

Abstract
Rubella virus is a small enveloped virus that assembles in association with Golgi membranes. Freeze-substitution electron microscopy of rubella virus-infected cells revealed a previously unrecognized virion polymorphism inside the Golgi stacks: homogeneously dense particles without a defined core coexisting with less dense, mature virions that contained assembled cores. The homogeneous particles appear to be a precursor form during the virion morphogenesis process as the forms with mature morphology were the only ones detected inside secretory vesicles and on the exterior of cells. In mature virions potential remnants of C protein membrane insertion were visualized as dense strips connecting the envelope with the internal core. In infected cells Golgi stacks were frequently seen close to cytopathic vacuoles, structures identified as the sites for viral RNA replication, along with the rough endoplasmic reticulum and mitochondria. These associations could facilitate the transfer of viral genomes from the cytopathic vacuoles to the areas of rubella assembly in Golgi membranes.
Keywords: Rubella virus, Viral assembly, Golgi complex
Introduction
Enveloped viruses use a variety of cellular membranes to support different processes in their life cycles. Recent studies have shown that enveloped viruses that assemble intracellularly sometimes induce the formation of large complexes through association of organelles. These “factories” are the framework for viral replication and/or assembly and the intracellular signals involved in their formation are poorly characterized Lee et al 1996, Mckenzie et al 1999, Risco et al 2002, Salanueva et al 2003. One of the less understood features of these factories is how viral components, physically anchored in different (apparently distant) organelles, are efficiently transfered between the organelles. One example would be the transport of replicated genomes from the replication complexes to the virion assembly sites. The superior preservation of small cellular and viral structures after freeze-substitution has been the key factor for identifying viral intermediates and maturation events in the morphogenic pathway of several viruses Risco et al 2002, Salanueva et al 1999, Salanueva et al 2003. Coronaviruses and bunyaviruses assemble in association with membranes of the endoplasmic reticulum–Golgi intermediate compartment (ERGIC) and the Golgi complex, respectively. For both, a major structural transformation of the virion inside the Golgi complex has been reported in that immature precursor viral particles change their organization inside the Golgi stack to produce mature virions; this maturation process requires a functional trans-Golgi subcompartment Salanueva et al 1999, Salanueva et al 2003.
Rubella virus is another enveloped virus that assembles in association with Golgi membranes. Rubella virus is the only member of the Rubivirus genus within the Togaviridae family, a group of single-stranded RNA(+) viruses that also includes the Alphavirus genus, a group of well-known pathogens. However, outside of genomic coding strategy, these genera are quite dissimilar (Frey, 1994). Based on computer-assisted homology analysis, it has been proposed that rubella virus originated by a complicated event including recombination with progenitors of the current alphaviruses, human hepatitis E virus, and possibly plant viruses (Frey and Wolinsky, 1999). Rubella virus is the causative agent of a generally benign disease known as rubella or German measles in children and adults but is highly teratogenic during the first trimester of pregnancy, leading to congenital rubella syndrome (CRS). Rubella virus has no known natural host other than humans. Despite routine vaccination programs which have been in place for 30 years, the virus persists in the human population and remains an important human pathogen (Frey, 1994).
The study of rubella virus has progressed slowly because of the slow replication rate, low virus yield in infected cell culture, and lack of synchronous infection in cell culture. Infection is known to induce a variety of morphological changes in the host cell, including the modification of lysosomes to produce enlarged structures filled with vesicles known as cytopathic vacuoles (CVs). CVs contain the viral “RNA-replication complexes,” or RC, as defined by the presence of double-stranded RNA Lee et al 1994, Magliano et al 1998. Infection also produces a significant alteration of mitochondrial morphology and distribution (Lee et al., 1999). Similar RC-containing structures were previously identified in cells infected with alphaviruses (Froshauer et al., 1988). Subsequent virion assembly, however, follows very different pathways for these genera. While alphaviruses assemble nucleocapsids in the cytoplasm which then bud through the plasma membrane, rubella virions assemble in the Golgi. The genomic RNA is a single molecule of positive-stranded RNA about 10 kb in length. The rubella virus structural proteins, capsid protein (C) and glycoproteins (E2 and E1), are translated from a 24S subgenomic RNA yielding a 110-kDa polyprotein precursor with the internal order NH2-C-E2-E1-COOH that is co-/posttranslationally processed by lumenal signalase to produce the three mature proteins. Rubella virus nucleocapsid assembly occurs in association with membranes and is synchronized with virus budding (Frey, 1994). Conversely, alphavirus nucleocapsid assembly occurs independently of membranes and virus budding (Strauss and Strauss, 1994). The structure of alpha and rubella virions is also different. As revealed by cryoelectron microscopy studies, alphaviruses are icosahedral enveloped particles with an internal core tightly attached to the envelope, building a double T = 4 icosahedral shell (Mancini et al., 2000). Rubella virus contains a smaller core, clearly separated from the viral envelope (Murphy, 1980). A T = 3 icosahedral symmetry has been proposed for rubella virus capsids (Chantler et al., 2001), although direct structural evidence is lacking. To acquire potential new data on rubella virion assembly in the Golgi complex, we have studied the morphogenesis of rubella virus by freeze-substitution electron microscopy in Vero and BHK-21 cells, the two cell types routinely used to grow rubella virus. One-step multiplication conditions are more closely approached in Vero cells than in BHK-21 cells (Hemphill et al., 1988) and the number of extracellular virions is higher for rubella virus-infected Vero cells, while the number of intracellular virions is higher in infected BHK-21 cells (Bardeletti et al., 1979).
Results
As shown in Fig. 1, Vero cells infected with rubella virus exhibit clear changes in organelle distribution and organization, most notably recruitment of mitochondria to the perinuclear region, dissociation of the nucleolar components, and formation of dense deposits on the cytoplasmic side of the plasma membrane, as reported previously (Lee et al., 1996). Characteristic CVs in association with mitochondria and cisternae of the rough endoplasmic reticulum (Magliano et al., 1998) were observed. While in control, noninfected cells, mitochondria of a medium electron density were randomly distributed (Fig. 1A), infected cells at 16 h.p.i. had elongated mitochondria with increased electron density that clustered around CVs formed in perinuclear regions (Fig. 1B). When low-magnification fields of cells sectioned through an equatorial plane were studied (approximately 100 control and 100 infected cells were analyzed) 73% of mitochondria in infected cells were seen attached to CVs, while only 12% of mitochondria in control cells were detected close to a vesicle or a vacuolar structure. We frequently detected Golgi stacks containing virions in close proximity to these CV–mitochondria–RER complexes; this was a novel observation (Fig. 1C). Higher magnification of CVs (Fig. 1D) revealed small vesicles attached to the internal membrane surface. These vesicles (arrows in Fig. 1D) had a diameter of roughly 30 nm and contained a dense core. The morphology and distribution of CVs were similar to those described by Kujala et al. (1999). By 48 h.p.i., noticeable cell degeneration had occurred. At this time p.i. intact Golgi stacks were difficult to detect and numerous cells contained “empty” mitochondria (Fig. 1E).
Fig. 1.
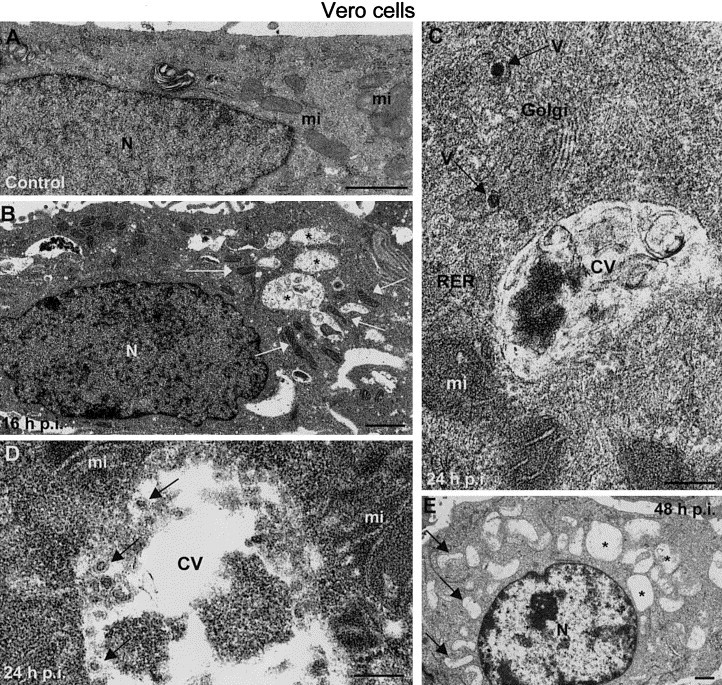
Associations of organelles in Vero cells infected with rubella virus. (A) Perinuclear areas in control, uninfected Vero cells, where mitochondria (mi) are randomly distributed. (B) Vero-infected cells at 16 h.p.i. exhibit cytopathic vacuoles (asterisks) surrounded by elongated, denser mitochondria (white arrows). (C) Higher magnification field showing a cytopathic vacuole (CV) attached to a mitochondrion (mi), rough endoplasmic reticulum (RER) cisternae, and a Golgi stack with viruses (V). (D) Cytopathic vacuole with vesicular structures containing dense spots (arrows). (E) Later in infection (48 h.p.i.) these associations of organelles mainly dissappear and in many cells both CVs (asterisks) and mitochondria (arrows) look empty. A, B, and D correspond to freeze-substituted cells, while C and E are conventionally processed samples. N, nucleus. Bars, 1 μm in A, B, and E; 200 nm in C and 100 nm in D.
Rubella virus replication in BHK cells is noncytopathic, although infected BHK-21 and Vero cells produce equivalent yields of virus. No significant alterations on cellular ultrastructure were distinguished at 24 or 48 h.p.i., with the exception of the formation of numerous characteristic cytopathic vacuoles (Figs. 2A and B) which were smaller and found in higher numbers than in infected Vero cells (Fig. 2B). These CVs were totally filled with vesicles 50–70 nm in diameter and contained an internal small dense spot (Fig. 2C). As in Vero cells, CVs accumulated on the perinuclear region, where the first viral particles were also distinguished (Fig. 2C).
Fig. 2.
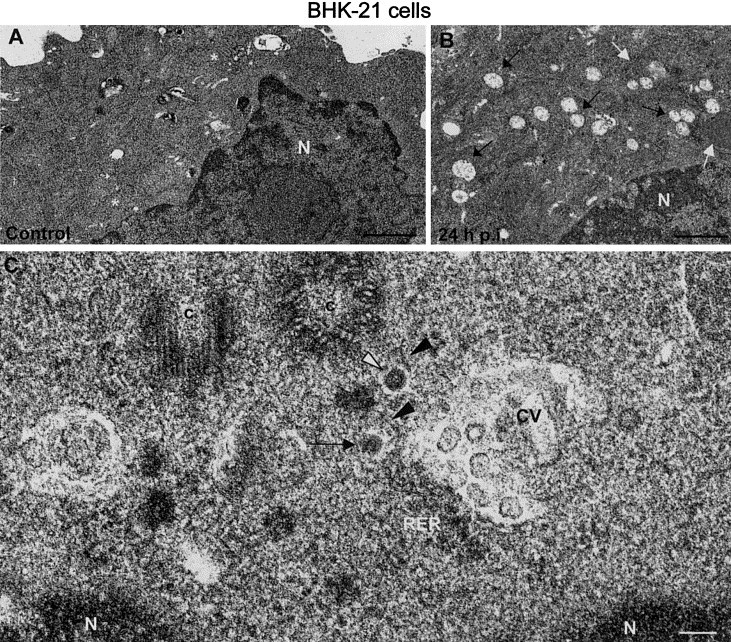
Ultrastructure of BHK-21 cells infected with rubella virus and processed by freeze-substitution. Low-magnification fields of (A) control uninfected BHK-21 cells and (B) rubella-infected cells (24 h.p.i.). Mitochondria are marked with white asterisks (A) or white arrowheads (B). Cytopathic vacuoles (black arrows in B) accumulate in the perinuclear region of infected cells. (C) Higher magnification field of the perinuclear region showing a budding profile (arrow) and a dense viral particle (white arrowhead) in smooth membranes (black arrowheads) close to a characteristic cytopathic vacuole (CV). N, nucleus; c, centrioles; RER, rough endoplasmic reticulum. Bars, 1 μm in A and B; 100 nm in C.
Analysis of viral particles in Vero cells was done at four different times p.i. representative of “early” (12 and 16 h.p.i.) and “late” (24 and 48 h.p.i.) replication. Because of the relative scarcity of virions at early times we had to scan a larger number of sections (around 500–700 infected cells per time point) than in later times. More than 400 control, noninfected Vero and BHK-21 cells were also studied to discard the potential presence of small structures of cellular origin in the locations of rubella replication and assembly (the whole study included observation of close to 5000 cells). At early times budding profiles and virus particles were detected inside Golgi stacks. These particles exhibited a predominant morphology, shown in Figs. 3A to E, with a diameter of 64.3 ± 3.8 nm and a homogeneously dense interior. Some particles were still attached to the budding membrane (Figs. 3C and D), while others showed fine contacts between the internal dense material and the particle membrane (Fig. 3E). Particles representative of a minor population are shown in Fig. 3F. In these, the majority of the interior of the particle was occupied by a dense material, but the center was less dense and had an annular structure, most probably representing an intermediate stage in the process of core formation. The third virion morphology inside the Golgi stack is shown in Fig. 3G and consists of viral particles with an internal core clearly separated from the viral envelope. This was the morphology previously described for rubella virions; in our sections it was the only virion morphology detected in secretory vesicles and the extracellular medium and thus represents the mature particle (Fig. 3H). At late times, coincident with significant deterioration of cell ultrastructure, the relative amount of intra-Golgi immature (uniformly dense) viral particles decreased and mature intra- and extracellular particles were the predominant class. By 48 h, the number of intracellular particles was very small and primarily extracellular virions were detected. All virion assembly detected in infected BHK cells was similar to that described in Vero cells, namely, homogeneously dense particles and particles with defined internal cores were observed inside Golgi stacks and in the perinuclear region (Figs. 3I and J), while extracellular particles were solely of the mature morphology (Fig. 3K).
Fig. 3.
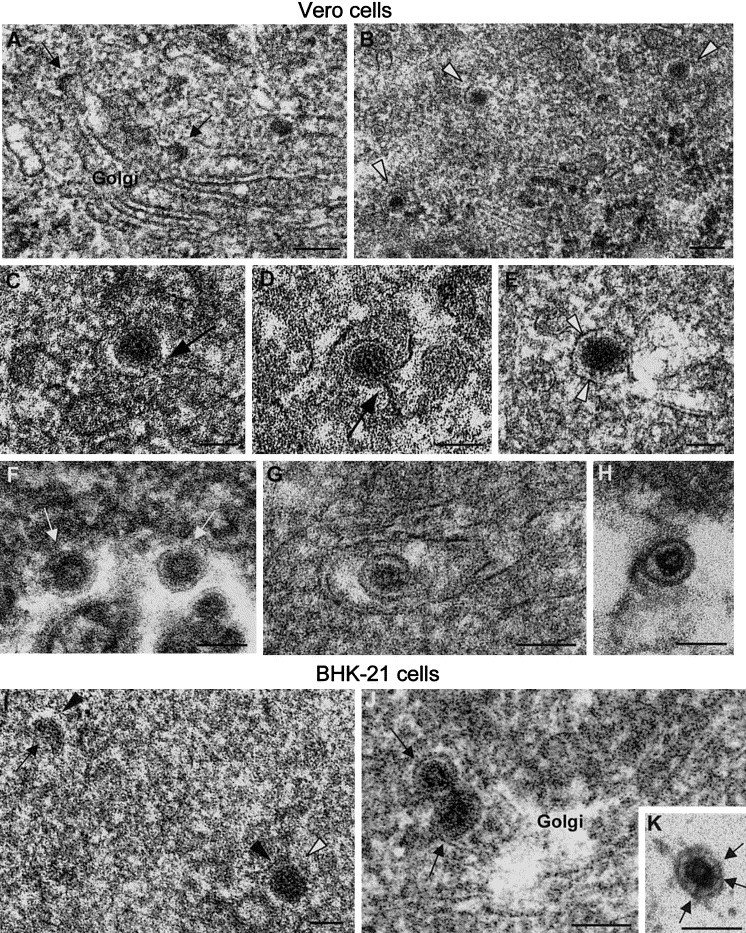
Rubella virus polymorphism in the Golgi complex. (A and B) Golgi membranes in infected Vero cells at 16 h.p.i. contain maturation arcs (arrows) and dense spherical particles (arrowheads). (C–E) Higher magnification fields showing the structure of dense rubella particles. Connections with budding membranes are marked with arrows. Focal contacts between the particle content and the envelope are marked in (E). (F) Viral particles (white arrows) with an annular-like morphology (dense periphery and less dense center) are seen as a minor class inside Golgi stacks. (G) Mature rubella virion inside the Golgi stack. Note the dense envelope and internal core well separated from the envelope, features also found in extracellular virions (H). (I–K) Rubella virus-infected BHK-21 cells. (I) Perinuclear budding profile (black arrowhead) and dense viral particle (white arrowhead) in smooth membranes (black arrowheads). (J) Mature viral particles (arrows) containing assembled cores inside a Golgi stack. (K) Extracellular virion attached to the cell surface containing a central dense core and connections between core and envelope (arrows). A conventionally processed sample is shown in A, while B to K correspond to freeze-substituted cells. Bars, 100 nm in A and B; 60 nm in C–K.
Fig. 4 shows a collection of images of representative extracellular virions. In all of the extracellular particles the viral envelope was very dense and clearly delineated. Small differences in density of the viral core content were apparent (compare, for example, Figs. 4A and B with C and D) and some cores exhibited a clear geometrical contour (Fig. 4B). In some virions some details of the core wall were distinguishable (arrow in Fig. 4E). Dense, fibrous connections between the internal core and the viral envelope were frequently observed (arrows in Fig. 3, Fig. 4). These connections were more evident in distorted viral particles (arrows in Figs. 4H and I). Extracellular virions were homogeneous in size, with a diameter of 61.5 ± 2.7 nm for the whole particle and 36.6 ± 2.1 nm for the internal core.
Fig. 4.
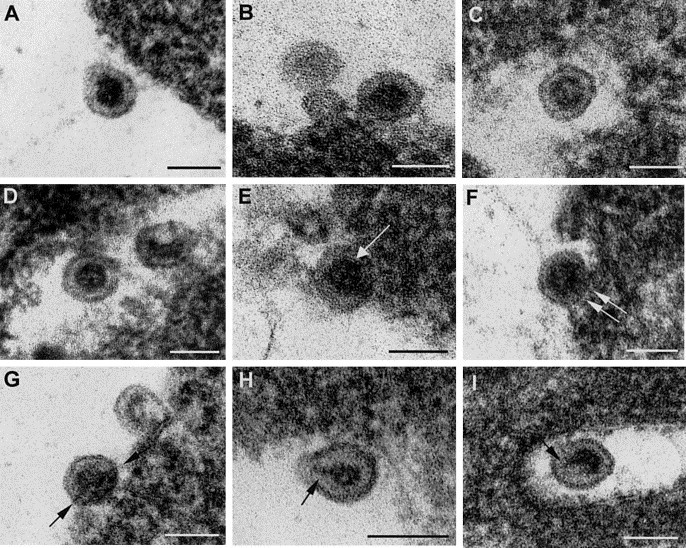
Structural details in rubella extracellular virions. Minor differences in the apparent density of the internal core are shown: (A and B) Virions with a denser core and (C and D) extracellular particles with less dense cores. (E–I) Shown are some internal details in extracellular virions. In (E) a layer (probably the core wall) is resolved (arrow), while in (F) and (G) envelope–core connections are clear (arrows). In distorted particles these contacts become more evident (arrows in H and I). All fields correspond to freeze-substituted samples. Bars, 60 nm.
Table 1 summarizes a quantitative analysis of rubella viral forms and their location in cells at different times postinfection.
Table 1.
Quantitation of viral forms in rubella virus-infected cells at different times postinfection
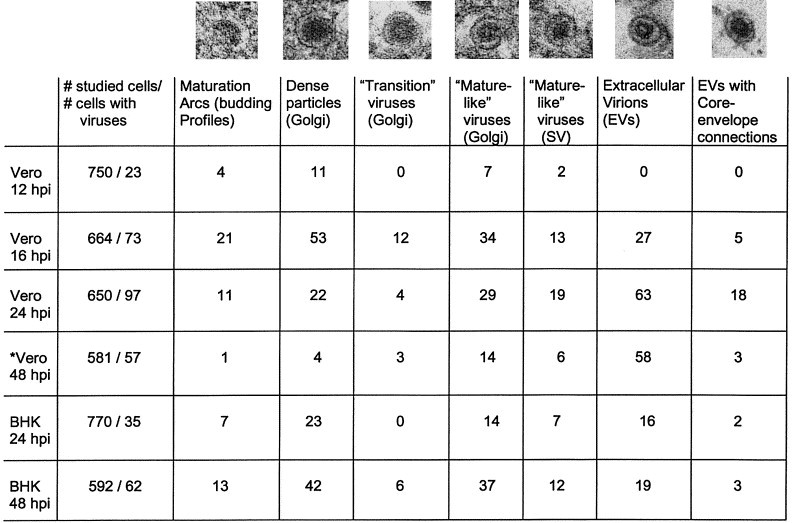
SV, secretory vesicles.
aExtensive cell lysis; 4007 sectioned cells were studied (347 of them contained viral particles). A total of 645 viral particles were counted and included in the table.
Discussion
This study revealed two novel aspects of rubella virus virion assembly: (1) the polymorphism of virus particles detected inside the Golgi complex strongly suggested the existence of a structural maturation process occurring inside the Golgi stack and (2) the association between Golgi stacks and CVs containing the viral RCs indicates a potential physical connection between the sites of viral replication and virion assembly that could mediate the transfer of replicated genomes to the assembly sites.
The presence of uniformly dense particles and the coexistence of these with “mature-like” viral forms containing a distinguishable core inside the Golgi stack, together with the exclusive presence of “mature-like” viruses inside secretory vesicles and in the extracellular space, strongly suggest that during rubella virion morphogenesis an immature dense precursor is initially assembled inside the Golgi which later transforms into the structurally mature particles with a distinguishable core. Intermediate stages could be also formed during maturation.
The rubella virus capsid protein associates with membranes due to the retention of the signal peptide from the E2 glycoprotein following cleavage (Baron et al., 1992). Thus, it has been hypothesized that the three structural proteins (C, E1, and E2) travel together from the RER to the Golgi complex, where virion assembly takes place. In contrast, the capsid protein of alphaviruses is cleaved by a viral endoproteolytic activity and released into the cytoplasm (Froshauer et al., 1988). Since the capsid protein molecules appear to be first associated with membranes, some reorganization inside the viral particle must take place to produce the mature core which resides in the center of the mature particle well separated from the virion envelope. The core–envelope fibrous connections detected in extracellular virions could be related to the early associations between the capsid protein and the membrane. However, this is only speculation and the nature of the mentioned connections remains to be defined.
Our original description of the structural maturation of coronaviruses (Salanueva et al., 1999) inside the Golgi stack were novel and we have since reported that bunyaviruses exhibit a similar process (Salanueva et al., 2003), although the morphogenetic pathway of these viruses is more complex, with two structural maturation steps. Rubella virus is then the third virus genus documented to undergo structural change inside the Golgi stacks. Rubella virus assembly would seem to be more similar to that of bunyaviruses than to coronaviruses since rubella immature particles transform rapidly after budding, as suggested by viral particles with assembled cores but still attached to the budding membrane (Fig. 3G). Another similarity in the maturation process of bunyaviruses and rubella virus is the efficiency of the mechanism that blocks the exit of immature viruses. While a significant number of coronavirus immature particles are seen inside secretory vesicles and outside of the cell, no immature viruses are seen on the surface of cells infected with bunyaviruses (Salanueva et al., 2003) or rubella virus (as seen in this work).
In the case of coronaviruses and bunyaviruses the use of drugs that affect transport along the secretory pathway in different locations allowed us to determine that the trans side of the Golgi stack is necessary for viral maturation. In the case of rubella virus the use of drugs for reversibly blocking maturation followed by EM studies would be difficult due to the low number of rubella virus particles accumulated inside infected cells (Table 1).
It is not known how the replicated genomes travel from the CVs to the sites of assembly in Golgi membranes; however, our detection of Golgi stacks near CVs suggested a physical association. Interestingly, in an immunocytochemical study Lee et al. (1999) reported that rubella virus capsid protein was localized on the periphery of mitochondria, and it was proposed that rubella virus cores were attached to the membrane of the CVs. In this analysis we could not confirm the existence of assembled cores on the periphery of CVs, mitochondria, or any location other than within mature virions. An alternative interpretation for the images reported by Lee et al. (1999) is that C protein is actually on the periphery of the CVs but not as assembled cores. It would be then interesting to define the role that C protein is playing there. Interestingly, in a study focused on the identification of host cell proteins involved in rubella virus assembly, Beatch and Hobman (2000) found that rubella virus capsid protein interacts with the host cell protein p32 on the periphery of mitochondria. P32 protein has been shown to bind a variety of other virus phosphoproteins that complex nucleic acids. Its interaction with rubella virus capsid protein could play a role in the regulation of nucleocapsid assembly (Beatch and Hobman, 2000).
Two experimental systems exist to disect, at least in part, those possibilities. First, replicons which express only the nonstructural proteins and replicate the replicon RNA (Tzeng et al., 2001) can be used to determined if the nonstructural proteins and the process of RNA replication is sufficient for attracting the RER, mitochondria, and Golgi to the CVs. Second, it has been found that expression of structural proteins results in formation of virus-like particles (VLPs) that are similar in all aspects, including morphology, to virions Hobman et al 1994, Qiu et al 1994. While VLPs do not contain genome RNA, it is not unequivocally clear whether they incorporate cellular RNA molecules. Study of VLP expressing cells would be of interest to determine if their morphogenesis is similar to the process observed during normal infection. Together with expression of rubella nonstructural proteins, these studies can help to characterize the factors involved in organelle recruitment and how this is involved in rubella replication and morphogenesis.
Materials and methods
Cells were infected with the F-Therien strain of rubella virus at a multiplicity of infection (m.o.i.) of 1 PFU/cell. Infected Vero cells were fixed at 12, 16, 24, and 48 h.p.i. while BHK-21 cells were fixed at 24 and 48 h.p.i. since infection progresses more slowly in this cell type and fewer cells are infected. After chemical fixation in a mixture of 2% glutaraldehyde and 2% tannic acid in HEPES buffer (pH 7.4), the cells were processed in parallel by fast freezing followed by freeze-substitution in a mixture of acetone and osmium tetroxide before embedding in an epoxy resin (Risco et al., 2002) or conventional dehydration in acetone at 4°C followed by embedding in an epoxy resin (Salanueva et al., 1999). Preservation of fine structural details is superior after freeze-substitution Risco et al 2002, Salanueva et al 2003 when compared with conventional processing, which, on the other hand, provides higher contrast of intracellular membranes. Ultrathin sections (30–40 nm) were stained following procedures for obtaining high contrast, such as using uranyl acetate in ethanol before treatment with lead citrate, as described (Risco et al., 2002). Sections were studied in a JEOL 1200-EX II electron microscope operating at 100 kV. Electron micrographs were scanned using an Epson Perfection 2450 PHOTO scanner and Picture Publisher 8 as software.
Acknowledgements
We express our gratitude to Dr. José Ruiz Castón for helpful discussions and to Emily S. Abernathy and Pilar Cabezas for technical assistance. This work has been supported by Grant BMC2000-0555 from the Ministerio de Ciencia y Tecnología of Spain (to C.R.) and by Grant AI-21389 from NIAID, NIH (to T.K.F.).
References
- Bardeletti G, Tektoff J, Gautheron D. Rubella virus maturation and production in two host cell systems. Intervirology. 1979;11:97–103. doi: 10.1159/000149019. [DOI] [PubMed] [Google Scholar]
- Baron M.D, Ebel T, Suomalainen M. Intracellular transport of rubella virus structural proteins expressed from cloned cDNA. J. Gen. Virol. 1992;73:1073–1086. doi: 10.1099/0022-1317-73-5-1073. [DOI] [PubMed] [Google Scholar]
- Beatch M.D, Hobman T.C. Rubella virus capsid associates with host cell protein p32 and localizes to mitochondria. J. Virol. 2000;74:5569–5576. doi: 10.1128/jvi.74.12.5569-5576.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chantler J, Wolinsky J.S, Tingle A. Rubella virus. In: Knipe D.M, Howley P.M, editors. Fields Virology. Lippincott Williams & Wilkins; Philadelphia, PA: 2001. pp. 963–990. [Google Scholar]
- Frey T.K. Molecular biology of rubella virus. Adv. Virus Res. 1994;44:69–160. doi: 10.1016/S0065-3527(08)60328-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frey T.K, Wolinsky J.S. Rubella virus (Togaviridae) In: Granoff A, Webster R.G, editors. Encyclopedia of Virology. Academic Press; San Diego, CA: 1999. pp. 1592–1601. [Google Scholar]
- Froshauer S, Kartenbeck J, Helenius A. Alphavirus RNA replicase is located on the cytoplasmic surface of endosomes and lysosomes. J. Cell Biol. 1988;107:2075–2986. doi: 10.1083/jcb.107.6.2075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hemphill M.L, Forng R.-Y, Abernathy E.S, Frey T.K. Time course of virus-specific macromolecular synthesis during rubella virus infection in Vero cells. Virology. 1988;162:65–75. doi: 10.1016/0042-6822(88)90395-9. [DOI] [PubMed] [Google Scholar]
- Hobman T.C, Lundstrom M.L, Mauracher C.A, Woodward L, Gillam S, Farquhar M.G. Assembly of rubella virus structural proteins into virus-like particles in transfected cells. Virology. 1994;202:574–585. doi: 10.1006/viro.1994.1379. [DOI] [PubMed] [Google Scholar]
- Kujala P, Ahola T, Ehsani N, Auvinen P, Vihinen H, Kääriänen L. Intracellular distribution of rubella virus nonstructural protein P150. J. Virol. 1999;73:7805–7811. doi: 10.1128/jvi.73.9.7805-7811.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J.-Y, Bowden D.S, Marshall J.A. Membrane junctions associated with rubella virus infected cells. J. Submicrosc. Cytol. Pathol. 1996;28:101–108. [PubMed] [Google Scholar]
- Lee J.-Y, Marshall J.A, Bowden D.S. Characterization of rubella virus replication complexes using antibodies to double-stranded RNA. Virology. 1994;200:307–312. doi: 10.1006/viro.1994.1192. [DOI] [PubMed] [Google Scholar]
- Lee J.-Y, Marshall J.A, Bowden D.S. Localization of rubella virus core particles in Vero cells. Virology. 1999;265:110–119. doi: 10.1006/viro.1999.0016. [DOI] [PubMed] [Google Scholar]
- Magliano D, Marshall J.A, Bowden D.S, Vardaxis N, Meanger J, Lee J.-Y. Rubella virus replication complexes are virus-modified lysosomes. Virology. 1998;240:57–63. doi: 10.1006/viro.1997.8906. [DOI] [PubMed] [Google Scholar]
- Mancini E.J, Clarke M, Gowen B.E, Rutten T, Fuller S.D. Cryo-electron microscopy reveals the functional organization of an enveloped virus, Semliki Forest Virus. Mol. Cell. 2000;5:255–266. doi: 10.1016/s1097-2765(00)80421-9. [DOI] [PubMed] [Google Scholar]
- Mckenzie J.M, Jones M.K, Westway E.G. Markers for trans-Golgi membranes and the intermediate compartment localize to induced membranes with distinct replication functions in flavivirus-infected cells. J. Virol. 1999;73:9555–9567. doi: 10.1128/jvi.73.11.9555-9567.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy F.A. Togavirus morphology and morphogenesis. In: Schlesinger R.W, editor. The Togaviruses. Academic Press; New York: 1980. pp. 241–316. [Google Scholar]
- Qiu Z, Ou D, Hobman T.C, Gillam S. Expression and characterization of virus-like particles containing rubella virus structural proteins. J. Virol. 1994;68:4086–4091. doi: 10.1128/jvi.68.6.4086-4091.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Risco C, Rodríguez J.R, López-Iglesias C, Carrascosa J.L, Esteban M, Rodriguez D. Endoplasmic reticulum-Golgi intermediate compartment membranes and vimentin filaments participate in vaccinia virus assembly. J. Virol. 2002;76:1839–1855. doi: 10.1128/JVI.76.4.1839-1855.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salanueva I.J, Carrascosa J.L, Risco C. Structural maturation of the transmissible gastroenteritis coronavirus. J. Virol. 1999;73:7952–7964. doi: 10.1128/jvi.73.10.7952-7964.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salanueva I.J, Novoa R.R, Cabezas P, López-Iglesias C, Carrascosa J.L, Elliott R.M, Risco C. Polymorphism and structural maturation of Bunyamwera virus in Golgi and post-Golgi compartments. J. Virol. 2003;77:1368–1381. doi: 10.1128/JVI.77.2.1368-1381.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strauss J.H, Strauss E.G. The alphaviruses: gene expression, replication, and evolution. Microbiol. Rev. 1994;58:491–562. doi: 10.1128/mr.58.3.491-562.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tzeng W.P, Chen M.-H, Derdeyn C.A, Frey T.K. Rubella virus DI RNAs and replicons: requirement for nonstructural proteins acting in cis for amplification by helper virus. Virology. 2001;289:63–73. doi: 10.1006/viro.2001.1088. [DOI] [PubMed] [Google Scholar]