

Abstract
For a virus to establish persistence in the host, it has to exploit the host immune system such that the active T-cell responses against the virus are curbed. On the other hand, the goal of the immune system is to clear the virus, following which the immune responses need to be downregulated, by a process known as immunoregulation. There are multiple known immunoregulatory mechanisms that appear to play a role in persistent viral infections. In the recent past, IL-10 and PD-1 have been identified to be playing a significant role in the regulation of antiviral immune responses. The evidence that viruses can escape immunologic attack by taking advantage of the host’s immune system is found in LCMV infection of mice and in humans persistently infected with HIV and HCV. The recent observation that the functionally inactive T-cells during chronic viral infections can be made to regain their cytokine secretion and cytolytic abilities is very encouraging. Thus, it would be likely that neutralization negative immune regulation during persistent viral infection would result in the preservation of effector T-cell responses against the virus, thereby resulting in the elimination of the persistent infection.
Key words: Immunoregulation, Persistent infection, IL-10, PD-1, LCMV
Virus and Disease
Most viruses that infect humans are usually cleared by the host immune system in a rapid (acute) manner, either (a) by the release of proinflammatory cytokines or (b) by killing the infected cells to limit the replication of the virus [1]. Such acute responses involve a first step innate immune response (macrophages, dendritic cells [DCs], and natural killer [NK] cells) [2], followed by the generation of adaptive immunity (T and B cells). Macrophages and DCs act as phagocytes thus eliminating the infected cells, and NK cells directly kill the infected cells. In adaptive immunity, which kicks in 3–5 days after infection, presentation of viral antigen by macrophages and/or DCs or by infected cells that present viral peptides on host MHC molecules results in the activation of T-cells. These T-cells then mount a viral antigen-specific immune response and kill the infected cells either in a cell–cell contact-dependent manner (perforin/granzyme mediated) or through the release of inflammatory cytokines TNF-α or IFN-γ. These cytokines can also act on the virus directly and prevent its replication. Some of the viral antigen-specific T-cells then survive for a very long time creating a memory pool.
Most, if not all, viral infections trigger the production of interferon’s (IFNs) by the host immune system. IFNs then primarily limit viral replication to prevent damage to the infected cell. Type-I IFNs are essential part of innate resistance to viral infections as is evident in the case B6 mice infected with MHV-1; absence of type I IFN-mediated signaling in IFN-αβR-KO mice results in progressive loss of body weight, culminating in the death of the mice by day 5 postinfection [3].
Intravenous infection of adult mice with lymphocytic choriomeningitis virus (LCMV)-Armstrong results in an acute infection, which is resolved by virus-specific CD8 and CD4 T cells within 8–10 days [4]. However, unlike in acute viral immune response where antiviral CTLs are present at all times examined; in persistent infections, whether it be the effector phase or memory phase, antiviral CD8 T-cells are rarely found in adult mice or in humans infected early in life [1]. A more detailed discussion of innate and adaptive immune responses in viral infections is beyond the scope of this chapter.
Some virus, for instance HIV and hepatitis B virus (HBV)/hepatitis C virus (HCV) in humans and LCMV in mice, escape killing by the host’s immune system, either by avoidance or manipulation and establish persistent infection in the host. To establish a persistent infection, the virus has to overcome several host-dependent factors. One unique strategy that viruses employ is that instead of killing its host cell, the virus causes little to no damage so that it can escape detection by the immune system. The virus then either continues to replicate inside the cell (HIV) or it establishes latency (EBV). Viruses can alter or interfere with the processing of viral peptides by professional antigen presenting cell (APC), a requirement for activation and expansion of the T cells that normally remove infected cells. Additionally, viruses can also inhibit the differentiation of antigen-presenting conventional DCs and can infect effector T and B cells directly [1]. Some viruses also encode miRNAs to regulate their replication or latency or to manipulate or evade host immune responses [5]. During latency, miRNAs keep protein levels of viral genes to a minimum, facilitating evasion of immune surveillance [6]. Finally, viruses also exploit the host’s immunoregulatory mechanisms such that the viral antigen-specific T-cells now become either anergic or are deleted and thus fail to clear the viral infection. This strategy is more detrimental to the host.
Persistent viral infections, as would be expected, are a severe problem to humans, as is evident in the case infection with HIV; persistent infection with HIV leads to the exhaustion of CD4 T-cells thereby rendering the host susceptible to a variety of secondary infections, which ultimately lead to the demise of the host. However, such persistent infections, if are actively sought by the host immune system, due to the continued activation of T-cells can lead to severe immune pathology, wherein the host’s immune system destroys the host’s own tissue.
There is a copious amount of data that are published, which provides evidence for the deleterious effects of an over-active immune system causing sever immune pathology in persistent viral infections. For instance, overexuberant immune response is implicated in fatal H5N1 influenza [7] and a dysregulated immune response contributes to clinical disease in patients with SARS [8, 9]. Similarly, in mice, excessive T-cell responses contribute to the tissue damage that occurs during the process of virus clearance in infections with LCMV, HSV, RSV [10–12], and MHV-1 [13].
LCMV as a Model to Study Immune Regulation
In our laboratory, we have extensively used LCMV as a model for establishing persistent infection in mice [14–20]. It is easier to dissect the various immunoregulatory mechanisms involved in viral infections using LCMV due to the availability of two different strains; one that establishes acute infection (LCMV-Armstrong) and is readily cleared from the host and the other that establishes persistent infection (LCMV-clone 13) for a very long time. Armstrong and clone 13 viruses share identical CD4 and CD8 T-cell epitopes but are phenotypically very distinct, replicate in different cell types, and have different clearance rates [21]. Further, since LCMV is not a cytolytic virus, it allows to distinctly examine the effects caused by the immune response-mediated damage to the host tissue [22].
Persistent viral infections ultimately result in immunosuppression in the host where the CTLs lose functionality by either functional inactivation and/or physical deletion [1]. Several different mechanisms, which act either individually or in combination, including clonal exhaustion, secretion of immunosuppressive cytokines such as IL-10 and TGF-β, and overexpression of programmed death 1 (PD-1) on activated T-cells, contribute to the failure of T-cell responses during persistent viral infections. In LCMV-clone 13 model of persistent infection, multiple immunoregulatory mechanisms are activated, including infection/impairment of DCs, and a global inactivation of the virus-specific T-cell response [23–27]. The antiviral CD4 and CD8 T-cells are either physically deleted or persist in an inactivated state and are nonresponsive toward viral antigens [23–27]. With respect to the significance of such inactivation in human patients, in HIV infection, CD4 T cells that can produce TNF-α, interleukin 2 (IL-2), and IFN-γ are conspicuously absent. Consequently, in HIV+ patients, the presence of functionally active CD4 T-cells that can produce the cytokines TNF-α, IFN-γ, and IL-2 directly correlates with low viremia [28]. Similarly, in mouse models of persistent infection, a significant three- and fourfold decrease in TNF-α and IL-2 producing cells, respectively, was observed in the spleens of clone 13 infected mice by day 100 postinfection [23].
However, unlike it was believed before, it is increasingly apparent that in persistent infections, the effector T-cells are not deleted but exist in a functionally inactive state. Thus, it was interesting to note that despite prolonged periods of inactivation, the remaining CD4 and CD8 T-cells still retain the ability to recover their functional activity [29]. This suggests that, in persistent viral infections, the host’s immune system can still be manipulated such that the host can reestablish immunity against the virus. Thus, functional preservation of CD4 and CD8 T-cells during viral persistence is the ultimate mode of achieving strong antiviral immunity and viral clearance.
Immune Regulation
General
As discussed previously, for a virus to establish persistence in the host, it has to exploit the host immune system such that the active T-cell responses against the virus are curbed. On the other hand, the goal of the immune system is to clear the virus, failing which the immune responses need to be downregulated so as to avoid unnecessary damage to the host tissue, by a process known as immunoregulation. The various known immunoregulatory mechanisms comprise (a) activation or induction of Tregs, (b) expression of inhibitory molecules such as programmed cell death-1 (PD-1) on activated T-cells, and (c) production of suppressor cytokines such as TGF-β or IL-10. In the recent past, among the major immunoregulatory molecules identified so far [15, 30–32], IL-10 [15, 32] and PD-1 [30] appear to be playing a significant role in the regulation of antiviral immune responses and are thus being actively investigated. The confounding aspect of all these different factors is that they not only regulate antiviral immune responses but they are also involved in the regulation of autoimmune responses.
The evidence that viruses can escape immunologic attack by taking advantage of the host’s immune system is found in LCMV infection of mice [15, 30, 32] and in persistently infected with HIV and HCV [33–37]. Thus, it is important to understand the effectors involved in active immunoregulation, such that we can define how a virus exploits the host’s immunoregulatory mechanism and thereby design drugs that can circumvent this virally induced immunoregulation.
Immune Regulation by Innate Immunity
This aspect of the immune system has been recently reviewed, for details refer McGill et al. [2]. The innate immune system mainly comprises macrophages, DCs, and NK cells. Macrophages and DCs are primarily involved in antigen presentation to the T-cells, with DCs usually being the more efficient APC. NK cells are primarily involved in the direct killing of virally infected cells as part of the innate immune protection. However, as with adaptive immune system, hyperactive innate immune system could very well lead to tissue/organ damage. A direct evidence for innate immune-mediated pathology is evident in the case of fatal H5N1 influenza [7]. Here, we will focus only on those cell populations that exhibit immune regulatory function.
Macrophages
Although macrophages are not the most efficient of APCs, they are very efficient phagocytes. And thus, it is believed that macrophages may be a predominant cell population contributing to IAV-associated immunopathology [39]. The first evidence for the role of macrophages in immune regulation was evident in alveolar macrophages that exist in a relatively quiescent state during homeostasis but have a regulatory phenotype in the lungs [40]. In addition, Dillon et al. showed that stimulation of macrophages through the TLR2 receptor induces macrophages that promote immunological tolerance through the production of TGF-β1 [41]. And finally, macrophages have been shown to suppress the induction of innate and adaptive immunity [42–44].
Dendritic Cells
DCs are probably the most efficient APCs of the immune system, both in animal models and humans. Therefore, the initiation of effective Ag-specific immunity to pathogens is a hallmark of DC function. However, it was also demonstrated that DCs can induce and maintain self-antigen-specific tolerance in the periphery [45]. Further, DCs can actively induce T-cell anergy [46, 47], T-cell suppression [47, 48], and generation of Tregs [49]. As they did for macrophages, Dillon et al. also showed that stimulation through the TLR2 receptor on DCs induces regulatory DC [41]. Further, TLR2 activation in mice results in IL-10 upregulation and Treg survival [50, 51]. Interestingly, incubation of DCs in the presence of IL-10 generates DC with tolerogenic properties [52]. Thus, DCs, when activated through the TLR2 receptor, appear to acquire regulatory phenotype and indeed are able to suppress the activation antigen-specific T-cells.
In support for the role of DC in immune regulation in viral infections, in in vitro experiments, it was found that the infection of DCs with high MOI results in truncated CD8 T-cell responses and increased IL-12p40 production [53]. Further, such high MOI infection also results in an increased production of the antiinflammatory cytokines IL-10 and TGF-β [54]. Finally, in IAV infections, rather than inducing T-cell activation, when influenza antigen-bearing immature DCs encounter naïve T cells, they induce tolerance to viral antigens [55, 56]. The immunoregulatory role played by DCs in other viral infections needs to be explored.
Thus, under certain conditions, for instance activation through TLR2, both DCs and macrophages appear to acquire an immunoregulatory function. However, whether macrophages and DCs can exhibit similar activity in a majority of persistent viral infections needs to be determined. Be as it may, it would be interesting to speculate that since viral infections result in TLR2 activation, at least in infections with some viruses (chronic infections), such TLR2 activation induces tolerogenic DCs and/or macrophages, which prevent further activation of antiviral T-cells.
Immune Regulation by Adaptive Immunity
CD4+ Tregs
One of the most extensively studied cell population in immunology is CD4+CD25+ regulatory T-cells (Tregs), because of their demonstrated importance in the events leading to autoimmunity. Thus, doing a formal detailed review of Tregs takes a very extensive review of literature and is therefore beyond the scope of this review article. But, for those who are interested, for a detailed review of Tregs, refer to Sakaguchi et al. [57].
There exist mainly two types of Tregs, the natural Tregs (nTregs) that originate from the thymus and the induced Tregs (iTregs) that arise in the periphery. Tregs, either natural or induced, are characterized by the expression of the transcription factor Foxp3. Thus, it was found that the ectopic Foxp3 expression in normal T-cells enables these cells to exhibit suppressor function in vitro and in vivo [58]. Although the significance still needs to be determined, it was recently demonstrated that the expression of Foxp3 in Treg cells does not destine the T-cell to be a Treg for life, since a subset of Treg cells appear to downregulate Foxp3 expression (exFoxp3 cells) [59].
nTregs constitute 4–5% of peripheral CD4 T cells and are important in the maintenance of immunological self-tolerance in the periphery [60]. CD25+ T cells appear in the periphery after day 3 and rapidly increase to adult levels within 2 weeks (reviewed in [61, 62]). CD4+CD25+ Tregs are actively involved in the regulation of antiviral immunity in order to limit immune pathology in the host [63–67] but thereby they also inhibit viral clearance [68–70].
Tregs have been implicated to play a role in a number of different viral infections, in both mice and humans, including but not limited to Friends’ virus or HSV (in mice) and HCV and HIV (in humans) [12, 68–72]. To further demonstrate the importance of Tregs in viral immunity (or immune regulation), Tregs are relatively deficient in patients with severe dengue infection when compared to those with mild disease [73].
Tregs exert their suppressive function through a combination of various mechanisms including cell-contact (killing and/or functional modulation of APC and/or responder T cells (reviewed in [74–76])) and soluble factors (secretion of immunosuppressive cytokines, IL-10 and TGF-β [77, 78]). However, it appears that direct cell-contact-mediated suppression is critical for the action of Tregs. Tregs inhibit, via CTLA-4, the upregulation of CD80 and CD86 by immature DC upon antigenic stimulation and also downregulate the expression of CD80/CD86 by mature DC [79, 80], thus indirectly affecting antiviral CD4 and CD8 T-cell responses. Another plausible mechanism by which Tregs mediate their regulatory activity has been proposed; since Tregs cannot produce IL-2 and thus cannot expand in an autocrine fashion [81], but constitutively express high levels of IL-2R (CD25), they likely deprive effector T-cells of IL-2 in the vicinity and thus provide hindrance to the activation/expansion of responder T cells [82].
As such, the take home point is that natural CD4+CD25+ Tregs have only a minimal impact on viral elimination per se but they play an important part in limiting collateral tissue damage usually caused by strong antiviral T-cell responses [12, 66]. Thus, they are one of the most important players in mediating immune regulation, either in antiviral immunity or in autoimmunity.
PD-1/PD-L1
Based on few recent findings [30, 31, 35, 36, 38, 83–86], it is increasingly apparent that the PD-1/PDL-1 axis plays a major role in regulating immune responses during antiviral T-cell responses. PD-1 is an inhibitory receptor of the CD28 family and has two ligands PD-L1 and PD-L2, but most of the immunoregulatory activity of PD-1 is brought upon by its interaction with PD-L1 (reviewed in [87]). Accordingly, PD-L1−/− mice infected with LCMV clone 13 died owing to immunopathologic damage [30].
During chronic infection with LCMV-clone 13, PD-1 is upregulated at both RNA and protein level, in exhausted LCMV-specific CD8 T cells. Further, PD-1 expression continued to increase in exhausted CD8 T cells and the high level of expression was sustained resulting in the failure of these to clear infection [30, 31]. Blocking the activity of PD-1, using an antibody against PD-L1, enhanced virus-specific CD8 T cell proliferation, production of IFN-γ, and their lytic ability [30]. And further, blocking PD-L1 results in enhanced epitope-specific responses to therapeutic vaccination [84].
In our laboratory, using LCMV-induced T1D model, the RIP-GP-LCMV mice, we found that the virus-induced upregulation of PD-L1 in prediabetic mice prevents the expansion of diabetogenic CD8+ T-cells, thus confirming the importance of PD-1 in regulating antiviral CD8 T-cell responses [85]. Interestingly, such PD-L1 upregulation also resulted in synergistic enhancement in the capacity of virally enhanced CD4+CD25+ Tregs to mediate immunoregulation [83].
Finally, in humans infected with HIV who have increased PD-1, an association exists between in vivo T-cell unresponsiveness and restoration of T-cell function in vitro with antibody to PD-L1 [36, 85, 86]. Thus, the PD-1–PD-L1 axis is crucial for sustaining suppression of CD8 T cells during persistent infection.
CTLA-4
Costimulation through the CD28 molecule is essential for the generation of efficient antiviral T-cell responses. Consequently, the prevention of costimulation during the chronic phase of clone 13 infections diminishes antiviral CD8 T cell responses and prevents control of viral replication [88].
CTLA-4, an inhibitory receptor belonging to the B7-CD28 family of costimulatory molecules (reviewed in [89]), is constitutively expressed on murine Foxp3+ Treg, whether thymus derived or induced in the periphery [90–92] and can mediate the suppression of effector T-cell responses. CTLA-4 is also upregulated on activated T cells, and competes with CD28 for binding to the same ligands, B7-1 (CD80) and B7-2 (CD86). However, instead of inducing activation of T-cells, such as CD28, CTLA-4 antagonizes the production of IL-2 and inhibits T-cell activation.
While the importance of CTLA-4-mediated regulation T-cell responses in tumor immunology is well known, its role in persistent viral infections is still debatable. In mice infected with LCMV, CTLA-4 expression or lack thereof by T cells has little or no effect on antiviral immunity, either in the effector or in memory generation during acute viral infection [30]. And, similar to acute infection, CTLA-4 was also not involved in T-cell exhaustion in mice chronically infected with LCMV clone 13 [30].
The strongest argument for the involvement of CTLA-4 in the regulation of antiviral immunity comes from the studies of HIV-infected humans (reviewed in [93]) or SIV-infected monkeys [94]. Arguing for a role of CTLA-4 in antiviral immunity, it was found that CTLA-4 expression was high in HIV-specific CD4 but not CD8 T cells in humans with acute HIV infection but low in people who were able to spontaneously control infection [95]. More specifically, consistent with the finding that CTLA-4 engagement downregulates IL-2 secretion [96], HIV-Gag-specific CD4 T cells that produced IL-2 in addition to IFN-γ had less CTLA-4, while those CD4 T cells that produced only IFN-γ had higher CTLA-4 expression [95]. And finally, it was shown that blockade of CTLA-4 with antibodies resulted in an augmentation of both SIV-specific CD4 and CD8 efffector T-cell responses [94].
Thus, CTLA-4 plays a major role in immunoregulation by preventing costimulation through CD28 receptor leading to inefficient activation of CD8 T-cells, in some, if not all, persistent viral infections. Hence, therapeutic approaches aimed at preventing the action of CTLA-4 during some persistent infections, where CTLA-4 is known to play a role, will be beneficial to a subgroup of patients.
TGF-β
With the discovery that Tregs make an immunoregulatory cytokine TGF-β, quite a few laboratories performed extensive analysis of the role played by this cytokine in immune regulation (reviewed in [97]). It is now well known that TGF-β controls various aspects of the inflammatory response through the regulation of chemotaxis, activation, and survival of T cells, NK cells, and APCs and is crucial for the control of antiviral immunity (reviewed in [97]). TGF-β also exhibits highly potent inhibitory effects on cytolytic functions, cytokine production, and proliferation of T-cells (reviewed in [98]). Conversely, TGF-β also exhibits antiapoptotic effects [99–101] on T-cells best exemplified in the case of the effector T-cells. When cultured in the presence of TGF-β and IL-2 effector, CD4 T-cells exhibit enhanced accumulation (owing to increased proliferation and survival) and even better prolonged expansion [99]. And finally, TGF-β, produced by Tregs, can act in an autocrine fashion to induce further expansion of Tregs from which it is produced. Also, TGF-β can induce the conversion of naïve CD4+CD25− T cells into CD4+CD25+ T cells by the induction of Foxp3 [102]. Thus TGF-β is extensively used to induce the expansion of Treg in in vitro cultures. Data from our laboratory as well as others have now shown that such in vitro expanded Tregs are able to adoptively confer protection in virus-induced T1D model (reviewed in [103]).
While the role of TGF-β in regulating immune responses is extensively known in a variety of autoimmune, infectious (bacterial) and other diseases evidence of TGF-β role in regulating antiviral immunity is scarce. Still, in chronic infections, such as influenza, HIV, HBV, HCV, and others, there is evidence of increased TGF-β production. In a virus-induced T1D mouse model, data from our laboratory showed that on one hand TGF-β suppresses naïve CD8+ T cell activation, while on the other hand it enhances the survival and function of antigen-experienced/memory CD8 T cells [104]. Most importantly, systemic administration of TGF-β was shown to protect from T1D [105].
Interestingly, several recent studies have convincingly demonstrated that TGF-β plays an essential role in the development of a newly discovered highly proinflammatory T-cells, the IL-17-secreting T cells (Th17) [106]. Although, TGF-β cannot induce the conversion of naïve cells to Th17 by itself, together with IL-6, TGF-β facilitates the differentiation of T-cells into Th17 cells [107]. The most important aspect of TGF-β-induced effect is that TGF-β in conjunction with IL-2 induces the differentiation of naïve T-cells into Foxp3+ Tregs and this IL-2 inhibits the differentiation into Th17 cells [108].
Thus TGF-β is a very interesting cytokine, in that, in the presence of IL-2, it induces the generation of regulatory T-cells, CD4+ Foxp3+ Tregs, but the same TGF-β, in the presence of IL-6, induces the generation of proinflammatory Th17 T-cells. Hence, TGF-β apparently is very important in the regulation of antiviral immunity because while Tregs (as discussed above) are clearly involved in preventing tissue damage due to immune pathology, Th17 cells are highly proinflammatory and are known to be playing a significant role in autoimmune diseases. Thus, this dual functionality of TGF-β needs further elucidation.
IL-10
One of the most studied regulatory cytokine other than TGF-β is IL-10. IL-10 exerts its suppressive effect on macrophages [109] and T cells by blocking CD28 and ICOS signals in a rapid signal transduction cascade [110].
The best example for the role of IL-10, in regulating immune responses in viral infections, was provided in the last few years by a couple of laboratories, including data from our own laboratory. It is known that by 9 days after infection with LCMV clone 13, which establishes persistent infection in adult mice, both virus-specific CD4+ and CD8+ T-cells become unresponsive. Data from our laboratory showed that the use of an antibody to the IL10 receptor results in very efficient control of viral persistence, even leading to the elimination of virus. By removing IL-10 inhibitory effect, in this case on T-cell anergy, we were able to induce host resistance such that the host could now resolve persistent LCMV infection [15]. This was followed by further proof that such IL-10 acts on effector T-cells [32]. When IL-10 was blocked with antibody to IL-10 receptors, T-cell numbers increase sufficiently to clear virus from blood and tissues; these T-cells that were anergized because of the persistent virus infection now regained their ability to produced proinflammatory cytokines like TNF-α, IFN-γ and thus were able to reacquire their normal effector T-cell functions [32].
There was a significant increase in the frequency of IL-10 producing cells in both spleen and liver during persistent clone 13 infection [23]. Accordingly, significantly more IL-10 RNA was found in the spleens of clone 13-infected mice than that of Armstrong-infected mice [32]. CD4+CD25+ Foxp3+ regulatory T-cells are thought to be the major producers of IL-10. However, in clone 13 persistent infection, although CD4 T-cells produced some IL-10 initially, they are not responsible for the majority of IL-10, since IL-10 was produced in significant quantities even after the functional inactivation of CD4 T-cells; further these CD4 T-cells stopped making any IL-10 after functional inactivation [32]. Interestingly, instead of the CD4 T-cells, it appears that DCs are the major producers of IL-10 in clone 13 infection [32]. Further, using IL-10−/− mice, the authors found that the lack of IL-10 resulted in the preservation of viral antigen (NP396-404)-specific CD8 T-cell responses that are normally lost during persistent infection [32]. And, as we had reported originally [15], antibody blockade of IL-10 prevented viral persistence [32], thus definitely showing that IL-10 plays a major role in immunoregulation during persistent viral infection with LCMV.
Although it appears that IL-10 may be acting directly on T-cells, the involvement of Tregs in immunoregulation but the their failure to make IL-10 during persistent LCMV infection suggests that other than acting on T-cells, IL-10 may also be playing an immunoregulatory role by directly altering the antigen-presenting capabilities of the APCs. However, this hypothesis needs further testing since it was found that APC functionality is normal in terms of antigen presentation in clone 13 infection [23].
Concluding Remarks
Persistent viral infections are a real threat to human health during a variety of viral infections with HIV, HBV, and HCV. The major role players responsible for the immunosuppression that happens during persistent infections have been identified as PD-1/PD-L1 and IL-10, with TGF-β and CTLA-4 also playing a role to some extent (Fig. 1). The recent observation that the functionally inactive T-cells during chronic viral infections can be made to regain their cytokine secretion and cytolytic abilities by neutralizing IL-10 is very encouraging [32]. More importantly, when IL-10 [111] or PD-L1 [84] was first neutralized, therapeutic vaccination against ongoing persistent LCMV infection was highly effective. Further, IL-10R blockade resulted in the ability of an otherwise ineffective DNA vaccination to be successful such that the DNA vaccination elicited a fourfold increase in the number of functional CD8 T cells [111]. Thus, it would be likely that neutralization of either IL-10 or PD-1/PD-L1 (depending on the virus) during the persistent viral infection would result in the preservation of effector T-cell responses against the virus thereby resulting in the elimination of the persistent infection.
Fig. 1.
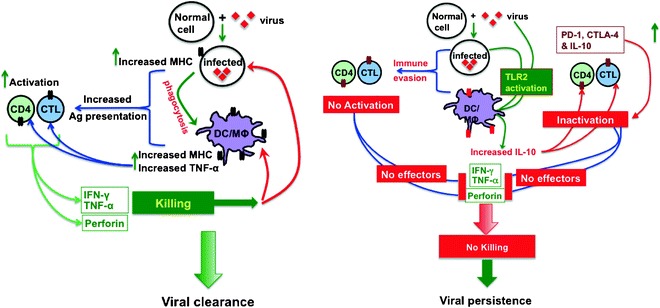
Immune regulation during persistent viral infection. In an acute infection (left), when a virus infects a normal cell, it is phagocytosed by antigen presenting cell (APC) (dendritic cell [DC] or MΦ). The APCs efficiently present the viral antigens to CD4 and CD8 cells, which either release proinflammatory cytokines (TNF-α, IFN-γ) or upregulate perforin, which kills the infected cells, ultimately leading to the elimination of the virus. However, in persistent infection (right), due to a number of reasons, including miRNAs and molecular mimicry, viruses evade immune recognition. Thus, the CD4 and CD8 cells are not sufficiently activated and the virus avoids immune-mediated elimination. On the other hand, APCs (MΦ and DCs), upon TLR2 activation, upregulate IL-10; IL-10 exhibits significant immunosuppressive effects leading to the inactivation of CD4 and CD8 T-cells. Further, activated T-cells also upregulate CTLA-4 that prevents costimulation, thereby leading to inactivation. And, finally, virus antigen-specific T-cells also upregulate PD-1 which, upon engagement with its ligand PD-L1, signals the death of that T-cell. These events, help to avoid immune response mediated host tissue damage but at the cost of establishing viral persistence.
Contributor Information
Maria Cristina Cuturi, Phone: (33)2 40 08 74 10, FAX: (33) 2 40 08 74 11, Email: Maria-Cristina.Cuturi@univ-nantes.fr.
Ignacio Anegon, Phone: (33) 2 40 08 74 15, FAX: (33) 2 40 08 74 11, Email: Ignacio.Anegon@univ-nantes.fr.
Matthias G. von Herrath, Email: matthias@liai.org
References
- 1.Oldstone MB. Anatomy of viral persistence. PLoS Pathog. 2009;5:e1000523. doi: 10.1371/journal.ppat.1000523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.McGill J, Heusel JW, Legge KL. Innate immune control and regulation of influenza virus infections. J Leukoc Biol. 2009;86:803–812. doi: 10.1189/jlb.0509368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Khanolkar A, Hartwig SM, Haag BA, Meyerholz DK, Epping LL, Haring JS, Varga SM, Harty JT. Protective and pathologic roles of the immune response to mouse hepatitis virus type 1: implications for severe acute respiratory syndrome. J Virol. 2009;83:9258–9272. doi: 10.1128/JVI.00355-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fung-Leung WP, Kundig TM, Zinkernagel RM, Mak TW. Immune response against lymphocytic choriomeningitis virus infection in mice without CD8 expression. J Exp Med. 1991;174:1425–1429. doi: 10.1084/jem.174.6.1425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gottwein E, Cullen BR. Viral and cellular microRNAs as determinants of viral pathogenesis and immunity. Cell Host Microbe. 2008;3:375–387. doi: 10.1016/j.chom.2008.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Huang J, Wang F, Argyris E, Chen K, Liang Z, Tian H, Huang W, Squires K, Verlinghieri G, Zhang H. Cellular microRNAs contribute to HIV-1 latency in resting primary CD4+ T lymphocytes. Nat Med. 2007;13:1241–1247. doi: 10.1038/nm1639. [DOI] [PubMed] [Google Scholar]
- 7.de Jong MD, Simmons CP, Thanh TT, Hien VM, Smith GJ, Chau TN, Hoang DM, Chau NV, Khanh TH, Dong VC, Qui PT, Cam BV, Ha do Q, Guan Y, Chinh JS, Hien NT, Farrar J. Fatal outcome of human influenza A (H5N1) is associated with high viral load and hypercytokinemia. Nat Med. 2006;12:1203–1207. doi: 10.1038/nm1477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cameron MJ, Ran L, Xu L, Danesh A, Bermejo-Martin JF, Cameron CM, Muller MP, Gold WL, Richardson SE, Poutanen SM, Willey BM, DeVries ME, Fang Y, Seneviratne C, Bosinger SE, Persad D, Wilkinson P, Greller LD, Somogyi R, Humar A, Keshavjee S, Louie M, Loeb MB, Brunton J, McGeer AJ, Kelvin DJ. Interferon-mediated immunopathological events are associated with atypical innate and adaptive immune responses in patients with severe acute respiratory syndrome. J Virol. 2007;81:8692–8706. doi: 10.1128/JVI.00527-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Jones BM, Ma ES, Peiris JS, Wong PC, Ho JC, Lam B, Lai KN, Tsang KW. Prolonged disturbances of in vitro cytokine production in patients with severe acute respiratory syndrome (SARS) treated with ribavirin and steroids. Clin Exp Immunol. 2004;135:467–473. doi: 10.1111/j.1365-2249.2003.02391.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hussell T, Pennycook A, Openshaw PJ. Inhibition of tumor necrosis factor reduces the severity of virus-specific lung immunopathology. Eur J Immunol. 2001;31:2566–2573. doi: 10.1002/1521-4141(200109)31:9<2566::AID-IMMU2566>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- 11.Oldstone MB. Biology and pathogenesis of lymphocytic choriomeningitis virus infection. Curr Top Microbiol Immunol. 2002;263:83–117. doi: 10.1007/978-3-642-56055-2_6. [DOI] [PubMed] [Google Scholar]
- 12.Suvas S, Azkur AK, Kim BS, Kumaraguru U, Rouse BT. CD4+CD25+ regulatory T cells control the severity of viral immunoinflammatory lesions. J Immunol. 2004;172:4123–4132. doi: 10.4049/jimmunol.172.7.4123. [DOI] [PubMed] [Google Scholar]
- 13.De Albuquerque N, Baig E, Ma X, Zhang J, He W, Rowe A, Habal M, Liu M, Shalev I, Downey GP, Gorczynski R, Butany J, Leibowitz J, Weiss SR, McGilvray ID, Phillips MJ, Fish EN, Levy GA. Murine hepatitis virus strain 1 produces a clinically relevant model of severe acute respiratory syndrome in A/J mice. J Virol. 2006;80:10382–10394. doi: 10.1128/JVI.00747-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bot A, Rodrigo E, Wolfe T, Bot S, Von Herrath MG. Infection-triggered regulatory mechanisms override the role of STAT 4 in control of the immune response to influenza virus antigens. J Virol. 2003;77:5794–5800. doi: 10.1128/JVI.77.10.5794-5800.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ejrnaes M, Filippi CM, Martinic MM, Ling EM, Togher LM, Crotty S, von Herrath MG. Resolution of a chronic viral infection after interleukin-10 receptor blockade. J Exp Med. 2006;203:2461–2472. doi: 10.1084/jem.20061462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ejrnaes M, von Herrath MG, Christen U. Cure of chronic viral infection and virus-induced type 1 diabetes by neutralizing antibodies. Clin Dev Immunol. 2006;13:337–347. doi: 10.1080/17402520600800721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Filippi CM, von Herrath MG. IL-10 and the resolution of infections. J Pathol. 2008;214:224–230. doi: 10.1002/path.2272. [DOI] [PubMed] [Google Scholar]
- 18.Homann D, Dummer W, Wolfe T, Rodrigo E, Theofilopoulos AN, Oldstone MB, von Herrath MG. Lack of intrinsic CTLA-4 expression has minimal effect on regulation of antiviral T-cell immunity. J Virol. 2006;80:270–280. doi: 10.1128/JVI.80.1.270-280.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.von Herrath MG, Berger DP, Homann D, Tishon T, Sette A, Oldstone MB. Vaccination to treat persistent viral infection. Virology. 2000;268:411–419. doi: 10.1006/viro.1999.0130. [DOI] [PubMed] [Google Scholar]
- 20.Wolfe T, Asseman C, Hughes A, Matsue H, Takashima A, von Herrath MG. Reduction of antiviral CD8 lymphocytes in vivo with dendritic cells expressing Fas ligand-increased survival of viral (lymphocytic choriomeningitis virus) central nervous system infection. J Immunol. 2002;169:4867–4872. doi: 10.4049/jimmunol.169.9.4867. [DOI] [PubMed] [Google Scholar]
- 21.Fuller MJ, Khanolkar A, Tebo AE, Zajac AJ. Maintenance, loss, and resurgence of T cell responses during acute, protracted, and chronic viral infections. J Immunol. 2004;172:4204–4214. doi: 10.4049/jimmunol.172.7.4204. [DOI] [PubMed] [Google Scholar]
- 22.Oldstone MB. A suspenseful game of “hide and seek” between virus and host. Nat Immunol. 2007;8:325–327. doi: 10.1038/ni0407-325. [DOI] [PubMed] [Google Scholar]
- 23.Brooks DG, Teyton L, Oldstone MB, McGavern DB. Intrinsic functional dysregulation of CD4 T cells occurs rapidly following persistent viral infection. J Virol. 2005;79:10514–10527. doi: 10.1128/JVI.79.16.10514-10527.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Moskophidis D, Lechner F, Pircher H, Zinkernagel RM. Virus persistence in acutely infected immunocompetent mice by exhaustion of antiviral cytotoxic effector T cells. Nature. 1993;362:758–761. doi: 10.1038/362758a0. [DOI] [PubMed] [Google Scholar]
- 25.Zajac AJ, Blattman JN, Murali-Krishna K, Sourdive DJ, Suresh M, Altman JD, Ahmed R. Viral immune evasion due to persistence of activated T cells without effector function. J Exp Med. 1998;188:2205–2213. doi: 10.1084/jem.188.12.2205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ou R, Zhou S, Huang L, Moskophidis D. Critical role for alpha/beta and gamma interferons in persistence of lymphocytic choriomeningitis virus by clonal exhaustion of cytotoxic T cells. J Virol. 2001;75:8407–8423. doi: 10.1128/JVI.75.18.8407-8423.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wherry EJ, Blattman JN, Murali-Krishna K, van der Most R, Ahmed R. Viral persistence alters CD8 T-cell immunodominance and tissue distribution and results in distinct stages of functional impairment. J Virol. 2003;77:4911–4927. doi: 10.1128/JVI.77.8.4911-4927.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kannanganat S, Kapogiannis BG, Ibegbu C, Chennareddi L, Goepfert P, Robinson HL, Lennox J, Amara RR. Human immunodeficiency virus type 1 controllers but not noncontrollers maintain CD4 T cells coexpressing three cytokines. J Virol. 2007;81:12071–12076. doi: 10.1128/JVI.01261-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Brooks DG, McGavern DB, Oldstone MB. Reprogramming of antiviral T cells prevents inactivation and restores T cell activity during persistent viral infection. J Clin Invest. 2006;116:1675–1685. doi: 10.1172/JCI26856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, Freeman GJ, Ahmed R. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature. 2006;439:682–687. doi: 10.1038/nature04444. [DOI] [PubMed] [Google Scholar]
- 31.Blackburn SD, Shin H, Haining WN, Zou T, Workman CJ, Polley A, Betts MR, Freeman GJ, Vignali DA, Wherry EJ. Coregulation of CD8+ T cell exhaustion by multiple inhibitory receptors during chronic viral infection. Nat Immunol. 2009;10:29–37. doi: 10.1038/ni.1679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Brooks DG, Trifilo MJ, Edelmann KH, Teyton L, McGavern DB, Oldstone MB. Interleukin-10 determines viral clearance or persistence in vivo. Nat Med. 2006;12:1301–1309. doi: 10.1038/nm1492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Brockman MA, Kwon DS, Tighe DP, Pavlik DF, Rosato PC, Sela J, Porichis F, Le Gall S, Waring MT, Moss K, Jessen H, Pereyra F, Kavanagh DG, Walker BD, Kaufmann DE. IL-10 is up-regulated in multiple cell types during viremic HIV infection and reversibly inhibits virus-specific T cells. Blood. 2009;114:346–356. doi: 10.1182/blood-2008-12-191296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Barrett L, Gallant M, Howley C, Bowmer MI, Hirsch G, Peltekian K, Grant M. Enhanced IL-10 production in response to hepatitis C virus proteins by peripheral blood mononuclear cells from human immunodeficiency virus-monoinfected individuals. BMC Immunol. 2008;9:28. doi: 10.1186/1471-2172-9-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Freeman GJ, Wherry EJ, Ahmed R, Sharpe AH. Reinvigorating exhausted HIV-specific T cells via PD-1-PD-1 ligand blockade. J Exp Med. 2006;203:2223–2227. doi: 10.1084/jem.20061800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Day CL, Kaufmann DE, Kiepiela P, Brown JA, Moodley ES, Reddy S, Mackey EW, Miller JD, Leslie AJ, DePierres C, Mncube Z, Duraiswamy J, Zhu B, Eichbaum Q, Altfeld M, Wherry EJ, Coovadia HM, Goulder PJ, Klenerman P, Ahmed R, Freeman GJ, Walker BD. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature. 2006;443:350–354. doi: 10.1038/nature05115. [DOI] [PubMed] [Google Scholar]
- 37.Landay AL, Clerici M, Hashemi F, Kessler H, Berzofsky JA, Shearer GM. In vitro restoration of T cell immune function in human immunodeficiency virus-positive persons: effects of interleukin (IL)-12 and anti-IL-10. J Infect Dis. 1996;173:1085–1091. doi: 10.1093/infdis/173.5.1085. [DOI] [PubMed] [Google Scholar]
- 38.Velu V, Titanji K, Zhu B, Husain S, Pladevega A, Lai L, Vanderford TH, Chennareddi L, Silvestri G, Freeman GJ, Ahmed R, Amara RR. Enhancing SIV-specific immunity in vivo by PD-1 blockade. Nature. 2009;458:206–210. doi: 10.1038/nature07662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Thepen T, Van Rooijen N, Kraal G. Alveolar macrophage elimination in vivo is associated with an increase in pulmonary immune response in mice. J Exp Med. 1989;170:499–509. doi: 10.1084/jem.170.2.499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Holt PG. Inhibitory activity of unstimulated alveolar macrophages on T-lymphocyte blastogenic response. Am Rev Respir Dis. 1978;118:791–793. doi: 10.1164/arrd.1978.118.4.791. [DOI] [PubMed] [Google Scholar]
- 41.Dillon S, Agrawal S, Banerjee K, Letterio J, Denning TL, Oswald-Richter K, Kasprowicz DJ, Kellar K, Pare J, van Dyke T, Ziegler S, Unutmaz D, Pulendran B. Yeast zymosan, a stimulus for TLR2 and dectin-1, induces regulatory antigen-presenting cells and immunological tolerance. J Clin Invest. 2006;116:916–928. doi: 10.1172/JCI27203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Holt PG, Oliver J, Bilyk N, McMenamin C, McMenamin PG, Kraal G, Thepen T. Downregulation of the antigen presenting cell function(s) of pulmonary dendritic cells in vivo by resident alveolar macrophages. J Exp Med. 1993;177:397–407. doi: 10.1084/jem.177.2.397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Bilyk N, Holt PG. Cytokine modulation of the immunosuppressive phenotype of pulmonary alveolar macrophage populations. Immunology. 1995;86:231–237. [PMC free article] [PubMed] [Google Scholar]
- 44.Strickland DH, Thepen T, Kees UR, Kraal G, Holt PG. Regulation of T-cell function in lung tissue by pulmonary alveolar macrophages. Immunology. 1993;80:266–272. [PMC free article] [PubMed] [Google Scholar]
- 45.Steinman RM, Hawiger D, Nussenzweig MC. Tolerogenic dendritic cells. Annu Rev Immunol. 2003;21:685–711. doi: 10.1146/annurev.immunol.21.120601.141040. [DOI] [PubMed] [Google Scholar]
- 46.Steinbrink K, Wolfl M, Jonuleit H, Knop J, Enk AH. Induction of tolerance by IL-10-treated dendritic cells. J Immunol. 1997;159:4772–4780. [PubMed] [Google Scholar]
- 47.Steinbrink K, Graulich E, Kubsch S, Knop J, Enk AH. CD4(+) and CD8(+) anergic T cells induced by interleukin-10-treated human dendritic cells display antigen-specific suppressor activity. Blood. 2002;99:2468–2476. doi: 10.1182/blood.V99.7.2468. [DOI] [PubMed] [Google Scholar]
- 48.Kubsch S, Graulich E, Knop J, Steinbrink K. Suppressor activity of anergic T cells induced by IL-10-treated human dendritic cells: association with IL-2- and CTLA-4-dependent G1 arrest of the cell cycle regulated by p27Kip1. Eur J Immunol. 2003;33:1988–1997. doi: 10.1002/eji.200323600. [DOI] [PubMed] [Google Scholar]
- 49.Penna G, Roncari A, Amuchastegui S, Daniel KC, Berti E, Colonna M, Adorini L. Expression of the inhibitory receptor ILT3 on dendritic cells is dispensable for induction of CD4+Foxp3+ regulatory T cells by 1, 25-dihydroxyvitamin D3. Blood. 2005;106:3490–3497. doi: 10.1182/blood-2005-05-2044. [DOI] [PubMed] [Google Scholar]
- 50.Netea MG, Sutmuller R, Hermann C, Van der Graaf CA, Van der Meer JW, van Krieken JH, Hartung T, Adema G, Kullberg BJ. Toll-like receptor 2 suppresses immunity against Candida albicans through induction of IL-10 and regulatory T cells. J Immunol. 2004;172:3712–3718. doi: 10.4049/jimmunol.172.6.3712. [DOI] [PubMed] [Google Scholar]
- 51.Sutmuller RP, den Brok MH, Kramer M, Bennink EJ, Toonen LW, Kullberg BJ, Joosten LA, Akira S, Netea MG, Adema GJ. Toll-like receptor 2 controls expansion and function of regulatory T cells. J Clin Invest. 2006;116:485–494. doi: 10.1172/JCI25439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Penna G, Adorini L. 1 Alpha, 25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J Immunol. 2000;164:2405–2411. doi: 10.4049/jimmunol.164.5.2405. [DOI] [PubMed] [Google Scholar]
- 53.Oh S, Eichelberger MC. Influenza virus neuraminidase alters allogeneic T cell proliferation. Virology. 1999;264:427–435. doi: 10.1006/viro.1999.0019. [DOI] [PubMed] [Google Scholar]
- 54.Oh S, McCaffery JM, Eichelberger MC. Dose-dependent changes in influenza virus-infected dendritic cells result in increased allogeneic T-cell proliferation at low, but not high, doses of virus. J Virol. 2000;74:5460–5469. doi: 10.1128/JVI.74.12.5460-5469.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Dhodapkar MV, Steinman RM. Antigen-bearing immature dendritic cells induce peptide-specific CD8(+) regulatory T cells in vivo in humans. Blood. 2002;100:174–177. doi: 10.1182/blood.V100.1.174. [DOI] [PubMed] [Google Scholar]
- 56.Dhodapkar MV, Steinman RM, Krasovsky J, Munz C, Bhardwaj N. Antigen-specific inhibition of effector T cell function in humans after injection of immature dendritic cells. J Exp Med. 2001;193:233–238. doi: 10.1084/jem.193.2.233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sakaguchi S, Wing K, Yamaguchi T. Dynamics of peripheral tolerance and immune regulation mediated by Treg. Eur J Immunol. 2009;39:2331–2336. doi: 10.1002/eji.200939688. [DOI] [PubMed] [Google Scholar]
- 58.Hori S, Nomura T, Sakaguchi S. Control of regulatory T cell development by the transcription factor Foxp3. Science. 2003;299:1057–1061. doi: 10.1126/science.1079490. [DOI] [PubMed] [Google Scholar]
- 59.Zhou X, Bailey-Bucktrout SL, Jeker LT, Penaranda C, Martinez-Llordella M, Ashby M, Nakayama M, Rosenthal W, Bluestone JA. Instability of the transcription factor Foxp3 leads to the generation of pathogenic memory T cells in vivo. Nat Immunol. 2009;10:1000–1007. doi: 10.1038/ni.1774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Toda A, Piccirillo CA. Development and function of naturally occurring CD4+CD25+ regulatory T cells. J Leukoc Biol. 2006;80:458–470. doi: 10.1189/jlb.0206095. [DOI] [PubMed] [Google Scholar]
- 61.Ozdemir C, Akdis M, Akdis CA. T regulatory cells and their counterparts: masters of immune regulation. Clin Exp Allergy. 2009;39:626–639. doi: 10.1111/j.1365-2222.2009.03242.x. [DOI] [PubMed] [Google Scholar]
- 62.Horai R, Asano M, Sudo K, Kanuka H, Suzuki M, Nishihara M, Takahashi M, Iwakura Y. Production of mice deficient in genes for interleukin (IL)-1alpha, IL-1beta, IL-1alpha/beta, and IL-1 receptor antagonist shows that IL-1beta is crucial in turpentine-induced fever development and glucocorticoid secretion. J Exp Med. 1998;187:1463–1475. doi: 10.1084/jem.187.9.1463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Aandahl EM, Michaelsson J, Moretto WJ, Hecht FM, Nixon DF. Human CD4+ CD25+ regulatory T cells control T-cell responses to human immunodeficiency virus and cytomegalovirus antigens. J Virol. 2004;78:2454–2459. doi: 10.1128/JVI.78.5.2454-2459.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Boettler T, Spangenberg HC, Neumann-Haefelin C, Panther E, Urbani S, Ferrari C, Blum HE, von Weizsacker F, Thimme R. T cells with a CD4+CD25+ regulatory phenotype suppress in vitro proliferation of virus-specific CD8+ T cells during chronic hepatitis C virus infection. J Virol. 2005;79:7860–7867. doi: 10.1128/JVI.79.12.7860-7867.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Robertson SJ, Messer RJ, Carmody AB, Hasenkrug KJ. In vitro suppression of CD8+ T cell function by Friend virus-induced regulatory T cells. J Immunol. 2006;176:3342–3349. doi: 10.4049/jimmunol.176.6.3342. [DOI] [PubMed] [Google Scholar]
- 66.Suvas S, Kumaraguru U, Pack CD, Lee S, Rouse BT. CD4+CD25+ T cells regulate virus-specific primary and memory CD8+ T cell responses. J Exp Med. 2003;198:889–901. doi: 10.1084/jem.20030171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Truong P, McGavern DB. A novel virus carrier state to evaluate immunotherapeutic regimens: regulatory T cells modulate the pathogenicity of antiviral memory cells. J Immunol. 2008;181:1161–1169. doi: 10.4049/jimmunol.181.2.1161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Belkaid Y. Regulatory T cells and infection: a dangerous necessity. Nat Rev Immunol. 2007;7:875–888. doi: 10.1038/nri2189. [DOI] [PubMed] [Google Scholar]
- 69.Belkaid Y, Rouse BT. Natural regulatory T cells in infectious disease. Nat Immunol. 2005;6:353–360. doi: 10.1038/ni1181. [DOI] [PubMed] [Google Scholar]
- 70.Robertson SJ, Hasenkrug KJ. The role of virus-induced regulatory T cells in immunopathology. Springer Semin Immunopathol. 2006;28:51–62. doi: 10.1007/s00281-006-0019-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Iwashiro M, Messer RJ, Peterson KE, Stromnes IM, Sugie T, Hasenkrug KJ. Immunosuppression by CD4+ regulatory T cells induced by chronic retroviral infection. Proc Natl Acad Sci U S A. 2001;98:9226–9230. doi: 10.1073/pnas.151174198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.MacDonald AJ, Duffy M, Brady MT, McKiernan S, Hall W, Hegarty J, Curry M, Mills KH. CD4 T helper type 1 and regulatory T cells induced against the same epitopes on the core protein in hepatitis C virus-infected persons. J Infect Dis. 2002;185:720–727. doi: 10.1086/339340. [DOI] [PubMed] [Google Scholar]
- 73.Luhn K, Simmons CP, Moran E, Dung NT, Chau TN, Quyen NT, Thao le TT, Van Ngoc T, Dung NM, Wills B, Farrar J, McMichael AJ, Dong T, Rowland-Jones S. Increased frequencies of CD4+ CD25(high) regulatory T cells in acute dengue infection. J Exp Med. 2007;204:979–985. doi: 10.1084/jem.20061381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Shevach EM. Mechanisms of foxp3+ T regulatory cell-mediated suppression. Immunity. 2009;30:636–645. doi: 10.1016/j.immuni.2009.04.010. [DOI] [PubMed] [Google Scholar]
- 75.Tang Q, Bluestone JA. The Foxp3+ regulatory T cell: a jack of all trades, master of regulation. Nat Immunol. 2008;9:239–244. doi: 10.1038/ni1572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Vignali DA, Collison LW, Workman CJ. How regulatory T cells work. Nat Rev Immunol. 2008;8:523–532. doi: 10.1038/nri2343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Miyara M, Sakaguchi S. Natural regulatory T cells: mechanisms of suppression. Trends Mol Med. 2007;13:108–116. doi: 10.1016/j.molmed.2007.01.003. [DOI] [PubMed] [Google Scholar]
- 78.Thornton AM, Shevach EM. CD4+CD25+ immunoregulatory T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. J Exp Med. 1998;188:287–296. doi: 10.1084/jem.188.2.287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Wing K, Onishi Y, Prieto-Martin P, Yamaguchi T, Miyara M, Fehervari Z, Nomura T, Sakaguchi S. CTLA-4 control over Foxp3+ regulatory T cell function. Science. 2008;322:271–275. doi: 10.1126/science.1160062. [DOI] [PubMed] [Google Scholar]
- 80.Onishi Y, Fehervari Z, Yamaguchi T, Sakaguchi S. Foxp3+ natural regulatory T cells preferentially form aggregates on dendritic cells in vitro and actively inhibit their maturation. Proc Natl Acad Sci U S A. 2008;105:10113–10118. doi: 10.1073/pnas.0711106105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.de la Rosa M, Rutz S, Dorninger H, Scheffold A. Interleukin-2 is essential for CD4+CD25+ regulatory T cell function. Eur J Immunol. 2004;34:2480–2488. doi: 10.1002/eji.200425274. [DOI] [PubMed] [Google Scholar]
- 82.Pandiyan P, Zheng L, Ishihara S, Reed J, Lenardo MJ. CD4+CD25+Foxp3+ regulatory T cells induce cytokine deprivation-mediated apoptosis of effector CD4+ T cells. Nat Immunol. 2007;8:1353–1362. doi: 10.1038/ni1536. [DOI] [PubMed] [Google Scholar]
- 83.Filippi CM, Estes EA, Oldham JE, von Herrath MG. Immunoregulatory mechanisms triggered by viral infections protect from type 1 diabetes in mice. J Clin Invest. 2009;119:1515–1523. doi: 10.1172/JCI38503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Ha SJ, Mueller SN, Wherry EJ, Barber DL, Aubert RD, Sharpe AH, Freeman GJ, Ahmed R. Enhancing therapeutic vaccination by blocking PD-1-mediated inhibitory signals during chronic infection. J Exp Med. 2008;205:543–555. doi: 10.1084/jem.20071949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Petrovas C, Casazza JP, Brenchley JM, Price DA, Gostick E, Adams WC, Precopio ML, Schacker T, Roederer M, Douek DC, Koup RA. PD-1 is a regulator of virus-specific CD8+ T cell survival in HIV infection. J Exp Med. 2006;203:2281–2292. doi: 10.1084/jem.20061496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Trautmann L, Janbazian L, Chomont N, Said EA, Gimmig S, Bessette B, Boulassel MR, Delwart E, Sepulveda H, Balderas RS, Routy JP, Haddad EK, Sekaly RP. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nat Med. 2006;12:1198–1202. doi: 10.1038/nm1482. [DOI] [PubMed] [Google Scholar]
- 87.Fife BT, Bluestone JA. Control of peripheral T-cell tolerance and autoimmunity via the CTLA-4 and PD-1 pathways. Immunol Rev. 2008;224:166–182. doi: 10.1111/j.1600-065X.2008.00662.x. [DOI] [PubMed] [Google Scholar]
- 88.Williams MA, Onami TM, Adams AB, Durham MM, Pearson TC, Ahmed R, Larsen CP. Cutting edge: persistent viral infection prevents tolerance induction and escapes immune control following CD28/CD40 blockade-based regimen. J Immunol. 2002;169:5387–5391. doi: 10.4049/jimmunol.169.10.5387. [DOI] [PubMed] [Google Scholar]
- 89.Rudd CE, Taylor A, Schneider H. CD28 and CTLA-4 coreceptor expression and signal transduction. Immunol Rev. 2009;229:12–26. doi: 10.1111/j.1600-065X.2009.00770.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Read S, Malmstrom V, Powrie F. Cytotoxic T lymphocyte-associated antigen 4 plays an essential role in the function of CD25(+)CD4(+) regulatory cells that control intestinal inflammation. J Exp Med. 2000;192:295–302. doi: 10.1084/jem.192.2.295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Takahashi T, Tagami T, Yamazaki S, Uede T, Shimizu J, Sakaguchi N, Mak TW, Sakaguchi S. Immunologic self-tolerance maintained by CD25(+)CD4(+) regulatory T cells constitutively expressing cytotoxic T lymphocyte-associated antigen 4. J Exp Med. 2000;192:303–310. doi: 10.1084/jem.192.2.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Salomon B, Lenschow DJ, Rhee L, Ashourian N, Singh B, Sharpe A, Bluestone JA. B7/CD28 costimulation is essential for the homeostasis of the CD4+CD25+ immunoregulatory T cells that control autoimmune diabetes. Immunity. 2000;12:431–440. doi: 10.1016/S1074-7613(00)80195-8. [DOI] [PubMed] [Google Scholar]
- 93.Simone R, Piatti G, Saverino D. The inhibitory co-receptors: a way to save from anergy the HIV-specific T cells. Curr HIV Res. 2009;7:266–272. doi: 10.2174/157016209788347949. [DOI] [PubMed] [Google Scholar]
- 94.Hryniewicz A, Boasso A, Edghill-Smith Y, Vaccari M, Fuchs D, Venzon D, Nacsa J, Betts MR, Tsai WP, Heraud JM, Beer B, Blanset D, Chougnet C, Lowy I, Shearer GM, Franchini G. CTLA-4 blockade decreases TGF-beta, IDO, and viral RNA expression in tissues of SIVmac251-infected macaques. Blood. 2006;108:3834–3842. doi: 10.1182/blood-2006-04-010637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Kaufmann DE, Kavanagh DG, Pereyra F, Zaunders JJ, Mackey EW, Miura T, Palmer S, Brockman M, Rathod A, Piechocka-Trocha A, Baker B, Zhu B, Le Gall S, Waring MT, Ahern R, Moss K, Kelleher AD, Coffin JM, Freeman GJ, Rosenberg ES, Walker BD. Upregulation of CTLA-4 by HIV-specific CD4+ T cells correlates with disease progression and defines a reversible immune dysfunction. Nat Immunol. 2007;8:1246–1254. doi: 10.1038/ni1515. [DOI] [PubMed] [Google Scholar]
- 96.Krummel MF, Allison JP. CTLA-4 engagement inhibits IL-2 accumulation and cell cycle progression upon activation of resting T cells. J Exp Med. 1996;183:2533–2540. doi: 10.1084/jem.183.6.2533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Li MO, Wan YY, Sanjabi S, Robertson AK, Flavell RA. Transforming growth factor-beta regulation of immune responses. Annu Rev Immunol. 2006;24:99–146. doi: 10.1146/annurev.immunol.24.021605.090737. [DOI] [PubMed] [Google Scholar]
- 98.Gorelik L, Flavell RA. Transforming growth factor-beta in T-cell biology. Nat Rev Immunol. 2002;2:46–53. doi: 10.1038/nri704. [DOI] [PubMed] [Google Scholar]
- 99.Zhang X, Giangreco L, Broome HE, Dargan CM, Swain SL. Control of CD4 effector fate: transforming growth factor beta 1 and interleukin 2 synergize to prevent apoptosis and promote effector expansion. J Exp Med. 1995;182:699–709. doi: 10.1084/jem.182.3.699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Genestier L, Kasibhatla S, Brunner T, Green DR. Transforming growth factor beta1 inhibits Fas ligand expression and subsequent activation-induced cell death in T cells via downregulation of c-Myc. J Exp Med. 1999;189:231–239. doi: 10.1084/jem.189.2.231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Cerwenka A, Kovar H, Majdic O, Holter W. Fas- and activation-induced apoptosis are reduced in human T cells preactivated in the presence of TGF-beta 1. J Immunol. 1996;156:459–464. [PubMed] [Google Scholar]
- 102.Chen W, Jin W, Hardegen N, Lei KJ, Li L, Marinos N, McGrady G, Wahl SM. Conversion of peripheral CD4+CD25-naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J Exp Med. 2003;198:1875–1886. doi: 10.1084/jem.20030152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Filippi C, Bresson D, von Herrath M. Antigen-specific induction of regulatory T cells for type 1 diabetes therapy. Int Rev Immunol. 2005;24:341–360. doi: 10.1080/08830180500371116. [DOI] [PubMed] [Google Scholar]
- 104.Filippi CM, Juedes AE, Oldham JE, Ling E, Togher L, Peng Y, Flavell RA, von Herrath MG. TGF-{beta} suppresses the activation of CD8+ T cells when naive but promotes their survival and function once antigen-experienced: a two-faced impact on autoimmunity. Diabetes. 2008;57:2684–2692. doi: 10.2337/db08-0609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Piccirillo CA, Chang Y, Prud’homme GJ. TGF-beta1 somatic gene therapy prevents autoimmune disease in nonobese diabetic mice. J Immunol. 1998;161:3950–3956. [PubMed] [Google Scholar]
- 106.Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, Weiner HL, Kuchroo VK. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441:235–238. doi: 10.1038/nature04753. [DOI] [PubMed] [Google Scholar]
- 107.Stockinger B, Veldhoen M. Differentiation and function of Th17 T cells. Curr Opin Immunol. 2007;19:281–286. doi: 10.1016/j.coi.2007.04.005. [DOI] [PubMed] [Google Scholar]
- 108.Laurence A, Tato CM, Davidson TS, Kanno Y, Chen Z, Yao Z, Blank RB, Meylan F, Siegel R, Hennighausen L, Shevach EM, O’Shea JJ. Interleukin-2 signaling via STAT5 constrains T helper 17 cell generation. Immunity. 2007;26:371–381. doi: 10.1016/j.immuni.2007.02.009. [DOI] [PubMed] [Google Scholar]
- 109.de Waal Malefyt R, Abrams J, Bennett B, Figdor CG, de Vries JE. Interleukin 10(IL-10) inhibits cytokine synthesis by human monocytes: an autoregulatory role of IL-10 produced by monocytes. J Exp Med. 1991;174:1209–1220. doi: 10.1084/jem.174.5.1209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Taylor A, Akdis M, Joss A, Akkoc T, Wenig R, Colonna M, Daigle I, Flory E, Blaser K, Akdis CA. IL-10 inhibits CD28 and ICOS costimulations of T cells via src homology 2 domain-containing protein tyrosine phosphatase 1. J Allergy Clin Immunol. 2007;120:76–83. doi: 10.1016/j.jaci.2007.04.004. [DOI] [PubMed] [Google Scholar]
- 111.Brooks DG, Lee AM, Elsaesser H, McGavern DB, Oldstone MB. IL-10 blockade facilitates DNA vaccine-induced T cell responses and enhances clearance of persistent virus infection. J Exp Med. 2008;205:533–541. doi: 10.1084/jem.20071948. [DOI] [PMC free article] [PubMed] [Google Scholar]