

Abstract
Canine coronavirus (CCoV) is an enveloped RNA virus, responsible for gastrointestinal infection in dogs. To date, two different CCoV genotypes have been recognized, CCoV type I and CCoV type II. Recently, CCoV type II strains of potential recombinant origin with transmissible gastroenteritis virus (TGEV) were detected and characterized as a new subtype (CCoV-IIb) of canine coronavirus, in order to be differentiated from the “classical” CCoV type II strains (CCoV-IIa). In the present study, two CCoV-IIb strains were detected in the faeces and internal organs of two puppies, which died after presenting gastrointestinal symptoms. Mixed infection of both subtypes (CCoV-IIa/IIb) was detected in the faeces, while only CCoV-IIb was detected in the organs. Puppies were also infected by canine parvovirus type 2 (CPV-2). Both CCoV-IIb strains were isolated on cell cultures and subjected to sequence analysis and phylogeny. By means of RT-PCR and real time RT-PCR assays, tissue distribution and quantitation of viral loads took place. These cases represent the first description of tissue distribution and quantitation of CCoV-IIb strains, detected in the organs. The detection of CCoV-IIa strains, which is restricted to the faeces, suggests that CCoV-IIb strains may have an advantage in disseminating throughout a dog with CPV-2 coinfection, in contrast to common enteric CCoV-IIa strains.
Keywords: CCoV, Recombination, TGEV-like, Dog, Distribution, Organs
1. Introduction
Canine coronavirus (CCoV; order Nidovirales, family Coronaviridae) is a large, enveloped, single stranded, RNA virus responsible for enteritis in dogs (Decaro and Buonavoglia, 2008). Recently, due to changes in virus classification, the virus was classified as a member of the genus Alphacoronavirus, species Alphacoronavirus-1, together with transmissible gastroenteritis virus of swine (TGEV) and feline coronavirus (FCoV) (Carstens, 2010). The genome, 27 kb in length, contains two large overlapping open reading frames (ORFs), ORF1a and ORF1b which encompass the 5 two thirds of the genomic RNA and encode polyproteins leading to the replicase complex. The ORFs, encoding for the structural spike (S), envelope (E), membrane (M) and nucleocapsid (N) proteins and the non-structural proteins (3a, 3b, 3c, 7a and 7b), are located downstream of the replicase gene (Decaro and Buonavoglia, 2008).
Coronaviruses are characterized by constant genetic evolution and diversity. To date, two different CCoV types have been recognized, CCoV type I (CCoV-I) and CCoV type II (CCoV-II), that share significant genetic similarity with FCoV type I (FCoV-I) and FCoV type II (FCoV-II), respectively (Decaro and Buonavoglia, 2008). Moreover, in 2009, TGEV-like CCoVs of potential recombinant origin were identified and characterized as a new CCoV subtype (CCoV-IIb) (Decaro et al., 2009, Decaro et al., 2010, Erles and Brownlie, 2009).
CCoV is the causative agent of gastroenteritis in dogs, characterized by high morbidity and low mortality. Clinical signs include anorexia, lethargy, vomiting, mild to severe diarrhoea (usually lasting 1–2 weeks) and occasionally death, mainly in puppies. The disease is more severe in young animals (Carmichael and Binn, 1981). Systemic infections are not usual; however, during the past few years, there have been reports of fatal disease, with CCoV strains detected in the enteric tract, as well as in the organs (Buonavoglia et al., 2006, Decaro et al., 2009).
In 2010, CCoV identification, molecular characterization and sequence analysis took place for the first time in Greece, regarding common enteric CCoV-II strains detected in a severe outbreak of diarrhoea in a kennel (Ntafis et al., 2010). In the current study we report the quantitation and molecular characterization of two TGEV-like CCoV strains, detected in the organs of two puppies displaying fatal enteritis.
2. Materials and methods
2.1. Clinical case
During summer of 2009, two dead dogs were submitted for laboratory investigation. The dogs were coming from two different pet shops of Thessaloniki, a city of northern Greece. Both dogs, a 6-week-old Yorkshire Terrier (66/09) and a 16-week-old Pomeranian (68/09), presented fever, lethargy, inappetence, severe haemorrhagic diarrhoea and vomiting leading to death, 2 days after the onset of the symptoms. The first puppy was vaccinated with a single dose of a polyvalent vaccine against all major infectious diseases (canine distemper, infectious hepatitis, parvoviral enteritis, parainfluenza and leptospirosis) 2 weeks before the symptoms, while the second one, had never been vaccinated.
Necropsy examination of both dogs revealed linear haemorrhages of the intestinal wall, haemorrhagic enteritis and ulcerated duodenum. Sero-sanguineous fluid was observed in the abdominal cavity of the Pomeranian. Lungs of both puppies were congested with multiple areas of emphysema. No lesions were observed at the heart. Liver of both puppies appeared enlarged, friable and yellow-brown in color with multifocal discolorated spots. Congested vessels in the dura mater of the brain were also observed.
2.2. Screening for viral pathogens
Samples from the faeces and the parenchymatous organs were subjected to virological investigations, using methods previously described, regarding common canine viral pathogens e.g., canine parvovirus type 2 (CPV-2) (PCR and real time PCR) (Decaro et al., 2005a, Decaro et al., 2006a, Decaro et al., 2006b), canine distemper virus (CDV) (RT-PCR) (Frisk et al., 1999), canine adenovirus type 1 and type 2 (CAV-1 and CAV-2) (PCR) (Hu et al., 2001) and CCoV (RT-PCR) (Pratelli et al., 1999).
2.3. Virus isolation
For virus isolation, A-72 cell line (canine fibrosarcoma) was used. The cells were grown in Dulbecco-Minimum Essential Medium (D-MEM) supplemented with 10% foetal bovine serum (FBS). Faecal and tissue samples were homogenized (10%, w/v) in D-MEM and centrifuged at 8000 × g for 10 min. Supernatants were treated with antibiotics (1000 IU/ml penicillin and 100 μg/ml streptomycin) for 30 min, inoculated on partially confluent A72 cell cultures and then, they were incubated at 37 °C in a 5% CO2 incubator. After an adsorption period of 30 min, D-MEM was added. Cells were daily observed for cytopathic effect (cpe) of CCoV for 5 days. An immunofluorescence (IF) assay was used for the detection of CCoV at the infected cells. For the IF assay a 1:100 dilution of cat polyclonal serum specific for Alphacoronavirus-1 and a 1:100 dilution of goat anti-cat IgG conjugated with fluorescein isothiocyanate (Sigma–Aldrich, USA). Each sample was considered negative after 3 passages.
2.4. CCoV characterization and quantitation
RNA was extracted from faecal and organ samples of both dogs using the QIAamp Viral RNA Mini Kit and the RNeasy Mini Kit (Qiagen GmbH, Hilden, Germany), respectively. For CCoV type I and II detection and quantitation in faecal and organ samples, two real time RT-PCR assays with the same sensitivity were used (Decaro et al., 2005b). Reverse transcription was performed using GeneAmp® RNA PCR (Applied Biosystems, Italy) according to the manufacturer's instructions.
For the discrimination of classical (subtype IIa) and TGEV-like (subtype IIb) CCoVs, two RT-PCR assays with comparable levels of sensitivity were performed, as previously described (Decaro et al., 2010). RT-PCRs with primers 20179/INS-R (CCoV-IIa) or 20179/174-268 (CCoV-IIb) were conducted using SuperScript One-Step RT-PCR for Long Templates (Invitrogen S.R.L.). In order to verify the absence of TGEV strains in the samples that were positive by CCoV-IIb specific assay, an RT-PCR, able to discriminate CCoV and TGEV according to the amplicon size was used (Wesley, 1999).
2.5. Sequencing and sequence analysis
The 3′ end of the genome of the CCoV-IIb strains was amplified as previously described, using viral RNA extracted from the lungs, SuperScript One-Step RT-PCR for Long Templates (Invitrogen S.R.L.) and six pairs of primers, specific for overlapping fragments, encompassing ORFs 2, 3a, 3b, 3c, 4, 5, 6, 7a and 7b (Decaro et al., 2007). The nucleotide sequences were determined in both directions by a commercial facility (Beckman Coulter Genomics, United Kingdom). Sequence assembling and analysis were carried out using the BioEdit software package (Hall, 1999) and the National Center for Biotechnology Information (NCBI; http://www.ncbi.nlm.nih.gov) and European Molecular Biology Laboratory (EMBL; http://www.ebi.ac.uk) analysis tools. Phylogenetic analysis was conducted using MEGA4 program (Tamura et al., 2007). Phylogenetic trees, based on the amino acid sequences of S, E, M and N proteins, were elaborated using neighbor-joining method, supplying a statistical support with bootstrapping over 1000 replicates. SimPlot was used for nucleotide sequence comparison of the two strains to Alphacoronavirus-1 reference strains (Lole et al., 1999). The sequences of strains 66/09 and 68/09 were registered in GenBank under the accession numbers HQ450376 and HQ450377, respectively.
3. Results
3.1. CCoV detection, characterization and isolation
By means of nested PCR assay for CCoV, viral RNA was detected in faeces, lungs, spleen, kidneys, pancreas, heart, and liver of both puppies. In addition, the brain of the Pomeranian (68/09) was tested positive, while the brain of the Yorkshire Terrier (66/09) was tested negative. By genotype specific real time RT-PCR assays, only CCoV-II was detected in all positive samples. CCoV-II RNA copies/μl of template in the samples are shown in Table 1 .
Table 1.
CCoV-II RNA copies/μl of template in the samples of the two puppies, tested by genotype-specific real time RT-PCR.
| Sample | 66/09 (Yorkshire Terrier) | 68/09 (Pomeranian) |
|---|---|---|
| Faeces | 3.59 × 103 | 7.22 × 105 |
| Liver | 4.64 × 104 | 3.21 × 105 |
| Spleen | 5.20 × 105 | 1.55 × 107 |
| Pancreas | 2.75 × 102 | 2.03 × 104 |
| Kidney | 1.23 × 105 | 3.37 × 106 |
| Lung | 5.99 × 106 | 4.10 × 106 |
| Heart | 1.14 × 105 | 7.08 × 106 |
| Brain | n.d. | 2.47 × 103 |
n.d., not detected.
In the faecal samples of the two puppies, both CCoV-II subtypes were detected, while in the organs which tested positive, only CCoV which was characterized as TGEV-like (CCoV-IIb) was detected. No TGEV strains were detected in the samples.
The CCoV-IIb strains (66/09 and 68/09) were isolated from the lung homogenates of both puppies. A-72 cells developed a cytopathic effect that consisted of cell rounding and lysis of the monolayer. In addition, cells were tested positive by the immunofluorescence assay. Viral titres on cell cultures were 104.25 (66/09) and 104 TCID50/50 μl (68/09) at the 3rd passage.
3.2. Detection of other viral pathogens
Both puppies were tested positive for CPV-2a field strains and negative for CDV, CAV-1 and CAV-2.
3.3. Sequencing results and phylogenetic analysis
A total of 8822 and 8828 nucleotides were determined for strains 66/09 and 68/09, respectively, encompassing ORFs 2 (S protein), 3a, 3b, 3c, 4 (E protein), 5 (M protein), 6 (N protein), 7a and 7b. Alignment of the sequences with TGEV, CCoV and FCoV reference strains available in GenBank showed the highest identity to CCoV-IIb reference strain 119/08 (EU924791) (98.2% and 98.9% for 66/09 and 68/09, respectively). The two Greek strains shared an identity of 98%.
The spike protein gene of both strains was 4374 nucleotides long, encoding a protein of 1457 amino acids. When compared to four TGEV-like reference strains (430/07, 119/08, 174/06 and 341/05), no insertions or deletions were observed. The two strains shared 97.6% aa identity to each other, while they showed the highest aa identity to CCoV-IIb reference strain 119/08 (98.3%). By Simplot analysis, the two strains displayed higher nucleotide conservation with the TGEV strain Purdue than with the pantropic CCoV-IIa strain CB/05, at the 5′-end of the S gene (Fig. 1 ). Phylogenetic analysis revealed that the two Greek strains were more closely related to the four CCoV-IIb reference strains detected in dogs’ organs (Fig. 2a).
Fig. 1.
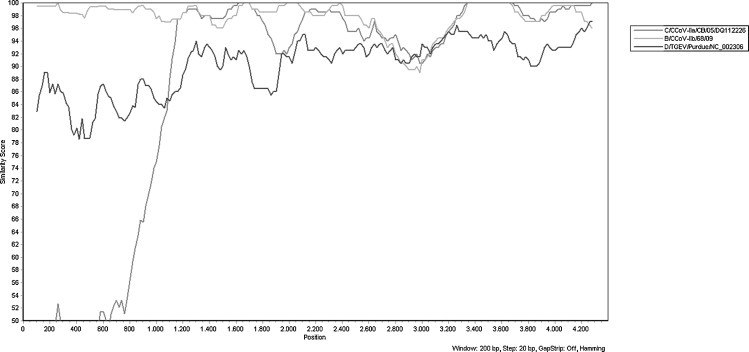
S gene sequences analysis with Simplot. The S gene of CCoV-IIb strain 68/09, TGEV strain Purdue and CCoV-IIa pantropic strain CB/05 were plotted against the S gene of CCoV-IIb strain 66/09.
Fig. 2.
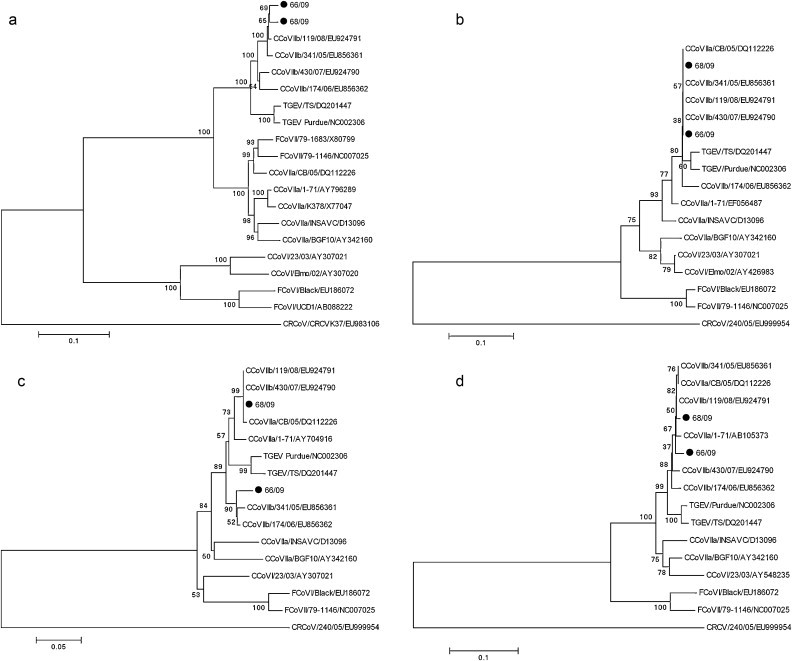
Neighbor-joining trees of the Greek strains, based on the S (a), E (b), M (c) and N (d) protein. The trees are rooted on the group 2 canine respiratory coronavirus (CRCoV). The numbers represent the percentage of replicate trees based on 1000 bootstrap replicates.
The envelope protein was found to be 82 amino acids in length, like in most canine coronavirus strains and in three TGEV-like reference strains, 119/08, 174/06 and 341/05, with the exception of 430/07, which is 7 amino acids shorter. The Greek strains had high amino acid identity to each other (98.7%). E protein of strains 66/09 and 68/09 had the highest amino acid identity (100% and 98.7%, respectively) to the CCoV-IIb strains 341/05, 119/08, and to CCoV-IIa CB/05. In the E protein, phylogenetic analysis revealed that the two strains were closely related to CCoV type II strains (Fig. 2b).
The membrane protein (M protein) of strains 66/09 and 68/09 was found to be 260 and 262 amino acids long, respectively. Two amino acids were missing from the N-terminal end of the M protein of strain 66/09 in positions 24 and 36, as it has been also observed in reference CCoV-IIb strains 174/06 and 341/05. The two strains shared high amino acid similarity (94.6%). M protein of strains 66/09 and 68/09 had the highest amino acid identity to the CCoV-IIb reference strains detected in the organs (97.3% and 100%, respectively). Phylogenetic analysis of the M protein showed that the two strains were closely related to CCoV-IIa and CCoV-IIb strains (Fig. 2c).
The N gene (nucleoprotein) was found to be 1149 nucleotides in length, coding for a polypeptide of 382 amino acids. The two proteins were 98.1% similar. The amino acid sequences had the highest identity with CCoV-IIb 119/08 (98.6% and 99.4%, for 66/09 and 68/09, respectively). Phylogenetic analysis revealed that the two Greek strains were more closely related to CCoV-II reference strains (Fig. 2d).
4. Discussion
Homologous RNA recombination consists one of the major “powers” of genetic evolution and diversity, regarding coronaviruses (Woo et al., 2009). Under field conditions, mixed infections are required to give rise to recombination events. So far, experimental infections of piglets (Woods and Wesley, 1992) and dogs (Larson et al., 1979) with CCoV and TGEV strains, respectively, and the fact that feline aminopeptidase N serves as a functional receptor for both CCoV and TGEV (Tresnan et al., 1996), strongly suggest that the two viruses can be found growing at the same “environment” in nature, although the exact host of recombination still remains unknown.
A canine coronavirus strain (UCD-1) of potential recombinant origin with TGEV, was identified for the first time in the late 90s (Wesley, 1999). Recently, TGEV-like strains were reported, circulating in dogs in different countries of Europe (Decaro et al., 2010). The strains were detected in faecal samples of dogs with gastroenteritis, they were classified as the new subtype CCoV-IIb and it was suggested that they were a result of recombination events, occurring at different times of these, regarding the old strain UCD-1 (Decaro et al., 2010). In the present study, sequence and phylogenetic analysis takes place for the first time in CCoV-IIb strains detected in Greece.
Moreover, our findings suggest that TGEV-like CCoV strains spreading to the internal organs are circulating in dogs, since so far, there has been only one report in Italy (Decaro et al., 2009). By means of real time RT-PCR, tissue distribution and quantitation of both strains were assessed for the first time, revealing the spreading of the virus to the internal organs. The CPV-2 coinfection may contribute to the spreading of TGEV-like CCoV strains, since so far, they have been only detected in organs of dogs infected also with CPV-2 (Decaro et al., 2009). However, the detection of CCoV-IIa strains strictly in the faeces, in both cases, suggests that CCoV-IIb may have an advantage in disseminating through the dog. In the first report of TGEV-like strains detected in the organs, CCoV-I was also detected strictly in the intestinal content in two cases (Decaro et al., 2009). These cases strongly suggest a difference in pathobiology of CCoV-IIb with respect to CCoV-I/IIa.
By sequence and phylogenetic analysis, it was shown that both strains segregate constantly with the CCoV-IIb reference strains detected in the organs of dogs. Accordingly, the strains were highly similar to TGEV in the 5′ end of the S gene, whereas they clustered with the pantropic CCoV variant CB/05 (subtype CCoV-IIa) in the E, M and N proteins. In a previous study, CCoV-IIb strains detected in the organs were found to share higher amino acid identity with CB/05 than with CCoV common enteric strains, at the level of the same proteins (Decaro et al., 2009). Whether the ability of CCoV-IIb strains to spread to the organs is related to the recently recognized recombinant protein S or to the CB/05-like proteins (E, M and N) needs further research. However, the S-protein “scenario” seems to be more possible, since in coronaviruses S protein mediates receptor attachment, and tissue tropism shift has been associated with mutations in the S gene (Masters, 2006).
In the last decade, new genotypes and subtypes of canine coronavirus have been recognized. Furthermore, a pantropic variant with the ability to cause fatal systemic infection was detected (Buonavoglia et al., 2006). Previous studies revealed that there are antigenic differences between CCoV-I and II (Pratelli et al., 2004). In addition, antigenic differences were observed between the two subtypes, CCoV-IIa and CCoV-IIb (TGEV-like CCoVs) (Decaro et al., 2009). Whether the currently circulating vaccines can protect against the TGEV-like recombinant isolates or not has to be verified via vaccinations and experimental infections.
5. Conclusion
In conclusion, this was the first report of CCoV-IIb tissue distribution. Up to now, there has been only one report of TGEV-like strains detected in internal organs of puppies in Italy. Based on sequence and phylogenetic analysis of the structural proteins, the two Greek isolates were found to be related to the Italian prototype CCoV-IIb strains. In addition, in all cases a mixed infection with CPV-2 was reported. However, the detection of CCoV-IIa strains, strictly at the faeces, suggests that CCoV-IIb strains may have an advantage in disseminating throughout a dog with CPV-2 coinfection, in contrast to common enteric CCoV-IIa strains.
Acknowledgement
Ntafis Vasileios is grateful to Alexander S. Onassis Public Benefit Foundation for doctoral funding.
References
- Buonavoglia C., Decaro N., Martella V., Elia G., Campolo M., Desario C., Castagnaro M., Tempesta M. Canine coronavirus highly pathogenic for dogs. Emerg. Infect. Dis. 2006;12:492–494. doi: 10.3201/eid1203.050839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmichael L.E., Binn L.N. New enteric viruses in the dog. Adv. Vet. Sci. Comp. Med. 1981;25:1–37. [PubMed] [Google Scholar]
- Carstens E.B. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses. Arch. Virol. 2010;155:133–146. doi: 10.1007/s00705-009-0547-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Decaro N., Elia G., Martella V., Desario C., Campolo M., Di Trani L., Tarsitano E., Tempesta M., Buonavoglia C. A real-time PCR assay for rapid detection and quantitation of canine parvovirus type 2 in the feces of dogs. Vet. Microbiol. 2005;105:19–28. doi: 10.1016/j.vetmic.2004.09.018. [DOI] [PubMed] [Google Scholar]
- Decaro N., Martella V., Ricci D., Elia G., Desario C., Campolo M., Cavaliere N., Trani L.D., Tempesta M., Buonavoglia C. Genotype-specific fluorogenic RT-PCR assays for the detection and the quantitation of canine coronavirus type I and type II RNA in faecal samples of dogs. J. Virol. Methods. 2005;130:72–78. doi: 10.1016/j.jviromet.2005.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Decaro N., Elia G., Desario C., Roperto S., Martella V., Campolo M., Lorusso A., Cavalli A., Buonavoglia C. A minor groove binder probe real-time PCR assay for discrimination between type 2-based vaccines and field strains of canine parvovirus. J. Virol. Methods. 2006;136:65–70. doi: 10.1016/j.jviromet.2006.03.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Decaro N., Elia G., Martella V., Campolo M., Desario C., Camero M., Cirone F., Lorusso E., Lucente M.S., Narcisi D., Scalia P., Buonavoglia C. Characterization of the canine parvovirus type 2 variants using minor groove binder probe technology. J. Virol. Methods. 2006;133:92–99. doi: 10.1016/j.jviromet.2005.10.026. [DOI] [PubMed] [Google Scholar]
- Decaro N., Martella V., Elia G., Campolo M., Desario C., Cirone F., Tempesta M., Buonavoglia C. Molecular characterization of the virulent canine coronavirus CB/05 strain. Virus Res. 2007;125:54–60. doi: 10.1016/j.virusres.2006.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Decaro N., Buonavoglia C. An update on canine coronaviruses: viral evolution and pathobiology. Vet. Microbiol. 2008;132:221–234. doi: 10.1016/j.vetmic.2008.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Decaro N., Mari V., Campolo M., Lorusso A., Camero M., Elia G., Martella V., Cordioli P., Enjuanes L., Buonavoglia C. Recombinant canine coronavirus related to transmissible gastroenteritis virus of swine are circulating in dogs. J. Virol. 2009;83(3):1532–1537. doi: 10.1128/JVI.01937-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Decaro N., Mari V., Elia G., Addie D.D., Camero M., Lucente M.S., Martella V., Buonavoglia C. Recombinant canine coronaviruses in dogs. Eur. Emerg. Infect. Dis. 2010;16:41–47. doi: 10.3201/eid1601.090726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erles K., Brownlie J. Sequence analysis of divergent canine coronavirus strains present in UK dog population. Virus Res. 2009;141:21–25. doi: 10.1016/j.virusres.2008.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frisk A.L., Konig M., Moritz A., Baumgartner W. Detection of canine distemper virus nucleoprotein RNA by reverse transcription-PCR using serum, whole blood, and cerebrospinal fluid from dogs with distemper. J. Clin. Microbiol. 1999;37:3634–3643. doi: 10.1128/jcm.37.11.3634-3643.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall T.A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. 1999;41:95–98. [Google Scholar]
- Hu R.L., Huang G., Qiu W., Zhong Z.H., Xia X.Z., Yin Z. Detection and differentiation of CAV-1 and CAV-2 by polymerase chain reaction. Vet. Res. Commun. 2001;25:77–84. doi: 10.1023/a:1006417203856. [DOI] [PubMed] [Google Scholar]
- Larson D.J., Morehouse L.G., Solorzano R.F., Kinden D.A. Transmissible gastroenteritis in neonatal dogs: experimental intestinal infection with transmissible gastroenteritis virus. Am. J. Vet. Res. 1979;40:477–486. [PubMed] [Google Scholar]
- Lole K.S., Bollinger R.C., Paranjape R.S., Gadkari D., Kulkarni S.S., Novak N.G., Ingersoll R., Sheppard H.W., Ray S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seronconverters in India, with evidence of intersubtype recombination. J. Virol. 1999;72:152–160. doi: 10.1128/jvi.73.1.152-160.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masters P.S. The molecular biology of coronaviruses. Adv. Virus Res. 2006;66:193–292. doi: 10.1016/S0065-3527(06)66005-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ntafis V., Mari V., Danika S., Fragkiadaki E., Buonavoglia C. An outbreak of canine coronavirus in a Greek kennel. J. Vet. Diagn. Invest. 2010;22:320–323. doi: 10.1177/104063871002200231. [DOI] [PubMed] [Google Scholar]
- Pratelli A., Tempesta M., Greco G., Martella V., Buonavoglia C. Development of a nested PCR assay for the detection of canine coronavirus. J. Virol. Methods. 1999;80:11–15. doi: 10.1016/S0166-0934(99)00017-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pratelli A., Elia G., Decaro N., Tola S., Tinelli A., Martella V., Rocca S., Tempesta M., Buonavoglia C. Cloning and expression of two fragments of the S gene of canine coronavirus type I. J. Virol. Methods. 2004;117:61–65. doi: 10.1016/j.jviromet.2003.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamura K., Dudley J., Nei M., Kumar S. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007;24:1596–1599. doi: 10.1093/molbev/msm092. [DOI] [PubMed] [Google Scholar]
- Tresnan D.B., Levis R., Holmes K.V. Feline aminopeptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I. J. Virol. 1996;70(12):8669–8674. doi: 10.1128/jvi.70.12.8669-8674.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wesley R.D. The S gene of canine coronavirus, strain UCD-1, is more closely related to the S gene of transmissible gastroenteritis virus than to that of feline infectious peritonitis virus. Virus Res. 1999;61:145–152. doi: 10.1016/S0168-1702(99)00032-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C.Y., Lau S.K.P., Huang Y., Yuen K.Y. Coronavirus diversity, phylogeny and interspecies jumping. Exp. Biol. Med. 2009;234:1117–1127. doi: 10.3181/0903-MR-94. [DOI] [PubMed] [Google Scholar]
- Woods R.D., Wesley R.D. Seroconversion of pigs in contact with dogs exposed to canine coronavirus. Can. J. Vet. Res. 1992;56:78–80. [PMC free article] [PubMed] [Google Scholar]