

Prevalence of neurologic disease in cats and the preponderance of cases with no known cause
Neurologic disease is seen commonly in cats. For example, neurologic cases make up approximately 10% of the case load of two separate feline medicine referral clinics (D.A. Gunn-Moore, BVM&S, PhD, Edinburgh University Feline Clinic data, 2004) [1]. There are many well-documented causes of neurologic disorder in cats [2], [3], and infectious causes are believed to account for 30% to 45% of cases [1], [4]. It is important to realize that a specific cause cannot be identified in 12% to 40% of cases, however. This holds true when looking at clinical cases [2], [5], [6] and histopathologic data ( Table 1) [1], [4]. In addition, although there are many known infectious causes ( Box 1), a large number of cases (35%–40%) are found to have histopathologic changes suggestive of viral infection (that is not consistent with feline infectious peritonitis [FIP]), but no causal agent can be identified ( Table 2) [1], [4]. This group becomes even more significant if you consider cats with particular clinical signs. For example, of 30 cats that were investigated for having recurrent seizures, all were found to have structural brain disease and 14 (47%) had nonsuppurative meningoencephalitis suggestive of a viral infection, but no infectious agent could be found [5].
Table 1.
Histopathologic diagnoses in 286 feline neurology cases (University of Bristol 1975–1998)
| Cause | Number | Percent |
|---|---|---|
| Inflammatory/infectious | 92 | 32 |
| No abnormalities detected | 51 | 18 |
| Degenerative | 42 | 15 |
| Neoplasia | 38 | 13 |
| Feline dysautonomia | 27 | 9 |
| Feline spongiform encephalopathy (FSE) | 24 | 8 |
| Congenital | 12 | 4 |
Data from Bradshaw JM, Pearson GR, Gruffydd-Jones TJ. A retrospective study of 286 cases of neurological disorders of the cat. J Comp Pathol 2004;131:112–20.
Box 1. Naturally occurring infectious causes of central nervous system disease in domestic cats.
- Viral
- Feline coronavirus (FCoV)a
- Feline panleukopenia virus (FPV)
- Feline immunodeficiency virus (FIV)
- Feline leukemia virus (FeLV)
- Rabies virus
- Aujeszky's disease virus
- Feline herpesvirus-1 (FHV-1)
- Borna disease virus (BDV)
- Certain arboviruses (see text)
- Bacterial
- Pasturella
- Staphylococcus
- Other aerobic organisms
- Anaerobic organisms
- Mycobacteria
- Protozoal
- Toxoplasmosisa
- Rickettsial
- Ehrlichiosis
- Fungal
- Cryptococcosisa
- Blastomycosis
- Histoplasmosis
- Aspergillosis
- Dematiaceous fungi
- Parasitic
- Cuterebra larval myiasis
- Visceral larva migrans (eg, Toxocara)
- Sarcocystis
- Dirofilaria immitis
- Probable and other
- Feline spongiform encephalopathy (FSE)
- Feline polioencephalomyelitis and miscellaneous nonsuppurative (meningo)encephalitidesa
aMost common causes of encephalitis in cats. Others are sporadic and rare.
Table 2.
Diagnoses in 92 feline neurologic cases found to have central nervous system histopathology consistent with inflammation and/or infection (University of Bristol 1975–1998)
| Cause | Number | Percent |
|---|---|---|
| Feline infectious peritonitis (FIP)a | 47 | 51 |
| Viral (non-FIP)b | 32 | 35 |
| Protozoal cysts (eg, toxoplasmosis) | 8 | 9 |
| Bacterial infection | 3 | 3 |
| Feline immunodeficiency virus (FIV) | 1 | 1 |
| Cryptococcosis | 1 | 1 |
Data from Bradshaw JM, Pearson GR, Gruffydd-Jones TJ. A retrospective study of 286 cases of neurological disorders of the cat. J Comp Pathol 2004;131:112–20.
One of the cats with FIP was also found to have an incidental nematode larvae (Toxocara) within its lateral ventricle.
Nonsuppurative meningitis and/or encephalitis was present, but no cause could be found. Five of these cats also had changes consistent with feline spongiform encephalopathy.
From this, we can see that infectious disease is a common cause of central nervous system (CNS) disorders in cats. In addition, the most common infectious agents are feline coronaviruses (FCoV, which can cause FIP) and some other, as yet unidentified, infectious agent(s), which are probably viruses [1], [5]. Unfortunately, because we cannot identify the cause, we do not know how best to treat these cases, nor do we know how to prevent them. It is therefore essential that we try harder to identify the etiology behind the pathologic findings. This means performing more detailed diagnostics on individual clinical cases, such as looking for potential infectious organisms by serology, cerebrospinal fluid (CSF) IgG quotient, and IgG index [6]; CSF polymerase chain reaction (PCR) assays to detect the infectious organism's DNA or RNA [7]; and immunohistochemistry or PCR on brain samples collected postmortem. In addition, it underlines the need for further experimental investigation into potential pathogens. It is only by adopting a more questioning approach to disease pathogenesis that we can hope to determine what may be causing neurologic disease in many of our pet cats.
Reasons for the increased recognition of infectious central nervous system disease in cats
Improved diagnostics and changing concepts in disease pathogenesis
Over the past 10 years, there has been a dramatic increase in the recognition and understanding of many different infectious diseases and of how they can affect the CNS. Advances in molecular technology have enabled the detection of pathogens within the CNS (eg, by using PCR assays), led to the recognition of new infectious diseases, and expanded our understanding of the etiopathogenesis of these infections.
With the changing understanding of etiopathogenesis, we have had to redefine our concept of “infectious disease.” No longer can we think that infectious agents can only cause acute disease that classically fulfills Koch's postulates of cause and effect. For example, the acute disease that is seen when a bacterial infection spreads from otitis media to cause suppurative meningoencephalitis [8]. We now know that some diseases result from chronic insidious infections and that this is particularly true within the relatively protected confines of the CNS. In addition, progressively more diseases are being identified that, although being associated with the presence of a particular pathogen, require a number of other factors to occur concurrently before disease becomes apparent. For these diseases to develop, there has to be a specific interaction between the infectious organism, host factors (especially genetics affecting the immune system), and the environment.
Unfortunately, establishing a causal relation can be difficult, particularly when the prevalence of the infection is high in the general population but only a few individuals have the necessary factors required for clinical signs to develop. Serologic surveys have been largely responsible for recognizing the role of infectious organisms in this type of disease. After detecting a serologic relation, it is then possible to use more complex molecular biology techniques to detect the pathogen within a particular individual or particular pathologic lesion. It is highly likely that it will be by using this type of approach that the causes for many feline CNS disorders will be found (eg, Borna disease [BD]).
Changing population dynamics
Populations are changing; people are living in progressively larger urban groups, and international travel is now commonplace. This allows for rapid spread of disease, not only among human beings but from human beings to other species. In addition, as the global human population increases, the demand for housing means that previously unexplored habitats are being developed, new pathogens are being exposed, and old pathogens are finding new hosts. There are a number of examples of infections that have crossed between species because of altered population dynamics, and many of them involve feline species. Examples include canine distemper virus (CDV), which is now causing disease in a number of large feline species, particularly lions in the Serengeti [9], but has not yet been detected in domestic cats [10]; severe acute respiratory syndrome (SARS), which is caused by a coronavirus that seems to have been passed from civet cats in China to human beings (although the civet cat is not actually a feline species) [11], [12]; avian influenza virus (H5N1), which has killed domestic and captive wild felids in Thailand [13]; and West Nile virus (WNV), which is now present in the United States, being spread by mosquitoes to many wild and captive birds as well as to horses and humans beings [14].
WNV is a particularly interesting infection to consider. This is because experimental studies have shown that it is relatively easy to transmit WNV to cats by a mosquito bite or orally via consumption of infected prey [15]. Experimental cats in one study showed only mild nonneurologic signs [15]. This does not mean that WNV cannot cause neurologic disease in cats, however. This is because dogs in the same study remained perfectly healthy [15] but have been shown to develop encephalitis and myocarditis in a separate study [16]. To date, domestic cats tested in New York City have not been found to be seropositive for WNV [17]. Seropositive cats have been identified in the United States, however, and several cases demonstrated seroconversion coincident with neurologic illness (A. Glaser, DVM, PhD, Cornell University Animal Health Diagnostic Laboratory, personal communication, 2004). The potential role that WNV may play in feline neurologic disease requires further investigation.
Increasing demand for inexpensive food
An increasing demand for inexpensive food has resulted in a growing number of food-related infections. In addition to classic types of food poisoning, the transmission of the transmissible spongiform encephalopathies (TSEs) should be considered in this group.
Increasing awareness of zoonotic infections
It is perhaps of some concern that there has been a particular increase in the recognition of zoonotic conditions (diseases that can be spread from animals to people). In fact, three quarters of all emerging human pathogens are zoonotic [18]. Because of this, it is important that we raise our general awareness of this type of disease and monitor closely for any evidence of interspecies transfer of infections.
By studying the genetic relation between pathogens and performing infectivity studies, a number of infectious agents have already been identified that can cross species barriers and raise the possibility of zoonotic infection. Two such diseases are feline spongiform encephalopathy (FSE) and BD. In both cases, the infectious agents can infect a number of mammalian species, including cats and people. In addition, both infections have a poorly understood etiopathogenesis and may result in terminal neurologic disease.
Known causes of central nervous system infection in cats
From the data shown in Table 1, Table 2, we can see that FIP and nonsuppurative encephalitides (also called viral non-FIP encephalitides) are the only two commonly recognized potentially infectious causes of CNS disease in the cat. Other infections, for example, toxoplasmosis, feline leukemia virus (FeLV), feline immunodeficiency virus (FIV), feline panleukopenia (FPV), and fungal and parasitic infections, are seen only rarely. Detailed summaries of the pathogenesis, clinical signs, diagnostic approach, treatment, and neuropathologic findings of these specific infections are readily available [1], [4], [6], [7], [19], [20], [21], [22], [23], [24]. The rest of this article therefore adds only selected comments in relation to these organisms and focuses on the nonsuppurative encephalitides and the more unusual infections for which information is less readily available.
Feline infectious peritonitis
FIP is the most commonly detected infectious cause of neurologic disease in cats. It accounts for 45% to 50% of all cases associated with inflammatory changes, which equates to 15% to 20% of all feline neurologic cases [1], [4]. It is essential to realize just how common this infection is and that although most clinical cases are seen in young pedigree cats, usually with obvious systemic involvement, this is not always the case.
As with many viral infections affecting the CNS, histopathologic examination reveals nonsuppurative meningoencephalomyelitis, with perivascular cuffing and meningeal infiltration with mononuclear cells, gliosis, and variable neuronal degeneration. The inflammation is often pyogranulomatous, is located around the lateral ventricles or in the meninges, or may affect the choroid plexus. In addition, vasculitis or acquired hydrocephalus may be present, and systemic changes are usually apparent [1], [25], [26].
Feline immunodeficiency virus
FIV can cause neurologic disease as a direct primary neurotropic effect of the virus or via secondary opportunistic infections, such as FIP, toxoplasmosis, or cryptococcosis [27], [28], [29], [30], [31], [32]. It is important to note that this virus cannot be excluded on the basis of negative serology. This is because several studies have failed to detect antibody in some virus-positive individuals [27], [33], and in one report, approximately 20% of cats naturally infected with FIV were antibody-negative [33]. Although the reason for this is unknown, it has been suggested that some cats were tested early in the course of infection, before the development of an antibody response. These cats are unlikely to show CNS disease. Alternatively, some cats may have been tested late in the disease, once antibody levels had fallen, along with the terminal decline of the immune system [27]. Other factors may also play a role, because some cats fail to produce a detectable antibody response to FIV at any time during infection [33]. Therefore, to confirm that a cat is not infected with FIV, it may be necessary to perform PCR on a blood or CSF sample or PCR or immunohistochemistry on brain sections collected postmortem [32].
Rare or experimental infectious causes of central nervous system disease in cats
Naturally occurring and clinically significant CNS infections have occasionally been seen in cats, resulting from a wide range of organisms. These include feline herpesvirus-1 (FHV-1) [34], feline calicivirus (FCV) [35], dematiaceous fungi (Cladophialophora bantiana) [36], and nematodes (eg, Sarcocystis neurona) [37].
It is also prudent to consider clinically significant CNS infections that have been produced experimentally in cats. These include Bartonella henselae [38], which typically causes cat scratch disease in people [39]; equine herpesvirus-9 [40]; Newcastle disease virus [41]; human poliovirus [42]; and simian cytomegalovirus-related stealth virus (which was taken from a human being with chronic fatigue syndrome) [43].
Much speculation has concerned the arboviruses (the arthropod-borne encephalomyelitis group) and whether or not they may be responsible for significant natural CNS infection in cats [44]. Many of these viruses have been shown to cause natural or experimental infection in cats. For example, natural infections have been seen in cats with BD virus (BDV), which can be subclinical or clinically significant. Antibodies to St. Louis encephalitis virus, Japanese encephalitis virus, yellow fever virus, Tenshaw virus, Snowshoe hare virus, Jamestown Canyon virus, and Powassan virus have been found in free-living cats, indicating previous subclinical infections [45], [46], [47]. Experimental infections have been documented with Near Eastern equine encephalitis virus [48], [49], Powassan virus [46], and Rift Valley fever virus in kittens [47]. Many of these viruses have a wide geographic distribution, including the United States and Europe, and some are endemically present in wild mammal populations to which free-living cats may become exposed [50]. Although these studies show that cats are potentially at risk of becoming infected with these organisms, their role in causing a significant incidence of naturally arising feline neurologic disease remains to be determined.
Borna disease virus
Epizootiology
Classical BD is a severe neurologic disease that is seen predominantly in horses and sheep in endemic areas of Germany and Switzerland. Natural infections have also been seen in cats and ostriches and, occasionally, in rabbits, cattle, goats, deer, foxes, and dogs [51], [52], [53], [54], [55], [56], [57], [58], [59]. Experimentally, BDV can also be transmitted to birds, rodents, and monkeys, and it is likely that the host range includes all warm-blooded animals and birds [52], [58], [60], [61]. The exact geographic distribution of the virus is uncertain, but serologic evidence has documented infection in Europe, the United States, and Asia [62], [63], [64], [65], [66], [67], [68], [69], [70].
BD in cats is also known as “staggering disease.” It was first described in Sweden [71] and later shown to be caused by BDV [72]. Based on serologic surveys or surveys looking for BDV RNA in peripheral blood samples, it is clear that BDV infection is usually asymptomatic. The prevalence of seropositivity increases steadily with age in cats [72], [73] and, interestingly, seems to be higher in cats that are also FIV antibody-positive [68], [74]. In the United Kingdom, 6% of cats with no evidence of CNS disease have antibodies against BDV [65], as do 9% of ill cats submitted for FIV, FeLV, and FIP virus testing (D.A. Harbour, PhD, unpublished results from 654 cat blood samples, 1999). In Japan, 13% to 22% of healthy randomly selected cats are seropositive for BDV [62], [75]. Antibodies against BDV or BDV RNA are seen most frequently in cats with neurologic disease, however. When cats with undefined neurologic disorders were investigated, 13% were found to be BDV antibody-positive in Germany [76] compared with 35% in the United Kingdom [65] and 67% in Japan [77]. Although most documented cases of feline BD have originated from northern and central Europe, probable cases have been seen in many other countries [65] (see section on nonsuppurative encephalomyelitis of unknown cause). Given the difficulty in making a premortem diagnosis (and even a postmortem diagnosis) and the low index of suspicion, it is likely that BD is underdiagnosed.
Pathogenesis
The source of BDV infection is rarely known. Infected cats usually, but not always, have outdoors access, particularly to rural or woodland areas, however [78], [79], [80]. This has led to the suggestion that rodents or wild birds may be viral carriers [80], [81], [82]. Natural infections are believed to be transmitted via saliva or nasal secretions [52], [73], [83].
BDV is a neurotropic RNA virus [84], [85] that is genetically stable [86], [87]. It is ubiquitously distributed and seems to have many well-adapted species-specific biotypes [54], [72], [88], [89], [90]; in most cases, infection causes little or no sign of disease. It is currently believed that clinical signs only develop when a host is exposed to a particular strain of BDV [57], [62], [73], [91] or is particularly susceptible and mounts an abnormal immune response to the virus (ie, disease seems to result from a T-cell–dependent immune mechanism) [60], [91], [92], [93].
Clinical signs
Natural BD has been reported in more than 100 cats [54], [78], [79], [80], [94]. It is seen most frequently in male cats, with no particular breed predisposition. Although a wide age range of cats may be affected (from 5 months to 11 years of age), young adults seem to be most at risk [54], [78], [79]. Disease is characterized by behavioral and motor disturbances resulting from meningoencephalomyelitis. In experimental infections, clinical signs included protrusion of the third eyelid, behavioral changes, circling, ataxia, and tremors [72]. Natural infections may present with progressive hind limb ataxia, loss of appetite, fever, increased affection toward the owner, unusual staring expression, apparent pain over the sacrum, increased salivation, aggression, an inability to retract claws, seizures, focal or generalized pruritus, hypersensitivity to light and sound, or constipation [76], [78], [79]. Occasional atypical cases have been seen, for example, causing muscle fasciculation and proprioceptive defects (without evidence of encephalitis) [95]. Disease is usually progressive, and mortality rates are high because affected cats usually warrant euthanasia within a week to 6 months [78], [79]. Cats that survive the initial episode may remain chronically infected or may experience recurrent episodes of disease [72]. Although fatal BD is seen most commonly as a rare isolated event, it can occasionally occur as a large outbreak, where as many as 30 to 40 cases may be seen in a week [78].
Diagnosis
Premortem diagnosis is difficult. In most cases, typical clinical signs in a cat from an endemic area result in a presumptive diagnosis of BD. Unfortunately, detection of serum antibodies is not reliable. This is because although raised serum antibodies are present in some cats with BD (∼40%) [76], particularly those with acute disease, others may be antibody-negative, particularly if they have subacute or chronic disease [54], [62], [65], [72], [79]. Although clinical signs of BD tend to develop at the same time that BDV RNA can be detected within the peripheral blood [89], this does not necessarily reflect the extent of the viral load in the CNS [60], and asymptomatic cats can also be positive [75]. Routine serum biochemistry and hematology are generally unremarkable, although some cats may show a leukopenia, and mild elevations in glucose or alanine aminotransferase (ALT) levels may also occur [72], [78]. CSF analysis may show a leukocytosis with mononuclear cells predominating, protein levels may be increased (Table 3) [78], and antibodies to BDV may be detected [92].
Table 3.
Infectious disease and typical cerebrospinal fluid changes
| Disease | CSF Pressure | CSF Appearance | WBC Counta | WBC Type | Total Protein concentrationb | Albumin | Globulin | CSF Antibodies detectable | Organisms visible |
|---|---|---|---|---|---|---|---|---|---|
| Feline infectious peritonitis (FIP) | WNL or ↑ | Clear or turbid | +++ (WNL- ++) | PMN-mono-mixed | +++ (WNL-+) | ++ | ++ | Yes | No |
| Other viral encephalitis (eg, Borna disease)c | WNL | Clear (turbid) | + (++) | Mono | + (++) | WNL | ? | No | No |
| Protozoal meningoencephalitis (eg, toxoplasmosis) | WNL or ↑ | Xanthochromic | + (++) | Mixed-PMN, eos, mono | + (++) | + | + | Variable | Rarely |
| Fungal meningoencephalitis (eg, cryptococcosis) | ↑ or viscous | Turbid, xanthochromic | ++ | Mixed-PMN, mono, eos | ++ | ++ | +(+) | Varies | Varies |
| Bacterial meningitis | WNL or ↑ | Turbid | ++ (+++) | PMN (mixed) | ++ (+++) | ++ | ++ | Varies | Yes (varies) |
Abbreviations: CSF, cerebrospinal fluid; eos, eosinophils predominate; FIP, feline infectious peritonitis; mono, mononuclear cells (ie, lymphocytes, monocytes, macrophages) predominate; PMN, polymorphonuclear cells (neutrophils) predominate; WBC, white blood cell; WNL, within normal limits; ↑ = increased.
Symbols in parentheses indicate less frequently seen variations.
Reference range for WBC count = <4 per microliter; + = 5–80 per microliter; ++ = 81–500 per microliter; +++ = >500 per microliter.
Reference range for total protein concentration = <25 mg/dL; + = 25–100 mg/dL; ++ = 100–300 mg/dL; +++ = >300mg/dL.
Some viral infections cause neuropathologic changes without inflammation, and these may alter the CSF little [43].
Histopathologically, BD usually results in nonsuppurative meningoencephalomyelitis, with neuronophagia, microgliosis, and heavy perivascular cuffing by mononuclear cells [10], [72], [78]. Occasional cases seem to result in neurologic signs without evidence of associated inflammation [95]. In most cases, lesions are particularly evident in the gray matter of the cerebral hemispheres, the limbic system, and the brain stem [10], [54], [78]. The cerebellum and spinal cord are less frequently affected [79].
Confirmation that BDV is the cause of this disease has been demonstrated by experimental transmission studies to cats and rabbits [72], [79]. In addition, BDV antigen may be detected by immunohistochemistry or enzyme-linked immunosorbent assay (ELISA) [54], and BDV RNA may be detected by PCR; all three methodologies can be performed on brain samples. Clinical cases are most easily confirmed by detecting BDV RNA within the inflamed areas of the brain using PCR [54], [65], [72], [96], [97].
Treatment
There is no specific treatment for BD. Supportive care and corticosteroids may help in some cases. Prednisolone may be given orally at a rate of 1 to 2 mg/kg every 24 hours until clinical signs regress, after which it should be reduced gradually over several weeks or months.
Zoonotic risk
It is currently unclear what role BDV may play in the induction of human disease. Antibodies against BDV, BD viral proteins, and BDV RNA have been found in people in Europe, the United States, and Asia. A higher prevalence of infection is seen in patients with neurologic or psychiatric disorders, particularly schizophrenia and uni- or bipolar disorders [61], [70], [98], [99], [100], [101], [102], [103], [104], [105], [106]. Because the virus has also been detected in clinically normal patients [67], [101], [107], however, its role in the development of these complex psychiatric disorders has still to be proven [61], [63].
The presence of BDV infection in many domestic species as well as evidence of cross-species transfer raises the possibility of zoonotic spread. Although animal species may pose a potential risk to people, finding BDV RNA in blood from normal human blood donors suggests that people may also be at risk from horizontal spread from person to person [64]. Considerably more investigation needs to be performed before the zoonotic potential of BDV can be determined.
Nonsuppurative encephalitides of unknown cause
Introduction and geographic distribution
A number of other nonsuppurative encephalitides have also been described in cats. These seem to comprise a group of diseases that are possibly related, and the histopathologic changes suggest a viral origin. They are geographically widespread and have been reported in Australia [108], the United States [109], Canada [4], [5], [23], Sweden [10], [71], [78], Norway [110], Switzerland [44], and the United Kingdom [1], [111], [112]; other potential cases have been seen as widely distributed as Morocco [113] and Sri Lanka [114]. A similar condition has been found in a number of large cats, including lions and tigers [115], [116], [117], [118]. In all cases, the reports state an unknown cause but comment that the histopathologic changes are suggestive of viral infection. Unfortunately, early studies performed few diagnostic investigations to try to determine the possible cause. Later studies usually assessed for FIV, FeLV, FIP, and Toxoplasma gondii and, in some cases, for FHV-1, FCV, FPV, and Borrelia burgdorferi. In almost all cases, the cats have been found to be negative for all these agents [1], [4], [5], [10], [23], [44], [112].
Reviewing the data available on these cases suggests that although these diseases generally affect cats of a similar age and sex and cause a range of rather similar clinical signs, they seem to separate into two groups based on histopathologic changes:
Group I: nonsuppurative encephalomyelitis
Clinical signs
These cats are of a wide age range (from a few months to >18 years), but young adults seem to be overrepresented. They show no sex or breed predisposition. They tend to have an acute duration of illness and typically develop ataxia, nystagmus, seizures, head tremor, anorexia, apathy, fever, and, occasionally, preceding vomiting and diarrhea (although not all cats show all signs) [4], [6], [10], [71], [78].
Diagnosis
A premortem diagnosis is typically based on the presence of suggestive clinical signs and typical CSF changes (see Table 3) [4], [6]. Some cats show leukopenia or a mildly increased ALT concentration [4], [78]. Neuroimaging of the brain may reveal multifocal areas of contrast enhancement suggestive of inflammatory disease.
Histopathologic examination reveals mild to severe nonsuppurative meningoencephalomyelitis characterized by mononuclear perivascular cuffing, inflammatory nodules of lymphocytes and macrophages, and neuronal degeneration. Changes can occur throughout the brain and spinal cord but are most prominent in the thalamocortex and brain stem [4], [78]. Lesions may be diffuse or focal [4].
Treatment
There is no curative treatment, but supportive therapies include anticonvulsants to control the seizures and, possibly, corticosteroids to reduce CNS inflammation. Because the disease is often self-limiting, the prognosis is quite good for those cats in which neurologic signs are not too severe [6].
Potential causes
Interestingly, when blood samples from affected Swedish and Austrian cats [10], [71], [78], [79] were retrospectively assessed by serologic testing for the organisms listed previously, they were found to be negative, as were their brain sections when assessed by immunohistochemistry for T gondii, CDV, FHV-1, tick-borne encephalitis, and Aujeszky's disease virus [10], [79]. After finding that the cats were seropositive for BDV and then performing experimental transmission studies, however, subsequent publications from the same authors determined that these cases had actually been caused by BDV [72], [79]. It has therefore been suggested that BDV may be responsible for more of these cases. Although this remains a possibility, it is also possible that there are a number of other previously unrecognized viruses. For this reason, it is important that these cases be fully studied before any causal relation can be proven (for diagnostic methods, see section on prevalence of neurological disease in cats and the preponderance of cases with no known cause) [6], [7]. In addition, because BDV can be found in the CNS of some clinically normal individuals [65], its presence, per se, within the CNS of a cat showing neurologic disease does not prove that it is the cause of the disorder.
Group II: polioencephalomyelitis or polioencephalitis
Clinical signs
These diseases also tend to affect younger cats (from a few months old to middle-aged), and there is no sex or breed predisposition. Affected individuals tend to have a subacute to chronic course that may last for months. Partial recovery may be seen in some cats, which can go on to live for many years [44], [109], [112]. Disease is most commonly sporadic [44], [109], [119]; however, there are also reports describing what seems to be the chronic form of this condition within large groups of research cats in the United Kingdom [111], [112].
Cats with polioencephalomyelitis or polioencephalitis present with problems of locomotion, including ataxia, paresis, and depressed postural reaction in all four limbs. Affected cats may occasionally show hyperesthesia or even lower motor neuron signs (muscle atrophy and decreased tendon reflexes). They can also show tremors, pupillary abnormalities, defective vision, nystagmus, and seizures [44], [109], [111], [112], [119]. When seizures occur, they do so as episodes of multiple short seizures [109]. Affected cats rarely show any other signs of systemic disease [109].
Diagnosis
A premortem diagnosis is typically based on the presence of suggestive clinical signs and typical CSF changes (see Table 3) [4], [6]. Some affected cats may be leukopenic or anemic (with myeloid hypoplasia) [109].
Early on in the disease, histopathologic examination reveals disseminated inflammatory lesions in the brain and spinal cord, with the spinal cord and medulla oblongata being most severely affected. The changes are those of polioencephalomyelitis or polioencephalitis. The lesions consist of perivascular mononuclear cuffing, gliosis, and neuronal degeneration, with the latter being most obvious in the ventral horns of the spinal cord [44], [109], [119].
In chronic cases, little inflammation remains. There is extensive neuronal loss and intense astrogliosis, however, particularly in the spinal cord. Wallerian degeneration arises secondary to the neuronal damage and is particularly evident in the lateral and ventral columns, where it may resemble a primary degenerative disorder [112], [119]. In addition, some cats have multifocal areas of Purkinje cell degeneration and gliosis in the cerebellar cortex [109], [112]. The changes are similar to those seen in human [120] and porcine poliomyelitis [121].
Treatment
There is no curative treatment. Supportive therapies are as previously discussed and include anticonvulsants to control seizures and, possibly, corticosteroids to reduce CNS inflammation. The prognosis can be good for those cats in which neurologic signs are not too severe.
Potential causes
The cause remains unknown. Genetic and nutritional causes seem unlikely [112], and a viral cause has been suggested by most authors [109], [112], [119]. Transmission studies have only been attempted occasionally and have been unsuccessful [108]. A number of infectious agents are known to be able to cause poliomyelitis or demyelination in cats. Some of these, including rabies virus, Aujeszky's disease virus, and Newcastle disease virus, have been ruled out on the basis of somewhat differing pathologic findings [109]. It has also been suggested that the condition may be an unusual manifestation of FPV infection [109], [112]. Although FPV classically causes cerebellar hypoplasia in kittens, with degeneration of the germinal and Purkinje cells [122], [123], it has occasionally been associated with inflammatory lesions within the brain, leukodystrophic lesions, neuronal degeneration, and gliosis of the spinal cord gray matter or spinal cord demyelination [124]. In addition, in FPV-vaccinated cats, FPV infection can still cause leukopenia and nonregenerative anemia [125]. Interestingly, 2 of 33 cases of clinical BD were seen in kittens that came from litters in which the rest of the litter had died of FPV [78]. Other suggested causes include FeLV [126] and arborviruses (see section on rare or experimental infectious causes of CNS disease in cats).
Although the pattern of disease is clinically and histopathologically distinct from that known to be caused by BDV, the author can find no evidence that this possibility has been investigated. Interestingly, the clinical signs and histopathologic findings of the chronic cases are somewhat similar to those of a case report in which the disease was attributed to an unusual case of BDV. It was an interesting case because it was found to have massive neuronal infection with BDV but lacked inflammatory change [95]. This poses an intriguing question as to whether this case was incorrectly attributed to BDV (with the BDV representing a striking but clinically insignificant finding), while also raising the possibility that most of the rest of the cases could actually be attributable to a second variant of BDV infection.
Central nervous system neuropathologic findings without inflammatory changes
Based on the histopathologic changes seen in the chronic cases of polioencephalomyelitis, it is important to realize that viral infections do not necessarily need to be associated with inflammatory changes within the CNS to cause clinically significant neurologic disease.
Neuropathologic changes, without inflammation, may be seen for a number of reasons. It may be that the inflammatory phase has passed and has been missed. Alternately, some viruses can induce direct neuropathic effects while hiding from the immune system within neurons or glial cells (eg, FIV [28]; CDV [127]; human stealth viruses [43]; many herpes viruses, such as chickenpox [128] and varicella zoster virus [129]). In addition, some perinatal infections can result in lasting CNS infection without the development of antibodies (circulating within the blood stream or within the CNS) or encephalitis, but they can still be associated with neurologic disease [95]. This has been seen with BDV infection of some rodents, where infected individuals develop subtle behavioral changes and defects in memory and learning [130], [131]. It is therefore possible that BDV infection or other viral infections could result in a number of different disease patterns, depending on differences in viral pathogenicity as well as on as yet unidentified host-specific factors.
Realizing that viral infections can cause CNS disease without obvious inflammatory change and recognizing that the changes can consist mainly of degenerative changes raise the possibility that a viral cause may be responsible for an even higher proportion of CNS disease in cats. Because approximately 35% of feline neurology cases are currently found to result from infectious or inflammatory causes and approximately 15% are degenerative, this could suggest that up to approximately 50% of all feline neurologic disease may potentially be caused by CNS infection (see Table 1) [1].
Unusual patterns of seizure activity
When seizures have been reported as part of the clinical syndromes described previously, the seizure pattern is rather striking; they occur in episodes of multiple short seizures [109]. When looking at studies focusing on all causes of recurrent seizures in cats, it is fascinating to see that they all result from structural brain disease and that the most common cause (∼50%) seems to be a result of nonsuppurative encephalitides [4], [5], [23]. Although the seizure pattern is similar, the cats in these studies were presented primarily for seizures rather than for ataxia, and despite the onset of seizures being rather dramatic, the prognosis was reasonably good [5].
Whether or not the same virus (or group of viruses) results in all forms of seizure-associated nonsuppurative encephalitides remains to be proven. Interestingly, experimental studies have shown that this pattern of seizure results from lesions in the periaqueductal gray matter of the midbrain [120]. It is therefore possible that it may simply be that this area is preferentially targeted by a number of different infectious agents in cats.
Feline spongiform encephalopathy
Epizootiology
FSE was first recognized in 1990 during the bovine spongiform encephalopathy (BSE) epidemic in the United Kingdom [132], [133], [134]. FSE is one of a group of naturally occurring TSEs. TSEs occur in many mammalian species [135], including scrapie in sheep and goats; BSE in cattle and captive exotic ungulates [136], [137]; FSE in domestic cats and captive exotic feline species, including the cheetah [137], [138], [139], puma [137], [140], and lion (A.L. Meredith, MA, VetMB, MRCVS, University of Edinburgh, personal communication, 1999); chronic wasting disease (CWD) of deer and elk [141]; transmissible mink encephalopathy (TME) in mink [142]; and Creutzfeldt-Jakob disease (CJD), variant CJD (vCJD), Gerstmann-Sträussler-Scheinker disease, and kuru in human beings [143]. Experimentally, TSEs can be transmitted to an even wider range of species, including rodents and nonhuman primates [144], [145], [146]. Although the widespread interest in TSEs developed only fairly recently, associated with the BSE epidemic and the recognition of vCJD, this type of disease is far from new. Historical records show that scrapie was first recognized approximately 300 years ago [147].
TSEs have been seen throughout the world. Although scrapie and human TSEs have a widespread distribution, BSE has been seen mainly in Europe, particularly in the United Kingdom. The situation is similar with FSE, with almost all cases having been seen in Britain; occasional cases have been seen in animals that had previously lived in the United Kingdom [148] or been fed on tissue from British cattle [135], [136], [137], [138], [139], [140], [141].
To understand FSE, it is necessary to know how BSE is believed to have originated. BSE was first reported in 1987 [149]. It is believed to have resulted from the inclusion of scrapie-infected sheep carcasses into feedstuffs for cattle [150]. This resulted in a change to the agent's pathogenicity, making it more infectious to cattle (and cats). Cattle succumbing to BSE were then included in cattle feed, thereby amplifying the transmission and spreading the infection [150], [151]. Once this epidemiologic pattern had been determined, the feeding of meat and bonemeal to ruminants was banned. Since then, the incidence of BSE first plateaued and then fell [152], [153].
The agent responsible for FSE is believed to be the same as for BSE [146]. It probably entered the feline population of the United Kingdom in contaminated pet food, and the temporal distribution of cases supports this hypothesis. Since its recognition in 1990, approximately 90 cases of FSE have been confirmed, mostly between 1990 and 1994 (J.W. Wilesmith, BVSc, PhD, J. Spriopoulos, DVM, PhD, Veterinary Laboratories Agency, Weybridge, UK, personal communication on confirmed cases up to the end of 2001, 2004) ( Fig. 1). In addition, retrospective study of brain tissue from cats with neurologic disease failed to find cases of FSE before 1990 [1], [154]. Although, like BSE, the peak of FSE seems to have passed, occasional cases are still seen (see Fig. 1) [153]. Because few domestic cats are subject to routine postmortem examination, it is likely that the total number of FSE cases has been underestimated.
Fig. 1.
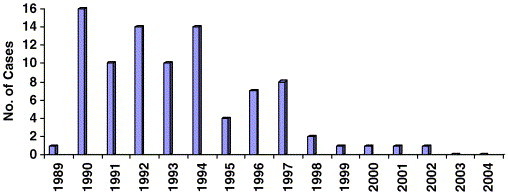
Graph showing the incidence of feline spongiform encephalopathy cases in cats by year of onset of clinical signs (cases confirmed to mid 2004; J.W. Wilesmith, BVSc, PhD, J. Spriopoulos, DVM, PhD, Veterinary Laboratories Agency, Weybridge, UK, personal communication).
Most TSEs, like FSE and BSE, seem to be transmitted by ingestion. Although maternal and even genetic transmission may occur in some species [135], [143], [155], there is no evidence of it occurring in cats or cattle [150], [152].
Pathogenesis
The TSE agents are unlike any other microorganisms. All TSE diseases are characterized by the accumulation of an abnormal isoform of a host-coded protein, the prion protein (PrP). PrP is found in all animals; it is a cell surface glycoprotein of unknown significance. Although the PrP isolated from normal individuals (PrPc) and the PrP isolated from TSE-infected individuals (PrP-res) have the same amino acid sequence and secondary structure, PrPc is totally degraded by proteinase K, whereas PrP-res resists digestion. Once present, PrP-res is believed to induce additional copies of itself by interacting with normal PrPc. In doing this, PrP-res acts as an infectious agent [156]. Once the host-coded PrPc has been transformed to PrP-res, it accumulates in fibrils (scrapie-associated fibrils [SAFs]), and this eventually leads to disease [135]. Because the process is slow, however, all TSEs have prolonged incubation periods. More detailed information on TSE pathogenesis is reviewed elsewhere [157], [158], [159]. Because PrP is host-coded, the accumulation of SAFs induces no immune response [160], [161].
Clinical signs
FSE shows no breed predisposition, and cats from all types of households have been affected. There seems to be a slight male predisposition [154]. The mean age at onset is approximately 5 to 7 years (range: 2–12 years) [132], [133], [134], [148].
FSE is characterized by progressive behavioral and motor disturbances. Affected cats present with progressive hind limb ataxia; increased aggression or affection; hyperesthesia to touch, sound, or light; altered grooming patterns; increased salivation; dilated pupils with an unusual staring expression; polyphagia or polydipsia; abnormal head posture; muscle fasciculations; or an inability to retract their claws [132], [133], [134], [148], [154]. Behavioral changes have usually been noted first, followed by progressive locomotor dysfunction. The cats tend to show ataxia, with dysmetria or hypermetria, which often leads to an erratic crouching gait [134], [148]. They also show an inability to judge distances. The disease is generally progressive, warranting euthanasia within 8 to 12 weeks of the onset of clinical signs [134].
Diagnosis
Premortem diagnosis is rarely possible. Although clinical signs may be suggestive of FSE, and nonspecific tests, such as electroencephalography (EEG) or MRI, may indicate the presence of diffuse CNS disease, specific tests are currently lacking. Significant abnormalities have not been detected on serum biochemistry, hematology, or CSF analysis [154]. Diagnosis of FSE is usually made by histopathologic examination of the brain (formalin-fixed tissue) and ultrastructural detection of SAFs in brain extracts (fresh-frozen brain or spinal cord) [162]. After euthanasia, any animal suspected of having FSE should have a full postmortem examination, which should be performed by a trained veterinary pathologist.
Pathologic changes are confined to the CNS and consist of variable degrees of neurophil vacuolation, vacuolation of the neuronal parenchyma, and an astrocytic response. Changes are particularly evident in the gray matter of the thalamus, basal ganglia, and cerebral and cerebellar cortices. More advanced cases may show neuronal loss and more striking gliosis. There are no inflammatory changes. Fibrils analogous to SAFs can be seen on electron microscopy [134], [163], [164].
Treatment
There is no effective treatment for FSE.
Zoonotic risk
Although it is generally difficult to transmit a TSE agent from one species to another by mouth, BSE seems to have been transmitted naturally, not only to cats, captive exotic felids, and captive exotic ungulates but to human beings, in the form of vCJD [137], [165]. Thankfully, with the introduction of strict laws regulating the slaughter and rendering of ruminants and the overall decline in the incidence of BSE, the possibility of the BSE agent continuing to be included in the food chain is extremely small. Because the incubation period is long and variable, however, we are likely to continue to see new cases of vCJD in the United Kingdom for a few years yet to come. That said, the increasing prevalence of CWD in deer and elk in the United States is of concern, and the potential for this TSE to be transmitted to human beings (or cats) is still to be determined [141].
It is unlikely that cats present a zoonotic risk. This is because the disease is now extremely rare (it was never common) and the likelihood of FSE-infected brain or spinal cord entering the human food chain is almost nonexistent. Although there has been one case of CJD and FSE occurring within the same household, the strain of TSE with which both individuals were affected seems to have been a variant more typically associated with spontaneous CJD rather than with BSE [166], and even that diagnosis has been questioned. The method of transmission in this case is not known.
Footnotes
The author received funding support for her lectureship from Nestlé Purina Petcare.
References
- 1.Bradshaw J.M., Pearson G.R., Gruffydd-Jones T.J. A retrospective study of 286 cases of neurological disorders of the cat. J Comp Pathol. 2004;131:112–120. doi: 10.1016/j.jcpa.2004.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hopkins A. Feline neurology. Part 1. Intracranial disorders. In Pract. 1992;14:59–65. [Google Scholar]
- 3.Summers B.A., Cummings J.F., deLahunta A. Mosby-Year Book; St. Louis: 1995. Veterinary neuropathology. [Google Scholar]
- 4.Rand J.S., Parent J., Percy D. Clinical, cerebrospinal fluid, and histological data from twenty-seven cats with primary inflammatory disease of the central nervous system. Can Vet J. 1994;35:103–110. [PMC free article] [PubMed] [Google Scholar]
- 5.Quesnel A.D., Parent J.M., McDonell W. Diagnostic evaluation of cats with seizure disorders: 30 cases(1991–1993) J Am Vet Med Assoc. 1997;210:65–71. [PubMed] [Google Scholar]
- 6.Munana K.R. Inflammatory disorders of the central nervous system. In: August J.R., editor. vol. 4. WB Saunders; Philadelphia: 2001. pp. 425–433. (Consultations in feline internal medicine). [Google Scholar]
- 7.Schatzberg S.J., Haley N.J., Barr S.C. Use of a multiplex polymerase chain reaction assay in the antemortem diagnosis of toxoplasmosis and Neospora in the central nervous system of cats and dogs. Am J Vet Res. 2003;64(12):1507–1513. doi: 10.2460/ajvr.2003.64.1507. [DOI] [PubMed] [Google Scholar]
- 8.Cook L.B., Bergman R.L., Bahr A. Inflammatory polyp in the middle ear with secondary suppurative meningoencephalitis in a cat. Vet Radiol Ultrasound. 2003;44(6):648–651. doi: 10.1111/j.1740-8261.2003.tb00525.x. [DOI] [PubMed] [Google Scholar]
- 9.Cleaveland S., Appel M.G.J., Chalmers W.S.K. Serological and demographic evidence for domestic dogs as a source of canine distemper virus infection for Serengeti wildlife. Vet Microbiol. 2000;72:217–227. doi: 10.1016/s0378-1135(99)00207-2. [DOI] [PubMed] [Google Scholar]
- 10.Lundgren A.L. Feline non-suppurative meningoencephalomyelitis. A clinical and pathological study. J Comp Pathol. 1992;107:411–425. doi: 10.1016/0021-9975(92)90015-M. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Guan Y., Zheng B.J., He Y.Q. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science. 2003;302(5643):276–278. doi: 10.1126/science.1087139. [DOI] [PubMed] [Google Scholar]
- 12.Martina B.E., Haagmans B.L., Kuiken T. Virology: SARS virus infection of cats and ferrets. Nature. 2003;425:915. doi: 10.1038/425915a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kuiken T., Rimmelzwaan G., Van Riel D. Avian H5N1 influenza in cats. Science. 2004;306:241. doi: 10.1126/science.1102287. [DOI] [PubMed] [Google Scholar]
- 14.Lanciotti R.S., Roehrig J.T., Deubel V. Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science. 1999;286(5448):2333–2337. doi: 10.1126/science.286.5448.2333. [DOI] [PubMed] [Google Scholar]
- 15.Austgen L.E., Bowen R.A., Bunning M.L. Experimental infection of cat and dogs with West Nile virus. Emerg Infect Dis. 2004;10(1):82–86. doi: 10.3201/eid1001.020616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lichtensteiger C.A., Heinz-Taheny K., Osborne T.S. West Nile virus encephalitis and myocarditis in wolf and dog. Emerg Infect Dis. 2003;9(10):1303–1306. doi: 10.3201/eid0910.020617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Komar N., Panella N.A., Boyce E. Exposure of domestic mammals to West Nile virus during an outbreak of human encephalitis, New York City, 1999. Emerg Infect Dis. 2001;7(4):736–738. doi: 10.3201/eid0704.010424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Woolhouse M.E.J. Population biology of emerging and re-emerging pathogens. Trends Microbiol. 2002;10(Suppl 10):s3–s7. doi: 10.1016/s0966-842x(02)02428-9. [DOI] [PubMed] [Google Scholar]
- 19.Luttgen P.J. Inflammatory disease of the central nervous system. Vet Clin N Am Small Anim Pract. 1988;18(3):623–640. doi: 10.1016/S0195-5616(88)50059-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Chrisman C.L., editor. Problems in small animal neurology. 2nd edition. Lea & Febiger; Philadelphia: 1991. [Google Scholar]
- 21.Skerritt G.C. Brain disorders in dogs and cats. In: Wheeler S.J., editor. Manual of small animal neurology. BSAVA; Cheltenham, UK: 1992. pp. 127–139. [Google Scholar]
- 22.Parent J.M., Quesnel A.D. Seizures in cats. Vet Clin N Am Small Anim Pract. 1996;26(4):811–825. [PubMed] [Google Scholar]
- 23.Munana K.R. Encephalitis and meningitis. Vet Clin N Am Small Anim Pract. 1996;26(4):857–876. [PubMed] [Google Scholar]
- 24.Greene C.E., editor. Infectious diseases of the dog and cat. 2nd edition. WB Saunders; Philadelphia: 1998. [Google Scholar]
- 25.Foley J.E., Leutenegger C. A review of coronavirus infection in the central nervous system of cats and mice. J Vet Intern Med. 2001;15:438–444. doi: 10.1111/j.1939-1676.2001.tb01572.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Foley J.E., Rand C., Leutenegger C. Inflammation and changes in cytokine levels in neurological feline infectious peritonitis. J Feline Med Surg. 2003;5(6):313–322. doi: 10.1016/S1098-612X(03)00048-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pedersen N.C., Yamamoto J.K., Ishida T. Feline immunodeficiency virus infection. Vet Immunol Immunopathol. 1989;21:111–129. doi: 10.1016/0165-2427(89)90134-7. [DOI] [PubMed] [Google Scholar]
- 28.Dow S.W., Poss M.L., Hoover E.A. Feline immunodeficiency virus: a neurotropic lentivirus. J Acquir Immune Defic Hum Retrovirol. 1990;3:658–668. [PubMed] [Google Scholar]
- 29.Dow S.W., Dreitz M.J., Hoover E.A. Exploring the link between feline immunodeficiency virus infection and neurologic disease in cats. Vet Med. 1992;87:1181–1184. [Google Scholar]
- 30.Hurtrel M., Ganiere J.P., Guelfi J.F. Comparison of early and late feline immunodeficiency virus encephalopathies. AIDS. 1992;6:399–406. doi: 10.1097/00002030-199204000-00007. [DOI] [PubMed] [Google Scholar]
- 31.Henriksen S.J., Prospero-Garcia O., Phillips T.R. Feline immunodeficiency virus as a model for study of lentivirus infection of the central nervous system. Curr Top Microbiol Immunol. 1995;202:167–186. doi: 10.1007/978-3-642-79657-9_12. [DOI] [PubMed] [Google Scholar]
- 32.Gunn-Moore D., Pearson G., Harbour D. Encephalitis with giant cells in a cat with naturally occurring feline immunodeficiency virus infection demonstrated by in situ hybridization. Vet Pathol. 1996;33:699–703. doi: 10.1177/030098589603300610. [DOI] [PubMed] [Google Scholar]
- 33.Hopper C.D., Sparkes A.H., Gruffydd-Jones T.J. Clinical and laboratory findings in cats infected with feline immunodeficiency virus. Vet Rec. 1989;125:341–346. doi: 10.1136/vr.125.13.341. [DOI] [PubMed] [Google Scholar]
- 34.Karpas A., Routledge J.K. Feline herpes virus: isolation and experimental studies. Zentralbl Veterinaermed. 1968;15:599–606. doi: 10.1111/j.1439-0450.1968.tb00333.x. [DOI] [PubMed] [Google Scholar]
- 35.Love D.N., Baker K.D. Sudden death in a kitten associated with a feline Picorna virus. Aust Vet J. 1972;48:643. doi: 10.1111/j.1751-0813.1972.tb05105.x. [DOI] [PubMed] [Google Scholar]
- 36.Bouljihad M., Lindeman C.J., Hayden D.W. Pyogranulomatous meningoencephalitis associated with dematiaceous fungal (Cladophialophora bantiana) infection in a domestic cat. J Vet Diagn Invest. 2002;14(1):70–72. doi: 10.1177/104063870201400116. [DOI] [PubMed] [Google Scholar]
- 37.Dubey J.P., Benson J., Larson M.A. Clinical Sarcocystis neurona encephalomyelitis in a domestic cat following routine surgery. Vet Parasitol. 2003;112(4):261–267. doi: 10.1016/s0304-4017(03)00019-0. [DOI] [PubMed] [Google Scholar]
- 38.Kordick D.L., Breitschwerdt E.B. Relapsing bacteremia after blood transmission of Bartonella henselae to cats. Am J Vet Res. 1997;58(5):492–497. [PubMed] [Google Scholar]
- 39.Marra C.M. Neurologic complications of Bartonella henselae infection. Curr Opin Neurol. 1995;8(3):164–169. doi: 10.1097/00019052-199506000-00002. [DOI] [PubMed] [Google Scholar]
- 40.Yanai T., Tujioka S., Sakai H. Experimental infection with equine herpesvirus 9 (EHV-9) in cats. J Comp Pathol. 2003;128(2–3):113–118. doi: 10.1053/jcpa.2002.0613. [DOI] [PubMed] [Google Scholar]
- 41.Luttrell C.N., Bang F.B. Newcastle disease encephalomyelitis in cats. I. Clinical and pathological features. Arch Neurol Psychiatr. 1958;79:647–657. doi: 10.1001/archneurpsyc.1958.02340060043005. [DOI] [PubMed] [Google Scholar]
- 42.Salvioli G., Gotti D., Sternini G. Effetti polimorfi nel gatto da inoculazione di materiale poliomielitico. Riv Ist Sieroter Ital. 1952;27:225–245. [PubMed] [Google Scholar]
- 43.Martin W.J., Glass R.T. Acute encephalopathy induced in cats with a stealth virus isolated from a patient with chronic fatigue syndrome. Pathobiology. 1995;63(3):115–118. doi: 10.1159/000163942. [DOI] [PubMed] [Google Scholar]
- 44.Hoff E.J., Vandevelde M. Non-suppurative encephalomyelitis in cats suggestive of a viral origin. Vet Pathol. 1981;18:170–180. doi: 10.1177/030098588101800204. [DOI] [PubMed] [Google Scholar]
- 45.Wilson M.S., Wherrett B.A., Mahdy M.S. Powassan virus meningoencephalitis: a case report. Can Med Assoc J. 1979;121(3):320–323. [PMC free article] [PubMed] [Google Scholar]
- 46.Keane D.P., Parent J., Little P.B. Californian subgroup: Powassan virus infection of cats. Can J Microbiol. 1987;33:693–697. doi: 10.1139/m87-121. [DOI] [PubMed] [Google Scholar]
- 47.Greene C.E., Baldwin C.A. Arboviral infections. In: Greene C.E., editor. Infectious diseases of the dog and cat. 2nd edition. WB Saunders; Philadelphia: 1998. pp. 131–133. [Google Scholar]
- 48.Daubney R. Viral encephalitis of equines and domestic ruminants in the Near East. Part II. Res Vet Sci. 1967;8:419–439. [PubMed] [Google Scholar]
- 49.Daubney R., Mahlau E.A. Viral encephalitis of equines and domestic ruminants in the Near East. Part I. Res Vet Sci. 1967;8:375–397. [PubMed] [Google Scholar]
- 50.Artsob H., Spence L., Th'ng C. Arbovirus infections in several Ontario mammals, 1975–1980. Can J Vet Res. 1986;50(1):42–46. [PMC free article] [PubMed] [Google Scholar]
- 51.Nicolau S., Galloway I.A. Borna disease and enzootic encephalomyelitis of sheep and cattle. Special Reports Series. Medical Research Council. 1928;121:7–90. [Google Scholar]
- 52.Ludwig H., Bode L., Gosztonyi G. Borna disease: a persistent virus infection of the central nervous system. Prog Med Virol. 1988;35:107–151. [PubMed] [Google Scholar]
- 53.Lundgren A.L., Czech G., Bode L., Ludwig H. Natural Borna disease in domestic animals other than horses and sheep. J Vet Med. 1993;B40:298–303. doi: 10.1111/j.1439-0450.1993.tb00142.x. [DOI] [PubMed] [Google Scholar]
- 54.Lundgren A.L., Zimmermann W., Bode L. Staggering disease in cats: isolation and characterization of the feline Borna virus. J Gen Virol. 1995;76(9):2215–2222. doi: 10.1099/0022-1317-76-9-2215. [DOI] [PubMed] [Google Scholar]
- 55.Malkinson M., Weisman Y., Ashash E. Borna disease in ostriches. Vet Rec. 1993;133:304. doi: 10.1136/vr.133.12.304-b. [DOI] [PubMed] [Google Scholar]
- 56.Malkinson M., Weisman Y., Perl S. A Borna-like disease of ostriches in Israel. Curr Top Microbiol Immunol. 1995;190:31–38. doi: 10.1007/978-3-642-78618-1_3. [DOI] [PubMed] [Google Scholar]
- 57.Bode L., Durrwald R., Ludwig H. Borna virus infections in cattle associated with fatal neurological disease. Vet Rec. 1994;135:283–284. doi: 10.1136/vr.135.12.283. [DOI] [PubMed] [Google Scholar]
- 58.Rott R., Becht H. Natural and experimental Borna disease in animals. Curr Top Microbiol Immunol. 1995;190:15–30. doi: 10.1007/978-3-642-78618-1_2. [DOI] [PubMed] [Google Scholar]
- 59.Weissenbock H., Nowotny N., Caplazi P. Borna disease in a dog with lethal meningoencephalitis. J Clin Microbiol. 1998;36:2127–2130. doi: 10.1128/jcm.36.7.2127-2130.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Narayan O., Herzog S., Frese K. Pathogenesis of Borna disease in rats: immune-mediated viral ophthalmoencephalopathy causing blindness and behavioural abnormalities. J Infect Dis. 1983;148:305–315. doi: 10.1093/infdis/148.2.305. [DOI] [PubMed] [Google Scholar]
- 61.Hatalski C.G., Lewis A.J., Lipkin W.I. Borna disease. Emerg Infect Dis. 1997;3:129–135. doi: 10.3201/eid0302.970205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Nakamura Y., Asahi S., Nakaya T. Demonstration of Borna disease virus RNA in peripheral blood mononuclear cells derived from domestic cats in Japan. J Clin Microbiol. 1996;34:188–198. doi: 10.1128/jcm.34.1.188-191.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Richt J.A., Pfeuffer I., Christ M. Borna disease virus infection in animals and humans. Emerg Infect Dis. 1997;3:343–352. doi: 10.3201/eid0303.970311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Takahashi H., Nakaya T., Nakamura Y. Higher prevalence of Borna disease virus infection in blood donors living near thoroughbred horse farms. J Med Virol. 1997;52:330–335. [PubMed] [Google Scholar]
- 65.Reeves N.A., Helps C.R., Gunn-Moore D.A. Natural Borna disease virus infection in cats in the United Kingdom. Vet Rec. 1998;143:523–526. doi: 10.1136/vr.143.19.523. [DOI] [PubMed] [Google Scholar]
- 66.Weissenbock H., Suchy A., Caplazi P. Borna disease in Austrian horses. Vet Rec. 1998;143:21–22. doi: 10.1136/vr.143.1.21. [DOI] [PubMed] [Google Scholar]
- 67.Cotto E., Neau D., Cransac-Neau M. Borna disease virus RNA in immunocompromised patients in southwestern France. J Clin Microbiol. 2003;41(12):5577–5581. doi: 10.1128/JCM.41.12.5577-5581.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Helps C.R., Turan N., Bilal T. Detection of antibodies to Borna disease virus in Turkish cats by using recombinant p40. Vet Rec. 2001;149:647–650. doi: 10.1136/vr.149.21.647. [DOI] [PubMed] [Google Scholar]
- 69.Dauphin G., Legay V., Pitel P.-H. Borna disease: current knowledge and virus detection in France. Vet Res. 2002;33:127–138. doi: 10.1051/vetres:2002002. [DOI] [PubMed] [Google Scholar]
- 70.Terayama H., Nishino Y., Kishi M. Detection of anti-Borna disease virus (BDV) antibodies from patients with schizophrenia and mood disorders in Japan. Psychiatry Res. 2003;120(2):201–206. doi: 10.1016/s0165-1781(03)00190-2. [DOI] [PubMed] [Google Scholar]
- 71.Kronevi T., Nordstrom M., Moreno W. Feline ataxia due to non-suppurative meningoencephalomyelitis of unknown aetiology. Nord Veterinmed. 1974;26:720–725. [PubMed] [Google Scholar]
- 72.Lundgren A.L., Johannisson A., Zimmermann W. Neurological disease and encephalitis in cats experimentally infected with Borna virus. Acta Neuropathol (Berl) 1997;93:391–401. doi: 10.1007/s004010050630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Ludwig H., Bode L. The neuropathogenesis of Borna disease virus infections. Intervirology. 1997;40:185–197. doi: 10.1159/000150545. [DOI] [PubMed] [Google Scholar]
- 74.Huebner J., Bode L., Ludwig H. Borna disease virus infection in FIV-positive cats in Germany. Vet Rec. 2001;149:152. doi: 10.1136/vr.149.5.152. [DOI] [PubMed] [Google Scholar]
- 75.Nishino Y., Funaba M., Fukushima R. Borna disease virus infection in domestic cats: evaluation by RNA and antibody detection. J Vet Med Sci. 1999;61(10):1167–1170. doi: 10.1292/jvms.61.1167. [DOI] [PubMed] [Google Scholar]
- 76.Lundgren A.L., Ludwig H. Clinically diseased cats with non-suppurative meningoencephalomyelitis have Borna disease virus-specific antibodies. Acta Vet Scand. 1993;34:101–103. doi: 10.1186/BF03548230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Nakamura Y., Watanabe M., Kamitani W. High prevalence of Borna disease virus in domestic cats with neurological disorders in Japan. Vet Microbiol. 1999;70(3–4):153–169. doi: 10.1016/s0378-1135(99)00135-2. [DOI] [PubMed] [Google Scholar]
- 78.Strom B., Andren B., Lundgren A.L. Idiopathic non-suppurative meningoencephalo-myelitis (Staggering disease) in the Swedish cat: a study of 33 cases. Svensk Veterinar Tindning. 1992;44(1):19–24. (translation published in the European Journal of Companion Animal Practice 1992;3(1):9–13) [Google Scholar]
- 79.Nowotny N., Weissenbock H. Description of feline nonsuppurative meningoencephalo-myelitis (“Staggering Disease”) and studies of its etiology. J Clin Microbiol. 1995;33:1668–1669. doi: 10.1128/jcm.33.6.1668-1669.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Berg A.L., Reid-Smith R., Larsson M. Case control study of feline Borna disease in Sweden. Vet Rec. 1998;142:715–717. doi: 10.1136/vr.142.26.715. [DOI] [PubMed] [Google Scholar]
- 81.Staeheli P., Sauder C., Hausmann J. Epidemiology of Borna disease virus. J Gen Virol. 2000;81:2123–2135. doi: 10.1099/0022-1317-81-9-2123. [DOI] [PubMed] [Google Scholar]
- 82.Berg M., Johansson M., Montell H. Wild birds as possible natural reservoirs of Borna disease virus. Epidemiol Infect. 2001;127:173–178. doi: 10.1017/s0950268801005702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Richt J.A., Herzog S., Haberzettl K., Rott R. Demonstration of Borna disease virus-specific RNA in secretions of naturally infected horses by the polymerase chain reaction. Med Microbiol Immunol (Berl) 1993;182:293–304. doi: 10.1007/BF00191945. [DOI] [PubMed] [Google Scholar]
- 84.Binz T., Lebelt J., Niemann H. Sequence analysis of the p24 gene of Borna disease virus in naturally infected horse, donkey and sheep. Vet Res. 1994;34:281–289. doi: 10.1016/0168-1702(94)90128-7. [DOI] [PubMed] [Google Scholar]
- 85.Schneider P.A., Briese T., Zimmermann W. Sequence conservation in field and experimental isolates of Borna disease virus. J Virol. 1994;68:63–68. doi: 10.1128/jvi.68.1.63-68.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Briese T., Lipkin W.I., de la Torre J.C. Molecular biology of Borna disease virus. Curr Top Microbiol Immunol. 1995;190:1–16. doi: 10.1007/978-3-642-78618-1_1. [DOI] [PubMed] [Google Scholar]
- 87.Cubitt B., de la Torre J.C. Borna disease virus (BDV), a nonsegmented RNA virus, replicates in the nucleus of infected cells where infectious BDV ribonucleoproteins are present. J Virol. 1994;68:1371–1381. doi: 10.1128/jvi.68.3.1371-1381.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Gonzalez-Dunia D., Sauder C., de la Torre J.C. Borna disease virus and the brain. Brain Res Bull. 1997;44(6):647–664. doi: 10.1016/S0361-9230(97)00276-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Bode L., Ludwig H. Clinical similarities and close genetic relationship of human and animal Borna disease virus. Arch Virol. 1997;(Suppl 13):167–182. doi: 10.1007/978-3-7091-6534-8_16. [DOI] [PubMed] [Google Scholar]
- 90.Cubitt B., Oldstone C., de la Torre J.C. Sequence and genome organisation of Borna disease virus. J Virol. 1994;68:1382–1396. doi: 10.1128/jvi.68.3.1382-1396.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Stitz L., Dietzschold B., Carbone K.M. Immunopathogenesis of Borna disease. Curr Top Microbiol Virol. 1995;190:75–92. doi: 10.1007/978-3-642-78618-1_5. [DOI] [PubMed] [Google Scholar]
- 92.Narayan O., Herzog S., Frese K. Behavioral disease in rats caused by immunopathological responses to persistent Borna virus in the brain. Science. 1983;220:1401–1403. doi: 10.1126/science.6602380. [DOI] [PubMed] [Google Scholar]
- 93.Berg A.L., Johansson A., Johansson M. Peripheral and intracerebral T cell immune response in cats naturally infected with Borna disease virus. Vet Immunol Immunopathol. 1999;68(2–4):241–253. doi: 10.1016/s0165-2427(99)00030-6. [DOI] [PubMed] [Google Scholar]
- 94.Weissenböck H., Nowotny N., Zoher J. Feline meningoencephalo-myelitis (“Staggering Disease”) Osterr Wien Tierarztl Mschr. 1994;81:195–201. [Google Scholar]
- 95.Berg A.L., Berg M. A variant form of feline Borna disease. J Comp Pathol. 1998;119:323–331. doi: 10.1016/s0021-9975(98)80054-6. [DOI] [PubMed] [Google Scholar]
- 96.Lundgren A.L., Lindberg R., Ludwig H. Immunoreactivity of the central nervous system in cats with a Borna disease-like meningoencephalomyelitis (staggering disease) Acta Neuropathol (Berl) 1995;90:184–193. doi: 10.1007/BF00294319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Lundgren A.L. Borna disease in cats. Vet Rec. 1999;145:87. [PubMed] [Google Scholar]
- 98.Bode L., Riegel S., Ludwig H. Borna disease virus-specific antibodies in patients with HIV and with mental disorders. Lancet. 1988;2:689. doi: 10.1016/s0140-6736(88)90505-3. [DOI] [PubMed] [Google Scholar]
- 99.Bode L., Zimmermann W., Ferszt R. Borna disease virus genome transcribed and expressed in psychiatric patients. Nat Med. 1995;1:232–236. doi: 10.1038/nm0395-232. [DOI] [PubMed] [Google Scholar]
- 100.Waltrip R.W., Buchanan R.W., Carpenter W.T. Borna disease antibodies and the deficit syndrome of schizophrenia. Schizophr Res. 1997;23:253–257. doi: 10.1016/s0920-9964(96)00114-4. [DOI] [PubMed] [Google Scholar]
- 101.Bode L., Ludwig H. Borna disease virus infection, a human mental-health risk. Clin Microbiol Rev. 2003;16(3):534–545. doi: 10.1128/CMR.16.3.534-545.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Rott R., Herzog S., Fleischer B. Detection of serum antibodies to Borna disease virus in patients with psychiatric disorders. Science. 1985;228:755–756. doi: 10.1126/science.3922055. [DOI] [PubMed] [Google Scholar]
- 103.Bode L., Riegel S., Lange W. Human infection with Borna disease virus: seroprevalence in patients with chronic diseases and healthy individuals. J Med Virol. 1992;36:309–315. doi: 10.1002/jmv.1890360414. [DOI] [PubMed] [Google Scholar]
- 104.Bode L., Durrwald R., Rantam F.A. First isolates of infectious human Borna disease virus from patients with mood disorders. Mol Psychiatry. 1996;1:200–212. [PubMed] [Google Scholar]
- 105.Salvatore M., Morzunov S., Schwemmle M. Borna disease virus in brains of North American and European people with schizophrenia and bipolar disorder. Lancet. 1997;349:1813–1814. doi: 10.1016/s0140-6736(05)61693-5. [DOI] [PubMed] [Google Scholar]
- 106.Iwahashi K., Watanabe M., Nakamura K. Clinical investigation of the relationship between Borna disease virus (BDV) infection and schizophrenia in 67 patients in Japan. Acta Psychiatr Scand. 1997;96(6):412–415. doi: 10.1111/j.1600-0447.1997.tb09941.x. [DOI] [PubMed] [Google Scholar]
- 107.Haga S., Yoshimura M., Motoi Y. Detection of Borna disease virus genome in normal human brain tissue. Brain Res. 1997;770:307–309. doi: 10.1016/s0006-8993(97)00903-7. [DOI] [PubMed] [Google Scholar]
- 108.Borland R., McDonald N. Feline encephalomyelitis. Br Vet J. 1965;121:479–483. doi: 10.1016/s0007-1935(17)40956-0. [DOI] [PubMed] [Google Scholar]
- 109.Vandevelde M., Braund K.G. Polioencephalomyelitis in cats. Vet Pathol. 1979;16:420–427. doi: 10.1177/030098587901600404. [DOI] [PubMed] [Google Scholar]
- 110.Kronevi T., Nordström M., Moreno W. Feline ataxia due to nonsuppurative meningoencephalomyelitis of unknown aetiology. Nord Vet Med. 1974;26:720–725. [PubMed] [Google Scholar]
- 111.Bleby J., Brierley J.B. An idiopathic demyelination in SPF cats. Vet Rec. 1972;91(Suppl):10. [abstract] [Google Scholar]
- 112.Palmer A.C., Cavanagh J.B. Encephalomyelopathy in young cats. J Soc Adm Pharm. 1995;36:57–64. doi: 10.1111/j.1748-5827.1995.tb02822.x. [DOI] [PubMed] [Google Scholar]
- 113.Martin L.A., Hintermann J. Une malade non décrite du chat; la myélite infectieuse. Bull de Arch Inst Pasteur Maroc. 1955;5:64–73. [Google Scholar]
- 114.McGaughey C.A. Infectious myelitis of cats: preliminary communication. Ceylon Veterinary Journal. 1953;1:34–40. [Google Scholar]
- 115.Flir K. Encephalomyelitis bei Grosskatzen. Dtsch Tierarztl Wochenschr. 1973;80:401–404. [PubMed] [Google Scholar]
- 116.Melchior G. Meningo-enzephalitis beim löwen and tiger. In: Ippen, Schröder, editors. Erkrankungen der Zootiere, Verhandlungsbericht des XV. Internationalen Symposiums uber die Erkrankungen der Zootiere. Akademie-Verlang; Berlin: 1973. pp. 245–254. [Google Scholar]
- 117.Gutter A, Wells S, Baskin G. Neurological disease in three cats at the Audubon Park Zoo. In: Proceedings of the Annual Meeting of the American Association of Zoo Veterinarians. Tampa; 1983. p. 21–6.
- 118.Truyen U., Stockhofe-Zurwieden N., Kaaden O.R. A case report: encephalitis in lions. Pathological and virological findings. Dtsch Tierarztl Wochenschr. 1990;97:89–91. [PubMed] [Google Scholar]
- 119.Vandevelde M. Neurologic diseases of suspected infectious origin. In: Greene C.D., editor. Infectious diseases of the dog and cat. WB Saunders; Philadelphia: 1998. pp. 530–539. [Google Scholar]
- 120.Greenfield J.G. 3rd edition. Edward Arnold; London: 1976. Neuropathology. p. 306–8. [Google Scholar]
- 121.Innes J.R.M., Saunders L.Z. Academic Press; New York: 1962. Comparative neuropathology. p. 361–4. [Google Scholar]
- 122.Kilham L., Margolis G. Viral etiology of spontaneous ataxia in cats. Am J Pathol. 1966;48:991–1011. [PMC free article] [PubMed] [Google Scholar]
- 123.Csiza C.K., deLahunta A., Scott F.W. Spontaneous feline ataxia. Cornell Vet. 1972;62:300–322. [PubMed] [Google Scholar]
- 124.Csiza C.K., Scott F.W., deLahunta A. Respiratory signs and central nervous system lesions in cats infected with panleukopenia virus. A case report. Cornell Vet. 1972;62:192–195. [PubMed] [Google Scholar]
- 125.Carlson J.H. Feline panleukopenia. In: Kirk R.W., editor. vol. VI. WB Saunders; Philadelphia: 1977. pp. 1292–1296. (Current veterinary therapy). [Google Scholar]
- 126.Mesfin G.M., Kusewitt D., Parker A. Degenerative myelopathy in a cat. J Am Vet Med Assoc. 1980;176:62–64. [PubMed] [Google Scholar]
- 127.Bollo E., Zurbriggen A., Vandevelde M. Canine distemper virus clearance in chronic inflammatory demyelination. Acta Neuropathol (Berl) 1986;72(1):69–73. doi: 10.1007/BF00687949. [DOI] [PubMed] [Google Scholar]
- 128.Shope T.C. Chickenpox encephalitis and encephalopathy: evidence for differing pathogenesis. Yale J Biol Med. 1982;55(3–4):321–327. [PMC free article] [PubMed] [Google Scholar]
- 129.Gray F., Mohr M., Rozenberg F. Varicella-zoster virus encephalitis in acquired immunodeficiency syndrome: report of four cases. Neuropathol Appl Neurobiol. 1992;18(5):502–514. doi: 10.1111/j.1365-2990.1992.tb00817.x. [DOI] [PubMed] [Google Scholar]
- 130.Dittrich W., Bode L., Ludwig H. Learning deficiencies in Borna disease virus-infected but clinically healthy rats. Biol Psychiatry. 1989;26:818–828. doi: 10.1016/0006-3223(89)90122-4. [DOI] [PubMed] [Google Scholar]
- 131.Bautista J.R., Schwartz G.J., de la Torre J.C. Early and persistent abnormalities in rats with neonatally acquired Borna virus infection. Brain Res Bull. 1994;34:31–40. doi: 10.1016/0361-9230(94)90183-x. [DOI] [PubMed] [Google Scholar]
- 132.Leggett M.M., Dukes J., Pirie H.M. A spongiform encephalopathy in a cat. Vet Rec. 1990;127:586–588. [PubMed] [Google Scholar]
- 133.Wyatt J.M., Pearson G.R., Smerdon T.N. Spongiform encephalopathy in a cat. Vet Rec. 1990;126:513. [PubMed] [Google Scholar]
- 134.Wyatt J.M., Pearson G.R., Smerdon T.N. Naturally occurring scrapie-like spongiform encephalopathy in five domestic cats. Vet Rec. 1991;129:233–236. doi: 10.1136/vr.129.11.233. [DOI] [PubMed] [Google Scholar]
- 135.Kimberlin R.H. Transmissible spongiform encephalopathies in animals. Can J Vet Res. 1990;54:30–37. [PMC free article] [PubMed] [Google Scholar]
- 136.Wells G.A.H., McGill I.S. Recently described scrapie-like encephalopathies in animals: case descriptions. In: Bradley R., Savey M., Marchant B., editors. Sub-acute spongiform encephalopathies. vol. 55. Kluwer Academic Publishers; Dordrecht: 1991. pp. 11–24. (Current topics in veterinary medicine and animal science). [Google Scholar]
- 137.Kirkwood J.K., Cunningham A.A. Epidemiological observations on spongiform encephalopathies in captive wild animals in the British Isles. Vet Rec. 1994;135:296–303. doi: 10.1136/vr.135.13.296. [DOI] [PubMed] [Google Scholar]
- 138.Kirkwood J.K., Cunningham A.A., Flach E.J. Spongiform encephalopathy in another captive cheetah (Acinonyx jubatus): evidence for variation in susceptibility or incubation periods between species? J Zoo Wildl Med. 1995;24:577–582. [Google Scholar]
- 139.Baron T., Belli P., Madec J.Y. Spongiform encephalopathy in an imported cheetah in France. Vet Rec. 1997;141:270–271. doi: 10.1136/vr.141.11.270. [DOI] [PubMed] [Google Scholar]
- 140.Willoughby K., Kelly D.F., Lyon D.G. Spongiform encephalopathy in a captive puma (Felis concolor) Vet Rec. 1992;131:431–434. doi: 10.1136/vr.131.19.431. [DOI] [PubMed] [Google Scholar]
- 141.Miller M.W., Willians E.S., Hobbs N.T. Environmental sources of prion transmission in mule deer. Emerg Infect Dis. 2004;10(6):1003–1006. doi: 10.3201/eid1006.040010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Hartsough G.R., Burger D. Encephalopathy of mink I. Epizootiology and clinical observations. J Infect Dis. 1965;115:387–392. doi: 10.1093/infdis/115.4.387. [DOI] [PubMed] [Google Scholar]
- 143.Brown P. Transmissible spongiform encephalopathies in humans: kuru, Creutzfeldt-Jakob disease, Gerstmann-Sträussler-Scheinker disease. Can J Vet Res. 1990;54:38–41. [PMC free article] [PubMed] [Google Scholar]
- 144.Gibbs C.J., Gajdusek D.C. Experimental subacute spongiform virus encephalopathies in primates and other laboratory animals. Science. 1973;182:67–69. doi: 10.1126/science.182.4107.67. [DOI] [PubMed] [Google Scholar]
- 145.Baker H.F., Ridley R.M., Wells G.A.H. Experimental transmission of BSE and scrapie to the common marmoset. Vet Rec. 1993;132:403–406. doi: 10.1136/vr.132.16.403. [DOI] [PubMed] [Google Scholar]
- 146.Fraser H., Pearson G.R., McConnell I. Transmission of feline spongiform encephalopathy to mice. Vet Rec. 1994;134:449. doi: 10.1136/vr.134.17.449. [DOI] [PubMed] [Google Scholar]
- 147.Parry H.B. Scrapie disease in sheep. In: Oppenheimer D.R., editor. Academic Press; London: 1983. p. 31. [Google Scholar]
- 148.Bratberg B., Ueland K., Wells G.A.H. Feline spongiform encephalopathy in a cat in Norway. Vet Rec. 1995;136:444. doi: 10.1136/vr.136.17.444. [DOI] [PubMed] [Google Scholar]
- 149.Wells G.A.H., Scott A.C., Johnson C.T. A novel progressive spongiform encephalopathy in cattle. Vet Rec. 1987;121:419–420. doi: 10.1136/vr.121.18.419. [DOI] [PubMed] [Google Scholar]
- 150.Wilesmith J.W., Wells G.A.H., Cranwell M.P. Bovine spongiform encephalopathy: epidemiological studies. Vet Rec. 1988;123:638–644. [PubMed] [Google Scholar]
- 151.Wilesmith J.W., Ryan J.B., Atkinson M.J. Bovine spongiform encephalopathy: epidemiological studies on the origin. Vet Rec. 1991;128:199–203. doi: 10.1136/vr.128.9.199. [DOI] [PubMed] [Google Scholar]
- 152.Hoinville L.J. Decline in the incidence of BSE in cattle born after the introduction of the ‘feed ban’. Vet Rec. 1994;134:274–275. doi: 10.1136/vr.134.11.274. [DOI] [PubMed] [Google Scholar]
- 153.Wilesmith J.W., Ryan J.B. Bovine spongiform encephalopathy: observations on incidence during 1992. Vet Rec. 1993;132:300–301. doi: 10.1136/vr.132.12.300. [DOI] [PubMed] [Google Scholar]
- 154.Gruffydd-Jones T.J., Galloway P.E., Pearson G.R. Feline spongiform encephalopathy. J Soc Adm Pharm. 1991;33:471–476. [Google Scholar]
- 155.Donnelly C.A. Maternal transmission of BSE: interpretation of the data on the offspring of BSE-affected pedigree suckler cows. Vet Rec. 1998;142:579–580. doi: 10.1136/vr.142.21.579. [DOI] [PubMed] [Google Scholar]
- 156.Prusiner S.B. Genetic and infectious prion diseases. Arch Virol. 1993;50:1129–1153. doi: 10.1001/archneur.1993.00540110011002. [DOI] [PubMed] [Google Scholar]
- 157.Gunn-Moore D.A., Harbour D.A. Feline spongiform encephalopathy and Borna disease. In: August J.R., editor. vol. 4. WB Saunders; Philadelphia: 2001. pp. 62–70. (Consultations in feline internal medicine). [Google Scholar]
- 158.Schreuder B.E.C., van Keulen L.J.M., Vromans M.E.W. Tonsillar biopsy and PrPSc detection in the preclinical diagnosis of scrapie. Vet Rec. 1998;142:564–568. doi: 10.1136/vr.142.21.564. [DOI] [PubMed] [Google Scholar]
- 159.Hadlow W.J., Kennedy R.C., Race R.E. Natural infection of Suffolk sheep with scrapie virus. J Infect Dis. 1982;146:657–664. doi: 10.1093/infdis/146.5.657. [DOI] [PubMed] [Google Scholar]
- 160.Bolton D.C., McKinley M.P., Prusiner S.B. Identification of a protein that purifies with scrapie prion. Science. 1982;218:1309–1311. doi: 10.1126/science.6815801. [DOI] [PubMed] [Google Scholar]
- 161.Chesebro B., Race R., Wehrly K. Identification of scrapie prion protein-specific mRNA in scrapie-infected and uninfected brain. Nature. 1985;315:331–333. doi: 10.1038/315331a0. [DOI] [PubMed] [Google Scholar]
- 162.Gibson P.H., Somerville R.A., Fraser H. Scrapie associated fibrils in the diagnosis of scrapie in sheep. Vet Rec. 1987;120:125–127. doi: 10.1136/vr.120.6.125. [DOI] [PubMed] [Google Scholar]
- 163.Pearson G.R., Gruffydd-Jones T.J., Wyatt J.M. Feline spongiform encephalopathy. Vet Rec. 1991;128:532. doi: 10.1136/vr.128.22.532-b. [DOI] [PubMed] [Google Scholar]
- 164.Pearson G.R., Wyatt J.M., Gruffydd-Jones T.J. Feline spongiform encephalopathy: fibril and PrP studies. Vet Rec. 1992;131:307–310. doi: 10.1136/vr.131.14.307. [DOI] [PubMed] [Google Scholar]
- 165.Collinge J., Sidle K.C.L., Meads J. Molecular analysis of prion strain variation and the etiology of new variant CJD. Nature. 1996;383:685–690. doi: 10.1038/383685a0. [DOI] [PubMed] [Google Scholar]
- 166.Zanusso G., Nardelli E., Zosati A. Simultaneous occurrence of spongiform encephalopathy in a man and his cat in Italy. Lancet. 1998;352:1116–1117. doi: 10.1016/S0140-6736(05)79756-7. [DOI] [PubMed] [Google Scholar]
- 167.Fenner W.R. Central nervous system infections. In: Greene C.E., editor. Infectious diseases of the dog and cat. 2nd edition. WB Saunders; Philadelphia: 1998. pp. 647–657. [Google Scholar]