

Abstract
The zwitterionic lysophospholipid Sphingosine 1-Phosphate (S1P) is a pleiotropic mediator of physiology and pathology. The synthesis, transport, and degradation of S1P are tightly regulated to ensure that S1P is present in the proper concentrations in the proper location. The binding of S1P to five G protein-coupled S1P receptors regulates many physiological systems, particularly the immune and vascular systems. Our understanding of the functions of S1P has been aided by the tractability of the system to both chemical and genetic manipulation. Chemical modulators have been generated to affect most of the known components of S1P biology, including agonists of S1P receptors and inhibitors of enzymes regulating S1P production and degradation. Genetic knockouts and manipulations have been similarly engineered to disrupt the functions of individual S1P receptors or enzymes involved in S1P metabolism. This chapter will focus on the development and utilization of these chemical and genetic tools to explore the complex biology surrounding S1P and its receptors, with particular attention paid to the in vivo findings that these tools have allowed for.
Keywords: Experimental Autoimmune Encephalomyelitis, Sphingosine Kinase, Knockin Mouse, Chemical Tool, Nonselective Agonist
Chemical Tools to Explore S1P Biology
Despite the relatively recent identification of S1P receptors pinpointing S1P as an important player in many physiological systems, a wide variety of chemical tools have been developed to understand the biology of S1P and its receptors. The S1P axis continues to be an area of significant drug discovery efforts. Chemical tools possess several benefits, including the ability to examine acute effects following treatment as opposed to genetic models where precise temporal control is not possible. This section will discuss three broad categories of chemical tools that have been generated and used to explore S1P biology: First, those that affect the normal production or degradation of S1P produced. Second, chemical agonists that activate S1P receptors. Third, chemical antagonists that inactivate S1P receptors. Special attention will be paid to the in vivo effects that these compounds have, and the relation of these compounds to treating human disease.
Chemical Modulators of Physiological S1P Levels
S1P levels are precisely controlled both in circulation, where S1P is present in high-nanomolar concentrations (Hla 2004), and in peripheral tissues, where S1P levels are significantly lower (Schwab et al. 2005) through the coordinated actions of sphingosine kinases, which produce S1P, S1P transporters, which export S1P into the extracellular environment, and S1P phosphatases and lyase, which degrade S1P. Two intracellular sphingosine kinases Sphingosine kinases, SphK1 and SphK2, act to phosphorylate the hydroxyl group of sphingosine to produce S1P. While S1P can act upon intracellular targets, its actions on S1P receptors requires transport to the extracellular environment by one or more S1P transporters, including Spns2 (Kawahara et al. 2009; Fukuhara et al. 2012; Mendoza et al. 2012; Kohama et al. 1998; Liu et al. 2000). S1P is degraded by either reversible dephosphorylation by two S1P-specific phosphatases and three nonspecific lipit phosphate phosphatases (Kai et al. 1997; Roberts et al. 1998; Mandala 2001) or by irreversible cleavage at the C2–3 carbon bond by S1P lyase (Zhou and Saba 1998). Chemical tools have been generated to affect several components of this pathway, and others remain possible targets for development.
Chemical modulation of sphingosine kinases began with the discovery that the sphingosine analogs d-, l-, and dl-threo-dihydrosphingosine and N, N-dimethylsphingosine inhibited the activity of sphingosine kinase in human platelets (Buehrer and Bell 1992; Yatomi et al. 1995). These sphingosine derivatives, though potent, exhibit significant nonselectivity, particularly the inhibition of protein kinase C (Merrill et al. 1989; Khan et al. 1990). Numerous other sphingosine analogs, including FTY720, a S1P receptor prodrug used clinically for the treatment of relapsing-remitting multiple sclerosis, also inhibit sphingosine kinases (Tonelli et al. 2010). In addition to binding competitively, several sphingosine kinase inhibitors also induce proteasomal degradation after binding, providing additional inhibition of the generation of S1P (Tonelli et al. 2010; Lim et al. 2011). Continued efforts have generated nanomolar potency, isoform-selective antagonists of both SphK1 (Paugh et al. 2008; Kennedy et al. 2011) and SphK2 (French et al. 2010).
Sphingosine kinase inhibitors have been investigated as potential treatments for a variety of diseases, particularly inflammatory disorders (Snider et al. 2010) and cancer (Maceyka et al. 2012). The non-S1P-like sphingosine kinase inhibitor ABC747080 was found to reduce inflammation and tissue S1P concentrations in an acute model of inflammatory bowel disease (Maines et al. 2008), while the selective SphK1 inhibitor SK1-i inhibited a mouse model of allergic asthma (Price et al. 2012). Selective antagonism of SphK2 by the antagonist ABC294640 has also demonstrated efficacy in mouse models of inflammatory Crohn’s disease (Maines et al. 2010) and osteoarthritis (Fitzpatrick et al. 2011). Sphingosine kinase inhibitors can also have antitumor effects, as SK1-i was originally found to inhibit leukemia cell proliferation in vitro and inhibit xenograft tumor growth in vivo (Paugh et al. 2008). While efforts have largely focused on the ability of sphingosine kinase inhibitors to reduce proliferation of tumor cells, SK1-i also has been found to inhibit breast cancer tumor growth and metastasis in mice by affecting angiogenesis and lymphangiogenesis (Nagahashi et al. 2012). Inhibition of SphK2 by ABC294640 has demonstrated a similar ability to inhibit tumor progression in hepatocellular carcinoma xenografts (Beljanski et al. 2011). The precise mechanisms by which chemical inhibition of sphingosine kinases can lead to either suppression of inflammation or tumor growth and/or metastasis remain unclear. Lowering S1P levels by inhibiting sphingosine kinases may have many effects, both extracellular through reducing signaling through S1P receptors, and intracellular through undetermined mechanisms (Strub et al. 2010), that may act in coordination to alleviate pathology (Table 1).
Table 1.
Summary of chemical and genetic tools for studying S1P biology
| Chemical | Genetic | |||||||
|---|---|---|---|---|---|---|---|---|
| In vivo agonist | In vivo antagonist | Knockout | Other | |||||
| Selective | Nonselective | Selective | Nonselective | Constitutive | Conditional | |||
| Genes affecting S1P synthesis, degradation, and transport | Sphingosine kinase 1 | ✓ | ✓ | ✓ | ✓ | Transgenic overexpression | ||
| Sphingosine kinase 2 | ✓ | ✓ | ✓ | ✓ | ||||
| S1P Lyase | ✓ | |||||||
| LPP3 | ✓ | ✓ | ||||||
| SPNS2 | ✓ | ✓ | ||||||
| S1P Receptors | S1P1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | Fluorescent and non-phosphorylable knockins, transgenic overexpression |
| S1P2 | ✓ | ✓ | ✓ | |||||
| S1P3 | ✓ | ✓ | ✓ | ✓ | Fluorescent knockin | |||
| S1P4 | ✓ | ✓ | ||||||
| S1P5 | ✓ | ✓ | ✓ | |||||
While inhibitors of sphingosine kinases effectively reduce the abundance of S1P, inhibitors of the enzymes that normally degrade S1P have also been designed to raise the concentrations of S1P present in both tissues and circulation. One such inhibitor is tetrahydroxybutylimidazole, or THI, a caramel food coloring that at high concentrations acts as a S1P lyase inhibitor, elevating the concentration of S1P 100-fold in lymphoid tissues and leading to the sequestration of lymphocytes from blood and lymph, similar to treatment with either S1P1 agonists or antagonists (Schwab et al. 2005). THI recently has also been shown to acutely alleviate cardiac ischemia-reperfusion injury concomitant with an increase in both tissue and plasma concentrations of S1P (Bandhuvula et al. 2011). S1P lyase inhibition has also been pursued clinically for the treatment of rheumatoid arthritis, presumably in part by sequestering lymphocytes from circulation (Bagdanoff et al. 2010).
While not a small molecule, one additional tool that has been developed to modulate the availability of S1P is sphingomab, a monoclonal antibody directed against S1P itself which is being developed as an anticancer drug. In animal models, treating mice with sphingomab can inhibit both growth and metastasis of tumors (Visentin et al. 2006; Ponnusamy et al. 2012). Additionally, treatment of mice with sphingomab can cause lymphopenia despite increasing blood and lymphatic concentrations of S1P, much of which is bound to sphingomab (Sensken et al. 2011). The continued development of tools that can increase, decrease, or bind S1P that is normally produced will help dissect out the many roles that S1P can play in physiology and may eventually lead to effective treatments for human diseases.
S1P Receptor Agonists
S1P receptor agonists have been heavily investigated following the discovery that FTY720 (fingolimod, Gilenya), an immunosuppressive derivative of the fungal metabolite myriocin, was phosphorylated in vivo to generate a highly potent, nonselective agonist of four of five S1P receptors (Mandala et al. 2002). In the past decade a remarkably wide array of compounds that activate one or more S1P receptors have been developed for use as chemical tools to investigate S1P receptor function, with FTY720 already progressing to clinical use.
Nonselective S1P Receptor Agonists
As previously mentioned, the phosphorylated form of FTY720, FTY720-P, is a highly potent yet nonselective agonist of S1P1, S1P3, S1P4, and S1P5 with EC50 values in the low nanomolar range (Brinkmann et al. 2002). FTY720 is an analog of sphingosine and is phosphorylated in vivo exclusively by Sphk2 to generate FTY720-P (Paugh et al. 2003; Kharel et al. 2005), which can be exported into the extracellular environment by Spns2 (Hisano et al. 2011). AAL-R, a close relative of FTY720 differing only in the lack of a hydroxyl group, is also efficiently phosphorylated by Sphk2 to generate AFD-R, a nonselective agonist of S1P receptors (Jary et al. 2010). Treatment with nonselective S1P receptor agonists leads to rapid, sustained sequestration of both T and B cells from the blood and the lymph and causes short-lasting bradycardia in both mice and humans (Luo et al. 1999; Budde et al. 2002; Sanna et al. 2004). The immunosuppressive properties of FTY720 and other nonselective S1P receptor agonists led to investigation into their potential as a therapy for both transplant rejection (Suzuki et al. 1996; Brinkmann et al. 2001; Shimizu et al. 2005) and autoimmune disorders, particularly multiple sclerosis (Fujino et al. 2003; Thomson 2006). While it has not progressed to clinical use for transplant rejection, FTY720 has been approved as a treatment for relapsing-remitting multiple sclerosis in humans.
While it may contribute to its clinical efficacy, the lack of selectivity of FTY720 makes it a less than ideal tool for dissecting the roles of individual S1P receptors play in physiology, FTY720 has been very useful in helping elucidate the differences in downstream signaling and receptor fate across S1P receptors. Treatment of cells expressing different S1P receptors with FTY720 leads to rapid and sustained internalization and degradation of S1P1, even after washout, but does not lead to the degradation of S1P3 or S1P4 (Graler and Goetzl 2004; Mullershausen et al. 2009). Given the high potency of FTY720-P across S1P receptors, most of the focus of generating new chemical tools to study S1P receptors has focused on improving selectivity, which has allowed for elucidating the role that individual S1P receptors can play in physiology and pathology.
Selective S1P1 Agonists
S1P1 has been by far the most studied S1P receptor both chemically and genetically due to its critical role in lymphocyte recirculation. S1P1-selective agonists, even from direct screening hits such as SEW2871, have generally been able to achieve strong potency, with EC50 values at least in the nanomolar range, and have been generated with a wide variety of chemical scaffolds (Zhang et al. 2009; Meng et al. 2012; Nakamura et al. 2012; Reed et al. 2012), of which several shown in Fig. 1. It is readily apparent that many S1P1 agonists exhibit long hydrophobic cores along with a polar or charged headgroup. These structures resemble the amphipathic nature of S1P, which consists of a long acyl chain and a zwitterionic headgroup consisting of a positively charged amine and a negatively charged phosphate. These groups in S1P interact with charged arginine and glutamate residues (Arg120 and Glu121) that are positioned within the binding pocket close to the extracellular side (Parrill et al. 2000). Despite the lack of a charged headgroup, SEW2871 still relies on interaction with Arg120 to activate S1P1 (Jo et al. 2005). One relatively unique agonist is CYM-5442, which does not possess a charged or highly polar headgroup, instead having a hydroxyl group. CYM-5442 has been shown to not require interactions with Arg120 or Glu121 of S1P1, and in fact binds S1P1 in a different manner, instead relying on interaction with a critical tryptophan residue (W269) deep in the S1P1 pocket for its potency (Gonzalez-Cabrera et al. 2008; Hanson et al. 2012). Modifications of CYM-5442 have led to a class of arylpropionic acids that activate S1P1 with picomolar EC50 values (Cahalan et al. 2011; Teijaro et al. 2011). These S1P1 agonists can vary significantly in their potency, pharmacokinetic profiles, and their effects on S1P1 following binding. Some S1P1 agonists, like SEW2871 or the native ligand S1P, cause S1P1 to be internalized from the cell surface but then recycled back to the surface, whereas others, such as the FTY720-P homolog AFD-R and CYM-5442, cause internalization, polyubiquitination, and degradation via the proteasome (Gonzalez-Cabrera et al. 2007).
Fig. 1.
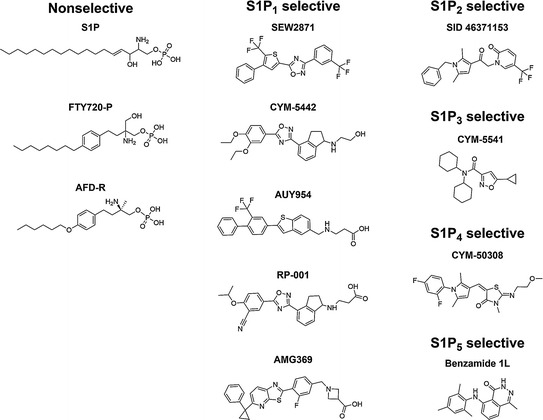
Chemical structures of S1P receptor agonists
The ability of FTY720 to sequester lymphocytes from the blood and the lymph relies on its activity on S1P1, as S1P1-selective agonists lead to similar sequestration of lymphocytes both in mice (Sanna et al. 2004) and in humans. Treatment of mice with SEW2871 causes the arrest of lymphocytes close to the lymph node medullary sinus but does not affect the velocity of lymphocytes found in the cortex (Wei et al. 2005; Sanna et al. 2006). While the receptor mediating lymphocyte sequestration induced by FTY720, S1P1, is the same between mice and humans, the receptors mediating bradycardia elicited by FTY720 differ significantly. Bradycardia caused by FTY720 in mice relies on its actions on S1P3 (Sanna et al. 2004), with S1P1-selective agonists causing negligible bradycardia, whereas S1P1-selective agonists can cause significant bradycardia in humans by activating the G protein-coupled inwardly rectifying potassium (GIRK) channel expressed in myocytes (Gergely et al. 2012).
Since the activation of S1P1 inhibits lymphocyte recirculation, selective S1P1 agonists have been investigated as treatments for autoimmune diseases as potential alternatives to nonselective agonists like FTY720-P. S1P1-selective agonists have demonstrated efficacy in several animal models, including multiple sclerosis (Galicia-Rosas et al. 2012; Gergely et al. 2012; Gonzalez-Cabrera et al. 2012), autoimmune arthritis (Piali et al. 2011), and ulcerative colitis (Sanada et al. 2011). Several S1P1 agonists have progressed into clinical trials focused on multiple sclerosis, with the goal of limiting the off-target effects resulting from treatment with FTY720. However, the possibility exists that the actions of FTY720 on other S1P receptors or other targets such as sphingosine kinases may aid in its efficacy. Whether these selective drugs can improve outcomes or minimize adverse events when compared to FTY720 remains to be seen.
Recent studies have also found that S1P1 agonists can have compelling effects in inhibiting cytokine production following infection by influenza virus. Local treatment with the nonselective S1P receptor proagonist AAL-R in the lungs of mice prior to infection with influenza virus leads to the suppression of viral specific T cell response and the suppression of cytokine production within the lungs (Marsolais et al. 2008; Marsolais et al. 2009). The suppression of cytokines by AAL-R is of particular interest, as excessive cytokine production has been implicated in the pathogenicity of several “pandemic” influenza strains, including H5N1 “avian” flu and H1N1 strains from both the 1918 pandemic and the more recent “swine” flu outbreak in 2009 (Arankalle et al. 2010; Lee et al. 2011). Indeed, mice treated with AAL-R exhibited significantly less mortality than untreated mice following infection with a 2009 H1N1 influenza strain, and this reduction in lethality was synergized by the administration of the clinically used neuraminidase inhibitor oseltamivir (Walsh et al. 2011). While the mechanisms by which AAL-R inhibits cytokine production aren’t fully understood, it appears that S1P1 expressed on endothelial cells plays a critical role in this suppression of cytokine production as well as inhibiting the infiltration of innate immune cells into the lung (Teijaro et al. 2011). The ability of S1P1 agonists to suppress cytokine production may be of much broader significance, as excessive cytokine production has also been implicated in other viral infections such as SARS (Huang et al. 2005; Nagata et al. 2008) or bacterial infections such as pneumococcal pneumonia (Fernandez-Serrano et al. 2003).
Selective S1P2, S1P3, S1P4, and S1P5 Agonists
Unlike S1P1, very few selective agonists have been developed against S1P2 (Rosen 2007), S1P3 (Jo et al. 2012), S1P4 (Urbano et al. 2011) or S1P5 (Mattes et al. 2010). In fact, no in vivo activity of any selective agonist of these receptors has been established to date, and only for a selective agonist of S1P5, which increased the number of mature oligodendrocytes obtained in vitro from neonatal rat cortices, has any physiological effect been determined in vitro (Mattes et al. 2010). While the abundance of S1P1-selective agonists in comparison to the dearth of selective agonists of other S1P receptors is largely due to the high-clinical relevance of S1P1, there may be additional factors at play, including the difficulty in actually discovering and generating compounds that fit specifically into the binding pocket of those receptors. For instance, S1P3 is thought to have a significantly smaller binding pocket than S1P1, which may constrain the amount of interaction between the agonist and the receptor; additionally compounds found in high-throughput screening libraries are more similar to identified S1P1 agonists than they are to S1P3 agonists (Schurer et al. 2008). The only published S1P3-selective agonist to date is in fact an allosteric agonist, CYM-5541, that does not compete with S1P binding. Such allosteric agonists may provide an alternative route to discovering selective agonists to the S1P receptors, relying predominantly on interactions deep within the binding pocket rather than interactions with charged residues located near the top of the binding pocket.
S1P Receptor Antagonists
While FTY720-P was initially characterized as a potent agonist of S1P receptors, it was soon determined that FTY720-P and other agonists could cause the internalization of S1P1 from the surface of the cell. This ability of FTY720-P led many to develop antagonists against S1P receptors in part to determine what the effect of in vivo blockade of normal S1P receptor signaling could have, and also to determine whether treatment with S1P receptor antagonists could replicate the effects seen following treatment with FTY720 or other S1P receptor agonists.
S1P1 Antagonists
The first published S1P1 antagonists were discovered following the finding that moving the substituents around the phenyl ring in FTY720-P from para- to ortho- caused a switch from agonism to antagonism on S1P1 (Davis et al. 2005), as shown in Fig. 2. The resulting compound, VPC23019, was a dual S1P1/S1P3 antagonist, though it exhibits significantly greater inhibition of S1P1 compare to S1P3. Another early S1P1 antagonist exhibiting a similar ortho- substitution, W123, was found to be selective for S1P1, with no activity on S1P3, and could reverse the arrest of lymphocytes in the medulla of explanted lymph nodes (Wei et al. 2005). However, both of these initial antagonists were unsuitable for in vivo work due to their instability when administered to mice.
Fig. 2.
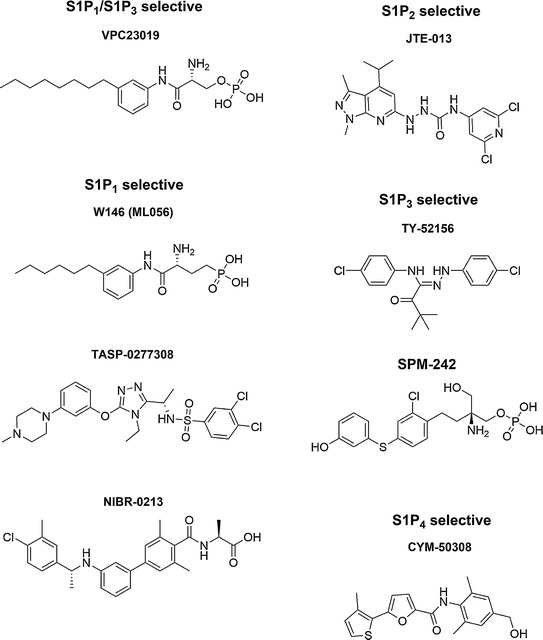
Chemical structures of S1P receptor antagonists
Optimization of these ortho-substituted FTY720-P analogs eventually led to the replacement of phosphate groups for a phosphonate group, yielding a chiral antagonist, W146, or ML056, that could act in vivo. While the R- enantiomer was found to be an antagonist, the S- enantiomer exhibited no activity on S1P1. When administered in vivo, W146 did not appear to cause lymphocyte sequestration and could, in fact, reverse lymphocyte sequestration induced by the S1P1 selective agonist SEW2871 when examined 4 h after treatment (Sanna et al. 2006). VPC44116, which is the equivalent phosphonate of VPC23019, had much the same effect in vivo, not causing lymphocyte sequestration in vivo but reversing the sequestration induced by another S1P1 agonist, VPC44152 (Foss et al. 2007). W146 also induce significant vascular leakage in the lung and exacerbated VEGF-induced leakage in the skin, and recently was shown to stabilize the binding pocket of S1P1 allowing for the crystallization and structural determination of S1P1 joined to T4 lysozyme (Hanson et al. 2012). The lack of lymphocyte sequestration and the reversal of agonist-induced sequestration originally observed with S1P1 antagonists led to the hypothesis that agonists acted not as functional antagonists of S1P1 expressed on lymphocytes, but as direct agonists of S1P1 expressed on endothelial cells leading to the tightening of cell–cell junctions and the inhibition of lymphocyte egress.
More recent work has demonstrated that selective S1P1 antagonists, including W146 if examined at earlier timepoints, can in fact cause lymphocyte sequestration by themselves (Tarrason et al. 2011). Additional newer antagonists exhibiting higher potency or improved pharmacokinetic properties can induce lymphocyte sequestration that can be sustained for many hours, including TASP0277308 (Fujii et al. 2012) and a series of biaryl benzylamine derivatives (Angst et al. 2010). These antagonists are structurally different from the previous S1P-like antagonists like W146 and VPC44116 and can upregulate S1P1 on lymphocytes (Cahalan et al. 2013; Fujii et al. 2012), opposite of the effect seen with S1P1 agonists. Furthermore, the ability to sequester lymphocytes by these antagonists has been accompanied by efficacy in animal models of allograft rejection (Angst et al. 2012), arthritis (Fujii et al. 2012), and multiple sclerosis(Quancard et al. 2012; Cahalan et al. 2013).
The ability of S1P-like antagonists such as W146 to reverse lymphocyte sequestration by S1P1 agonists at some timepoints while causing lymphocyte sequestration at earlier timepoints raises some interesting possibilities regarding the competition within the binding pocket and downstream signaling, which may be an area for further study. Future work should also determine whether antagonists may be useful as potential therapeutics for autoimmune disease, or whether they elicit significantly worse adverse events, such as pulmonary or peripheral edema, compared to S1P1 agonists. Additionally, the development of longer-lasting antagonists may allow for extended studies of other physiological and pathological conditions.
S1P2, S1P3, S1P4, and S1P5 Antagonists
Similar to what has occurred with S1P receptor agonists, studies using S1P receptor antagonists have predominantly focused on S1P1, with fewer antagonists against S1P2, S1P3, S1P4, and S1P5. A specific antagonist to S1P2, JTE-013, has been widely used to study the role of S1P2 both in vitro and in vivo, particularly the role that S1P2 can play in vascular physiology. Treatment of vascular endothelial cells with JTE-013 can improve their barrier integrity in vitro (Sanchez et al. 2007) and inhibit S1P-induced vasoconstriction in excised arteries (Kono et al. 2007). Some questions have been raised about the specificity of JTE-013, as it may have effects independent of S1P2 (Salomone and Waeber 2011; Li et al. 2012).
Two selective S1P3 antagonists have been described: TY-52156, which could inhibit S1P-dependent coronary flow within isolated rat hearts in vitro and inhibit FTY720-induced bradycardia in vivo (Murakami et al. 2010), and SPM-242, which is able to compete both with the native agonist S1P and with an allosteric agonist to S1P3, thus making it a “bitopic” antagonist, although no in vivo actions of this antagonist have been established to date (Jo et al. 2012). Additionally an anti-S1P3 antibody has been described that blocks the activation of S1P3 (Harris et al. 2012). This antibody could block lethality in mice caused by systemic LPS treatment and inhibit the growth of breast cancer xenografts, providing potential avenues for further examination of the roles of S1P3 in two distinct pathological conditions. S1P4 antagonists have been described in vitro (Guerrero et al. 2011), but with no relevant physiological data published to date. With regard to S1P5, no data has been published describing the inhibition of S1P5 by selective antagonists, though a patent application was filed describing scaffolds that may act as either agonists or antagonists of S1P5 (Harris et al. 2010).
Genetic Tools to Explore S1P Biology
In addition to the numerous chemical tools that have been generated to modulate the levels of S1P produced and the activity of the five S1P receptors, genetically engineered mice have been generated to examine the complex physiological processes controlled by S1P. Genetic knockouts have the benefit of completely eliminating the gene of interest, with potentially fewer off-target effects, though the potential for compensation by other genes remain possible. Additionally, cell-type specific deletion or overexpression can be achieved by genetic modification, something not able to be done by chemical treatment. Some significant drawbacks exist, such as the difficulty to control precisely when the gene of interest is inhibited or reversing such inhibition. This section will review briefly three general types of mouse models that have been generated; significantly more detail on several of these mouse models will be discussed in later chapters. First, mice in which the generation, degradation, or transport of S1P has been modified. Second, mice where the expression of S1P receptors have been deleted, either globally or conditionally. Lastly, mice where a tagged or mutated S1P receptor replaces the endogenous S1P receptor.
Genetic Modification of Physiological S1P Production, Degradation, and Transport
Similar to the variety of chemical modulators of S1P levels in vivo, a number of genetic mouse models have been generated lacking or overexpressing the genes that are involved in regulating the physiological levels of S1P.
Sphingosine Kinase Deficient Mice
Genetic knockouts to both the sphingosine kinases, SphK1 and SphK2, alone and in combination have been generated and have revealed many important roles for the generation of S1P in physiology. While they play many unique roles, they can also act somewhat redundantly, as either SphK1-deficient or SphK2-deficient mice are viable and fertile (Allende et al. 2004; Kharel et al. 2005; Mizugishi et al. 2005). SphK1-deficient mice display significantly reduced SphK activity in some tissues, particularly the spleen, but only in the serum is a significant reduction in S1P observed. SphK2-deficient mice also display only a partial reduction in circulating S1P, but display a complete inability to phosphorylate FTY720. However, mice that are deficient in both SphK1 and SphK2 are rendered completely unable to produce S1P and begin to exhibit severe vascular hemorrhage in utero, with no embryos surviving past E13.5 (Mizugishi et al. 2005).
The lethality of SphK1/SphK2 double knockout mice has been circumvented using mice where SphK2 is constitutively deleted while SphK1 is deleted shortly after birth in many tissues by using inducible Mx1-Cre driven excision of SphK1 (Pappu et al. 2007). Such “S1P-less” mice are viable and lack detectable amounts of circulating S1P in either the blood or the lymph. The sources of the S1P found in the blood and in the lymph are distinct, with S1P in the blood being produced by hematopoieitic cells, largely erythrocytes, while S1P in the lymph is produced in large part by lymphatic endothelial cells (Pham et al. 2010). S1P-less mice also display significant basal vascular leakage in the lung, similar to what is observed in mice treated with S1P1 antagonists, and also display greatly increased sensitivity to passive systemic anaphylaxis, a sensitivity that could be largely reversed by agonism of S1P1 (Camerer et al. 2009).
Despite playing somewhat redundant roles in the production of circulating S1P, deletion of either of the sphingosine kinases in mice can have effects by themselves. SphK1 can play an important role in inflammation induced by TNF. TNF leads to the activation SphK1, which can eventually lead to production of prostaglandin E2 by COX2 (Pettus et al. 2003). Mice deficient in SphK1 display significantly reduced pathology in several experimental inflammatory pathologies, including dextran sulfate sodium-induced colitis (Snider et al. 2009) and arthritis induced by transgenic expression of TNF (Baker et al. 2010), in keeping with the findings mentioned earlier utilizing the selective sphingosine kinase inhibitor SK1-i. These findings would suggest that inhibiting SphK1 might be a good treatment for these diseases, but this is complicated by the finding that SphK1−/− mice display significantly increased pulmonary edema in response to either inflammatory LPS or PAR-1 agonist peptide treatment, potentially by lowering the amount of S1P that is able to activate S1P1, which normally acts to tighten vessel permeability.
Recent work utilizing SphK2-deficient mice has also suggested a protective role of SphK2 following ischemia. In a model of cerebral ischemia, mice deficient in SphK2 exhibited significantly larger ischemic lesion sizes 24 h following a 2 h occlusion of the middle cerebral artery (Pfeilschifter et al. 2011). FTY720 treatment was found to have a protective effect following ischemic injury, but this protection was abolished in SphK2-deficient mice, pointing toward this protection as a function of FTY720-P. Hearts from SphK2-deficient mice are also sensitized to ischemia/reoxygenation injury within the heart, exhibiting significantly larger infarct sizes (Vessey et al. 2011). Hearts from SphK2-deficient mice were also resistant to protection usually offered by ischemic preconditioning, but the addition of exogenous S1P to SphK2-deficient hearts could offer protection.
S1P Lyase and Phosphatase Deficient Mice
On the opposing side of S1P metabolism, mice deficient in some of the enzymes that normally degrade S1P have been generated to study what role the proper degradation of S1P can play. Mice deficient in S1P lyase, encoded by the gene Sgpl1, display significantly reduced viability, size, and weight, and all mice were dead by 8 weeks of age (Schmahl et al. 2007). When examined at early ages, sphingolipid levels of not only S1P, but of ceramide and sphingomyelin, were elevated in plasma, and levels of dihydrosphingosine, ceramide, sphingosine, dihydro-S1P, and S1P were elevated within the liver (Bektas et al. 2010). These mice also display higher levels of cholesterol in the serum and exhibited significant differences in the metabolism of many other lipids in the liver, pointing toward an important role for S1P degradation in total lipid metabolism. The broad elevation of several other sphingolipids points to a potential increase in the recycling of S1P by S1P phosphatases, which may provide more sphingosine that can act as a substrate for ceramide synthase to in turn produce more ceramide (Hagen-Euteneuer et al. 2012). Mice deficient in S1P lyase have significant changes in immune cell development and function, as they have reduced T and B cell numbers in the blood and in the spleen while exhibiting elevated numbers of neutrophils and monocytes in the blood and elevated proinflammatory cytokines in the serum (Allende et al. 2011). The elevated numbers of neutrophils and increase in proinflammatory cytokines could be partially rescued by also deleting S1P4 but not S1P1.
As opposed to the irreversible cleavage of S1P caused by S1P lyase, several phosphatases, either S1P-specific S1P phosphatases (SPPs) or nonspecific lipid phosphate phosphatases (LPPs), can reversibly dephosphorylate S1P to yield sphingosine. While mice lacking either of the specific SPPs have not been described to date, mice lacking each of the LPPs have been generated. Mice deficient in LPP1 have been described to have deficient clearance of lysophosphatidic acid, but were otherwise phenotypically normal (Tomsig et al. 2009). Likewise, mice deficient in LPP2 were found to be viable and did not display any significant phenotype to date (Zhang et al. 2000). Mice deficient in LPP3, in contrast, die in utero prior to E10.5 due to both gastrulation deficits and inefficient vascularization of the yolk sac (Escalante-Alcalde et al. 2003). Conditional deletion of LPP3 in specialized astrocytes known as Bergmann glia can lead to alterations of S1P metabolism within the cerebellum, resulting in motor coordination deficits (Lopez-Juarez et al. 2011). In keeping with the important role for S1P in lymphocyte development, inducible deletion of LPP3 leads to inefficient egress of mature thymocytes into the periphery (Breart et al. 2011). Deletion of LPP3 on either endothelial cells or on epithelial cells leads to a similar inefficiency of egress, concomitant with a downregulation of CD69 that would be expected to be a result of exposure of mature thymocytes to higher concentrations of S1P. LPP3 deletion also has effects in smooth muscle cells, as mice where LPP3 has been deleted in smooth muscle exhibit higher inflammation after vascular injury (Panchatcharam et al. 2013).
S1P Transporter Deficient Mice
Since S1P is produced largely inside the cell, it must be transported to the outside of the cell in order to act on extracellular targets such as S1P receptors. S1P is thought to be transported in part by ABC type transporters such as ABCA1 and ABCC1 (Mitra et al. 2006; Sato et al. 2007), but mice deficient in these transporters display no changes in circulating S1P. Recently, a separate S1P transporter, Spns2, was identified in a zebrafish mutant that displayed impaired migration of myocardial precursors, similar to the deletion of the zebrafish homolog of S1P2 (Kawahara et al. 2009). Several groups have generated mice lacking Spns2, identifying that it is indeed an S1P transporter that when deleted can lead to deficiencies in thymocyte egress and B cell development (Fukuhara et al. 2012; Hisano et al. 2012; Mendoza et al. 2012; Nagahashi et al. 2012; Nijnik et al. 2012). Spns2 expressed on endothelial cells, not erythrocytes, is critical for the efficient egress of mature thymocytes. The presence of significant concentrations of circulating S1P even in the absence of Spns2 points toward some level of redundancy, either through the functions of the ABC type transporters or another S1P transporter that has yet to be determined.
Mice Deficient in S1P Receptors
S1P1 Deficient Mice
S1P1 was originally identified as an endothelial differentiation gene, and thus was given the gene name Edg1. Mice deficient in S1P1 die between E12.5 and E14.5 due to vascular hemorrhage and an inability of smooth muscles to migrate to enclose blood vessels (Liu et al. 2000). Mice that lack S1P1 solely on endothelial cells exhibit a similar phenotype, pointing to the critical role that S1P1 plays in vascular development (Allende et al. 2003). S1P1 expressed on endothelial cells can also play an important role in the development of a mature vascular network, particularly the inhibition of excess sprouting. Abnormal sprouting of vascular networks is seen in the embryos where S1P1 has been deleted only in endothelial cells (Shoham et al. 2012), and a similar increase in branching is seen in mice where S1P1 is deleted inducibly shortly after birth, either in all tissues, or solely in endothelial cells (Jung et al. 2012). Interestingly, S1P1 expression seems to be highest in areas of vasculature with fluid flow, and S1P1 may function as a sensor of shear flow, providing another way that S1P1 can regulate vascular function (Jung et al. 2012). The roles of S1P1 specifically within the vascular system will be discussed in significantly more detail in a later chapter.
Following the finding that FTY720-P acted as an agonist to S1P receptors, many became interested in elucidating the functions of different S1P receptors in lymphocyte function. Deletion of S1P1 in developing T cells, either by conditional knockout or bone marrow chimeric approaches, leads to the retention of mature CD4+ and CD8+ T cells within the thymus, leaving the blood and lymph depleted of lymphocytes (Allende et al. 2004; Matloubian et al. 2004). Mice in which B cells are deficient S1P1 have also been generated, and such mice display impaired egress of immature B cells from the bone marrow into the blood as well as a decrease in splenic B cell numbers (Allende et al. 2010). A more detailed examination of the roles of lymphocytic S1P1, particularly the functions S1P1 can have in lymphocyte recirculation and the positioning of lymphocytes within the lymphoid tissues, will also follow in a later chapter.
S1P1, in addition to its critical roles in vascular and lymphocyte physiology, plays important roles within the central nervous system. S1P1 is highly expressed within the brain and the spinal cord, across multiple cell types including neurons and astrocytes, and FTY720-P and other S1P1 agonists can penetrate efficiently into the CNS, giving rise to the potential for actions of these agonists directly on CNS tissue. Mice in which S1P1 has been deleted in astrocytes are refractory to developing experimental autoimmune encephalomyelitis (EAE), a mouse model of multiple sclerosis, whereas mice in which S1P1 was deleted only in neurons develop EAE similar to wild-type mice (Choi et al. 2011). Interestingly, whereas wild-type mice with EAE displayed improvements in clinical scores following treatment with FTY720, mice with EAE that lacked S1P1 on astrocytes did not display any additional improvement in their symptoms following FTY720 treatment. S1P1 may also play a role in inflammatory pain, as S1P and the S1P-selective agonist SEW2871 can cause thermal hyperalgesia following intraplanar injection. This hyperalgesia is dependent in part on S1P1 expressed on nociceptive neurons, as mice where S1P1 has been deleted in Nav1.8-expressing neurons have reduced hyperalgesia following S1P administration (Mair et al. 2011).
S1P2 Deficient Mice
Like S1P1, S1P2 has a critical role in vascular physiology. Unlike mice lacking S1P1, mice lacking S1P2 are viable (MacLennan et al. 2001; Ishii et al. 2002; Kono et al. 2004); however, S1P2 deficient mice are deaf, potentially as a result of malformed stria vascularis in the inner ear or defects in hair cells (Herr et al. 2007; Kono et al. 2007). S1P2 deficient mice in some genetic backgrounds can also exhibit spontaneous seizures (MacLennan et al. 2001). S1P2 may play somewhat overlapping roles with both S1P1 and S1P3, as mice lacking both S1P1 and S1P2 display a more severe embryonic lethal phenotype than mice lacking S1P1 alone while mice lacking S1P2 and S1P3 display significant perinatal lethality (Ishii et al. 2002; Kono et al. 2004). Mice deficient in both S1P2 and S1P3 also display significantly larger myocardial infarct sizes following ischemia/reperfusion injury, but mice lacking either display no difference, once again pointing to the significant overlap in function between the two receptors (Means et al. 2007). Another role of S1P2 within the vascular system is seen when examining neovascularization within the eye, as S1P2 deficient mice displayed significantly less intravitreal angiogenesis following ischemia (Skoura et al. 2007). Conditional knockout mouse lines for S1P2 have yet to be reported, but could potentially determine which cell types are responsible for the phenotypes seen in mice completely lacking S1P2.
S1P3 Deficient Mice
Viable S1P3 deficient mice have also been generated which display no significant abnormalities, but display marginal deficits in reproduction when deficient mice intercrossed (Ishii et al. 2001). S1P3 deficient mice have been used to show roles for S1P3 in cardiac physiology, as S1P3 deficient mice display resistance to bradycardia induced by the nonselective proagonist AAL-R (Sanna et al. 2004) and are resistant to developing cardiac fibrosis observed when SphK1 is transgenically overexpressed (Takuwa et al. 2010). S1P3 deficient mice additionally exhibit a lack of vasorelaxation caused by S1P and show diminished vasorelaxation induced by injection of high-density lipoprotein (HDL), presumably as a result of the high amounts of S1P found within HDL (Nofer et al. 2004).
S1P3 has also been studied for its role in the immune system, particularly marginal zone (MZ) B cells and dendritic cells. While no conditional knockout mouse strain has been described, reciprocal bone marrow chimera experiments and adoptive transfer experiments have been used with great success between wild-type and S1P3 deficient mice. Such approaches have suggested that S1P3 expressed on MZ B cells helps properly position them within the white pulp, and that migration of MZ B cells to S1P is mediated in large part by S1P3 (Cinamon et al. 2004; Cinamon et al. 2008). S1P3 expressed on dendritic cells can regulate their migration to S1P (Maeda et al. 2007) and mediates inflammation induced through various pathways. First, S1P3 deficient mice display significantly less systemic inflammatory cytokine production following LPS treatment than do wild-type mice, a reduction that was reversed by adoptively transferring wild-type dendritic cells (Niessen et al. 2008). S1P3 deficient mice also displayed improved survival compared to wild-type mice following LPS treatment and delayed lethality in a model of cecal ligation and puncture that could be reversed by adding in exogenous wild-type dendritic cells. S1P3 deficient mice are also resistant to kidney ischemia reperfusion injury, and this resistance is dependent on the deletion of S1P3 on dendritic cells, as injection of wild-type dendritic cells into S1P3 deficient mice prior to injury could lead to a reversal in the protection granted by S1P3 deficient mice (Bajwa et al. 2012).
S1P4 Deficient Mice
S1P4 was the last S1P receptor to be described and likewise was the last to have any in vivo phenotype ascribed to its deletion. Like S1P1 and S1P3, S1P4 plays an important role in several different cell types in the immune system. S1P4 knockout mice display impaired terminal differentiation of megakaryocytes, which leads to a deficit in platelet repopulation following administration of an antiplatelet antibody but not any observable differences in platelet counts under normal conditions (Golfier et al. 2010). S1P4 deficient mice also may help regulate neutrophil inflammation that can be caused by high levels of S1P and other sphingolipids in S1P lyase deficient mice (Allende et al. 2011). S1P4 is also expressed on dendritic cells and may affect both their migration and their activation of T cells, as dendritic cells in mice lacking S1P4 display accelerated in vivo migration to draining lymph nodes while S1P4 deficient dendritic cells may have a reduced ability to cause T cells to secrete IL-17 (Schulze et al. 2011). No conditional S1P4 mice have been described to date.
S1P5 Deficient Mice
S1P5 deficient mice, like mice lacking either S1P3 or S1P4, are viable and fertile. S1P5 knockout mice were first described in the context of the function of S1P5 on oligodendrocytes, as S1P5 deficient oligodendrocytes did not exhibit process retraction following treatment with S1P (Jaillard et al. 2005). The most noted function of S1P5 has also been found in the immune system, particularly its functions in natural killer (NK) cell migration, similar to the way other S1P receptors can regulate the migration of other types of immune cells. S1P5 deficient mice display significant reductions in NK cells found within the blood and the spleen, but increased numbers in the bone marrow and lymph nodes, and NK cells lacking S1P5 do not migrate to S1P as do wild-type NK cells (Walzer et al. 2007; Jenne et al. 2009; Mayol et al. 2011). Like all the other S1P receptors other than S1P1, no S1P5 conditional knockout mice have yet been described.
Knockin Mice Expressing Tagged or Mutated S1P Receptors
Fluorescence-Tagged Knockin Mice
While many genetically modified mice have been generated to delete S1P receptors in cells of interest, recent work has generated mice in which a fluorescence-tagged version of an individual S1P receptor replaces that S1P receptor. These mice allow for the clarification of the expression of S1P receptors in vivo by both flow cytometry and classic biochemistry, and can also be used in intravital two-photon microscopy to directly visualize changes in the subcellular localization of S1P receptors in real-time. Mice in which S1P1 fused to the fluorescent protein eGFP have been used to study how treatment with different S1P receptor agonists and antagonists can affect the expression of S1P1 on lymphocytes (Cahalan et al. 2011), endothelial cells, and within the central nervous system (Gonzalez-Cabrera et al. 2012), have been used to visualize the differential labeling of distinct vascular types with fluorescent particles (Sarkisyan et al. 2012), and have been critical in the identification of endothelial cells as important regulators of cytokine production following influenza infection (Teijaro et al. 2011). A similar approach has also been taken in generating viable mice that express S1P3 fused to the fluorescent protein mCherry (Hugh Rosen, personal communication). While many potential avenues of research may derive from native fluorescence of the fusion proteins, many may also come from the ability to immunoprecipitate the receptors using antibodies against these fluorescent proteins in order to determine posttranslational modifications such as phosphorylation and polyubiquitination of the receptors, as well as potentially determining associated proteins.
S1P1 Mutant Knockin Mice
As mentioned previously, S1P1 becomes rapidly internalized following the binding of agonists. As lymphocyte sequestration induced by S1P1 agonists is thought to occur due to this internalization, the mechanisms by which this internalization occurs have been studied in detail. The internalization of S1P1 is mediated in part by the phosphorylation of a number of serine and threonine residues located in C-terminal tail by G protein-coupled receptor kinases, or GRKs, leading to β-arrestin binding and clathrin-mediated endocytosis (Oo et al. 2007). Recently, knockin mice have been generated where native S1P1 has been replaced by S1P1 in which five of these C-terminal serines have been mutated to alanines (S1P1-S5A), thus preventing the internalization of S1P1 (Thangada et al. 2010). While S1P1-S5A expressed on lymphocytes could still be internalized following either agonist treatment or exposure to blood, S1P1-S5A exhibited delayed internalization in response to S1P1 agonist treatment compared to wild-type S1P1. This resistance to internalization paralleled a delay in lymphocyte sequestration in response to either S1P1 agonism or S1P lyase inhibition. S1P1-S5A expressing lymphocytes show increased migration toward high concentrations of S1P, suggesting that higher concentrations of S1P may normally internalize S1P, preventing migration. A similar knockin mouse in which a threonine and two serines in the C-terminal tail have been replaced by alanines (S1P1-TSS) has also been generated that displays similar resistance to internalization and an increase in migration to S1P but not to other chemokines (Arnon et al. 2011). These two knockin mice, which appear to cause similar downstream signaling to wild-type mice, can help separate out which events following S1P1 agonist treatment are a result of S1P1 signaling, and which events may be affected by the internalization of the receptor.
Conclusions
The pleiotropic functions and complex biology of S1P make it a delicate system to study. Studying the actions of S1P on its receptors has yielded significant insights into a broad range of physiological systems. S1P will continue to be an exciting area of study, particularly because of its relevance in human disease. Despite the relatively recent discovery of S1P receptors, modulation of S1P biology has already progressed to clinical treatment of disease. This progression has been aided significantly by the generation of an incredible diversity of both chemical and genetic tools that affect all aspects of S1P biology. While significant progress has been made, several areas of development of these tools remain, particularly the generation of selective in vivo chemical modulators and conditional knockouts. A summary of Additionally, it is likely that other genes regulating S1P function have yet to be found Truly, the combined use of chemical and the genetic tools within a single experiment provides the most useful and conclusive findings into the functions of S1P and its receptors. The continued development and refinement of both will hopefully continue to aid in our understanding of physiology and in the treatment of human disease.
Contributor Information
Michael B. A. Oldstone, Phone: +1858-784-8054, Email: mbaobo@scripps.edu
Hugh Rosen, Phone: +18587842396, Email: hrosen@scripps.edu.
Stuart M. Cahalan, Email: scahalan@scripps.edu
References
- Allende ML, Bektas M, Lee BG, Bonifacino E, Kang J, Tuymetova G, Chen W, Saba JD, Proia RL. Sphingosine-1-phosphate lyase deficiency produces a pro-inflammatory response while impairing neutrophil trafficking. J Biol Chem. 2011;286:7348–7358. doi: 10.1074/jbc.M110.171819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allende ML, Dreier JL, Mandala S, Proia RL. Expression of the sphingosine 1-phosphate receptor, S1P1, on T-cells controls thymic emigration. J Biol Chem. 2004;279:15396–15401. doi: 10.1074/jbc.M314291200. [DOI] [PubMed] [Google Scholar]
- Allende ML, Sasaki T, Kawai H, Olivera A, Mi Y, van Echten-Deckert G, Hajdu R, Rosenbach M, Keohane CA, Mandala S, Spiegel S, Proia RL. Mice deficient in sphingosine kinase 1 are rendered lymphopenic by FTY720. J Biol Chem. 2004;279:52487–52492. doi: 10.1074/jbc.M406512200. [DOI] [PubMed] [Google Scholar]
- Allende ML, Tuymetova G, Lee BG, Bonifacino E, Wu YP, Proia RL. S1P1 receptor directs the release of immature B cells from bone marrow into blood. J Exp Med. 2010;207:1113–1124. doi: 10.1084/jem.20092210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allende ML, Yamashita T, Proia RL. G-protein-coupled receptor S1P1 acts within endothelial cells to regulate vascular maturation. Blood. 2003;102:3665–3667. doi: 10.1182/blood-2003-02-0460. [DOI] [PubMed] [Google Scholar]
- Angst D, Bollbuck B, Janser P, Quancard J (2010) Biaryl benzylamine derivatives. World Intellect Property Organ, Pub. No. WO/2010/072712
- Angst D, Janser P, Quancard J, Buehlmayer P, Berst F, Oberer L, Beerli C, Streiff M, Pally C, Hersperger R, Bruns C, Bassilana F, Bollbuck B. An oral Sphingosine 1-phosphate receptor 1 (S1P(1)) antagonist prodrug with efficacy in vivo: discovery, synthesis, and evaluation. J Med Chem. 2012;55:9722–9734. doi: 10.1021/jm3009508. [DOI] [PubMed] [Google Scholar]
- Arankalle VA, Lole KS, Arya RP, Tripathy AS, Ramdasi AY, Chadha MS, Sangle SA, Kadam DB (2010) Role of host immune response and viral load in the differential outcome of pandemic H1N1 (2009) influenza virus infection in Indian patients. PLoS One 5 [DOI] [PMC free article] [PubMed]
- Arnon TI, Xu Y, Lo C, Pham T, An J, Coughlin S, Dorn GW, Cyster JG. GRK2-dependent S1PR1 desensitization is required for lymphocytes to overcome their attraction to blood. Science. 2011;333:1898–1903. doi: 10.1126/science.1208248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bagdanoff JT, Donoviel MS, Nouraldeen A, Carlsen M, Jessop TC, Tarver J, Aleem S, Dong L, Zhang H, Boteju L, Hazelwood J, Yan J, Bednarz M, Layek S, Owusu IB, Gopinathan S, Moran L, Lai Z, Kramer J, Kimball SD, Yalamanchili P, Heydorn WE, Frazier KS, Brooks B, Brown P, Wilson A, Sonnenburg WK, Main A, Carson KG, Oravecz T, Augeri DJ. Inhibition of sphingosine 1-phosphate lyase for the treatment of rheumatoid arthritis: discovery of (E)-1-(4-((1R,2S,3R)-1,2,3,4-tetrahydroxybutyl)-1H-imidazol-2-yl)ethanone oxime (LX2931) and (1R,2S,3R)-1-(2-(isoxazol-3-yl)-1H-imidazol-4-yl)butane-1,2,3,4-tetraol (LX2932) J Med Chem. 2010;53:8650–8662. doi: 10.1021/jm101183p. [DOI] [PubMed] [Google Scholar]
- Bajwa A, Huang L, Ye H, Dondeti K, Song S, Rosin DL, Lynch KR, Lobo PI, Li L, Okusa MD. Dendritic cell sphingosine 1-phosphate receptor-3 regulates Th1-Th2 polarity in kidney ischemia-reperfusion injury. J Immunol. 2012;189:2584–2596. doi: 10.4049/jimmunol.1200999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker DA, Barth J, Chang R, Obeid LM, Gilkeson GS. Genetic sphingosine kinase 1 deficiency significantly decreases synovial inflammation and joint erosions in murine TNF-alpha-induced arthritis. J Immunol. 2010;185:2570–2579. doi: 10.4049/jimmunol.1000644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bandhuvula P, Honbo N, Wang GY, Jin ZQ, Fyrst H, Zhang M, Borowsky AD, Dillard L, Karliner JS, Saba JD. S1P lyase: a novel therapeutic target for ischemia-reperfusion injury of the heart. Am J Physiol Heart Circ Physiol. 2011;300:H1753–H1761. doi: 10.1152/ajpheart.00946.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bektas M, Allende ML, Lee BG, Chen W, Amar MJ, Remaley AT, Saba JD, Proia RL. Sphingosine 1-phosphate lyase deficiency disrupts lipid homeostasis in liver. J Biol Chem. 2010;285:10880–10889. doi: 10.1074/jbc.M109.081489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beljanski V, Lewis CS, Smith CD. Antitumor activity of sphingosine kinase 2 inhibitor ABC294640 and sorafenib in hepatocellular carcinoma xenografts. Cancer Biol Ther. 2011;11:524–534. doi: 10.4161/cbt.11.5.14677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breart B, Ramos-Perez WD, Mendoza A, Salous AK, Gobert M, Huang Y, Adams RH, Lafaille JJ, Escalante-Alcalde D, Morris AJ, Schwab SR. Lipid phosphate phosphatase 3 enables efficient thymic egress. J Exp Med. 2011;208:1267–1278. doi: 10.1084/jem.20102551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brinkmann V, Davis MD, Heise CE, Albert R, Cottens S, Hof R, Bruns C, Prieschl E, Baumruker T, Hiestand P, Foster CA, Zollinger M, Lynch KR. The immune modulator FTY720 targets sphingosine 1-phosphate receptors. J Biol Chem. 2002;277:21453–21457. doi: 10.1074/jbc.C200176200. [DOI] [PubMed] [Google Scholar]
- Brinkmann V, Pinschewer DD, Feng L, Chen S. FTY720: altered lymphocyte traffic results in allograft protection. Transplantation. 2001;72:764–769. doi: 10.1097/00007890-200109150-00002. [DOI] [PubMed] [Google Scholar]
- Budde K, Schmouder RL, Brunkhorst R, Nashan B, Lucker PW, Mayer T, Choudhury S, Skerjanec A, Kraus G, Neumayer HH. First human trial of FTY720, a novel immunomodulator, in stable renal transplant patients. J Am Soc Nephrol. 2002;13:1073–1083. doi: 10.1681/ASN.V1341073. [DOI] [PubMed] [Google Scholar]
- Buehrer BM, Bell RM. Inhibition of sphingosine kinase in vitro and in platelets. Implications for signal transduction pathways. J Biol Chem. 1992;267:3154–3159. [PubMed] [Google Scholar]
- Cahalan SM, Gonzalez-Cabrera PJ, Nguyen N, Guerrero M, Cisar EA, Leaf NB, Brown SJ, Roberts E, Rosen H. Sphingosine 1-phosphate receptor 1 (S1P(1)) upregulation and amelioration of experimental autoimmune encephalomyelitis by an S1P(1) antagonist. Mol Pharmacol. 2013;83:316–321. doi: 10.1124/mol.112.082958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cahalan SM, Gonzalez-Cabrera PJ, Sarkisyan G, Nguyen N, Schaeffer MT, Huang L, Yeager A, Clemons B, Scott F, Rosen H. Actions of a picomolar short-acting S1P agonist in S1P-eGFP knock-in mice. Nat Chem Biol. 2011;7:254–256. doi: 10.1038/nchembio.547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Camerer E, Regard JB, Cornelissen I, Srinivasan Y, Duong DN, Palmer D, Pham TH, Wong JS, Pappu R, Coughlin SR. Sphingosine-1-phosphate in the plasma compartment regulates basal and inflammation-induced vascular leak in mice. J Clin Invest. 2009;119:1871–1879. doi: 10.1172/JCI38575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi JW, Gardell SE, Herr DR, Rivera R, Lee CW, Noguchi K, Teo ST, Yung YC, Lu M, Kennedy G, Chun J. FTY720 (fingolimod) efficacy in an animal model of multiple sclerosis requires astrocyte sphingosine 1-phosphate receptor 1 (S1P1) modulation. Proc Natl Acad Sci USA. 2011;108:751–756. doi: 10.1073/pnas.1014154108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cinamon G, Matloubian M, Lesneski MJ, Xu Y, Low C, Lu T, Proia RL, Cyster JG. Sphingosine 1-phosphate receptor 1 promotes B cell localization in the splenic marginal zone. Nat Immunol. 2004;5:713–720. doi: 10.1038/ni1083. [DOI] [PubMed] [Google Scholar]
- Cinamon G, Zachariah MA, Lam OM, Foss FW, Jr, Cyster JG. Follicular shuttling of marginal zone B cells facilitates antigen transport. Nat Immunol. 2008;9:54–62. doi: 10.1038/ni1542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis MD, Clemens JJ, Macdonald TL, Lynch KR. Sphingosine 1-phosphate analogs as receptor antagonists. J Biol Chem. 2005;280:9833–9841. doi: 10.1074/jbc.M412356200. [DOI] [PubMed] [Google Scholar]
- Escalante-Alcalde D, Hernandez L, Le Stunff H, Maeda R, Lee HS, Jr, Gang C, Sciorra VA, Daar I, Spiegel S, Morris AJ, Stewart CL. The lipid phosphatase LPP3 regulates extra-embryonic vasculogenesis and axis patterning. Development. 2003;130:4623–4637. doi: 10.1242/dev.00635. [DOI] [PubMed] [Google Scholar]
- Fernandez-Serrano S, Dorca J, Coromines M, Carratala J, Gudiol F, Manresa F. Molecular inflammatory responses measured in blood of patients with severe community-acquired pneumonia. Clin Diagn Lab Immunol. 2003;10:813–820. doi: 10.1128/CDLI.10.5.813-820.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fitzpatrick LR, Green C, Maines LW, Smith CD. Experimental osteoarthritis in rats is attenuated by ABC294640, a selective inhibitor of sphingosine kinase-2. Pharmacology. 2011;87:135–143. doi: 10.1159/000323911. [DOI] [PubMed] [Google Scholar]
- Foss FW, Jr, Snyder AH, Davis MD, Rouse M, Okusa MD, Lynch KR, Macdonald TL. Synthesis and biological evaluation of gamma-aminophosphonates as potent, subtype-selective sphingosine 1-phosphate receptor agonists and antagonists. Bioorg Med Chem. 2007;15:663–677. doi: 10.1016/j.bmc.2006.10.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- French KJ, Zhuang Y, Maines LW, Gao P, Wang W, Beljanski V, Upson JJ, Green CL, Keller SN, Smith CD. Pharmacology and antitumor activity of ABC294640, a selective inhibitor of sphingosine kinase-2. J Pharmacol Exp Ther. 2010;333:129–139. doi: 10.1124/jpet.109.163444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujii Y, Hirayama T, Ohtake H, Ono N, Inoue T, Sakurai T, Takayama T, Matsumoto K, Tsukahara N, Hidano S, Harima N, Nakazawa K, Igarashi Y, Goitsuka R. Amelioration of collagen-induced arthritis by a novel S1P1 antagonist with immunomodulatory activities. J Immunol. 2012;188:206–215. doi: 10.4049/jimmunol.1101537. [DOI] [PubMed] [Google Scholar]
- Fujino M, Funeshima N, Kitazawa Y, Kimura H, Amemiya H, Suzuki S, Li XK. Amelioration of experimental autoimmune encephalomyelitis in Lewis rats by FTY720 treatment. J Pharmacol Exp Ther. 2003;305:70–77. doi: 10.1124/jpet.102.045658. [DOI] [PubMed] [Google Scholar]
- Fukuhara S, Simmons S, Kawamura S, Inoue A, Orba Y, Tokudome T, Sunden Y, Arai Y, Moriwaki K, Ishida J, Uemura A, Kiyonari H, Abe T, Fukamizu A, Hirashima M, Sawa H, Aoki J, Ishii M, Mochizuki N. The sphingosine-1-phosphate transporter Spns2 expressed on endothelial cells regulates lymphocyte trafficking in mice. J Clin Invest. 2012;122:1416–1426. doi: 10.1172/JCI60746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galicia-Rosas G, Pikor N, Schwartz JA, Rojas O, Jian A, Summers-Deluca L, Ostrowski M, Nuesslein-Hildesheim B, Gommerman JL. A sphingosine-1-phosphate receptor 1-directed agonist reduces central nervous system inflammation in a plasmacytoid dendritic cell-dependent manner. J Immunol. 2012;189:3700–3706. doi: 10.4049/jimmunol.1102261. [DOI] [PubMed] [Google Scholar]
- Gergely P, Nuesslein-Hildesheim B, Guerini D, Brinkmann V, Traebert M, Bruns C, Pan S, Gray NS, Hinterding K, Cooke NG, Groenewegen A, Vitaliti A, Sing T, Luttringer O, Yang J, Gardin A, Wang N, Crumb WJ, Jr, Saltzman M, Rosenberg M, Wallstrom E. The selective sphingosine 1-phosphate receptor modulator BAF312 redirects lymphocyte distribution and has species-specific effects on heart rate. Br J Pharmacol. 2012;167:1035–1047. doi: 10.1111/j.1476-5381.2012.02061.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golfier S, Kondo S, Schulze T, Takeuchi T, Vassileva G, Achtman AH, Graler MH, Abbondanzo SJ, Wiekowski M, Kremmer E, Endo Y, Lira SA, Bacon KB, Lipp M. Shaping of terminal megakaryocyte differentiation and proplatelet development by sphingosine-1-phosphate receptor S1P4. FASEB J. 2010;24:4701–4710. doi: 10.1096/fj.09-141473. [DOI] [PubMed] [Google Scholar]
- Gonzalez-Cabrera PJ, Cahalan SM, Nguyen N, Sarkisyan G, Leaf NB, Cameron MD, Kago T, Rosen H. S1P(1) receptor modulation with cyclical recovery from lymphopenia ameliorates mouse model of multiple sclerosis. Mol Pharmacol. 2012;81:166–174. doi: 10.1124/mol.111.076109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalez-Cabrera PJ, Hla T, Rosen H. Mapping pathways downstream of sphingosine 1-phosphate subtype 1 by differential chemical perturbation and proteomics. J Biol Chem. 2007;282:7254–7264. doi: 10.1074/jbc.M610581200. [DOI] [PubMed] [Google Scholar]
- Gonzalez-Cabrera PJ, Jo E, Sanna MG, Brown S, Leaf N, Marsolais D, Schaeffer MT, Chapman J, Cameron M, Guerrero M, Roberts E, Rosen H. Full pharmacological efficacy of a novel S1P1 agonist that does not require S1P-like headgroup interactions. Mol Pharmacol. 2008;74:1308–1318. doi: 10.1124/mol.108.049783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graler MH, Goetzl EJ. The immunosuppressant FTY720 down-regulates sphingosine 1-phosphate G-protein-coupled receptors. FASEB J. 2004;18:551–553. doi: 10.1096/fj.03-0910fje. [DOI] [PubMed] [Google Scholar]
- Guerrero M, Urbano M, Velaparthi S, Zhao J, Schaeffer MT, Brown S, Rosen H, Roberts E. Discovery, design and synthesis of the first reported potent and selective sphingosine-1-phosphate 4 (S1P4) receptor antagonists. Bioorg Med Chem Lett. 2011;21:3632–3636. doi: 10.1016/j.bmcl.2011.04.097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagen-Euteneuer N, Lutjohann D, Park H, Merrill AH, Jr, van Echten-Deckert G. Sphingosine 1-phosphate (S1P) lyase deficiency increases sphingolipid formation via recycling at the expense of de novo biosynthesis in neurons. J Biol Chem. 2012;287:9128–9136. doi: 10.1074/jbc.M111.302380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanson MA, Roth CB, Jo E, Griffith MT, Scott FL, Reinhart G, Desale H, Clemons B, Cahalan SM, Schuerer SC, Sanna MG, Han GW, Kuhn P, Rosen H, Stevens RC. Crystal structure of a lipid G protein-coupled receptor. Science. 2012;335:851–855. doi: 10.1126/science.1215904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris CM, Hobson AD, Wilson NS (2010) Agonists and antagonists of the S1P(5) receptor, and methods of uses thereof. World Intellect Property Organ, Pub. No. WO/2010/093704
- Harris GL, Creason MB, Brulte GB, Herr DR. In vitro and in vivo antagonism of a G protein-coupled receptor (S1P3) with a novel blocking monoclonal antibody. PLoS ONE. 2012;7:e35129. doi: 10.1371/journal.pone.0035129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herr DR, Grillet N, Schwander M, Rivera R, Muller U, Chun J. Sphingosine 1-phosphate (S1P) signaling is required for maintenance of hair cells mainly via activation of S1P2. J Neurosci. 2007;27:1474–1478. doi: 10.1523/JNEUROSCI.4245-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hisano Y, Kobayashi N, Kawahara A, Yamaguchi A, Nishi T. The sphingosine 1-phosphate transporter, SPNS2, functions as a transporter of the phosphorylated form of the immunomodulating agent FTY720. J Biol Chem. 2011;286:1758–1766. doi: 10.1074/jbc.M110.171116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hisano Y, Kobayashi N, Yamaguchi A, Nishi T. Mouse SPNS2 functions as a sphingosine-1-phosphate transporter in vascular endothelial cells. PLoS ONE. 2012;7:e38941. doi: 10.1371/journal.pone.0038941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hla T. Physiological and pathological actions of sphingosine 1-phosphate. Semin Cell Dev Biol. 2004;15:513–520. doi: 10.1016/j.semcdb.2004.05.002. [DOI] [PubMed] [Google Scholar]
- Huang KJ, Su IJ, Theron M, Wu YC, Lai SK, Liu CC, Lei HY. An interferon-gamma-related cytokine storm in SARS patients. J Med Virol. 2005;75:185–194. doi: 10.1002/jmv.20255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishii I, Friedman B, Ye X, Kawamura S, McGiffert C, Contos JJ, Kingsbury MA, Zhang G, Brown JH, Chun J. Selective loss of sphingosine 1-phosphate signaling with no obvious phenotypic abnormality in mice lacking its G protein-coupled receptor, LP(B3)/EDG-3. J Biol Chem. 2001;276:33697–33704. doi: 10.1074/jbc.M104441200. [DOI] [PubMed] [Google Scholar]
- Ishii I, Ye X, Friedman B, Kawamura S, Contos JJ, Kingsbury MA, Yang AH, Zhang G, Brown JH, Chun J. Marked perinatal lethality and cellular signaling deficits in mice null for the two sphingosine 1-phosphate (S1P) receptors, S1P(2)/LP(B2)/EDG-5 and S1P(3)/LP(B3)/EDG-3. J Biol Chem. 2002;277:25152–25159. doi: 10.1074/jbc.M200137200. [DOI] [PubMed] [Google Scholar]
- Jaillard C, Harrison S, Stankoff B, Aigrot MS, Calver AR, Duddy G, Walsh FS, Pangalos MN, Arimura N, Kaibuchi K, Zalc B, Lubetzki C. Edg8/S1P5: an oligodendroglial receptor with dual function on process retraction and cell survival. J Neurosci. 2005;25:1459–1469. doi: 10.1523/JNEUROSCI.4645-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jary E, Bee T, Walker SR, Chung SK, Seo KC, Morris JC, Don AS. Elimination of a hydroxyl group in FTY720 dramatically improves the phosphorylation rate. Mol Pharmacol. 2010;78:685–692. doi: 10.1124/mol.110.064873. [DOI] [PubMed] [Google Scholar]
- Jenne CN, Enders A, Rivera R, Watson SR, Bankovich AJ, Pereira JP, Xu Y, Roots CM, Beilke JN, Banerjee A, Reiner SL, Miller SA, Weinmann AS, Goodnow CC, Lanier LL, Cyster JG, Chun J. T-bet-dependent S1P5 expression in NK cells promotes egress from lymph nodes and bone marrow. J Exp Med. 2009;206:2469–2481. doi: 10.1084/jem.20090525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jo E, Bhhatarai B, Repetto E, Guerrero M, Riley S, Brown SJ, Kohno Y, Roberts E, Schurer SC, Rosen H. Novel Selective Allosteric and Bitopic Ligands for the S1P(3) Receptor. ACS Chem Biol. 2012;7:1975–1983. doi: 10.1021/cb300392z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jo E, Sanna MG, Gonzalez-Cabrera PJ, Thangada S, Tigyi G, Osborne DA, Hla T, Parrill AL, Rosen H. S1P1-selective in vivo-active agonists from high-throughput screening: off-the-shelf chemical probes of receptor interactions, signaling, and fate. Chem Biol. 2005;12:703–715. doi: 10.1016/j.chembiol.2005.04.019. [DOI] [PubMed] [Google Scholar]
- Jung B, Obinata H, Galvani S, Mendelson K, Ding BS, Skoura A, Kinzel B, Brinkmann V, Rafii S, Evans T, Hla T. Flow-regulated endothelial S1P receptor-1 signaling sustains vascular development. Dev Cell. 2012;23:600–610. doi: 10.1016/j.devcel.2012.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kai M, Wada I, Imai S, Sakane F, Kanoh H. Cloning and characterization of two human isozymes of Mg2+-independent phosphatidic acid phosphatase. J Biol Chem. 1997;272:24572–24578. doi: 10.1074/jbc.272.39.24572. [DOI] [PubMed] [Google Scholar]
- Kawahara A, Nishi T, Hisano Y, Fukui H, Yamaguchi A, Mochizuki N. The sphingolipid transporter spns2 functions in migration of zebrafish myocardial precursors. Science. 2009;323:524–527. doi: 10.1126/science.1167449. [DOI] [PubMed] [Google Scholar]
- Kennedy AJ, Mathews TP, Kharel Y, Field SD, Moyer ML, East JE, Houck JD, Lynch KR, Macdonald TL. Development of amidine-based sphingosine kinase 1 nanomolar inhibitors and reduction of sphingosine 1-phosphate in human leukemia cells. J Med Chem. 2011;54:3524–3548. doi: 10.1021/jm2001053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan WA, Dobrowsky R, el Touny S, Hannun YA. Protein kinase C and platelet inhibition by D-erythro-sphingosine: comparison with N, N-dimethylsphingosine and commercial preparation. Biochem Biophys Res Commun. 1990;172:683–691. doi: 10.1016/0006-291x(90)90728-6. [DOI] [PubMed] [Google Scholar]
- Kharel Y, Lee S, Snyder AH, Sheasley-O’neill SL, Morris MA, Setiady Y, Zhu R, Zigler MA, Burcin TL, Ley K, Tung KS, Engelhard VH, Macdonald TL, Pearson-White S, Lynch KR. Sphingosine kinase 2 is required for modulation of lymphocyte traffic by FTY720. J Biol Chem. 2005;280:36865–36872. doi: 10.1074/jbc.M506293200. [DOI] [PubMed] [Google Scholar]
- Kohama T, Olivera A, Edsall L, Nagiec MM, Dickson R, Spiegel S. Molecular cloning and functional characterization of murine sphingosine kinase. J Biol Chem. 1998;273:23722–23728. doi: 10.1074/jbc.273.37.23722. [DOI] [PubMed] [Google Scholar]
- Kono M, Belyantseva IA, Skoura A, Frolenkov GI, Starost MF, Dreier JL, Lidington D, Bolz SS, Friedman TB, Hla T, Proia RL. Deafness and stria vascularis defects in S1P2 receptor-null mice. J Biol Chem. 2007;282:10690–10696. doi: 10.1074/jbc.M700370200. [DOI] [PubMed] [Google Scholar]
- Kono M, Mi Y, Liu Y, Sasaki T, Allende ML, Wu YP, Yamashita T, Proia RL. The sphingosine-1-phosphate receptors S1P1, S1P2, and S1P3 function coordinately during embryonic angiogenesis. J Biol Chem. 2004;279:29367–29373. doi: 10.1074/jbc.M403937200. [DOI] [PubMed] [Google Scholar]
- Lee N, Wong CK, Chan PK, Chan MC, Wong RY, Lun SW, Ngai KL, Lui GC, Wong BC, Lee SK, Choi KW, Hui DS. Cytokine response patterns in severe pandemic 2009 H1N1 and seasonal influenza among hospitalized adults. PLoS ONE. 2011;6:e26050. doi: 10.1371/journal.pone.0026050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C, Chi XX, Xie W, Strong JA, Zhang JM, Nicol GD. Sphingosine 1-phosphate receptor 2 antagonist JTE-013 increases the excitability of sensory neurons independently of the receptor. J Neurophysiol. 2012;108:1473–1483. doi: 10.1152/jn.00825.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim KG, Tonelli F, Li Z, Lu X, Bittman R, Pyne S, Pyne NJ. FTY720 analogues as sphingosine kinase 1 inhibitors: enzyme inhibition kinetics, allosterism, proteasomal degradation, and actin rearrangement in MCF-7 breast cancer cells. J Biol Chem. 2011;286:18633–18640. doi: 10.1074/jbc.M111.220756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu H, Sugiura M, Nava VE, Edsall LC, Kono K, Poulton S, Milstien S, Kohama T, Spiegel S. Molecular cloning and functional characterization of a novel mammalian sphingosine kinase type 2 isoform. J Biol Chem. 2000;275:19513–19520. doi: 10.1074/jbc.M002759200. [DOI] [PubMed] [Google Scholar]
- Liu Y, Wada R, Yamashita T, Mi Y, Deng CX, Hobson JP, Rosenfeldt HM, Nava VE, Chae SS, Lee MJ, Liu CH, Hla T, Spiegel S, Proia RL. Edg-1, the G protein-coupled receptor for sphingosine-1-phosphate, is essential for vascular maturation. J Clin Invest. 2000;106:951–961. doi: 10.1172/JCI10905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Juarez A, Morales-Lazaro S, Sanchez-Sanchez R, Sunkara M, Lomeli H, Velasco I, Morris AJ, Escalante-Alcalde D. Expression of LPP3 in Bergmann glia is required for proper cerebellar sphingosine-1-phosphate metabolism/signaling and development. Glia. 2011;59:577–589. doi: 10.1002/glia.21126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo ZJ, Tanaka T, Kimura F, Miyasaka M. Analysis of the mode of action of a novel immunosuppressant FTY720 in mice. Immunopharmacology. 1999;41:199–207. doi: 10.1016/s0162-3109(99)00004-1. [DOI] [PubMed] [Google Scholar]
- Maceyka M, Harikumar KB, Milstien S, Spiegel S. Sphingosine-1-phosphate signaling and its role in disease. Trends Cell Biol. 2012;22:50–60. doi: 10.1016/j.tcb.2011.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacLennan AJ, Carney PR, Zhu WJ, Chaves AH, Garcia J, Grimes JR, Anderson KJ, Roper SN, Lee N. An essential role for the H218/AGR16/Edg-5/LP(B2) sphingosine 1-phosphate receptor in neuronal excitability. Eur J Neurosci. 2001;14:203–209. doi: 10.1046/j.0953-816x.2001.01634.x. [DOI] [PubMed] [Google Scholar]
- Maeda Y, Matsuyuki H, Shimano K, Kataoka H, Sugahara K, Chiba K. Migration of CD4 T cells and dendritic cells toward sphingosine 1-phosphate (S1P) is mediated by different receptor subtypes: S1P regulates the functions of murine mature dendritic cells via S1P receptor type 3. J Immunol. 2007;178:3437–3446. doi: 10.4049/jimmunol.178.6.3437. [DOI] [PubMed] [Google Scholar]
- Maines LW, Fitzpatrick LR, French KJ, Zhuang Y, Xia Z, Keller SN, Upson JJ, Smith CD. Suppression of ulcerative colitis in mice by orally available inhibitors of sphingosine kinase. Dig Dis Sci. 2008;53:997–1012. doi: 10.1007/s10620-007-0133-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maines LW, Fitzpatrick LR, Green CL, Zhuang Y, Smith CD. Efficacy of a novel sphingosine kinase inhibitor in experimental Crohn’s disease. Inflammopharmacology. 2010;18:73–85. doi: 10.1007/s10787-010-0032-x. [DOI] [PubMed] [Google Scholar]
- Mair N, Benetti C, Andratsch M, Leitner MG, Constantin CE, Camprubi-Robles M, Quarta S, Biasio W, Kuner R, Gibbins IL, Kress M, Haberberger RV. Genetic evidence for involvement of neuronally expressed S1P(1) receptor in nociceptor sensitization and inflammatory pain. PLoS ONE. 2011;6:e17268. doi: 10.1371/journal.pone.0017268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandala S, Hajdu R, Bergstrom J, Quackenbush E, Xie J, Milligan J, Thornton R, Shei GJ, Card D, Keohane C, Rosenbach M, Hale J, Lynch CL, Rupprecht K, Parsons W, Rosen H. Alteration of lymphocyte trafficking by sphingosine-1-phosphate receptor agonists. Science. 2002;296:346–349. doi: 10.1126/science.1070238. [DOI] [PubMed] [Google Scholar]
- Mandala SM. Sphingosine-1-phosphate phosphatases. Prostaglandins. 2001;64:143–156. doi: 10.1016/s0090-6980(01)00111-3. [DOI] [PubMed] [Google Scholar]
- Marsolais D, Hahm B, Edelmann KH, Walsh KB, Guerrero M, Hatta Y, Kawaoka Y, Roberts E, Oldstone MB, Rosen H. Local not systemic modulation of dendritic cell S1P receptors in lung blunts virus-specific immune responses to influenza. Mol Pharmacol. 2008;74:896–903. doi: 10.1124/mol.108.048769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsolais D, Hahm B, Walsh KB, Edelmann KH, McGavern D, Hatta Y, Kawaoka Y, Rosen H, Oldstone MB. A critical role for the sphingosine analog AAL-R in dampening the cytokine response during influenza virus infection. Proc Natl Acad Sci USA. 2009;106:1560–1565. doi: 10.1073/pnas.0812689106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matloubian M, Lo CG, Cinamon G, Lesneski MJ, Xu Y, Brinkmann V, Allende ML, Proia RL, Cyster JG. Lymphocyte egress from thymus and peripheral lymphoid organs is dependent on S1P receptor 1. Nature. 2004;427:355–360. doi: 10.1038/nature02284. [DOI] [PubMed] [Google Scholar]
- Mattes H, Dev KK, Bouhelal R, Barske C, Gasparini F, Guerini D, Mir AK, Orain D, Osinde M, Picard A, Dubois C, Tasdelen E, Haessig S. Design and synthesis of selective and potent orally active S1P5 agonists. Chem Med Chem. 2010;5:1693–1696. doi: 10.1002/cmdc.201000253. [DOI] [PubMed] [Google Scholar]
- Mayol K, Biajoux V, Marvel J, Balabanian K, Walzer T. Sequential desensitization of CXCR4 and S1P5 controls natural killer cell trafficking. Blood. 2011;118:4863–4871. doi: 10.1182/blood-2011-06-362574. [DOI] [PubMed] [Google Scholar]
- Means CK, Xiao CY, Li Z, Zhang T, Omens JH, Ishii I, Chun J, Brown JH. Sphingosine 1-phosphate S1P2 and S1P3 receptor-mediated Akt activation protects against in vivo myocardial ischemia-reperfusion injury. Am J Physiol Heart Circ Physiol. 2007;292:H2944–H2951. doi: 10.1152/ajpheart.01331.2006. [DOI] [PubMed] [Google Scholar]
- Mendoza A, Breart B, Ramos-Perez WD, Pitt LA, Gobert M, Sunkara M, Lafaille JJ, Morris AJ, Schwab SR. The transporter spns2 is required for secretion of lymph but not plasma sphingosine-1-phosphate. Cell Rep. 2012;2:1104–1110. doi: 10.1016/j.celrep.2012.09.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng Q, Zhao B, Xu Q, Xu X, Deng G, Li C, Luan L, Ren F, Wang H, Xu H, Xu Y, Zhang H, Xiang JN, Elliott JD, Guo TB, Zhao Y, Zhang W, Lu H, Lin X. Indole-propionic acid derivatives as potent, S1P3-sparing and EAE efficacious sphingosine-1-phosphate 1 (S1P1) receptor agonists. Bioorg Med Chem Lett. 2012;22:2794–2797. doi: 10.1016/j.bmcl.2012.02.083. [DOI] [PubMed] [Google Scholar]
- Merrill AH, Jr, Nimkar S, Menaldino D, Hannun YA, Loomis C, Bell RM, Tyagi SR, Lambeth JD, Stevens VL, Hunter R, et al. Structural requirements for long-chain (sphingoid) base inhibition of protein kinase C in vitro and for the cellular effects of these compounds. Biochemistry. 1989;28:3138–3145. doi: 10.1021/bi00434a004. [DOI] [PubMed] [Google Scholar]
- Mitra P, Oskeritzian CA, Payne SG, Beaven MA, Milstien S, Spiegel S. Role of ABCC1 in export of sphingosine-1-phosphate from mast cells. Proc Natl Acad Sci USA. 2006;103:16394–16399. doi: 10.1073/pnas.0603734103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mizugishi K, Yamashita T, Olivera A, Miller GF, Spiegel S, Proia RL. Essential role for sphingosine kinases in neural and vascular development. Mol Cell Biol. 2005;25:11113–11121. doi: 10.1128/MCB.25.24.11113-11121.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mullershausen F, Zecri F, Cetin C, Billich A, Guerini D, Seuwen K. Persistent signaling induced by FTY720-phosphate is mediated by internalized S1P1 receptors. Nat Chem Biol. 2009;5:428–434. doi: 10.1038/nchembio.173. [DOI] [PubMed] [Google Scholar]
- Murakami A, Takasugi H, Ohnuma S, Koide Y, Sakurai A, Takeda S, Hasegawa T, Sasamori J, Konno T, Hayashi K, Watanabe Y, Mori K, Sato Y, Takahashi A, Mochizuki N, Takakura N. Sphingosine 1-phosphate (S1P) regulates vascular contraction via S1P3 receptor: investigation based on a new S1P3 receptor antagonist. Mol Pharmacol. 2010;77:704–713. doi: 10.1124/mol.109.061481. [DOI] [PubMed] [Google Scholar]
- Nagahashi M, Kim EY, Yamada A, Ramachandran S, Allegood JC, Hait NC, Maceyka M, Milstien S, Takabe K, Spiegel S. Spns2, a transporter of phosphorylated sphingoid bases, regulates their blood and lymph levels and the lymphatic network. FASEB J. 2012;27:1001–1011. doi: 10.1096/fj.12-219618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagahashi M, Ramachandran S, Kim EY, Allegood JC, Rashid OM, Yamada A, Zhao R, Milstien S, Zhou H, Spiegel S, Takabe K. Sphingosine-1-phosphate produced by sphingosine kinase 1 promotes breast cancer progression by stimulating angiogenesis and lymphangiogenesis. Cancer Res. 2012;72:726–735. doi: 10.1158/0008-5472.CAN-11-2167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagata N, Iwata N, Hasegawa H, Fukushi S, Harashima A, Sato Y, Saijo M, Taguchi F, Morikawa S, Sata T. Mouse-passaged severe acute respiratory syndrome-associated coronavirus leads to lethal pulmonary edema and diffuse alveolar damage in adult but not young mice. Am J Pathol. 2008;172:1625–1637. doi: 10.2353/ajpath.2008.071060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura T, Asano M, Sekiguchi Y, Mizuno Y, Tamaki K, Kimura T, Nara F, Kawase Y, Shimozato T, Doi H, Kagari T, Tomisato W, Inoue R, Nagasaki M, Yuita H, Oguchi-Oshima K, Kaneko R, Watanabe N, Abe Y, Nishi T. Discovery of CS-2100, a potent, orally active and S1P3-sparing S1P1 agonist. Bioorg Med Chem Lett. 2012;22:1788–1792. doi: 10.1016/j.bmcl.2011.12.019. [DOI] [PubMed] [Google Scholar]
- Niessen F, Schaffner F, Furlan-Freguia C, Pawlinski R, Bhattacharjee G, Chun J, Derian CK, Andrade-Gordon P, Rosen H, Ruf W. Dendritic cell PAR1-S1P3 signalling couples coagulation and inflammation. Nature. 2008;452:654–658. doi: 10.1038/nature06663. [DOI] [PubMed] [Google Scholar]
- Nijnik A, Clare S, Hale C, Chen J, Raisen C, Mottram L, Lucas M, Estabel J, Ryder E, Adissu H, Sanger Mouse Genetics P, Adams NC, Ramirez-Solis R, White JK, Steel KP, Dougan G, Hancock RE. The role of sphingosine-1-phosphate transporter Spns2 in immune system function. J Immunol. 2012;189:102–111. doi: 10.4049/jimmunol.1200282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nofer JR, van der Giet M, Tolle M, Wolinska I, von Wnuck Lipinski K, Baba HA, Tietge UJ, Godecke A, Ishii I, Kleuser B, Schafers M, Fobker M, Zidek W, Assmann G, Chun J, Levkau B. HDL induces NO-dependent vasorelaxation via the lysophospholipid receptor S1P3. J Clin Invest. 2004;113:569–581. doi: 10.1172/JCI18004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oo ML, Thangada S, Wu MT, Liu CH, Macdonald TL, Lynch KR, Lin CY, Hla T. Immunosuppressive and anti-angiogenic sphingosine 1-phosphate receptor-1 agonists induce ubiquitinylation and proteasomal degradation of the receptor. J Biol Chem. 2007;282:9082–9089. doi: 10.1074/jbc.M610318200. [DOI] [PubMed] [Google Scholar]
- Panchatcharam M, Miriyala S, Salous A, Wheeler J, Dong A, Mueller P, Sunkara M, Escalante-Alcalde D, Morris AJ, Smyth SS. Lipid phosphate phosphatase 3 negatively regulates smooth muscle cell phenotypic modulation to limit intimal hyperplasia. Arterioscler Thromb Vasc Biol. 2013;33:52–59. doi: 10.1161/ATVBAHA.112.300527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pappu R, Schwab SR, Cornelissen I, Pereira JP, Regard JB, Xu Y, Camerer E, Zheng YW, Huang Y, Cyster JG, Coughlin SR. Promotion of lymphocyte egress into blood and lymph by distinct sources of sphingosine-1-phosphate. Science. 2007;316:295–298. doi: 10.1126/science.1139221. [DOI] [PubMed] [Google Scholar]
- Parrill AL, Wang D, Bautista DL, Van Brocklyn JR, Lorincz Z, Fischer DJ, Baker DL, Liliom K, Spiegel S, Tigyi G. Identification of Edg1 receptor residues that recognize sphingosine 1-phosphate. J Biol Chem. 2000;275:39379–39384. doi: 10.1074/jbc.M007680200. [DOI] [PubMed] [Google Scholar]
- Paugh SW, Paugh BS, Rahmani M, Kapitonov D, Almenara JA, Kordula T, Milstien S, Adams JK, Zipkin RE, Grant S, Spiegel S. A selective sphingosine kinase 1 inhibitor integrates multiple molecular therapeutic targets in human leukemia. Blood. 2008;112:1382–1391. doi: 10.1182/blood-2008-02-138958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paugh SW, Payne SG, Barbour SE, Milstien S, Spiegel S. The immunosuppressant FTY720 is phosphorylated by sphingosine kinase type 2. FEBS Lett. 2003;554:189–193. doi: 10.1016/s0014-5793(03)01168-2. [DOI] [PubMed] [Google Scholar]
- Pettus BJ, Bielawski J, Porcelli AM, Reames DL, Johnson KR, Morrow J, Chalfant CE, Obeid LM, Hannun YA. The sphingosine kinase 1/sphingosine-1-phosphate pathway mediates COX-2 induction and PGE2 production in response to TNF-alpha. FASEB J. 2003;17:1411–1421. doi: 10.1096/fj.02-1038com. [DOI] [PubMed] [Google Scholar]
- Pfeilschifter W, Czech-Zechmeister B, Sujak M, Mirceska A, Koch A, Rami A, Steinmetz H, Foerch C, Huwiler A, Pfeilschifter J. Activation of sphingosine kinase 2 is an endogenous protective mechanism in cerebral ischemia. Biochem Biophys Res Commun. 2011;413:212–217. doi: 10.1016/j.bbrc.2011.08.070. [DOI] [PubMed] [Google Scholar]
- Pham TH, Baluk P, Xu Y, Grigorova I, Bankovich AJ, Pappu R, Coughlin SR, McDonald DM, Schwab SR, Cyster JG. Lymphatic endothelial cell sphingosine kinase activity is required for lymphocyte egress and lymphatic patterning. J Exp Med. 2010;207(17–27):S11–S14. doi: 10.1084/jem.20091619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piali L, Froidevaux S, Hess P, Nayler O, Bolli MH, Schlosser E, Kohl C, Steiner B, Clozel M. The selective sphingosine 1-phosphate receptor 1 agonist ponesimod protects against lymphocyte-mediated tissue inflammation. J Pharmacol Exp Ther. 2011;337:547–556. doi: 10.1124/jpet.110.176487. [DOI] [PubMed] [Google Scholar]
- Ponnusamy S, Selvam SP, Mehrotra S, Kawamori T, Snider AJ, Obeid LM, Shao Y, Sabbadini R, Ogretmen B. Communication between host organism and cancer cells is transduced by systemic sphingosine kinase 1/sphingosine 1-phosphate signalling to regulate tumour metastasis. EMBO Mol Med. 2012;4:761–775. doi: 10.1002/emmm.201200244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price MM, Oskeritzian CA, Falanga YT, Harikumar KB, Allegood JC, Alvarez SE, Conrad D, Ryan JJ, Milstien S, Spiegel S. A specific sphingosine kinase 1 inhibitor attenuates airway hyperresponsiveness and inflammation in a mast cell-dependent murine model of allergic asthma. J Allergy Clin Immunol. 2012;131:501–511. doi: 10.1016/j.jaci.2012.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quancard J, Bollbuck B, Janser P, Angst D, Berst F, Buehlmayer P, Streiff M, Beerli C, Brinkmann V, Guerini D, Smith PA, Seabrook TJ, Traebert M, Seuwen K, Hersperger R, Bruns C, Bassilana F, Bigaud M. A potent and selective S1P(1) antagonist with efficacy in experimental autoimmune encephalomyelitis. Chem Biol. 2012;19:1142–1151. doi: 10.1016/j.chembiol.2012.07.016. [DOI] [PubMed] [Google Scholar]
- Reed AB, Lanman BA, Neira S, Harrington PE, Sham KK, Frohn M, Pickrell AJ, Tasker AS, Gore A, Fiorino M, Itano A, McElvain M, Middleton S, Morrison H, Xu H, Xu Y, Wong M, Cee VJ. Isoform-selective thiazolo[5,4-b]pyridine S1P1 agonists possessing acyclic amino carboxylate head-groups. Bioorg Med Chem Lett. 2012;22:1779–1783. doi: 10.1016/j.bmcl.2011.12.073. [DOI] [PubMed] [Google Scholar]
- Roberts R, Sciorra VA, Morris AJ. Human type 2 phosphatidic acid phosphohydrolases. Substrate specificity of the type 2a, 2b, and 2c enzymes and cell surface activity of the 2a isoform. J Biol Chem. 1998;273:22059–22067. doi: 10.1074/jbc.273.34.22059. [DOI] [PubMed] [Google Scholar]
- Rosen H (2007) Probe report: high throughput screening for S1P receptor agonists and antagonists, S1P2 agonist. Scripps, Florida
- Salomone S, Waeber C. Selectivity and specificity of sphingosine-1-phosphate receptor ligands: caveats and critical thinking in characterizing receptor-mediated effects. Front Pharmacol. 2011;2:9. doi: 10.3389/fphar.2011.00009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanada Y, Mizushima T, Kai Y, Nishimura J, Hagiya H, Kurata H, Mizuno H, Uejima E, Ito T. Therapeutic effects of novel sphingosine-1-phosphate receptor agonist W-061 in murine DSS colitis. PLoS ONE. 2011;6:e23933. doi: 10.1371/journal.pone.0023933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez T, Skoura A, Wu MT, Casserly B, Harrington EO, Hla T. Induction of vascular permeability by the sphingosine-1-phosphate receptor-2 (S1P2R) and its downstream effectors ROCK and PTEN. Arterioscler Thromb Vasc Biol. 2007;27:1312–1318. doi: 10.1161/ATVBAHA.107.143735. [DOI] [PubMed] [Google Scholar]
- Sanna MG, Liao J, Jo E, Alfonso C, Ahn MY, Peterson MS, Webb B, Lefebvre S, Chun J, Gray N, Rosen H. Sphingosine 1-phosphate (S1P) receptor subtypes S1P1 and S1P3, respectively, regulate lymphocyte recirculation and heart rate. J Biol Chem. 2004;279:13839–13848. doi: 10.1074/jbc.M311743200. [DOI] [PubMed] [Google Scholar]
- Sanna MG, Wang SK, Gonzalez-Cabrera PJ, Don A, Marsolais D, Matheu MP, Wei SH, Parker I, Jo E, Cheng WC, Cahalan MD, Wong CH, Rosen H. Enhancement of capillary leakage and restoration of lymphocyte egress by a chiral S1P1 antagonist in vivo. Nat Chem Biol. 2006;2:434–441. doi: 10.1038/nchembio804. [DOI] [PubMed] [Google Scholar]
- Sarkisyan G, Cahalan SM, Gonzalez-Cabrera PJ, Leaf NB, Rosen H. Real-time differential labeling of blood, interstitium, and lymphatic and single-field analysis of vasculature dynamics in vivo. Am J Physiol Cell Physiol. 2012;302:C1460–C1468. doi: 10.1152/ajpcell.00382.2011. [DOI] [PubMed] [Google Scholar]
- Sato K, Malchinkhuu E, Horiuchi Y, Mogi C, Tomura H, Tosaka M, Yoshimoto Y, Kuwabara A, Okajima F. Critical role of ABCA1 transporter in sphingosine 1-phosphate release from astrocytes. J Neurochem. 2007;103:2610–2619. doi: 10.1111/j.1471-4159.2007.04958.x. [DOI] [PubMed] [Google Scholar]
- Schmahl J, Raymond CS, Soriano P. PDGF signaling specificity is mediated through multiple immediate early genes. Nat Genet. 2007;39:52–60. doi: 10.1038/ng1922. [DOI] [PubMed] [Google Scholar]
- Schulze T, Golfier S, Tabeling C, Rabel K, Graler MH, Witzenrath M, Lipp M. Sphingosine-1-phospate receptor 4 (S1P(4)) deficiency profoundly affects dendritic cell function and TH17-cell differentiation in a murine model. FASEB J. 2011;25:4024–4036. doi: 10.1096/fj.10-179028. [DOI] [PubMed] [Google Scholar]
- Schurer SC, Brown SJ, Gonzalez-Cabrera PJ, Schaeffer MT, Chapman J, Jo E, Chase P, Spicer T, Hodder P, Rosen H. Ligand-binding pocket shape differences between sphingosine 1-phosphate (S1P) receptors S1P1 and S1P3 determine efficiency of chemical probe identification by ultrahigh-throughput screening. ACS Chem Biol. 2008;3:486–498. doi: 10.1021/cb800051m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwab SR, Pereira JP, Matloubian M, Xu Y, Huang Y, Cyster JG. Lymphocyte sequestration through S1P lyase inhibition and disruption of S1P gradients. Science. 2005;309:1735–1739. doi: 10.1126/science.1113640. [DOI] [PubMed] [Google Scholar]
- Sensken SC, Nagarajan M, Bode C, Graler MH. Local inactivation of sphingosine 1-phosphate in lymph nodes induces lymphopenia. J Immunol. 2011;186:3432–3440. doi: 10.4049/jimmunol.1002169. [DOI] [PubMed] [Google Scholar]
- Shimizu H, Takahashi M, Kaneko T, Murakami T, Hakamata Y, Kudou S, Kishi T, Fukuchi K, Iwanami S, Kuriyama K, Yasue T, Enosawa S, Matsumoto K, Takeyoshi I, Morishita Y, Kobayashi E. KRP-203, a novel synthetic immunosuppressant, prolongs graft survival and attenuates chronic rejection in rat skin and heart allografts. Circulation. 2005;111:222–229. doi: 10.1161/01.CIR.0000152101.41037.AB. [DOI] [PubMed] [Google Scholar]
- Shoham AB, Malkinson G, Krief S, Shwartz Y, Ely Y, Ferrara N, Yaniv K, Zelzer E. S1P1 inhibits sprouting angiogenesis during vascular development. Development. 2012;139:3859–3869. doi: 10.1242/dev.078550. [DOI] [PubMed] [Google Scholar]
- Skoura A, Sanchez T, Claffey K, Mandala SM, Proia RL, Hla T. Essential role of sphingosine 1-phosphate receptor 2 in pathological angiogenesis of the mouse retina. J Clin Invest. 2007;117:2506–2516. doi: 10.1172/JCI31123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snider AJ, Kawamori T, Bradshaw SG, Orr KA, Gilkeson GS, Hannun YA, Obeid LM. A role for sphingosine kinase 1 in dextran sulfate sodium-induced colitis. FASEB J. 2009;23:143–152. doi: 10.1096/fj.08-118109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snider AJ, Orr Gandy KA, Obeid LM. Sphingosine kinase: role in regulation of bioactive sphingolipid mediators in inflammation. Biochimie. 2010;92:707–715. doi: 10.1016/j.biochi.2010.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strub GM, Maceyka M, Hait NC, Milstien S, Spiegel S. Extracellular and intracellular actions of sphingosine-1-phosphate. Adv Exp Med Biol. 2010;688:141–155. doi: 10.1007/978-1-4419-6741-1_10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki S, Enosawa S, Kakefuda T, Shinomiya T, Amari M, Naoe S, Hoshino Y, Chiba K. A novel immunosuppressant, FTY720, with a unique mechanism of action, induces long-term graft acceptance in rat and dog allotransplantation. Transplantation. 1996;61:200–205. doi: 10.1097/00007890-199601270-00006. [DOI] [PubMed] [Google Scholar]
- Takuwa N, Ohkura S, Takashima S, Ohtani K, Okamoto Y, Tanaka T, Hirano K, Usui S, Wang F, Du W, Yoshioka K, Banno Y, Sasaki M, Ichi I, Okamura M, Sugimoto N, Mizugishi K, Nakanuma Y, Ishii I, Takamura M, Kaneko S, Kojo S, Satouchi K, Mitumori K, Chun J, Takuwa Y. S1P3-mediated cardiac fibrosis in sphingosine kinase 1 transgenic mice involves reactive oxygen species. Cardiovasc Res. 2010;85:484–493. doi: 10.1093/cvr/cvp312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarrason G, Auli M, Mustafa S, Dolgachev V, Domenech MT, Prats N, Dominguez M, Lopez R, Aguilar N, Calbet M, Pont M, Milligan G, Kunkel SL, Godessart N. The sphingosine-1-phosphate receptor-1 antagonist, W146, causes early and short-lasting peripheral blood lymphopenia in mice. Int Immunopharmacol. 2011;11:1773–1779. doi: 10.1016/j.intimp.2011.07.004. [DOI] [PubMed] [Google Scholar]
- Teijaro JR, Walsh KB, Cahalan S, Fremgen DM, Roberts E, Scott F, Martinborough E, Peach R, Oldstone MB, Rosen H. Endothelial cells are central orchestrators of cytokine amplification during influenza virus infection. Cell. 2011;146:980–991. doi: 10.1016/j.cell.2011.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thangada S, Khanna KM, Blaho VA, Oo ML, Im DS, Guo C, Lefrancois L, Hla T. Cell-surface residence of sphingosine 1-phosphate receptor 1 on lymphocytes determines lymphocyte egress kinetics. J Exp Med. 2010;207:1475–1483. doi: 10.1084/jem.20091343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomson A. FTY720 in multiple sclerosis: the emerging evidence of its therapeutic value. Core Evid. 2006;1:157–167. [PMC free article] [PubMed] [Google Scholar]
- Tomsig JL, Snyder AH, Berdyshev EV, Skobeleva A, Mataya C, Natarajan V, Brindley DN, Lynch KR. Lipid phosphate phosphohydrolase type 1 (LPP1) degrades extracellular lysophosphatidic acid in vivo. Biochem J. 2009;419:611–618. doi: 10.1042/BJ20081888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tonelli F, Lim KG, Loveridge C, Long J, Pitson SM, Tigyi G, Bittman R, Pyne S, Pyne NJ. FTY720 and (S)-FTY720 vinylphosphonate inhibit sphingosine kinase 1 and promote its proteasomal degradation in human pulmonary artery smooth muscle, breast cancer and androgen-independent prostate cancer cells. Cell Signal. 2010;22:1536–1542. doi: 10.1016/j.cellsig.2010.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urbano M, Guerrero M, Velaparthi S, Crisp M, Chase P, Hodder P, Schaeffer MT, Brown S, Rosen H, Roberts E. Discovery, synthesis and SAR analysis of novel selective small molecule S1P4-R agonists based on a (2Z,5Z)-5-((pyrrol-3-yl)methylene)-3-alkyl-2-(alkylimino)thiazolidin-4-one chemotype. Bioorg Med Chem Lett. 2011;21:6739–6745. doi: 10.1016/j.bmcl.2011.09.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vessey DA, Li L, Jin ZQ, Kelley M, Honbo N, Zhang J, Karliner JS. A sphingosine kinase form 2 knockout sensitizes mouse myocardium to ischemia/reoxygenation injury and diminishes responsiveness to ischemic preconditioning. Oxid Med Cell Longev. 2011;2011:961059. doi: 10.1155/2011/961059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visentin B, Vekich JA, Sibbald BJ, Cavalli AL, Moreno KM, Matteo RG, Garland WA, Lu Y, Yu S, Hall HS, Kundra V, Mills GB, Sabbadini RA. Validation of an anti-sphingosine-1-phosphate antibody as a potential therapeutic in reducing growth, invasion, and angiogenesis in multiple tumor lineages. Cancer Cell. 2006;9:225–238. doi: 10.1016/j.ccr.2006.02.023. [DOI] [PubMed] [Google Scholar]
- Walsh KB, Teijaro JR, Wilker PR, Jatzek A, Fremgen DM, Das SC, Watanabe T, Hatta M, Shinya K, Suresh M, Kawaoka Y, Rosen H, Oldstone MB. Suppression of cytokine storm with a sphingosine analog provides protection against pathogenic influenza virus. Proc Natl Acad Sci USA. 2011;108:12018–12023. doi: 10.1073/pnas.1107024108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walzer T, Chiossone L, Chaix J, Calver A, Carozzo C, Garrigue-Antar L, Jacques Y, Baratin M, Tomasello E, Vivier E. Natural killer cell trafficking in vivo requires a dedicated sphingosine 1-phosphate receptor. Nat Immunol. 2007;8:1337–1344. doi: 10.1038/ni1523. [DOI] [PubMed] [Google Scholar]
- Wei SH, Rosen H, Matheu MP, Sanna MG, Wang SK, Jo E, Wong CH, Parker I, Cahalan MD. Sphingosine 1-phosphate type 1 receptor agonism inhibits transendothelial migration of medullary T cells to lymphatic sinuses. Nat Immunol. 2005;6:1228–1235. doi: 10.1038/ni1269. [DOI] [PubMed] [Google Scholar]
- Yatomi Y, Ruan F, Hakomori S, Igarashi Y. Sphingosine-1-phosphate: a platelet-activating sphingolipid released from agonist-stimulated human platelets. Blood. 1995;86:193–202. [PubMed] [Google Scholar]
- Zhang N, Sundberg JP, Gridley T. Mice mutant for Ppap2c, a homolog of the germ cell migration regulator wunen, are viable and fertile. Genesis. 2000;27:137–140. doi: 10.1002/1526-968x(200008)27:4<137::aid-gene10>3.0.co;2-4. [DOI] [PubMed] [Google Scholar]
- Zhang ZY, Zhang Z, Zug C, Nuesslein-Hildesheim B, Leppert D, Schluesener HJ. AUY954, a selective S1P(1) modulator, prevents experimental autoimmune neuritis. J Neuroimmunol. 2009;216:59–65. doi: 10.1016/j.jneuroim.2009.09.010. [DOI] [PubMed] [Google Scholar]
- Zhou J, Saba JD. Identification of the first mammalian sphingosine phosphate lyase gene and its functional expression in yeast. Biochem Biophys Res Commun. 1998;242:502–507. doi: 10.1006/bbrc.1997.7993. [DOI] [PubMed] [Google Scholar]