

Abstract
The 2009 pandemic H1N1 infection in humans has been one of the greatest concerns for public health in recent years. However, influenza in pigs is a zoonotic viral disease well-known to virologists for almost one century with the classical H1N1 subtype the only responsible agent for swine influenza in the United States for many decades. Swine influenza was first recognized clinically in pigs in the Midwestern U.S. in 1918 and since that time it has remained important to the swine industry throughout the world. Since 1998, however, the epidemiology of swine influenza changed dramatically. A number of emerging subtypes and genotypes have become established in the U.S. swine population. The ability of multiple influenza virus lineages to infect pigs is associated with the emergence of reassortant viruses with new genomic arrangements, and the introduction of the 2009 pandemic H1N1 from humans to swine represents a well-known example. The recent epidemiological data regarding the current state of influenza A virus subtypes circulating in the Canadian and American swine population is discussed in this review.
Keywords: Influenza Virus, H1N1 Virus, Swine Influenza, Pandemic H1N1, Swine Influenza Virus
Brief Introduction to Influenza A Viruses
Influenza is a zoonotic viral disease that represents a health and economic threat to both human and animals worldwide. Influenza A viruses are the most studied of the Orthomyxoviridae since they can infect a large variety of birds and mammals including humans, pigs, horses, domestic poultry, marine mammals, cats, dogs and wild carnivores (Webster 2002; Thiry 2007). Wild aquatic birds were shown to be an asymptomatic reservoir for most subtypes of influenza A viruses (Scholtissek 1978; Fouchier et al. 2005). Moreover, influenza A virus ecology is intricate due to the high number of possible reassortment events and cross-species jumps that lead to their evolution (Webster et al. 1992). The hemagglutinin (HA) and the neuraminidase (NA) proteins encoded by gene segments 4 and 6, respectively, play a key role in the influenza life cycle and represent the primary targets of the host humoral immune response (Skehel and Wiley 2000). The HA protein is the most important determinant of virulence and host specificity as it binds to sialic acid-containing cell surface receptors on host epithelial cells (Shinya et al. 2006; Nicholls et al. 2008; Ayora-Talavera et al. 2009; de Wit et al. 2010). The HA mediates virus binding to N-acetylneuraminic acid-2,3-galactose (2,3-sialic acid) or N-acetylneuraminic acid-2,6-galactose (2,6-sialic acid) terminal residues on sialyloligosaccharides for avian and mammalian virus primary binding predilection, respectively (Rogers and Paulson 1983). However, receptor binding restriction has been shown to be more complicated than previously understood, with tissues from human, swine and Japanese quail expressing both 2,3- and 2,6-sialic acid receptor types (Ito et al. 1998; Suzuki et al. 2000; Shinya et al. 2006; Wan and Perez 2006). Additionally, glycan array analysis has demonstrated that avian and mammalian adapted flu viruses can have binding spillover to the opposing receptor linkage type and that different strains bind preferentially to novel structures (such as sulphated and sialylated glycans) (Stevens et al. 2006). The NA is responsible for cleaving terminal sialic acid residues from carbohydrate moieties on the surfaces of the host cell and virus (Gottschalk 1957), thus assisting in virus cell entry by mucus degradation (Matrosovich et al. 2004) and the release and spread of progeny virions (Palese et al. 1974). The remaining six segments encode for the following structural and accessory proteins: PB2 (segment 1), PB1 (segment 2), PA (segment 3), NP (segment 5), M1 and M2 (segment 7), NS1 and NEP (segment 8) (Lamb and Krug 2007). Both HA and NA genes undergo two types of variation called antigenic drift and antigenic shift. Antigenic drift involves minor changes in the HA and NA due to polymerase errors during replication, whereas antigenic shift involves major changes in these molecules resulting from replacement of the entire gene segment as a consequence of reassortment events in the event that two (or more) unique viruses infect the same cell (Webster 1971). Based upon the major differences within the HA and NA proteins, 16 HA and 9 NA subtypes, naturally paired in different combinations, have been identified thus far (Webster et al. 1992; Rohm et al. 1996; Fouchier et al. 2005). Only a limited number of subtypes have been established in mammals. For example, only viruses of H1, H2, H3, N1 and N2 subtypes have circulated widely in the human population (Webster et al. 1992; Alexander and Brown 2000) and only H1, H3, N1 and N2 subtypes have been consistently isolated from pigs (Webster et al. 1992; Olsen 2002).
Influenza A Virus in Pigs
First Detection in the United States
Swine influenza was first recognized in pigs in the Midwestern U.S. in 1918 (Fig. 1) as a respiratory disease that coincided with the human pandemic known as the Spanish flu (Koen 1919). Since then, it has become an important disease to the swine industry throughout the world. The first influenza virus was isolated in 1930 by Shope (1931) and was demonstrated to cause respiratory disease in swine that was similar to human influenza. This strain was subsequently recognized as an H1N1 influenza virus, and swine were utilized in the following years as a model to study influenza pathogenesis in a natural host.
Fig. 1.
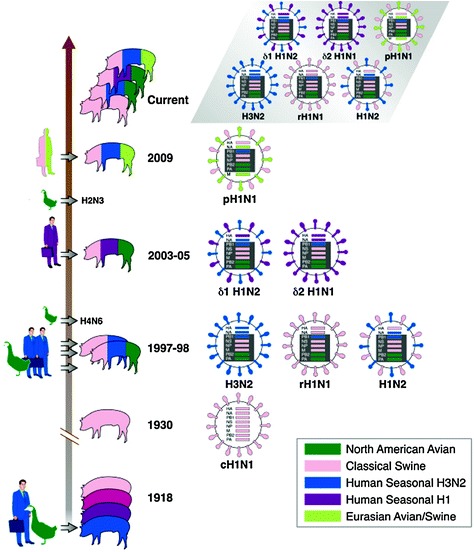
Epidemiology and genetic composition of influenza viruses from U.S. and Canadian pigs. Swine virus lineage is color coded pink, avian lineage is coded green, human lineage is coded blue or purple. The chronology of transmission events leading to reassortant viruses with genes from swine, human and avian influenza virus lineages is visualized by the vertical arrow. The “Spanish flu” virus was transmitted from avian/human origin to pigs and evolved into the cH1N1, as indicated by the transition in color of pigs from blue to light blue to red to pink. The human and avian images to the left of the vertical timeline represent the species origin of viral gene segments donated to give rise to the swine influenza virus reassortants listed on the right side of the vertical timeline. Time line is not drawn to scale. Each viral subtype is represented with its eight gene segment arrangement. The triple reassortant H3N2 reassorted with the cH1N1 to produce rH1N1 and H1N2 subtypes with the triple reassortant internal gene (TRIG) cassette. Further reassortment events with two independent human H1 subtype viruses led to the δ1 H1N2 and δ2 H1N1. The source of the reassortment event producing the combination of gene segments in the 2009 pandemic H1N1 prior to its emergence in human and subsequent transmission from humans to pigs in 2009 is currently unknown. Light green indicates the Eurasian avian/swine lineage. The gray highlighted area illustrates the currently circulating influenza A subtypes in Canadian and American pigs
Introduction of the Triple Reassortant H3N2 in Pigs
Among the RNA viruses affecting mammals, influenza viruses and coronaviruses represent, as a consequence of different molecular mechanisms, two of the best examples of viruses with exceptionally plastic genomes. Thus, we should not be surprised that the high mutation and reassortment rates have propelled the evolution of influenza viruses in pigs in recent years. However, from the first characterization of swine influenza virus until the late 1990s, the classical swine lineage H1N1 (cH1N1) was relatively stable at the genetic and antigenic levels in U.S. swine. Based on phylogenetic analysis, the cH1N1 lineage is closely related to the 1918 H1N1 Spanish flu virus (Easterday and van Reeth 1999) and other human influenza viruses isolated in the 1930s following the discovery of SIV. Although the cH1N1 was the predominant subtype causing disease in pigs until the late 1990s, there was serological evidence that human subtype H3 influenza viruses were circulating at a low frequency in U.S. pigs, but failed to establish a lineage with sustained transmission among swine (Chambers et al. 1991).
The epidemiology of influenza in pigs dramatically changed after the events of 1997–1998 (Fig. 1). In 1998, a severe influenza-like disease was observed in pigs in North Carolina with additional outbreaks in swine herds in Minnesota, Iowa and Texas. The causative agents for these outbreaks were identified as influenza A viruses of the H3N2 subtype. Genetic analysis of these H3N2 viruses showed that at least two different genotypes were present. The initial North Carolina isolate was a double reassortant and contained gene segments similar to those of the classical swine lineage (PB2, PA, NP, M, NS) combined with gene segments from a human seasonal H3N2 influenza virus circulating in 1995 (PB1, HA, NA). The isolates from Minnesota, Iowa and Texas were triple reassortants containing gene segments from the classical swine virus (NP, M, NS,) and the same human virus (PB1, HA, NA) in combination with an avian virus (PB2, PA) (Zhou et al. 1999). By the end of 1999, viruses antigenically and genetically related to the triple reassortant lineage were widespread in the U.S. swine population (Webby et al. 2000) whereas the double reassortant virus did not become established. Interestingly, the double and triple reassortant H3N2 viruses were shown to possess a similar HA encoding gene with identical residues in critical receptor binding regions, suggesting that their different successes were due to factors not associated with the HA and receptor binding pocket. The major difference between the two viruses was the acquisition of two avian polymerase genes (PB2 and PA) in the triple reassortant virus. The human lineage PB1, avian lineage PB2 and PA and swine lineage NP, M and NS found in contemporary swine influenza viruses are referred to as the triple reassortant internal gene (TRIG) constellation (Vincent et al. 2008). Genetic and antigenic evaluation of H3N2 swine influenza isolates since 1998 (Richt et al. 2003; Webby et al. 2004) indicate at least three introductions of human H3 subtype viruses became established in swine, leading to phylogenetic clusters I, II and III. The cluster III viruses have become dominant in North America (Gramer et al. 2007) and have continued to evolve into cluster III variants, also known as cluster IV (Olsen et al. 2006).
The H3N2 viruses not only evolved and became endemic in pigs but also reassorted with extant cH1N1 swine influenza viruses. The vast majority of the resulting reassortant and drift variant viruses since 1998 contain the TRIG. The H1N1 viruses containing the HA and NA from the cH1N1 virus and the TRIG from triple reassortant H3N2 viruses are referred as reassortant H1N1 (rH1N1) and the viruses containing the HA from the classical swine virus and the NA and TRIG from the triple reassortant H3N2 virus are H1N2 viruses (Karasin et al. 2002; Webby et al. 2004) (Fig. 1). Reassortant viruses have become endemic and co-circulate in most major swine producing regions of the U.S. and Canada, including further drift variants of H3N2 (Webby et al. 2000,2004; Richt et al. 2003; Olsen et al. 2006), H1N2 (Choi et al. 2002; Karasin et al. 2002), and rH1N1 (Webby et al. 2004). H3N1 viruses have occasionally been identified in limited outbreaks but do not appear to circulate widely (Lekcharoensuk et al. 2006; Ma et al. 2006). Moreover, the TRIG was shown to have accepted an avian lineage H2 and N3, producing a novel triple reassortant swine H2N3 in 2006 (Ma et al. 2007). More recently, introduction of H1 viruses with the HA gene of human H1N2 seasonal influenza virus origin (hu-like H1) that are genetically and antigenically distinct from the classical swine H1 lineage were reported in pigs in Canada (Karasin et al. 2006) (Fig. 1). Since 2005, hu-like H1N1 and H1N2 viruses containing the TRIG have emerged in swine herds across the U.S. (Vincent et al. 2009b) that have HA and NA segments most similar to H1N1 and H1N2 human seasonal influenza virus lineages from around 2003.
Evolution of the H1 Subtype
The well characterized contemporary swine influenza reassortant viruses possessing the ability to spread and become established in U.S. and Canadian swine populations have contained similar TRIG constellations. This would suggest that the TRIG constellation can accept multiple HA and NA types and may confer a selective advantage to viruses possessing this gene cassette (Bastien et al. 2010; Vijaykrishna et al. 2010). Moreover, since the acquisition of TRIG, an increase in the rate of mutation in North American swine influenza isolates appears to have occurred in H1 subtype hemagglutinins. Genetic mutation may be related to antigenic changes if mutations occur in antigenic sites of the HA, potentially resulting in escape from herd immunity. This scenario is in stark contrast with that observed with the cH1N1 viruses prior to acquiring TRIG. Indeed, cH1N1 viruses remained relatively stable genetically and antigenically for at least seven decades (Sheerar et al. 1989; Luoh et al. 1992; Noble et al. 1993; Olsen et al. 1993).
For best representing the evolution of the currently circulating H1 viruses, a cluster classification has been proposed (Fig. 2a). Viruses from the classical H1N1 lineage-HA acquired from the TRIG cassette evolved to form α-, β-, and γ-clusters based on the genetic makeup of the HA gene; whereas H1 subtypes strains with HA genes most similar to human seasonal H1 viruses form the δ-cluster (Vincent et al. 2009b). All four HA gene cluster types can be found with NA genes of either the N1 or N2 subtype. In order to study the evolution and the antigenic relationships among the H1 swine influenza virus subtypes, we recently analyzed 12 different strains, selected from the University of Minnesota Veterinary Diagnostic Laboratory (UMVDL) diagnostic case database (Lorusso et al. 2011). The viruses were isolated from outbreaks of respiratory disease in pigs from diagnostic cases submitted to the UMVDL in 2008 and are representative of each of the postulated four H1 clusters. All gene segments were sequenced and analyzed, and antigenic changes were measured for all twelve viruses using the hemagglutination inhibition (HI) assay and mapped by antigenic cartography. All 2008 H1 viruses contained the North American TRIG. Furthermore, variation was demonstrated in the six genes that make up the TRIG, but no HA cluster-specific patterns were detected among the genes composing the TRIG constellation. In contrast, an HA cluster-specific pattern was observed for the NA gene. The N1 gene of the α, β and γ cluster of the 2008 H1 viruses and of sequences publicly available each formed a separate clade within the North American N1 cluster. We speculate that the evolution of the H1 gene drives that of the N1 gene as well. Indeed, antigenic drifts that characterize the evolutionary history of the antigenic and phylogenetic clusters of H1 influenza virus in U.S. swine isolates were accompanied by changes in the N1 genes, thus allowing a parallel sub-cluster classification (Fig. 2b). A proper HA/NA pairing in association with the TRIG could optimize viral transmission and replication as shown by recent experiments in pigs. Indeed, experimental coinfection in the lower respiratory tract of inoculated pigs with two phylogenetically and genetically distant viruses, a triple reassortant H3N2 and cH1N1, resulted in the genesis, of all possible HA/NA combinations but only the parental H3N2 was found in two consecutive direct contact pig groups (Ma et al. 2010). These results confirm that multiple reassortments can occur but not all reassortants are readily transmissible.
Fig. 2.
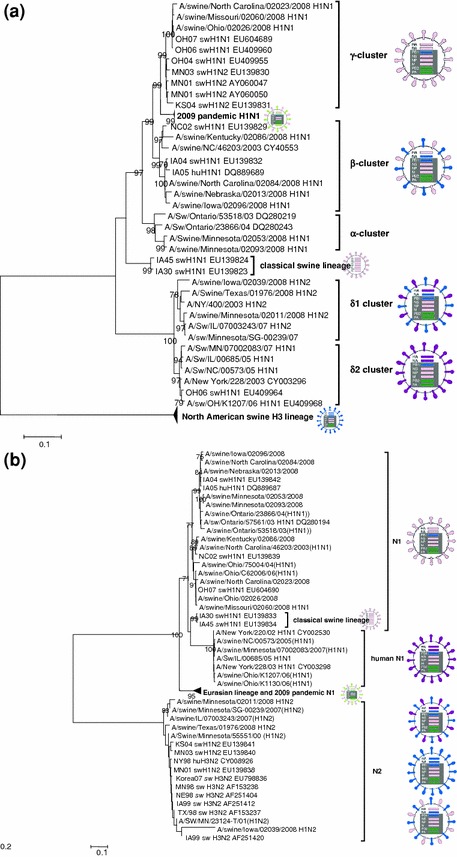
Neighbor-joining trees inferred from multiple nucleotide sequence alignment of segment 4 (HA, a) and segment 6 (NA, 2b). a shows four H1 clusters of viruses, H1α, H1β, H1γ and H1δ (human-like H1) as indicated by the bars on the right of the tree. In both trees, the HA cluster specificity is indicated. The genomic constellation of each clade is indicated by the images on the right side of the tree. Classical swine lineage is color coded pink, avian lineage is coded green, human lineage is coded blue or purple. Light green indicates the Eurasian avian/swine lineage. Classical swine lineage-HA gene (a) was acquired by the TRIG cassette and evolved overtime to form α-, β- and γ- clusters. The introduction of human seasonal HA from H1N2 and H1N1 gave rise to δ cluster viruses differentiated phylogenetically by two distinct sub-clusters, δ1 and δ2 (a). Similar to the δ-cluster viruses in the HA phylogenetic analysis, β-viruses have split into two sub-clusters (b). Phylogenetic analyzes were conducted in MEGA4. Statistical support was provided by bootstrapping over 1,000 replicates and bootstrap values >70 are indicated at the correspondent node. The scale bars indicate the estimated numbers of nucleotide substitutions per site. human (Hu), swine (Sw)
The viruses representing the classical swine H1 lineage, phylogenetic clusters α, β and γ, had moderate to strong cross-reactivity within a cluster, especially within recent β- and γ-cluster viruses. However, cross-reactivity between clusters was more variable, ranging from no cross-reactivity to strong cross-reactivity, such as between α- and β-cluster viruses. This study suggested that the H1 is evolving by drift while maintaining the TRIG backbone, and that the resulting viruses differ genetically and antigenically with obvious consequences for vaccine and diagnostic test development. In 2008 and 2009, α cluster H1 viruses were rarely isolated from influenza outbreaks in pigs in North America, and while β cluster H1 viruses are still common, they occur with less frequency than the more dominant subtypes from the the γ and δ viruses. Since the acquisition of TRIG, the H1 of the classical swine lineage, under apparent evolutionary pressure, has developed multiple amino acid changes in the putative antigenic sites. The γ viruses are chronologically the newest H1 variants and it cannot be ruled out that the same mechanisms will be responsible for further H1 cluster variants. The genetic diversity within the H1 clusters was confirmed functionally by the demonstrated loss in cross-reactivity in the HI assay between H1 clusters overall. It is likely that, as a consequence of evolutionary and immunogical pressures, the H1 will continue to mutate in the future, allowing evasion of the immune system of the host or only partially protective immunity.
Human-Like H1 Viruses
Since 2005, H1N1 and H1N2 viruses with the HA gene derived from human viruses have spread across the U.S. in swine herds forming the δ-cluster H1 (Vincent et al. 2009b) (Fig. 1). The HAs from the human-like (hu) swine H1 viruses are genetically and antigenically distinct from classical swine lineage and derivatives. Indeed the putative antigenic site in the HA1 of the hu-like viruses possesses typical human lineage residues in contrast to that found in the HA1 of the α-, β- and γ-clusters (Lorusso et al. 2011). However, their TRIG genes are similar to those found in the TRIG cassette of the contemporary swine triple reassortant viruses (Vincent et al. 2009b). The HA from the δ-cluster viruses were shown to have most likely emerged from at least two separate introductions of human seasonal HA from H1N2 and H1N1 viruses being differentiated phylogenetically by two distinct sub-clusters, δ1 and δ2, respectively, (Lorusso et al. 2011; Vincent et al. 2009a). Viruses belonging to the δ-cluster were shown to be paired either with a N1 or N2 gene consistently of human lineage and not of swine lineage N1. δ1-subcluster viruses, first detected in 2003, showed an N2 gene preference whereas δ2-subcluster viruses, first detected in 2005, showed an N1 preference (Fig. 2b) initially but have subsequently begun to reassort. Limited HI cross-reactivity was demonstrated between the δ1 and δ2 viruses thus supporting the scenario assumed by the phylogenetic analysis (Fig. 2a). The hu-H1 viruses have become one of the major subtypes of influenza virus isolated and characterized from swine respiratory disease outbreaks. Indeed, if we consider the time period 2008–2010, the incidence of hu-H1 in swine respiratory disease outbreaks has dramatically increased. In 2008, 85% of the influenza viruses isolated from swine diagnostic cases submitted to the UMVDL were shown to be of the H1 subtype. Most of the H1 isolates (up to 78%) were of the γ- and β-cluster with the γ-cluster viruses found in slightly higher numbers, whereas δ-cluster viruses represented approximately 20% of the total. However, in 2009 the epidemiologic scenario changed. While the influenza A viruses isolated were mostly H1 subtype (five-fold more than the H3 subtype), the number of δ viruses now represented 40% of the total, thus quickly becoming the dominant subtype isolated from cases of respiratory disease. β- and γ-cluster viruses were 35 and 23% of the total H1 clusters represented, respectively. The same trend was shown in the early months of 2010 as well, with a slight increase in the number of δ-cluster viruses compared to the γ-cluster viruses, cluster IV H3 subtype viruses, and the newly emerged 2009 pandemic H1N1. An experimental in vivo study in 4-week-old pigs with an H1N1 isolate of the δ2-subcluster demonstrated differences in kinetics of lung lesion development, viral load in the lung and nasal shedding when compared to a virulent rH1N1 in the β-cluster. This study suggested the emerging virus genotype may not have been fully adapted to the swine host since virus replication in the lung and virus shedding from the nose were reduced compared to a contemporary rH1N1 (Vincent et al. 2009b). A more recent pathogenesis and transmission study in pigs comparing viruses in the δ1- and δ2-subclusters recapitulated the phenotypic differences seen in the initial study; however, the δ1-subcluster virus studied demonstrated increased virulence and nasal shedding over the δ2-subcluster viruses (Ciacci-Zanella, unpublished). Further studies are warranted in order to monitor the evolution of δ-cluster viruses. The presence of typical “human-like” residues in the receptor binding pocket in the HA of two of the δ-cluster viruses isolated in 2008 demonstrates that although these viruses have replicated in pigs for over five years, the swine viruses may preserve human-adapted receptor binding phenotypes (Lorusso et al. 2011). This preservation of human-like residues in the swine host may allow potential novel reassortant influenza viruses, including the δ-cluster swine viruses, to spill back into the human population. Escaping the immune response by changing the external makeup is a well-known strategy that influenza viruses adopt. The acquisition of human HA segments by the TRIG cassette platform were shown to be entirely different from those of the classical swine lineage and further drift derivatives provided an important antigenic advantage for these reassortant viruses. Indeed, the number of influenza outbreaks in which δ-cluster viruses were recognized as causative agents increased in the recent years. Moreover, geographical regions have differing cluster variants circulating, thus further complicating vaccine strain selection.
2009 Pandemic H1N1 in Pigs
In the early spring of 2009, the United States, Canada and Mexico reported community outbreaks of pneumonia in humans caused by a novel H1N1 influenza A virus. This virus subsequently spread across the globe at a high rate, prompting the WHO to declare a pandemic in June 2009 (Garten et al. 2009). Retrospectively, the earliest known case was identified February 24, 2009, in a baby from San Louis Potosi, Mexico (http://news.sciencemag.org/scienceinsider/2009/07/yet-another-new.html). This novel pandemic H1N1 possesses a unique genome with six gene segments (PB2, PB1, PA, HA, NP and NS) most closely related to the triple reassortant influenza viruses of the North American swine lineage, and the M and NA genes derived from a Eurasian lineage of swine influenza viruses (Dawood et al. 2009). The 2009 pandemic influenza became infamously known as “swine flu” due to the phylogenetic origin of the gene segments. However, since the recognition of the outbreak, infection in humans has not been connected to pig exposure (Dawood et al. 2009). Indeed, as it was believed to have occurred in 1918 (Webster 2002), humans transmitted the novel virus to pigs, as most of the initial documented swine outbreaks were preceded by reported human influenza-like illness during the human pandemic (Cohen 2010). The 2009 pandemic H1N1 was promptly shown to replicate efficiently in the lower and upper respiratory tract of infected pigs and to cause a clinical disease comparable to that typically observed during common, enzootic influenza virus infection in swine (Lange et al. 2009; Vincent et al. 2009a). Early reference to the 2009 pandemic H1N1 as “swine flu” led to unnecessary alarm over the safety of pork meat products and culminated in the ban of exported pork from the U.S. by several countries, resulting in billions of dollars in lost revenue for the swine industry (http://agriculture.house.gov/testimony/111/h102209/Butler.pdf). However, contamination of fresh pork meat with the novel virus was experimentally excluded (Vincent et al. 2009a). Immediately after the onset in humans, cases of infection of pigs with the pandemic 2009 H1N1 were reported in different areas of the world (http://www.oie.int/wahis/public.php?page=weekly_report_index&admin=0). The first case was detected on April 28, 2009 in Canada (Leslieville, Alberta) in a farm with pigs that were not previously vaccinated against swine influenza (Howden et al. 2009; Weingartl et al. 2010). The source of the outbreak was linked to a worker who showed symptoms of influenza-like disease (Howden et al. 2009). Pigs infected with the 2009 pandemic H1N1 were first detected in the U.S. in a farm in Indiana in November 2009 (Lowe et al. 2010). Based on recent data the 2009 H1N1 continued to spread from humans to susceptible pigs with subsequent sustained pig-to-pig transmission and, thus establishing yet another endemic virus in swine populations. Importantly, none of the eight genes of the 2009 pandemic H1N1 cluster tightly with the genes of swine influenza viruses circulating in the U.S. prior of the outbreak in humans (Lorusso et al. 2011; Smith et al. 2009). In the phylogenetic analyzes of each gene segment, the 2009 pandemic H1N1 formed a distinct and independent branch from the U.S. swine lineage genes of the 2008 H1 isolates evaluated as well as swine virus sequences available from GenBank. This suggests that neither the 2009 pandemic H1N1 nor closely related progenitor viral genes were present in U.S. swine influenza viruses prior to 2009 (Fig. 2a, Lorusso et al. 2011). A closely related progenitor virus with the same 8-gene constellation has yet to be identified in swine or other species, although a 2004 swine virus with 7/8 of the 2009 pandemic H1N1 genome was identified in Hong Kong, China (Smith et al. 2009).
The HA of the 2009 pandemic H1N1 is most closely related to the North American swine γ-cluster H1 lineage viruses (Fig. 2a). Limited serologic cross-reactivity was demonstrated in HI tests using two 2009 pandemic H1N1 human strains as antigens against sera from pigs immunized with 2007–2008 (notably γ-cluster) swine viruses (Vincent et al. 2010). Thus, prior exposure to some H1 subtypes is likely to provide swine with some level of protection against infection with the 2009 human pandemic H1N1. This is also suggested by data from human epidemiological studies that showed high prevalence of neutralizing antibodies against 2009 pandemic H1N1 in people born before 1930 (Itoh et al. 2009; Munster et al. 2009). Moreover, immunization in mice with human H1N1 viruses that circulated before 1945 (e.g. specific antibodies against 1918 H1N1 or related viruses) is sufficient for immune protection from the 2009 pandemic H1N1 (Manicassamy et al. 2010). Furthermore, 2009 pandemic H1N1 viruses cause sustained human-to-human transmission and there are several case reports and experimental studies demonstrating human-to-swine (Howden et al. 2009), swine-to-swine (Vincent unpublished; Lange et al. 2009; Brookes et al. 2010) and swine-to-human transmission (Weingartl et al. 2010). Thus, the 2009 pandemic H1N1, being a virus shared between people and pigs, has the potential to further change the epidemiology of influenza viruses in human and swine populations.
Exceptional Influenza A Subtypes: H2N3 and H4N6
Two H2N3 influenza viruses were isolated in 2006 from clinically affected pigs from two different farms in the central U.S. (Ma et al. 2007). The epidemiologic link identified between the two farms, besides the geographical location, was the use of water collected from open-air ponds for pig drinking water and cleaning the barns. This subtype was not reported in pigs before and has not been identified since. Sequencing demonstrated both viruses were H2N3 influenza A viruses sharing 99.3–99.9% genetic homology. Although the H2N3 viruses contained the typical North American TRIG, the HA segment was similar to an avian influenza virus H2N3 isolated from mallards and the NA sequence was similar to an avian influenza virus H4N3 isolated from blue-winged teal. Importantly, the predicted HA1 protein displayed an amino acid constellation in the receptor binding area suggesting a preferential affinity to the mammalian receptor. This HA mutation resembled that of the initial reassortant human influenza isolates found in the beginning of the 1957 H2N2 pandemic. To investigate the transmission and pathogenesis features of the novel virus, in vivo studies with pigs, mice and ferrets were conducted. Both swine and ferrets were shown to efficiently transmit virus to contact animals; moreover the virus was lethal to young mice. The combined data demonstrating the ability of the H2N3 viruses with a typical avian-origin HA to replicate in three different animal models confirmed adaptation to the mammalian host, indicating a potential risk to the human population. However, serological studies conducted on workers exposed to H2N3 infected pigs showed no evidence of zoonotic transmission (Beaudoin et al. 2010).
A similar scenario was described in October 1999 in a swine farm in Canada. This farm was located near a lake on which large numbers of waterfowl congregate each fall and from which the farm drew water. The source of infection was postulated to have come from ducks on the adjacent lake. Viruses isolated from infected pigs, were shown genetically and antigenically to be wholly avian H4N6 viruses of the North American avian lineage (Karasin et al. 2000). Moreover, the wholly avian H4N6 virus spread from pig-to-pig (Olsen 2002), but has not been detected outside the primary swine farm system. Interestingly, even in this case of a wholly avian virus, the receptor binding pocket of the putative HA protein displayed residues postulated to be associated with mammalian receptor binding (Karasin et al. 2000).
Triple Reassortant Swine Viruses Found Outside North America
China is a major area of swine and poultry production. Influenza virus infection in pigs was first described in 1918 in China coincident, as in other areas of the world, with the Spanish flu pandemic in humans (Brown 2000). It has been documented that four subtypes (H1N1, H1N2, H3N1, and H3N2) are circulating in Chinese pigs (Shu et al. 1994; Guan et al. 1996; Peiris et al. 2001; Xu et al. 2004; Qi and Lu 2006; Qi et al. 2009). Studies from 1976 to 1982 revealed cocirculation of classical swine H1N1 and human-like H3N2 and double reassortant H3N2 viruses containing the surface genes HA and NA from the human viruses and the internal genes from swine H1N1 viruses (Shortridge et al. 1979, 1987; Xu et al. 2004). In 1993–1994, avian-like H1N1 viruses were detected in pigs in southern China (Guan et al. 1996). In 2004, a reassortant H1N2 virus containing the NA gene of a human H3N2 and the remaining seven genes of the classical H1N1 swine viruses emerged (Qi and Lu 2006). Recently, interspecies transmission of human H1N1, avian H5N1 and avian H9N2 to pigs has been reported (Peiris et al. 2001; Yu et al. 2007; Zhu et al. 2008). Moreover, recent studies revealed the presence of novel avian-like H1N1strains that seem to be derived from the European swine H1N1 viruses and novel triple reassortant H1N2 carrying the TRIG. These novel triple reassortant viruses have the HA of the classical swine lineage whereas the NA is of human origin (Yu et al. 2009). The epidemiology of influenza viruses in the Chinese swine population is complex and worthy of increased monitoring and reporting.
Currently, at least four different subtypes of swine influenza viruses, classical H1N1, H1N2, H3N1 and H3N2, are circulating in South Korean swine herds with H1N2 being the dominant subtype (Jung and Chae 2005; Shin et al. 2006; Jung et al. 2007; Song et al. 2007). Multiple HA lineages are present for the H1 and H3 subtypes (Pascua et al. 2008). H1N1 viruses currently circulating show the HA is closely related to that of the first H1N2 isolated in South Korea in 2002, A/swine/Korea/CY02/02. However, more recent H1N2 isolates contained HA genes closely related to an H1N2 isolate (A/swine/MN/16419/01) reported in USA. These findings suggest that the HA genes from recent H1N2 viruses originated from viral sources other than the pre-existing H1N2 isolates in Korea, such as those from USA. The phylogeny of the H3 subtype is more complicated. A recent study showed that four different H3 lineages cocirculate in Korean swine. The first lineage displayed an H3 identical to that described in USA in the earliest H3N2 isolates containing the TRIG constellation, A/swine/TX/4199/2/98 (cluster I), whereas the HA gene of the second lineage appeared to be derived from the American cluster III H3N2. Another strain was found to possess an H3 similar to those of the American cluster II H3N2. The remaining lineage contains new emerging reassortant viruses in which the HA gene originated from an earlier human-like isolate (A/NY/647/95). All the viruses characterized from the period 2002–2007 in South Korea were shown to possess the TRIG. Most recently, two H5N2 viruses with the surface glycoproteins from the Eurasian avian lineage were isolated from swine. While the first of the two was a wholly avian virus, the second was a double reassortant between an avian virus (PB1, HA, NA, NS) and a triple reassortant swine H3N1 (PB2, PA, NP, M) (Lee et al. 2009).
The TRIG genome constellation of influenza viruses from Canadian and American pigs has not been described in Europe, whereas recent studies reveal its presence in China and Korea. Many major swine producing regions of the world are under-represented in the influenza genome databases and reports in the literature documenting the status of influenza in swine are likewise limited or absent. However, the global spread of the 2009 pandemic H1N1 carrying a modified TRIG constellation represents a new viral source for the TRIG with an unpredictable consequence to the epidemiology of swine influenza viruses worldwide. Thus, increased and sustainable global surveillance for influenza viruses in swine is needed in order to develop control and prevention programs for swine health as well as rapid detection and reporting of emerging zoonotic influenza threats.
Conclusions
Currently, up to seven antigenic HA clusters and subtypes are cocirculating in pigs in Canada and the United States: α-, β-, γ-, δ1- and δ2-cluster viruses of the H1 subtype, 2009 pandemic H1N1 and the cluster IV triple reassortant H3N2. The HA of α-, β- or γ-cluster H1 viruses can be paired with either N1 or N2 genes. The N2 was introduced from humans in the 1990s with the H3N2 and is well-established in the swine population. However, δ1- and δ2-subcluster human-like viruses where shown to be preferentially paired with an N1 or N2 genes consistently of human lineage. δ1-subcluster viruses showed an N2 gene preference whereas δ2-subcluster viruses showed an N1 preference.
The key element in understanding influenza virus epidemiology in all of the influenza A viruses resides in the fascinating plastic nature of their genome. Whereas pandemic influenza outbreaks occur following rare antigenic shifts, established influenza viruses perpetually undergo antigenic drift allowing survival in the host population. The 2009 pandemic H1N1 underscores the potential risk to the human population from additional influenza virus subtypes and genotypes with the swine influenza TRIG backbone and demonstrates the potential for viruses with genes from swine lineages to emerge and cause illness in the human population. On the other hand, from the swine industry point of view, pigs have been the victim of human-adapted seasonal influenza viruses, as well as the 2009 pandemic H1N1. Although the 2009 pandemic H1N1 does not pose a greater risk in terms of clinical disease over viruses endemic prior to 2009, its emergence in the swine populations makes influenza control strategies exceedingly difficult. Additionally, there is a concern for the novel pandemic H1N1 virus to pick up or contribute genes of different sources in the swine host, potentially generating further reassortant viruses with potential implications for the human and swine populations. Indeed, a novel reassortant virus has been recently found in pigs in Hong Kong. It was shown to have the NA gene of 2009 pandemic H1N1 origin, the HA gene of the avian-like H1N1 swine isolates, whereas the remaining genes are TRIG-derived (Vijaykrishna et al. 2010). This is the first documented event of reassortment involving the 2009 pandemic H1N1 but not likely the last.
Contributor Information
Jürgen A. Richt, Phone: 785-532 4408, Email: jricht@vet.k-state.edu
Richard J. Webby, Email: richard.webby@stjude.org
Amy L. Vincent, Email: amy.vincent@ars.usda.gov
References
- Alexander DJ, Brown IH. Recent zoonoses caused by influenza A viruses. Rev Sci Tech. 2000;19:197–225. doi: 10.20506/rst.19.1.1220. [DOI] [PubMed] [Google Scholar]
- Ayora-Talavera G, Shelton H, Scull MA, Ren J, Jones IM, Pickles RJ, Barclay WS. Mutations in H5N1 influenza virus hemagglutinin that confer binding to human tracheal airway epithelium. PLoS One. 2009;4:e7836. doi: 10.1371/journal.pone.0007836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bastien N, Antonishyn NA, Brandt K, Wong CE, Chokani K, Vegh N, Horsman GB, Tyler S, Graham MR, Plummer FA, Levett PN, Li Y. Human infection with a triple-reassortant swine influenza A(H1N1) virus containing the hemagglutinin and neuraminidase genes of seasonal influenza virus. J Infect Dis. 2010;201:1178–1182. doi: 10.1086/651507. [DOI] [PubMed] [Google Scholar]
- Beaudoin A, Gramer M, Gray GC, Capuano A, Setterquist S, Bender J. Serologic survey of swine workers for exposure to H2N3 swine influenza A. Influenza Other Respi Viruses. 2010;4:163–170. doi: 10.1111/j.1750-2659.2009.00127.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brookes SM, Núñez A, Choudhury B, Matrosovich M, Essen SC, Clifford D, Slomka MJ, Kuntz-Simon G, Garcon F, Nash B, Hanna A, Heegaard PM, Quéguiner S, Chiapponi C, Bublot M, Garcia JM, Gardner R, Foni E, Loeffen W, Larsen L, Van Reeth K, Banks J, Irvine RM, IH B (2010) Replication, pathogenesis and transmission of pandemic (H1N1) 2009 virus in non-immune pigs. PLoS One 5:e9068 [DOI] [PMC free article] [PubMed]
- Brown IH. The epidemiology and evolution of influenza viruses in pigs. Vet Microbiol. 2000;74:29–46. doi: 10.1016/S0378-1135(00)00164-4. [DOI] [PubMed] [Google Scholar]
- Chambers TM, Hinshaw VS, Kawaoka Y, Easterday BC, Webster RG. Influenza viral infection of swine in the United States 1988–1989. Arch Virol. 1991;116:261–265. doi: 10.1007/BF01319247. [DOI] [PubMed] [Google Scholar]
- Choi YK, Goyal SM, Farnham MW, Joo HS. Phylogenetic analysis of H1N2 isolates of influenza A virus from pigs in the United States. Virus Res. 2002;87:173–179. doi: 10.1016/S0168-1702(02)00053-9. [DOI] [PubMed] [Google Scholar]
- Cohen J. Swine flu pandemic. What’s old is new: 1918 virus matches 2009 H1N1 strain. Science. 2010;327:1563–1564. doi: 10.1126/science.327.5973.1563. [DOI] [PubMed] [Google Scholar]
- Dawood FS, Jain S, Finelli L, Shaw MW, Lindstrom S, Garten RJ, Gubareva LV, Xu X, Bridges CB, Uyeki TM. Emergence of a novel swine-origin influenza A (H1N1) virus in humans. N Engl J Med. 2009;360:2605–2615. doi: 10.1056/NEJMoa0903810. [DOI] [PubMed] [Google Scholar]
- de Wit E, Munster VJ, van Riel D, Beyer WE, Rimmelzwaan GF, Kuiken T, Osterhaus AD, Fouchier RA. Molecular determinants of adaptation of highly pathogenic avian influenza H7N7 viruses to efficient replication in the human host. J Virol. 2010;84:1597–1606. doi: 10.1128/JVI.01783-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Easterday BC, Reeth KV. Swine Influenza. In: Straw BE, D’Allaire S, Mengeling WL, Taylor DJ, editors. Disease of swine. 8. Ames: Iowa State University; 1999. [Google Scholar]
- Fouchier RA, Munster V, Wallensten A, Bestebroer TM, Herfst S, Smith D, Rimmelzwaan GF, Olsen B, Osterhaus AD. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J Virol. 2005;79:2814–2822. doi: 10.1128/JVI.79.5.2814-2822.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garten RJ, Davis CT, Russell CA, Shu B, Lindstrom S, Balish A, Sessions WM, Xu X, Skepner E, Deyde V, Okomo-Adhiambo M, Gubareva L, Barnes J, Smith CB, Emery SL, Hillman MJ, Rivailler P, Smagala J, de Graaf M, Burke DF, Fouchier RA, Pappas C, Alpuche-Aranda CM, Lopez-Gatell H, Olivera H, Lopez I, Myers CA, Faix D, Blair PJ, Yu C, Keene KM, Dotson PD, Jr, Boxrud D, Sambol AR, Abid SH, St George K, Bannerman T, Moore AL, Stringer DJ, Blevins P, Demmler-Harrison GJ, Ginsberg M, Kriner P, Waterman S, Smole S, Guevara HF, Belongia EA, Clark PA, Beatrice ST, Donis R, Katz J, Finelli L, Bridges CB, Shaw M, Jernigan DB, Uyeki TM, Smith DJ, Klimov AI, Cox NJ. Antigenic and genetic characteristics of swine-origin 2009 A(H1N1) influenza viruses circulating in humans. Science. 2009;325:197–201. doi: 10.1126/science.1176225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottschalk A. Neuraminidase: the specific enzyme of influenza virus and Vibrio cholerae. Biochim Biophys Acta. 1957;23:645–646. doi: 10.1016/0006-3002(57)90389-X. [DOI] [PubMed] [Google Scholar]
- Gramer MR, Lee JH, Choi YK, Goyal SM, Joo HS. Serologic and genetic characterization of North American H3N2 swine influenza A viruses. Can J Vet Res. 2007;71:201–206. [PMC free article] [PubMed] [Google Scholar]
- Guan Y, Shortridge KF, Krauss S, Li PH, Kawaoka Y, Webster RG. Emergence of avian H1N1 influenza viruses in pigs in China. J Virol. 1996;70:8041–8046. doi: 10.1128/jvi.70.11.8041-8046.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howden KJ, Brockhoff EJ, Caya FD, McLeod LJ, Lavoie M, Ing JD, Bystrom JM, Alexandersen S, Pasick JM, Berhane Y, Morrison ME, Keenliside JM, Laurendeau S, Rohonczy EB. An investigation into human pandemic influenza virus (H1N1) 2009 on an Alberta swine farm. Can Vet J. 2009;50:1153–1161. [PMC free article] [PubMed] [Google Scholar]
- Ito T, Couceiro JN, Kelm S, Baum LG, Krauss S, Castrucci MR, Donatelli I, Kida H, Paulson JC, Webster RG, Kawaoka Y. Molecular basis for the generation in pigs of influenza A viruses with pandemic potential. J Virol. 1998;72:7367–7373. doi: 10.1128/jvi.72.9.7367-7373.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Itoh Y, Shinya K, Kiso M, Watanabe T, Sakoda Y, Hatta M, Muramoto Y, Tamura D, Sakai-Tagawa Y, Noda T, Sakabe S, Imai M, Hatta Y, Watanabe S, Li C, Yamada S, Fujii K, Murakami S, Imai H, Kakugawa S, Ito M, Takano R, Iwatsuki-Horimoto K, Shimojima M, Horimoto T, Goto H, Takahashi K, Makino A, Ishigaki H, Nakayama M, Okamatsu M, Warshauer D, Shult PA, Saito R, Suzuki H, Furuta Y, Yamashita M, Mitamura K, Nakano K, Nakamura M, Brockman-Schneider R, Mitamura H, Yamazaki M, Sugaya N, Suresh M, Ozawa M, Neumann G, Gern J, Kida H, Ogasawara K, Kawaoka Y. In vitro and in vivo characterization of new swine-origin H1N1 influenza viruses. Nature. 2009;460:1021–1025. doi: 10.1038/nature08260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jung K, Chae C. First outbreak of respiratory disease associated with swine influenza H1N2 virus in pigs in Korea. J Vet Diagn Invest. 2005;17:176–178. doi: 10.1177/104063870501700213. [DOI] [PubMed] [Google Scholar]
- Jung K, Song DS, Kang BK, Oh JS, Park BK. Serologic surveillance of swine H1 and H3 and avian H5 and H9 influenza A virus infections in swine population in Korea. Prev Vet Med. 2007;79:294–303. doi: 10.1016/j.prevetmed.2006.12.005. [DOI] [PubMed] [Google Scholar]
- Karasin AI, Brown IH, Carman S, Olsen CW. Isolation and characterization of H4N6 avian influenza viruses from pigs with pneumonia in Canada. J Virol. 2000;74:9322–9327. doi: 10.1128/JVI.74.19.9322-9327.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karasin AI, Landgraf J, Swenson S, Erickson G, Goyal S, Woodruff M, Scherba G, Anderson G, Olsen CW. Genetic characterization of H1N2 influenza A viruses isolated from pigs throughout the United States. J Clin Microbiol. 2002;40:1073–1079. doi: 10.1128/JCM.40.3.1073-1079.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karasin AI, Carman S, Olsen CW. Identification of human H1N2 and human-swine reassortant H1N2 and H1N1 influenza A viruses among pigs in Ontario, Canada (2003 to 2005) J Clin Microbiol. 2006;44:1123–1126. doi: 10.1128/JCM.44.3.1123-1126.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koen JS. A practical method for field diagnosis of swine diseases. Am J Vet Med. 1919;14:468–470. [Google Scholar]
- Lamb RA, Krug MA. Orthomyxoviridae: the viruses and their replication. In: Knipe D, Howley PM, Griffin E, Lamb RA, Martin MA, editors. Fields Virology. 5. Philadelphia: Lippincott Williams and Wilkins; 2007. [Google Scholar]
- Lange E, Kalthoff D, Blohm U, Teifke JP, Breithaup tA, Maresch C, Starick E, Fereidouni S, Hoffmann B, Mettenleiter TC, Beer M, Vahlenkamp TW (2009) Pathogenesis and transmission of the novel swine-origin influenza virus A/H1N1 after experimental infection of pigs. J Gen Virol 90:2119–2123 [DOI] [PubMed]
- Lee JH, Pascua PN, Song MS, Baek YH, Kim CJ, Choi HW, Sung MH, Webby RJ, Webster RG, Poo H, Choi YK. Isolation and genetic characterization of H5N2 influenza viruses from pigs in Korea. J Virol. 2009;83:4205–4215. doi: 10.1128/JVI.02403-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lekcharoensuk P, Lager KM, Vemulapalli R, Woodruff M, Vincent AL, Richt JA. Novel swine influenza virus subtype H3N1, United States. Emerg Infect Dis. 2006;12:787–794. doi: 10.3201/eid1205.051060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lorusso A, Vincent AL, Harland ML, Bayles DO, Swenson SL, Gramer MR, Russell CA, Smith DJ, Lager KM, Lewis NS. Genetic and antigenic characterization of H1 influenza viruses from United States swine from 2008. J Gen Virol. 2011;92:919–930. doi: 10.1099/vir.0.027557-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowe JF, Probst-Miller S., Augsburger DM, Gramer MR (2010) Clinical presentation and diagnosis of 2009 pandemic H1N1 influenza in two commercial swine herds located in the Midwestern United States. In: Proceedings of the neglected influenza virus symposium, Feb 2010, Amelia Island, Florida
- Luoh SM, McGregor MW, Hinshaw VS. Hemagglutinin mutations related to antigenic variation in H1 swine influenza viruses. J Virol. 1992;66:1066–1073. doi: 10.1128/jvi.66.2.1066-1073.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma W, Gramer M, Rossow K, Yoon KJ. Isolation and genetic characterization of new reassortant H3N1 swine influenza virus from pigs in the midwestern United States. J Virol. 2006;80:5092–5096. doi: 10.1128/JVI.80.10.5092-5096.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma W, Vincent AL, Gramer MR, Brockwell CB, Lager KM, Janke BH, Gauger PC, Patnayak DP, Webby RJ, Richt JA. Identification of H2N3 influenza A viruses from swine in the United States. Proc Natl Acad Sci USA. 2007;104:20949–20954. doi: 10.1073/pnas.0710286104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma W, Lager KM, Lekcharoensuk P, Ulery ES, Janke BH, Solorzano A, Webby RJ, Garcia-Sastre A, Richt JA. Viral reassortment and transmission after coinfection of pigs with classical H1N1 and triple reassortant H3N2 swine influenza viruses. J Gen Virol. 2010;91:2314–2321. doi: 10.1099/vir.0.021402-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manicassamy B, Medina RA, Hai R, Tsibane T, Stertz S, Nistal-Villan E, Palese P, Basler CF, Garcia-Sastre A. Protection of mice against lethal challenge with 2009 H1N1 influenza A virus by 1918-like and classical swine H1N1 based vaccines. PLoS Pathog. 2010;6:e1000745. doi: 10.1371/journal.ppat.1000745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matrosovich MN, Matrosovich TY, Gray T, Roberts NA, Klenk HD. Neuraminidase is important for the initiation of influenza virus infection in human airway epithelium. J Virol. 2004;78:12665–12667. doi: 10.1128/JVI.78.22.12665-12667.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munster VJ, de Wit E, van den Brand JM, Herfst S, Schrauwen EJ, Bestebroer TM, van de Vijver D, Boucher CA, Koopmans M, Rimmelzwaan GF, Kuiken T, Osterhaus AD, Fouchier RA. Pathogenesis and transmission of swine-origin 2009 A (H1N1) influenza virus in ferrets. Science. 2009;325:481–483. doi: 10.1126/science.1177127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicholls JM, Chan RW, Russell RJ, Air GM, Peiris JS. Evolving complexities of influenza virus and its receptors. Trends Microbiol. 2008;16:149–157. doi: 10.1016/j.tim.2008.01.008. [DOI] [PubMed] [Google Scholar]
- Noble S, McGregor MS, Wentworth DE, Hinshaw VS. Antigenic and genetic conservation of the haemagglutinin in H1N1 swine influenza viruses. J Gen Virol. 1993;74:1197–1200. doi: 10.1099/0022-1317-74-6-1197. [DOI] [PubMed] [Google Scholar]
- Olsen CW. The emergence of novel swine influenza viruses in North America. Vir Res. 2002;85:199–210. doi: 10.1016/S0168-1702(02)00027-8. [DOI] [PubMed] [Google Scholar]
- Olsen CW, McGregor MW, Cooley AJ, Schantz B, Hotze B, Hinshaw VS. Antigenic and genetic analysis of a recently isolated H1N1 swine influenza virus. Am J Vet Res. 1993;54:1630–1636. [PubMed] [Google Scholar]
- Olsen CW, Karasin AI, Carman S, Li Y, Bastien N, Ojkic D, Alves D, Charbonneau G, Henning BM, Low DE, Burton L, Broukhanski G. Triple reassortant H3N2 influenza A viruses, Canada, 2005. Emerg Infect Dis. 2006;12:1132–1135. doi: 10.3201/eid1207.060268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palese P, Schulman JL, Bodo G, Meindl P. Inhibition of influenza and parainfluenza virus replication in tissue culture by 2-deoxy-2,3-dehydro-N-trifluoroacetylneuraminic acid (FANA) Virology. 1974;59:490–498. doi: 10.1016/0042-6822(74)90458-9. [DOI] [PubMed] [Google Scholar]
- Pascua PN, Song MS, Lee JH, Choi HW, Han JH, Kim JH, Yoo GJ, Kim CJ, Choi YK. Seroprevalence and genetic evolutions of swine influenza viruses under vaccination pressure in Korean swine herds. Virus Res. 2008;138:43–49. doi: 10.1016/j.virusres.2008.08.005. [DOI] [PubMed] [Google Scholar]
- Peiris JS, Guan Y, Markwell D, Ghose P, Webster RG, Shortridge KF. Cocirculation of avian H9N2 and contemporary “human” H3N2 influenza A viruses in pigs in southeastern China: potential for genetic reassortment? J Virol. 2001;75:9679–9686. doi: 10.1128/JVI.75.20.9679-9686.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi X, Lu CP. Genetic characterization of novel reassortant H1N2 influenza A viruses isolated from pigs in southeastern China. Arch Virol. 2006;151:2289–2299. doi: 10.1007/s00705-006-0796-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi X, Pang B, Lu CP. Genetic characterization of H1N1 swine influenza A viruses isolated in eastern China. Virus Genes. 2009;39:193–199. doi: 10.1007/s11262-009-0375-9. [DOI] [PubMed] [Google Scholar]
- Richt JA, Lager KM, Janke BH, Woods RD, Webster RG, Webby RJ. Pathogenic and antigenic properties of phylogenetically distinct reassortant H3N2 swine influenza viruses cocirculating in the United States. J Clin Microbiol. 2003;41:3198–3205. doi: 10.1128/JCM.41.7.3198-3205.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers GN, Paulson JC. Receptor determinants of human and animal influenza virus isolates: differences in receptor specificity of the H3 hemagglutinin based on species of origin. Virology. 1983;127:361–373. doi: 10.1016/0042-6822(83)90150-2. [DOI] [PubMed] [Google Scholar]
- Rohm C, Zhou N, Suss J, Mackenzie J, Webster RG. Characterization of a novel influenza hemagglutinin, H15: criteria for determination of influenza A subtypes. Virology. 1996;217:508–516. doi: 10.1006/viro.1996.0145. [DOI] [PubMed] [Google Scholar]
- Scholtissek C, Rohde W, Von-Hoyningen V, Rott R. On the origin of the human influenza virus subtypes H2N2 and H3N2. Virology. 1978;87:13–20. doi: 10.1016/0042-6822(78)90153-8. [DOI] [PubMed] [Google Scholar]
- Sheerar MG, Easterday BC, Hinshaw VS. Antigenic conservation of H1N1 swine influenza viruses. J Gen Virol. 1989;70:3297–3303. doi: 10.1099/0022-1317-70-12-3297. [DOI] [PubMed] [Google Scholar]
- Shin JY, Song MS, Lee EH, Lee YM, Kim SY, Kim HK, Choi JK, Kim CJ, Webby RJ, Choi YK. Isolation and characterization of novel H3N1 swine influenza viruses from pigs with respiratory diseases in Korea. J. Clin. Microbiol. 2006;44:3923–3927. doi: 10.1128/JCM.00904-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinya K, Ebina M, Yamada S, Ono M, Kasai N, Kawaoka Y. Avian flu: influenza virus receptors in the human airway. Nature. 2006;440:435–436. doi: 10.1038/440435a. [DOI] [PubMed] [Google Scholar]
- Shortridge KF, Cherry A, Kendal AP. Further studies of the antigenic properties of H3N2 strains of influenza A isolated from swine in South East Asia. J Gen Virol. 1979;44:251–254. doi: 10.1099/0022-1317-44-1-251. [DOI] [PubMed] [Google Scholar]
- Shortridge KF, King AP, Webster RG. Monoclonal antibodies for characterizing H3N2 influenza viruses that persist in pigs in China. J Infect Dis. 1987;755:577–581. doi: 10.1093/infdis/155.3.577. [DOI] [PubMed] [Google Scholar]
- Shu LL, Lin YP, Wright SM, Shortridge KF, Webster RG. Evidence for interspecies transmission and reassortment of influenza A viruses in pigs in southern China. Virology. 1994;202:825–833. doi: 10.1006/viro.1994.1404. [DOI] [PubMed] [Google Scholar]
- Skehel JJ, Wiley DC. Receptor binding and membrane fusion in virus entry: the influenza hemagglutinin. Annu Rev Biochem. 2000;69:531–569. doi: 10.1146/annurev.biochem.69.1.531. [DOI] [PubMed] [Google Scholar]
- Smith GJ, Vijaykrishna D, Bahl J, Lycett SJ, Worobey M, Pybus OG, Ma SK, Cheung CL, Raghwani J, Bhatt S, Peiris JS, Guan Y, Rambaut A. Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. Nature. 2009;459:1122–1125. doi: 10.1038/nature08182. [DOI] [PubMed] [Google Scholar]
- Song DS, Lee CS, Jung K, Kang BK, Oh JS, Yoon YD, Lee JH, Park BK. Isolation and phylogenetic analysis of H1N1 swine influenza virus isolated in Korea. Virus Res. 2007;125:98–103. doi: 10.1016/j.virusres.2006.11.008. [DOI] [PubMed] [Google Scholar]
- Stevens J, Blixt O, Glaser L, Taubenberger JK, Palese P, Paulson JC, Wilson IA. Glycan microarray analysis of the hemagglutinins from modern and pandemic influenza viruses reveals different receptor specificities. J Mol Biol. 2006;355:1143–1155. doi: 10.1016/j.jmb.2005.11.002. [DOI] [PubMed] [Google Scholar]
- Suzuki Y, Ito T, Suzuki T, Holland RE, Jr, Chambers TM, Kiso M, Ishida H, Kawaoka Y. Sialic acid species as a determinant of the host range of influenza A viruses. J Virol. 2000;74:11825–11831. doi: 10.1128/JVI.74.24.11825-11831.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiry E, Zicola A, Addie D, Egberink H, Hartmann K, Lutz H, Poulet H, Horzinek MC. Highly pathogenic avian influenza H5N1 virus in cats and other carnivores. Vet Microbiol. 2007;122:25–31. doi: 10.1016/j.vetmic.2006.12.021. [DOI] [PubMed] [Google Scholar]
- Vijaykrishna D, Poon LLM, Zhu HC, Ma SK, Li OTW, Cheung CL, Smith GJD, Peiris JSM, Guan Y. Reassortment of pandemic H1N1/2009 influenza A virus in swine. Science. 2010;328:1529. doi: 10.1126/science.1189132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vincent AL, Ma W, Lager KM, Janke BH, Richt JA. Swine influenza viruses a North American perspective. Adv Virus Res. 2008;72:127–154. doi: 10.1016/S0065-3527(08)00403-X. [DOI] [PubMed] [Google Scholar]
- Vincent AL, Lager KM, Harland M, Lorusso A, Zanella E, Ciacci-Zanella JR, Kehrli ME, Klimov A. Absence of 2009 pandemic H1N1 influenza A virus in fresh pork. PLoS One. 2009;4:e8367. doi: 10.1371/journal.pone.0008367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vincent AL, Ma W, Lager KM, Gramer MR, Richt JA, Janke BH. Characterization of a newly emerged genetic cluster of H1N1 and H1N2 swine influenza virus in the United States. Virus Genes. 2009;39:176–185. doi: 10.1007/s11262-009-0386-6. [DOI] [PubMed] [Google Scholar]
- Vincent AL, Lager KM, Faaberg KS, Harland M, Zanella EL, Ciacci-Zanella JR, Kehrli ME, Jr, Janke BH, Klimov A. Experimental inoculation of pigs with pandemic H1N1 2009 virus and HI cross-reactivity with contemporary swine influenza virus antisera. Influenza Other Respi Viruses. 2010;4:53–60. doi: 10.1111/j.1750-2659.2009.00121.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wan H, Perez DR. Quail carry sialic acid receptors compatible with binding of avian and human influenza viruses. Virology. 2006;346:278–286. doi: 10.1016/j.virol.2005.10.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webby RJ, Swenson SL, Krauss SL, Gerrish PJ, Goyal SM, Webster RG. Evolution of swine H3N2 influenza viruses in the United States. J Virol. 2000;74:8243–8251. doi: 10.1128/JVI.74.18.8243-8251.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webby RJ, Rossow K, Erickson G, Sims Y, Webster R. Multiple lineages of antigenically and genetically diverse influenza A virus co-circulate in the United States swine population. Virus Res. 2004;103:67–73. doi: 10.1016/j.virusres.2004.02.015. [DOI] [PubMed] [Google Scholar]
- Webster RG. Antigenic variation in influenza virus. Biology and chemistry. Prog Med Virol. 1971;13:271–338. [PubMed] [Google Scholar]
- Webster RG. The importance of animal influenza for human disease. Vaccine. 2002;20:S16–S20. doi: 10.1016/S0264-410X(02)00123-8. [DOI] [PubMed] [Google Scholar]
- Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y. Evolution and ecology of influenza A viruses. Microbiol Rev. 1992;56:152–179. doi: 10.1128/mr.56.1.152-179.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weingartl HM, Berhane Y, Hisanaga T, Neufeld J, Kehler H, Emburry-Hyatt C, Hooper-McGreevy KKS, Dalman B, Bystrom J, Alexandersen S, Li Y, Pasick J. Genetic and pathobiologic characterization of pandemic H1N1 2009 influenza viruses from a naturally infected swine herd. J Virol. 2010;84:2245–2256. doi: 10.1128/JVI.02118-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu C, Fan W, Wei R, Zhao H. Isolation and identification of swine influenza recombinant A/Swine/Shandong/1/2003(H9N2) virus. Microbes Infect. 2004;6:919–925. doi: 10.1016/j.micinf.2004.04.015. [DOI] [PubMed] [Google Scholar]
- Yu H, Zhang GH, Hua RH, Zhang Q, Liu TQ, Liao M, Tong GZ. Isolation and genetic analysis of human origin H1N1 and H3N2 influenza viruses from pigs in China. Biochem Biophys Res Commun. 2007;356:91–96. doi: 10.1016/j.bbrc.2007.02.096. [DOI] [PubMed] [Google Scholar]
- Yu H, Zhang PC, Zhou YJ, Li GX, Pan J, Yan LP, Shi XX, Liu HL, Tong GZ. Isolation and genetic characterization of avian-like H1N1 and novel ressortant H1N2 influenza viruses from pigs in China. Biochem Biophys Res Commun. 2009;386:278–283. doi: 10.1016/j.bbrc.2009.05.056. [DOI] [PubMed] [Google Scholar]
- Zhou NN, Senne DA, Landgraf JS, Swenson SL, Erickson G, Rossow K, Liu L, Yoon K, Krauss S, Webster RG. Genetic reassortment of avian, swine, and human influenza A viruses in American pigs. J Virol. 1999;73:8851–8856. doi: 10.1128/jvi.73.10.8851-8856.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Q, Yang H, Chen W, Cao W, Zhong G, Jiao P, Deng G, Yu K, Yang C, Bu Z, Kawaoka Y, Chen H. A naturally occurring deletion in its NS gene contributes to the attenuation of an H5N1 swine influenza virus in chickens. J Virol. 2008;82:220–228. doi: 10.1128/JVI.00978-07. [DOI] [PMC free article] [PubMed] [Google Scholar]