., Sardesai N.Y., Muthumani K., Weiner D.B. A DNA vaccine against chikungunya virus is protective in mice and induces neutralizing antibodies in mice and nonhuman primates. PLoS Negl. Trop. Dis. 2011;5:e928. doi: 10.1371/journal.pntd.0000928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manimunda S.P., Mavalankar D., Bandyopadhyay T., Sugunan A.P. Chikungunya epidemic-related mortality. Epidemiol. Infect. 2011;139:1410–1412. doi: 10.1017/S0950268810002542. [DOI] [PubMed] [Google Scholar]
- Manimunda S.P., Vijayachari P., Uppoor R., Sugunan A.P., Singh S.S., Rai S.K., Sudeep A.B., Muruganandam N., Chaitanya I.K., Guruprasad D.R. Clinical progression of chikungunya fever during acute and chronic arthritic stages and the changes in joint morphology as revealed by imaging. Trans. R. Soc. Trop. Med. Hyg. 2010;104:392–399. doi: 10.1016/j.trstmh.2010.01.011. [DOI] [PubMed] [Google Scholar]
- Marchette N.J., Rudnick A., Garcia R., MacVean D.W. Alphaviruses in Peninusular Malaysia: I. Virus isolations and animal serology. Southeast Asian J. Trop. Med. Public Health. 1978;9:317–329. [PubMed] [Google Scholar]
- Marimoutou C., Vivier E., Oliver M., Boutin J.P., Simon F. Morbidity and impaired quality of life 30 months after chikungunya infection: comparative cohort of infected and uninfected French military policemen in Reunion Island. Medicine (Baltimore) 2012;91:212–219. doi: 10.1097/MD.0b013e318260b604. [DOI] [PubMed] [Google Scholar]
- Mason P.J., Haddow A.J. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952–53: an additional note on Chikungunya virus isolations and serum antibodies. Trans. R. Soc. Trop. Med. Hyg. 1957;51:238–240. doi: 10.1016/0035-9203(57)90022-6. [DOI] [PubMed] [Google Scholar]
- McClain D.J., Pittman P.R., Ramsburg H.H., Nelson G.O., Rossi C.A., Mangiafico J.A., Schmaljohn A.L., Malinoski F.J. Immunologic interference from sequential administration of live attenuated alphavirus vaccines. J. Infect. Dis. 1998;177:634–641. doi: 10.1086/514240. [DOI] [PubMed] [Google Scholar]
- McCrae A.W., Henderson B.E., Kirya B.G., Sempala S.D. Chikungunya virus in the Entebbe area of Uganda: isolations and epidemiology. Trans. R. Soc. Trop. Med. Hyg. 1971;65:152–168. doi: 10.1016/0035-9203(71)90212-4. [DOI] [PubMed] [Google Scholar]
- McIntosh B.M., Harwin R.M., Paterson H.E., Westwater M.L. An Epidemic of Chikungunya in South–Eastern Southern Rhodesia. Cent. Afr. J. Med. 1963;43:351–359. [PubMed] [Google Scholar]
- Mendis N.M. Epidemiology of dengue-like fever in Ceylon. Ceylon Med. J. 1967;12:67–74. [PubMed] [Google Scholar]
- Metz S.W., Geertsema C., Martina B.E., Andrade P., Heldens J.G., van Oers M.M., Goldbach R.W., Vlak J.M., Pijlman G.P. Functional processing and secretion of Chikungunya virus E1 and E2 glycoproteins in insect cells. Virol. J. 2011;8:353. doi: 10.1186/1743-422X-8-353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metz S.W., Pijlman G.P. Arbovirus vaccines; opportunities for the baculovirus-insect cell expression system. J. Invertebrate Pathol. 2011;107(Suppl):S16–S30. doi: 10.1016/j.jip.2011.05.002. [DOI] [PubMed] [Google Scholar]
- Mirabel M., Vignaux O., Lebon P., Legmann P., Weber S., Meune C. Acute myocarditis due to Chikungunya virus assessed by contrast-enhanced MRI. Int. J. Cardiol. 2007;121:e7–e8. doi: 10.1016/j.ijcard.2007.04.153. [DOI] [PubMed] [Google Scholar]
- Mohd Zim M.A., Sam I.C., Omar S.F., Chan Y.F., Abubakar S., Kamarulzaman A. Chikungunya infection in Malaysia: comparison with dengue infection in adults and predictors of persistent arthralgia. J. Clin. Virol. 2013;56:141–145. doi: 10.1016/j.jcv.2012.10.019. [DOI] [PubMed] [Google Scholar]
- Moore D.L., Reddy S., Akinkugbe F.M., Lee V.H., David-West T.S., Causey O.R., Carey D.E. An epidemic of chikungunya fever at Ibadan, Nigeria, 1969. Ann. Trop. Med. Parasitol. 1974;68:59–68. doi: 10.1080/00034983.1974.11686925. [DOI] [PubMed] [Google Scholar]
- Moro M.L., Gagliotti C., Silvi G., Angelini R., Sambri V., Rezza G., Massimiliani E., Mattivi A., Grilli E., Finarelli A.C., Spataro N., Pierro A.M., Seyler T., Macini P. Chikungunya virus in North-Eastern Italy: a seroprevalence survey. Am. J. Trop. Med. Hyg. 2010;82:508–511. doi: 10.4269/ajtmh.2010.09-0322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moro M.L., Grilli E., Corvetta A., Silvi G., Angelini R., Mascella F., Miserocchi F., Sambo P., Finarelli A.C., Sambri V., Gagliotti C., Massimiliani E., Mattivi A., Pierro A.M., Macini P. Long-Term chikungunya infection clinical manifestations after an outbreak in Italy: a prognostic cohort study. J. Infect. 2012;65:165–172. doi: 10.1016/j.jinf.2012.04.005. [DOI] [PubMed] [Google Scholar]
- Morrison T.E., Oko L., Montgomery S.A., Whitmore A.C., Lotstein A.R., Gunn B.M., Elmore S.A., Heise M.T. A mouse model of chikungunya virus-induced musculoskeletal inflammatory disease: evidence of arthritis, tenosynovitis, myositis, and persistence. Am. J. Pathol. 2011;178:32–40. doi: 10.1016/j.ajpath.2010.11.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller S., Coleman J.R., Papamichail D., Ward C.B., Nimnual A., Futcher B., Skiena S., Wimmer E. Live attenuated influenza virus vaccines by computer-aided rational design. Nat. Biotechnol. 2010;28:723–726. doi: 10.1038/nbt.1636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller S., Papamichail D., Coleman J.R., Skiena S., Wimmer E. Reduction of the rate of poliovirus protein synthesis through large-scale codon deoptimization causes attenuation of viral virulence by lowering specific infectivity. J. Virol. 2006;80:9687–9696. doi: 10.1128/JVI.00738-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muthumani K., Lankaraman K.M., Laddy D.J., Sundaram S.G., Chung C.W., Sako E., Wu L., Khan A., Sardesai N., Kim J.J., Vijayachari P., Weiner D.B. Immunogenicity of novel consensus-based DNA vaccines against Chikungunya virus. Vaccine. 2008;26:5128–5134. doi: 10.1016/j.vaccine.2008.03.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muyembe-Tamfum J.J., Peyrefitte C.N., Yogolelo R., Mathina Basisya E., Koyange D., Pukuta E., Mashako M., Tolou H., Durand J.P. Epidemic of Chikungunya virus in 1999 and 200 in the Democratic Republic of the Congo. Med. Trop. (Mars) 2003;63:637–638. [PubMed] [Google Scholar]
- Nair A.G., Biswas J., Bhende M.P. A case of bilateral Chikungunya neuroretinitis. J. Ophthalmic Inflamm. Infect. 2012;2:39–40. doi: 10.1007/s12348-011-0038-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nair P.M. Chikungunya in neonates. Indian Pediatr. 2008;45:605. [PubMed] [Google Scholar]
- Nakaya H.I., Gardner J., Poo Y.S., Major L., Pulendran B., Suhrbier A. Gene profiling of Chikungunya virus arthritis in a mouse model reveals significant overlap with rheumatoid arthritis. Arthritis Rheum. 2012;64:3553–3563. doi: 10.1002/art.34631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narsimulu G., Prabhu N. Post-chikungunya chronic arthritis. J. Assoc. Physicians India. 2011;59:81. [PubMed] [Google Scholar]
- Ng L.C., Hapuarachchi H.C. Tracing the path of Chikungunya virus – evolution and adaptation. Infect. Genet. Evol. 2010;10:876–885. doi: 10.1016/j.meegid.2010.07.012. [DOI] [PubMed] [Google Scholar]
- Ng L.F., Chow A., Sun Y.J., Kwek D.J., Lim P.L., Dimatatac F., Ng L.C., Ooi E.E., Choo K.H., Her Z., Kourilsky P., Leo Y.S. IL-1beta, IL-6, and RANTES as biomarkers of Chikungunya severity. PLoS One. 2009;4:e4261. doi: 10.1371/journal.pone.0004261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng L.C., Lam S., Teo D. Epidemiology of dengue and chikungunya viruses and their potential impact on the blood supply. ISBT. Sci. Ser. 2009;4:357–367. [Google Scholar]
- Nkoghe D., Kassa R.F., Caron M., Grard G., Mombo I., Bikie B., Paupy C., Becquart P., Bisvigou U., Leroy E.M. Clinical forms of chikungunya in Gabon, 2010. PLoS Negl. Trop. Dis. 2012;6:e1517. doi: 10.1371/journal.pntd.0001517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nougairede, A., De Fabritus, L., Aubry, F., Gould, E.A., Holmes, E.C. and de Lamballerie, X., 2013. Random codon re-encoding induces stable reduction of replicative fitness of Chikungunya virus in primate and mosquito cells. PLoS pathogens 9, e1003172. [DOI] [PMC free article] [PubMed]
- Ozden S., Huerre M., Riviere J.P., Coffey L.L., Afonso P.V., Mouly V., de Monredon J., Roger J.C., El Amrani M., Yvin J.L., Jaffar M.C., Frenkiel M.P., Sourisseau M., Schwartz O., Butler-Browne G., Despres P., Gessain A., Ceccaldi P.E. Human muscle satellite cells as targets of Chikungunya virus infection. PLoS One. 2007;2:e527. doi: 10.1371/journal.pone.0000527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozden S., Lucas-Hourani M., Ceccaldi P.E., Basak A., Valentine M., Benjannet S., Hamelin J., Jacob Y., Mamchaoui K., Mouly V., Despres P., Gessain A., Butler-Browne G., Chretien M., Tangy F., Vidalain P.O., Seidah N.G. Inhibition of Chikungunya virus infection in cultured human muscle cells by furin inhibitors: impairment of the maturation of the E2 surface glycoprotein. J. Biol. Chem. 2008;283:21899–21908. doi: 10.1074/jbc.M802444200. [DOI] [PubMed] [Google Scholar]
- Paessler S., Rijnbrand R., Stein D.A., Ni H., Yun N.E., Dziuba N., Borisevich V., Seregin A., Ma Y., Blouch R., Iversen P.L., Zacks M.A. Inhibition of alphavirus infection in cell culture and in mice with antisense morpholino oligomers. Virology. 2008;376:357–370. doi: 10.1016/j.virol.2008.03.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pages F., Peyrefitte C.N., Mve M.T., Jarjaval F., Brisse S., Iteman I., Gravier P., Tolou H., Nkoghe D., Grandadam M. Aedes albopictus mosquito: the main vector of the 2007 Chikungunya outbreak in Gabon. PLoS One. 2009;4:e4691. doi: 10.1371/journal.pone.0004691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paquet C., Quatresous I., Solet J.L., Sissoko D., Renault P., Pierre V., Cordel H., Lassalle C., Thiria J., Zeller H., Schuffnecker I. Chikungunya outbreak in Reunion: epidemiology and surveillance, 2005 to early January 2006. Euro Surveill. 2006;11:E060202.3. doi: 10.2807/esw.11.05.02891-en. [DOI] [PubMed] [Google Scholar]
- Parola P., de Lamballerie X., Jourdan J., Rovery C., Vaillant V., Minodier P., Brouqui P., Flahault A., Raoult D., Charrel R.N. Novel chikungunya virus variant in travelers returning from Indian Ocean islands. Emerg. Infect. Dis. 2006;12:1493–1499. doi: 10.3201/eid1210.060610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Partidos C.D., Paykel J., Weger J., Borland E.M., Powers A.M., Seymour R., Weaver S.C., Stinchcomb D.T., Osorio J.E. Cross-protective immunity against o’nyong–nyong virus afforded by a novel recombinant chikungunya vaccine. Vaccine. 2012;30:4638–4643. doi: 10.1016/j.vaccine.2012.04.099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Partidos C.D., Weger J., Brewoo J., Seymour R., Borland E.M., Ledermann J.P., Powers A.M., Weaver S.C., Stinchcomb D.T., Osorio J.E. Probing the attenuation and protective efficacy of a candidate chikungunya virus vaccine in mice with compromised interferon (IFN) signaling. Vaccine. 2011;29:3067–3073. doi: 10.1016/j.vaccine.2011.01.076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pastorino B., Muyembe-Tamfum J.J., Bessaud M., Tock F., Tolou H., Durand J.P., Peyrefitte C.N. Epidemic resurgence of Chikungunya virus in democratic Republic of the Congo: identification of a new central African strain. J. Med. Virol. 2004;74:277–282. doi: 10.1002/jmv.20168. [DOI] [PubMed] [Google Scholar]
- Paterson H.E., McIntosh B.M. Further Studies on the Chikungunya Outbreak in Southern Rhodesia in 1962. Ii. Transmission Experiments with the Aedes Furcifer–Taylori Group of Mosquitoes and with a Member of the Anopheles Gambiae Complex. Ann. Trop. Med. Parasitol. 1964;58:52–55. doi: 10.1080/00034983.1964.11686214. [DOI] [PubMed] [Google Scholar]
- Paul S.D., Singh K.R. Experimental infection of Macaca radiata with Chikungunya virus and transmission of virus by mosquitoes. Indian J. Med. Res. 1968;56:802–811. [PubMed] [Google Scholar]
- Peiris J.S., Dittus W.P., Ratnayake C.B. Seroepidemiology of dengue and other arboviruses in a natural population of toque macaques (Macaca sinica) at Polonnaruwa, Sri Lanka. J. Med. Primatol. 1993;22:240–245. [PubMed] [Google Scholar]
- Pialoux G., Gauzere B.A., Jaureguiberry S., Strobel M. Chikungunya, an epidemic arbovirosis. Lancet Infect. Dis. 2007;7:319–327. doi: 10.1016/S1473-3099(07)70107-X. [DOI] [PubMed] [Google Scholar]
- Pijlman, G.P., 2012. Chikungunya virus nsP2 and nsP3 differentially anagonize host responses. In: 11th ARA, 10th MCAA, Symposium, vol. 1.
- Pinto A.J., Morahan P.S., Brinton M., Stewart D., Gavin E. Comparative therapeutic efficacy of recombinant interferons-alpha, -beta, and -gamma against alphatogavirus, bunyavirus, flavivirus, and herpesvirus infections. J. Interferon Res. 1990;10:293–298. doi: 10.1089/jir.1990.10.293. [DOI] [PubMed] [Google Scholar]
- Pinto M.R., Filipe A.R. Arbovirus studies in Luanda, Angola. 1. Virological and serological studies during a yellow fever epidemic. Bull. World Health Organ. 1973;49:31–35. [PMC free article] [PubMed] [Google Scholar]
- Plante K., Wang E., Partidos C.D., Weger J., Gorchakov R., Tsetsarkin K., Borland E.M., Powers A.M., Seymour R., Stinchcomb D.T., Osorio J.E., Frolov I., Weaver S.C. Novel chikungunya vaccine candidate with an IRES-based attenuation and host range alteration mechanism. PLoS Pathog. 2011;7:e1002142. doi: 10.1371/journal.ppat.1002142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porter K.R., Tan R., Istary Y., Suharyono W., Sutaryo, Widjaja S., Ma’Roef C., Listiyaningsih E., Kosasih H., Hueston L., McArdle J., Juffrie M. A serological study of Chikungunya virus transmission in Yogyakarta, Indonesia: evidence for the first outbreak since 1982. Southeast Asian J. Trop. Med. Public Health. 2004;35:408–415. [PubMed] [Google Scholar]
- Powers A.M., Brault A.C., Tesh R.B., Weaver S.C. Re-emergence of Chikungunya and O’nyong–nyong viruses: evidence for distinct geographical lineages and distant evolutionary relationships. J. Gen. Virol. 2000;81:471–479. doi: 10.1099/0022-1317-81-2-471. [DOI] [PubMed] [Google Scholar]
- Powers A.M., Logue C.H. Changing patterns of chikungunya virus: re-emergence of a zoonotic arbovirus. J. Gen. Virol. 2007;88:2363–2377. doi: 10.1099/vir.0.82858-0. [DOI] [PubMed] [Google Scholar]
- Pulmanausahakul R., Roytrakul S., Auewarakul P., Smith D.R. Chikungunya in Southeast Asia: understanding the emergence and finding solutions. Int. J. Infect. Dis. 2011;15:e671–e676. doi: 10.1016/j.ijid.2011.06.002. [DOI] [PubMed] [Google Scholar]
- Queyriaux B., Simon F., Grandadam M., Michel R., Tolou H., Boutin J.P. Clinical burden of chikungunya virus infection. Lancet Infect. Dis. 2008;8:2–3. doi: 10.1016/S1473-3099(07)70294-3. [DOI] [PubMed] [Google Scholar]
- Rajapakse S., Rodrigo C., Rajapakse A. Atypical manifestations of chikungunya infection. Trans. R. Soc. Trop. Med. Hyg. 2010;104:89–96. doi: 10.1016/j.trstmh.2009.07.031. [DOI] [PubMed] [Google Scholar]
- Ramful D., Carbonnier M., Pasquet M., Bouhmani B., Ghazouani J., Noormahomed T., Beullier G., Attali T., Samperiz S., Fourmaintraux A., Alessandri J.L. Mother-to-child transmission of Chikungunya virus infection. Pediatr. Infect. Dis. J. 2007;26:811–815. doi: 10.1097/INF.0b013e3180616d4f. [DOI] [PubMed] [Google Scholar]
- Randall R.E., Goodbourn S. Interferons and viruses: an interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008;89:1–47. doi: 10.1099/vir.0.83391-0. [DOI] [PubMed] [Google Scholar]
- Rao G., Khan Y.Z., Chitnis D.S. Chikungunya infection in neonates. Indian Pediatr. 2008;45:240–242. [PubMed] [Google Scholar]
- Ravichandran R., Manian M. Ribavirin therapy for Chikungunya arthritis. J. Infect. Dev. Ctries. 2008;2:140–142. [PubMed] [Google Scholar]
- Renault P., Balleydier E., D’Ortenzio E., Baville M., Filleul L. Epidemiology of chikungunya infection on Reunion Island, Mayotte, and neighboring countries. Med. Mal. Infect. 2012;42:93–101. doi: 10.1016/j.medmal.2011.12.002. [DOI] [PubMed] [Google Scholar]
- Renault P., Solet J.L., Sissoko D., Balleydier E., Larrieu S., Filleul L., Lassalle C., Thiria J., Rachou E., de Valk H., Ilef D., Ledrans M., Quatresous I., Quenel P., Pierre V. A major epidemic of chikungunya virus infection on Reunion Island, France, 2005–2006. Am. J. Trop. Med. Hyg. 2007;77:727–731. [PubMed] [Google Scholar]
- Rezza G., Nicoletti L., Angelini R., Romi R., Finarelli A.C., Panning M., Cordioli P., Fortuna C., Boros S., Magurano F., Silvi G., Angelini P., Dottori M., Ciufolini M.G., Majori G.C., Cassone A. Infection with chikungunya virus in Italy: an outbreak in a temperate region. Lancet. 2007;370:1840–1846. doi: 10.1016/S0140-6736(07)61779-6. [DOI] [PubMed] [Google Scholar]
- Ribera A., Degasne I., Jaffar Bandjee M.C., Gasque P. [Chronic rheumatic manifestations following chikungunya virus infection: clinical description and therapeutic considerations] Med. Trop. (Mars) 2012 72 Spec No, 83-5. [PubMed] [Google Scholar]
- Rinaldo C.R., Jr., Overall J.C., Jr., Glasgow L.A. Viral replication and interferon production in fetal and adult ovine leukocytes and spleen cells. Infect. Immun. 1975;12:1070–1077. doi: 10.1128/iai.12.5.1070-1077.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riyaz N., Riyaz A., Abdul Latheef E.N., Anitha P.M., Aravindan K.P., Nair A.S., Shameera P. Cutaneous manifestations of chikungunya during a recent epidemic in Calicut, north Kerala, south India. Indian J. Dermatol. Venereol. Leprol. 2010;76:671–676. doi: 10.4103/0378-6323.72466. [DOI] [PubMed] [Google Scholar]
- Robert E., Johnston R.E., Peters C.J. Alphaviruses. In: Fields B.N., Knipe D.M., Howley P.M., editors. Vol. 1. Lippincott-Raven Publishers; 1996. pp. 843–898. (Fields virology). [Google Scholar]
- Robin S., Ramful D., Le Seach F., Jaffar-Bandjee M.C., Rigou G., Alessandri J.L. Neurologic manifestations of pediatric chikungunya infection. J. Child. Neurol. 2008;23:1028–1035. doi: 10.1177/0883073808314151. [DOI] [PubMed] [Google Scholar]
- Robin S., Ramful D., Zettor J., Benhamou L., Jaffar-Bandjee M.C., Riviere J.P., Marichy J., Ezzedine K., Alessandri J.L. Severe bullous skin lesions associated with Chikungunya virus infection in small infants. Eur. J. Pediatr. 2010;169:67–72. doi: 10.1007/s00431-009-0986-0. [DOI] [PubMed] [Google Scholar]
- Robinson M.C. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952–53. I. Clinical features. Trans. R. Soc. Trop. Med. Hyg. 1955;49:28–32. doi: 10.1016/0035-9203(55)90080-8. [DOI] [PubMed] [Google Scholar]
- Roche S., Robin Y. Human infections by Chikungunya virus in Rufisque (Senegal), October–November, 1966. Bull. Soc. Med. Afr. Noire Lang. Fr. 1967;12:490–496. [PubMed] [Google Scholar]
- Roques P., Gras G., Labadie K., Larcher T., Cherel Y., Suhrbier A., Le Grand R. Chikungunya virus infection involved monocytes and during chronic phase of the disease persisted in tissue macrophages. 45th Annual Scientific Meeting of the European Society for Clinical Investigation. Crete, Greece, 13–16 April 2011. Eur. J. Clin. Invest. 2011;41:23–52. [Google Scholar]
- Rose N., Anoop T.M., John A.P., Jabbar P.K., George K.C. Acute optic neuritis following infection with chikungunya virus in southern rural India. Int. J. Infect. Dis. 2011;15:e147–e150. doi: 10.1016/j.ijid.2010.10.004. [DOI] [PubMed] [Google Scholar]
- Ross R.W. The Newala epidemic. III. The virus: isolation, pathogenic properties and relationship to the epidemic. J. Hyg. (Lond.) 1956;54:177–191. doi: 10.1017/s0022172400044442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roussel A., Lescar J., Vaney M.C., Wengler G., Wengler G., Rey F.A. Structure and interactions at the viral surface of the envelope protein E1 of Semliki Forest virus. Structure. 2006;14:75–86. doi: 10.1016/j.str.2005.09.014. [DOI] [PubMed] [Google Scholar]
- Rudd P.A., Wilson J., Gardner J., Larcher T., Babarit C., Le T.T., Anraku I., Kumagai Y., Loo Y.M., Gale M., Jr., Akira S., Khromykh A.A., Suhrbier A. Interferon response factors 3 and 7 protect against chikungunya virus hemorrhagic fever and shock. J. Virol. 2012;86:9888–9898. doi: 10.1128/JVI.00956-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rulli N.E., Guglielmotti A., Mangano G., Rolph M.S., Apicella C., Zaid A., Suhrbier A., Mahalingam S. Amelioration of alphavirus-induced arthritis and myositis in a mouse model by treatment with bindarit, an inhibitor of monocyte chemotactic proteins. Arthritis Rheum. 2009;60:2513–2523. doi: 10.1002/art.24682. [DOI] [PubMed] [Google Scholar]
- Rulli N.E., Rolph M.S., Srikiatkhachorn A., Anantapreecha S., Guglielmotti A., Mahalingam S. Protection from arthritis and myositis in a mouse model of acute chikungunya virus disease by bindarit, an inhibitor of monocyte chemotactic protein-1 synthesis. J. Infect. Dis. 2011;204:1026–1030. doi: 10.1093/infdis/jir470. [DOI] [PubMed] [Google Scholar]
- Russo A.T., White M.A., Watowich S.J. The crystal structure of the Venezuelan equine encephalitis alphavirus nsP2 protease. Structure. 2006;14:1449–1458. doi: 10.1016/j.str.2006.07.010. [DOI] [PubMed] [Google Scholar]
- Saluzzo J.F., Gonzalez J.P., Herve J.P., Georges A.J. Epidemiological study of arboviruses in the Central African Republic: demonstration of Chikungunya virus during 1978 and 1979. Bull. Soc. Pathol. Exot. Filiales. 1980;73:390–399. [PubMed] [Google Scholar]
- Saluzzo J.F., Gonzalez J.P., Herve J.P., Georges A.J. Serological survey for the prevalence of certain arboviruses in the human population of the south-east area of Central African Republic (author’s transl) Bull. Soc. Pathol. Exot. Filiales. 1981;74:490–499. [PubMed] [Google Scholar]
- Saluzzo J.F., Ivanoff B., Languillat G., Georges A.J. Serological survey for arbovirus antibodies in the human and simian populations of the South-East of Gabon (author’s transl) Bull. Soc. Pathol. Exot. Filiales. 1982;75:262–266. [PubMed] [Google Scholar]
- Sarkar J.K. Calcutta’s experience and findings in haemorrhagic fever and Chikungunya fever epidemic. Jpn. J. Med. Sci. Biol. 1967;20(Suppl.):88–90. [PubMed] [Google Scholar]
- Savarino A., Boelaert J.R., Cassone A., Majori G., Cauda R. Effects of chloroquine on viral infections: an old drug against today’s diseases? Lancet Infect. Dis. 2003;3:722–727. doi: 10.1016/S1473-3099(03)00806-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schilte C., Buckwalter M.R., Laird M.E., Diamond M.S., Schwartz O., Albert M.L. Cutting edge: independent roles for IRF-3 and IRF-7 in hematopoietic and nonhematopoietic cells during host response to Chikungunya infection. J. Immunol. 2012;188:2967–2971. doi: 10.4049/jimmunol.1103185. [DOI] [PubMed] [Google Scholar]
- Schilte C., Couderc T., Chretien F., Sourisseau M., Gangneux N., Guivel-Benhassine F., Kraxner A., Tschopp J., Higgs S., Michault A., Arenzana-Seisdedos F., Colonna M., Peduto L., Schwartz O., Lecuit M., Albert M.L. Type I IFN controls chikungunya virus via its action on nonhematopoietic cells. J. Exp. Med. 2010;207:429–442. doi: 10.1084/jem.20090851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuffenecker I., Iteman I., Michault A., Murri S., Frangeul L., Vaney M.C., Lavenir R., Pardigon N., Reynes J.M., Pettinelli F., Biscornet L., Diancourt L., Michel S., Duquerroy S., Guigon G., Frenkiel M.P., Brehin A.C., Cubito N., Despres P., Kunst F., Rey F.A., Zeller H., Brisse S. Genome microevolution of chikungunya viruses causing the Indian Ocean outbreak. PLoS Med. 2006;3:e263. doi: 10.1371/journal.pmed.0030263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sebastian M.R., Lodha R., Kabra S.K. Chikungunya infection in children. Indian J. Pediatr. 2009;76:185–189. doi: 10.1007/s12098-009-0049-6. [DOI] [PubMed] [Google Scholar]
- Sergon K., Njuguna C., Kalani R., Ofula V., Onyango C., Konongoi L.S., Bedno S., Burke H., Dumilla A.M., Konde J., Njenga M.K., Sang R., Breiman R.F. Seroprevalence of Chikungunya virus (CHIKV) infection on Lamu Island, Kenya, October 2004. Am. J. Trop. Med. Hyg. 2008;78:333–337. [PubMed] [Google Scholar]
- Sergon K., Yahaya A.A., Brown J., Bedja S.A., Mlindasse M., Agata N., Allaranger Y., Ball M.D., Powers A.M., Ofula V., Onyango C., Konongoi L.S., Sang R., Njenga M.K., Breiman R.F. Seroprevalence of Chikungunya virus infection on Grande Comore Island, union of the Comoros, 2005. Am. J. Trop. Med. Hyg. 2007;76:1189–1193. [PubMed] [Google Scholar]
- Shah K.V., Gilotra S.K., Gibbs C.J., Jr., Rozeboom L.E. Laboratory studies of transmission of chikungunya virus by mosquitoes: a preliminary report. Indian J. Med. Res. 1964;52:703–709. [PubMed] [Google Scholar]
- Simizu B., Yamamoto K., Hashimoto K., Ogata T. Structural proteins of Chikungunya virus. J. Virol. 1984;51:254–258. doi: 10.1128/jvi.51.1.254-258.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon F., Javelle E., Oliver M., Leparc-Goffart I., Marimoutou C. Chikungunya virus infection. Curr. Infect. Dis. Rep. 2011;13:218–228. doi: 10.1007/s11908-011-0180-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon F., Parola P., Grandadam M., Fourcade S., Oliver M., Brouqui P., Hance P., Kraemer P., Ali Mohamed A., de Lamballerie X., Charrel R., Tolou H. Chikungunya infection: an emerging rheumatism among travelers returned from Indian Ocean islands. Report of 47 cases. Medicine (Baltimore) 2007;86:123–137. doi: 10.1097/MD/0b013e31806010a5. [DOI] [PubMed] [Google Scholar]
- Simon F., Paule P., Oliver M. Chikungunya virus-induced myopericarditis: toward an increase of dilated cardiomyopathy in countries with epidemics? Am. J. Trop. Med. Hyg. 2008;78:212–213. [PubMed] [Google Scholar]
- Singh Kh.D., Kirubakaran P., Nagarajan S., Sakkiah S., Muthusamy K., Velmurgan D., Jeyakanthan J. Homology modeling, molecular dynamics, e-pharmacophore mapping and docking study of Chikungunya virus nsP2 protease. J. Mol. Model. 2012;18:39–51. doi: 10.1007/s00894-011-1018-3. [DOI] [PubMed] [Google Scholar]
- Singh S.K., Unni S.K. Chikungunya virus: host pathogen interaction. Rev. Med. Virol. 2011;21:78–88. doi: 10.1002/rmv.681. [DOI] [PubMed] [Google Scholar]
- Sissoko D., Ezzedine K., Moendandze A., Giry C., Renault P., Malvy D. Field evaluation of clinical features during chikungunya outbreak in Mayotte, 2005–2006. Trop. Med. Int. Health. 2010;15:600–607. doi: 10.1111/j.1365-3156.2010.02485.x. [DOI] [PubMed] [Google Scholar]
- Sissoko D., Malvy D., Ezzedine K., Renault P., Moscetti F., Ledrans M., Pierre V. Post-epidemic Chikungunya disease on Reunion Island: course of rheumatic manifestations and associated factors over a 15-month period. PLoS Negl. Trop. Dis. 2009;3:e389. doi: 10.1371/journal.pntd.0000389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sissoko D., Moendandze A., Malvy D., Giry C., Ezzedine K., Solet J.L., Pierre V. Seroprevalence and risk factors of chikungunya virus infection in Mayotte, Indian Ocean, 2005–2006: a population-based survey. PLoS One. 2008;3:e3066. doi: 10.1371/journal.pone.0003066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smee D.F., Alaghamandan H.A., Kini G.D., Robins R.K. Antiviral activity and mode of action of ribavirin 5′-sulfamate against Semliki Forest virus. Antiviral Res. 1988;10:253–262. doi: 10.1016/0166-3542(88)90044-7. [DOI] [PubMed] [Google Scholar]
- Solignat M., Gay B., Higgs S., Briant L., Devaux C. Replication cycle of chikungunya: a re-emerging arbovirus. Virology. 2009;393:183–197. doi: 10.1016/j.virol.2009.07.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Y., Liu Y., Ward C.B., Mueller S., Futcher B., Skiena S., Paul A.V., Wimmer E. Identification of two functionally redundant RNA elements in the coding sequence of poliovirus using computer-generated design. Proc. Natl. Acad. Sci. U.S.A. 2012;109:14301–14307. doi: 10.1073/pnas.1211484109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sourisseau M., Schilte C., Casartelli N., Trouillet C., Guivel-Benhassine F., Rudnicka D., Sol-Foulon N., Le Roux K., Prevost M.C., Fsihi H., Frenkiel M.P., Blanchet F., Afonso P.V., Ceccaldi P.E., Ozden S., Gessain A., Schuffenecker I., Verhasselt B., Zamborlini A., Saib A., Rey F.A., Arenzana-Seisdedos F., Despres P., Michault A., Albert M.L., Schwartz O. Characterization of reemerging chikungunya virus. PLoS Pathog. 2007;3:e89. doi: 10.1371/journal.ppat.0030089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sourisseau M., Schilte C., Casartelli N., Trouillet C., Guivel-Benhassine F., Rudnicka D., Sol-Foulon N., Roux K.L., Prevost M.C., Fsihi H., Frenkiel M.P., Blanchet F., Afonso P.V., Ceccaldi P.E., Ozden S., Gessain A., Schuffenecker I., Verhasselt B., Zamborlini A., Saib A., Rey F.A., Arenzana-Seisdedos F., Despres P., Michault A., Albert M.L., Schwartz O. Characterization of reemerging Chikungunya virus. PLoS Pathog. 2007;3:e89. doi: 10.1371/journal.ppat.0030089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staikowsky F., Talarmin F., Grivard P., Souab A., Schuffenecker I., Le Roux K., Lecuit M., Michault A. Prospective study of Chikungunya virus acute infection in the Island of La Reunion during the 2005–2006 outbreak. PLoS One. 2009;4:e7603. doi: 10.1371/journal.pone.0007603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stein D.A. Inhibition of RNA virus infections with peptide-conjugated morpholino oligomers. Curr. Pharm. Des. 2008;14:2619–2634. doi: 10.2174/138161208786071290. [DOI] [PubMed] [Google Scholar]
- Taubitz W., Cramer J.P., Kapaun A., Pfeffer M., Drosten C., Dobler G., Burchard G.D., Loscher T. Chikungunya fever in travelers: clinical presentation and course. Clin. Infect. Dis. 2007;45:e1–e4. doi: 10.1086/518701. [DOI] [PubMed] [Google Scholar]
- Teng T.S., Foo S.S., Simamarta D., Lum F.M., Teo T.H., Lulla A., Yeo N.K., Koh E.G., Chow A., Leo Y.S., Merits A., Chin K.C., Ng L.F. Viperin restricts chikungunya virus replication and pathology. J. Clin. Invest. 2012;122:4447–4460. doi: 10.1172/JCI63120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teo T.H., Lum F.M., Lee W.W., Ng L.F. Mouse models for Chikungunya virus: deciphering immune mechanisms responsible for disease and pathology. Immunol. Res. 2012;53:136–147. doi: 10.1007/s12026-012-8266-x. [DOI] [PubMed] [Google Scholar]
- Thiberville S.D., Boisson V., Gaudart J., Simon F., Flahault A., de Lamballerie X. Chikungunya Fever: a clinical and virological investigation of outpatients on reunion island, South-west Indian ocean. PLoS Negl. Trop. Dis. 2013;7:e2004. doi: 10.1371/journal.pntd.0002004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thonnon J., Spiegel A., Diallo M., Diallo A., Fontenille D. Chikungunya virus outbreak in Senegal in 1996 and 1997. Bull. Soc. Pathol. Exot. 1999;92:79–82. [PubMed] [Google Scholar]
- Tilston N., Skelly C., Weinstein P. Pan-European Chikungunya surveillance: designing risk stratified surveillance zones. Int. J. Health Geogr. 2009;8:61. doi: 10.1186/1476-072X-8-61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomori O., Fagbami A., Fabiyi A. The 1974 epidemic of Chikungunya fever in children in Ibadan. Trop. Geogr. Med. 1975;27:413–417. [PubMed] [Google Scholar]
- Tsetsarkin K.A., Chen R., Leal G., Forrester N., Higgs S., Huang J., Weaver S.C. Chikungunya virus emergence is constrained in Asia by lineage-specific adaptive landscapes. Proc. Natl. Acad. Sci. U.S.A. 2011;108:7872–7877. doi: 10.1073/pnas.1018344108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsetsarkin K.A., Chen R., Sherman M.B., Weaver S.C. Chikungunya virus: evolution and genetic determinants of emergence. Curr. Opin. Virol. 2011;1:310–317. doi: 10.1016/j.coviro.2011.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsetsarkin K.A., McGee C.E., Volk S.M., Vanlandingham D.L., Weaver S.C., Higgs S. Epistatic roles of E2 glycoprotein mutations in adaption of chikungunya virus to Aedes albopictus and Ae. aegypti mosquitoes. PLoS One. 2009;4:e6835. doi: 10.1371/journal.pone.0006835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsetsarkin K.A., Vanlandingham D.L., McGee C.E., Higgs S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007;3:e201. doi: 10.1371/journal.ppat.0030201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsetsarkin K.A., Weaver S.C. Sequential adaptive mutations enhance efficient vector switching by Chikungunya virus and its epidemic emergence. PLoS Pathog. 2011;7:e1002412. doi: 10.1371/journal.ppat.1002412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turell M.J., Beaman J.R. Experimental transmission of Venezuelan equine encephalomyelitis virus by a strain of Aedes albopictus (Diptera: Culicidae) from New Orleans, Louisiana. J. Med. Entomol. 1992;29:802–805. doi: 10.1093/jmedent/29.5.802. [DOI] [PubMed] [Google Scholar]
- Turell M.J., Malinoski F.J. Limited potential for mosquito transmission of a live, attenuated chikungunya virus vaccine. Am. J. Trop. Med. Hyg. 1992;47:98–103. doi: 10.4269/ajtmh.1992.47.98. [DOI] [PubMed] [Google Scholar]
- Valamparampil J.J., Chirakkarot S., Letha S., Jayakumar C., Gopinathan K.M. Clinical profile of Chikungunya in infants. Indian J. Pediatr. 2009;76:151–155. doi: 10.1007/s12098-009-0045-x. [DOI] [PubMed] [Google Scholar]
- Vanlandingham D.L., Tsetsarkin K., Klingler K.A., Hong C., McElroy K.L., Lehane M.J., Higgs S. Determinants of vector specificity of o’nyong nyong and chikungunya viruses in Anopheles and Aedes mosquitoes. Am. J. Trop. Med. Hyg. 2006;74:663–669. [PubMed] [Google Scholar]
- Vazeille M., Moutailler S., Coudrier D., Rousseaux C., Khun H., Huerre M., Thiria J., Dehecq J.S., Fontenille D., Schuffenecker I., Despres P., Failloux A.B. Two Chikungunya isolates from the outbreak of La Reunion (Indian Ocean) exhibit different patterns of infection in the mosquito, Aedes albopictus. PLoS One. 2007;2:e1168. doi: 10.1371/journal.pone.0001168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vega-Rua A., Zouache K., Caro V., Diancourt L., Delaunay P., Grandadam M., Failloux A.B. High efficiency of temperate Aedes albopictus to transmit chikungunya and dengue viruses in the Southeast of France. PLoS One. 2013;8:e59716. doi: 10.1371/journal.pone.0059716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vesenjak-Hirjan J., Hermon Y., Vitarana T. Arbovirus infections in Ceylon. Bull. World Health Organ. 1969;41:243–249. [PMC free article] [PubMed] [Google Scholar]
- Vierboom M.P., Jonker M., Tak P.P., t Hart B.A. Preclinical models of arthritic disease in non-human primates. Drug. Discov. Today. 2007;12:327–335. doi: 10.1016/j.drudis.2007.02.012. [DOI] [PubMed] [Google Scholar]
- Vilain P., Larrieu S., Renault P., Baville M., Filleul L. How to explain the re-emergence of chikungunya infection in Reunion Island in 2010? Acta Trop. 2012;123:85–90. doi: 10.1016/j.actatropica.2012.03.009. [DOI] [PubMed] [Google Scholar]
- Volk S.M., Chen R., Tsetsarkin K.A., Adams A.P., Garcia T.I., Sall A.A., Nasar F., Schuh A.J., Holmes E.C., Higgs S., Maharaj P.D., Brault A.C., Weaver S.C. Genome-scale phylogenetic analyses of chikungunya virus reveal independent emergences of recent epidemics and various evolutionary rates. J. Virol. 2010;84:6497–6504. doi: 10.1128/JVI.01603-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voss J.E., Vaney M.C., Duquerroy S., Vonrhein C., Girard-Blanc C., Crublet E., Thompson A., Bricogne G., Rey F.A. Glycoprotein organization of Chikungunya virus particles revealed by X-ray crystallography. Nature. 2010;468:709–712. doi: 10.1038/nature09555. [DOI] [PubMed] [Google Scholar]
- Vu Qui D., Nguyen-Thi K.-T. Hemorrhagic fever in Vietnam in 1964–1965. Serologic study with a brief clinica and epidemiologic note. Bull. Soc. Pathol. Exot. Filiales. 1967;60:21–33. [PubMed] [Google Scholar]
- Walsh G.P., Tan E.V., dela Cruz E.C., Abalos R.M., Villahermosa L.G., Young L.J., Cellona R.V., Nazareno J.B., Orwitz M.A. The Philippine cynomolgus monkey (Macaca fasicularis) provides a new nonhuman primate model of tuberculosis that resembles human disease. Nat. Med. 1996;2:430–436. doi: 10.1038/nm0496-430. [DOI] [PubMed] [Google Scholar]
- Wang B.X., Fish E.N. The yin and yang of viruses and interferons. Trends Immunol. 2012;33:190–197. doi: 10.1016/j.it.2012.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang D., Suhrbier A., Penn-Nicholson A., Woraratanadharm J., Gardner J., Luo M., Le T.T., Anraku I., Sakalian M., Einfeld D., Dong J.Y. A complex adenovirus vaccine against chikungunya virus provides complete protection against viraemia and arthritis. Vaccine. 2011;29:2803–2809. doi: 10.1016/j.vaccine.2011.01.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang E., Kim D.Y., Weaver S.C., Frolov I. Chimeric Chikungunya viruses are nonpathogenic in highly sensitive mouse models but efficiently induce a protective immune response. J. Virol. 2011;85:9249–9252. doi: 10.1128/JVI.00844-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warter L., Lee C.Y., Thiagarajan R., Grandadam M., Lebecque S., Lin R.T., Bertin-Maghit S., Ng L.F., Abastado J.P., Despres P., Wang C.I., Nardin A. Chikungunya virus envelope-specific human monoclonal antibodies with broad neutralization potency. J. Immunol. 2011;186:3258–3264. doi: 10.4049/jimmunol.1003139. [DOI] [PubMed] [Google Scholar]
- Watts D.M., el-Tigani A., Botros B.A., Salib A.W., Olson J.G., McCarthy M., Ksiazek T.G. Arthropod-borne viral infections associated with a fever outbreak in the northern province of Sudan. J. Trop. Med. Hyg. 1994;97:228–230. [PubMed] [Google Scholar]
- Weinbren M.P. The occurrence of Chikungunya virus in Uganda. II. In man on the Entebbe peninsula. Trans. R. Soc. Trop. Med. Hyg. 1958;52:258–259. [PubMed] [Google Scholar]
- Weinbren M.P., Haddow A.J., Williams M.C. The occurrence of Chikungunya virus in Uganda. I. Isolation from mosquitoes. Trans. R. Soc. Trop. Med. Hyg. 1958;52:253–257. doi: 10.1016/0035-9203(58)90084-1. [DOI] [PubMed] [Google Scholar]
- Werneke S.W., Schilte C., Rohatgi A., Monte K.J., Michault A., Arenzana-Seisdedos F., Vanlandingham D.L., Higgs S., Fontanet A., Albert M.L., Lenschow D.J. ISG15 is critical in the control of Chikungunya virus infection independent of UbE1L mediated conjugation. PLoS Pathog. 2011;7:e1002322. doi: 10.1371/journal.ppat.1002322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White A., Berman S., Lowenthal J.P. Comparative immunogenicities of Chikungunya vaccines propagated in monkey kidney monolayers and chick embryo suspension cultures. Appl. Microbiol. 1972;23:951–952. doi: 10.1128/am.23.5.951-952.1972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO Outbreak news. Chikungunya and dengue, south-west Indian Ocean. Wkly. Epidemiol. Rec. 2006;81:106–108. [PubMed] [Google Scholar]
- WHO Outbreak and spread of chikungunya. Wkly. Epidemiol. Rec. 2007;82:409–415. [PubMed] [Google Scholar]
- Win M.K., Chow A., Dimatatac F., Go C.J., Leo Y.S. Chikungunya fever in Singapore: acute clinical and laboratory features, and factors associated with persistent arthralgia. J. Clin. Virol. 2010;49:111–114. doi: 10.1016/j.jcv.2010.07.004. [DOI] [PubMed] [Google Scholar]
- Wolfe N.D., Kilbourn A.M., Karesh W.B., Rahman H.A., Bosi E.J., Cropp B.C., Andau M., Spielman A., Gubler D.J. Sylvatic transmission of arboviruses among Bornean orangutans. Am. J. Trop. Med. Hyg. 2001;64:310–316. doi: 10.4269/ajtmh.2001.64.310. [DOI] [PubMed] [Google Scholar]
- Wu D., Wu J., Zhang Q., Zhong H., Ke C., Deng X., Guan D., Li H., Zhang Y., Zhou H., He J., Li L., Yang X. Chikungunya outbreak in Guangdong province, China, 2010. Emerg. Infect. Dis. 2012;18:493–495. doi: 10.3201/eid1803.110034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu C., Guo T.C., Mutoloki S., Haugland O., Marjara I.S., Evensen O. Alpha interferon and not gamma interferon inhibits salmonid alphavirus subtype 3 replication in vitro. J. Virol. 2010;84:8903–8912. doi: 10.1128/JVI.00851-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yadav P., Gokhale M.D., Barde P.V., Singh D.K., Mishra A.C., Mourya D.T. Experimental transmission of Chikungunya virus by Anopheles stephensi mosquitoes. Acta Virol. 2003;47:45–47. [PubMed] [Google Scholar]
- Yoosuf A.A., Shiham I., Mohamed A.J., Ali G., Luna J.M., Pandav R., Gongal G.N., Nisaluk A., Jarman R.G., Gibbons R.V. First report of chikungunya from the Maldives. Trans. R. Soc. Trop. Med. Hyg. 2009;103:192–196. doi: 10.1016/j.trstmh.2008.09.006. [DOI] [PubMed] [Google Scholar]
- Zaid A., Rulli N.E., Rolph M.S., Suhrbier A., Mahalingam S. Disease exacerbation by etanercept in a mouse model of alphaviral arthritis and myositis. Arthritis Rheum. 2011;63:488–491. doi: 10.1002/art.30112. [DOI] [PubMed] [Google Scholar]
- Zhang R., Hryc C.F., Cong Y., Liu X., Jakana J., Gorchakov R., Baker M.L., Weaver S.C., Chiu W. 4.4 a cryo-EM structure of an enveloped alphavirus Venezuelan equine encephalitis virus. EMBO J. 2011;30:3854–3863. doi: 10.1038/emboj.2011.261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ziegler S.A., Lu L., da Rosa A.P., Xiao S.Y., Tesh R.B. An animal model for studying the pathogenesis of chikungunya virus infection. Am. J. Trop. Med. Hyg. 2008;79:133–139. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Epidemiological record and outbreak bulletin of chikungunya disease from the World Health Organisation (WHO). The weekly epidemiological records are available every Friday on the website: http://www.who.int/wer/en/. The outbreak bulletins of the WHO Regional Office for Africa are available on the website http://www.afro.who.int. We present here all epidemiological records and outbreak bulletins that include the keyword “chikungunya”.
ProMED-Mail alerts of chikungunya disease. Alerts are available on the website: http://www.promedmail.org. ProMED (Program for Monitoring Emerging Diseases) is an internet-based reporting system dedicated to rapid global dissemination of information on infectious disease outbreaks. Sources of information include media reports, official reports, online summaries, local observations, and others. A team of expert moderators screens, reviews and investigates reports before posting to the network. ProMED-mail currently reaches over 60,000 subscribers in at least 185 countries. This table summarises ProMED-mail alerts relating to chikungunya disease.
CHIKV sequences available from the NIH GenBank genetic sequence database. GenBank is a comprehensive database that contains publicly available nucleotide sequences for more than 380,000 organisms named at the genus level or lower, obtained primarily through submissions from individual laboratories and batch submissions from large-scale sequencing projects. GenBank is built and distributed by the National centre for Biotechnology Information, a division of the National Library of Medicine, located on the campus of the US National Institutes of Health in Bethesda, MD, USA. We summarise here the GenBank sequences of CHIKV available on the website: http://www.ncbi.nlm.nih.gov/nuccore and with nucleotide size equal or higher than 200 bp.
PubMed references of chikungunya cases. PubMed is a free database accessing primarily the MEDLINE database of references and abstracts on life sciences and biomedical topics. This database is maintained by the United States National Library of Medicine at the National Institutes of Health in Bethesda, MD, USA. We summarise here the main publications that report outbreaks or imported cases of chikungunya disease referenced on the website: http://www.ncbi.nlm.nih.gov/pubmed.
PubMed references of seroprevalence studies of chikungunya virus. The table summarises studies that report serologic surveys (IgG) within human populations and referenced on NCBI, available on the website: http://www.ncbi.nlm.nih.gov/pubmed.
Summary of the Pubmed references, WHO publications, GenBank sequences and ProMED alerts for chikungunya virus. We summarise here (i) PubMed references [excluding imported cases], (ii) WHO publications [excluding imported cases], (iii) GenBank sequences and (iv) ProMED-mail alerts, relevant to chikungunya virus, described in Supplemental Tables 1–5.