

Abstract
The outcomes of feline coronavirus (FCoV) infection vary greatly from asymptomatic or mild enteric infection to fatal feline infectious peritonitis (FIP). On the basis of in vitro neutralization tests, FCoVs can be divided into two serotypes. To explore the correlation between different types of FCoV and FIP, clinical specimens collected from 363 naturally infected cats during 2003–2007 were analyzed. Amplification of a portion of the S gene from the FCoV was performed and a total of 222 cases were differentiated. Among them, 197 (88.7%) cats were type I-positive, 13 (5.9%) were type II-positive, and 12 (5.4%) were positive for both types. Irrespective of the predominance of type I FCoV infection in Taiwan, type II FCoV demonstrated a significantly higher correlation with FIP (p < 0.01). Analysis of partial S gene sequences of the local type I and II FCoVs strains revealed that type I viruses were more genetically divergent (6.2–11.7%) than type II viruses (0.6–3.2%) within the 5-year study period. The higher genetic diversity of type I FCoVs might be due to the larger infected cat population and to the long period of viral persistence in asymptomatic cats in comparison to type II viruses.
Keywords: Feline infectious peritonitis, Feline coronavirus, Virus type, Genetic diversity
1. Introduction
Feline infectious peritonitis (FIP) is one of the major infectious cause of mortality in kitten (Cave et al., 2002). The causative agent, feline coronavirus (FCoV), is an enveloped, positive-sense, single-stranded RNA virus belonging to family Coronaviridae within the order Nidovirales (Lai et al., 2007). FCoVs are also associated with mild or subclinical enteric infections (Pedersen et al., 1981). Despite the high prevalence of FCoVs in feline populations around the world (Bell et al., 2006, Horzinek and Osterhaus, 1979, Kiss et al., 2000, Pedersen, 1976, Sparkes et al., 1992), only 5–12% of seropositive cats develop FIP (Addie and Jarrett, 1992, Addie et al., 1995).
FCoVs are further classified into two serotypes that differ in their growth characteristics in cell culture and antigenicity relative to canine coronaviruses (CCoVs) (Pedersen et al., 1984). Type II FCoVs are the result of recombination between a type I FCoV and CCoV (Herrewegh et al., 1998). Both types of FCoV can cause FIP (Pedersen et al., 1984). Type I and II FCoVs have different cell tropisms that can be explained by differences in the binding specificity of their respective spike proteins (Hohdatsu et al., 1998). Based upon recognized differences in the spike genes of these two virus types, reverse transcription-polymerase chain reaction (RT-PCR) analyses has been employed as an alternative method to distinguish between type I and II viruses (Addie et al., 2003, Benetka et al., 2004). The prevalence of type I and II FCoV has been surveyed in many countries, including the USA (Vennema, 1999), UK (Addie et al., 2003), Japan (Hohdatsu et al., 1992, Shiba et al., 2007), Austria (Benetka et al., 2004), and Switzerland (Kummrow et al., 2005). Regardless of which assays were used, the majority of infections were attributed to type I FCoV. Although both type I and II FCoV can cause FIP, their likelihood of association with FIP was controversial in different studies (Kummrow et al., 2005, Hohdatsu et al., 1992).
Coronaviruses (CoVs) have been identified in a variety of host species, including birds, humans, livestock, and other mammals (Lai et al., 2007). Recently, the evolution of CoVs has been extensively studied (Dong et al., 2007, Vijaykrishna et al., 2007, Woo et al., 2007) and the prototype FCoV strain 79-1146 was placed in group 1a together with transmissible gastroenteritis virus (TGEV) and two wild animal CoVs (Vijaykrishna et al., 2007). As strain 79-1146 is a type II virus isolated long time ago (McKeirnan et al., 1981), the genetic information carrying in current FCoVs might be quite different. To explore the relationship between FIP and different FCoV serotypes and to better understand the prevalence and genetic diversity of the recent type I and II viruses, naturally infected cases collected over the past 5 years from our teaching hospital were investigated. Despite the higher prevalence and genetic diversity of type I FCoV, infection with type II FCoV appears to be highly significantly correlated with FIP.
2. Materials and methods
2.1. Animal and specimen collection
Clinical specimens including body effusions, whole blood, rectal swabs, and conjunctival swabs were collected from cats presenting at the Animal Hospital of National Taiwan University over a 5-year period (2003–2007). All specimens were screened for FCoV by RT-PCR, as described by Herrewegh et al. (1995). A total of 363 cases that showed positive RT-PCR from either one of the specimens were enrolled in this study. These include 127 healthy cats (83 admitted for routine health check and 44 FIP-cohabitated cats) together with 236 clinically FIP-suspected cats that showed either a clinical history of anorexia, weight loss, lethargy, icterus, mild antibiotic-unresponsive fever, abdominal distension and/or thoracic effusion with a low albumin to globulin ratio. The specimens that tested positively were subsequently subjected to FCoV typing.
2.2. Sample preparation and reverse transcription
Rectal swab samples were suspended in 1 ml of 0.1% DEPC-treated water. Total RNA was extracted from 300 μl of whole blood, body effusion, or rectal swab suspension using Trizol (Chomczynski and Sacchi, 1987). Eleven microlitre of isolated RNA were added to the premix, consisting of 4 μl of 5× RT buffer, 1 μl of 2.5 mM dNTPs (GeneTeks Bioscience, Inc., Taipei), 1 μl of 50 μM oligo dT, 2 μl of 0.1 M dithiothreitol, and 1 μl containing 200 U Moloney murine leukemia virus reverse transcriptase (Invitrogen, CA, USA) in a 0.6 ml reaction tube. This reaction mixture was incubated at 37 °C for 60 min, then at 72 °C for 15 min, and finally at 94 °C for 5 min.
2.3. Multiplex-nested PCR amplification of the S gene
For typing of FCoV, multiplex-nested PCR was performed according to the procedures outlined in Addie et al. (2003) with slight modifications. Following reverse transcription, 2 μl of the product was added to 28 μl of the PCR mixture. The PCR mixture consisted of 3 μl of 10× Taq buffer, 0.5 μl of each primer (10 μM) (S1: 5′-CCACACATACCAAGGCCA-3′, S2: 5′-CTTAATGC(A/T)T(A/T)TGTGTCTC-3′), 1 μl dNTPs (2.5 mM), 0.5 μl Taq DNA polymerase (GeneTeks BioScience, Inc., Taipei), and 22.5 μl of 0.1% DEPC-treated water. The temperature cycling (Mastercycler Personal, Eppendorf), consisted of 3 min of preheating at 94 °C; 35 cycles of denaturation at 94 °C for 30 s, annealing at 50 °C for 30 s, and extension at 72 °C for 40 s, with a final extension at 72 °C for 5 min. Multiplex-nested PCR was performed on 2 μl of the first PCR product using nested primers (Addie et al., 2003). The temperature cycling protocol consisted of 3 min of preheating at 94 °C, 35 cycles of denaturation at 94 °C for 30 s, annealing at 47 °C for 30 s, and extension at 72 °C for 30 s, with a final extension at 72 °C for 5 min. The expected size of the product from the first PCR was 702 bp. The second PCR yielded the expected products for type I and II FCoV: 360 and 218 bp, respectively.
2.4. Sequencing and analysis of PCR-amplified products
A 10 μl aliquot of each PCR product was analyzed by electrophoresis through a 1.5% agarose gel (Viogene, Taipei). Amplification products were visualized using UV illumination after ethidium bromide staining. The targeted DNA fragments were purified (Geneaid Biotech, Ltd., Taipei), and the nucleic acid sequences were determined from both orientations, sense and antisense, using an automated DNA sequencer (ABI 3730XL, USA). The cDNA sequences of a region of the S gene were then compared with those of reference FCoVs, CCoV, and wild animal CoVs. Multiple alignments of nucleic acid sequences were performed by the Clustal W method using the MegAlign program (DNASTAR Inc., WI, USA).
2.5. Statistical analysis
All cases were divided into three groups, namely FIP, ill, and healthy, according to the laboratory findings. A definitive diagnosis of FIP was made by a postmortem histopathological examination with typical pyogranulomatous lesions (Hartmann et al., 2003). The grouping criteria for ill were made for cases lacking the final histopathological confirmation irrespective of how strong their clinical and/or laboratory evidences of FIP may be. Healthy cats refer to those that were first presented to our hospital either for a routine health check or cohabitated with a FIP-diagnosed cat, and have never been readmitted for FIP during the past years. Correlation of virus genotypes with FIP disease was made by analyzing cases from FIP and healthy groups but not ill ones. The statistical evaluation was carried out with the chi-square test with Yate's correction. The difference in values was considered statistically significant and highly significant if the associated p value was <0.05 and <0.01, respectively.
3. Results
3.1. Prevalence of FCoV types
Of the 363 cases, 197 (54.3%) tested positively for type I, 13 (3.6%) for type II, and 12 (3.3%) for both types; however, there were 141 cases (38.8%) for which differentiation was not possible by our RT-nPCR (Table 1 ). Among the 222 differentiable cases, type I appeared to be the main type (88.7%, 197/222), type II accounted for 5.9% (13/222), and cats infected with both types of virus made up the rest 5.4% (12/222). Only the differentiable cases were subjected to further analysis.
Table 1.
Typing of feline coronavirus by RT-PCR and its prevalence in Taiwan.
| Clinical statusa | Number of cases tested |
||||
|---|---|---|---|---|---|
| Typable |
Untypable | Total | |||
| Type I | Type II | Type I and II | |||
| FIP | 28/38 (73.7%) | 4/38 (10.5%) | 6/38 (15.8%) | 10 | 48/363 (13.2%) |
| Ill | 98/110 (89.0%) | 6/110 (5.5%) | 6/110 (5.5%) | 78 | 188/363 (51.8%) |
| Healthy | 71/74 (95.9%) | 3/74 (4.1%) | 0/74 (0%) | 53 | 127/363 (35.0%) |
| Total | 197/363 (54.3%) | 13/363 (3.6%) | 12/363 (3.3%) | 141/363 (38.8%) | 363 |
FIP: FIP confirmed by FCoV positive results from body effusion and necropsy; Ill: cat was present either with the anorexia, weight loss, lethargy, icterus, or diarrhea but lack of final histopathological confirmation.
3.2. Correlation of FCoV type with FIP disease
According to the grouping criteria given above, 110 FIP-suspected cats of which necropsy were unavailable after death were excluded from our analysis and were put into ill group (Table 1). Among the 74 FCoV strains from healthy cats, 71 (95.9%) were type I and 3 (4.1%) were type II. For the 38 cats with definite diagnosis of FIP, 28 (73.7%) were type I, 4 (10.5%) were type II, and 6 (15.8%) had both types of FCoVs (Table 1). Surprisingly, among the 13 cats infected with type II viruses, 10 (76.9%) were associated with FIP whereas only 32.4% (34/105) of type I FCoV-infected cats succumbed to FIP. Using a chi-square test the correlation of FCoV type with FIP disease was calculated. Among the 118 FCoV strains (105 type I and 13 type II) analyzed, type II virus appeared to be highly significantly correlated with the disease (p = 0.0046) (Table 2 ).
Table 2.
Disease correlation of type I and II FCoV.
| Group | Number of FCoV strain |
||
|---|---|---|---|
| Type I | Type II | Total | |
| FIP | 34 (32.4%) | 10 (76.9%) | 44 |
| Healthy | 71 (67.6%) | 3 (23.1%) | 74 |
| Total | 105 | 13 | 118 |
3.3. Genetic comparison and phylogenetic analysis of the local type I and II FCoVs
FCoV originated from 10 different households were chosen to further understand the evolutionary track of our local type I and II FCoVs (Table 3 ), a region of the S gene from local strains that were circulating yearly from 2003 to 2007 were sequenced and compared with that of reference FCoV, CCoV and wild animal CoV strains. In comparison to the low nucleotide sequence similarity between type I and II FCoV (56.2–63.6%), homology within all analyzed type I viruses (83.3–95.2%) and within all type II viruses (91.7–99.4%) appeared to be much higher (Fig. 1, Fig. 2 ). The nucleotide divergence between local FCoVs was between 6.2% and 11.7% for five type I viruses (Table 4 ) and between 0.6% and 3.1% for five type II viruses (Table 5 ). These findings revealed that, within the time frame of this study, type I FCoVs displayed more genetic diversity than type II viruses. The phylogenetic trees constructed using the partial S gene sequences are shown in Fig. 3 . Apart from one local type I virus (NTU2R) and local CCoVs (NTU2 and 9) that originated from the same lineage, other type I viruses displayed divergence. Surprisingly, four of our local type II viruses (NTU30A, 39A, 46Ce and 205C) and a Japanese FCoV (F21-1) clustered with two wild animal CoVs from China (raccoon dog CoV/GZ43/03 and Chinese ferret badger CoV/DM95/03), and they were distinct from the reference type II FCoVs strains 79-1146 and 79-1683.
Table 3.
Signalment of 10 cats stratified according to different type of FCoV analysis.
| Animal |
FCoVs |
||||||
|---|---|---|---|---|---|---|---|
| Cat no.a | Year | Age | Gender | Breed | Type | Denominationb | Denomination in brief |
| 1 | 2003 | 6.5 months | F | Domestic short hair | I | FCoV/NTU2/R/’03 | NTU2R |
| 2 | 2004 | 1 year | F | Domestic short hair | I | FCoV/NTU22/P/’04 | NTU22P |
| 3 | 2005 | 5 months | M | Domestic short hair | I | FCoV/NTU45/P/’05 | NTU45P |
| 4 | 2006 | 4.5 months | M | Persian | I | FCoV/NTU47/A/’06 | NTU47A |
| 5 | 2007 | 5 months | M | Domestic short hair | I | FCoV/NTU48/A/’07 | NTU48A |
| 6 | 2004 | 3 months | M | America short hair | II | FCoV/NTU30/A/’04 | NTU30A |
| 7 | 2005 | 3 months | F | Scottish Fold | II | FCoV/NTU39/A/’05 | NTU39A |
| 8 | 2006 | 5 months | F | Domestic short hair | II | FCoV/NTU46/Ce/’06 | NTU46Ce |
| 9 | 2007 | 3.5 months | M | Scottish Fold | II | FCoV/NTU156/P/’07 | NTU156P |
| 10 | 2007 | 5 months | F | Scottish Fold | II | FCoV/NTU205/C/’07 | NTU205C |
All but no. 1 of 10 cats were diagnosed FIP through postmortem histopathological examination.
Specimen source: R, rectal swab; P, pleural effusion; A, ascites; Ce, cerebellum; C, conjunctival swab.
Fig. 1.

Multiple nucleotide sequence alignment of the 168 bp fragment of the S gene from 12 type I FCoVs and 3 type I CCoVs. Only those nucleotides differing from the overall majority sequence are shown. The Taiwanese FCoV and CCoV GenBank accession numbers are EU513381–EU513384 and EU513390–EU513391, respectively. The reference strains include FCoV F13-27 (EF408013), FCoV Ku-2 (D32400), FCoV 99LVIPortugal_05A (EU327752), FCoV Black (DQ122859), FCoV UCD1 (AB088222), FCoV J10_0998 (AY159748), FCoV C1Je (DQ848678), and CCoV C15-60 (EF407998).
Fig. 2.

Multiple nucleotide sequence alignment of the 168 bp fragment of the S gene from 11 type II FCoVs, 2 type II CCoVs and 2 wild animal CoVs. The Taiwanese FCoV GenBank accession number is EU513385–EU513389. The reference strains include FCoV F21-1 (EF408020), FCoV F20-54-II (EF408022), FCoV HABER (DQ122859), FCoV DF-2 (DQ286389), FCoV 79-1146 (DQ010921), FCoV 79-1683 (X80799), CCoV C8-45 (EF408003), CCoV Chiba-40 (EF408004), RDCoV/GZ43/2003 (EF192155), and CFBCoV/DM95/2003 (EF192156).
Table 4.
Sequence divergence of local type I viruses.
| Strains | Strains |
|||
|---|---|---|---|---|
| NTU2 | NTU22 | NTU45 | NTU47 | |
| NTU22 | 6.2 | |||
| NTU45 | 8.9 | 10.3 | ||
| NTU47 | 6.2 | 10.9 | 10.9 | |
| NTU48 | 8.3 | 11.7 | 7.6 | 8.2 |
Table 5.
Sequence divergence of local type II viruses.
| Strains | Strains |
|||
|---|---|---|---|---|
| NTU30 | NTU39 | NTU46 | NTU156 | |
| NTU39 | 0.6 | |||
| NTU46 | 1.8 | 1.2 | ||
| NTU156 | 1.2 | 1.8 | 3.1 | |
| NTU205 | 0.6 | 1.2 | 2.4 | 0.6 |
Fig. 3.
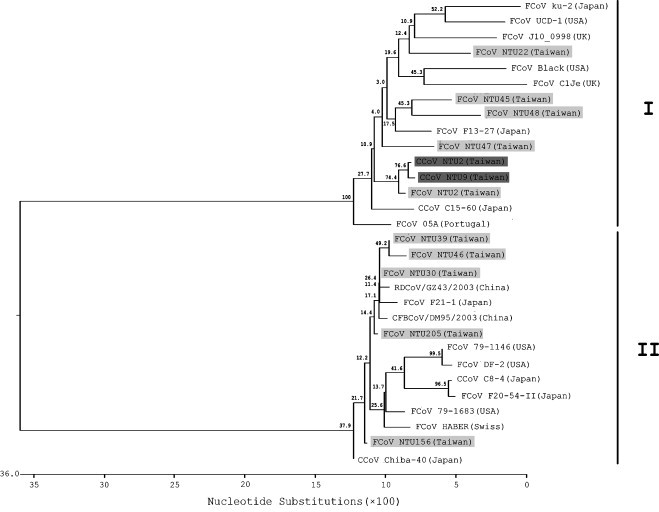
Phylogenetic relationships determined by the S gene sequences of the local FCoV and CCoV strains and reference FCoV, CCoV and wild animal CoVs strains. The phylogenetic trees were generated using the DNASTAR MegAlign program. The number of branches were calculated using bootstrapped values from 1000 replicates. The scale beneath the tree provides a key for the distance between sequences and the units at the bottom of the tree indicate the number of substitution events. Light grey underlay: local FCoV strains; dark grey underlay: local CCoV strains.
4. Discussion
The prevalence of type I and II FCoVs has been surveyed in many countries using serological and molecular assays (Addie et al., 2003, Benetka et al., 2004, Hohdatsu et al., 1992, Kummrow et al., 2005, Shiba et al., 2007, Vennema, 1999). In concordance with others findings, our results indicate that the majority of FCoV infection in Taiwan is due to type I viruses (88.7%). Our sequence analyses revealed that the type I virus is more genetically diverse than type II virus, a phenomenon that might be due to the large type I-infected population around the world and the possibility of a lengthy persistence of the virus in the asymptomatic cats (Addie et al., 2003, Herrewegh et al., 1997). Cats that are co-infected with both type I and II viruses were detected at a rather high frequency in this study (15.8%, 6 among a total of 38 FIP cases) (Table 1). Similar findings (6.8%, 5/74) were reported in a study that employed RT-PCR to type FCoV using organ pools from cats with FIP in Austria (Benetka et al., 2004). Did both types of viruses participate equally in the pathogenesis of FIP in these co-infected cats? Or one type plays a more significant role than the other? The question is currently under investigation.
A study conducted in Swiss cats using an IFA assay reported that infection with type I FCoV was more frequently associated with clinical signs and/or FIP (Kummrow et al., 2005). In contrast, using a different assay (RT-PCR), our results revealed that it is type II virus that significantly correlated with FIP (p = 0.0046). A previous seroprevalence study conducted by Hohdatsu et al. (1992) in Japan also concluded that FIP due to type II FCoV might be higher than type I viruses. In our study, type II FCoV was detected on the rectal swabs from three healthy cats using RT-PCR. Follow-ups were possible for two of these three cats. None of the rectal swabs collected during the past 3 years for these two cats were positive for FCoV (data not shown). Similar finding was observed from one specific pathogen-free cat experimentally infected with a type II virus (strain Wellcome) (Stoddart et al., 1988). Unlike other seven cats that died from the infection, this cat did not develop clinical signs of FIP and the viral shedding ceased at day 14 post-infection. Taken together with the observations from the aforementioned studies, infection with type II FCoV seems to follow a different course than infection with type I virus. Type I virus was demonstrated to be able to persist in cats living in multi-cat households for more than 6 years (Addie et al., 2003) and for at least 7 months in a single experimentally infected cat (Herrewegh et al., 1997). To our knowledge none of the similar finding has been reported in the naturally occurring type II FCoV infection. Type II virus is more likely to cause acute infections that result in either FIP or recovery without persistence. This distinction might explain why, in the epidemiological studies from various countries, the prevalence of type II virus is consistently lower than that of type I virus (Addie et al., 2003, Benetka et al., 2004, Hohdatsu et al., 1992, Kummrow et al., 2005, Shiba et al., 2007, Vennema, 1999).
A recent study indicated that FCoV (FCoV 79-1146 strain) and TGEV (Purdue strain) were most closely related to the wild animal CoV (Vijaykrishna et al., 2007). Since type II FCoV originated from a double recombination between type I FCoV and type II CCoV (Herrewegh et al., 1998, Pratelli et al., 2003), it was believed that the S gene of type II FCoV would be more closely related to type II CCoV. When the wild animal CoV, type II CCoV, and type II FCoV were aligned (Fig. 2), however, type II FCoVs had higher similarity to a raccoon dog virus strain CoV/GZ43/03 and a Chinese ferret badger CoV strain CoV/DM95/03 (97.6–99.4%) than to either reference type II FCoV (93.5–97.6%) or type II CCoV (95.2–98.8%). This finding indicates that, in addition to CCoV, there are viruses from other families of animals (e.g., Mustelidae) that share a common evolutionary origin in the S gene with type II FCoVs.
Acknowledgements
The authors would like to thank the cat owners without whose help this study could not be completed.
References
- Addie D.D., Jarrett O. A study of naturally occurring feline coronavirus infections in kittens. Vet. Rec. 1992;130:133–137. doi: 10.1136/vr.130.7.133. [DOI] [PubMed] [Google Scholar]
- Addie D.D., Toth S., Murray G.D., Jarrett O. Risk of feline infectious peritonitis in cats naturally infected with feline coronavirus. Am. J. Vet. Res. 1995;56:429–434. [PubMed] [Google Scholar]
- Addie D.D., Schaap I.A., Nicolson L., Jarrett O. Persistence and transmission of natural type I feline coronavirus infection. J. Gen. Virol. 2003;84:2735–2744. doi: 10.1099/vir.0.19129-0. [DOI] [PubMed] [Google Scholar]
- Bell E.T., Toribio J.A., White J.D., Malik R., Norris J.M. Seroprevalence study of feline coronavirus in owned and feral cats in Sydney, Australia. Aust. Vet. J. 2006;84:74–81. doi: 10.1111/j.1751-0813.2006.tb12231.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benetka V., Kubber-Heiss A., Kolodziejek J., Nowotny N., Hofmann-Parisot M., Mostl K. Prevalence of feline coronavirus types I and II in cats with histopathologically verified feline infectious peritonitis. Vet. Microbiol. 2004;99:31–42. doi: 10.1016/j.vetmic.2003.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cave T.A., Thompson H., Reid S.W., Hodgson D.R., Addie D.D. Kitten mortality in the United Kingdom: a retrospective analysis of 274 histopathological examinations (1986 to 2000) Vet. Rec. 2002;151:497–501. doi: 10.1136/vr.151.17.497. [DOI] [PubMed] [Google Scholar]
- Chomczynski P., Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate–phenol–chloroform extraction. Anal. Biochem. 1987;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- Dong B.Q., Liu W., Fan X.H., Vijaykrishna D., Tang X.C., Gao F., Li L.F., Li G.J., Zhang J.X., Yang L.Q., Poon L.L., Zhang S.Y., Peiris J.S., Smith G.J., Chen H., Guan Y. Detection of a novel and highly divergent coronavirus from Asian leopard cats and Chinese ferret badgers in Southern China. J. Virol. 2007;81:6920–6926. doi: 10.1128/JVI.00299-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartmann K., Binder C., Hirschberger J., Cole D., Reinacher M., Schroo S., Frost J., Egberink H., Lutz H., Hermanns W. Comparison of different tests to diagnose feline infectious peritonitis. J. Vet. Intern. Med. 2003;17:781–790. doi: 10.1111/j.1939-1676.2003.tb02515.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrewegh A.A., de Groot R.J., Cepica A., Egberink H.F., Horzinek M.C., Rottier P.J. Detection of feline coronavirus RNA in feces, tissues, and body fluids of naturally infected cats by reverse transcriptase PCR. J. Clin. Microbiol. 1995;33:684–689. doi: 10.1128/jcm.33.3.684-689.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrewegh A.A.P.M., Mahler M., Hedrich H.J., Haagmans B.L., Egberink H.F., Horzinek M.C., Rottier P.J.M., de Groot R.J. Persistence and evolution of feline coronavirus in a closed cat-breeding colony. Virology. 1997;234:349–363. doi: 10.1006/viro.1997.8663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrewegh A.A., Smeenk I., Horzinek M.C., Rottier P.J., de Groot R.J. Feline coronavirus type II strains 79-1683 and 79-1146 originate from a double recombination between feline coronavirus type I and canine coronavirus. J. Virol. 1998;72:4508–4514. doi: 10.1128/jvi.72.5.4508-4514.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hohdatsu T., Okada S., Ishizuka Y., Yamada H., Koyama H. The prevalence of types I and II feline coronavirus infections in cats. J. Vet. Med. Sci. 1992;54:557–562. doi: 10.1292/jvms.54.557. [DOI] [PubMed] [Google Scholar]
- Hohdatsu T., Izumiya Y., Yokoyama Y., Kida K., Koyama H. Differences in virus receptor for type I and type II feline infectious peritonitis virus. Arch. Virol. 1998;143:839–850. doi: 10.1007/s007050050336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horzinek M.C., Osterhaus A.D. Feline infectious peritonitis: a worldwide serosurvey. Am. J. Vet. Res. 1979;40:1487–1492. [PubMed] [Google Scholar]
- Kiss I., Kecskemeti S., Tanyi J., Klingeborn B., Belak S. Prevalence and genetic pattern of feline coronaviruses in urban cat populations. Vet. J. 2000;159:64–70. doi: 10.1053/tvjl.1999.0402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kummrow M., Meli M.L., Haessig M., Goenczi E., Poland A., Pedersen N.C., Hofmann-Lehmann R., Lutz H. Feline coronavirus serotypes 1 and 2: seroprevalence and association with disease in Switzerland. Clin. Diagn. Lab. Immunol. 2005;12:1209–1215. doi: 10.1128/CDLI.12.10.1209-1215.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai M.M.C., Perlman S., Anderson L.J. Coronaviridae. In: Knipe D.M., Howley P.M., Griffin D.E., Lamb R.A., Martin M.A., Roizman B., Straus S.E., editors. Fields Virology. Lippincott Williams & Wilkins; Philadelphia, PA: 2007. pp. 1305–1335. [Google Scholar]
- McKeirnan A.J., Evermann J.F., Hargis A., Miller L.M., Ott R.L. Isolation of feline coronavirus from two cats with diverse disease manifestation. Feline Pract. 1981;11:16–20. [Google Scholar]
- Pedersen N.C. Serologic studies of naturally occurring feline infectious peritonitis. Am. J. Vet. Res. 1976;37:1449–1453. [PubMed] [Google Scholar]
- Pedersen N.C., Boyle J.F., Floyd K., Fudge A., Barker J. An enteric coronavirus infection of cats and its relationship to feline infectious peritonitis. Am. J. Vet. Res. 1981;42:368–377. [PubMed] [Google Scholar]
- Pedersen N.C., Black J.W., Boyle J.F., Evermann J.F., McKeirnan A.J., Ott R.L. Pathogenic differences between various feline coronavirus isolates. Adv. Exp. Med. Biol. 1984;173:365–380. doi: 10.1007/978-1-4615-9373-7_36. [DOI] [PubMed] [Google Scholar]
- Pratelli A., Martella V., Decaro N., Tinelli A., Camero M., Cirone F., Elia G., Cavalli A., Corrente M., Greco G., Buonavoglia D., Gentile M., Tempesta M., Buonavoglia C. Genetic diversity of a canine coronavirus detected in pups with diarrhoea in Italy. J. Virol. Methods. 2003;110:9–17. doi: 10.1016/S0166-0934(03)00081-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiba N., Maeda K., Kato H., Mochizuki M., Iwata H. Differentiation of feline coronavirus type I and II infections by virus neutralization test. Vet. Microbiol. 2007;124:348–352. doi: 10.1016/j.vetmic.2007.04.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sparkes A.H., Gruffydd-Jones T.J., Howard P.E., Harbour D.A. Coronavirus serology in healthy pedigree cats. Vet. Rec. 1992;131:35–36. doi: 10.1136/vr.131.2.35. [DOI] [PubMed] [Google Scholar]
- Stoddart M.E., Gaskell R.M., Harbour D.A., Gaskell C.J. Virus shedding and immune responses in cats inoculated with cell culture-adapted feline infectious peritonitis virus. Vet. Microbiol. 1988;16:145–158. doi: 10.1016/0378-1135(88)90039-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vennema H. Genetic drift and genetic shift during feline coronavirus evolution. Vet. Microbiol. 1999;69:139–141. doi: 10.1016/S0378-1135(99)00102-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vijaykrishna D., Smith G.J., Zhang J.X., Peiris J.S., Chen H., Guan Y. Evolutionary insights into the ecology of coronaviruses. J. Virol. 2007;81:4012–4020. doi: 10.1128/JVI.02605-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woo P.C., Wang M., Lau S.K., Xu H., Poon R.W., Guo R., Wong B.H., Gao K., Tsoi H.W., Huang Y., Li K.S., Lam C.S., Chan K.H., Zheng B.J., Yuen K.Y. Comparative analysis of twelve genomes of three novel group 2c and group 2d coronaviruses reveals unique group and subgroup features. J. Virol. 2007;81:1574–1585. doi: 10.1128/JVI.02182-06. [DOI] [PMC free article] [PubMed] [Google Scholar]