

Abstract
The persistent public health threat of infection with the Middle East respiratory syndrome coronavirus (MERS-CoV) highlights the need for an effective MERS-CoV vaccine. Previous studies have focused mainly on the receptor-binding domain (RBD) on the spike protein of MERS-CoV. Herein, we investigated the immunogenicity and protective potential of the recombinant N-terminal domain (rNTD) of spike proteins as a vaccine candidate. BALB/c mice vaccinated with 5 or 10 μg of rNTD protein demonstrated a significant humoral immune response (serum IgG and neutralizing activity). Additionally, according to the enzyme-linked immunospot, intracellular cytokine staining, and cytometric bead array assays, significant and functional T-cell immunity was induced by 10 μg of the rNTD vaccination with aluminum and CpG adjuvant. Furthermore, rNTD-immunized mice showed reduced lung abnormalities in a MERS-CoV-challenge mouse model transfected with an adenoviral vector expressing human DPP4, showing protection consistent with that found with rRBD vaccination. These data show that rNTD induced potent cellular immunity and antigen-specific neutralizing antibodies in mice and that it demonstrated protective capacity against a viral challenge, indicating that rNTD is a vaccine candidate against MERS-CoV infection.
Keywords: MERS-CoV, Vaccine, Mice, NTD, RBD, Animal model
1. Introduction
Middle East respiratory syndrome coronavirus (MERS-CoV) was first isolated in 2012 from a patient suffering from severe respiratory illness in Saudi Arabia [1]. Phylogenetic analysis showed that the virus represented a novel member of the betacoronavirus and was most closely related to bat coronaviruses HKU4 and HKU5 [2], which were distinct from severe acute respiratory syndrome coronavirus (SARS-CoV) [3]. Since then, MERS-CoV continued to circulate and cause human infection in different countries [4], including 27 countries in Europe, North Africa, Asia, USA, and the Middle East. From September 2012 to September 2016, WHO documented 1806 laboratory-confirmed cases of infection with MERS-CoV globally, including at least 643 related deaths with a case fatality rate of 35.6% (http://www.who.int/csr/don/21-September-2016-mers-saudi-arabia/en/). A number of clusters, including a recent outbreak in the Republic of Korea, indicated human-to-human transmission of the virus, evoking global concern and intensive discussion worldwide [4]. Although many therapeutic antibodies, including humanized neutralizing antibody, have been developed [5], [6], [7], [8], [9], none has been approved for clinical application. Therefore, the development of an effective vaccine against MERS-CoV infection is urgent.
Spike (S) protein mediates coronavirus entry into host-cells by first binding to the receptor on the host-cell surface [10]. As a class I viral membrane fusion protein, the ectodomain of S can be divided into receptor-binding S1 and membrane-fusion S2 subunits. S1 contains two independent domains, an N-terminal domain (NTD) and a C-terminal domain (CTD) [11], either or both of which could function as the receptor-binding domain (RBD) of various coronaviruses [12]. The NTD of mouse hepatitis coronavirus (MHV) recognizes carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM1) [13], and the CTD of SARS-CoV recognizes angiotensin-converting enzyme 2 (ACE2) [14]. The NTD and CTD of porcine transmissible gastroenteritis coronavirus (TGEV) recognize the sugar and aminopeptidase N (APN) moieties, respectively [12], [15]. MERS-CoV utilizes the CTD as its RBD to bind the dipeptidyl peptidase 4 (DPP4) of permissive cells [16], [17]. As the RBD contains major neutralization epitopes and induces most of the host immune responses [12], most studies have focused on the RBD [18], [19], [20], [21] for the development of a vaccine against MERS-CoV infection [22], [23], [24], [25]. However, rRBD vaccination induced only partial protection against the MERS-CoV challenge in mice or non-human primates (NHPs) [19], [26]. Additionally, the nature of the rapid variation in the RNA virus increases the likelihood of immunologic escape, especially with regard to countermeasures directed at the RBD. Therefore, developing a different immunogen to replace or use in combination with the RBD as a vaccine candidate is a promising strategy against MERS-CoV infection.
Vaccines against MERS-CoV infection have been developed using purified coronavirus S protein [27], as well as DNA or viral vector-based vaccines expressing full-length MERS-CoV S protein; these have been tested for their ability to induce virus-neutralizing antibodies in mice [22], [23], [24], [28], [29]. A recent study using serological analysis to develop murine monoclonal antibodies showed that immunization elicits neutralizing antibodies to the non-RBD portions of S1 and S2 subunits [30]. These results indicated that protective antigenic elements might exist on the S glycoprotein in addition to the RBD. In this study, we explored the NTD of MERS-CoV S as a novel subunit vaccine and evaluated its protective effects against MERS-CoV infection in a mouse model.
2. Materials and methods
2.1. Ethics statement
Animal studies were carried out in strict compliance with the guidelines for the Care and Use of Laboratory Animals of the People’s Republic of China. The protocol of the study was approved by the Committee on the Ethics of Animal Experiments of the Chinese Center for Disease Control and Prevention. All procedures were performed under ethyl ether anesthesia, and all efforts were made to minimize animal suffering.
2.2. rNTD vaccine preparation
As shown in Fig.1 A, the rNTD protein consists of a gp67 signal peptide (SP) and 336 amino acid (aa) residues (18–353 aa) of S derived from MERS-CoV (hCoV-EMC strain, Genbank ID: JX869059). The coding sequences of MERS-CoV NTD were cloned into the pFastBac1 vector. Transfection and virus amplification were conducted with Sf9 cells, and the recombinant protein was produced in High5 cells and purified using the same approach employed in the rRBD preparation [16]. The purified rNTD was resolved by 10% SDS-PAGE followed by Western blot analysis (Fig.1B) as described previously [16].
Fig. 1.
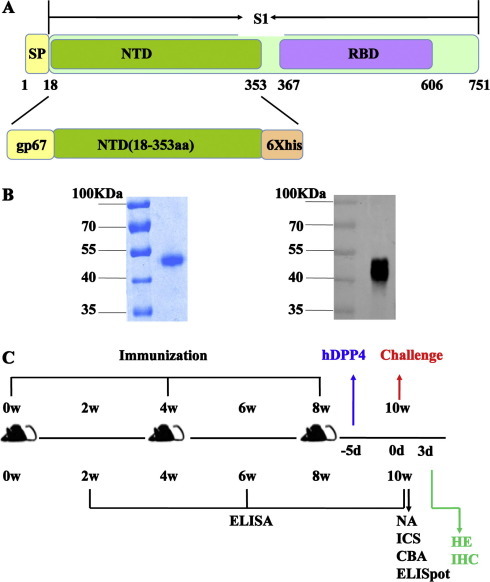
Description of the N-terminal domain (NTD) immunogen and vaccination schedule. (A) The location of the NTD protein on the Middle East respiratory syndrome coronavirus MERS-CoV spike (S) protein. The recombinant (r)NTD protein consists of 336 amino acid (aa) residues (18–353) of S protein. A gp67 signal peptide (SP) was added to the N terminus for expression of the rNTD protein. (B) Purified rNTD protein detected by SDS-PAGE (left) and Western blot (right). The purified rNTD protein was separated by a 10% SDS-PAGE and stained with 0.25% Coomassie brilliant blue. Anti-NTD polyclonal antibody and infrared ray-labeled secondary antibody were used for the Western blot assay. Lane 1: protein molecular weight marker; lane 2: purified rNTD protein. (C). Vaccination schedule and detection. Mice received three vaccinations consisting of 5 or 10 μg of rNTD protein combined with adjuvants at 4-week intervals. Sera were collected at the indicated times to analyze the humoral immune response. Six mice from each group were sacrificed 2 weeks after the last immunization. The spleens were harvested for enzyme-linked immunospot (ELISpot), intracellular cytokine staining (ICS), and cytometric bead array (CBA) assays. In parallel experiments, the remaining mice were challenged with MERS-CoV to detect the protective effect elicited by the rNTD protein. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
2.3. Mice immunization and sampling
Female BALB/c mice aged 6–8 weeks were injected intramuscularly (i.m.) with rNTD protein at a dose of 5 or 10 μg with 100 μL of aluminum hydroxide and 10 μg of CpG adjuvant. The control group was immunized with aluminum and CpG without immunogen. The day before vaccination, aluminum was mixed with the protein at a final concentration of 1 mg/ml. For a thorough mixture, aluminum hydroxide and the protein were mixed every few hours. As shown in Fig.1C, all mice were immunized three times at 4-week intervals. For detection of total IgG level and neutralization activity, the mice were bled from the vena angularis 2 weeks after each immunization (2W, 6W, 10W) and 14 weeks after the last immunization (22W). To evaluate the cellular-mediated immunity (CMI), the mice were euthanized and splenocytes were isolated at 2 weeks after the last immunization (10W) or 14 weeks after the last immunization (22W). Additionally, vaccination with rRBD protein at a dose of 10 μg with 100 μL of aluminum and 10 μg of CpG adjuvant was applied for challenge control as described previously [19].
2.4. MERS-CoV challenge
Nine days after the last immunization with 10 μg of rNTD, rRBD, or adjuvant control, mice were anesthetized lightly with isoflurane and transduced intranasally with 2.5 × 108 plaque-forming units (pfu) of Ad5-hDPP4 for rapid generation of a mouse model of MERS-CoV infection as described previously [31]. Five days later, the transduced mice were infected intranasally with MERS-CoV (1 × 105 pfu) in a total volume of 50 μL of DMEM. Three days’ post-infection (dpi) with MERS-CoV, the mice were euthanized, and the lungs and trachea were harvested for pathology detection. All experiments with MERS-CoV were conducted in an animal biosafety level 3 (ABSL-3) facility.
2.5. Enzyme-linked immunosorbent assay (ELISA)
The antigen-specific IgG antibody responses were detected by ELISA as described previously [22]. Briefly, serially diluted mouse sera were added to 96-well micro-well plates pre-coated with rNTD protein at 100 ng per well. After blocking with FBS and washing with PBST, bound antibodies were reacted with horseradish peroxidase-conjugated anti-mouse IgG (1:5000, Sigma-Aldrich, St. Louis, MO, US) at 37 °C for 1 h. The samples were then washed with PBST, after which 3,3′,5,5′-tetramethylbenzidine was added to the plates. The reaction was stopped by adding 2 M H2SO4. The absorbance was read at 450 nm using an ELISA plate reader (Thermo, Waltham, MA,US). The cut-off value was set to 2.1-fold above that of the negative control.
2.6. Pseudovirus neutralization assay
The neutralizing activity of mouse sera was determined using the MERS-CoV pseudovirus system as described previously [32]. In brief, serum samples were serially diluted twofold in 96-well tissue culture plates and incubated at 37 °C for 2 h with the addition of MERS-CoV pseudovirus to each well. The mixtures were then transferred to duplicate wells containing confluent Huh7 cells. After incubation for 72 h, a luciferase assay was performed using an Ultra 384 luminometer (Tecan, Hombrechtikon, CH). The neutralizing antibody efficiency was calculated as: (relative luciferase units of mock sera – relative luciferase units of immune serum for a given dilution)/relative luciferase units of mock sera.
2.7. Plaque reduction neutralization (PRNT) assay
The PRNT assay was conducted in a BSL-3. Briefly, sera from immunized mice were serially diluted twofold. After incubation with the same volume containing MERS-CoV (hCoV-EMC), the mixtures were transferred to a 12-well plate with confluent Vero cells for 1 h. As a control, the virus was likewise incubated with DMEM, and cells were cultured at 37 °C for an additional 72 h. The cells were fixed with paraformaldehyde for 20 min and dyed with Gentian violet for an additional 5 min. The plates were washed with tap water, and the viral plaques that formed were counted. Plaque reduction was calculated as follows: inhibition percentage = 100 × [1 − (plaque number incubated with sera/plaque number with virus)]. The titer is expressed as PRNT50 (the serum dilution resulting in a 50% plaque reduction compared with plaque formation in virus-only controls).
2.8. Enzyme-linked immunospot (ELISpot) assay
To evaluate the antigen-specific T-cell response induced by the vaccination regime, an IFN-γ ELISpot assay was performed as described previously [19]. Briefly, 96-well plates were coated with 100 μL per well of 5 mg/mL anti-mouse IFN-γ antibody (BD Pharmingen, San Diego, CA, USA) overnight at 4 °C and then blocked for 2 h at room temperature (RT). Freshly harvested splenocytes (5 × 105/well) were stimulated in triplicate for 24 h with 4 mg/mL of a synthesized 18-mer peptide library, which overlapped with the NTD of MERS-CoV. Next, a biotinylated detection antibody (BD Pharmingen) and streptavidin-horseradish peroxidase were added. Blots were developed by the addition of an AEC (3-amino-9-ethylcarbazole) substrate solution, which produced a colored spot after 5-min RT incubation in the dark. Finally, IFN-γ spot-forming cells (SFCs) were counted by a Bio-reader (BIOSYS, Pasadena, CA, US).
2.9. Intracellular cytokine staining (ICS)
The ICS assay was performed as described previously [33]. In brief, splenocytes (2.5 × 106/sample) were cultured for 4 h at 37 °C in a 96-well round-bottom micro-well plate with complete RPMI 1640 culture supplemented with peptide pools at a concentration of 4 μg/mL to stimulate the cells. Control cells were incubated with an unrelated peptide or without peptide. Brefeldin A was then added and incubated with the cells for 2 h before staining. Cells were washed and incubated for 30 min at 4 °C with 50 μL of a 1/100 dilution of FITC-labeled antibody targeting mouse CD8 and PerCP-labeled CD4. Cells were washed again, permeabilized in 1× cytofix/cytoperm for 20 min at 4 °C, washed three times with perm/wash buffer, and incubated in the same buffer for 30 min at 4 °C with 50 μL of a 1/100 dilution of APC-labeled antibody targeting mouse IFN-γ and PE-labeled antibody targeting mouse IL-2. After washing, cells were examined by three-color flow cytometry. The above reagents and antibodies were purchased from BD Pharmingen (San Diego, CA, USA).
2.10. Cytometric bead array (CBA)
To investigate the levels of Th1-, Th2-, and Th17-type cytokine secretion, CBA analysis was conducted as described previously [22]. Splenocytes (5 × 105/well) were distributed in 96-well plates and stimulated with 4 mg/mL of pooled peptides including the NTD. Plates were incubated for 24 h at 37 °C, and supernatants were harvested. The concentrations of cytokines, including IL-2, IL-4, IL-6, IL-10, TNF-α, IL-17A, and IFN-γ, were measured using a mouse Th1/Th2/Th17 cytokine kit (Becton Dickinson, New Jersey, US) and a FACS Calibur flow cytometer (Becton Dickinson, Franklin Lakes, NJ, US). Data were analyzed using the FCAP Array software (Becton Dickinson, Franklin Lakes, NJ, US).
2.11. Hematoxylin-eosin (HE) and immunohistochemistry (IHC) staining
After euthanizing the mice, the lungs and trachea were collected for HE and IHC examination. The tissues were fixed using 10% neutral buffered formalin, embedded in paraffin, sectioned sequentially to 4-μm thickness, and stained with HE prior to examination by light microscopy. For IHC analysis, staining methods included antigen retrieval with citrate buffer. Rabbit-serum-derived polyclonal antibody against nucleoprotein (cat: 100213-RP02; Sino Biological Inc., Beijing, CHN) was incubated with the sections at 1:1000 dilution; goat anti-rabbit (cat: pv-9001; ZSGB-Bio, Beijing, CHN) secondary antibody was used at 1:2000, and sections were evaluated using light microscopy. Various sites of the lungs and trachea tissue samples were collected for histopathology and IHC analyses. Finally, 4–5 slides per animal were used for quantification to assess the effect and impact of the vaccine on the immunized animals.
3. Statistical analysis
Statistical analyses were conducted using one-way ANOVA with a Bonferroni post-test using the SPSS for Windows software package (ver. 17, SPSS Inc., Chicago, IL, USA). Unpaired two-tailed Student’s t-tests were used to compare the means of different groups. A value of P < 0.05 was considered to indicate statistical significance. Results are expressed as means ± standard deviation (s.d.). All figures were rendered using the Prism 5 software package (GraphPad Software, San Diego,CA, USA).
4. Results
4.1. Expression and purification of rNTD protein
The rNTD spanning 18–353 aa of MERS-CoV S protein was produced using the Bac-to-Bac system, and rNTD was purified by SDS-PAGE and Coomassie brilliant blue staining as shown in Fig.1B (left). rNTD was recognized by a polyclonal anti-NTD mouse serum (Fig.1B) at the expected molecular weight of 45 kDa (right). This purified rNTD protein was evaluated at a dose of 5 or 10 μg with 100 μL of aluminum and 10 μg of CpG adjuvant as a subunit vaccine candidate in mice [19].
4.2. Significant and sustained humoral immune response was induced by the rNTD vaccination
Two weeks after immunization, sera of the immunized mice were collected, and antigen-specific IgG was detected by ELISA. The results showed detectable IgG in the sera of rNTD-immunized mice after the first immunization, although the antibody titers were low. After the second immunization, the titer of the NTD group peaked as high as 1:105, which could not be boosted by the third immunization (Fig.2 A). Furthermore, IgG was detected at 22 weeks without an obvious decline. At all three detection time points, the IgG titer induced by 10 μg of rNTD did not exceed that induced by 5 μg of rNTD protein.
Fig. 2.
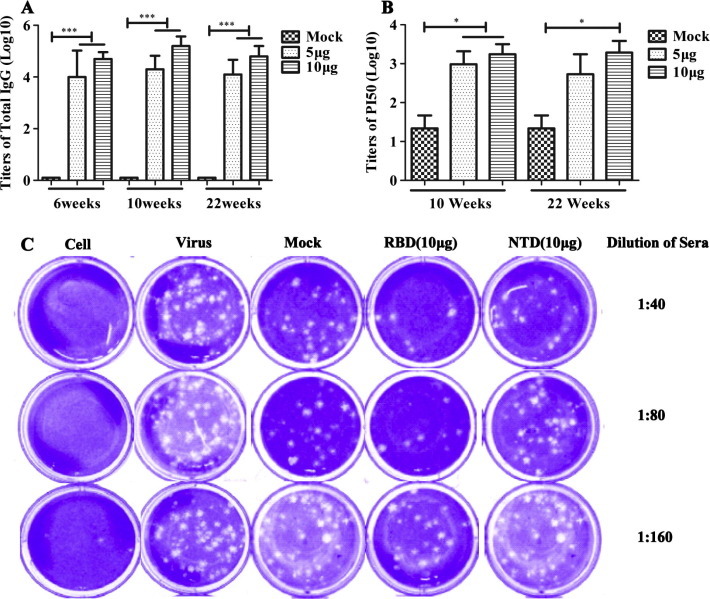
Humoral immunity response induced in rNTD-immunized mice. (A) NTD-specific IgG titers were determined by enzyme-linked immunosorbent assay (ELISA) at 6, 10, and 22 weeks (14 weeks after the last immunization). (B) The neutralization activity in the sera of mice was detected using the pseudovirus neutralization assay at 10 or 22 weeks. (C) Representative results of the plaque reduction neutralization (PRNT) assay for the detection of neutralization activity in the sera of mice. Approximately 30 pfu virus stock (hCoV-EMC) was used to infect Vero cells in 12-well plates with or without heat-inactivated sera from immunized mice 2 weeks (10W) after the third immunization. PRNT50 was calculated after the plaques were counted. Statistical significance was set at ∗P < 0.05 and ∗∗∗P < 0.001.
Sera from the immunized mice were evaluated for neutralizing activity against MERS-CoV using a pseudotype reporter virus neutralization assay. There were no detectable neutralizing antibodies in the sera of immunized mice 2 weeks after the second immunization (6W). However, a high titer of neutralization activity was induced at 2 weeks after the third immunization (10 W) with either 5 or 10 μg of rNTD protein as shown in Fig.2B. Moreover, the neutralizing activity in the sera of both immunized groups was sustained up to 14 weeks after the last immunization (22W) without an obvious decline (Fig.2B).
To investigate the neutralization antibodies induced by the rNTD protein, a PRNT assay was conducted using MERS-CoV (hCoV-EMC); in this procedure, the positive control consisted of sera obtained from mice immunized with the same dose of rRBD protein. A representative result, shown in Fig.2C, demonstrates that the neutralization of NTD was evident compared with that of the groups of adjuvants, although it was slightly lower than that obtained with RBD. When diluted to 1:80 or 1:160, the sera from the NTD-immunized mice showed significant difference in neutralizing activity compared with the RBD protein. Therefore, the rNTD protein induced effective neutralization antibodies (PRNT50 = 1:40) in mice, and the titer of neutralization antibodies was lower than that obtained with the same dose of rRBD protein (PRNT50 = 1:160).
4.3. Significant and functional CMI induced by vaccination with 10 μg of rNTD
To evaluate the cellular immunological response, splenocytes of the immunized mice were harvested 2 weeks after the last immunization, and the IFN-γ-secreting cells were counted using the ELISpot assay. As shown in Fig.3 A, at a dose of 5 μg, rNTD induced low levels of CMI, whereas the dose of 10 μg of rNTD protein induced significant CMI levels, with a higher frequency of IFN-γ-secreting cells. Additionally, the induced T-cell immunity was sustained up to 14 weeks after the last immunization (22W) without an obvious decline.
Fig. 3.
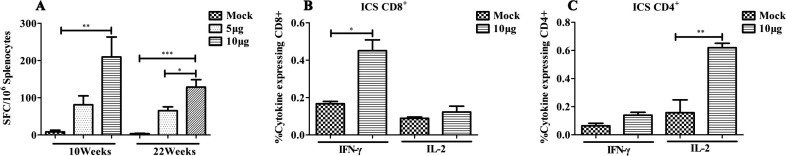
Cellular immune response induced by the rNTD-immunized mice detected by ELISpot and ICS. (A) ELISpot analysis of IFN-γ secretion cells in spleen. Data are expressed as spot-forming cells (SFCs) responding to peptide-specific IFN-γ secretion and presented as means with standard deviation. (B) ICS assay to detect specific IFN-γ and IL-2 expression in CD8+ T-cells of the splenocytes in mice. (C) ICS assay to detect specific IFN-γ and IL-2 expression in the CD4+ T-cells of the splenocytes in mice. The thresholds for statistically significant differences between groups were set at ∗P < 0.05, ∗∗P < 0.01, and ∗∗∗P < 0.001.
To evaluate the functionality of T-lymphocyte responses in the spleens of mice immunized with 10 μg of rNTD, an ICS assay was performed 2 weeks after the third immunization. The secretion of IFN-γ was observed mainly in the CD8+ T-cells (Fig.3B, left) compared with in the CD4+ T-cells (Fig.3C, left). Additionally, the splenocytes of rNTD-immunized mice secreted IL-2, which was due mainly to the CD4+ T-cells (Fig.3C, right) compared with the CD8+ T-cells (Fig.3B, right).
Cytokines of Th1 (Fig.4 A), Th2 (Fig.4B) and Th17 (Fig.4C) produced by splenocytes from mice immunized with 10 μg rNTD were analyzed by CBA assay after stimulation with protein-specific peptides. Higher concentrations of IL-2 and IL-17A were induced in the splenocytes of immunized mice compared with control groups. Additionally, the splenocytes of rNTD-immunized mice secreted higher concentrations of IL-6 than of IL-10. Thus, rNTD protein immunization induced a composite of Th1, Th2, and Th17 T-cell responses.
Fig. 4.
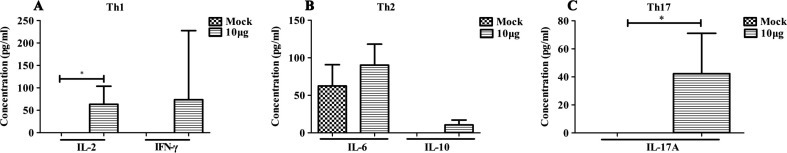
Cytokine production of splenocytes among immunized mice was determined by CBA 2 weeks after the third immunization. Splenocytes were stimulated for 24 h with pooled peptides consisting of the NTD, and the supernatants were harvested. Cytokine concentration was detected using a CBA kit. The cytokines of Th1, Th2, and Th17 are shown in the figures. (A) IL-2 and IFN-γ. (B) IL-6 and IL-10. (C) IL-17A. Statistical significance was set at ∗P < 0.05.
4.4. Protection from MERS-CoV challenge induced by either rNTD or rRBD vaccination in mice
To further elucidate the protective effect of the vaccination, histopathological and IHC analyses were conducted on mice challenged with MERS-CoV. As shown in Fig.5 A, the lung tissue of mice in the control group exhibited severe lesions, including the loss of pulmonary alveolus and diffuse inflammatory cell infiltration. In contrast, milder lesions were observed in mice immunized with rNTD or rRBD, as the pulmonary alveolus was highly visible with lower infiltration of inflammatory cells (Fig.5B and C). Similarly, compared with the control group, mice immunized with the recombinant protein showed slightly lower pathologic damage to the trachea (Fig.5D–F). As shown in Fig. 6 , IHC revealed that the MERS-CoV antigen was located mainly in the trachea. Additionally, MERS-CoV expression was detected in the lung tissue of all immunized groups. These results indicate that rNTD or rRBD vaccination reduced the respiratory tract pathology in mice after challenge with the MERS-CoV. However, this protection was incomplete, as MERS-CoV expression was detected in the immunized groups.
Fig. 5.
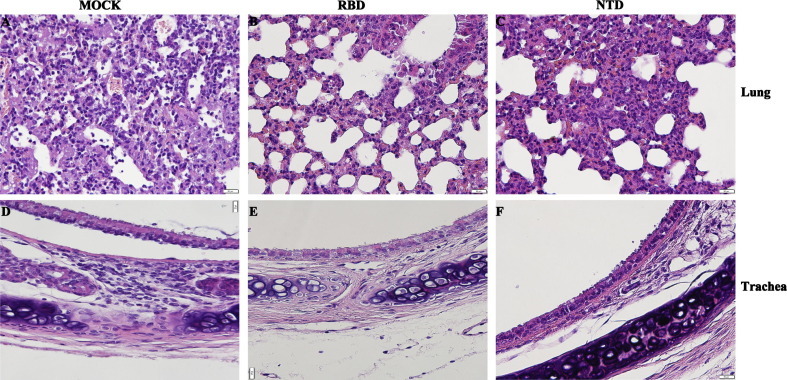
rNTD or rRBD vaccination reduced respiratory tract pathology in mice after MERS-CoV challenge. Representative results of hematoxylin-eosin (HE) staining in the lung (A–C) and trachea (D–F) of mock-treated or immunized mice. Severe lesions including the loss of pulmonary alveolus (represented by the white vacuole) and diffuse inflammatory cell infiltration (represented by the dark purple point) are shown (figure A). In contrast, milder lesions were observed among mice immunized with rRBD (figure B) or NTD (figure C), as the pulmonary alveolus was highly visible with less inflammatory cell infiltration. Inflammatory cell infiltration and impaired epithelium of the tunica mucosa bronchiorum were seen in the mock group (D). rRBD (E) or rNTD (F) alleviated the pathologic damage in the trachea of immunized mice. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Fig. 6.
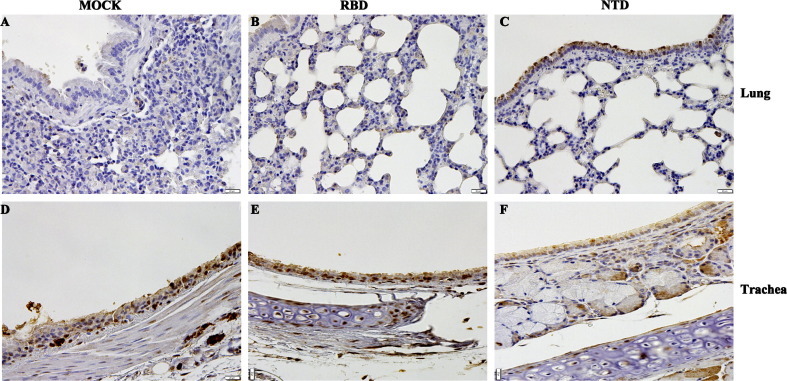
IHC detection of virus antigen expression in mouse tissue after challenge with MERS-CoV. Lung (A–C) and trachea (D–F) sections were assessed using rabbit polyclonal antibody to MERS-CoV nucleoprotein (NP) 3 days after the MERS-CoV challenge. The dark purple spot marked the inflammatory cell infiltration, and the brown particle marked the antigen of MERS-CoV. The MERS-CoV was located mainly in the trachea. Additionally, the lung tissue showed MERS-CoV expression in all immunized groups.
5. Discussion
The high case fatality rate, vaguely defined epidemiology, and absence of prophylactic or therapeutic measures against the MERS-CoV have created an urgent need for an effective vaccine [30], [34]. In this study, the rNTD protein located on the N terminal of S protein was investigated as a potential prophylactic vaccine against MERS-CoV infection. Our data showed that rNTD vaccination induced a sustained significant titer of IgG and neutralizing activity in the sera of immunized mice, although the titers were not as high as that found in rRBD-immunized mice as assessed by PRNT assay. Additionally, robust and functional T-cell-mediated immunity was achieved in rNTD-vaccinated mice, which was much higher than that in rRBD-vaccinated mice. This effect might be attributable to the prominent T-cell epitope located on the NTD, as Zhao et al. [31] showed KYYSIIPHSI (located in NTD) comprised the potential CD8+ T-cell epitope of the MERS-CoV S protein. Previously, we verified that the S protein peptide TIKYYSIIPHSIRSIQSD, located at aa residues 289–306, was the strongest T-cell epitope (supplementary Fig. 1) among 161 peptide pools overlapping with the S protein of MERS-CoV in BALB/c mice. As cytokines play an important role in the clearance of viruses in the body, we evaluated the expression of various cytokines in the rNTD-immunized mice using ICS and CBA assays. ICS revealed that rNTD vaccination elicited high levels of IL-2-positive CD4+ T-cells and IFN-γ-positive CD8+ T-cells in splenocytes, and CBA revealed evidence of a Th1, Th2, and Th17 T-cell response. As treatment with IFN improved the outcome in MERS-CoV-infected rhesus macaques [35], [36], we investigated whether rNTD-immunized mice benefited from the IFN-γ detected by ICS and CBA in the splenocytes of mice.
The immunity protection elicited by rNTD vaccination was evaluated in a MERS-CoV challenge mouse model that had been transfected with an adenoviral vector expressing human DPP4. As a control, the rRBD-immunized mice were challenged with MERS-CoV. The reduced abnormalities in the lung and trachea in recombinant protein-immunized mice suggested that either the rNTD or rRBD induced effective protection against the MERS-CoV infection in mice. Although both the rNTD and rRBD proteins induced significant immunity in mice, the orientation of the immunologic response differed in the two models. The rRBD protein induced a higher level of neutralizing activity by PRNT assay, whereas the rNTD protein induced a higher CMI level than did the rRBD. The challenge results showed that both rRBD and rNTD proteins could partially protect the immunized mice against MERS-CoV infection. The immunity protection elicited by the rNTD protein coincided with that elicited by the rRBD protein, suggesting that both the T-cell and antibody responses contributed to MERS-CoV clearance, a finding supported by other studies. Zhao et al. [31] showed no clearance of MERS-CoV in mice that lacked T-cells or in mice with severe combined immunodeficiency disease. In contrast, MERS-CoV was cleared in B cell-deficient mice, suggesting that T-cells were important for acute virus clearance. Additionally, the authors showed the accelerated kinetics of virus clearance in Ad5-hDPP4-transduced mice immunized with sera from Venezuelan equine encephalitis replicon particles expressing the MERS-CoV S protein, proving that the anti-S antibody blockage of virus attachment was largely sufficient for the MERS-CoV clearance effect. Finally, the study concluded that T-cells and antibodies were required for MERS-CoV clearance and protection from subsequent challenge.
Although the rNTD protein induced a humoral and cellular immune response in mice, there are several limitations in interpreting the data in this study. First, the mechanism of neutralization by rNTD vaccination in mice is unknown and requires further investigation. For example, the nature of the neutralization antibodies detected by the pseudovirus neutralization assay and PRNT assay remains unclear. Second, pathological lesions and antigen expression indicated by IHC were detected after the mice challenged by MERS-CoV, although significant humoral and cellular immunity was elicited by the rNTD vaccination. Whether this was the result of a memory response need further study. Additionally, the limitations of the animal models restricted the assessment of the protective efficacy of rNTD vaccine in mice [37]. The MERS-CoV infection animal model used in this study was developed by transducing mice with an adenovirus vector expressing human DPP4 [31]. MERS-CoV infection in this model is highly dependent on the transduction of cells and the level of DPP4 expression from the adenovirus vector; therefore, the model may not reflect the natural disease process [38]. Additionally, the 5-day animal model imposed the limitation that the viral titer data could not be collected for further investigation of the immune system response following clearance of the virus after rNTD vaccination in the mice. Other animal models, such as the transgenic mouse, might overcome this shortcoming. Whereas several studies have developed transgenic mouse models of MERS-CoV infection [39], [40], the non-specific damage and virus detection in the brain of transgenic mice restricted the application of the model. Other small animal models, such as ferrets [41], immune-deficient mice [42], or hamsters [43], were not susceptible to MERS-CoV infection.
Rodent animal models offer several advantages to the evaluation of vaccines, including the availability of animals, ease of handling, low cost, and the ability to use sufficient numbers for statistical analysis [44]. Thus, most investigators screen vaccines in small animal models first, and such screenings can prompt subsequent primate studies [44]. Several previous studies reported SARS vaccine candidates that elicited neutralizing antibodies and showed effectiveness in protecting young mice or hamsters from challenge [45], [46], [47], [48], [49]. However, reports of immunopathologic reactions in older mice and in NHPs vaccinated with SARS-CoV vaccines that were subsequently challenged with SARS-CoV [50], [51], [52] raised concerns of vaccine-associated disease enhancement before performing clinical trials with SARS-CoV vaccines [45]. Similar concerns have been reported in the vaccine-associated disease enhancement of feline infectious peritonitis virus [45], [53] and respiratory syncytial virus [54] vaccines. Therefore, it is necessary to evaluate the protection of the rNTD protein vaccine in a NHP model before applying it to humans to protect against MERS-CoV infection. Although there are many limitations, the rhesus macaque [55], [56] and common marmoset [57] have been developed as the NHP animal models of MERS-CoV infection.
This study showed that rNTD is a promising potential vaccine candidate against the MERS-CoV infection. To our knowledge, this is the first report examining a target other than RBD in S as a subunit vaccine against MERS-CoV infection. These findings contribute to our understanding of the function of NTD in the S protein of MERS-CoV and to the development of preventive or therapeutic strategies against MERS-CoV infection. This study also contributes to the development of a more suitable animal model for the evaluation of the protective efficiency of the rNTD protein vaccine and investigated a more effective target antigen to protect against MERS-CoV infection.
Competing interests
The authors have declared that no competing interests exist.
Funding
This work was supported by the National Key Research and Development Program of China (2016YFD0500301), the Megaproject for Infectious Disease Research of China (2014ZX10004001-002, 2012ZX10004501-004), National Natural Sciences Fund of China (81302607, 31400787) and Natural Sciences Fund of Hebei Province (H2013206108). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
The authors would like to thank Drs. Bart L.Haagmans and Ron A.M.Fouchier (Erasmus MC, Rotterdam, the Netherlands) for providing MERS-CoV (isolate hCoV-EMC/2012). We also thank Drs. Zhao Jincun and Perlman S (University of Iowa, Iowa City, IA) for providing rAd5-hDPP4 stock.
Footnotes
Supplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/j.vaccine.2016.11.064.
Appendix A. Supplementary material
Supplementary Fig. 1.
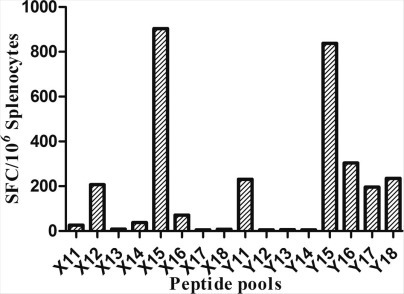
Synthesized peptide pools spanned the entire S protein were consisted of 161 short peptides. Each peptide was 18-mer and overlapped by 10 amino acids. Splenocytes of mice immunized with the recombinant Ad5 vector-based vaccines expressing full-length MERS-CoV S protein were collected for the detection of T lymphocyte immunodominant epitopes by an ELISpot assay. The represented results were shown in this figure. The highest level of T cell responses was induced by the peptide pools of X15 and Y15. Both of the two peptide pools contained the peptide of 37 which sequence of aa was TIKYYSIIPHSIRSIQSD. Thus this peptide was one of the T lymphocyte immunodominant epitopes of S protein.
References
- 1.Zaki A.M., van Boheemen S., Bestebroer T.M., Osterhaus A.D., Fouchier R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med. 2012;367:1814–1820. doi: 10.1056/NEJMoa1211721. [DOI] [PubMed] [Google Scholar]
- 2.van Boheemen S., de Graaf M., Lauber C., Bestebroer T.M., Raj V.S., Zaki A.M. Genomic characterization of a newly discovered coronavirus associated with acute respiratory distress syndrome in humans. MBio. 2012;3 doi: 10.1128/mBio.00473-12. [pii: e00473-12] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Peiris J.S., Yuen K.Y., Osterhaus A.D., Stohr K. The severe acute respiratory syndrome. N Engl J Med. 2003;349:2431–2441. doi: 10.1056/NEJMra032498. [DOI] [PubMed] [Google Scholar]
- 4.Zumla A., Hui D.S., Perlman S. Middle East respiratory syndrome. The Lancet. 2015;386:995–1007. doi: 10.1016/S0140-6736(15)60454-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Li Y., Wan Y., Liu P., Zhao J., Lu G., Qi J. A humanized neutralizing antibody against mers-cov targeting the receptor-binding domain of the spike protein. Cell Res. 2015;25:1237–1249. doi: 10.1038/cr.2015.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Tang X.C., Agnihothram S.S, Jiao Y., Stanhope J., Graham R.L., Peterson E.C. Identification of human neutralizing antibodies against MERS-CoV and their role in virus adaptive evolution. Proc Natl Acad Sci USA. 2014;111:E2018–E2026. doi: 10.1073/pnas.1402074111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Pascal K.E., Coleman C.M., Mujica A.O., Kamat V., Badithe A., Fairhurst J. Pre- and postexposure efficacy of fully human antibodies against spike protein in a novel humanized mouse model of MERS-CoV infection. Proc Natl Acad Sci USA. 2015;112:8738–8743. doi: 10.1073/pnas.1510830112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ying T., Du L., Ju T.W., Prabakaran P., Lau C.C., Lu L. Exceptionally potent neutralization of Middle East respiratory syndrome coronavirus by human monoclonal antibodies. J Virol. 2014;88:7796–7805. doi: 10.1128/JVI.00912-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Johnson R.F., Bagci U., Keith L., Tang X., Mollura D.J., Zeitlin L. 3B11-N, a monoclonal antibody against MERS-CoV, reduces lung pathology in rhesus monkeys followingintratracheal inoculation of MERS-CoV Jordan-n3/2012. Virology. 2016;490:49–58. doi: 10.1016/j.virol.2016.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Taguchi F. Coronaviruses. Uirusu. 2011;61:205–210. doi: 10.2222/jsv.61.205. [DOI] [PubMed] [Google Scholar]
- 11.Li F. Evidence for a common evolutionary origin of coronavirus spike protein receptor-binding subunits. J Virol. 2012;86:2856–2858. doi: 10.1128/JVI.06882-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Li F. Receptor recognition mechanisms of coronaviruses: a decade of structural studies. J Virol. 2015;89:1954–1964. doi: 10.1128/JVI.02615-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Williams R.K., Jiang G.S., Holmes K.V. Receptor for mouse hepatitis virus is a member of the carcinoembryonic antigen family of glycoproteins. Proc Natl Acad Sci USA. 1991;88:5533–5536. doi: 10.1073/pnas.88.13.5533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Li W., Moore M.J., Vasilieva N., Sui J., Wong S.K., Berne M.A. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426:450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Delmas B., Gelfi J., L’Haridon R., Vogel L.K., Sjöström H., Norén O. Aminopeptidase N is a major receptor for the entero-pathogenic coronavirus TGEV. Nature. 1992;357:417–420. doi: 10.1038/357417a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lu G., Hu Y., Wang Q., Qi J., Gao F., Li Y. Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor cd26. Nature. 2013;500:227–231. doi: 10.1038/nature12328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Raj V.S., Mou H., Smits S.L., Dekkers D.H., Müller M.A., Dijkman R. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature. 2013;495:251–254. doi: 10.1038/nature12005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Du L., Zhao G., Kou Z., Ma C., Sun S., Poon V.K. Identification of a receptor-binding domain in the s protein of the novel human coronavirus Middle East respiratory syndrome coronavirus as an essential target for vaccine development. J Virol. 2013;87:9939–9942. doi: 10.1128/JVI.01048-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lan J., Deng Y., Chen H., Lu G., Wang W., Guo X. Tailoring subunit vaccine immunity with adjuvant combinations and delivery routes using the Middle East respiratory coronavirus (MERS-CoV) receptor-binding domain as an antigen. PLoS ONE. 2014;9:e112602. doi: 10.1371/journal.pone.0112602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Tang J., Zhang N., Tao X., Zhao G., Guo Y., Tseng C.T. Optimization of antigen dose for a receptor-binding domain-based subunit vaccine against MERS coronavirus. Hum Vaccin Immunother. 2015;11:1244–1250. doi: 10.1080/21645515.2015.1021527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhang N., Tang J., Lu L., Jiang S., Du L. Receptor-binding domain-based subunit vaccines against MERS-CoV. Virus Res. 2015;202:151–159. doi: 10.1016/j.virusres.2014.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Guo X., Deng Y., Chen H., Lan J., Wang W., Zou X. Systemic and mucosal immunity in mice elicited by a single immunization with human adenovirus type 5 or 41 vector-based vaccines carrying the spike protein of Middle East respiratory syndrome coronavirus. Immunology. 2015;145:476–484. doi: 10.1111/imm.12462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kim E., Okada K., Kenniston T., Raj V.S., AlHajri M.M., Farag E.A. Immunogenicity of an adenoviral-based middle east respiratory syndrome coronavirus vaccine in balb/c mice. Vaccine. 2014;32:5975–5982. doi: 10.1016/j.vaccine.2014.08.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Song F., Fux R., Provacia L.B., Volz A., Eickmann M., Becker S. Middle East respiratory syndrome coronavirus spike protein delivered by modified vaccinia virus ankara efficiently induces virus-neutralizing antibodies. J Virol. 2013;87:11950–11954. doi: 10.1128/JVI.01672-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Malczyk A.H., Kupke A., Prufer S., Scheuplein V.A., Hutzler S., Kreuz D. A highly immunogenic and protective Middle East respiratory syndrome coronavirus vaccine based on a recombinant measles virus vaccine platform. J Virol. 2015;89:11654–11667. doi: 10.1128/JVI.01815-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lan J., Yao Y., Deng Y., Chen H., Lu G., Wang W. Recombinant receptor binding domain protein induces partial protective immunity in rhesus macaques against Middle East respiratory syndrome coronavirus challenge. EBioMedicine. 2015;2:1438–1446. doi: 10.1016/j.ebiom.2015.08.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Coleman C.M., Liu Y.V., Mu H., Taylor J.K., Massare M., Flyer D.C. Purified coronavirus spike protein nanoparticles induce coronavirus neutralizing antibodies in mice. Vaccine. 2014;32:3169–3174. doi: 10.1016/j.vaccine.2014.04.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Volz A., Kupke A., Song F., Jany S., Fux R., Shams-Eldin H. Protective efficacy of recombinant modified vaccinia virus Ankara delivering middle east respiratory syndrome coronavirus spike glycoprotein. J Virol. 2015;89:8651–8656. doi: 10.1128/JVI.00614-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Haagmans B.L., van den Brand J.M., Raj V.S., Volz A., Wohlsein P., Smits S.L. An orthopoxvirus-based vaccine reduces virus excretion after MERS-CoV infection in dromedary camels. Science. 2016;351:77–81. doi: 10.1126/science.aad1283. [DOI] [PubMed] [Google Scholar]
- 30.Wang L., Shi W., Joyce M.G., Modjarrad K., Zhang Y., Leung K. Evaluation of candidate vaccine approaches for MERS-CoV. Nat Commun. 2015;6:7712. doi: 10.1038/ncomms8712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhao J., Li K., Wohlford-Lenane C., Agnihothram S.S., Fett C., Zhao J. Rapid generation of a mouse model for middle east respiratory syndrome. Proc Natl Acad Sci USA. 2014;111:4970–4975. doi: 10.1073/pnas.1323279111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yang Y., Deng Y., Wen B., Wang H., Meng X., Lan J. The amino acids 736–761 of the MERS-CoV spike protein induce neutralizing antibodies: implications for the development of vaccines and antiviral agents. Viral Immunol. 2014;27:543–550. doi: 10.1089/vim.2014.0080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Guan J., Deng Y., Chen H., Yin X., Yang Y., Tan W. Priming with two DNA vaccines expressing hepatitis c virus ns3 protein targeting dendritic cells elicits superior heterologous protective potential in mice. Arch Virol. 2015;160:2517–2524. doi: 10.1007/s00705-015-2535-7. [DOI] [PubMed] [Google Scholar]
- 34.Muthumani K., Falzarano D., Reuschel E.L., Tingey C., Flingai S., Villarreal D.O. A synthetic consensus anti-spike protein DNA vaccine induces protective immunity against Middle East respiratory syndrome coronavirus in nonhuman primates. Sci Transl Med. 2015;7:301ra132. doi: 10.1126/scitranslmed.aac7462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Falzarano D., de Wit E., Rasmussen A.L., Feldmann F., Okumura A., Scott D.P. Treatment with interferon-α2b and ribavirin improves outcome in MERS-CoV-infected rhesus macaques. Nat Med. 2013;19:1313–1317. doi: 10.1038/nm.3362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Omrani A.S., Saad M.M., Baig K., Bahloul A., Abdul-Matin M., Alaidaroos A.Y. Ribavirin and interferon alfa-2a for severe Middle East respiratory syndrome coronavirus infection: a retrospective cohort study. Lancet Infect Dis. 2014;14:1090–1095. doi: 10.1016/S1473-3099(14)70920-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Du L., Jiang S. Middle East respiratory syndrome: current status and future prospects for vaccine development. Exp Opin Biol Ther. 2015;15:1647–1651. doi: 10.1517/14712598.2015.1092518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Falzarano D., de Wit E., Feldmann F., Rasmussen A.L., Okumura A., Peng X. Infection with MERS-CoV causes lethal pneumonia in the common marmoset. PLoS Pathog. 2014;10:e1004250. doi: 10.1371/journal.ppat.1004250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Agrawal A.S., Garron T., Tao X., Peng B.H., Wakamiya M., Chan T.S. Generation of a transgenic mouse model of Middle East respiratory syndrome coronavirus infection and disease. J Virol. 2015;89:3659–3670. doi: 10.1128/JVI.03427-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tao X., Garron T., Agrawal A.S., Algaissi A., Peng B.H., Wakamiya M. Characterization and demonstration of the value of a lethal mouse model of Middle East respiratory syndrome coronavirus infection and disease. J Virol. 2015;90:57–67. doi: 10.1128/JVI.02009-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Raj V.S., Smits S.L., Provacia L.B., van den Brand J.M., Wiersma L., Ouwendijk W.J. Adenosine deaminase acts as a natural antagonist for dipeptidyl peptidase 4-mediated entry of the Middle East respiratory syndrome corona-virus. J Virol. 2014;88:1834–1838. doi: 10.1128/JVI.02935-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Coleman C.M., Matthews K.L., Goicochea L., Frieman M.B. Wild-type and innate immune-deficient mice are not susceptible to the Middle East respiratory syndrome coronavirus. J Gen Virol. 2014;95:408–412. doi: 10.1099/vir.0.060640-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.de Wit E., Prescott J., Baseler L., Bushmaker T., Thomas T., Lackemeyer M.G. The Middle East respiratory syndrome coronavirus (MERS-CoV) does not replicate in Syrian hamsters. PloS One. 2013:e69127. doi: 10.1371/journal.pone.0069127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sutton T.C., Subbarao K. Development of animal models against emerging coronaviruses: from SARS to MERS coronavirus. Virology. 2015;479–480:247–258. doi: 10.1016/j.virol.2015.02.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gretebeck L.M., Subbarao K. Animal models for SARS and MERS coronaviruses. Curr Opin Virol. 2015;13:123–129. doi: 10.1016/j.coviro.2015.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lamirande E.W., DeDiego M.L., Roberts A., Jackson J.P., Alvarez E., Sheahan T. A live attenuated severe acute respiratory syndrome coronavirus is immunogenic and efficacious in golden Syrian hamsters. J Virol. 2008;82:7721–7724. doi: 10.1128/JVI.00304-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Enjuanes L., Dediego M.L., Alvarez E., Deming D., Sheahan T., Baric R. Vaccines to prevent severe acute respiratory syndrome coronavirus-induced disease. Virus Res. 2008;133:45–62. doi: 10.1016/j.virusres.2007.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.See R.H., Zakhartchouk A.N., Petric M., Lawrence D.J., Mok C.P., Hogan R.J. Comparative evaluation of two severe acute respiratory syndrome (SARS) vaccine candidates in mice challenged with SARS coronavirus. J Gen Virol. 2006;87:641–650. doi: 10.1099/vir.0.81579-0. [DOI] [PubMed] [Google Scholar]
- 49.Spruth M., Kistner O., Savidis-Dacho H., Hitter E., Crowe B., Gerencer M. A double-inactivated whole virus candidate SARS coronavirus vaccine stimulates neutralising and protective antibody responses. Vaccine. 2006;24:652–661. doi: 10.1016/j.vaccine.2005.08.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lokugamage K.G., Yoshikawa-Iwata N., Ito N., Watts D.M., Wyde P.R., Wang N. Chimeric coronavirus-like particles carrying severe acute respiratory syndrome coronavirus (SCoV) S protein protect mice against challenge with SCoV. Vaccine. 2008;26:797–808. doi: 10.1016/j.vaccine.2007.11.092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yasui F., Kai C., Kitabatake M., Inoue S., Yoneda M., Yokochi S. Prior immunization with severe acute respiratory syndrome (SARS)-associated coronavirus (SARS-CoV) nucleocapsid protein causes severe pneumonia in mice infected with SARS-CoV. J Immunol. 2008;181:6337–6348. doi: 10.4049/jimmunol.181.9.6337. [DOI] [PubMed] [Google Scholar]
- 52.Tseng C.T., Sbrana E., Iwata-Yoshikawa N., Newman P.C., Garron T., Atmar R.L. Immunization with SARS coronavirus vaccines leads to pulmonary immunopathology on challenge with the SARS virus. PLoS ONE. 2012;7:e35421. doi: 10.1371/journal.pone.0035421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Takano T., Kawakami C., Yamada S., Satoh R., Hohdatsu T. Antibody-dependent enhancement occurs upon re-infection with the identical serotype virus in feline infectious peritonitis virus infection. J Vet Med Sci. 2008;70:1315–1321. doi: 10.1292/jvms.70.1315. [DOI] [PubMed] [Google Scholar]
- 54.Graham B.S. Vaccines against respiratory syncytial virus: the time has finally come. Vaccine. 2016;34:3535–3541. doi: 10.1016/j.vaccine.2016.04.083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.de Wit E., Rasmussen A.L., Falzarano D., Bushmaker T., Feldmann F., Brining D.L. Middle East respiratory syndrome coronavirus (MERS-CoV) causes transient lower respiratory tract infection in rhesus macaques. Proc Natl Acad Sci USA. 2013;110:16598–16603. doi: 10.1073/pnas.1310744110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Munster V.J., de Wit E., Feldmann H. Pneumonia from human coronavirus in a Macaque model. New Engl J Med. 2013;368:1560–1562. doi: 10.1056/NEJMc1215691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Falzarano D. Infection with MERS-CoV causes lethal pneumonia in the common Marmoset. PLoS Pathog. 2014;10:e1004250. doi: 10.1371/journal.ppat.1004250. [DOI] [PMC free article] [PubMed] [Google Scholar]