

Abstract
A significant consequence of equine arteritis virus (EAV) infection of horses is persistence of the virus in a variable percentage of infected stallions. We recently established an in vitro model of EAV persistence in cell culture for the purpose of furthering our understanding of EAV biology in general and viral persistence in the stallion in particular. In this study we investigated whether persistently infected HeLa cells could be cured of EAV infection by treatment with an antisense peptide-conjugated phosphorodiamidate morpholino oligomer (PPMO) designed to target the 5′-terminal region of the EAV genome. We found that persistently infected HeLa cells passaged three times in the presence of 5–10 μM EAV-specific PPMO produced no detectable virus. The PPMO-cured HeLa cells were free of infectious virus, viral antigen and EAV RNA as measured by plaque assay, indirect immunofluorescence assay and RT-PCR, respectively. Furthermore, when re-challenged with EAV at several passages after discontinuation of PPMO treatments, PPMO-cured HeLa cells were found to be refractory to re-infection and to the re-establishment of viral persistence. While these findings demonstrate that PPMO can be used to eliminate persistent EAV infection in cell culture, the efficacy of PPMO against EAV in vivo remains to be addressed.
Keywords: Equine arteritis virus, Persistent infection, Antisense, Antiviral, Morpholino oligomer, PPMO
Equine arteritis virus (EAV) is an enveloped virus with a single-stranded, positive-sense RNA genome of approximately 12.7 kb and belongs to the family Arteriviridae in the order Nidovirales (Cavanagh, 1997, Snijder and Meulenberg, 1998). The genomic RNA comprises nine known open reading frames (ORFs) flanked by 5′- and 3′-untranslated regions (UTRs) (Snijder and Meulenberg, 1998, Snijder et al., 1999). The two most 5′-proximal ORFs (1a and 1b) encode two replicase polyproteins (pp1a and pp1ab) and the remaining seven ORFs (2a, 2b, and 3-7) located in the 3′-proximal one-quarter of the genome encode the envelope proteins E, GP2, GP3, GP4, GP5, M, and the nucleocapsid protein (N), respectively (Snijder and Spaan, 2006).
EAV is the causative agent of equine viral arteritis (EVA) and is widely distributed in equine populations throughout the world (Timoney and McCollum, 1993). A variable percentage (10–70%) of stallions acutely infected with EAV can become persistently infected carriers, shedding the virus constantly in semen (Timoney and McCollum, 1993). Persistently infected stallions are the principal reservoir of EAV and are responsible for perpetuation and dissemination of EAV in equine populations (Timoney, 1988, Timoney and McCollum, 1993, Timoney et al., 1997). Further, the long-term carrier state in the stallion allows the generation of genetic and phenotypic diversity of EAV (Balasuriya et al., 1999, Balasuriya et al., 2004, Hedges et al., 1999). There is evidence that establishment and maintenance of the carrier state in stallions is testosterone-dependent. However, many aspects of the biology and immunology underlying viral persistence remain to be elucidated (Holyoak et al., 1993, Little et al., 1991, McCollum et al., 1994). Despite the significant biological and economic consequences of persistent EAV infection, there is to date no fully validated therapeutic strategy, except surgical castration, to eliminate the carrier state (Timoney and McCollum, 1993).
Phosphorodiamidate morpholino oligomers (PMO) comprise a class of single-stranded oligonucleotide analogues that contain the same purine and pyrimidine bases as DNA but possess a backbone composed of morpholine rings and phosphorodiamidate intersubunit linkages instead of the deoxyribose and phosphodiester components of DNA (Summerton and Weller, 1997). PMO can act as antisense agents by Watson–Crick base pairing with complementary RNA target sequence, thus forming a steric blockade and interfering with gene expression (Stein et al., 1997, Summerton, 1999). PMO are water-soluble, nuclease-resistant and are typically synthesized to be 20–25 subunits in length (Summerton, 1999). The covalent conjugation of an arginine-rich cell-penetrating peptide (CPP) to the PMO 5′-terminus has been found to greatly improve delivery of PMO into cells (Abes et al., 2006, Deas et al., 2005, Moulton et al., 2004). Peptide-conjugated PMO (PPMO) have been shown to inhibit amplification of a number of RNA viruses in cell culture and mouse models (reviewed by Stein, 2008), including the arterivirus, porcine reproductive and respiratory syndrome virus (Han et al., 2009, Patel et al., 2008, Patel et al., 2009, Zhang et al., 2006) and the coronaviruses, mouse hepatitis virus and SARS-CoV (Burrer et al., 2007, Neuman et al., 2005, Neuman et al., 2004). Previous studies with PRRSV (Patel et al., 2008) and EAV (van den Born et al., 2005) have shown that PPMO targeting the 5′-terminal region of the respective arteriviral genomes were particularly effective at inhibiting virus infection. van den Born et al. (2005) concluded that the mechanism of action by which EAV 5′-UTR-directed PPMO suppressed virus production was likely by inhibiting the translation of the pp1a and pp1ab.
We recently established an in vitro model of EAV persistence in HeLa cells for the purpose of furthering our understanding of EAV biology in general and viral persistence in the stallion in particular (Zhang et al., 2008a, Zhang et al., 2008b). The objective of the current study was to investigate whether an EAV-specific PPMO could effectively eliminate virus from persistently infected HeLa cells.
Our study employed an antisense PPMO targeting the 5′-terminal region of the EAV genome (EAV5′24: 5′-TATGGCACCATACACACTTCGAGC-3′, nucleotide positions 1–24 of the VB strain of EAV, GenBank accession number DQ846750) and a random sequence negative control 22mer PPMO (SCR: 5′-TGCTCTGTCTACAGTAGTGTCA-3′). The CPP (RXR)4XB (where R = arginine, X = 6-aminohexanoic acid and B = beta-alanine) was covalently conjugated to the 5′-end of each PMO. The synthesis, purification and analysis of EAV5′24 PPMO and SCR PPMO were performed at AVI BioPharma Inc. (Corvallis, OR) using methods previously described (Abes et al., 2006, Summerton and Weller, 1997). The potential cytotoxicity of each PPMO was evaluated in normal HeLa-H cells and in HeLa-H cells persistently infected with EAV. Briefly, normal HeLa-H and persistently infected HeLa-H cells (approximately 3 × 105 cells) were subcultured in 2 ml of serum-free Eagle's minimum essential medium (EMEM) containing the EAV5′24 PPMO or SCR PPMO at concentrations of 1 μM, 5 μM, or 10 μM in 6-well plates under conditions designed to mirror the antiviral experiments below. After 6 h treatment at 37 °C, 2 ml of EMEM containing 10% bovine calf serum was added. Cell viability was determined by comparing PPMO-treated to mock-treated cells at 24 h and 48 h post-treatment using the Vi-Cell XR (Beckman Coulter, California) which automates cell viability analysis based on the trypan blue vital dye exclusion method. Under these conditions, no cytotoxicity was observed in normal HeLa-H and persistently infected HeLa-H cells following treatment with either of the PPMO at any of the three concentrations tested (data not shown).
Subsequently, the effect of PPMO on HeLa-H cells persistently infected with EAV was investigated. The 83rd serial passage of the EAV-persistently infected HeLa-H cell line was grown in 6-well plates and subcultured (once every 3 days) for 10 passages (through the 92nd passage) in the presence of EAV5′24 PPMO or SCR PPMO at concentrations of 1 μM, 5 μM, or 10 μM. At each passage level, cells were first subcultured for 6 h in 2 ml of serum-free EMEM containing the respective concentration of each PPMO, or no PPMO, followed by addition of 2 ml of EMEM containing 10% bovine calf serum. Tissue culture supernatants at each passage level were harvested and evaluated for infectious virus by plaque assay in rabbit kidney cells (RK-13, ATCC CCL-37) as described previously (McCollum et al., 1962). Both standard RT-PCR (sRT-PCR) and real-time RT-PCR (rRT-PCR) analyses of RNA extracted from cell lysates were also carried out at each passage level following previously described procedures (Balasuriya et al., 2002, Gilbert et al., 1997, Lu et al., 2008). An indirect immunofluorescence assay (IFA) was performed at selected passage levels of cells using monoclonal antibody (MAb) 12A4 against the EAV nonstructural protein-1 (nsp1; Wagner et al., 2003) and MAb 3E2 against the EAV nucleocapsid (N) protein (MacLachlan et al., 1998) followed by fluorescein-conjugated goat anti-mouse immunoglobulin (Pierce), according to previously described procedures (Zhang et al., 2008b).
As shown in Fig. 1A, persistently infected HeLa-H cells receiving treatments of 1 μM EAV5′24 PPMO produced infectious virus with titers similar to untreated cells, throughout the observation period (83rd to 92nd passages). In contrast, virus in persistently infected cell cultures receiving treatments of 5 μM or 10 μM EAV5′24 PPMO gradually decreased in titer over the first three passages, after which no infectious virus was detectable, i.e. from the 87th passage onwards. Parallel results were obtained when EAV RNA was measured by sRT-PCR and rRT-PCR. Cell lysates from persistently infected HeLa-H cells either untreated or treated with 1 μM EAV5′24 PPMO were positive for EAV RNA from the 83rd to 92nd passages (Fig. 1A), whereas cells treated with 5 μM or 10 μM EAV5′24 PPMO were positive for EAV RNA only from the 83rd to 86th passages, with no EAV RNA detectable from the 87th passages onwards (Fig. 1A). Representative sRT-PCR and rRT-PCR results are shown in Fig. 1C and D, respectively. Persistently infected HeLa-H cells receiving various concentrations (1 μM, 5 μM, and 10 μM) of SCR PPMO or no PPMO continued to produce EAV RNA and infectious virus throughout the duration of the study, although some fluctuations in viral titers were observed over time (Fig. 1B). IFA evaluation was carried out on select passage levels of cells receiving EAV5′24 PPMO, SCR PPMO or no PPMO. The IFA results from the 92nd passage are depicted in Fig. 2A–F. Cells receiving no PPMO or 5 μM SCR PPMO robustly expressed EAV nonstructural (Fig. 2A and C) and structural (Fig. 2B and D) proteins, whereas cells receiving 5 μM EAV5′24 PPMO produced no detectable nonstructural (Fig. 2E) or structural (Fig. 2F) proteins. To determine if virus production would resume in cultures after discontinuation of the effective PPMO treatment described above, PPMO treatment was stopped after the 90th passage in a set of replicate cultures that had been receiving 5 μM or 10 μM EAV5′24 PPMO. These cultures were passed seven more times without PPMO treatment (through passage 97). Absence of PPMO treatment did not result in the production of renewed viral RNA, viral antigen or infectious virus as measured by sRT-PCR and rRT-PCR, IFA and plaque assay, respectively. Representative RT-PCR and IFA results for the PPMO-cured PI97 cells are shown in Fig. 1C and D and Fig. 2G and H. Together, these findings demonstrate that EAV infection was eliminated from persistently infected HeLa-H cells passaged in the presence of 5–10 μM EAV5′24 PPMO. Our results also further confirm the importance of the 5′-terminal region of the genome in the amplification-cycle of EAV (van den Born et al., 2005).
Fig. 1.
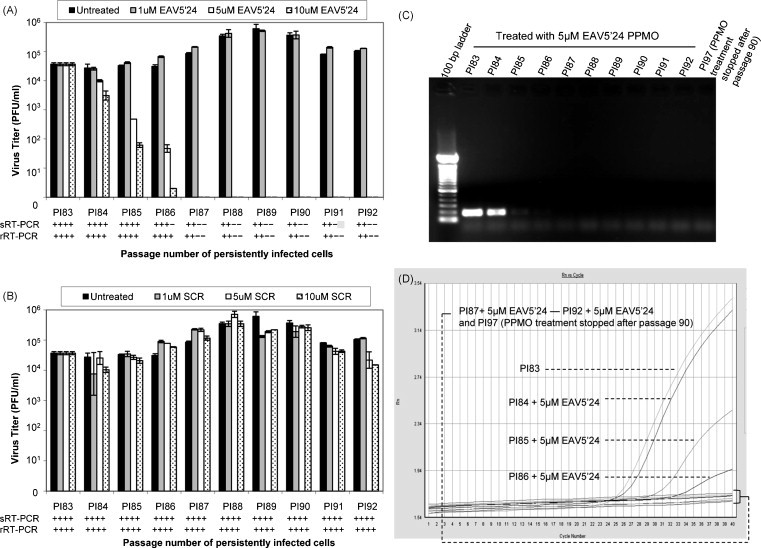
Effect of PPMO on HeLa-H cells persistently infected with EAV (strain VB). Persistently infected HeLa-H cells (83rd passage, PI83) were subcultured for 10 passages in the presence of 0 μM, 1 μM, 5 μM, or 10 μM of EAV5′24 PPMO or SCR PPMO. Supernatants from each passage were harvested and titrated by plaque assay as described in the text. The average results from two experiments are presented (A, EAV5′24 PPMO; B, SCR PPMO). In parallel, standard RT-PCR (sRT-PCR) and real-time RT-PCR (rRT-PCR) analysis were carried out with RNA extracted from cell lysates at each passage and the results are shown below each graph (+RT-PCR, positive; −RT-PCR, negative). Representative sRT-PCR (expected PCR product size is 185 bp) and rRT-PCR results of persistently infected HeLa-H cells treated with 5 μM EAV5′24 PPMO (PI83 to PI92) are shown in C and D, respectively. Also included is RT-PCR analysis of PPMO-cured PI97 HeLa-H cells in which the 5 μM EAV5′24 PPMO treatment was stopped after passage 90.
Fig. 2.
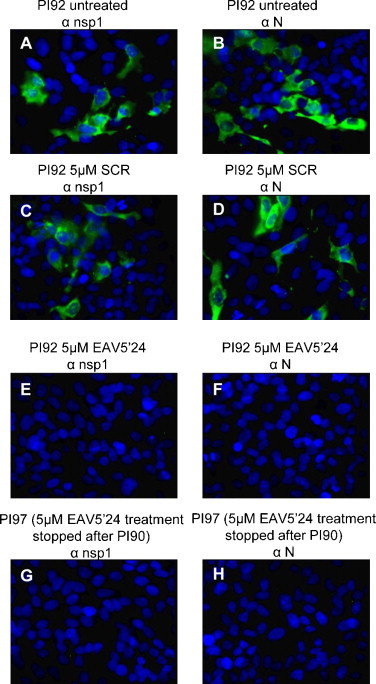
Representative IFA images. PI92 HeLa cells having received treatments with no PPMO (A and B), 5 μM SCR PPMO (C and D), or 5 μM EAV5′24 PPMO (E and F) between passages 84 and 92 were subject to IFA, respectively using MAb 12A4 against EAV nsp1 protein (αnsp1) and MAb 3E2 against EAV nucleocapsid protein (αN) followed by fluorescein-conjugated goat anti-mouse immunoglobulin. The cell nucleus was stained by Hoechst 33342 (blue color). Also included is the IFA image of PPMO-cured PI97 HeLa-H cells in which 5 μM EAV5′24 PPMO treatment was stopped after passage 90 (G and H). (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)
In a previous study we demonstrated that both the virulent Bucyrus (VB) strain of EAV and the EAVP80 virus recovered from the 80th passage of persistently infected HeLa-H cells could readily establish persistent infection in normal HeLa-H cells (Zhang et al., 2008b). With this in mind, we decided to investigate the growth kinetics of VB and EAVP80 viruses in HeLa-H cells cured of virus persistence by PPMO treatment. In addition, we investigated if persistent infection with either of these two viruses could be re-established in PPMO-cured HeLa-H cells. As shown in Fig. 3A, a substantial increase in infectious virus titer was observed in both VB- and EAVP80-infected HeLa-H cells (VB: ∼1.6 log increase; EAVP80: ∼2.7 log increase). In contrast, virus titers did not increase substantially in PPMO-cured HeLa-H cells inoculated with either VB or EAVP80 virus. Consistent with our previous findings, persistent infection could be established in normal HeLa-H cells with VB and EAVP80 viruses (Fig. 3B). In contrast, persistent infection could not be established with either virus strain in PPMO-cured HeLa-H cells, although low titers of virus could be detected for two passages in VB-inoculated and for five passages in EAVP80-inoculated cells (Fig. 3). These results demonstrate that the PPMO-cured HeLa-H cells were significantly less receptive than normal HeLa-H cells to re-infection with EAV. Factors contributing to the greatly reduced susceptibility of PPMO-cured HeLa-H cells to re-infection with EAV and to the re-establishment of virus persistence remain to be elucidated. The PPMO-cured PI97 HeLa-H cells evaluated in this study were obtained after treatment of persistently infected cells with 5 μM EAV5′24 PPMO for seven passages (84–90th) followed by seven more passages (91st to 97th) without PPMO treatment; it is therefore unlikely that residual PPMO made the cells resistant to re-infection with EAV. It would be of interest to compare expression levels of EAV receptors on PPMO-cured HeLa-H cells to those present on normal HeLa-H cells, but unfortunately, the cellular receptors involved in EAV infection have not as yet been clearly identified.
Fig. 3.
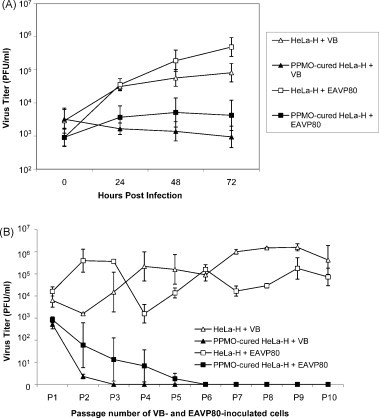
Comparison of the VB and EAVP80 viruses in normal HeLa-H cells and in PPMO-cured HeLa-H cells. (A) Growth characteristics of the VB and EAVP80 viruses in normal HeLa-H cells and in PPMO-cured HeLa-H cells. Subconfluent monolayers of normal HeLa-H and PPMO-cured PI97 HeLa-H cells were grown in 6-well plates and inoculated with each virus (moi of 3) at 37 °C for 1 h. After removal of the inoculum, the cells were rinsed three times with PBS and overlaid with 4 ml of EMEM. At indicated time points, supernatants were harvested and titrated by plaque assay. The average results from four experiments are shown. (B) Attempt to establish persistent infection with the VB and EAVP80 viruses in normal HeLa-H cells and PPMO-cured HeLa-H cells. Subconfluent monolayers of normal HeLa-H and PPMO-cured PI97 HeLa-H cells grown in T-25 flasks were inoculated with each virus at a moi of 3. Following 1 h adsorption at 37 °C, cell monolayers were washed three times with PBS and 10 ml of fresh culture medium was added. Inoculated cultures were incubated at 37 °C and subcultured once every 4 days. Tissue culture supernatants from serial subcultures up to the 10th passage were harvested and titrated for virus by plaque assay. The average results from three experiments are shown.
PPMO targeting viral sequences have been shown to inhibit amplification of a number of RNA viruses in vitro and in vivo (reviewed by Stein, 2008). However, antiviral studies with PPMO reported to date have been limited to acute viral infection models. In this study, we investigated the effects of PPMO on HeLa-H cells persistently infected with EAV. Treatment with an antisense PPMO targeting the EAV 5′-terminus at concentrations of 5–10 μM eliminated the virus from persistently infected HeLa-H cells, and the cells were confirmed free of EAV and EAV nucleic acid on subsequent passages in the absence of PPMO. Although our results indicate that PPMO can eliminate persistent EAV infection in cell culture, the efficacy of PPMO against EAV in vivo remains to be evaluated. Unfortunately no satisfactory small animal model for EAV infection exists. To investigate possible elimination of the carrier state in the stallion by PPMO, several complex issues would need to be addressed, including the potential toxicity of therapeutically relevant levels of PPMO in equines and the delivery of effective dosing of PPMO to the equine male reproductive tract. Nevertheless, the results of this in vitro study suggest that consideration of such in vivo investigation may be warranted.
Acknowledgements
We are grateful to the Chemistry Group at AVI BioPharma in Corvallis, OR for production of the PPMO used in this study. This study was supported by the Grayson Jockey Club Research Foundation Inc. in Lexington, KY, the Frederick Van Lennep Chair endowment fund at the Maxwell H. Gluck Equine Research Center, and the Kentucky Agricultural Experiment Station, College of Agriculture, University of Kentucky. D.A.S. was supported by funding from the Pacific Northwest Regional Center of Excellence for Biodefense and Emerging Infectious Diseases.
References
- Abes S., Moulton H.M., Clair P., Prevot P., Youngblood D.S., Wu R.P., Iversen P.L., Lebleu B. Vectorization of morpholino oligomers by the (R-Ahx-R)4 peptide allows efficient splicing correction in the absence of endosomolytic agents. J. Control. Release. 2006;116(3):304–313. doi: 10.1016/j.jconrel.2006.09.011. [DOI] [PubMed] [Google Scholar]
- Balasuriya U.B., Hedges J.F., Nadler S.A., McCollum W.H., Timoney P.J., MacLachlan N.J. Genetic stability of equine arteritis virus during horizontal and vertical transmission in an outbreak of equine viral arteritis. J. Gen. Virol. 1999;80(Pt 8):1949–1958. doi: 10.1099/0022-1317-80-8-1949. [DOI] [PubMed] [Google Scholar]
- Balasuriya U.B., Hedges J.F., Smalley V.L., Navarrette A., McCollum W.H., Timoney P.J., Snijder E.J., MacLachlan N.J. Genetic characterization of equine arteritis virus during persistent infection of stallions. J. Gen. Virol. 2004;85(Pt 2):379–390. doi: 10.1099/vir.0.19545-0. [DOI] [PubMed] [Google Scholar]
- Balasuriya U.B., Leutenegger C.M., Topol J.B., McCollum W.H., Timoney P.J., MacLachlan N.J. Detection of equine arteritis virus by real-time TaqMan reverse transcription-PCR assay. J. Virol. Methods. 2002;101(1–2):21–28. doi: 10.1016/s0166-0934(01)00416-5. [DOI] [PubMed] [Google Scholar]
- Burrer R., Neuman B.W., Ting J.P., Stein D.A., Moulton H.M., Iversen P.L., Kuhn P., Buchmeier M.J. Antiviral effects of antisense morpholino oligomers in murine coronavirus infection models. J. Virol. 2007;81(11):5637–5648. doi: 10.1128/JVI.02360-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavanagh D. Nidovirales: a new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 1997;142(3):629–633. [PubMed] [Google Scholar]
- Deas T.S., Binduga-Gajewska I., Tilgner M., Ren P., Stein D.A., Moulton H.M., Iversen P.L., Kauffman E.B., Kramer L.D., Shi P.Y. Inhibition of flavivirus infections by antisense oligomers specifically suppressing viral translation and RNA replication. J. Virol. 2005;79(8):4599–4609. doi: 10.1128/JVI.79.8.4599-4609.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert S.A., Timoney P.J., McCollum W.H., Deregt D. Detection of equine arteritis virus in the semen of carrier stallions by using a sensitive nested PCR assay. J. Clin. Microbiol. 1997;35(8):2181–2183. doi: 10.1128/jcm.35.8.2181-2183.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han X., Fan S., Patel D., Zhang Y.J. Enhanced inhibition of porcine reproductive and respiratory syndrome virus replication by combination of morpholino oligomers. Antiviral Res. 2009;82(1):59–66. doi: 10.1016/j.antiviral.2009.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedges J.F., Balasuriya U.B., Timoney P.J., McCollum W.H., MacLachlan N.J. Genetic divergence with emergence of novel phenotypic variants of equine arteritis virus during persistent infection of stallions. J. Virol. 1999;73(5):3672–3681. doi: 10.1128/jvi.73.5.3672-3681.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holyoak G.R., Little T.V., McCollam W.H., Timoney P.J. Relationship between onset of puberty and establishment of persistent infection with equine arteritis virus in the experimentally infected colt. J. Comp. Pathol. 1993;109(1):29–46. doi: 10.1016/S0021-9975(08)80238-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Little T.V., Holyoak G.R., McCollum W.H., Timoney P.J. Output of equine arteritis virus from persistently infected stallion is testosterone dependent. Proceedings of the 6th International Conference of Equine Infectious Diseases; Cambridge, UK; 1991. pp. 225–229. [Google Scholar]
- Lu Z., Branscum A.J., Shuck K.M., Zhang J., Dubovi E.J., Timoney P.J., Balasuriya U.B. Comparison of two real-time reverse transcription polymerase chain reaction assays for the detection of Equine arteritis virus nucleic acid in equine semen and tissue culture fluid. J. Vet. Diagn. Invest. 2008;20(2):147–155. doi: 10.1177/104063870802000202. [DOI] [PubMed] [Google Scholar]
- MacLachlan N.J., Balasuriya U.B., Hedges J.F., Schweidler T.M., McCollum W.H., Timoney P.J., Hullinger P.J., Patton J.F. Serologic response of horses to the structural proteins of equine arteritis virus. J. Vet. Diagn. Invest. 1998;10(3):229–236. doi: 10.1177/104063879801000302. [DOI] [PubMed] [Google Scholar]
- McCollum W.H., Doll E.R., Wilson J.C., Cheatham J. Isolation and propagation of equine arteritis virus in monolayer cell cultures of rabbit kidney. Cornell Vet. 1962;52:452–458. [PubMed] [Google Scholar]
- McCollum W.H., Little T.V., Timoney P.J., Swerczek T.W. Resistance of castrated male horses to attempted establishment of the carrier state with equine arteritis virus. J. Comp. Pathol. 1994;111(4):383–388. doi: 10.1016/s0021-9975(05)80096-9. [DOI] [PubMed] [Google Scholar]
- Moulton H.M., Nelson M.H., Hatlevig S.A., Reddy M.T., Iversen P.L. Cellular uptake of antisense morpholino oligomers conjugated to arginine-rich peptides. Bioconjug. Chem. 2004;15(2):290–299. doi: 10.1021/bc034221g. [DOI] [PubMed] [Google Scholar]
- Neuman B.W., Stein D.A., Kroeker A.D., Churchill M.J., Kim A.M., Kuhn P., Dawson P., Moulton H.M., Bestwick R.K., Iversen P.L., Buchmeier M.J. Inhibition, escape, and attenuated growth of severe acute respiratory syndrome coronavirus treated with antisense morpholino oligomers. J. Virol. 2005;79(15):9665–9676. doi: 10.1128/JVI.79.15.9665-9676.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neuman B.W., Stein D.A., Kroeker A.D., Paulino A.D., Moulton H.M., Iversen P.L., Buchmeier M.J. Antisense morpholino-oligomers directed against the 5′ end of the genome inhibit coronavirus proliferation and growth. J. Virol. 2004;78(11):5891–5899. doi: 10.1128/JVI.78.11.5891-5899.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel D., Opriessnig T., Stein D.A., Halbur P.G., Meng X.J., Iversen P.L., Zhang Y.J. Peptide-conjugated morpholino oligomers inhibit porcine reproductive and respiratory syndrome virus replication. Antiviral Res. 2008;77(2):95–107. doi: 10.1016/j.antiviral.2007.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel D., Stein D.A., Zhang Y.J. Morpholino oligomer-mediated protection of porcine pulmonary alveolar macrophages from arterivirus-induced cell death. Antivir. Ther. 2009;14(7):899–909. doi: 10.3851/IMP1409. [DOI] [PubMed] [Google Scholar]
- Snijder E.J., Meulenberg J.J. The molecular biology of arteriviruses. J. Gen. Virol. 1998;79(Pt 5):961–979. doi: 10.1099/0022-1317-79-5-961. [DOI] [PubMed] [Google Scholar]
- Snijder E.J., Spaan W.J. Arteriviruses. In: Knipe D.M., Howley P.M., editors. Fields Virology. 5th ed. Lippincott Williams & Wilkins; Philadelphia: 2006. pp. 1337–1355. [Google Scholar]
- Snijder E.J., van Tol H., Pedersen K.W., Raamsman M.J., de Vries A.A. Identification of a novel structural protein of arteriviruses. J. Virol. 1999;73(8):6335–6345. doi: 10.1128/jvi.73.8.6335-6345.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stein D., Foster E., Huang S.B., Weller D., Summerton J. A specificity comparison of four antisense types: morpholino, 2′-O-methyl RNA, DNA, and phosphorothioate DNA. Antisense Nucleic Acid Drug Dev. 1997;7(3):151–157. doi: 10.1089/oli.1.1997.7.151. [DOI] [PubMed] [Google Scholar]
- Stein D.A. Inhibition of RNA virus infections with peptide-conjugated morpholino oligomers. Curr. Pharm. Des. 2008;14(25):2619–2634. doi: 10.2174/138161208786071290. [DOI] [PubMed] [Google Scholar]
- Summerton J. Morpholino antisense oligomers: the case for an RNase H-independent structural type. Biochim. Biophys. Acta. 1999;1489(1):141–158. doi: 10.1016/s0167-4781(99)00150-5. [DOI] [PubMed] [Google Scholar]
- Summerton J., Weller D. Morpholino antisense oligomers: design, preparation, and properties. Antisense Nucleic Acid Drug Dev. 1997;7(3):187–195. doi: 10.1089/oli.1.1997.7.187. [DOI] [PubMed] [Google Scholar]
- Timoney P.J. Equine viral arteritis: epidemiology and control. J. Equine Vet. Sci. 1988;8:54–59. [Google Scholar]
- Timoney P.J., McCollum W.H. Equine viral arteritis. Vet. Clin. North Am. Equine Pract. 1993;9(2):295–309. doi: 10.1016/S0749-0739(17)30397-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Timoney P.J., McCollum W.H., Vickers M.L. The carrier stallion as a reservoir of equine arteritis virus. Equine Dis. Q. 1997;6(1):2. [Google Scholar]
- van den Born E., Stein D.A., Iversen P.L., Snijder E.J. Antiviral activity of morpholino oligomers designed to block various aspects of Equine arteritis virus amplification in cell culture. J. Gen. Virol. 2005;86(Pt 11):3081–3090. doi: 10.1099/vir.0.81158-0. [DOI] [PubMed] [Google Scholar]
- Wagner H.M., Balasuriya U.B., James MacLachlan N. The serologic response of horses to equine arteritis virus as determined by competitive enzyme-linked immunosorbent assays (c-ELISAs) to structural and non-structural viral proteins. Comp. Immunol. Microbiol. Infect. Dis. 2003;26(4):251–260. doi: 10.1016/S0147-9571(02)00054-1. [DOI] [PubMed] [Google Scholar]
- Zhang J., Timoney P.J., Maclachlan N.J., Balasuriya U.B. Identification of an additional neutralization determinant of equine arteritis virus. Virus Res. 2008;138(1–2):150–153. doi: 10.1016/j.virusres.2008.09.003. [DOI] [PubMed] [Google Scholar]
- Zhang J., Timoney P.J., MacLachlan N.J., McCollum W.H., Balasuriya U.B. Persistent equine arteritis virus infection in HeLa cells. J. Virol. 2008;82(17):8456–8464. doi: 10.1128/JVI.01249-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y.J., Stein D.A., Fan S.M., Wang K.Y., Kroeker A.D., Meng X.J., Iversen P.L., Matson D.O. Suppression of porcine reproductive and respiratory syndrome virus replication by morpholino antisense oligomers. Vet. Microbiol. 2006;117(2–4):117–129. doi: 10.1016/j.vetmic.2006.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]