

Abstract
Vaccine-induced antibodies can prevent or, in the case of feline infectious peritonitis virus, aggravate infections by coronaviruses. We investigated whether a recombinant native full-length S-protein trimer (triSpike) of severe acute respiratory syndrome coronavirus (SARS-CoV) was able to elicit a neutralizing and protective immune response in animals and analyzed the capacity of anti-S antibodies to mediate antibody-dependent enhancement (ADE) of virus entry in vitro and enhancement of replication in vivo. SARS-CoV-specific serum and mucosal immunoglobulins were readily detected in immunized animals. Serum IgG blocked binding of the S-protein to the ACE2 receptor and neutralized SARS-CoV infection in vitro. Entry into human B cell lines occurred in a FcγRII-dependent and ACE2-independent fashion indicating that ADE of virus entry is a novel cell entry mechanism of SARS-CoV. Vaccinated animals showed no signs of enhanced lung pathology or hepatitis and viral load was undetectable or greatly reduced in lungs following challenge with SARS-CoV. Altogether our results indicate that a recombinant trimeric S protein was able to elicit an efficacious protective immune response in vivo and warrant concern in the safety evaluation of a human vaccine against SARS-CoV.
Keywords: SARS-CoV, Protection, ADE
1. Introduction
Coronavirus infections of the human respiratory tract occur frequently. Infections by HCoV-229E and HCoV-OC43 account for 15% of all common colds in humans and recently identified human coronavirus NL63 and HKU1 are associated with acute respiratory distress syndrome and pneumonia [1], [2]. While most coronavirus infections are mild and self-limiting, infections by severe acute respiratory syndrome coronavirus (SARS-CoV) result in high mortality and morbidity [3]. There is currently no vaccine available against any human coronavirus infection.
The viral surface Spike glycoprotein (S) is the key target for protective neutralizing antibodies against coronaviruses. The S-protein is a 150–180 kDa highly glycosylated trimeric class-I fusion protein [4], [5] responsible for receptor binding, virus-membrane fusion and tissue tropism. Angiotensin converting enzyme 2 (ACE2) has been identified as the functional receptor for virus entry into susceptible target cells [6], [7], but C-type lectin L-SIGN (CD209L) has also been shown to mediate virus uptake [8]. S-mediated viral entry occurs in a pH-dependent manner [9], [10] and can be inhibited by S-specific sera [11]. Neutralizing antibodies are broadly elicited in patients with SARS [12] and passive transfer of S-specific antibodies, including monoclonal antibodies directed against the receptor-binding domain (RBD) of S [13], to naïve animals confer protection to SARS-CoV challenge indicating that the humoral immune response alone can protect against SARS-CoV [14], [15], [16], [17], [18].
SARS vaccine candidates should induce strong protective immunity against infection and person-to-person transmission which requires a humoral immune response at both systemic and mucosal levels and absence of immunopathology. Different vaccine candidates have been explored for SARS including DNA [19], Modified Vaccinia virus Ankara (MVA) [16] subunit vaccine [20] or inactivated virus vaccine [21], [22], [23] and they all have been shown to induce neutralizing and protective responses. While protection was achieved in the mouse with a recombinant MVA virus expressing S protein [16], vaccination with a different MVA-S construct failed to elicit a sustained serum neutralizing antibody titer to SARS-CoV in ferrets and subsequent challenge with SARS-CoV was associated with hepatitis [24]. In vitro data also demonstrate that anti-S immune responses can enhance SARS-CoV entry into target cells. Yang and colleagues showed that entry of SARS-CoV pseudotype viruses, expressing the palm civet S-protein, into a human renal epithelial cell-line was enhanced by human S-specific neutralizing antibodies [25]. Such in vitro detectable facilitating antibodies can be correlated with disease enhancement in feline infectious peritonitis virus-infected cats (FIPV) and it has been shown that vaccine-induced anti-S antibodies and macrophage-expressed Fc receptors (FcR) are key factors of antibody-dependent enhancement (ADE) [26], [27] in cats.
Animal models for SARS are useful to evaluate the neutralizing or enhancing capacity of antibodies in vivo. Several animal models have been proposed for SARS including mice, ferrets, hamsters, rhesus and cynomologus macaques [15], [28], [29], [30]. The hamster is an interesting small animal model as it reproduces several pathological features of human SARS including high titer replication of SARS-CoV in lungs and nasal turbinates and marked lung pathology including cellular necrosis and inflammatory cell infiltrates [30]. In the present study, we analyzed the immune response to a recombinant native full-length S protein trimer of SARS-CoV. Neutralizing and enhancing activity could be detected in sera of immunized animals and human convalescent SARS patient sera in vitro while challenge experiments in hamsters indicate complete protection and absence of disease enhancement in vivo.
2. Materials and methods
2.1. Cell lines and expression vectors
Human Raji B and BHK-21 cells were cultured as described previously [31], [32]. HEK293T, VeroE6, FRhK-4 and murine macrophagic J774A1 cell lines were cultured at 37 °C, 5% CO2, in Dulbecco's modified Eagle medium supplemented with 10% FBS, 100 U penicillin/ml, 100 μg streptomycin/ml. Optimized DNA and Semliki Forest Virus expression vectors encoding SARS-CoV S-protein fused to a C-terminal FLAG-sequence were described previously [33].
2.2. Immunoaffinity purification and biochemical analysis of S-protein
For protein production BHK cells were cultured in the presence or absence of cycloheximide (100 μg/ml), added at 7 h pre-harvest. Fourteen hours post-infection/transfection BHK-21 cells were lysed (20 mM Tris–HCl pH 7.5, 150 mM NaCl, 2 mM EDTA, 1% Triton X-100) and incubated for 5 min on ice. The collected lysate was vortexed, incubated for another 15 min on ice prior to centrifugation at 13,000 rpm for 15 min. Recombinant S-protein was immunopurified using anti-FLAG M2 mAb-coated agarose beads (Sigma) as described previously [31]. The quantity and quality of recombinant S-protein was assessed by SDS-PAGE and Western Blot using BAP-FLAG protein and microBSA methods as standards for protein quantification [31], [32]. Mouse sera were diluted (1/500) prior to Western Blot analysis and immune complexes detected with HRP-conjugated anti-mouse IgG (H + L) (1/1000) (Zymed), followed by visualization of the bands on X-ray film (Kodak) using chemiluminescence (Amersham Biosciences).
2.3. Immunization with triSpike
Balb/c mice 6–8 weeks old (n = 3 per group) were immunized intraperitoneally (i.p.) with 20 μg of purified triSpike or triSpike in 1 mg of aluminium hydroxide gel (alum) on days 0, 16 and 32. Animals in the control group received PBS with 1 mg of alum on the same days. Blood samples were collected by saphenous vein bleeding at indicated time points in accordance with local guidelines and sera were prepared and heat-inactivated. Fecal samples [34] and nasal lavages [35] were prepared as described previously. HRP-conjugated anti-mouse IgG (H + L) or IgA (1/1000) (Zymed) were used to detect the existence of IgG or IgA antibody against triSpike in immunized mice fecal and nasal lavage sample. Absence of blood contamination was confirmed using the Hema-Screen occult blood detection kit (Stanbio Laboratory). Antibodies from fecal samples were used at a final dilution of 1/500 and nasal lavage sample at 1/25 dilution for Western Blot analysis.
Six to eight week old female golden Syrian hamsters (n = 4 per group) were immunized subcutaneously (s.c.) with 2, 10 or 20 μg of purified triSpike in 2 mg of alum on d0, d21 and d42 at HKU-Pasteur Research Centre, Hong Kong, China. A parallel experiment in 5-week-old hamsters was performed at the NIAID, NIH, Bethesda, Maryland with immunization of 2, 10 or 50 μg of purified triSpike in 1 mg of alum on d0, d21 and d42.
2.4. Flow cytometry and ACE2 binding assay
Recombinant S-protein expressing BHK-21 cells were detached with 5 mM EDTA and incubated for 45 min at 4 °C with the diluted hamster sera (1/100), washed, fixed with 3.2% of PFA for 5 min at 4 °C and labeled with the FITC-conjugated goat anti-Syrian hamster IgG (Santa Cruz Biotechnology) for 30 min at 4 °C. Cells were analyzed by flow cytometer (FACSCalibur, BD). All steps were blocked with 3% normal goat serum (Zymed). Fluorescence values are expressed as mean fluorescence intensity (MFI). ACE2 binding assays were performed by incubating soluble ACE2 receptor with S-protein coated on agarose beads as described previously [33]. For inhibition of binding analysis, protein-coated beads were pre-incubated with sera for 1 h at 4 °C before incubation with soluble ACE2. The beads were washed four times with lysis buffer (20 mM Tris–HCl 7.5, 150 mM NaCl, 2 mM EDTA, 1% Triton X-100) prior to SDS-PAGE and Western Blot analysis.
2.5. Serum-neutralization assay
Neutralization assays were carried on FRhK-4 cells using SARS-CoV strain HKU-39849 (Department of Microbiology, HKU) as follows. One hundred TCID50 of SARS-CoV were incubated for 2 h at 37 °C with serial 2-fold dilutions of heat-inactivated mouse sera in quadruplicate. Virus antibody mix was then added to cells in 96-well plates and plates were incubated at 37 °C with microscopic examination for cytopathic effect (cpe) after a 4-day incubation. Neutralization titers were calculated by the Reed & Muench formula and are expressed as the reciprocal of the serum dilution which neutralized cpe in 50% of the wells [36].
2.6. SARS-CoV pseudotype particles
Recombinant SARS-CoVpp lentiviral vectors expressing a luciferase reporter gene were produced from HEK293T cells as described previously [37] using 10 μg of plasmid pNL4.3.Luc R−E−pro− [11] and 10 μg of plasmid pCDNA-S-FLAG encoding codon-optimized SARS-CoV S protein [33]. For ADE assays, 25 μL of serial 2-fold dilutions of heat-inactivated murine or hamster sera were incubated for 1 h at 37 °C with 25 μL of pseudovirus (50 ng p24). Fifty microliters of Raji cells, at 2 × 106 cell/mL previously washed three times with serum-free RPMI, were added to the antibody-SARS-CoVpp mixture in a 96-well plate. After adsorption for 1 h at 37 °C, 100 μL of RPMI 1640 containing 5% FCS were added. Medium was renewed 16 h later and incubated for an additional 48 h, washed in PBS, lysed and luciferase activity measured for 10 sec in a MicroBeta Jet Counter (Perkin Elmer) according to the instructions provided by the supplier (Promega). For ADE blocking assays, Raji cells were pre-incubated for 15 min at 4 °C with 10 μg/mL of murine monoclonal antibody directed against human CD32 (FcγRII, BD Pharmingen) or goat polyclonal antibody directed against human ACE2 (R&D Systems) prior to infection with SARS-CoVpp.
2.7. SARS-CoV challenge and viral load titration in hamsters, histopathology and blood chemistry
On day 56 (14 days following the last immunization), 48 anesthetized hamsters received 100 μl (103 TCID50) of SARS-CoV (strain Urbani) intranasally (Table 1 ). Hamsters were euthanatized by lethal intraperitoneal injection with sodium pentobarbital (200 μl/hamster) on days 2 and 5 following challenge when peak virus replication, and pneumonitis are anticipated and on day 21 post challenge to look for possible late changes in histopathology. Sixteen of forty eight SARS-CoV-inoculated hamsters were sacrificed 2 days post-challenge. Lungs were harvested and processed for viral titration. Groups of 16 SARS-CoV-inoculated hamsters were sacrificed 5 and 21 days post-challenge and lungs and livers were harvested and processed for pathology studies. Tissue samples were homogenized and virus titers were determined in VeroE6 cell monolayers as described previously [30].
Table 1.
| Group (# Hms) | Days 0, 21, 42 Inoculum (dose) | Day 56 SARS | Day of sacrifice | Organs harvested for |
|---|---|---|---|---|
| 1 (4) | PBS | IN | 58 | Viral titers |
| 2 (4) | ST 2 μg | IN | 58 | Viral titers |
| 3 (4) | ST 10 μg | IN | 58 | Viral titers |
| 4 (4) | ST 50 μg | IN | 58 | Viral titers |
| 5 (4) | PBS | IN | 61 | Pathology (in 10% buffered formalin) |
| 6 (4) | ST 2 μg | IN | 61 | Pathology (in 10% buffered formalin) |
| 7 (4) | ST 10 μg | IN | 61 | Pathology (in 10% buffered formalin) |
| 8 (4) | ST 50 μg | IN | 61 | Pathology (in 10% buffered formalin) |
| 9 (4) | PBS | IN | 77 | Pathology (in 10% buffered formalin) |
| 10 (4) | ST 2 μg | IN | 77 | Pathology (in 10% buffered formalin) |
| 11 (4) | ST 10 μg | IN | 77 | Pathology (in 10% buffered formalin) |
| 12 (4) | ST 50 μg | IN | 77 | Pathology (in 10% buffered formalin) |
Four hamsters per group, immunized and challenged as described above, were sacrificed at 5 days post-challenge and an additional four hamsters per group (with one additional hamster in the 50 μg/dose group) were sacrificed 21 days following challenge. Lungs were evaluated for pneumonitis and consolidation as previously described [30]. A colorimetric immunoalkaline phosphatase IHC method was used to identify SARS-CoV antigens [30]. Hamster serum samples were analyzed for the levels of aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), gamma-glutamyl transferase (GGT), blood urea nitrogen (BUN) and total bilirubin using blood chemistry analyzer (Analytics, MedTec Lab).
3. Results
3.1. Production of immunopurified trimeric S-protein with native antigenicity
Full-length, codon-optimized SARS-CoV S-protein fused to a C-terminal FLAG was purified by a single-step immunoaffinity protocol to over 90% purity with an average yield of 3 μg of pure S-protein per 106 cells, i.e. 1.5 mg/L (Fig. 1A). Analysis of the apparent molecular weight of the protein by SDS-PAGE and Western Blot under non-reducing conditions revealed the predominant trimeric nature of the antigen (Fig. 1B, lane 1). Trimers dissociate into monomers when the protein is heat-denatured in the presence of SDS indicating that monomers are associated non-covalently (Fig. 1B, lane 2). Higher molecular weight DTT-sensitive aggregates were observed when the protein was not heat denatured prior to SDS-PAGE (Fig. 1B, lane 3). As expected, trimers and aggregates dissociate completely into monomers when heat-denatured in SDS and DTT (Fig. 1B, lane 4). Monomers derived from SDS/heat-denatured trimers have a slightly greater electrophoretic mobility than monomers derived from SDS/heat/DTT denatured trimers indicating that intramolecular disulfide bonds lead to a more compact three dimensional conformation of the Spike monomer. The trimeric and monomeric S-protein frequently migrate as doublets (Fig. 1B) [33]. These represent high-mannose glycoforms from proteins that reside in the ER at the time of lysis and glycoforms from proteins that have acquired complex N-glycans in the median-Golgi [33]. triSpike has native antigenicity shown by reactivity with sera from 10 convalescent SARS patients by Western Blot and FACS (data not shown). The native fold was further underscored by the specific binding of the triSpike protein with soluble ACE2 receptor (Fig. 2E, lane 2). Altogether our results strongly argue that purified triSpike molecules mimic the native trimeric S-protein on the virion surface.
Fig. 1.
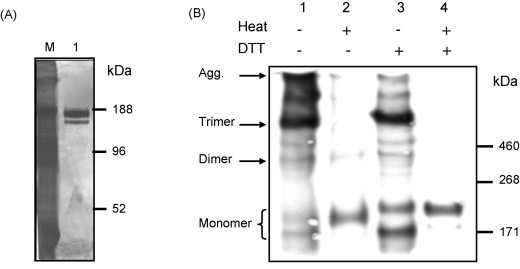
Biochemical characterization of purified trimeric SARS-CoV S-protein (triSpike). (A) Silver staining of immunopurified triSpike, lane M contains a pre-stained protein marker and lane 1 had 720 ng of purified triSpike. (B) triSpike was expressed in BHK-21 cells in the presence of cycloheximide. Recombinant triSpike purification was performed by immunoaffinity as described in Section 2. Eluted protein was treated as indicated and analyzed by 4–8% Tris-Acetate SDS-PAGE gel and Western Blot using M2 monoclonal antibody against the FLAG peptide. Sizes of molecular weight markers are indicated on the right (HiMark™ Pre-stained HMW Protein Standard, Invitrogen).
Fig. 2.
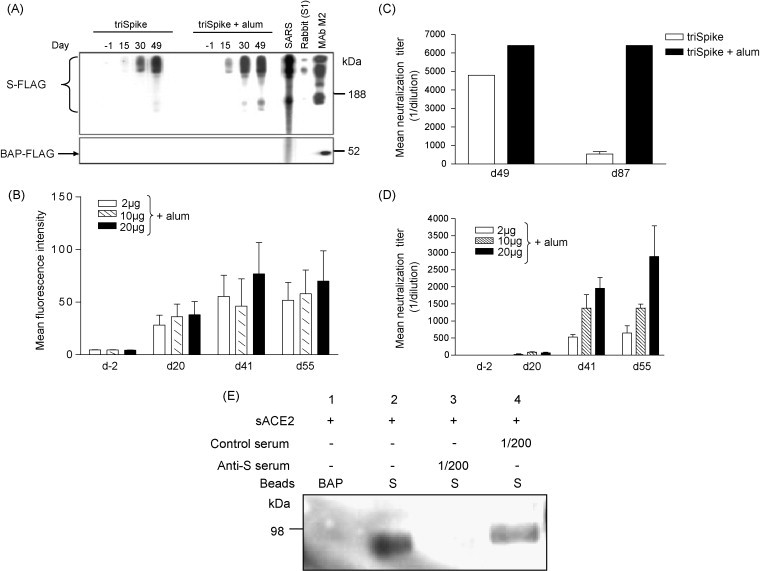
Immunogenicity of triSpike. Sera collected at indicated time points from vaccinated mice or hamsters were analyzed for reactivity with triSpike. (A) Western Blot analysis of pooled sera from immunized mice with or without alum adjuvant. Sera were collected and used at 1/1000 dilution against S-FLAG that was separated under conditions that allowed simultaneous detection of mono-, di-, and trimers of S-protein. A SARS patient serum (SARS), a rabbit serum against S1 and M2 monoclonal antibody against the FLAG peptide was used as control. FLAG-tagged bacterial alkaline phosphatase (BAP-FLAG) was used to assess the presence of antibodies against the FLAG tag. Immune complexes were detected with HRP-conjugated goat anti-mouse, -human or -rabbit IgG polyclonal antibody. Sizes of molecular weight markers are indicated on the right. (B) Reactivity of immune sera from pooled immunized hamster with live BHK-21 cells expressing S-protein at the plasma membrane using FACS analysis. Values were expressed as mean ± standard deviations. (C) Effect of alum adjuvant on longevity of neutralizing response in mouse sera against SARS-CoV (determined on FRhK-4 cells). Values were expressed as mean ± standard deviations. (D) Dose effect of triSpike immunization with alum adjuvant on neutralizing response against SARS-CoV in hamsters (determined on FRhK-4 cells). Values were expressed as mean ± standard deviations. (E) Inhibition of S-protein binding to ACE2 by sera from immunized mice. S-protein coated beads were pre-incubated with sera prior to incubation with soluble ACE2 (sACE2) and detection of the receptor was performed with a polyclonal goat-anti human ACE2 antibody. BAP-FLAG coated beads were used as control.
3.2. triSpike induces antibodies against SARS-CoV S-protein in mice and hamsters
Immunogenicity of triSpike was assessed by immunization with three different doses with or without adjuvant (alum) in mice or hamsters (Fig. 2). Immunization of mice with triSpike alone induced low levels of anti-S antibodies (Fig. 2A, dl5). However a single subsequent injection of triSpike induced detectable levels of antibodies against S-protein (Fig. 2A, d30) which could be further boosted by a third injection (Fig. 2A, d49). Sera were reactive against mono-, di- and trimers of S-proteins carrying either high-mannose or complex N-glycosylation. Analysis of individual serum samples from d49 confirmed the homogeneity of the antibody response in individual mice (data not shown). When triSpike was used in combination with alum adjuvant (Fig. 2A) a very similar response was observed, except that the level of anti-S IgG was slightly higher after the second injection (Fig. 2A, d30) and a third injection did not increase the anti-S serum antibody level (Fig. 2A, d49). Neither triSpike nor triSpike with alum adjuvant immunization induced antibodies directed against the FLAG peptide (Fig. 2A, lower panels).
In a next step we analyzed the dose response and booster immunization effect in hamsters, immunized with 2, 10 or 20 μg triSpike plus alum, by FACScan. The first immunization with all three doses resulted in a moderate anti-S antibody response (Fig. 2B, d20). A second injection strongly increased anti-S antibody levels (Fig. 2B, d41), while no further increase in anti-S antibody level was seen after a third injection (Fig. 2B, d55). There is no significant dose-dependent antibody response as 2 μg triSpike plus alum immunization seems to induce maximum antibody response in hamster model. Analysis of individual serum samples from d55 confirmed the homogeneity of the antibody response in individual hamster (data not shown). Altogether these results indicate that an optimal anti-S antibody response might require two doses of triSpike (2 μg) immunization in alum adjuvant.
3.3. Neutralizing antibodies in sera from triSpike vaccinated mice and hamsters
We next evaluated the neutralizing activity of sera from triSpike vaccinated mice. Serial dilutions of sera were tested for their neutralizing activity of cpe induced by SARS-CoV replication in FRhK-4 cells (neutralization of 100 TCID50). Injection of triSpike with alum adjuvant induces higher neutralization titers compared to triSpike alone (Fig. 2C). Antibody titers in triSpike with alum immunized animals remained at high levels until day 87 (1/6400) while neutralizing titers dropped rapidly from day 49 to day 87 in animals injected with triSpike alone (from 1/4800 to 1/600). Subsequently, a dose–response experiment was performed in hamsters. Sera from hamsters immunized with at least two doses as low as 2 μg effectively neutralized 100 TCID50 of SARS-CoV on FRhK-4 cells (Fig. 2D). Boosting effect with a third injection was limited with all three doses. Maximum antibody levels were obtained in animals immunized with 20 μg.
3.4. Neutralizing sera block spike binding to the ACE2 receptor
We next investigated the mechanism of neutralization by analyzing the capacity of sera to block the interaction between immunopurified triSpike coated on agarose beads with purified soluble human ACE2, the SARS-CoV entry receptor [6], [7]. Fig. 2E shows that sera from triSpike + alum immunized mice (lane 3) but not from control animals (lane 4) neutralized S-protein binding to ACE2 receptor. These results suggest inhibition of receptor binding as the key mechanism of neutralization in triSpike immunized animals.
3.5. Mucosal IgG and IgA response in mice
Detection of SARS-CoV in intestinal tissue of fatal SARS cases indicated that the gastrointestinal tract is another target for SARS-CoV infection [38]. Hence, a vaccine candidate should ideally induce protective response in mucosal sites. Previous study suggested the induction of IgG response in serum (systemic response) and IgA response in stool (mucosal response) samples against Yersinia pestis using intraperitoneal immunization [39]. To address this question antibody were prepared from fecal samples of immunized animals. Occult blood detection showed absence of blood contamination (data not shown) indicating that the samples were not contaminated by serum antibodies. Western Blot using antibodies isolated from fecal samples revealed that mucosal IgG and IgA were induced by triSpike plus alum but not by triSpike alone (Fig. 3A). These antibodies were able to neutralize SARS-CoV infection of FRhK-4 cells (Fig. 3C). Altogether, we conclude that triSpike plus alum immunization induced antigen-specific IgG and IgA in the gastrointestinal tract.
Fig. 3.
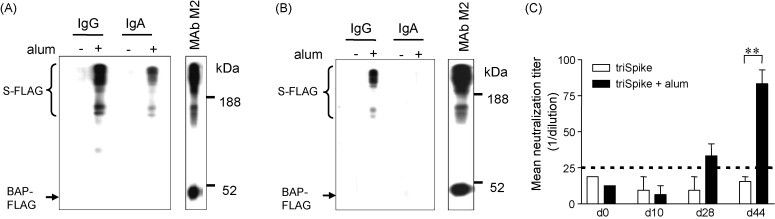
Induction of mucosal immune response in triSpike plus alum vaccinated mice. (A) Antibodies prepared from fecal samples collected at day 44 post-immunization were reacted against S-protein as described in Fig. 2A. Immune complexes were detected with HRP-conjugated goat anti-mouse IgG or IgA polyclonal antibody. (B) Similar as (A) except that Western Blot analysis was performed with pooled nasal lavage samples collected at day 65. (C). Antibodies prepared from fecal samples of vaccinated mice were analyzed for neutralizing activity against SARS-CoV infection on FRhK-4 cells. Weak neutralizing activity was detected after the third immunization only. Asterisk (**) indicates the value of p < 0.01 in two-tailed t tests. Values were expressed as mean ± standard deviations.
Mice were also analyzed for the presence of anti-S antibodies in nasal lavage samples. Occult blood detection indicated the absence of blood contamination in the sample. Western Blot analysis using antibodies isolated from nasal lavage revealed the induction of a mucosal IgG response only after triSpike plus alum immunization but not after triSpike immunization alone (Fig. 3B). We were unable to detect any IgA response in either group and anti-S antibodies in nasal lavage samples did not neutralize SARS-CoV (Fig. 3B). Further studies are required to address the respective role of IgA and IgG in the neutralizing activity, epitope specificity and antibody titer in the neutralization mechanism.
3.6. Antibody-dependent enhancement (ADE) of viral entry into human B cells
We investigated whether vaccine-induced anti-S antibodies against triSpike could mediate ADE of SARS-CoV entry into FcR expressing cell lines. SARS pseudotype particles (SARS-CoVpp) were generated for this study. These recombinant viruses encoding the luciferase reporter gene and expressing the SARS-CoV Spike protein at the virion surface have been shown to faithfully mimic the SARS-CoV entry process [9], [10], [11]. We infected cells with SARS-CoVpp in the presence or absence of heat-inactivated serum from triSpike immunized mice and hamsters as well as sera from ten convalescent SARS patients. Transduction of FcR-negative VeroE6 cells with SARS-CoVpp was reduced in a dose-dependent manner when pseudovirus was pre-incubated with S-specific antisera (Fig. 4B & C). When SARS-CoVpp were pre-incubated with heat-inactivated S-specific mouse or hamster sera a dose-dependent enhancement of ACE2-negative Raji B and Daudi cell transduction was observed (Fig. 4A–C). Only background level of transduction was detected in the absence of serum or in the presence of sera from the naïve control group. When Raji B cells were pre-incubated with a monoclonal antibody (mAb) against human FcγRII (CD32) SARS-CoVpp transduction was reduced to background levels demonstrating that pseudovirus entry was FcγRII dependent (Fig. 4D). Furthermore ADE could be abrogated by pre-incubating the pseudovirus-antibody mix with protein G, demonstrating that the process is mediated by the Fc portion of the antibody (data not shown). Interestingly, a mAb against ACE2 blocked transduction of VeroE6 cells by SARS-CoVpp but not on Raji B cells by SARS-CoVpp-antibody complexes indicating that FcγRII mediated virus entry does not require ACE2 for virus-cell membrane fusion (Fig. 4D). Only a low level of SARS-CoVpp transduction was observed with the J774A1 murine macrophage cell line despite the presence of FcγRII receptor. ADE was not limited to vaccine-induced anti-S sera from mice and hamsters. Eight out of ten sera from convalescent SARS patients also showed enhancement of Raji B cell transduction despite the capacity of 10/10 sera to neutralize virus entry into VeroE6 cells (data not shown). Our results demonstrate that S-specific antibodies can induce a shift in cell tropism of SARS-CoV towards human FcγRII expressing cells. Antibody- and FcγRII-dependent but ACE2-independent virus entry and fusion is a novel mechanism by which SARS-CoV can enter into target cells.
Fig. 4.
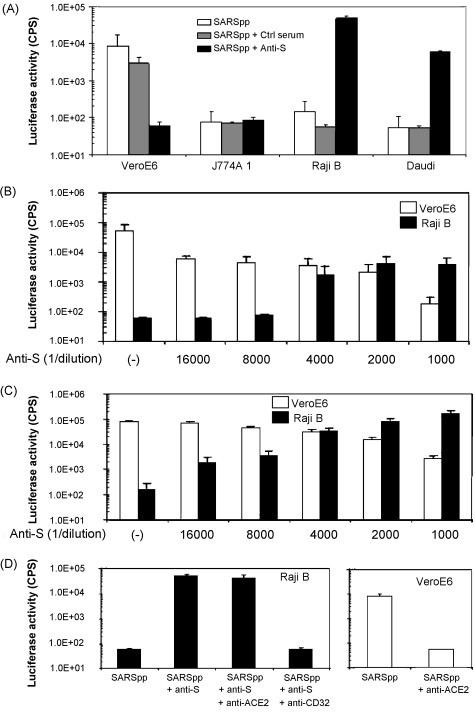
Analysis of antibody-dependent enhancement of SARS-CoVpp entry. (A) Analysis of various cell lines for ADE of Spike-mediated viral entry. SARS-CoVpp were incubated with a 1/1000 dilution of sera from mice immunized with triSpike and transduction was measured as SARS-CoVpp-expressed luciferase activity. (B) Serial dilutions of sera from mice immunized with triSpike neutralize SARS-CoVpp transduction of VeroE6 cells and enhance transduction of Raji B cells. (C) Same as (B) except that sera from hamsters immunized with triSpike were used. (D) FcγRII (CD32) involvement in antibody-dependent enhancement of SARS-CoVpp entry. Same as (C), SARS-CoVpp were incubated with a 1/1000 dilution of sera and Raji cells pre-incubated with 10 mg/mL of a mAb against ACE2 or CD32 (left panel) prior to infection. Inhibition of SARS-CoVpp entry by the mAb against ACE2 was controlled on VeroE6 cells (right panel). All experiments were performed in triplicates and data are presented as mean ± standard deviations.
3.7. Protection of immunized hamsters from SARS-CoV challenge
Syrian golden hamsters were immunized 3 times at bi-weekly intervals with 2 μg, 10 μg or 50 μg of triSpike plus alum and challenged intranasally with 103 TCID50/hamster of SARS-CoV. Hamsters receiving triSpike vaccine were significantly protected from SARS-CoV challenge compared to mock immunized controls (Fig. 5 ). Complete protection was observed following three doses of 50 μg (mean viral titer below the limit of detection) and partial protection was observed in hamsters receiving the lowest dose. Half the hamsters receiving three doses of 2 μg triSpike vaccine showed complete protection from challenge. Mean viral titers for the group were 102.3 TCID50/g tissue. Seventy-five percent of hamsters receiving three doses of 10 μg triSpike vaccine showed complete protection from challenge. One hamster for this group had viral titers just above the limit of detection at 102 TCIDso/g tissue. In comparison of four mock immunized hamsters had similar high titers of SARS-CoV present with a mean viral titer of 107.2 TCIDso/g tissue. Patterns or consistent elevations of liver enzymes were not observed in any group following any immunization or SARS-CoV challenge (data not shown).
Fig. 5.
Protection of triSpike-immunized hamsters from SARS-CoV challenge. Hamsters were inoculated intranasally with 103 TCID50 of SARS-CoV (Urbani strain) and virus titers in lung homogenates were determined. Error bars indicated standard errors and asterisk (***) indicate the value of p < 0.001 in two-tailed t tests. Values were expressed as TCID50 per g of lung tissue.
3.8. Histopathology
Five days following SARS-CoV challenge mock immunized hamsters had severe pneumonitis and consolidation in comparison to triSpike immunized hamsters where 9/12 had mild, 2/12 had moderate and only 1/12 had severe pneumonitis and none had pulmonary consolidation. The data indicate that regardless of the dose administered the vaccine significantly protects from the occurrence and severity of pneumonitis following SARS-CoV challenge. In contrast to previously published data where pneumonitis and consolidation were completely cleared by day 14 post-challenge, moderate (1/4) to severe pneumonitis (3/4) was observed 21 days post-challenge in mock immunized hamsters. In triSpike immunized hamsters 21 days post-challenge, pneumonitis was completely cleared (3/13) or reduced to mild (8/12) or moderate (2/12) levels. The variation between these observations and previously reported observations in the time to resolution of pneumonitis is likely due to changes in tissue processing. Lungs were inflated with formalin in these studies and were not in the previous report [30]. Nevertheless, consistent with previously reported data, 21 days after challenge pulmonary consolidation was absent in all groups including the mock immunized controls.
Histopathological analyses of livers at both days 5 (3/16) and 21 (4/17) post-challenge revealed very rare foci of hepatic cellular necrosis. These foci were observed in both mock immunized and triSpike immunized hamsters post-challenge and did not show any correlation with either pre-challenge neutralizing antibody titers or liver enzyme levels (pre- or post-challenge). Furthermore, SARS-CoV antigen was not detected by immunohistochemical staining.
4. Discussion
There is currently no licensed vaccine against a human coronavirus. Vaccines against animal coronaviruses have been successfully generated, e.g. transmissible gastroenteritis virus (TGEV) [40]. However, the development of a vaccine against FIPV has proven difficult due to immune enhancement of disease in vaccinated cats. Receptors binding viral envelope glycoproteins are prime targets for neutralizing antibodies [41]. We hypothesized that a SARS-CoV envelope with native trimeric conformation would be optimal for induction of conformation-dependent neutralizing antibodies targeting multiple epitopes. Full-length coronavirus Spike glycoproteins with an intact transmembrane domain were previously shown to assemble into rosettes generating highly immunogenic enveloped lipid spheres [42]. Biochemical characterization of the S-protein immunogen showed that purified recombinant S protein migrates in SDS-PAGE with an apparent molecular weight >500 kDa, suggesting that it is a trimer. The correctly folded S protein is a complex-glycosylated, non-covalently linked homotrimeric glycoprotein which is not cleaved by intracellular proteases during exit to the cell surface. These results are in agreement with data obtained by others for SARS [5] and other coronaviruses [4]. Immunopurified preparations of full-length S trimer (triSpike) contain correctly folded S protein but also mannosylated high molecular weight forms, which are likely ER retained proteins that are not yet correctly folded or being degraded. Indeed, high-mannose forms are purged from the ER, as evidenced by pulse-chase experiments [33] and by treatment of S expressing cells with translation inhibitor cycloheximide. Correct folding and native conformation of triSpike was confirmed by binding to soluble ACE2 receptor. Preliminary results obtained with two soluble forms of the S-protein ectodomain, Secd (aa1-1184) and S1 (aa1-757), show that Secd migrates with the apparent molecular weight of a trimer (>500 kDa), consistent with the model that trimerization domains are located within the membrane proximal S2 domain, while S1 migrates with the apparent molecular weight of a monomer (data not shown). These findings are in agreement with previous reports showing that coronavirus MHV and TGEV S proteins are trimers [4], [43].
Immunization studies in mice and hamsters demonstrated that triSpike is highly immunogenic and induces a long-lasting neutralizing response which remains stable for up to 6 months (neutralization titer 1/800) when the immunogen is administered with alum, an adjuvant licensed for use in humans [44]. Mucosal immunity can provide a first-line defense mechanism against viral infection in the respiratory tract or intestinal lumen where SARS-CoV replicates [45], [46]. Our results show that a neutralizing IgG and IgA response can be induced through immunization with S protein adjuvanted in alum. The induction of anti-S IgA, absent from nasal lavage samples in this study, might be critical for improved neutralization activity [47].
Several studies have previously pointed towards the receptor binding domain (RBD) within S1 as key target for neutralizing antibodies [48], [49]. A human mAb from a non-immune human antibody library was described which blocked association of S-protein with ACE2 [50]. In our experiment sera collected from Secd and S1 immunized animals also neutralized SARS-CoV (unpublished data) suggesting that the key neutralization epitopes are located in the S1 domain and that correct conformation of the monomer alone is sufficient to achieve a neutralizing response. Our data show that a purified protein vaccine can induce such blocking antibodies with high efficiency. Indeed neutralizing sera from our vaccinated mice prevented triSpike binding to soluble human ACE2 receptor indicating that the RBD is accessible to antibodies and suggesting receptor binding inhibition as key mechanism of virus neutralization in triSpike immunized animals. However, blocking of receptor binding might not be the sole mechanism of virus neutralization. Neutralization with antibodies against the putative S2 protein [51] suggest that antibodies can also block post binding steps, e.g. conformational transitions of the S2 subunit required for membrane fusion.
Antibody-dependent enhancement (ADE) of infection has been described for several viruses, e.g. Dengue virus, Human Immunodeficiency Virus, Ebola virus and FIPV [26], [52], [53], [54], [55]. ADE occurs when a virus-antibody complex interacts with FcR or complement to trigger virus uptake or alternatively when antibodies induce conformational changes in envelope glycoproteins that are required for virus-cell membrane fusion [56]. Antibody-dependent conformational changes have previously been reported for a civet cat strain of SARS-CoV which showed enhanced entry into a human renal epithelial cell-line in the presence of human monoclonal antibodies against human strain of SARS-CoV Spike [25]. We could exclude this mechanism of enhancement as sera from triSpike vaccinated mice did neutralize, rather than enhance, virus infection on VeroE6 cells. However, we observed that sera from triSpike vaccinated mice or hamsters showed a 100–1000 fold increase in virus entry into FcγRII positive, ACE2-negative human B cells. ADE was still observed when the complement was inactivated by heat treatment of sera but could be abrogated by protein G and an anti-FcγRII antibody, demonstrating that ADE is mediated by the Fc region of the antibody and the FcγRII receptor. Interestingly, the R/R131 genotype of the human FcγRIIA receptor was recently correlated with severity of disease in SARS patients [57]. Our results show that anti-ACE2 antibody did not reduce transduction by pseudovirus-antibody complexes indicating that Spike-mediated virus cell entry and membrane fusion can occur through an ACE2-independent mechanism. Together with previously described data, our results suggest that pseudovirus-antibody complexes might be endocytosed via FcγRII, followed by protease cleavage of S in the endosome [58], [59] and S2-mediated membrane fusion in a low pH endosomal compartment [9], [60].
ADE in the FIPV model has been linked to specific enhancing non-neutralizing epitopes and enhancing non-neutralizing IgG2a antibodies [61], [62]. Analysis of monoclonal antibodies against S should provide evidence whether antibody subclass or specific epitopes are involved in the enhancement mechanism. Interestingly, we observed the enhancement phenotype only in human B cell lines but not in mouse macrophages despite the presence of FcγRII. Reasons for this are unclear, but the respective role of activating and inhibiting human FcγRII which have identical extracellular but heterologous intracellular domains and signaling capacities might play a role [63]. ADE in SARS-CoV entry is not restricted to vaccine-induced anti-S antibodies but can also be observed with SARS patient sera. Eight out of ten sera capable of neutralizing infection on VeroE6 cells were found to enhance transduction of B cell lines. B cells have been shown to be only occasionally infected in patients with SARS [64]. However, enhanced viral entry into B cells could have a profound effect on pathogenesis even if ensuing viral replication in this cell type is abortive. Vaccine-induced antibodies that enhance entry of virus could increase the frequency of B cell infection in vaccinated subjects and alter disease on subsequent exposure to SARS-CoV.
Vaccine-mediated protection against SARS-CoV infection in mice has been demonstrated using different vaccine approaches [19], [20], [65]. However, mice do not replicate SARS-CoV to high titers and do not show pronounced pathology in lungs [15]. On the contrary, ferrets [28], cynomolgus macaques [29] and hamsters [30] have been shown to support high titer replication of SARS-CoV in the respiratory tract associated with pneumonitis. In order to assess the respective role of neutralization or ADE in vivo we performed SARS-CoV challenge experiments in hamsters vaccinated with three doses of 2, 10 or 50 μg of protein in alum adjuvant. All vaccinated groups showed a significant decrease of viral load at 3 days post challenge by at least four orders of magnitude without signs of enhanced lung pathology. The rare foci of hepatocyte necrosis that we observed in these hamsters did not correlate with pre-challenge neutralizing antibody titers, liver enzyme levels (pre- or post-challenge), or immunohistochemical detection of SARS-CoV antigen suggesting that hepatic cellular necrosis is unrelated to SARS-CoV immunization or infection in Syrian golden hamsters. This contrasts with a report of hepatitis 28 days following challenge with SARS-CoV in ferrets vaccinated with a poorly immunogenic recombinant MVA vaccine expressing SARS-CoV spike protein [24].
Altogether our results indicate that a recombinant trimeric S protein was able to elicit a protective immune response in vivo but if the observation of enhanced entry into B cells in vitro can be shown to have an in vivo correlate, this would warrant concern in the safety evaluation of a human vaccine against SARS-CoV.
Acknowledgements
We wish to thank Cheman Chan, Pierre Corby, Jean-Claude Manuguerra, Isabelle Staropoli and Marc Daeron for expert advice and helpful discussions. This work was supported by the European Commission (EPISARS contract SP22-CT-2004-511063), the “Programme de Recherche en Réseaux Franco-Chinois -Epidémie du SARS: de 1’émergence au contrôle and the Public Health Research Grant A195357 from the National Institute of Allergy and Infectious Disease, USA.
References
- 1.van der Hoek L., Pyrc K., Jebbink M.F., Vermeulen-Oost W., Berkhout R.J., Wolthers K.C. Identification of a new human coronavirus. Nat Med. 2004;10(4):368–373. doi: 10.1038/nm1024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Woo P.C., Lau S.K., Chu C.M., Chan K.H., Tsoi H.W., Huang Y. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. J Virol. 2005;79(2):884–895. doi: 10.1128/JVI.79.2.884-895.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Peiris J.S., Guan Y., Yuen K.Y. Severe acute respiratory syndrome. Nat Med. 2004;10(12 Suppl):S88–S97. doi: 10.1038/nm1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bosch B.J., van der Zee R., de Haan C.A., Rottier P.J. The coronavirus spike protein is a class I virus fusion protein: structural and functional characterization of the fusion core complex. J Virol. 2003;77(16):8801–8811. doi: 10.1128/JVI.77.16.8801-8811.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Song H.C., Seo M.Y., Stadler K., Yoo B.J., Choo Q.L., Coates S.R. Synthesis and characterization of a native, oligomeric form of recombinant severe acute respiratory syndrome coronavirus spike glycoprotein. J Virol. 2004;78(19):10328–10335. doi: 10.1128/JVI.78.19.10328-10335.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li W., Moore M.J., Vasilieva N., Sui J., Wong S.K., Berne M.A. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426(6965):450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang P., Chen J., Zheng A., Nie Y., Shi X., Wang W. Expression cloning of functional receptor used by SARS coronavirus. Biochem Biophys Res Commun. 2004;315(2):439–444. doi: 10.1016/j.bbrc.2004.01.076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Jeffers S.A., Tusell S.M., Gillim-Ross L., Hemmila E.M., Achenbach J.E., Babcock G.J. CD209L (L-SIGN) is a receptor for severe acute respiratory syndrome coronavirus. Proc Natl Acad Sci USA. 2004;101(44):15748–15753. doi: 10.1073/pnas.0403812101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yang Z.Y., Huang Y., Ganesh L., Leung K., Kong W.P., Schwartz O. pH-dependent entry of severe acute respiratory syndrome coronavirus is mediated by the spike glycoprotein and enhanced by dendritic cell transfer through DC-SIGN. J Virol. 2004;78(11):5642–5650. doi: 10.1128/JVI.78.11.5642-5650.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Simmons G., Reeves J.D., Rennekamp A.J., Amberg S.M., Piefer A.J., Bates P. Characterization of severe acute respiratory syndrome-associated coronavirus (SARS-CoV) spike glycoprotein-mediated viral entry. Proc Natl Acad Sci USA. 2004;101(12):4240–4245. doi: 10.1073/pnas.0306446101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hofmann H., Hattermann K., Marzi A., Gramberg T., Geier M., Krumbiegel M. S protein of severe acute respiratory syndrome-associated coronavirus mediates entry into hepatoma cell lines and is targeted by neutralizing antibodies in infected patients. J Virol. 2004;78(12):6134–6142. doi: 10.1128/JVI.78.12.6134-6142.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Nie Y., Wang G., Shi X., Zhang H., Qiu Y., He Z. Neutralizing antibodies in patients with severe acute respiratory syndrome-associated coronavirus infection. J Infect Dis. 2004;190(6):1119–1126. doi: 10.1086/423286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Greenough T.C., Babcock G.J., Roberts A., Hernandez H.J., Thomas W.D., Jr., Coccia J.A. Development and characterization of a severe acute respiratory syndrome-associated coronavirus-neutralizing human monoclonal antibody that provides effective immunoprophylaxis in mice. J Infect Dis. 2005;191(4):507–514. doi: 10.1086/427242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.ter Meulen J., Bakker A.B., van den Brink E.N., Weverling G.J., Martina B.E., Haagmans B.L. Human monoclonal antibody as prophylaxis for SARS coronavirus infection in ferrets. Lancet. 2004;363(9427):2139–2141. doi: 10.1016/S0140-6736(04)16506-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Subbarao K., McAuliffe J., Vogel L., Fahle G., Fischer S., Tatti K. Prior infection and passive transfer of neutralizing antibody prevent replication of severe acute respiratory syndrome coronavirus in the respiratory tract of mice. J Virol. 2004;78(7):3572–3577. doi: 10.1128/JVI.78.7.3572-3577.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bisht H., Roberts A., Vogel L., Bukreyev A., Collins P.L., Murphy B.R. Severe acute respiratory syndrome coronavirus spike protein expressed by attenuated vaccinia virus protectively immunizes mice. Proc Natl Acad Sci USA. 2004;101(17):6641–6646. doi: 10.1073/pnas.0401939101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Traggiai E., Becker S., Subbarao K., Kolesnikova L., Uematsu Y., Gismondo M.R. An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus. Nat Med. 2004;10(8):871–875. doi: 10.1038/nm1080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sui J., Li W., Roberts A., Matthews L.J., Murakami A., Vogel L. Evaluation of human monoclonal antibody 80R for immunoprophylaxis of severe acute respiratory syndrome by an animal study, epitope mapping, and analysis of spike variants. J Virol. 2005;79(10):5900–5906. doi: 10.1128/JVI.79.10.5900-5906.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yang Z.Y., Kong W.P., Huang Y., Roberts A., Murphy B.R., Subbarao K. A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. Nature. 2004;428(6982):561–564. doi: 10.1038/nature02463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bisht H., Roberts A., Vogel L., Subbarao K., Moss B. Neutralizing antibody and protective immunity to SARS coronavirus infection of mice induced by a soluble recombinant polypeptide containing an N-terminal segment of the spike glycoprotein. Virology. 2005;334(2):160–165. doi: 10.1016/j.virol.2005.01.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhou Z., Post P., Chubet R., Holtz K., McPherson C., Petric M. A recombinant baculovirus-expressed S glycoprotein vaccine elicits high titers of SARS-associated coronavirus (SARS-CoV) neutralizing antibodies in mice. Vaccine. 2006;24(17):3624–3631. doi: 10.1016/j.vaccine.2006.01.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.See R.H., Zakhartchouk A.N., Petric M., Lawrence D.J., Mok C.P., Hogan R.J. Comparative evaluation of two severe acute respiratory syndrome (SARS) vaccine candidates in mice challenged with SARS coronavirus. J Gen Virol. 2006;87(Pt 3):641–650. doi: 10.1099/vir.0.81579-0. [DOI] [PubMed] [Google Scholar]
- 23.Qin E., Shi H., Tang L., Wang C., Chang G., Ding Z. Immunogenicity and protective efficacy in monkeys of purified inactivated Vero-cell SARS vaccine. Vaccine. 2006;24(7):1028–1034. doi: 10.1016/j.vaccine.2005.06.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Weingartl H., Czub M., Czub S., Neufeld J., Marszal P., Gren J. Immunization with modified vaccinia virus Ankara-based recombinant vaccine against severe acute respiratory syndrome is associated with enhanced hepatitis in ferrets. J Virol. 2004;78(22):12672–12676. doi: 10.1128/JVI.78.22.12672-12676.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yang Z.Y., Werner H.C., Kong W.P., Leung K., Traggiai E., Lanzavecchia A. Evasion of antibody neutralization in emerging severe acute respiratory syndrome coronaviruses. Proc Natl Acad Sci USA. 2005;102(3):797–801. doi: 10.1073/pnas.0409065102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Vennema H., de Groot R.J., Harbour D.A., Dalderup M., Gruffydd-Jones T., Horzinek M.C. Early death after feline infectious peritonitis virus challenge due to recombinant vaccinia virus immunization. J Virol. 1990;64(3):1407–1409. doi: 10.1128/jvi.64.3.1407-1409.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Olsen C.W., Corapi W.V., Ngichabe C.K., Baines J.D., Scott F.W. Monoclonal antibodies to the spike protein of feline infectious peritonitis virus mediate antibody-dependent enhancement of infection of feline macrophages. J Virol. 1992;66(2):956–965. doi: 10.1128/jvi.66.2.956-965.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Martina B.E., Haagmans B.L., Kuiken T., Fouchier R.A., Rimmelzwaan G.F., Van Amerongen G. Virology: SARS virus infection of cats and ferrets. Nature. 2003;425(6961):915. doi: 10.1038/425915a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fouchier R.A., Kuiken T., Schutten M., van Amerongen G., van Doornum G.J., van den Hoogen B.G. Aetiology: Koch's postulates fulfilled for SARS virus. Nature. 2003;423(6937):240. doi: 10.1038/423240a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Roberts A., Vogel L., Guarner J., Hayes N., Murphy B., Zaki S. Severe acute respiratory syndrome coronavirus infection of golden Syrian hamsters. J Virol. 2005;79(1):503–511. doi: 10.1128/JVI.79.1.503-511.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lozach P.Y., Lortat-Jacob H., de Lacroix de Lavalette A., Staropoli I., Foung S., Amara A. DC-SIGN and L-SIGN are high affinity binding receptors for hepatitis C virus glycoprotein E2. J Biol Chem. 2003;278(22):20358–20366. doi: 10.1074/jbc.M301284200. [DOI] [PubMed] [Google Scholar]
- 32.Staropoli I., Chanel C., Girard M., Altmeyer R. Processing, stability, and receptor binding properties of oligomeric envelope glycoprotein from a primary HIV-1 isolate. J Biol Chem. 2000;275(45):35137–35145. doi: 10.1074/jbc.M003868200. [DOI] [PubMed] [Google Scholar]
- 33.Nal B., Chan C., Kien F., Siu L., Tse J., Chu K. Differential maturation and subcellular localization of severe acute respiratory syndrome coronavirus surface proteins S, M and E. J Gen Virol. 2005;86(Pt 5):1423–1434. doi: 10.1099/vir.0.80671-0. [DOI] [PubMed] [Google Scholar]
- 34.Matoba N., Magerus A., Geyer B.C., Zhang Y., Muralidharan M., Alfsen A. A mucosally targeted subunit vaccine candidate eliciting HIV-1 transcytosis-blocking Abs. Proc Natl Acad Sci USA. 2004;101(37):13584–13589. doi: 10.1073/pnas.0405297101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wu H.Y., Russell M.W. Induction of mucosal and systemic immune responses by intranasal immunization using recombinant cholera toxin B subunit as an adjuvant. Vaccine. 1998;16(2–3):286–292. doi: 10.1016/s0264-410x(97)00168-0. [DOI] [PubMed] [Google Scholar]
- 36.Reed L.J., Munich H A simple method for estimating fifty percent endpoints. Am J Hyg. 1938;27:493–497. [Google Scholar]
- 37.Lozach P.Y., Amara A., Bartosch B., Virelizier J.L., Arenzana-Seisdedos F., Cosset F.L. C-type Lectins L-SIGN and DC-SIGN capture and transmit infectious hepatitis C virus pseudotype particles. J Biol Chem. 2004;279(31):32035–32045. doi: 10.1074/jbc.M402296200. [DOI] [PubMed] [Google Scholar]
- 38.Shi X., Gong E., Gao D., Zhang B., Zheng J., Gao Z. Severe acute respiratory syndrome associated coronavirus is detected in intestinal tissues of fatal cases. Am J Gastroenterol. 2005;100(1):169–176. doi: 10.1111/j.1572-0241.2005.40377.x. [DOI] [PubMed] [Google Scholar]
- 39.Reddin K.M., Easterbrook T.J., Eley S.M., Russell P., Mobsby V.A., Jones D.H. Comparison of the immunological and protective responses elicited by microencapsulated formulations of the F1 antigen from Yersinia pestis. Vaccine. 1998;16(8):761–767. doi: 10.1016/s0264-410x(97)00305-8. [DOI] [PubMed] [Google Scholar]
- 40.Gomez N., Carrillo C., Salinas J., Parra F., Borca M.V., Escribano J.M. Expression of immunogenic glycoprotein S polypeptides from transmissible gastroenteritis coronavirus in transgenic plants. Virology. 1998;249(2):352–358. doi: 10.1006/viro.1998.9315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.He Y., Zhou Y., Liu S., Kou Z., Li W., Farzan M. Receptor-binding domain of SARS-CoV spike protein induces highly potent neutralizing antibodies: implication for developing subunit vaccine. Biochem Biophys Res Commun. 2004;324(2):773–781. doi: 10.1016/j.bbrc.2004.09.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Anton I.M., Gonzalez S., Bullido M.J., Corsin M., Risco C., Langeveld J.P. Cooperation between transmissible gastroenteritis coronavirus (TGEV) structural proteins in the in vitro induction of virus-specific antibodies. Virus Res. 1996;46(1–2):111–124. doi: 10.1016/S0168-1702(96)01390-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Delmas B., Laude H. Assembly of coronavirus spike protein into trimers and its role in epitope expression. J Virol. 1990;64(11):5367–5375. doi: 10.1128/jvi.64.11.5367-5375.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gupta R.K., Siber G.R. Adjuvants for human vaccines-current status, problems and future prospects. Vaccine. 1995;13(14):1263–1276. doi: 10.1016/0264-410x(95)00011-o. [DOI] [PubMed] [Google Scholar]
- 45.van Ginkel F.W., Nguyen H.H., McGhee J.R. Vaccines for mucosal immunity to combat emerging infectious diseases. Emerg Infect Dis. 2000;6(2):123–132. doi: 10.3201/eid0602.000204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lee J.S., Poo H., Han D.P., Hong S.P., Kim K., Cho M.W. Mucosal immunization with surface-displayed severe acute respiratory syndrome coronavirus spike protein on Lactobacillus casei induces neutralizing antibodies in mice. J Virol. 2006;80(8):4079–4087. doi: 10.1128/JVI.80.8.4079-4087.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Renegar K.B., Small P.A., Jr., Boykins L.G., Wright P.F. Role of IgA versus IgG in the control of influenza viral infection in the murine respiratory tract. J Immunol. 2004;173(3):1978–1986. doi: 10.4049/jimmunol.173.3.1978. [DOI] [PubMed] [Google Scholar]
- 48.He Y., Lu H., Siddiqui P., Zhou Y., Jiang S. Receptor-binding domain of severe acute respiratory syndrome coronavirus spike protein contains multiple conformation-dependent epitopes that induce highly potent neutralizing antibodies. J Immunol. 2005;174(8):4908–4915. doi: 10.4049/jimmunol.174.8.4908. [DOI] [PubMed] [Google Scholar]
- 49.He Y., Zhu Q., Liu S., Zhou Y., Yang B., Li J. Identification of a critical neutralization determinant of severe acute respiratory syndrome (SARS)-associated coronavirus: importance for designing SARS vaccines. Virology. 2005;334(1):74–82. doi: 10.1016/j.virol.2005.01.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sui J., Li W., Murakami A., Tamin A., Matthews L.J., Wong S.K. Potent neutralization of severe acute respiratory syndrome (SARS) coronavirus by a human mAb to S1 protein that blocks receptor association. Proc Natl Acad Sci USA. 2004;101(8):2536–2541. doi: 10.1073/pnas.0307140101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhang H., Wang G., Li J., Nie Y., Shi X., Lian G. Identification of an antigenic determinant on the S2 domain of the severe acute respiratory syndrome coronavirus spike glycoprotein capable of inducing neutralizing antibodies. J Virol. 2004;78(13):6938–6945. doi: 10.1128/JVI.78.13.6938-6945.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Halstead S.B. Neutralization and antibody-dependent enhancement of dengue viruses. Adv Virus Res. 2003;60:421–467. doi: 10.1016/s0065-3527(03)60011-4. [DOI] [PubMed] [Google Scholar]
- 53.Halstead S.B., Shotwell H., Casals J. Studies on the pathogenesis of dengue infection in monkeys. II. Clinical laboratory responses to heterologous infection. J Infect Dis. 1973;128(1):15–22. doi: 10.1093/infdis/128.1.15. [DOI] [PubMed] [Google Scholar]
- 54.Fust G. Enhancing antibodies in HIV infection. Parasitology. 1997;115(Suppl):S127–S140. doi: 10.1017/s0031182097001819. [DOI] [PubMed] [Google Scholar]
- 55.Takada A., Watanabe S., Okazaki K., Kida H., Kawaoka Y. Infectivity-enhancing antibodies to Ebola virus glycoprotein. J Virol. 2001;75(5):2324–2330. doi: 10.1128/JVI.75.5.2324-2330.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Takada A., Kawaoka Y. Antibody-dependent enhancement of viral infection: molecular mechanisms and in vivo implications. Rev Med Virol. 2003;13(6):387–398. doi: 10.1002/rmv.405. [DOI] [PubMed] [Google Scholar]
- 57.Yuan F.F., Tanner J., Chan P.K., Biffin S., Dyer W.B., Geczy A.F. Influence of FcgammaRIIA and MBL polymorphisms on severe acute respiratory syndrome. Tissue Antigens. 2005;66(4):291–296. doi: 10.1111/j.1399-0039.2005.00476.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Simmons G., Gosalia D.N., Rennekamp A.J., Reeves J.D., Diamond S.L., Bates P. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc Natl Acad Sci USA. 2005;102(33):11876–11881. doi: 10.1073/pnas.0505577102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Matsuyama S., Ujike M., Morikawa S., Tashiro M., Taguchi F. Protease-mediated enhancement of severe acute respiratory syndrome coronavirus infection. Proc Natl Acad Sci USA. 2005;102(35):12543–12547. doi: 10.1073/pnas.0503203102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Bosch B.J., Martina B.E., Van Der Zee R., Lepault J., Haijema B.J., Versluis C. Severe acute respiratory syndrome coronavirus (SARS-CoV) infection inhibition using spike protein heptad repeat-derived peptides. Proc Natl Acad Sci USA. 2004;101(22):8455–8460. doi: 10.1073/pnas.0400576101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Olsen C.W., Corapi W.V., Jacobson R.H., Simkins R.A., Saif L.J., Scott F.W. Identification of antigenic sites mediating antibody-dependent enhancement of feline infectious peritonitis virus infectivity. J Gen Virol. 1993;74(Pt 4):745–749. doi: 10.1099/0022-1317-74-4-745. [DOI] [PubMed] [Google Scholar]
- 62.Corapi W.V., Darteil R.J., Audonnet J.C., Chappuis G.E. Localization of antigenic sites of the S glycoprotein of feline infectious peritonitis virus involved in neutralization and antibody-dependent enhancement. J Virol. 1995;69(5):2858–2862. doi: 10.1128/jvi.69.5.2858-2862.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Daeron M. Structural bases of Fc gamma R functions. Int Rev Immunol. 1997;16(1–2):1–27. doi: 10.3109/08830189709045701. [DOI] [PubMed] [Google Scholar]
- 64.Gu J., Gong E., Zhang B., Zheng J., Gao Z., Zhong Y. Multiple organ infection and the pathogenesis of SARS. J Exp Med. 2005;202(3):415–424. doi: 10.1084/jem.20050828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Stadler K., Roberts A., Becker S., Vogel L., Eickmann M., Kolesnikova L. SARS vaccine protective in mice. Emerg Infect Dis. 2005;11(8):1312–1314. doi: 10.3201/eid1108.041003. [DOI] [PMC free article] [PubMed] [Google Scholar]