

Abstract
The lung, like many other organs, is innervated by a variety of sensory nerves and by nerves of the parasympathetic and sympathetic nervous systems that regulate the function of cells within the respiratory tract. Activation of sensory nerves by both mechanical and chemical stimuli elicits a number of defensive reflexes, including cough, altered breathing pattern, and altered autonomic drive, which are important for normal lung homeostasis. However, diseases that afflict the lung are associated with altered reflexes, resulting in a variety of symptoms, including increased cough, dyspnea, airways obstruction, and bronchial hyperresponsiveness. This review summarizes the current knowledge concerning the physiological role of different sensory nerve subtypes that innervate the lung, the factors which lead to their activation, and pharmacological approaches that have been used to interrogate the function of these nerves. This information may potentially facilitate the identification of novel drug targets for the treatment of respiratory disorders such as cough, asthma, and chronic obstructive pulmonary disease.
Keywords: Rapidly adapting receptors, C-fibers, Cough receptor, Cough, Parasympathetic nervous system, Sympathetic nervous system, Cough, Bronchoconstriction, Mucus secretion, Bronchial hyperresponsiveness
Introduction
The primary function of the lung is gas exchange. The airways serve as a conduit for moving inspired air to the gas-exchanging regions of the lungs, and for expiration of CO2. Airway and lung reflexes optimize lung capacity for gas exchange in response to a continually changing demand. Airway reflexes also serve to preserve airway patency. These reflexes can become aberrant, however, and may worsen the symptoms of diseases such as asthma and chronic obstructive pulmonary disease (COPD). Multiple afferent nerve subtypes regulate these homeostatic and defensive reflexes, each subtype with unique physiological, anatomical, and pharmacological attributes. The properties of airway afferent nerve subtypes will be reviewed, as will their role in regulating bronchopulmonary reflexes and bronchial responsiveness.
Airway and Lung Afferent Nerve Subtypes
Airway afferent nerve subtypes have been defined by their chemical and physical sensitivity, adaptation to mechanical stimulation, origin, myelination, conduction velocity, neurochemistry, basal activity, reflexes associated with their activation, and sites of termination in the airways, lungs, and brain stem. These various approaches to characterizing airway afferent nerves are hampered by their lack of specificity. But when used in combination, patterns of physiological and pharmacological attributes have emerged to help define at least four distinct subtypes of airway afferent nerves (Canning et al. 2006b).
Slowly Adapting Receptors
Slowly adapting receptors (SARs) are the prototypical airway mechanoreceptors. The mechanical forces produced during breathing are the primary stimulus for SAR activation, with SAR activity increasing during inspiration and peaking prior to expiration (Miserocchi and Sant'Ambrogio 1974; Ho et al. 2001; Schelegle and Green 2001). SARs regulate the Hering-Breuer reflex, which terminates inspiration and initiates expiration when the lungs are adequately inflated (Schelegle and Green 2001). SARs thus play a primary role in regulating respiratory rate.
SARs can be differentiated from rapidly adapting receptors (RARs) in some species on the basis of action potential conduction velocity, and in most species by their modest adaptation to sustained lung inflation (Fig. 1). SARs may be differentially distributed in the airways of commonly studied mammalian species (Schelegle and Green 2001). In cats, guinea pigs, and rats, few SARs but many RAR-like receptors and C-fibers can be found in the extrapulmonary airways. In dogs, SARs may also be localized to the extrapulmonary airways (Miserocchi and Sant'Ambrogio 1974; Sant'Ambrogio et al. 1988). SARs also differ from RARs with respect to the reflexes they precipitate (see later). Subtypes of SARs have been described (Miserocchi and Sant'Ambrogio 1974; Schelegle and Green 2001).
Fig. 1.

The characteristic features of airway and lung vagal afferent nerve subtypes are shown in these single-fiber recordings in the rat. C-fibers are generally unresponsive to mechanical stimulation, including the mechanical consequences of lung inflation and deflation, but are vigorously activated by capsaicin. The rapidly adapting stretch receptors (RARs) and the slowly adapting stretch receptors (SARs) are largely insensitive to capsaicin. Both lung stretch receptor subtypes are responsive to lung inflation, with RAR activity more prominent in species with higher respiratory rates. RARs and SARs are differentiated in part by their responses to sustained lung inflation. These vagal afferent nerve subtypes differentially regulate airway autonomic outflow, respiratory pattern, respiratory sensations, and cough. Subtypes of each afferent class have been described and are found in all species thus far studied. (Reproduced with permission from Ho et al. 2001)
SARs are generally unresponsive to chemical stimuli. With the exception of the small population of SARs terminating in airway smooth muscle, this also includes an insensitivity to stimuli that initiate bronchospasm, pulmonary edema, pulmonary vascular congestion, or any stimulus that decreases lung compliance. The ion channels regulating the mechanical sensitivity of SARs are also poorly defined. Gadolinium (20 mM applied repeatedly for 30 min to SAR receptive fields) slightly (10–40%) reduced rabbit bronchial SAR discharge to different levels of inflation pressures (Ma et al. 2004). Other drugs reported to modify SAR discharge include the voltage-sensitive K+-channel blocker 4-aminopyridine, the voltage-sensitive Na+-channel opener veratradine, the Na+–K+–2Cl− transporter inhibitor furosemide, sulfur dioxide, and the Na+–K+–ATPase inhibitor ouabain (Davies et al. 1978; Matsumoto et al. 1998, 1999, 2000, 2005, 2006; Sudo et al. 2000; Guardiola et al. 2007).
An additional and unique physiological and pharmacological property of SARs is their sensitivity to alveolar CO2 concentrations (Coleridge et al. 1978; Fisher and Sant'Ambrogio 1982; Green et al. 1986). As alveolar CO2 increases, SAR activity decreases. This contrasts sharply with bronchopulmonary C-fibers, which may be activated or at least sensitized by elevated alveolar CO2 and/or decreases in extracellular pH (Delpierre et al. 1981; Lin et al. 2005). The inhibitory effect of CO2 on SARs contributes in part to the hyperpnea associated with hypercapnea. The actions of CO2 on SAR excitability may occur secondary to effects on nerve terminal pH as evidenced by the preventive effect of the carbonic anhydrase inhibitor acetazolamide during CO2 challenges (Ravi 1985; Matsumoto 1996; Hempleman et al. 2000). The expression of a carbonic anhydrase isozyme in SARs has not been systematically evaluated.
Rapidly Adapting Receptors
The term “rapidly adapting receptor” (RAR) describes a subtype of airway and lung stretch receptor that are activated during the dynamic phase of lung inflation, but unlike SARs become quiescent during static lung inflation (Knowlton and Larrabee 1946; Widdicombe 1954a). Often inappropriately, airway afferents that rapidly adapt to any stimulus are grouped into a broad and heterogeneous class, all of which are called “RARs.” This has proven misleading (Canning and Chou 2009). In this review, the term “RAR” refers to those intrapulmonary stretch receptors that rapidly adapt to sustained lung inflation (Fig. 1).
RARs are considerably less active than SARs during eupnea but more active than C-fibers. RARs are also activated (either directly or indirectly) by a variety of mechanical stimuli in the lung, including airway smooth muscle contraction, pulmonary edema, decreased lung compliance, lung collapse, and negative airway luminal pressures (Mills et al. 1970; Armstrong and Luck 1974; Bergren 1997; Ho et al. 2001; Canning et al. 2004; Canning and Chou 2009).
RARs are generally more responsive to chemical stimuli than SARs, prompting use of the term “irritant receptor” to describe this airway afferent nerve subtype. What has been less clear, however, is whether the ability of these chemical stimuli to activate RARs is through a direct action or secondary to a mechanical effect evoked in the lung. As mentioned above, RARs are exquisitely sensitive to decreases in lung compliance (Jonzon et al. 1986; Yu et al. 1987, 1989). Accordingly, bronchoconstrictors such as serotonin, methacholine, histamine, and substance P and many other stimuli that initiate bronchospasm likely activate RARs at least partly through their effects on lung mechanics (Mills et al. 1970; Mohammed et al. 1993; Bergren 1997; Canning et al. 2004; Chou et al. 2008). Similar mechanisms may underlie the ability of pulmonary embolism and pulmonary vascular congestion to activate RARs (Mills et al. 1969; Sellick and Widdicombe 1969; Ravi and Kappagoda 1992; Bonham et al. 1996) (Fig. 2). But there are several reports suggesting a direct effect of autacoids such as ATP, histamine, and serotonin on RARs (Vidruk et al. 1977; Dixon et al. 1979; Matsumoto and Shimizu 1989; Canning et al. 2004). Unlike SARs and C-fibers, however, RARs are largely unaffected by changes in alveolar CO2 concentrations (Sampson and Vidruk 1975; Ravi 1985).
Fig. 2.
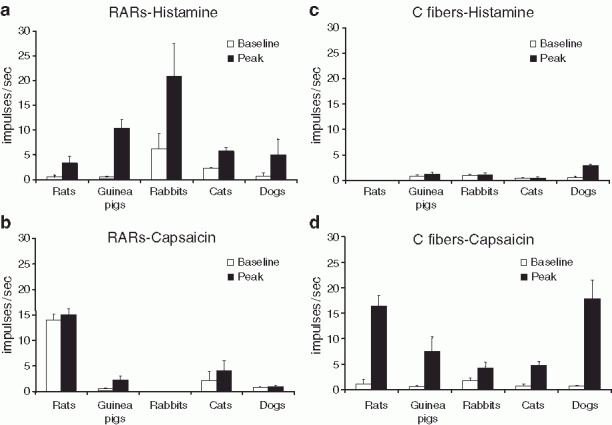
Responsiveness to histamine and capsaicin differentiates RARs from C-fibers. (Reproduced from Canning and Chou 2009 and summarizes results published elsewhere)
As with SARs, the ion channels regulating RAR activation secondary to mechanical stimulation are poorly defined. Gadolinium (20 mM applied repeatedly for 30 min to RAR receptive fields) is reported to have a profound effect on RAR excitation by lung inflation and pulmonary vascular congestion (Ma et al. 2004). Given the distinctive accommodative responses of RARs to sustained lung inflation, it seems possible that they express a unique set of ion channels that may be amenable to selective pharmacological modulation. This may prove useful in developing more selective drugs for treating respiratory disorders.
C-Fibers
With their defining physiological attribute of an axonal conduction velocity of 1 ms−1 or less, bronchopulmonary C-fibers are the most readily identifiable vagal afferent nerve subtype innervating the airways. C-fibers can be activated by several chemical and mechanical stimuli, with responses depending upon the stimulus and the C-fiber subtype studied (Coleridge and Coleridge 1984; Ricco et al. 1996; Lee and Pisarri 2001; Undem et al. 2004). The majority of C-fibers innervating the airways and lungs of all species are activated by the TRPV1 receptor agonist capsaicin (Figs. 1, 2), a predictable observation, given the known expression patterns of TRPV1 in afferent C-fibers throughout the body of most species (Caterina et al. 1997). But it is inappropriate to conclude from these data that responsiveness to capsaicin is the defining characteristic of airway C-fibers. C-fibers in dogs, rats, and mice that are not activated by lung capsaicin challenge have been described (Coleridge and Coleridge 1984; Ho et al. 2001; Kollarik et al. 2003). Moreover, perhaps secondary to the end organ effects associated with C-fiber activation (mucus secretion, vascular engorgement, airway smooth muscle contraction, altered respiratory pattern, and cough), other afferent nerve subtypes, especially intrapulmonary RARs, can be activated by capsaicin challenge (Mohammed et al. 1993; Bergren 1997; Morikawa et al. 1997). A lack of responsiveness to mechanical stimulation and basal activity may also fail to differentiate C-fibers from other subtypes of bronchopulmonary afferent nerves. While C-fibers are generally less responsive to mechanical stimulation, they can be activated by punctate mechanical stimulation or lung inflation, and can have basal activity comparable to that of some RARs (Coleridge and Coleridge 1984; Fox et al. 1993; Ricco et al. 1996; Lee and Pisarri 2001).
Subtypes of bronchopulmonary C-fibers have been described. The Coleridges defined C-fiber subtypes in dogs by their responsiveness to stimulants administered via the pulmonary or bronchial circulation (Coleridge and Coleridge 1984). More recently, Undem (Kollarik et al. 2003; Undem et al. 2004) described bronchopulmonary C-fiber subtypes in both guinea pigs and mice. In guinea pigs, C-fiber subtypes are differentiated on the basis of their ganglionic origin (nodose vs. jugular ganglia) and, by extension, their embryological origin, their sites of peripheral termination (extrapulmonary and intrapulmonary vs. exclusively intrapulmonary), expression of neurokinins, and responsiveness to adenosine, serotonin 5-HT3, and ATP receptor agonists (Undem et al. 2004; Chuaychoo et al. 2005, 2006). These subtypes are known to have opposing effects on both cough and respiration, but both subtypes may initiate reflex bronchospasm upon activation (Canning et al. 2006a, b; Chou et al. 2008; Reynolds et al. 2008) (Fig. 3). C-fibers arising from dorsal root ganglia also innervate the airways (Martling 1987; Kummer et al. 1992; Dinh et al. 2004; Oh et al. 2006; Kwong et al. 2008b). Their physiological properties and reflex actions have been only partially described.
Fig. 3.

Respiratory reflex effects evoked by histamine, adenosine, and capsaicin reveal the differential distribution of airway vagal afferent nerve subtypes and their distinct effects on respiratory pattern. Histamine selectively activates intrapulmonary RARs and initiates tachypnea. Adenosine selectively activates pulmonary C-fibers and also initiates tachypnea. Capsaicin activates both bronchial and pulmonary type C-fibers, initiating a profound slowing of respiration upon laryngeal challenge, tachypnea when capsaicin is inhaled (not shown), and both tachypnea and respiratory slowing following intravenous administration. (Data adapted from Chou et al. 2008)
C-fibers are found throughout the airways and lungs of all species. In guinea pigs, the jugular-type C-fibers have been localized to both intrapulmonary and extrapulmonary airways, while the nodose-type C-fibers are found predominantly in the peripheral airways and lungs (Undem et al. 2004). The extensively branched terminals of C-fibers in guinea pig and rat tracheae can be immunohistochemically labeled for the neuropeptides calcitonin gene-related peptide (CGRP), substance P, and neurokinin A (McDonald et al. 1988; Baluk et al. 1992; Kummer et al. 1992; Hunter and Undem 1999; Yamamoto et al. 2007). Comparable structures can be found in the airways of other species and in the peripheral airways of guinea pigs (Dey et al. 1990; Yamamoto et al. 1998; Lamb and Sparrow 2002; Watanabe et al. 2006). C-fiber terminals can also be found in the airway microvasculature and airway smooth muscle layer, and comprise at least a portion of Paintal's J-receptors, suggesting peripheral/interstitial lung terminations (Paintal 1973; McDonald et al. 1988; Baluk et al. 1992).
A myelinated (based on an axonal conduction velocity of 5 ms−1) afferent nerve subtype with many shared physiological and pharmacological attributes of jugular C-fibers has also been described in guinea pigs (Ricco et al. 1996). These afferents have their cell bodies in the jugular ganglia and are activated by acid, hypertonic saline, bradykinin, and capsaicin. Unlike jugular C-fibers, these capsaicin-sensitive Aδ-fibers terminate exclusively in the large airways (larynx, trachea, mainstem bronchi) and do not normally express the neuropeptide substance P (but can be labeled immunohistochemically for the structural protein neurofilament). TRPV1-positive, substance P-negative nerve terminals have been described in the airway epithelium of guinea pigs and may correspond to this afferent subtype (Watanabe et al. 2005, 2006). The existence of an Aδ afferent subpopulation expressing TRPV1 in other species and their reflex effects in any species upon activation are unknown.
In contrast to the indirect effects of autacoids and irritants thought to account for their activation of RARs, there is molecular, immunohistochemical, and electrophysiological evidence to suggest that many mediators associated with airway inflammation act directly on bronchopulmonary C-fibers. Stimuli known to activate airway and lung C-fibers include capsaicin and other TRPV1 receptor ligands, acid, cationic proteins, bradykinin, thrombin, and other protease-activated receptor 1 (PAR1) agonists, adenosine, 5-HT3 receptor agonists, nicotine, ATP, prostanoids, and isoprostanes, and a variety of environmental irritants including acrolein, toluene diisocyanate, and ozone (Coleridge and Coleridge 1984; Lee and Pisarri 2001; Undem et al. 2004; Chuaychoo et al. 2005, 2006; Nassenstein et al. 2008; Taylor-Clark et al. 2008). Many of these stimuli work partly or entirely through gating of the ion channels TRPV1 and TRPA1. PCR analyses confirm the expression of TRPV1 and TRPA1, but also adenosine A1, adenosine A2, PAR1, and multiple subunits of nicotinic receptors in bronchopulmonary C-fibers (Chuaychoo et al. 2006; Gu et al. 2008; Kwong et al. 2008a, b; Nassenstein et al. 2008). The responsiveness to such a variety of inflammatory mediators and environmental toxins and the reflexes initiated upon the activation of C-fibers lends credence to the notion that bronchopulmonary C-fibers are analogous to the nociceptors innervating somatic tissues.
TRPV1-dependent signaling is not the same in all bronchopulmonary C-fibers and is at least suggestive of the differential expression of a ligand-transporting system in some C-fibers or perhaps unique gating mechanisms for TRPV1 in the various bronchopulmonary C-fiber subtypes. Olvanil and anandamide are reasonably effective and potent activators of intrapulmonary C-fibers in rats and in guinea pigs, but are minimally effective at evoking tracheal/bronchial C-fiber action potential discharge or tachykinin release from the peripheral terminals of bronchial and tracheal C-fibers (Tucker et al. 2001; Lin and Lee 2002; Kollarik and Undem 2004; Lee et al. 2005). This inability to activate bronchial C-fibers is overcome with sustained incubation times, suggesting an impaired access to the intracellular binding site of TRPV1. Conversely, cooling the terminals of pulmonary C-fibers rendered them considerably less responsive to olvanil and anandamide, but equally responsive to capsaicin. These data may predict the expression of an anandamide-transporting system in pulmonary C-fibers that is absent in bronchial and tracheal C-fibers (Ligresti et al. 2004). To date, however, no protein subserving this transporting function has been identified (Glaser et al. 2005). It is thus interesting that activation of TRPV1 has been shown to promote the movement of extraordinarily large molecules from the extracellular to the intracellular space through the open TRPV1 channel (Meyers et al. 2003; Binshtok et al. 2007). The Hill coefficient for TRPV1 activation is significantly different from unity, suggestive of cooperative binding properties (Szallasi 1994; Welch et al. 2000; Undem and Kollarik 2002). It seems possible that threshold TRPV1 activation resulting in transient channel opening promotes additional agonist influx and further receptor activation. Perhaps some subtle modification of TRPV1 channel gating in C-fiber subtypes determines the ability of anandamide and olvanil to move through the open TRPV1 channel.
In addition to the autacoids listed above that activate bronchopulmonary C-fibers, many other mediators can sensitize them to subsequent activation. These include histamine via H1 receptors, cysteinyl leukotrienes via cysLT1 receptors, epinephrine via β3 receptors, and prostaglandin EP and TP receptor agonists (Karla et al. 1992; Lee and Morton 1993, 1995; McAlexander et al. 1998; Xiang et al. 2002; Gu et al. 2007). Prostaglandins also likely account for the sensitizing effects of protease-activated receptor 2 (PAR2) agonists on bronchopulmonary C-fibers (Gatti et al. 2006). Some mechanistic studies of these sensitizing effects have been carried out in patch-clamp analyses. Other unique characteristics regulating airway C-fiber activation include sensitivity to changes in extracellular Cl− and Ca2+ concentrations, changes in airway surface liquid osmolarity, TRPV1-independent activation by acid (perhaps involving acid-sensing ion channels), and activation/sensitization by CO2 (Delpierre et al. 1981; Pisarri et al. 1992; Fox et al. 1995; Pedersen et al. 1998; Kollarik and Undem 2002; Undem et al. 2003; Lin et al. 2005; Gu and Lee 2006).
Cough Receptors
C-fiber-selective stimulants that readily initiate coughing in awake human subjects and in awake guinea pigs have consistently failed to initiate cough in anesthetized cats, dogs, or guinea pigs. On the basis of the studies of Widdicombe (1954a, b) published in 1954 and the results of vagal cooling studies in cats and dogs by Tatar et al. (1988, 1994), it had become almost dogma that cough is initiated by activation of RARs. But many well-known and even selective stimuli for RARs, including a variety of bronchoconstrictors, negative airway luminal pressures, or inspiratory efforts against a closed glottis, have been consistently ineffective at evoking cough in either awake or anesthetized animals or humans. Recent studies carried out in guinea pigs and a reappraisal of Widdicombe's studies in cats suggest that a vagal afferent nerve subtype distinct from both C-fibers and RARs plays an essential role in regulating the cough reflex in anesthetized guinea pigs and cats and likely in any species that has a well-defined cough reflex. These afferents have thus been called “cough receptors” (Canning et al. 2004, 2006a, b; Canning and Chou 2009).
Cough receptors are differentiated from C-fibers and RARs in guinea pigs by conduction velocity. With a conduction velocity of approximately 5 ms−1, these afferents conduct action potentials considerably faster than C-fibers (1 ms−1 or less) but considerably slower than either RARs or SARs (more than 20 ms−1). Cough receptors are also differentiated from C-fibers and RARs by mechanical sensitivity, being exquisitely sensitive to punctate mechanical stimulation (5–10 times more sensitive than C-fibers) but utterly insensitive to changes in airway luminal pressure or airway smooth muscle contraction, both of which activate RARs. Also unlike C-fibers, the cough receptors are insensitive to capsaicin and bradykinin (Fig. 4). Cough receptors are activated by acid but entirely through TRPV1-independent mechanisms (Canning et al. 2004, 2006a, b).
Fig. 4.
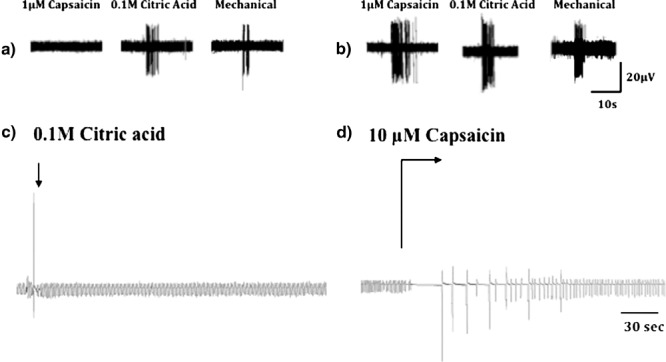
Electrophysiological characteristics of the extrapulmonary vagal afferent nerves regulating cough of guinea pigs. Cough receptors and C-fibers are both activated by punctate mechanical stimulation and by acid, but the cough receptors are insensitive to capsaicin. Capsaicin and other C-fiber-selective stimulants initiate coughing in awake animals and in awake human subjects, but have consistently failed to initiate coughing in anesthetized animals. In anesthetized guinea pigs, topical acid challenge of the tracheal mucusa initiates coughing, while topical capsaicin challenge does not evoke coughing. Rather, capsaicin challenge in anesthetized guinea pigs evokes respiratory slowing and, occasionally, a profound apnea followed by gasping and a gradual recovery of a normal respiratory pattern. (Reproduced with permission from Canning et al. 2004)
By combination of electrophysiological studies with intravital labeling methods, retrograde neuronal tracing, organotypic cultures, and immunohistochemistry, the peripheral terminals of cough receptors in the guinea pig trachea and bronchus have been identified (Canning et al. 2006a, b). Terminating between the epithelium and smooth muscle layers of the airways mucosa, the cough receptors assume a circumferential position in the extracellular matrix. Branching is extensive at the terminals, with axons projecting from longitudinal nerve bundles through the smooth muscle layer. Similar structures have been described in the airway mucosa of other species but their identity as “cough receptors” is unclear (Larsell 1921, 1922; Gaylor 1934; Yamamoto et al. 1995; Yu 2005; De Proost et al. 2007). Immunohistochemistry confirms the selective expression of subtypes of Na+–K+–ATPase and Na+–K+–2Cl− transporter in guinea pig cough receptors (Canning et al. 2006a, b; Mazzone and McGovern 2006, 2008). More recently, tetrodotoxin-insensitive Na+ channels have been localized to these cough receptors (Kwong et al. 2008a, b). Pharmacological analyses suggest that these regulators of ion flux and gradients, as well as Cl− channels and voltage-sensitive K+ channels, may be critical to the regulation of cough receptor responsiveness to chemical (acid) and punctate mechanical stimuli (Fox et al. 1995; McAlexander and Undem 2000; Canning et al. 2006a, b; Mazzone and McGovern 2006; Canning 2007). No other stimuli thus far studied, including a variety of autacoids and neurotransmitters and ion channel modulators, alter cough receptor excitability or the ability of acid or mechanical stimuli to initiate coughing in guinea pigs.
Autonomic Reflexes
Parasympathetic Nerve Regulation of Airway and Vascular Smooth Muscle and Mucus Secretion
Parasympathetic nerves play a primary role in regulating airway smooth muscle tone and glandular secretion in the airways and also regulate pulmonary and bronchial vascular tone (Canning 2006; Wine 2007). There are two anatomically, physiologically, and pharmacologically distinct parasympathetic pathways projecting to the airways with opposing effects on airway smooth muscle but synergistic effects on airway mucus secretion. Parasympathetic-cholinergic nerves initiate airway smooth muscle contraction, pulmonary vascular dilatation, and mucus secretion upon activation, with acetylcholine acting in each target tissue via muscarinic M3 receptors. Parasympathetic noncholinergic nerves also innervate the airways of most species, including humans. Noncholinergic parasympathetic nerves utilize the peptide transmitter vasoactive intestinal peptide and related peptides (pituitary adenylate cyclase activating peptide, peptide histidine isoleucine, peptide histidine methionine) as well as the gaseous transmitter nitric oxide (formed from arginine by the neuronal isoform of nitric oxide synthase). Upon activation, noncholinergic parasympathetic nerves evoke bronchodilatation, airway vascular dilatation, and mucus secretion. Coincident activation of cholinergic and noncholinergic parasympathetic nerves may have synergistic effects on airway glandular secretion (Choi et al. 2007; Wine 2007).
Airway and lung afferent nerve activation initiates myriad patterns of airway parasympathetic nerve responses (Canning 2006). At eupnea, basal parasympathetic tone appears to be necessarily dependent upon the ongoing activity of airway vagal afferent nerves, either RARs or C-fibers (Jammes and Mei 1979; Kesler and Canning 1999). With challenge, activation of bronchopulmonary C-fibers or RARs increases airway cholinergic and noncholinergic parasympathetic nerve activity (Fig. 5). Activation of intrapulmonary stretch receptors (SARs) by lung inflation or during the hyperpnea associated with exercise induces a withdrawal of parasympathetic cholinergic nerve activity and bronchodilatation, but has no effect on parasympathetic noncholinergic nerves.
Fig. 5.

Reflex-evoked, airway parasympathetic nerve-dependent regulation of airway smooth muscle tone in guinea pigs in situ. (a) The C-fiber-selective stimulant bradykinin evokes reflex bronchospasm largely independent of any direct effects on airway smooth muscle. Histamine-evoked reflex bronchospasm occurs secondary to its direct effects on airway smooth muscle, which in turn activates intrapulmonary RARs. Evidence for the selective effects of bradykinin and histamine on C-fibers and RARs, respectively, is apparent from the marked inhibition of bradykinin-evoked reflex bronchospasm by intravenous or intracerebroventricular administration of neurokinin receptor antagonists, which are without effect on histamine-evoked reflexes. Neurokinins are selectively expressed by C-fibers in guinea pigs. (b) When RARs and C-fibers are activated simultaneously, marked synergism is apparent. This synergistic effect of RAR and C-fiber activation on airway parasympathetic tone may result from central convergence in the nucleus of the solitary tract of these afferent nerve subtypes. (c, d) The mean data for reflex bronchospasm and whole-lung-inflation pressures evoked by histamine, bradykinin, or the combination of histamine and bradykinin. (Reproduced with permission from Canning et al. 2001 and Mazzone and Canning 2002a, b)
Reflex regulation of airway parasympathetic nerves by vagal afferents may not be entirely unidirectional. Secondary to the end-organ effects precipitated by parasympathetic nerve stimulation (e.g., mucus secretion, bronchospasm), action potential patterning in airway mechanoreceptors may change dramatically (Coleridge et al. 1982; Richardson et al. 1984). This is especially true under conditions in which tidal volumes are held constant (e.g., mechanical ventilation). An increase in parasympathetic cholinergic tone will decrease airway volume and deadspace, resulting in an increase in end-inspiratory pressure with mechanical ventilation and an increase in alveolar stretch under any mode of static volume ventilation. The increase in alveolar distension will favor an increase in SAR activation and a resulting withdrawal of cholinergic tone. In this way, airway afferent and efferent nerves may work in concert to establish a set point for airway parasympathetic tone (Fisher and Sant'Ambrogio 1982; Richardson et al. 1984; Matsumoto 1996). Perhaps in COPD, with alveolar destruction and increases in lung compliance, SAR activation may be diminished, prompting the elevation in airway cholinergic tone observed in this disease (Gross et al. 1989; Canning 2006).
Reflexes regulating noncholinergic airway parasympathetic nerves have been studied in guinea pigs, cats, and human subjects (Szarek et al. 1986; Ichinose et al. 1987, 1988; Michoud et al. 1987; Inoue et al. 1989; Lammers et al. 1989; Canning et al. 2001; Kesler et al. 2002; Mazzone and Canning 2002a). Unlike cholinergic contractions of the airway smooth muscle, which reach a near maximum within 30 s and can reverse at the same rate, noncholinergic parasympathetic nerve mediated relaxations of airway smooth muscle are slow in both onset and reversal (Chesrown et al. 1980; Diamond and O'Donnell 1980; Irvin et al. 1982; Matsumoto et al. 1985; Lama et al. 1988; Canning and Undem 1993; Canning et al. 2001; Kesler et al. 2002; Mazzone and Canning 2002a, b). Perhaps noncholinergic parasympathetic nerves function to restore or maintain airway patency during or at the conclusion of defensive reflexes (Coburn and Tomita 1973; Canning et al. 2006a). Consistent with this hypothesis, noncholinergic parasympathetic nerve activation is only modestly effective at preventing bronchospasm mediated reflexively or by direct actions on smooth muscle, but can gradually reverse an evoked contraction and modulate sustained cholinergic tone at eupnea (Aizawa et al. 1982, 1997, 1999; Bai et al. 1986; Szarek et al. 1986; Clerici et al. 1989; Miura et al. 1990; Inoue et al. 1991; Matsumoto et al. 1999; Canning et al. 2001; Kesler et al. 2002).
Reflex Regulation of Airway Sympathetic Nerves
Sympathetic nerves innervate the airways and lungs of all species. In most species, including humans, sympathetic-adrenergic innervation of intrapulmonary airway smooth muscle is limited or nonexistent (Canning 2006). In all species, sympathetic aderenergic nerves have been found innervating the airway vascular smooth muscle. Until recently, however, no study has directly addressed the reflex mechanisms controlling airway sympathetic nerve activity. We recently studied reflex regulation of airway sympathetic nerves innervating the trachealis of guinea pigs (Oh et al. 2006). The vagus nerves were cut bilaterally to limit the influence of airway parasympathetic nerves on smooth muscle tone. With the trachealis precontracted with histamine, capsaicin inhalation evoked a marked relaxation of the trachealis that was prevented by sympathetic denervation of the trachealis, propranolol, or dorsal rhizotomy (T1-T4). Retrograde tracing and electrophysiological analyses identified a population of capsaicin-sensitive spinal afferent nerves innervating the intrapulmonary airways and lungs. The majority of these spinal afferent nerves expressed substance P. Not surprisingly, then, neurokinin receptor antagonists prevented the reflex-mediated relaxations evoked by capsaicin inhalation.
Interestingly, we found that the sympathetic reflexes evoked in the airways by capsaicin inhalation occurred without any coincident cardiovascular responses (Oh et al. 2006). This adds further evidence against historical notions regarding sympathetic nerve function in homeostatic and defensive settings (Morrison 2001; Janig and Habler 2003). We also observed that stimulating the central cut ends of the vagus nerves evoked propranolol-sensitive relaxations of the trachealis (Oh et al. 2006). Vagal afferents are known to regulate sympathetic outflow to multiple organs, including the airways (Barman and Gebber 1976; Bachoo and Polosa 1987; Habler et al. 1994; Huang et al. 2000).
The Axon Reflex
In rats and in guinea pigs, bronchopulmonary C-fiber activation can also initiate an axon reflex, characterized by the peripheral release of neuropeptides that produce a variety of end-organ effects within the airways and lungs, including bronchospasm, mucus secretion, vascular engorgement, inflammatory cell recruitment, and plasma extravasation (Barnes 1986, 2001; Canning et al. 2006a, b). The prominent role of the axon reflex in the response to a variety of experimental challenges in rats and guinea pigs prompted a nearly two decade effort to address the hypothesis that respiratory disorders such as asthma and COPD were due in part to an axon reflex. This notion did not live up to its promise in rats and guinea pigs when evaluated in the human airways, in large part owing to the relative paucity of neuropeptide-containing afferent nerve terminals in the airways and lungs of humans (Hislop et al. 1990; Howarth et al. 1995; Chanez et al. 1998; Lamb and Sparrow 2002). It is nevertheless possible that axonal reflexes regulate human airway function, but through the actions of transmitters (e.g., ATP, glutamate) other than substance P, neurokinin A, and CGRP.
The most effective stimulants of the axon reflex work through the gating of the ion channel TRPV1. Capsaicin, for example, evokes a profound C-fiber discharge and an axon reflex, all of which are abolished when TRPV1 gating is prevented. By contrast, bradykinin, which acts only partially through TRPV1 gating on bronchopulmonary C-fibers, evokes little if any axon reflex (Mizrahi et al. 1982; Bramley et al. 1990; Schlemper and Calixto 2002). Other stimuli evoking an axon reflex include hypertonic saline, cold, dry air, PAR2 agonists, nicotine, immunosuppressants (cyclosporin A, FK 506), and TRPA1 receptor activation (Lundberg et al. 1983; Umeno et al. 1990; Mapp et al. 1991; Harrison et al. 1998; Pedersen et al. 1998; Yoshihara et al. 1998; Carr et al. 2000; Ricciardolo et al. 2000; Andresen and Saugstad 2008; Taylor-Clark et al. 2008).
A variety of stimuli have also been reported to inhibit the axon reflex through effects on the airway C-fiber terminal, including α2 adrenoceptor agonists, β2 adrenoceptor agonists, μ-opioid receptor agonists, GABAB receptor agonists, nociceptin, neurotensin, galanin, serotonin (via 5-HT1 receptors), prostaglandin E1, adenosine, phosphodiesterase type 4 inhibitors, neuropeptide Y, vasoactive intestinal peptide/pituitary adenylate cyclase activating peptide, dopamine D2 receptor agonists, bradykinin channel openers, and histamine H3 receptor agonists (Grundstrom et al. 1984; Belvisi et al. 1988, 1989; Giuliani et al. 1989; Kamikawa 1989; Matran et al. 1989; Aikawa et al. 1990; Stretton 1991; Verleden et al. 1993; Takahashi et al. 1994; Undem et al. 1994; Spina et al. 1995; Fox et al. 1997; Fischer et al. 1998; Shah et al. 1998; Birrell et al. 2002). It is tempting to speculate that the ability of these agents to inhibit the action-potential-independent axon reflex predicts a peripheral site of action of these drugs on bronchopulmonary C-fiber activation. This seems unlikely. Thus, prostaglandin E and adenosine both inhibit the axon reflex but activate and/or sensitize C-fibers to action potential formation (Kamikawa and Shimo 1989; Aikawa et al. 1990; Hong et al. 1998; Ho et al. 2000). The PAR2 agonist initiates an axon reflex but fails to initiate action potentials on airway C-fibers (Carr et al. 2000). Removal of extracellular Ca2+ reduces neuropeptide release from capsaicin-sensitive nerves, but enhances airway C-fiber excitability (Hua et al. 1992; Undem et al. 2003). Together, the data argue for an almost complete dissociation of the axon reflex from C-fiber action potential formation.
Respiratory Reflexes
Respiratory Pattern Changes and Respiratory Sensations
Changes in respiratory pattern attributable to airway afferent nerve activation have been studied extensively in animals (Fig. 3). Respiratory sensations such as dyspnea are less amenable to study in animals, but have been studied in human subjects. The classic triad of the pulmonary chemoreflex includes bradycardia and apnea followed by rapid shallow breathing (Green and Jackman 1984; Lee et al. 1995). Both the apnea and the rapid shallow breathing depend upon pulmonary C-fiber activation (Green and Jackman 1984). Apnea/respiratory slowing can also be evoked by C-fiber activation in the extrapulmonary airways of anesthetized animals (Palecek et al. 1989; Chou et al. 2008). In both animals and humans, activation of intrapulmonary C-fibers and RARs can initiate tachypnea (Mills et al. 1969; Green and Jackman 1984; Chou et al. 2008). In humans, the increase in respiratory rate evoked by pulmonary C-fiber activation with adenosine is accompanied by a sensation of dyspnea (Burki et al. 2005). Dyspnea and “breathlessness” can be reduced by airway or vagus nerve anesthesia or transection (Winning et al. 1985; Davies et al. 1987b; Taguchi et al. 1991). Prostaglandin E2 worsens the sensation of dyspnea (Taguchi et al. 1992). Bradykinin, a selective stimulant for airway C-fibers, reproduces the sensation of “sore throat” associated with upper respiratory tract infections (Proud and Kaplan 1988). Enhanced breaths (or sighs) become more frequent as airway lung compliance decreases. These have been attributed to the activation of RARs and may serve to open closed airways during tidal breathing at rest or during bronchospasm (Matsumoto et al. 1998; Dybas et al. 2006).
For good reason, much of the focus on respiratory sensations in disease has been directed to the activation of pulmonary C-fibers. But a role for SARs in respiratory sensations should not be discounted. The accumulation of CO2 in the alveoli would limit SAR discharge, delaying inspiratory termination and thus prompting hyperpnea. In COPD, with alveolar destruction, the lung stretch associated with a normal tidal volume may have limited stretching effects in the peripheral airways and thus may limit SAR discharge, prompting a compensatory increase in end expiratory lung volume or an enhanced sensation of air hunger despite normal or near-normal blood gases. The Na–K+–2Cl transport inhibitor furosemide is reported to diminish air hunger sensation during breath hold, perhaps owing to an inhibition of RAR discharge but an enhancement of SAR discharge (Nishino 2000; Sudo et al. 2000).
Cough
The cough reflex is initiated by activation of the cough receptors and by activation of a C-fiber subtype innervating the large airways (Canning and Chou 2009). The role of C-fibers in cough has been the subject of considerable debate. The chemical stimuli most effective at activating bronchopulmonary C-fibers, including capsaicin, bradykinin, and acid, are similarly very effective at initiating cough in conscious human subjects and in conscious animals (Forsberg et al. 1988; Laude et al. 1993; Karlsson and Fuller 1999; Jia et al. 2002; Trevisani et al. 2004; Dicpinigaitis 2007). These stimuli work entirely or partly through TRPV1, and immunohistochemical and single-cell PCR confirms expression of TRPV1 in airway C-fibers (Myers et al. 2002; Groneberg et al. 2004; Watanabe et al. 2006; Kwong et al. 2008a, b). Prior capsaicin desensitization prevents citric acid induced coughing in awake guinea pigs, as does pretreatment with TRPV1 receptor antagonists (Forsberg et al. 1988; Bolser et al. 1991; Lalloo et al. 1995; Trevisani et al. 2004; Gatti et al. 2006; Leung et al. 2007). Taken together, these and other observations argue strongly for a role of bronchopulmonary C-fibers in cough (Canning et al. 2006a, b). But C-fiber-selective stimuli have consistently failed to evoke coughing in anesthetized animals (Tatar et al. 1988; Karlsson et al. 1993; Tatar et al. 1994; Canning et al. 2004, 2006a, b). Anesthesia has no effect on coughing evoked by mechanical or acid stimulation of the airway mucosa and does not prevent C-fiber activation or other C-fiber-dependent reflexes, and yet capsaicin and bradykinin do not evoke cough in anesthetized animals (Coleridge and Coleridge 1984; Tatar et al. 1988; Canning et al. 2006a, b).
Perhaps it should be expected that C-fiber-selective stimulants would fail to evoke coughing in anesthetized animals. Airway and lung C-fibers share many characteristics with somatosensory nociceptors, and it is the objective of general anesthesia to prevent the sensations and reflexes associated with nociceptor activation. But while the effects of anesthesia on nociceptor signaling may explain the inability of C-fiber-selective stimulants to evoke coughing in anesthetized animals, anesthesia cannot account for the known acute inhibitory effects C-fiber activation may have on cough in anesthetized animals, or the inability of some C-fiber stimuli to evoke coughing in conscious animals and in conscious human subjects (Tatar et al. 1988, 1994). We have recently addressed the hypothesis that C-fiber subtypes might account for these opposing effects on cough. Subtypes have been described in several species (Coleridge and Coleridge 1984; Kollarik et al. 2003; Undem et al. 2004). In guinea pigs, airway vagal C-fiber subtypes can be differentiated by their ganglionic origin, distribution in the airways, and responsiveness to ATP, adenosine, and serotonin 5-HT3 receptor agonists (Undem et al. 2004; Chuaychoo et al. 2005, 2006). The ability of C-fiber activation to evoke coughing in awake guinea pigs is reasonably well established, and we also reported a facilitating effect of C-fiber activation on cough (Mazzone et al. 2005; Canning et al. 2006a, b). In these latter studies, capsaicin or bradykinin applied topically to the tracheal mucosa greatly enhanced sensitivity to subsequent tussive stimuli. On the basis of the location of these bradykinin and capsaicin challenges, C-fibers arising from the jugular ganglia likely promote coughing. By inference, then, we further speculated that nodose C-fiber activation might acutely inhibit coughing. Consistent with this hypothesis, we found that selective activation of nodose C-fibers with adenosine or 2-methyl-5-hydroxytryptamine did not evoke coughing but greatly reduced the ability of citric acid to evoke coughing in anesthetized animals. Prior adenosine inhalation also inhibited capsaicin-induced coughing in conscious guinea pigs.
The results of studies carried out in other species are at least consistent with the notion that C-fiber subtypes may have opposing effects on cough. In anesthetized dogs and cats, C-fiber activation by bradykinin, capsaicin, or phenyldiguanide (a 5-HT3 receptor agonist) does not induce cough but can inhibit cough (Tatar et al. 1988, 1994; Karlsson et al. 1993). In rabbits, a species in which cough can be evoked by citric acid aerosol inhalation (consistent with a TRPV1- and C-fiber-dependent mechanism; Tatar et al. 1997, Adcock et al. 2003), it has also been reported that sulfur dioxide inhalation is acutely inhibitory for cough (Hanacek et al. 1984). Sulfur dioxide is known to activate lung C-fibers (Ho et al. 2001). Adcock et al. (2003) speculated that the inhibitory effects of the compound RSD931 in cough induced in rabbits might be due to its ability to activate pulmonary C-fibers. Humans readily cough to capsaicin and bradykinin challenge, but are refractory to serotonin and adenosine challenge (Stone et al. 1993; Burki et al. 2005) while intravenous capsaicin infusion is only minimally effective at evoking cough (Winning et al. 1986). There is also a report of serotonin-mediated inhibition of cough in human subjects (Stone et al. 1993). A comparable inability of intravenously capsaicin to evoke coughing has been reported in studies using conscious nonhuman primates (Deep et al. 2001).
CNS Pharmacology and Central Interactions Between Airway Afferent Nerve Subtypes
Studies of airway reflexes in response to stimuli known to be selective for the various airway afferent nerve subtypes largely substantiate the accepted classification schemes for afferent nerves. Implicit in the observation that afferent nerve subtypes subserve distinct reflex functions is that central termination sites of the various afferent nerve subpopulations must diverge to some extent, allowing for reflex specificity. From the little published evidence available, this notion would seem to be substantiated. Most of the work on central terminations of airway sensory nerves has been carried out in cats and rats. Bronchopulmonary C-fibers and RARs terminate extensively and often bilaterally in the nucleus of the solitary tract (nTS), particularly in the commissural and medial subnuclei (Davies and Kubin 1986; Kalia and Richter 1988; Bonham and Joad 1991; Ezure et al. 1991; Kubin et al. 1991; Lipski et al. 1991; Otake et al. 1992; Mazzone and Canning 2002a, b; Kubin et al. 2006). SARs terminate primarily ipsilateral to their vagal origin, rostral to obex in the lateral and interstitial subnuclei (Kalia and Richter 1985; Davies et al. 1987a,b; Bonham and McCrimmon 1990; Ezure et al. 2002; Kubin et al. 2006). No attempt at differentiating termination sites of RAR, SAR, or C-fiber subtypes has been described. In addition to the studies of SAR, RAR, and bronchopulmonary C-fiber termination sites, some work has been done to identify the nTS subnuclei regulating the cough reflex (Gestreau et al. 1997; Ohi et al. 2005; Jakus et al. 2008).
Electrophysiological and functional studies show evidence for bronchopulmonary afferent nerve convergence in the CNS (Takagi et al. 1995; Paton 1998; Silva-Carvalho et al. 1998). Coincident activation of airway afferent nerve subtypes can have synergistic effects on airway reflexes, including reflex bronchospasm and cough (Mazzone and Canning 2002a, b; Mazzone et al. 2005) (Fig. 5). Such synergistic interactions may explain the association between extrapulmonary disorders (e.g., gastroesophageal reflux disease, allergic rhinitis) and cough.
Several studies have characterized the pharmacology of the primary central synapses for airway vagal afferent nerves and have revealed a prominent role for glutamate acting via non-NMDA receptors (Bonham et al. 1993; Vardhan et al. 1993; Karius et al. 1994; Chianca and Machado 1996; Wilson et al. 1996; Aylwin et al. 1997; Ezure et al. 1999; Haxhiu et al. 2000; Mutolo et al. 2007, 2008). Notably, however, NMDA receptor activation plays an essential role in the initiation of cough, explaining in part the ability of the antitussive agent dextromethorphan to prevent coughing in animals and in human subjects (Canning et al. 2004, 2006a, b; Mutolo et al. 2007). Other agents shown to act centrally in nTS to regulate airway vagal reflexes include μ-opioid receptor agonists (codeine, DAMGO), GABAB receptor agonists, sigma agonists, and TRPV1 receptor agonists (Mazzone and Geraghty 1999; Mazzone et al. 2005; Ohi et al. 2005, 2007; Mutolo et al. 2007, 2008). Serotonin (5-HT) receptor antagonists have also been shown to act centrally to modulate airway reflexes, but their site of action has not been determined (Bootle et al. 1996).
The tachykinins substance P and neurokinin A have been localized to airway afferent neurons, and tachykinin receptor antagonists have been shown to reduce or abolish coughing evoked in guinea pigs, dogs, rabbits, cats, and pigs (Advenier and Emonds-Alt 1996; Bolser et al. 1997; Moreaux et al. 2000; House et al. 2004; Mutolo et al. 2008). Capsaicin microinjection in nTS evokes respiratory reflexes in rats that are abolished by neurokinin receptor antagonists, while coughing evoked in rabbits and sensitization of cough induced in guinea pigs is markedly inhibited or abolished by nTS microinjection of neurokinin receptor antagonists (Mazzone and Geraghty 1999; Mazzone et al. 2005; Mutolo et al. 2008). A central site of action for neuroknin receptor antagonists in cough in cats and in guinea pigs has also been suggested (Bolser et al. 1997). Reflex bronchospasm evoked by laryngeal capsaicin and by intravenous bradykinin in guinea pigs is also prevented by centrally acting neurokinin receptor antagonists (Canning et al. 2001; Mazzone and Canning 2002a, b) (Fig. 5). Neurokinin-1 receptor antagonists are also used clinically to treat emesis, a vagal reflex in humans that has many similarities to the cough reflex (Hornby 2001; Warr 2006). It seems likely then that neurokinins released from the central terminals of airway afferent nerves may also modulate airway reflexes in humans and in other species. It is thus interesting and confusing that in electrophysiological recordings of nTS neurons receiving synaptic input from airway afferent nerves, little evidence for an excitatory effect of neurokinins in otherwise healthy animals has been reported. Indeed, in one study, exogenously administered substance P was found to act presynaptically to depress synaptic transmission in nTS (Sekizawa et al. 2003). Many of these studies involved recording from unidentified synapses or the synapses of RARs or SARs, which are unlikely to express substance P under normal conditions. But even in recordings in C-fiber relay neurons, synaptic transmission has been explained entirely by the actions of glutamate (Wilson et al. 1996; Mutoh et al. 2000). This suggests that under the experimental conditions used for the electrophysiological recordings done to date, solitary tract stimulation is subthreshold in intensity, frequency, or duration for tachykinin release, the neurons selected for recording (i.e., neurons receiving monosynaptic input) are typically devoid of direct tachykinin input, or the process of tissue harvest and slice preparation effectively silences neurokinin-mediated effects in nTS.
Airway Sensory Nerves and Bronchial Hyperresponsiveness
Defining Characteristics of Bronchial Hyperresponsiveness
A number of clinical features distinguish asthmatic subjects from other respiratory diseases and may be considered characteristic of this phenotype (Avital et al. 1995). These include an exacerbation of disease following exposure to β-adrenoceptor antagonists (Bond et al. 2007), an impairment in the ability to bronchodilate following deep inspiration (Slats et al. 2007), and their bronchoconstrictor sensitivity to a wide range of innocuous stimuli (Cockcroft and Davis 2006; Van Schoor et al. 2002).
It is well established that asthmatic subjects are invariably more responsive to a range of stimuli, as expressed by an increase in provocative concentration that induces a 20% fall in forced expiratory volume in 1 s termed “bronchial hyperresponsiveness” (BHR). However, not only is there an increase in the sensitivity of the airways to a stimulus, but there is also an increase in the maximum degree of airway narrowing for a given dose of agonist (Fig. 6). The importance of understanding the underlying mechanism contributing toward BHR is confirmed by a study showing that treating the underlying hyperresponsiveness leads to a better improvement in asthma symptoms (Sont et al. 1999). A number of mechanisms have been proposed to account for why asthmatic subjects are invariably more responsive to the external environment. These include an alteration in airway geometry due to an increase in airway smooth muscle thickness that would lead to a greater degree of airway narrowing for a given dose of agonist and/or perturbations in myosin-actin function resulting in a loss in the ability of smooth muscle to dilate in response to deep inspiration, thereby leading to enhanced bronchoconstrictor responses (An et al. 2007; Gil and Lauzon 2007); the release of cytokines and growth factors from epithelial cells which stimulate mesenchymal cells and promote structural changes in the airways leading to airway remodeling, airway inflammation, and BHR (Holgate 2007); and recruitment and activation of dendritic cells, T lymphocytes, and eosinophils whose cell-derived products trigger a cascade of events within the lung leading to epithelial cell damage, increased smooth muscle contractility, and airway remodeling (Beier et al. 2007; Hammad and Lambrecht 2007; Jacobsen et al. 2007; Kallinich et al. 2007; Lloyd and Robinson 2007; Rosenberg et al. 2007). These mechanisms are all thought to contribute toward BHR in asthma, are likely to be interrelated, and contribute to the overall expression of BHR. However, there is also good evidence for the contribution of airway sensory nerves in this phenomenon (Spina and Page 2002) that might be likened to allodynia and/or hyperalgesia, which are characteristic of pain syndromes (Carr and Undem 2003; Undem et al. 2002).
Fig. 6.

Bronchial hyperresponsiveness (BHR) in asthma. It is convenient to measure changes in forced expiratory volume in 1 s (FEV1) to increasing doses of methacholine. In asthma, there is an increase in sensitivity (leftward position of the dose–response curve) often measured in terms of PC20 (dotted line) and reactivity (increase in slope) and in severe cases of the disease an inability to define the maximum degree value for airway narrowing compared with healthy subjects. However, BHR as measured by changes in FEV1 to increasing doses of methacholine may not be a sensitive indicator of the asthma phenotype (see the text). An increase in BHR can occur during exacerbation of disease as observed naturally during the pollen season, in the case of an allergic asthmatic, or following the deliberate exposure to a relevant antigen (arrow). However, asthmatic subjects are invariably responsive to a wide range of physiological stimuli that are otherwise refractory in healthy subjects. An understanding of the mechanisms by which these stimuli induce bronchoconstriction suggests that sensitization of afferent pathways may underlie this phenomenon
Bronchial Hyperresponsiveness and Sensory Nerves
It is common for clinicians to use stimuli such as methacholine and histamine to induce bronchoconstriction because these agents are relatively convenient to use. However, although there is a separation in airways responsiveness to these agents between asthmatic subjects and healthy individuals, there is a considerable degree of overlap and it has been suggested that these agents may not be sensitive indicators of the asthma phenotype (Avital et al. 1995; O'Connor et al. 1999). In contrast, asthmatic subjects invariably bronchoconstrict in response to the indirect-acting stimuli described earlier, which provoke little if any response in otherwise healthy individuals or in subjects with other respiratory diseases (Avital et al. 1995; Van Schoor et al. 2000).
Asthmatic subjects bronchoconstrict in response to a number of physiological stimuli such as exercise, distilled water, cold air, and hypertonic saline which are otherwise refractory in healthy subjects. Similarly, acidification, pollutants such as sulfur dioxide, and chemical substances, including adenosine, bradykinin, and neuropeptides, evoke bronchoconstriction in asthma but have little if any effect in nondiseased individuals. These agents are commonly referred to as “indirect-acting stimuli,” since they do not appear to mediate bronchoconstriction by direct activation of airway smooth muscle. They are thought to elicit bronchospasm by activating a number of different cell types, including mast cells, vascular smooth muscle cells, and vascular endothelial cells, and/or airway nerves (Spina and Page 1996, 2002; Van Schoor et al. 2000). A number of studies which measured the generation of action potentials from individual afferent nerves using well-established electrophysiological techniques have shown that stimuli including sulfur dioxide, acidification, distilled water, bradykinin, neuropeptides, and adenosine can activate C-fiber and Aδ-fibers in vivo (Table 1). It is therefore of interest that asthmatic subjects are sensitive to such stimuli, whereas healthy subjects are invariably unresponsive to these agents (Van Schoor et al. 2000).
Table 1.
Electrophysiological evidence for activation of afferent nerves in vivo by substances that elicit bronchoconstriction in asthmatic subjects
| Stimulus | RARs | C-fibers | References |
|---|---|---|---|
| Sulfur dioxide | Cat, rabbit | Dog, rat | Widdicombe (1954a, b), Boushey et al. (1974), Ho et al. (2001), Matsumoto et al. (1997), Roberts et al. (1982) |
| Distilled water | Dog | Dog, guinea pig | Fox et al. (1995), Pisarri et al. (1992) |
| Bradykinin | Dog | Kaufman et al. (1980) | |
| Guinea pig | Guinea pig | Bergren (1997), Fox et al. (1993), Ricco et al. (1996) | |
| Mouse | Kollarik and Undem (2004) | ||
| Neuropeptides | Rabbit | Rabbit, guinea pig | Bergren (2006), Bonham et al. (1996), Prabhakar et al. (1987) |
| Capsaicin | Cat, guinea pig | Cat, dog, guinea pig, rat, mouse | Armstrong and Luck (1974), Bergren (1997) |
| Mohammed et al. (1993), Morikawa et al. (1997) | |||
| Coleridge and Coleridge (1977), Dixon et al. (1980), Fox et al. (1993), Ho et al. (2001), Jackson et al. (1989), Kollarik and Undem (2004), Ricco et al. (1996) | |||
| Adenosine | Rat, guinea pig | Chuaychoo et al. (2006), Hong et al. (1998), Kwong et al. (1998) | |
| Endotoxin | Rat | Rat | Lai et al. (2005), Ruan et al. (2005) |
RARs rapidly adapting receptors
This suggests that the mechanisms by which these stimuli provoke bronchoconstriction are upregulated in asthma and characteristic of this phenotype. Furthermore, airways inflammation appears to be correlated better with BHR to indirect stimuli such as adenosine (van den Berge et al. 2001), bradykinin (Polosa et al. 1998; Roisman et al. 1996), and hypertonic saline (Sont et al. 1993) than it is to more direct acting stimuli such as methacholine. Similarly, during an exacerbation of BHR following the deliberate exposure of an asthmatic subject to an environmental allergen (e.g., house dust mite) there is a preferential increase in BHR to an indirect-acting stimulus such as bradykinin compared with methacholine (Berman et al. 1995). On the other hand, a number of pharmacological drugs used to treat asthma, including nedocromil sodium and ipratropium bromide, suppress airways responsiveness to these indirect-acting stimuli, suggesting the likely involvement of neural reflexes (Van Schoor et al. 2000). Furthermore, it is now recognized that glucocorticosteroids preferentially suppress BHR to adenosine (Ketchell et al. 2002; van den Berge et al. 2001) and bradykinin (Reynolds et al. 2002) compared with methacholine.
It is also noted that there is often a wide variability in airway sensitivity to spasmogens in subjects with mild asthma and there is little or very poor correlation between airway sensitivity to indirect-acting bronchoconstrictor agents such as adenosine and sensitivity to direct-acting stimuli such as methacholine. Also, BHR to an indirect-acting stimulus is greater during an exacerbation of asthma and lower following anti-inflammatory treatment compared with BHR to methacholine, which directly activates airway smooth muscle (O'Connor et al. 1999; Van Schoor et al. 2000). This apparent lack of correlation between bronchoconstrictor potency of these different types of stimuli suggest that alteration in the thickness of the airway wall (i.e., airway remodeling) alone cannot account for these discrepancies. If airway remodeling were responsible, then there would be a better correlation between bronchoconstrictor potency of indirect-acting and direct-acting stimuli. Furthermore, inflammatory insults to the airway wall would cause a similar change in BHR to these different agents and, finally, there would be no preferential effect of drug treatment on BHR to different stimuli.
Together, the findings of clinical studies support the notion that inflammatory insults to the lung might increase the activity of neuronal pathways, thereby resulting in heightened sensitivity of the lungs to these indirect-acting stimuli. Furthermore, one cannot view BHR as a nonspecific phenomenon, that is to say, that asthmatics are hyperresponsive to all stimuli, but rather it is increasingly apparent that BHR is more heterogeneous than is widely appreciated (O'Connor et al. 1999) and sensory nerves might be a common pathway through which BHR is manifested in respiratory diseases such as asthma.
TRPV1 and Bronchial Hyperresponsiveness
Transient receptor potential (TRP) channels are protein sensors for the perception of pain, taste, hearing, and smell and comprise at least six subfamilies (Nilius et al. 2007). One member of this superfamily (TRPV1) is predominantly localized to small-diameter afferent neurons in dorsal and vagal sensory ganglia (Szallasi and Blumberg 1999) and activation by capsaicin gives rise to feelings of warmth, heat, and pain. The cloning and expression of TRPV1 has increased our understanding of the role of this protein in neurogenic pain, but also with migraine, cough, irritable bladder disease, and gastrointestinal inflammation (Cortright et al. 2007; Geppetti and Trevisani 2004; Jia and Lee 2007; Kollarik et al. 2007; Liddle 2007; Ma and Quirion 2007; Okajima and Harada 2006; Storr 2007). The role of these proteins in contributing to BHR has been investigated in a number of experimental models. It has long been recognized that capsaicin selectively activates a subpopulation of afferent nerves, the neuropeptide containing C-fibers. However, it is now well established that capsaicin may also target a subset of airway Aδ-fibers whose cell nuclei reside within the jugular but not nodose ganglion (Myers et al. 2002). The activation of both of these nerve types can lead to a number of physiological changes within the airways, including reflex bronchoconstriction, release of sensory neuropeptides, edema, cough, and submucosal gland secretion (De Swert and Joos 2006).
Chronic treatment with capsaicin in various animal species leads to an impairment of somatosensory function as a consequence of depletion of sensory neuropeptide content, downregulation of TRPV1 receptor expression, and/or destruction and loss of sensory nerves (Watanabe et al. 2005, 2006). A consequence of chronic treatment with capsaicin upon neural function in the lung is an attenuation of BHR induced by a range of stimuli (Table 2). Thus, BHR induced by exposing nonallergic animals to lipid mediators, including platelet-activating factor and 15-hydroperoxyeicosatetraenoic acid, is attenuated. A similar observation was noted when BHR was elicited following exposure of nonallergic animals to lipopolysaccharide, ozone, citric acid, parainfluenza-3 virus, and poly(l-lysine) or exposure of allergic animals to inhaled antigens (Table 2). It has been concluded that the peripheral release of sensory neuropeptides per se was responsible for inducing BHR because depletion of sensory neuropeptides within the airways was a natural consequence of chronic treatment with capsaicin. Indeed a variety of animal experimental data have provided a wealth of information concerning the potential role of tachykinins such as substance P, neurokinin A, and CGRP in altering both resident lung and inflammatory cells, thereby perpetuating the inflammatory process in the lung and contributing to this process and inducing BHR (De Swert and Joos 2006).
Table 2.
Studies demonstrating a role for sensory nerves in bronchial hyperresponsiveness (BHR)
| Stimulusa | Spasmogen | Species | Effect of chronic capsaicin treatment on BHR | Effect of chronic capsaicin treatment on inflammatory cell recruitment | References |
|---|---|---|---|---|---|
| Antigen | Acetylcholine | Guinea pig | Inhibited | No effect | Matsuse et al. (1991) |
| Histamine | Rabbit | Inhibited | No effect | Herd et al. (1995) | |
| Histamine | Rabbit | Inhibited | No effect | Riccio et al. (1993)f | |
| Serotonin | Rat (BNxWi/Fu) | Increasedc | Increasede | Ahlstedt et al. (1986)f | |
| TDI | Acetylcholine | Guinea pig | Inhibited | Not measured | Thompson et al. (1987) |
| Acetylcholine | Rabbit | Inhibited | Not measured | Marek et al. (1996) | |
| SO2 | Methacholine | Rat (Sprague-Dawley) | Augmentedc | Not measured | Long et al. (1997)f |
| Not measured | Augmented | Long et al. (1999)f | |||
| LPS | Histamine | Guinea pig | Inhibited | Inhibited | Jarreau et al. (1994) |
| Histamine | Guinea pig | Augmentedd | Not measured | Loeffler et al. (1997) | |
| Ozone | Histamine | Guinea pig | Inhibited | Not measured | Tepper et al. (1993) |
| Histamine | Guinea pig | Inhibited | Inhibited | Koto et al. (1995) | |
| Methacholine | Rat (Sprague-Dawley) | Augmentedc | Not measured | Jimba et al. (1995)f | |
| Rat (Wistar) | Not measured | Augmented | Vesely et al. (1999)f | ||
| EFSb | Ferret | Inhibited | Not measured | Wu et al. (2003) | |
| Citric acid | Acetylcholine | Guinea pig | Inhibited | Not measured | Girard et al. (1996) |
| Parainfluenza-3 | Acetylcholine | Guinea pig | Inhibited | Not measured | Riedel et al. (1997) |
| Cigarette smoke | Acetylcholine | Guinea pig | Inhibited | Not measured | Daffonchio et al. (1990) |
| PAF | Histamine | Rabbit | Inhibited | Neutrophils inhibited; eosinophils not inhibited | Spina et al. (1991) |
| Acetylcholine | Guinea pig | Not measured | Perretti and Manzini (1993) | ||
| 15-HPETE | Histamine | Rabbit | Inhibited | No effect | Riccio et al. (1997) |
| Poly(l-lysine) | Methacholine | Rat (Sprague-Dawley) | Inhibited | Not determined | Coyle et al. (1994)f |
| Nonallergic chronic rhinitis | Nasal resistance inhibited; symptom scores reduced | Lacroix et al. (1991) | |||
| Allergic rhinitis | Hyperosmolar response inhibited; symptom scores reduced | Sanico et al. (1999) |
15-HPETE 15-hydroperoxyeicosatetraenoic acid, LPS lipopolysaccharide, PAF platelet-activating factor, TDI toluene diioscyanate, EFS electrical field stimulation
aStimulus denotes a substance used to induce bronchial hyperresponsiveness
bEFS and BHR was measured in vitro
cUnlike in guinea pigs, rabbits, ferrets and humans, neurokinins released from capsaicin-sensitive nerves are bronchodilators in rats (and mice), which may explain in part the augmentation of bronchial responsiveness seen with capsaicin pretreatment in rats (Manzini 1992; Szarek et al. 1995)
dBHR augmented at 2 h but not at 1 and 3 h after LPS challenge
eData not analyzed statistically
fTreatment of neonates with capsaicin but measurement of bronchial hyperresponsiveness and inflammation performed in adult animals
It is therefore surprising that neurokinin antagonists have thus far proved disappointing in clinical trials in asthma (De Swert and Joos 2006). However, the rationale for the development of tachykinin antagonists was based on the assumption that local release of tachykinins from C-fibers was sufficient to perpetuate the inflammatory response. One can only conclude that other mediators released within the airways contribute to the inflammatory process, or the possibility that capsaicin is selective for C-fiber afferents in the airways needs to be revised since TRPV1 may also be localized to non-C-fiber afferents, or is not necessarily colocalized with sensory neuropeptides (Guo et al. 1999; Myers et al. 2002; Tominaga et al. 1998; Watanabe et al. 2005, 2006), and, therefore, sensory-neuropeptide-independent mechanisms may also operate in the lung. Furthermore, most studies showing an importance for tachykinins in mediating BHR stem from studies conducted in the guinea pig, an animal rich in sensory neuropeptide innervation in the lung. In contrast, the rabbit is relatively resistant to the neuropeptide-depleting effects of capsaicin despite the inhibition of BHR induced by nonimmunological and immunological methods (Herd et al. 1995; Riccio et al. 1997)and, therefore, mechanisms other than neuropeptide depletion must account for this phenomenon, and the peripheral release of sensory neuropeptides is not obligatory for the development of BHR. This may be one reason why, thus far, selective neurokinin antagonists have proved disappointing in the treatment of asthma (Spina and Page 2002).
Studies using capsaicin have shown that the functional effect of neuropeptides on airway smooth muscle is species-dependent. Rabbit, monkey, and human airways contract weakly on exposure to capsaicin in vitro (Ellis et al. 1997; Spina et al. 1998), whereas guinea-pig airways are very sensitive (Grundstrom et al. 1981). These variable functional effects are superficially consistent with the differential localization of substance P and CGRP in the airways across species, as the occurrence of these neuropeptide-containing nerves in human and rabbit tends to be sparse (Hislop et al. 1990; Howarth et al. 1995; Laitinen et al. 1983; Lundberg et al. 1984), whereas neuropeptides are found widely throughout the airways of guinea pigs (Hua et al. 1985; Nohr and Weihe 1991; Saria et al. 1985).
Hyperalgesia induced by chemical and thermal stimuli is suppressed in TRPV1 knockout mice, suggesting that this protein is an important transducer of pain (Gunthorpe et al. 2002; Julius and Basbaum 2001). It is therefore of interest that chronic treatment with capsaicin in humans can lead to a suppression of allodynia and hyperalgesia induced by intradermal injection of capsaicin (Davis et al. 1995) and when applied topically to the nose, capsaicin reduces nasal hyperresponsiveness in allergic rhinitis patients (Sanico et al. 1999). A loss in TRPV1 signaling might help explain the loss in BHR observed in various experimental animal models (Spina and Page 2002).
TRPV1 Antagonist and Knockout Studies
If the activation of TRPV1 is important for the sensitization of primary afferent nerves in the lung, then it would seem reasonable to suggest that pharmacological antagonism of this protein might reduce BHR. Unfortunately, very few studies have specifically addressed this question in the context of BHR. In one study, BHR was induced in the guinea pig by acute challenge with platelet-activating factor, which was inhibited following pretreatment with ruthenium red, a nonselective TRP channel blocker (Perretti and Manzini 1993). Studies using TRPV1 antagonists may require an additional control to rule out any possible bronchorelaxant capabilities that may confound any potential effect on BHR (Skogvall et al. 2007).
The effect of TRPV1 antagonists on cellular recruitment has been investigated in a number of inflammatory models. The TRPV1 antagonist N-(4-chlorobenzyl)-N′-(4-hydroxy-3-iodo-5-methoxybenzyl) thiourea had no effect on neutrophil recruitment induced by injection of complete Freund's adjuvant into the hindpaw of mice (Tang et al. 2007). In contrast, capsazepine inhibited neutrophil recruitment to the lung induced by systemic administration of hydrogen sulfide donor (Bhatia et al. 2006) and inhibited neutrophil recruitment in a model of colitis (Kihara et al. 2003) and pancreatitis (Hutter et al. 2005). Differences in the severity of the inflammatory models utilized in these studies may account for the lack of general consensus concerning the role of TRPV1 in inflammatory cell recruitment; however, as indicated earlier, the findings of most chemical ablation studies using chronic treatment with capsaicin are consistent with the peripheral involvement of sensory neuropeptides in neutrophil recruitment to sites of inflammation (Table 2).
There has also been a paucity of studies utilizing TRPV1 gene deficient mice to study the role of this protein in BHR and inflammation. In one study, BHR in response to lipopolysaccharide challenge was significantly augmented in TRPV1 knockout mice, and highlighted the existence of an anti-inflammatory substance (e.g., somatostatin) released from TRPV1-positive cells, which could act in a negative-feedback mechanism to limit the inflammatory response (Helyes et al. 2007). However, these data are not consistent with those form chemical ablation studies showing that BHR to lipopolysaccharide is inhibited in the guinea pig (Jarreau et al. 1994). This discrepancy may be due to the two different methods employed to “impair” TRPV1 signaling. It could be envisaged that sensory nerves would be bombarded by multiple signals by a plethora of mediators released following the initial insult, resulting in the activation of various receptor proteins (e.g., bradykinin and NGF receptor, other TRPs) on primary afferent terminals. These signals would be processed at the level of the nTS, but would also require the activation of TRPV1 for a facilitated response (i.e., gain in function). However, the interpretation of these signals by the nTS would be lost following chemical ablation with capsaicin, owing to destruction of the peripheral terminations of C-fibers in the airway, but these would be retained in TRPV1 knockout mice with an intact afferent nervous system and therefore would still able to signal to the nTS. Alternatively, compensatory mechanisms during the development of TRPV1 knockout mice might account for this anomaly. The implication is that impairment of sensory nerve function (e.g., via TRPV1 desensitization) may be required instead of TRPV1 antagonism to completely suppress BHR.
In terms of the inflammatory response, it appears that neutrophil recruitment in joint inflammation was either unaffected (Keeble et al. 2005) or augmented during inflammatory insults to the lung (Helyes et al. 2007) and gastrointestinal tract (Massa et al. 2006). Similarly, the amounts of TNF-α released within the extracellular space at sites of inflammation were either increased (Clark et al. 2007) or unaffected (Keeble et al. 2005) by gene deletion of TRPV1 in different inflammatory models. Hence, these murine models have been inconclusive concerning the role of TRPV1 in mediating BHR and/or inflammation. The observation that activation of TRPV1 may stimulate the release of an anti-inflammatory substance in the mouse also makes it difficult to elucidate the role of TRPV1 in BHR and inflammation in this species (Helyes et al. 2007).
Conclusions
Airway sensory nerves play an essential role in regulating airway and lung defensive and homeostatic reflexes. The afferent nerve subtypes regulating these reflexes have unique physiological and pharmacological attributes that are amenable to selective therapeutic interventions. There is extensive evidence to suggest that airway sensory nerves are dysregulated in disease. Therapeutic strategies that target the excitability of airway sensory nerves at their central and peripheral terminations may provide symptom relief in conditions such as cough, asthma, and COPD.
Contributor Information
Brendan J. Canning, Phone: +1410550-2130, FAX: +1410550-2130, Email: bjc@jhmi.edu
Domenico Spina, Email: domenico.spina@kcl.ac.uk.
B.J. Canning, Email: bjc@jhmi.edu
D. Spina, Email: domenico.spina@kcl.ac.uk
References
- Adcock JJ, Douglas GJ, Garabette M, Gascoigne M, Beatch G, Walker M, Page CP. RSD931, a novel anti-tussive agent acting on airway sensory nerves. Br J Pharmacol. 2003;138:407–416. doi: 10.1038/sj.bjp.0705056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Advenier C, Emonds-Alt X. Tachykinin receptor antagonists and cough. Pulm Pharmacol. 1996;9:329–333. doi: 10.1006/pulp.1996.0043. [DOI] [PubMed] [Google Scholar]
- Ahlstedt S, Alving K, Hesselmar B, Olaisson E. Enhancement of the bronchial reactivity in immunized rats by neonatal treatment with capsaicin. Int Arch Allergy Appl Immunol. 1986;80:262–266. doi: 10.1159/000234063. [DOI] [PubMed] [Google Scholar]
- Aikawa T, Sekizawa K, Itabashi S, Sasaki H, Takishima T. Inhibitory actions of prostaglandin E1 on non-adrenergic non-cholinergic contraction in guinea-pig bronchi. Br J Pharmacol. 1990;101(1):13–14. doi: 10.1111/j.1476-5381.1990.tb12080.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aizawa H, Matsuzaki Y, Ishibashi M, Domae M, Hirose T, Shigematsu N, Tanaka K. A possible role of a nonadrenergic inhibitory nervous system in airway hyperreactivity. Respir Physiol. 1982;50:187–196. doi: 10.1016/0034-5687(82)90017-2. [DOI] [PubMed] [Google Scholar]
- Aizawa H, Tanaka H, Sakai J, Takata S, Hara N, Ito Y. L-NAME-sensitive and -insensitive nonadrenergic noncholinergic relaxation of cat airway in vivo and in vitro. Eur Respir J. 1997;10:314–321. doi: 10.1183/09031936.97.10020314. [DOI] [PubMed] [Google Scholar]
- Aizawa H, Takata S, Inoue H, Matsumoto K, Koto H, Hara N. Role of nitric oxide released from iNANC neurons in airway responsiveness in cats. Eur Respir J. 1999;13(4):775–780. doi: 10.1034/j.1399-3003.1999.13d13.x. [DOI] [PubMed] [Google Scholar]
- An SS, Bai TR, Bates JH, Black JL, Brown RH, Brusasco V, Chitano P, Deng L, Dowell M, Eidelman DH, Fabry B, Fairbank NJ, Ford LE, Fredberg JJ, Gerthoffer WT, Gilbert SH, Gosens R, Gunst SJ, Halayko AJ, Ingram RH, Irvin CG, James AL, Janssen LJ, King GG, Knight DA, Lauzon AM, Lakser OJ, Ludwig MS, Lutchen KR, Maksym GN, Martin JG, Mauad T, McParl BE, Mijailovich SM, Mitchell HW, Mitchell RW, Mitzner W, Murphy TM, Pare PD, Pellegrino R, Sanderson MJ, Schellenberg RR, Seow CY, Silveira PS, Smith PG, Solway J, Stephens NL, Sterk PJ, Stewart AG, Tang DD, Tepper RS, Tran T, Wang L. Airway smooth muscle dynamics: a common pathway of airway obstruction in asthma. Eur Respir J. 2007;29:834–860. doi: 10.1183/09031936.00112606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andresen JH, Saugstad OD. Effects of nicotine infusion on striatal glutamate and cortical non-protein-bound iron in hypoxic newborn piglets. Neonatology. 2008;94:284–292. doi: 10.1159/000151648. [DOI] [PubMed] [Google Scholar]
- Armstrong DJ, Luck JC. A comparative study of irritant and type J receptors in the cat. Respir Physiol. 1974;21:47–60. doi: 10.1016/0034-5687(74)90006-1. [DOI] [PubMed] [Google Scholar]
- Avital A, Springer C, Bar-Yishay E, Godfrey S. Adenosine, methacholine, and exercise challenges in children with asthma or paediatric chronic obstructive pulmonary disease. Thorax. 1995;50:511–516. doi: 10.1136/thx.50.5.511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aylwin ML, Horowitz JM, Bonham AC. NMDA receptors contribute to primary visceral afferent transmission in the nucleus of the solitary tract. J Neurophysiol. 1997;77:2539–2548. doi: 10.1152/jn.1997.77.5.2539. [DOI] [PubMed] [Google Scholar]
- Bachoo M, Polosa C. Properties of the inspiration-related activity of sympathetic preganglionic neurones of the cervical trunk in the cat. J Physiol. 1987;385:545–564. doi: 10.1113/jphysiol.1987.sp016507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bai TR, Macklem PT, Martin JG. The effects of parasympathectomy on serotonin-induced bronchoconstriction in the cat. Am Rev Respir Dis. 1986;133:110–115. doi: 10.1164/arrd.1986.133.1.110. [DOI] [PubMed] [Google Scholar]
- Baluk P, Nadel JA, McDonald DM. Substance P-immunoreactive sensory axons in the rat respiratory tract: a quantitative study of their distribution and role in neurogenic inflammation. J Comp Neurol. 1992;319:586–598. doi: 10.1002/cne.903190408. [DOI] [PubMed] [Google Scholar]
- Barman SM, Gebber GL. Basis for synchronization of sympathetic and phrenic nerve discharges. Am J Physiol. 1976;231:1601–1607. doi: 10.1152/ajplegacy.1976.231.5.1601. [DOI] [PubMed] [Google Scholar]
- Barnes PJ. Asthma as an axon reflex. Lancet. 1986;1(8475):242–245. doi: 10.1016/s0140-6736(86)90777-4. [DOI] [PubMed] [Google Scholar]
- Barnes PJ. Neurogenic inflammation in the airways. Respir Physiol. 2001;125:145–154. doi: 10.1016/s0034-5687(00)00210-3. [DOI] [PubMed] [Google Scholar]
- Barnes PJ, Adcock IM. NF-kappa B: a pivotal role in asthma and a new target for therapy. Trends Pharmacol Sci. 1997;18:46–50. doi: 10.1016/s0165-6147(97)89796-9. [DOI] [PubMed] [Google Scholar]
- Beier KC, Kallinich T, Hamelmann E. T-cell co-stimulatory molecules: novel targets for the treatment of allergic airway disease. Eur Respir J. 2007;30:383–390. doi: 10.1183/09031936.00094406. [DOI] [PubMed] [Google Scholar]
- Belvisi MG, Chung KF, Jackson DM, Barnes PJ. Opioid modulation of non-cholinergic neural bronchoconstriction in guinea-pig in vivo. Br J Pharmacol. 1988;95:413–418. doi: 10.1111/j.1476-5381.1988.tb11661.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belvisi MG, Ichinose M, Barnes PJ. Modulation of non-adrenergic, non-cholinergic neural bronchoconstriction in guinea-pig airways via GABAB-receptors. Br J Pharmacol. 1989;97:1225–1231. doi: 10.1111/j.1476-5381.1989.tb12582.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergren DR. Sensory receptor activation by mediators of defense reflexes in guinea-pig lungs. Respir Physiol. 1997;108:195–204. doi: 10.1016/s0034-5687(97)00030-3. [DOI] [PubMed] [Google Scholar]
- Bergren DR. Prostaglandin involvement in lung C-fiber activation by substance P in guinea pigs. J Appl Physiol. 2006;100(6):1918–1927. doi: 10.1152/japplphysiol.01276.2005. [DOI] [PubMed] [Google Scholar]
- Berman AR, Togias AG, Skloot G, Proud D. Allergen-induced hyperresponsiveness to bradykinin is more pronounced than that to methacholine. J Appl Physiol. 1995;78:1844–1852. doi: 10.1152/jappl.1995.78.5.1844. [DOI] [PubMed] [Google Scholar]
- Bhatia M, Zhi L, Zhang H, Ng SW, Moore PK. Role of substance P in hydrogen sulfide-induced pulmonary inflammation in mice. Am J Physiol Lung Cell Mol Physiol. 2006;291:L896–L904. doi: 10.1152/ajplung.00053.2006. [DOI] [PubMed] [Google Scholar]
- Binshtok AM, Bean BP, Woolf CJ. Inhibition of nociceptors by TRPV1-mediated entry of impermeant sodium channel blockers. Nature. 2007;449:607–610. doi: 10.1038/nature06191. [DOI] [PubMed] [Google Scholar]
- Birrell MA, Crispino N, Hele DJ, Patel HJ, Yacoub MH, Barnes PJ, Belvisi MG. Effect of dopamine receptor agonists on sensory nerve activity: possible therapeutic targets for the treatment of asthma and COPD. Br J Pharmacol. 2002;136:620–628. doi: 10.1038/sj.bjp.0704758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolser DC, Aziz SM, Chapman RW. Ruthenium red decreases capsaicin and citric acid-induced cough in guinea pigs. Neurosci Lett. 1991;126:131–133. doi: 10.1016/0304-3940(91)90536-3. [DOI] [PubMed] [Google Scholar]
- Bolser DC, DeGennaro FC, O'Reilly S, McLeod RL, Hey JA. Central antitussive activity of the NK1 and NK2 tachykinin receptor antagonists, CP-99,994 and SR 48968, in the guinea-pig and cat. Br J Pharmacol. 1997;121:165–170. doi: 10.1038/sj.bjp.0701111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bond RA, Spina D, Parra S, Page CP. Getting to the heart of asthma: can “beta blockers” be useful to treat asthma? Pharmacol Ther. 2007;115:360–374. doi: 10.1016/j.pharmthera.2007.04.009. [DOI] [PubMed] [Google Scholar]
- Bonham AC, Joad JP. Neurones in commissural nucleus tractus solitarii required for full expression of the pulmonary C fibre reflex in rat. J Physiol. 1991;441:95–112. doi: 10.1113/jphysiol.1991.sp018740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonham AC, McCrimmon DR. Neurones in a discrete region of the nucleus tractus solitarius are required for the Breuer-Hering reflex in rat. J Physiol. 1990;427:261–280. doi: 10.1113/jphysiol.1990.sp018171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonham AC, Coles SK, McCrimmon DR. Pulmonary stretch receptor afferents activate excitatory amino acid receptors in the nucleus tractus solitarii in rats. J Physiol. 1993;464:725–745. doi: 10.1113/jphysiol.1993.sp019660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonham AC, Kott KS, Ravi K, Kappagoda CT, Joad JP. Substance P contributes to rapidly adapting receptor responses to pulmonary venous congestion in rabbits. J Physiol. 1996;493(Pt 1):229–238. doi: 10.1113/jphysiol.1996.sp021378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bootle DJ, Adcock JJ, Ramage AG. Involvement of central 5-HT1A receptors in the reflex activation of pulmonary vagal motoneurones by inhaled capsaicin in anaesthetized cats. Br J Pharmacol. 1996;117:724–728. doi: 10.1111/j.1476-5381.1996.tb15250.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boushey HA, Richardson PS, Widdicombe JG, Wise JC. The response of laryngeal afferent fibres to mechanical and chemical stimuli. J Physiol Lond. 1974;240:153–175. doi: 10.1113/jphysiol.1974.sp010605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bramley AM, Samhoun MN, Piper PJ. The role of the epithelium in modulating the responses of guinea-pig trachea induced by bradykinin in vitro. Br J Pharmacol. 1990;99:762–766. doi: 10.1111/j.1476-5381.1990.tb13003.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burki NK, Dale WJ, Lee LY. Intravenous adenosine and dyspnea in humans. J Appl Physiol. 2005;98:180–185. doi: 10.1152/japplphysiol.00913.2004. [DOI] [PubMed] [Google Scholar]
- Canning BJ. Reflex regulation of airway smooth muscle tone. J Appl Physiol. 2006;101:971–985. doi: 10.1152/japplphysiol.00313.2006. [DOI] [PubMed] [Google Scholar]
- Canning BJ. Encoding of the cough reflex. Pulm Pharmacol Ther. 2007;20:396–401. doi: 10.1016/j.pupt.2006.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canning BJ, Chou YL (2009) Cough sensors. I. Physiological and pharmacological properties of the afferent nerves regulating cough. Handb Exp Pharmacol:23–47 [DOI] [PubMed]
- Canning BJ, Undem BJ. Relaxant innervation of the guinea-pig trachealis: demonstration of capsaicin-sensitive and -insensitive vagal pathways. J Physiol. 1993;460:719–739. doi: 10.1113/jphysiol.1993.sp019496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canning BJ, Reynolds SM, Mazzone SB. Multiple mechanisms of reflex bronchospasm in guinea pigs. J Appl Physiol. 2001;91:2642–2653. doi: 10.1152/jappl.2001.91.6.2642. [DOI] [PubMed] [Google Scholar]
- Canning BJ, Mazzone SB, Meeker SN, Mori N, Reynolds SM, Undem BJ. Identification of the tracheal and laryngeal afferent neurones mediating cough in anaesthetized guinea-pigs. J Physiol. 2004;557:543–558. doi: 10.1113/jphysiol.2003.057885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canning BJ, Farmer DG, Mori N. Mechanistic studies of acid-evoked coughing in anesthetized guinea pigs. Am J Physiol Regul Integr Comp Physiol. 2006;291:R454–R463. doi: 10.1152/ajpregu.00862.2005. [DOI] [PubMed] [Google Scholar]
- Canning BJ, Mori N, Mazzone SB. Vagal afferent nerves regulating the cough reflex. Respir Physiol Neurobiol. 2006;152:223–242. doi: 10.1016/j.resp.2006.03.001. [DOI] [PubMed] [Google Scholar]
- Carr MJ, Undem BJ. Pharmacology of vagal afferent nerve activity in guinea pig airways. Pulm Pharmacol Ther. 2003;16:45–52. doi: 10.1016/S1094-5539(02)00179-7. [DOI] [PubMed] [Google Scholar]
- Carr MJ, Schechter NM, Undem BJ. Trypsin-induced, neurokinin-mediated contraction of guinea pig bronchus. Am J Respir Crit Care Med. 2000;162(5):1662–1667. doi: 10.1164/ajrccm.162.5.9912099. [DOI] [PubMed] [Google Scholar]
- Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389:816–824. doi: 10.1038/39807. [DOI] [PubMed] [Google Scholar]
- Chanez P, Springall D, Vignola AM, Moradoghi-Hattvani A, Polak JM, Godard P, Bousquet J. Bronchial mucosal immunoreactivity of sensory neuropeptides in severe airway diseases. Am J Respir Crit Care Med. 1998;158:985–990. doi: 10.1164/ajrccm.158.3.9608104. [DOI] [PubMed] [Google Scholar]
- Chesrown SE, Venugopalan CS, Gold WM, Drazen JM. In vivo demonstration of nonadrenergic inhibitory innervation of the guinea pig trachea. J Clin Invest. 1980;65:314–320. doi: 10.1172/JCI109674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chianca DA, Jr, Machado BH. Microinjection of NMDA antagonist into the NTS of conscious rats blocks the Bezold–Jarisch reflex. Brain Res. 1996;718:185–188. doi: 10.1016/0006-8993(96)00010-8. [DOI] [PubMed] [Google Scholar]
- Choi JY, Joo NS, Krouse ME, Wu JV, Robbins RC, Ianowski JP, Hanrahan JW, Wine JJ. Synergistic airway gland mucus secretion in response to vasoactive intestinal peptide and carbachol is lost in cystic fibrosis. J Clin Invest. 2007;117(10):3118–3127. doi: 10.1172/JCI31992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou YL, Scarupa MD, Mori N, Canning BJ. Differential effects of airway afferent nerve subtypes on cough and respiration in anesthetized guinea pigs. Am J Physiol Regul Integr Comp Physiol. 2008;295:R1572–R1584. doi: 10.1152/ajpregu.90382.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuaychoo B, Lee MG, Kollarik M, Undem BJ. Effect of 5-hydroxytryptamine on vagal C-fiber subtypes in guinea pig lungs. Pulm Pharmacol Ther. 2005;18:269–276. doi: 10.1016/j.pupt.2004.12.010. [DOI] [PubMed] [Google Scholar]
- Chuaychoo B, Lee MG, Kollarik M, Pullmann R, Jr, Undem BJ. Evidence for both adenosine A1 and A2A receptors activating single vagal sensory C-fibres in guinea pig lungs. J Physiol. 2006;575:481–490. doi: 10.1113/jphysiol.2006.109371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark N, Keeble J, Fernandes ES, Starr A, Liang L, Sugden D, de WP, Brain SD. The transient receptor potential vanilloid 1 (TRPV1) receptor protects against the onset of sepsis after endotoxin. FASEB J. 2007;21:3747–3755. doi: 10.1096/fj.06-7460com. [DOI] [PubMed] [Google Scholar]
- Clerici C, Macquin-Mavier I, Harf A. Nonadrenergic bronchodilation in adult and young guinea pigs. J Appl Physiol. 1989;67(5):1764–1769. doi: 10.1152/jappl.1989.67.5.1764. [DOI] [PubMed] [Google Scholar]
- Coburn RF, Tomita T. Evidence for nonadrenergic inhibitory nerves in the guinea pig trachealis muscle. Am J Physiol. 1973;224:1072–1080. doi: 10.1152/ajplegacy.1973.224.5.1072. [DOI] [PubMed] [Google Scholar]
- Cockcroft DW, Davis BE. Mechanisms of airway hyperresponsiveness. J Allergy Clin Immunol. 2006;118:551–559. doi: 10.1016/j.jaci.2006.07.012. [DOI] [PubMed] [Google Scholar]
- Coleridge HM, Coleridge JC. Impulse activity in afferent vagal C-fibres with endings in the intrapulmonary airways of dogs. Respir Physiol. 1977;29:125–142. doi: 10.1016/0034-5687(77)90086-x. [DOI] [PubMed] [Google Scholar]
- Coleridge JC, Coleridge HM. Afferent vagal C fibre innervation of the lungs and airways and its functional significance. Rev Physiol Biochem Pharmacol. 1984;99:1–110. doi: 10.1007/BFb0027715. [DOI] [PubMed] [Google Scholar]
- Coleridge HM, Coleridge JC, Banzett RB. II. Effect of CO2 on afferent vagal endings in the canine lung. Respir Physiol. 1978;34:135–151. doi: 10.1016/0034-5687(78)90053-1. [DOI] [PubMed] [Google Scholar]
- Coleridge JC, Coleridge HM, Roberts AM, Kaufman MP, Baker DG. Tracheal contraction and relaxation initiated by lung and somatic afferents in dogs. J Appl Physiol. 1982;52:984–990. doi: 10.1152/jappl.1982.52.4.984. [DOI] [PubMed] [Google Scholar]
- Cortright DN, Krause JE, Broom DC. TRP channels and pain. Biochim Biophys Acta. 2007;1772:978–988. doi: 10.1016/j.bbadis.2007.03.003. [DOI] [PubMed] [Google Scholar]
- Coyle AJ, Perretti F, Manzini S, Irvin CG. Cationic protein-induced sensory nerve activation: role of substance P in airway hyperresponsiveness and plasma protein extravasation. J Clin Invest. 1994;94:2301–2306. doi: 10.1172/JCI117594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daffonchio L, Hernandez A, Gallico L, Omini C. Airway hyperreactivity induced by active cigarette smoke exposure in guinea-pigs: possible role of sensory neuropeptides. Pulm Pharmacol. 1990;3:161–166. doi: 10.1016/0952-0600(90)90048-n. [DOI] [PubMed] [Google Scholar]
- Davies RO, Kubin L. Projection of pulmonary rapidly adapting receptors to the medulla of the cat: an antidromic mapping study. J Physiol. 1986;373:63–86. doi: 10.1113/jphysiol.1986.sp016035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies A, Dixon M, Callanan D, Huszczuk A, Widdicombe JG, Wise JC. Lung reflexes in rabbits during pulmonary stretch receptor block by sulphur dioxide. Respir Physiol. 1978;34:83–101. doi: 10.1016/0034-5687(78)90050-6. [DOI] [PubMed] [Google Scholar]
- Davies RO, Kubin L, Pack AI. Pulmonary stretch receptor relay neurones of the cat: location and contralateral medullary projections. J Physiol. 1987;383:571–585. doi: 10.1113/jphysiol.1987.sp016429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies SF, McQuaid KR, Iber C, McArthur CD, Path MJ, Beebe DS, Helseth HK. Extreme dyspnea from unilateral pulmonary venous obstruction. Demonstration of a vagal mechanism and relief by right vagotomy. Am Rev Respir Dis. 1987;136(1):184–188. doi: 10.1164/ajrccm/136.1.184. [DOI] [PubMed] [Google Scholar]
- Davis KD, Meyer RA, Turnquist JL, Filloon TG, Pappagallo M, Campbell JN. Cutaneous pretreatment with the capsaicin analog NE-21610 prevents the pain to a burn and subsequent hyperalgesia. Pain. 1995;62:373–378. doi: 10.1016/0304-3959(95)00007-F. [DOI] [PubMed] [Google Scholar]
- Deep V, Singh M, Ravi K. Role of vagal afferents in the reflex effects of capsaicin and lobeline in monkeys. Respir Physiol. 2001;125:155–168. doi: 10.1016/s0034-5687(00)00223-1. [DOI] [PubMed] [Google Scholar]
- Delpierre S, Grimaud C, Jammes Y, Mei N. Changes in activity of vagal bronchopulmonary C fibres by chemical and physical stimuli in the cat. J Physiol. 1981;316:61–74. doi: 10.1113/jphysiol.1981.sp013772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Proost I, Pintelon I, Brouns I, Timmermans JP, Adriaensen D. Selective visualisation of sensory receptors in the smooth muscle layer of ex-vivo airway whole-mounts by styryl pyridinium dyes. Cell Tissue Res. 2007;329:421–431. doi: 10.1007/s00441-007-0431-5. [DOI] [PubMed] [Google Scholar]
- De Swert KO, Joos GF. Extending the understanding of sensory neuropeptides. Eur J Pharmacol. 2006;533:171–181. doi: 10.1016/j.ejphar.2005.12.066. [DOI] [PubMed] [Google Scholar]
- Dey RD, Altemus JB, Zervos I, Hoffpauir J. Origin and colocalization of CGRP- and SP-reactive nerves in cat airway epithelium. J Appl Physiol. 1990;68:770–778. doi: 10.1152/jappl.1990.68.2.770. [DOI] [PubMed] [Google Scholar]
- Diamond L, O'Donnell M. A nonadrenergic vagal inhibitory pathway to feline airways. Science. 1980;208:185–188. doi: 10.1126/science.7361114. [DOI] [PubMed] [Google Scholar]
- Dicpinigaitis PV. Experimentally induced cough. Pulm Pharmacol Ther. 2007;20:319–24. doi: 10.1016/j.pupt.2006.10.003. [DOI] [PubMed] [Google Scholar]
- Dinh QT, Groneberg DA, Peiser C, Mingomataj E, Joachim RA, Witt C, Arck PC, Klapp BF, Fischer A. Substance P expression in TRPV1 and trkA-positive dorsal root ganglion neurons innervating the mouse lung. Respir Physiol Neurobiol. 2004;144(1):15–24. doi: 10.1016/j.resp.2004.08.001. [DOI] [PubMed] [Google Scholar]
- Dixon M, Jackson DM, Richards IM. The effects of histamine, acetylcholine and 5-hydroxytryptamine on lung mechanics and irritant receptors in the dog. J Physiol. 1979;287:393–403. doi: 10.1113/jphysiol.1979.sp012666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dixon M, Jackson DM, Richards IM. The action of sodium cromoglycate on ‘C' fibre endings in the dog lung. Br J Pharmacol. 1980;70:11–13. doi: 10.1111/j.1476-5381.1980.tb10898.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dybas JM, Andresen CJ, Schelegle ES, McCue RW, Callender NN, Jackson AC. Deep-breath frequency in bronchoconstricted monkeys (Macaca fascicularis) J Appl Physiol. 2006;100:786–791. doi: 10.1152/japplphysiol.01014.2004. [DOI] [PubMed] [Google Scholar]
- Ellis JL, Sham JS, Undem BJ. Tachykinin-independent effects of capsaicin on smooth muscle in human isolated bronchi. Am J Respir Crit Care Med. 1997;155:751–755. doi: 10.1164/ajrccm.155.2.9032224. [DOI] [PubMed] [Google Scholar]
- Ezure K, Otake K, Lipski J, She RB. Efferent projections of pulmonary rapidly adapting receptor relay neurons in the cat. Brain Res. 1991;564:268–278. doi: 10.1016/0006-8993(91)91463-b. [DOI] [PubMed] [Google Scholar]
- Ezure K, Tanaka I, Miyazaki M. Electrophysiological and pharmacological analysis of synaptic inputs to pulmonary rapidly adapting receptor relay neurons in the rat. Exp Brain Res. 1999;128:471–480. doi: 10.1007/s002210050870. [DOI] [PubMed] [Google Scholar]
- Ezure K, Tanaka I, Saito Y, Otake K. Axonal projections of pulmonary slowly adapting receptor relay neurons in the rat. J Comp Neurol. 2002;446:81–94. doi: 10.1002/cne.10185. [DOI] [PubMed] [Google Scholar]
- Fischer A, Forssmann WG, Undem BJ. Nociceptin-induced inhibition of tachykinergic neurotransmission in guinea pig bronchus. J Pharmacol Exp Ther. 1998;285:902–907. [PubMed] [Google Scholar]
- Fisher JT, Sant'Ambrogio G. Effects of inhaled CO2 on airway stretch receptors in the newborn dog. J Appl Physiol. 1982;53(6):1461–1465. doi: 10.1152/jappl.1982.53.6.1461. [DOI] [PubMed] [Google Scholar]
- Forsberg K, Karlsson JA, Theodorsson E, Lundberg JM, Persson CG. Cough and bronchoconstriction mediated by capsaicin-sensitive sensory neurons in the guinea-pig. Pulm Pharmacol. 1988;1:33–39. doi: 10.1016/0952-0600(88)90008-7. [DOI] [PubMed] [Google Scholar]
- Fox AJ, Barnes PJ, Urban L, Dray A. An in vitro study of the properties of single vagal afferents innervating guinea-pig airways. J Physiol. 1993;469:21–35. doi: 10.1113/jphysiol.1993.sp019802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox AJ, Barnes PJ, Dray A. Stimulation of guinea-pig tracheal afferent fibres by non-isosmotic and low-chloride stimuli and the effect of frusemide. J Physiol. 1995;482(Pt 1):179–187. doi: 10.1113/jphysiol.1995.sp020508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox AJ, Barnes PJ, Venkatesan P, Belvisi MG. Activation of large conductance potassium channels inhibits the afferent and efferent function of airway sensory nerves in the guinea pig. J Clin Invest. 1997;99:513–519. doi: 10.1172/JCI119187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gatti R, Andre E, Amadesi S, Dinh TQ, Fischer A, Bunnett NW, Harrison S, Geppetti P, Trevisani M. Protease-activated receptor-2 activation exaggerates TRPV1-mediated cough in guinea pigs. J Appl Physiol. 2006;101:506–511. doi: 10.1152/japplphysiol.01558.2005. [DOI] [PubMed] [Google Scholar]
- Gaylor JB. The intrinsic nervous mechanism of the human lung. Brain. 1934;57:143–160. [Google Scholar]
- Geppetti P, Trevisani M. Activation and sensitisation of the vanilloid receptor: role in gastrointestinal inflammation and function. Br J Pharmacol. 2004;141:1313–1320. doi: 10.1038/sj.bjp.0705768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gestreau C, Bianchi AL, Grelot L. Differential brainstem fos-like immunoreactivity after laryngeal-induced coughing and its reduction by codeine. J Neurosci. 1997;17:9340–9352. doi: 10.1523/JNEUROSCI.17-23-09340.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gil FR, Lauzon AM. Smooth muscle molecular mechanics in airway hyperresponsiveness and asthma. Can J Physiol Pharmacol. 2007;85:133–140. doi: 10.1139/y06-096. [DOI] [PubMed] [Google Scholar]
- Girard V, Yavo JC, Emonds-Alt X, Advenier C. The tachykinin NK2 receptor antagonist SR 48968 inhibits citric acid-induced airway hyperresponsiveness in guinea-pigs. Am J Respir Crit Care Med. 1996;153:1496–1502. doi: 10.1164/ajrccm.153.5.8630592. [DOI] [PubMed] [Google Scholar]
- Giuliani S, Amann R, Papini AM, Maggi CA, Meli A. Modulatory action of galanin on responses due to antidromic activation of peripheral terminals of capsaicin-sensitive sensory nerves. Eur J Pharmacol. 1989;163:91–96. doi: 10.1016/0014-2999(89)90399-3. [DOI] [PubMed] [Google Scholar]
- Glaser ST, Kaczocha M, Deutsch DG. Anandamide transport: a critical review. Life Sci. 2005;77:1584–1604. doi: 10.1016/j.lfs.2005.05.007. [DOI] [PubMed] [Google Scholar]
- Green JF, Schmidt ND, Schultz HD, Roberts AM, Coleridge HM, Coleridge JC. Pulmonary C-fibers evoke both apnea and tachypnea of pulmonary chemoreflex. J Appl Physiol. 1984;57(2):562–567. doi: 10.1152/jappl.1984.57.2.562. [DOI] [PubMed] [Google Scholar]
- Green JF, Schertel ER, Coleridge HM, Coleridge JC. Effect of pulmonary arterial PCO2 on slowly adapting pulmonary stretch receptors. J Appl Physiol. 1986;60:2048–2055. doi: 10.1152/jappl.1986.60.6.2048. [DOI] [PubMed] [Google Scholar]
- Groneberg DA, Niimi A, Dinh QT, Cosio B, Hew M, Fischer A, Chung KF. Increased expression of transient receptor potential vanilloid-1 in airway nerves of chronic cough. Am J Respir Crit Care Med. 2004;170:1276–1280. doi: 10.1164/rccm.200402-174OC. [DOI] [PubMed] [Google Scholar]
- Gross NJ, Co E, Skorodin MS. Cholinergic bronchomotor tone in COPD. Estimates of its amount in comparison with that in normal subjects. Chest. 1989;96:984–987. doi: 10.1378/chest.96.5.984. [DOI] [PubMed] [Google Scholar]
- Grundstrom N, Andersson RG, Wikberg JE. Prejunctional alpha 2 adrenoceptors inhibit contraction of tracheal smooth muscle by inhibiting cholinergic neurotransmission. Life Sci. 1981;28:2981–2986. doi: 10.1016/0024-3205(81)90275-7. [DOI] [PubMed] [Google Scholar]
- Grundstrom N, Andersson RG, Wikberg JE. Inhibition of the excitatory non-adrenergic, non-cholinergic neurotransmission in the guinea pig tracheo-bronchial tree mediated by alpha 2-adrenoceptors. Acta Pharmacol Toxicol (Copenh) 1984;54:8–14. doi: 10.1111/j.1600-0773.1984.tb01889.x. [DOI] [PubMed] [Google Scholar]
- Gu Q, Lee LY. Characterization of acid signaling in rat vagal pulmonary sensory neurons. Am J Physiol Lung Cell Mol Physiol. 2006;291:L58–L65. doi: 10.1152/ajplung.00517.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu Q, Lin YS, Lee LY. Epinephrine enhances the sensitivity of rat vagal chemosensitive neurons: role of beta3-adrenoceptor. J Appl Physiol. 2007;102:1545–1555. doi: 10.1152/japplphysiol.01010.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu Q, Ni D, Lee LY. Expression of neuronal nicotinic acetylcholine receptors in rat vagal pulmonary sensory neurons. Respir Physiol Neurobiol. 2008;161:87–91. doi: 10.1016/j.resp.2007.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guardiola J, Proctor M, Li H, Punnakkattu R, Lin S, Yu J. Airway mechanoreceptor deactivation. J Appl Physiol. 2007;103:600–607. doi: 10.1152/japplphysiol.01286.2006. [DOI] [PubMed] [Google Scholar]
- Gunthorpe MJ, Benham CD, Randall A, Davis JB. The diversity in the vanilloid (TRPV) receptor family of ion channels. Trends Pharmacol Sci. 2002;23:183–191. doi: 10.1016/s0165-6147(02)01999-5. [DOI] [PubMed] [Google Scholar]
- Guo A, Vulchanova L, Wang J, Li X, Elde R. Immunocytochemical localization of the vanilloid receptor 1 (VR1): relationship to neuropeptides, the P2X3 purinoceptor and IB4 binding sites. Eur J Neurosci. 1999;11:946–958. doi: 10.1046/j.1460-9568.1999.00503.x. [DOI] [PubMed] [Google Scholar]
- Habler HJ, Janig W, Michaelis M. Respiratory modulation in the activity of sympathetic neurones. Prog Neurobiol. 1994;43:567–606. doi: 10.1016/0301-0082(94)90053-1. [DOI] [PubMed] [Google Scholar]
- Hammad H, Lambrecht BN. Lung dendritic cell migration. Adv Immunol. 2007;93:265–278. doi: 10.1016/S0065-2776(06)93007-7. [DOI] [PubMed] [Google Scholar]
- Hanacek J, Davies A, Widdicombe JG. Influence of lung stretch receptors on the cough reflex in rabbits. Respiration. 1984;45:161–168. doi: 10.1159/000194614. [DOI] [PubMed] [Google Scholar]
- Harrison S, Reddy S, Page CP, Spina D. Stimulation of airway sensory nerves by cyclosporin A and FK506 in guinea-pig isolated bronchus. Br J Pharmacol. 1998;125:1405–1412. doi: 10.1038/sj.bjp.0702198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haxhiu MA, Chavez JC, Pichiule P, Erokwu B, Dreshaj IA. The excitatory amino acid glutamate mediates reflexly increased tracheal blood flow and airway submucosal gland secretion. Brain Res. 2000;883:77–86. doi: 10.1016/s0006-8993(00)02890-0. [DOI] [PubMed] [Google Scholar]
- Helyes Z, Elekes K, Nemeth J, Pozsgai G, Sandor K, Kereskai L, Borzsei R, Pinter E, Szabo A, Szolcsanyi J. Role of transient receptor potential vanilloid 1 receptors in endotoxin-induced airway inflammation in the mouse. Am J Physiol Lung Cell Mol Physiol. 2007;292:L1173–L1181. doi: 10.1152/ajplung.00406.2006. [DOI] [PubMed] [Google Scholar]
- Hempleman SC, Rodriguez TA, Bhagat YA, Begay RS. Benzolamide, acetazolamide, and signal transduction in avian intrapulmonary chemoreceptors. Am J Physiol Regul Integr Comp Physiol. 2000;279:R1988–R1995. doi: 10.1152/ajpregu.2000.279.6.R1988. [DOI] [PubMed] [Google Scholar]
- Herd CM, Gozzard N, Page CP. Capsaicin pretreatment prevents the development of antigen induced airway hyperresponsiveness in neonatally immunized rabbits. Eur J Pharmacol. 1995;282:111–119. doi: 10.1016/0014-2999(95)00291-r. [DOI] [PubMed] [Google Scholar]
- Hislop AA, Wharton J, Allen KM, Polak JM, Haworth SG. Immunohistochemical localization of peptide-containing nerves in human airways: age-related changes. Am J Respir Cell Mol Biol. 1990;3:191–198. [PubMed] [Google Scholar]
- Ho CY, Gu Q, Hong JL, Lee LY. Prostaglandin E(2) enhances chemical and mechanical sensitivities of pulmonary C fibers in the rat. Am J Respir Crit Care Med. 2000;162:528–533. doi: 10.1164/ajrccm.162.2.9910059. [DOI] [PubMed] [Google Scholar]
- Ho CY, Gu Q, Lin YS, Lee LY. Sensitivity of vagal afferent endings to chemical irritants in the rat lung. Respir Physiol. 2001;127:113–124. doi: 10.1016/s0034-5687(01)00241-9. [DOI] [PubMed] [Google Scholar]
- Holgate ST. The epithelium takes centre stage in asthma and atopic dermatitis. Trends Immunol. 2007;28:248–251. doi: 10.1016/j.it.2007.04.007. [DOI] [PubMed] [Google Scholar]
- Hong JL, Ho CY, Kwong K, Lee LY. Activation of pulmonary C fibres by adenosine in anaesthetized rats: role of adenosine A1 receptors. J Physiol. 1998;508(Pt 1):109–118. doi: 10.1111/j.1469-7793.1998.109br.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hornby PJ. Central neurocircuitry associated with emesis. Am J Med. 2001;111(Suppl 8A):106S–112S. doi: 10.1016/s0002-9343(01)00849-x. [DOI] [PubMed] [Google Scholar]
- House A, Celly C, Skeans S, Lamca J, Egan RW, Hey JA, Chapman RW. Cough reflex in allergic dogs. Eur J Pharmacol. 2004;492:251–258. doi: 10.1016/j.ejphar.2004.03.053. [DOI] [PubMed] [Google Scholar]
- Howarth PH, Springall DR, Redington AE, Djukanovic R, Holgate ST, Polak JM. Neuropeptide-containing nerves in endobronchial biopsies from asthmatic and nonasthmatic subjects. Am J Respir Cell Mol Biol. 1995;13:288–296. doi: 10.1165/ajrcmb.13.3.7654385. [DOI] [PubMed] [Google Scholar]
- Hua XY, Yaksh TL. Release of calcitonin gene-related peptide and tachykinins from the rat trachea. Peptides. 1992;13(1):113–120. doi: 10.1016/0196-9781(92)90148-v. [DOI] [PubMed] [Google Scholar]
- Hua XY, Theodorsson-Norheim E, Brodin E, Lundberg JM, Hokfelt T. Multiple tachykinins (neurokinin A, neuropeptide K and substance P) in capsaicin-sensitive sensory neurons in the guinea-pig. Regul Pept. 1985;13:1–19. doi: 10.1016/0167-0115(85)90082-5. [DOI] [PubMed] [Google Scholar]
- Huang WX, Yu Q, Cohen MI. Fast (3 Hz and 10 Hz) and slow (respiratory) rhythms in cervical sympathetic nerve and unit discharges of the cat. J Physiol. 2000;523(Pt 2):459–477. doi: 10.1111/j.1469-7793.2000.00459.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunter DD, Undem BJ. Identification and substance P content of vagal afferent neurons innervating the epithelium of the guinea pig trachea. Am J Respir Crit Care Med. 1999;159:1943–1948. doi: 10.1164/ajrccm.159.6.9808078. [DOI] [PubMed] [Google Scholar]
- Hutter MM, Wick EC, Day AL, Maa J, Zerega EC, Richmond AC, Jordan TH, Grady EF, Mulvihill SJ, Bunnett NW, Kirkwood KS. Transient receptor potential vanilloid (TRPV-1) promotes neurogenic inflammation in the pancreas via activation of the neurokinin-1 receptor (NK-1R) Pancreas. 2005;30:260–265. doi: 10.1097/01.mpa.0000153616.63384.24. [DOI] [PubMed] [Google Scholar]
- Ichinose M, Inoue H, Miura M, Yafuso N, Nogami H, Takishima T. Possible sensory receptor of nonadrenergic inhibitory nervous system. J Appl Physiol. 1987;63:923–929. doi: 10.1152/jappl.1987.63.3.923. [DOI] [PubMed] [Google Scholar]
- Ichinose M, Inoue H, Miura M, Takishima T. Nonadrenergic bronchodilation in normal subjects. Am Rev Respir Dis. 1988;138:31–34. doi: 10.1164/ajrccm/138.1.31. [DOI] [PubMed] [Google Scholar]
- Inoue H, Ichinose M, Miura M, Katsumata U, Takishima T. Sensory receptors and reflex pathways of nonadrenergic inhibitory nervous system in feline airways. Am Rev Respir Dis. 1989;139(5):1175–1178. doi: 10.1164/ajrccm/139.5.1175. [DOI] [PubMed] [Google Scholar]
- Inoue H, Aizawa H, Miyazaki N, Ikeda T, Shigematsu N. Possible roles of the peripheral vagal nerve in histamine-induced bronchoconstriction in guinea-pigs. Eur Respir J. 1991;4:860–866. [PubMed] [Google Scholar]
- Irvin CG, Martin RR, Macklem PT. Nonpurinergic nature and efficacy of nonadrenergic bronchodilation. J Appl Physiol. 1982;52:562–569. doi: 10.1152/jappl.1982.52.3.562. [DOI] [PubMed] [Google Scholar]
- Jackson DM, Norris AA, Eady RP. Nedocromil sodium and sensory nerves in the dog lung. Pulm Pharmacol. 1989;2:179–184. doi: 10.1016/0952-0600(89)90018-5. [DOI] [PubMed] [Google Scholar]
- Jacobsen EA, Ochkur SI, Lee NA, Lee JJ. Eosinophils and asthma. Curr Allergy Asthma Rep. 2007;7:18–26. doi: 10.1007/s11882-007-0026-y. [DOI] [PubMed] [Google Scholar]
- Jakus J, Poliacek I, Halasova E, Murin P, Knocikova J, Tomori Z, Bolser DC. Brainstem circuitry of tracheal-bronchial cough: c-fos study in anesthetized cats. Respir Physiol Neurobiol. 2008;160:289–300. doi: 10.1016/j.resp.2007.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jammes Y, Mei N. Assessment of the pulmonary origin of bronchoconstrictor vagal tone. J Physiol. 1979;291:305–316. doi: 10.1113/jphysiol.1979.sp012814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janig W, Habler HJ. Neurophysiological analysis of target-related sympathetic pathways--from animal to human: similarities and differences. Acta Physiol Scand. 2003;177:255–274. doi: 10.1046/j.1365-201X.2003.01088.x. [DOI] [PubMed] [Google Scholar]
- Jarreau PH, D'Ortho MP, Boyer V, Harf A, quin Mavier I. Effects of capsaicin on the airway responses to inhaled endotoxin in the guinea pig. Am J Respir Crit Care Med. 1994;149:128–133. doi: 10.1164/ajrccm.149.1.8111569. [DOI] [PubMed] [Google Scholar]
- Jia Y, Lee LY. Role of TRPV receptors in respiratory diseases. Biochim Biophys Acta. 2007;1772:915–927. doi: 10.1016/j.bbadis.2007.01.013. [DOI] [PubMed] [Google Scholar]
- Jia Y, McLeod RL, Wang X, Parra LE, Egan RW, Hey JA. Anandamide induces cough in conscious guinea-pigs through VR1 receptors. Br J Pharmacol. 2002;137:831–836. doi: 10.1038/sj.bjp.0704950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jimba M, Skornik WA, Killingsworth CR, Long NC, Brain JD, Shore SA. Role of C fibers in physiological responses to ozone in rats. J Appl Physiol. 1995;78:1757–1763. doi: 10.1152/jappl.1995.78.5.1757. [DOI] [PubMed] [Google Scholar]
- Jonzon A, Pisarri TE, Coleridge JC, Coleridge HM. Rapidly adapting receptor activity in dogs is inversely related to lung compliance. J Appl Physiol. 1986;61:1980–1987. doi: 10.1152/jappl.1986.61.5.1980. [DOI] [PubMed] [Google Scholar]
- Julius D, Basbaum AI. Molecular mechanisms of nociception. Nature. 2001;413:203–210. doi: 10.1038/35093019. [DOI] [PubMed] [Google Scholar]
- Kalia M, Richter D. Morphology of physiologically identified slowly adapting lung stretch receptor afferents stained with intra-axonal horseradish peroxidase in the nucleus of the tractus solitarius of the cat. I. A light microscopic analysis. J Comp Neurol. 1985;241:503–520. doi: 10.1002/cne.902410409. [DOI] [PubMed] [Google Scholar]
- Kalia M, Richter D. Rapidly adapting pulmonary receptor afferents: II. Fine structure and synaptic organization of central terminal processes in the nucleus of the tractus solitarius. J Comp Neurol. 1988;274:574–594. doi: 10.1002/cne.902740407. [DOI] [PubMed] [Google Scholar]
- Kallinich T, Beier KC, Wahn U, Stock P, Hamelmann E. T-cell co-stimulatory molecules: their role in allergic immune reactions. Eur Respir J. 2007;29:1246–1255. doi: 10.1183/09031936.00094306. [DOI] [PubMed] [Google Scholar]
- Kamikawa Y, Shimo Y. Adenosine selectively inhibits noncholinergic transmission in guinea pig bronchi. J Appl Physiol. 1989;66:2084–2091. doi: 10.1152/jappl.1989.66.5.2084. [DOI] [PubMed] [Google Scholar]
- Karius DR, Ling L, Speck DF. Nucleus tractus solitarius and excitatory amino acids in afferent-evoked inspiratory termination. J Appl Physiol. 1994;76:1293–1301. doi: 10.1152/jappl.1994.76.3.1293. [DOI] [PubMed] [Google Scholar]
- Karla W, Shams H, Orr JA, Scheid P. Effects of the thromboxane A2 mimetic, U46,619, on pulmonary vagal afferents in the cat. Respir Physiol. 1992;87:383–396. doi: 10.1016/0034-5687(92)90019-s. [DOI] [PubMed] [Google Scholar]
- Karlsson JA, Fuller RW. Pharmacological regulation of the cough reflex--from experimental models to antitussive effects in Man. Pulm Pharmacol Ther. 1999;12:215–228. doi: 10.1006/pupt.1999.0207. [DOI] [PubMed] [Google Scholar]
- Karlsson JA, Sant'Ambrogio FB, Forsberg K, Palecek F, Mathew OP, Sant'Ambrogio G. Respiratory and cardiovascular effects of inhaled and intravenous bradykinin, PGE2, and PGF2 alpha in dogs. J Appl Physiol. 1993;74:2380–2386. doi: 10.1152/jappl.1993.74.5.2380. [DOI] [PubMed] [Google Scholar]
- Kaufman MP, Coleridge HM, Coleridge JC, Baker DG. Bradykinin stimulates afferent vagal C-fibers in intrapulmonary airways of dogs. J Appl Physiol. 1980;48:511–517. doi: 10.1152/jappl.1980.48.3.511. [DOI] [PubMed] [Google Scholar]
- Keeble J, Russell F, Curtis B, Starr A, Pinter E, Brain SD. Involvement of transient receptor potential vanilloid 1 in the vascular and hyperalgesic components of joint inflammation. Arthritis Rheum. 2005;52:3248–3256. doi: 10.1002/art.21297. [DOI] [PubMed] [Google Scholar]
- Kesler BS, Canning BJ. Regulation of baseline cholinergic tone in guinea-pig airway smooth muscle. J Physiol. 1999;518(Pt 3):843–855. doi: 10.1111/j.1469-7793.1999.0843p.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kesler BS, Mazzone SB, Canning BJ. Nitric oxide-dependent modulation of smooth-muscle tone by airway parasympathetic nerves. Am J Respir Crit Care Med. 2002;165:481–488. doi: 10.1164/ajrccm.165.4.2004005. [DOI] [PubMed] [Google Scholar]
- Ketchell RI, Jensen MW, Lumley P, Wright AM, Allenby MI, O'Connor BJ. Rapid effect of inhaled fluticasone propionate on airway responsiveness to adenosine 5′-monophosphate in mild asthma. J Allergy Clin Immunol. 2002;110:603–606. doi: 10.1067/mai.2002.128486. [DOI] [PubMed] [Google Scholar]
- Kihara N, de la Fuente SG, Fujino K, Takahashi T, Pappas TN, Mantyh CR. Vanilloid receptor-1 containing primary sensory neurones mediate dextran sulphate sodium induced colitis in rats. Gut. 2003;52:713–719. doi: 10.1136/gut.52.5.713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knowlton GC, Larrabee MG. A unitary analysis of pulmonary volume receptors. Am J Physiol Lung Cell Mol Physiol. 1946;147:100–114. doi: 10.1152/ajplegacy.1946.147.1.100. [DOI] [PubMed] [Google Scholar]
- Kollarik M, Undem BJ. Mechanisms of acid-induced activation of airway afferent nerve fibres in guinea-pig. J Physiol. 2002;543:591–600. doi: 10.1113/jphysiol.2002.022848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kollarik M, Undem BJ. Activation of bronchopulmonary vagal afferent nerves with bradykinin, acid and vanilloid receptor agonists in wild-type and TRPV1−/− mice. J Physiol. 2004;555:115–123. doi: 10.1113/jphysiol.2003.054890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kollarik M, Dinh QT, Fischer A, Undem BJ. Capsaicin-sensitive and -insensitive vagal bronchopulmonary C-fibres in the mouse. J Physiol. 2003;551:869–879. doi: 10.1113/jphysiol.2003.042028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kollarik M, Ru F, Undem BJ. Acid-sensitive vagal sensory pathways and cough. Pulm Pharmacol Ther. 2007;20:402–411. doi: 10.1016/j.pupt.2006.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koto H, Aizawa H, Takata S, Inoue H, Hara N. An important role of tachykinins in ozone-induced airway hyperresponsiveness. Am J Respir Crit Care Med. 1995;151:1763–1769. doi: 10.1164/ajrccm.151.6.7767518. [DOI] [PubMed] [Google Scholar]
- Kubin L, Kimura H, Davies RO. The medullary projections of afferent bronchopulmonary C fibres in the cat as shown by antidromic mapping. J Physiol. 1991;435:207–228. doi: 10.1113/jphysiol.1991.sp018506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubin L, Alheid GF, Zuperku EJ, McCrimmon DR. Central pathways of pulmonary and lower airway vagal afferents. J Appl Physiol. 2006;101:618–627. doi: 10.1152/japplphysiol.00252.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kummer W, Fischer A, Kurkowski R, Heym C. The sensory and sympathetic innervation of guinea-pig lung and trachea as studied by retrograde neuronal tracing and double-labelling immunohistochemistry. Neuroscience. 1992;49:715–737. doi: 10.1016/0306-4522(92)90239-x. [DOI] [PubMed] [Google Scholar]
- Kwong K, Hong JL, Morton RF, Lee LY. Role of pulmonary C fibres in adenosine-induced respiratory inhibition in anesthetized rats. J Appl Physiol. 1998;84:417–424. doi: 10.1152/jappl.1998.84.2.417. [DOI] [PubMed] [Google Scholar]
- Kwong K, Carr MJ, Gibbard A, Savage TJ, Singh K, Jing J, Meeker S, Undem BJ. Voltage-gated sodium channels in nociceptive versus non-nociceptive nodose vagal sensory neurons innervating guinea pig lungs. J Physiol. 2008;586:1321–1336. doi: 10.1113/jphysiol.2007.146365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwong K, Kollarik M, Nassenstein C, Ru F, Undem BJ. P2X2 receptors differentiate placodal vs. neural crest C-fiber phenotypes innervating guinea pig lungs and esophagus. Am J Physiol Lung Cell Mol Physiol. 2008;295:L858–L865. doi: 10.1152/ajplung.90360.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lacroix JS, Buvelot JM, Polla BS, Lundberg JM. Improvement of symptoms of non-allergic chronic rhinitis by local treatment with capsaicin. Clin Exp Allergy. 1991;21:595–600. doi: 10.1111/j.1365-2222.1991.tb00852.x. [DOI] [PubMed] [Google Scholar]
- Lai CJ, Ruan T, Kou YR. The involvement of hydroxyl radical and cyclooxygenase metabolites in the activation of lung vagal sensory receptors by circulatory endotoxin in rats. J Appl Physiol. 2005;98:620–628. doi: 10.1152/japplphysiol.00539.2004. [DOI] [PubMed] [Google Scholar]
- Laitinen LA, Laitinen A, Panula PA, Partanen M, Tervo K, Tervo T. Immunohistochemical demonstration of substance P in the lower respiratory tract of the rabbit and not of man. Thorax. 1983;38:531–536. doi: 10.1136/thx.38.7.531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lalloo UG, Fox AJ, Belvisi MG, Chung KF, Barnes PJ. Capsazepine inhibits cough induced by capsaicin and citric acid but not by hypertonic saline in guinea pigs. J Appl Physiol. 1995;79:1082–1087. doi: 10.1152/jappl.1995.79.4.1082. [DOI] [PubMed] [Google Scholar]
- Lama A, Delpierre S, Jammes Y. The effects of electrical stimulation of myelinated and non-myelinated vagal motor fibres on airway tone in the rabbit and the cat. Respir Physiol. 1988;74:265–274. doi: 10.1016/0034-5687(88)90035-7. [DOI] [PubMed] [Google Scholar]
- Lamb JP, Sparrow MP. Three-dimensional mapping of sensory innervation with substance p in porcine bronchial mucosa: comparison with human airways. Am J Respir Crit Care Med. 2002;166:1269–1281. doi: 10.1164/rccm.2112018. [DOI] [PubMed] [Google Scholar]
- Lammers JW, Minette P, McCusker MT, Chung KF, Barnes PJ. Capsaicin-induced bronchodilation in mild asthmatic subjects: possible role of nonadrenergic inhibitory system. J Appl Physiol. 1989;67(2):856–861. doi: 10.1152/jappl.1989.67.2.856. [DOI] [PubMed] [Google Scholar]
- Larsell O. Nerve termination in the lung of the rabbit. J Comp Neurol. 1921;33:105–131. [Google Scholar]
- Larsell O. The ganglia, plexuses and nerve-termination of the mammalian lung and pleura pulmonis. J Comp Neurol. 1922;35:97–132. [Google Scholar]
- Laude EA, Higgins KS, Morice AH. A comparative study of the effects of citric acid, capsaicin and resiniferatoxin on the cough challenge in guinea-pig and man. Pulm Pharmacol. 1993;6:171–175. doi: 10.1006/pulp.1993.1023. [DOI] [PubMed] [Google Scholar]
- Lee LY, Morton RF. Histamine enhances vagal pulmonary C-fiber responses to capsaicin and lung inflation. Respir Physiol. 1993;93:83–96. doi: 10.1016/0034-5687(93)90070-q. [DOI] [PubMed] [Google Scholar]
- Lee LY, Morton RF. Pulmonary chemoreflex sensitivity is enhanced by prostaglandin E2 in anesthetized rats. J Appl Physiol. 1995;79:1679–1686. doi: 10.1152/jappl.1995.79.5.1679. [DOI] [PubMed] [Google Scholar]
- Lee LY, Pisarri TE. Afferent properties and reflex functions of bronchopulmonary C-fibers. Respir Physiol. 2001;125:47–65. doi: 10.1016/s0034-5687(00)00204-8. [DOI] [PubMed] [Google Scholar]
- Lee MG, Weinreich D, Undem BJ. Effect of olvanil and anandamide on vagal C-fiber subtypes in guinea pig lung. Br J Pharmacol. 2005;146:596–603. doi: 10.1038/sj.bjp.0706339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung SY, Niimi A, Williams AS, Nath P, Blanc FX, Dinh QT, Chung KF. Inhibition of citric acid- and capsaicin-induced cough by novel TRPV-1 antagonist, V112220, in guinea-pig. Cough. 2007;3:10. doi: 10.1186/1745-9974-3-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liddle RA. The role of Transient Receptor Potential Vanilloid 1 (TRPV1) channels in pancreatitis. Biochim Biophys Acta. 2007;1772:869–878. doi: 10.1016/j.bbadis.2007.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ligresti A, Morera E, Van Der Stelt M, Monory K, Lutz B, Ortar G, Di Marzo V. Further evidence for the existence of a specific process for the membrane transport of anandamide. Biochem J. 2004;380:265–272. doi: 10.1042/BJ20031812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin YS, Lee LY. Stimulation of pulmonary vagal C-fibres by anandamide in anaesthetized rats: role of vanilloid type 1 receptors. J Physiol. 2002;539:947–955. doi: 10.1113/jphysiol.2001.013290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin RL, Gu Q, Lin YS, Lee LY. Stimulatory effect of CO2 on vagal bronchopulmonary C-fiber afferents during airway inflammation. J Appl Physiol. 2005;99:1704–1711. doi: 10.1152/japplphysiol.00532.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipski J, Ezure K, Wong She RB. Identification of neurons receiving input from pulmonary rapidly adapting receptors in the cat. J Physiol. 1991;443:55–77. doi: 10.1113/jphysiol.1991.sp018822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lloyd CM, Robinson DS. Allergen-induced airway remodelling. Eur Respir J. 2007;29:1020–1032. doi: 10.1183/09031936.00150305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loeffler BS, Arden WA, Fiscus RR, Lee LY. Involvement of tachykinins in endotoxin-induced airway hyperresponsiveness. Lung. 1997;175:253–263. doi: 10.1007/pl00007572. [DOI] [PubMed] [Google Scholar]
- Long NC, Martin JG, Pantano R, Shore SA. Airway hyperresponsiveness in a rat model of chronic bronchitis: role of C fibers. Am J Respir Crit Care Med. 1997;155:1222–1229. doi: 10.1164/ajrccm.155.4.9105058. [DOI] [PubMed] [Google Scholar]
- Long NC, Abraham J, Kobzik L, Weller EA, Krishna Murthy GG, Shore SA. Respiratory tract inflammation during the induction of chronic bronchitis in rats: role of C-fibres. Eur Respir J. 1999;14:46–56. doi: 10.1034/j.1399-3003.1999.14a10.x. [DOI] [PubMed] [Google Scholar]
- Lundberg JM, Martling CR, Saria A, Folkers K, Rosell S. Cigarette smoke-induced airway oedema due to activation of capsaicin-sensitive vagal afferents and substance P release. Neuroscience. 1983;10:1361–1368. doi: 10.1016/0306-4522(83)90117-3. [DOI] [PubMed] [Google Scholar]
- Lundberg JM, Hokfelt T, Martling CR, Saria A, Cuello C. Substance P-immunoreactive sensory nerves in the lower respiratory tract of various mammals including man. Cell Tissue Res. 1984;235:251–261. doi: 10.1007/BF00217848. [DOI] [PubMed] [Google Scholar]
- Ma AA, Ravi K, Bravo EM, Kappagoda CT. Effects of gadolinium chloride on slowly adapting and rapidly adapting receptors of the rabbit lung. Respir Physiol Neurobiol. 2004;141: 125–135. doi: 10.1016/j.resp.2004.04.008. [DOI] [PubMed] [Google Scholar]
- Ma W, Quirion R. Inflammatory mediators modulating the transient receptor potential vanilloid 1 receptor: therapeutic targets to treat inflammatory and neuropathic pain. Expert Opin Ther Targets. 2007;11:307–320. doi: 10.1517/14728222.11.3.307. [DOI] [PubMed] [Google Scholar]
- Manzini S. Bronchodilatation by tachykinins and capsaicin in the mouse main bronchus. Br J Pharmacol. 1992;105(4):968–972. doi: 10.1111/j.1476-5381.1992.tb09086.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mapp CE, Boniotti A, Graf PD, Chitano P, Fabbri LM, Nadel JA. Bronchial smooth muscle responses evoked by toluene diisocyanate are inhibited by ruthenium red and by indomethacin. Eur J Pharmacol. 1991;200:73–76. doi: 10.1016/0014-2999(91)90667-f. [DOI] [PubMed] [Google Scholar]
- Marek W, Potthast JJW, Marczynski B, Baur X. Role of substance P and neurokinin A in toluene diisocyanate- induced increased airway responsiveness in rabbits. Lung. 1996;174:83–97. doi: 10.1007/BF00177703. [DOI] [PubMed] [Google Scholar]
- Martling CR. Sensory nerves containing tachykinins and CGRP in the lower airways. Functional implications for bronchoconstriction, vasodilatation and protein extravasation. Acta Physiol Scand Suppl. 1987;563:1–57. [PubMed] [Google Scholar]
- Massa F, Sibaev A, Marsicano G, Blaudzun H, Storr M, Lutz B. Vanilloid receptor (TRPV1)-deficient mice show increased susceptibility to dinitrobenzene sulfonic acid induced colitis. J Mol Med. 2006;84:142–146. doi: 10.1007/s00109-005-0016-2. [DOI] [PubMed] [Google Scholar]
- Matran R, Martling CR, Lundberg JM. Inhibition of cholinergic and non-adrenergic, non-cholinergic bronchoconstriction in the guinea pig mediated by neuropeptide Y and alpha 2-adrenoceptors and opiate receptors. Eur J Pharmacol. 1989;163:15–23. doi: 10.1016/0014-2999(89)90390-7. [DOI] [PubMed] [Google Scholar]
- Matsumoto S. Effects of vagal stimulation on slowly adapting pulmonary stretch receptors and lung mechanics in anesthetized rabbits. Lung. 1996;174:333–344. doi: 10.1007/BF00176192. [DOI] [PubMed] [Google Scholar]
- Matsumoto S. Effects of sustained constant artificial ventilation on rapidly adapting pulmonary stretch receptors and lung mechanics in rabbits. Life Sci. 1998;62(4):319–325. doi: 10.1016/s0024-3205(97)01113-2. [DOI] [PubMed] [Google Scholar]
- Matsumoto S, Shimizu T. Effects of 5-hydroxytryptamine on rapidly adapting pulmonary stretch receptor activity in the rabbit. J Auton Nerv Syst. 1989;27:35–38. doi: 10.1016/0165-1838(89)90126-4. [DOI] [PubMed] [Google Scholar]
- Matsumoto N, Inoue H, Ichinose M, Ishii M, Inoue C, Sasaki H, Takishima T. Effective sites by sympathetic beta-adrenergic and vagal nonadrenergic inhibitory stimulation in constricted airways. Am Rev Respir Dis. 1985;132:1113–1117. doi: 10.1164/arrd.1985.132.5.1113. [DOI] [PubMed] [Google Scholar]
- Matsumoto S, Okamura H, Suzuki K, Sugai N, Shimizu T. Inhibitory mechanism of CO2 inhalation on slowly adapting pulmonary stretch receptors in the anesthetized rabbit. J Pharmacol Exp Ther. 1996;279(1):402–409. [PubMed] [Google Scholar]
- Matsumoto S, Takeda M, Saiki C, Takahashi T, Ojima K. Effects of vagal and carotid chemoreceptor afferents on the frequency and pattern of spontaneous augmented breaths in rabbits. Lung. 1997;175:175–186. doi: 10.1007/pl00007565. [DOI] [PubMed] [Google Scholar]
- Matsumoto S, Takahashi T, Tanimoto T, Saiki C, Takeda M, Ojima K. Excitatory mechanism of veratridine on slowly adapting pulmonary stretch receptors in anesthetized rabbits. Life Sci. 1998;63:1431–1437. doi: 10.1016/s0024-3205(98)00410-x. [DOI] [PubMed] [Google Scholar]
- Matsumoto S, Takahashi T, Tanimoto T, Saiki C, Takeda M. Effects of potassium channel blockers on CO2-induced slowly adapting pulmonary stretch receptor inhibition. J Pharmacol Exp Ther. 1999;290:974–979. [PubMed] [Google Scholar]
- Matsumoto S, Ikeda M, Nishikawa T. Effects of sodium and potassium channel blockers on hyperinflation-induced slowly adapting pulmonary stretch receptor stimulation in the rat. Life Sci. 2000;67:2167–2175. doi: 10.1016/s0024-3205(00)00813-4. [DOI] [PubMed] [Google Scholar]
- Matsumoto S, Ikeda M, Yoshida S, Nishikawa T, Itoh Y, Fujimi Y, Tanimoto T, Saiki C, Takeda M. The inhibitory effect of ouabain on the response of slowly adapting pulmonary stretch receptors to hyperinflation in the rabbit. Life Sci. 2005;78:112–120. doi: 10.1016/j.lfs.2005.04.036. [DOI] [PubMed] [Google Scholar]
- Matsumoto S, Saiki C, Yoshida S, Takeda M, Kumagai Y. Effect of ouabain on the afterhyperpolarization of slowly adapting pulmonary stretch receptors in the rat lung. Brain Res. 2006;1107:131–139. doi: 10.1016/j.brainres.2006.06.015. [DOI] [PubMed] [Google Scholar]
- Matsuse T, Thomson RJ, Chen XR, Salari H, Schellenberg RR. Capsaicin inhibits airway hyperresponsiveness but not lipoxygenase activity or eosinophilia after repeated aerosolized antigen in guinea pigs. Am Rev Respir Dis. 1991;144:368–372. doi: 10.1164/ajrccm/144.2.368. [DOI] [PubMed] [Google Scholar]
- Mazzone SB, Canning BJ. Evidence for differential reflex regulation of cholinergic and noncholinergic parasympathetic nerves innervating the airways. Am J Respir Crit Care Med. 2002;165(8):1076–1083. doi: 10.1164/ajrccm.165.8.2001121270c. [DOI] [PubMed] [Google Scholar]
- Mazzone SB, Canning BJ. Synergistic interactions between airway afferent nerve subtypes mediating reflex bronchospasm in guinea pigs. Am J Physiol Regul Integr Comp Physiol. 2002;283:R86–R98. doi: 10.1152/ajpregu.00007.2002. [DOI] [PubMed] [Google Scholar]
- Mazzone SB, Geraghty DP. Respiratory action of capsaicin microinjected into the nucleus of the solitary tract: involvement of vanilloid and tachykinin receptors. Br J Pharmacol. 1999;127:473–481. doi: 10.1038/sj.bjp.0702522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazzone SB, McGovern AE. Na+−K+−2Cl− cotransporters and Cl− channels regulate citric acid cough in guinea pigs. J Appl Physiol. 2006;101:635–643. doi: 10.1152/japplphysiol.00106.2006. [DOI] [PubMed] [Google Scholar]
- Mazzone SB, McGovern AE. Immunohistochemical characterization of nodose cough receptor neurons projecting to the trachea of guinea pigs. Cough. 2008;4:9. doi: 10.1186/1745-9974-4-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazzone SB, Mori N, Canning BJ. Synergistic interactions between airway afferent nerve subtypes regulating the cough reflex in guinea-pigs. J Physiol. 2005;569:559–573. doi: 10.1113/jphysiol.2005.093153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McAlexander MA, Undem BJ. Potassium channel blockade induces action potential generation in guinea-pig airway vagal afferent neurones. J Auton Nerv Syst. 2000;78:158–164. doi: 10.1016/s0165-1838(99)00075-2. [DOI] [PubMed] [Google Scholar]
- McAlexander MA, Myers AC, Undem BJ. Inhibition of 5-lipoxygenase diminishes neurally evoked tachykinergic contraction of guinea pig isolated airway. J Pharmacol Exp Ther. 1998;285:602–607. [PubMed] [Google Scholar]
- McDonald DM, Mitchell RA, Gabella G, Haskell A. Neurogenic inflammation in the rat trachea. II. Identity and distribution of nerves mediating the increase in vascular permeability. J Neurocytol. 1988;17:605–628. doi: 10.1007/BF01260989. [DOI] [PubMed] [Google Scholar]
- Meyers JR, MacDonald RB, Duggan A, Lenzi D, Standaert DG, Corwin JT, Corey DP. Lighting up the senses: FM1-43 loading of sensory cells through nonselective ion channels. J Neurosci. 2003;23:4054–4065. doi: 10.1523/JNEUROSCI.23-10-04054.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michoud MC, Amyot R, Jeanneret-Grosjean A, Couture J. Reflex decrease of histamine-induced bronchoconstriction after laryngeal stimulation in humans. Am Rev Respir Dis. 1987;136:618–622. doi: 10.1164/ajrccm/136.3.618. [DOI] [PubMed] [Google Scholar]
- Mills JE, Sellick H, Widdicombe JG. Activity of lung irritant receptors in pulmonary microembolism, anaphylaxis and drug-induced bronchoconstrictions. J Physiol. 1969;203:337–357. doi: 10.1113/jphysiol.1969.sp008867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mills JE, Sellick H, Widdicombe JG. Epithelial irritant receptors in the lungs. In: Ruth P, editor. Breathing: Hering–Breuer Centenary Symposium. London: J. & A. Churchill; 1970. pp. 77–99. [Google Scholar]
- Miserocchi G, Sant'Ambrogio G. Responses of pulmonary stretch receptors to static pressure inflations. Respir Physiol. 1974;21:77–85. doi: 10.1016/0034-5687(74)90009-7. [DOI] [PubMed] [Google Scholar]
- Miura M, Inoue H, Ichinose M, Kimura K, Katsumata U, Takishima T. Effect of nonadrenergic noncholinergic inhibitory nerve stimulation on the allergic reaction in cat airways. Am Rev Respir Dis. 1990;141:29–32. doi: 10.1164/ajrccm/141.1.29. [DOI] [PubMed] [Google Scholar]
- Mizrahi J, D'Orleans-Juste P, Caranikas S, Regoli D. Effects of peptides and amines on isolated guinea pig tracheae as influenced by inhibitors of the metabolism of arachidonic acid. Pharmacology. 1982;25:320–326. doi: 10.1159/000137758. [DOI] [PubMed] [Google Scholar]
- Mohammed SP, Higenbottam TW, Adcock JJ. Effects of aerosol-applied capsaicin, histamine and prostaglandin E2 on airway sensory receptors of anaesthetized cats. J Physiol Lond. 1993;469:51–66. doi: 10.1113/jphysiol.1993.sp019804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreaux B, Nemmar A, Vincke G, Halloy D, Beerens D, Advenier C, Gustin P. Role of substance P and tachykinin receptor antagonists in citric acid-induced cough in pigs. Eur J Pharmacol. 2000;408:305–312. doi: 10.1016/s0014-2999(00)00763-9. [DOI] [PubMed] [Google Scholar]
- Morikawa T, Gallico L, Widdicombe J. Actions of moguisteine on cough and pulmonary rapidly adapting receptor activity in the guinea pig. Pharmacol Res. 1997;35:113–118. doi: 10.1006/phrs.1996.0122. [DOI] [PubMed] [Google Scholar]
- Morrison SF. Differential control of sympathetic outflow. Am J Physiol Regul Integr Comp Physiol. 2001;281:R683–R698. doi: 10.1152/ajpregu.2001.281.3.R683. [DOI] [PubMed] [Google Scholar]
- Mutoh T, Joad JP, Bonham AC. Chronic passive cigarette smoke exposure augments bronchopulmonary C-fibre inputs to nucleus tractus solitarii neurones and reflex output in young guinea-pigs. J Physiol. 2000;523(Pt 1):223–233. doi: 10.1111/j.1469-7793.2000.00223.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mutolo D, Bongianni F, Fontana GA, Pantaleo T. The role of excitatory amino acids and substance P in the mediation of the cough reflex within the nucleus tractus solitarii of the rabbit. Brain Res Bull. 2007;74:284–293. doi: 10.1016/j.brainresbull.2007.07.001. [DOI] [PubMed] [Google Scholar]
- Mutolo D, Bongianni F, Cinelli E, Fontana GA, Pantaleo T. Modulation of the cough reflex by antitussive agents within the caudal aspect of the nucleus tractus solitarii in the rabbit. Am J Physiol Regul Integr Comp Physiol. 2008;295:R243–R251. doi: 10.1152/ajpregu.00184.2008. [DOI] [PubMed] [Google Scholar]
- Myers AC, Kajekar R, Undem BJ. Allergic inflammation-induced neuropeptide production in rapidly adapting afferent nerves in guinea pig airways. Am J Physiol Lung Cell Mol Physiol. 2002;282:L775–L781. doi: 10.1152/ajplung.00353.2001. [DOI] [PubMed] [Google Scholar]
- Nassenstein C, Kwong K, Taylor-Clark T, Kollarik M, Macglashan DM, Braun A, Undem BJ. Expression and function of the ion channel TRPA1 in vagal afferent nerves innervating mouse lungs. J Physiol. 2008;586:1595–1604. doi: 10.1113/jphysiol.2007.148379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nilius B, Owsianik G, Voets T, Peters JA. Transient receptor potential cation channels in disease. Physiol Rev. 2007;87:165–217. doi: 10.1152/physrev.00021.2006. [DOI] [PubMed] [Google Scholar]
- Nishino T. Physiological and pathophysiological implications of upper airway reflexes in humans. Jpn J Physiol. 2000;50:3–14. doi: 10.2170/jjphysiol.50.3. [DOI] [PubMed] [Google Scholar]
- Nohr D, Weihe E. Tachykinin-, calcitonin gene-related peptide-, and protein gene product 9.5-immunoreactive nerve fibers in alveolar walls of mammals. Neurosci Lett. 1991;134:17–20. doi: 10.1016/0304-3940(91)90498-i. [DOI] [PubMed] [Google Scholar]
- O’Connor BJ, Crowther SD, Costello JF, Morley J. Selective airway responsiveness in asthma. Trends Pharmacol Sci. 1999;20:9–11. doi: 10.1016/s0165-6147(98)01280-2. [DOI] [PubMed] [Google Scholar]
- Oh EJ, Mazzone SB, Canning BJ, Weinreich D. Reflex regulation of airway sympathetic nerves in guinea-pigs. J Physiol. 2006;573:549–564. doi: 10.1113/jphysiol.2005.104661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohi Y, Yamazaki H, Takeda R, Haji A. Functional and morphological organization of the nucleus tractus solitarius in the fictive cough reflex of guinea pigs. Neurosci Res. 2005;53:201–209. doi: 10.1016/j.neures.2005.06.016. [DOI] [PubMed] [Google Scholar]
- Ohi Y, Kato F, Haji A. Codeine presynaptically inhibits the glutamatergic synaptic transmission in the nucleus tractus solitarius of the guinea pig. Neuroscience. 2007;146(3):1425–1433. doi: 10.1016/j.neuroscience.2007.02.052. [DOI] [PubMed] [Google Scholar]
- Okajima K, Harada N. Regulation of inflammatory responses by sensory neurons: molecular mechanism(s) and possible therapeutic applications. Curr Med Chem. 2006;13:2241–2251. doi: 10.2174/092986706777935131. [DOI] [PubMed] [Google Scholar]
- Otake K, Ezure K, Lipski J, Wong She RB. Projections from the commissural subnucleus of the nucleus of the solitary tract: an anterograde tracing study in the cat. J Comp Neurol. 1992;324:365–378. doi: 10.1002/cne.903240307. [DOI] [PubMed] [Google Scholar]
- Paintal AS. Vagal sensory receptors and their reflex effects. Physiol Rev. 1973;53:159–227. doi: 10.1152/physrev.1973.53.1.159. [DOI] [PubMed] [Google Scholar]
- Palecek F, Sant'Ambrogio G, Sant'Ambrogio FB, Mathew OP. Reflex responses to capsaicin: intravenous, aerosol, and intratracheal administration. J Appl Physiol. 1989;67:1428–1437. doi: 10.1152/jappl.1989.67.4.1428. [DOI] [PubMed] [Google Scholar]
- Paton JF. Pattern of cardiorespiratory afferent convergence to solitary tract neurons driven by pulmonary vagal C-fiber stimulation in the mouse. J Neurophysiol. 1998;79:2365–2373. doi: 10.1152/jn.1998.79.5.2365. [DOI] [PubMed] [Google Scholar]
- Pedersen KE, Meeker SN, Riccio MM, Undem BJ. Selective stimulation of jugular ganglion afferent neurons in guinea pig airways by hypertonic saline. J Appl Physiol. 1998;84:499–506. doi: 10.1152/jappl.1998.84.2.499. [DOI] [PubMed] [Google Scholar]
- Perretti F, Manzini S. Activation of capsaicin-sensitive sensory fibers modulates PAF-induced bronchial hyperresponsiveness in anesthetized guinea pigs. Am Rev Respir Dis. 1993;148:927–931. doi: 10.1164/ajrccm/148.4_Pt_1.927. [DOI] [PubMed] [Google Scholar]
- Pisarri TE, Jonzon A, Coleridge HM, Coleridge JC. Vagal afferent and reflex responses to changes in surface osmolarity in lower airways of dogs. J Appl Physiol. 1992;73:2305–2313. doi: 10.1152/jappl.1992.73.6.2305. [DOI] [PubMed] [Google Scholar]
- Polosa R, Renaud L, Cacciola R, Prosperini G, Crimi N, Djukanovic R. Sputum eosinophilia is more closely associated with airway responsiveness to bradykinin than methacholine in asthma. Eur Respir J. 1998;12:551–556. doi: 10.1183/09031936.98.12030551. [DOI] [PubMed] [Google Scholar]
- Prabhakar NR, Runold M, Yamamoto Y, Lagercrantz H, Cherniack NS, von Euler C. Role of the vagal afferents in substance P-induced respiratory responses in anaesthetized rabbits. Acta Physiol Scand. 1987;131:63–71. doi: 10.1111/j.1748-1716.1987.tb08206.x. [DOI] [PubMed] [Google Scholar]
- Proud D, Reynolds CJ, Lacapra S, Kagey-Sobotka A, Lichtenstein LM, Naclerio RM. Nasal provocation with bradykinin induces symptoms of rhinitis and a sore throat. Am Rev Respir Dis. 1988;137(3):613–616. doi: 10.1164/ajrccm/137.3.613. [DOI] [PubMed] [Google Scholar]
- Ravi K. Effect of carbon dioxide on the activity of slowly and rapidly adapting pulmonary stretch receptors in cats. J Auton Nerv Syst. 1985;12:267–277. doi: 10.1016/0165-1838(85)90067-0. [DOI] [PubMed] [Google Scholar]
- Ravi K, Kappagoda CT. Responses of pulmonary C-fibre and rapidly adapting receptor afferents to pulmonary congestion and edema in dogs. Can J Physiol Pharmacol. 1992;70:68–76. doi: 10.1139/y92-010. [DOI] [PubMed] [Google Scholar]
- Reynolds CJ, Togias A, Proud D. Airways hyper-responsiveness to bradykinin and methacholine: effects of inhaled fluticasone. Clin Exp Allergy. 2002;32:1174–1179. doi: 10.1046/j.1365-2745.2002.01443.x. [DOI] [PubMed] [Google Scholar]
- Reynolds SM, Docherty R, Robbins J, Spina D, Page CP. Adenosine induces a cholinergic tracheal reflex contraction in guinea pigs in vivo via an adenosine A1 receptor-dependent mechanism. J Appl Physiol. 2008;105:187–196. doi: 10.1152/japplphysiol.01048.2007. [DOI] [PubMed] [Google Scholar]
- Ricciardolo FL, Steinhoff M, Amadesi S, Guerrini R, Tognetto M, Trevisani M, Creminon C, Bertr C, Bunnett NW, Fabbri LM, Salvadori S, Geppetti P. Presence and bronchomotor activity of protease-activated receptor-2 in guinea pig airways. Am J Respir Crit Care Med. 2000;161:1672–1680. doi: 10.1164/ajrccm.161.5.9907133. [DOI] [PubMed] [Google Scholar]
- Riccio MM, Manzini S, Page CP. The effect of neonatal capsaicin on the development of bronchial hyperresponsiveness in allergic rabbits. Eur J Pharmacol. 1993;232:89–97. doi: 10.1016/0014-2999(93)90732-w. [DOI] [PubMed] [Google Scholar]
- Riccio MM, Matsumoto T, Adcock JJ, Douglas GJ, Spina D, Page CP. The effect of 15-HPETE on airway responsiveness and pulmonary cell recruitment in rabbits. Br J Pharmacol. 1997;122:249–256. doi: 10.1038/sj.bjp.0701379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ricco MM, Kummer W, Biglari B, Myers AC, Undem BJ. Interganglionic segregation of distinct vagal afferent fibre phenotypes in guinea-pig airways. J Physiol. 1996;496(Pt 2): 521–530. doi: 10.1113/jphysiol.1996.sp021703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson CA, Herbert DA, Mitchell RA. Modulation of pulmonary stretch receptors and airway resistance by parasympathetic efferents. J Appl Physiol. 1984;57:1842–1849. doi: 10.1152/jappl.1984.57.6.1842. [DOI] [PubMed] [Google Scholar]
- Riedel F, Benden C, Philippou S, Streckert HJ, Marek W. Role of sensory neuropeptides in PIV-3-infection-induced airway hyperresponsiveness in guinea pigs. Respiration. 1997;64:211–219. doi: 10.1159/000196673. [DOI] [PubMed] [Google Scholar]
- Roberts AM, Hahn HL, Schultz HD, Nadel JA, Coleridge HM, Coleridge J. Afferent vagal C-fibres are responsible for the reflex airway constriction and secretion evoked by pulmonary administration of SO2 in dogs. Physiologist. 1982;250:226. [Google Scholar]
- Roisman GL, Lacronique JG, Desmazes DN, Carre C, Le-Cae A, Dusser DJ. Airway responsiveness to bradykinin is related to eosinophilic inflammation in asthma. Am J Respir Crit Care Med. 1996;153:381–390. doi: 10.1164/ajrccm.153.1.8542147. [DOI] [PubMed] [Google Scholar]
- Rosenberg HF, Phipps S, Foster PS. Eosinophil trafficking in allergy and asthma. J Allergy Clin Immunol. 2007;119:1303–1310. doi: 10.1016/j.jaci.2007.03.048. [DOI] [PubMed] [Google Scholar]
- Ruan T, Lin YS, Lin KS, Kou YR. Sensory transduction of pulmonary reactive oxygen species by capsaicin-sensitive vagal lung afferent fibres in rats. J Physiol. 2005;565:563–578. doi: 10.1113/jphysiol.2005.086181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sampson SR, Vidruk EH. Properties of ‘irritant’ receptors in canine lung. Respir Physiol. 1975;25:9–22. doi: 10.1016/0034-5687(75)90047-x. [DOI] [PubMed] [Google Scholar]
- Sanico AM, Philip G, Lai GK, Togias A. Hyperosmolar saline induces reflex nasal secretions, evincing neural hyperresponsiveness in allergic rhinitis. J Appl Physiol. 1999;86: 1202–1210. doi: 10.1152/jappl.1999.86.4.1202. [DOI] [PubMed] [Google Scholar]
- Sant'Ambrogio FB, Sant'Ambrogio G, Fisher JT. Lung mechanics and activity of slowly adapting airway stretch receptors. Eur Respir J. 1988;1:685–690. [PubMed] [Google Scholar]
- Saria A, Martling CR, Dalsgaard CJ, Lundberg JM. Evidence for substance P-immunoreactive spinal afferents that mediate bronchoconstriction. Acta Physiol Scand. 1985;125:407–414. doi: 10.1111/j.1748-1716.1985.tb07736.x. [DOI] [PubMed] [Google Scholar]
- Schelegle ES, Green JF. An overview of the anatomy and physiology of slowly adapting pulmonary stretch receptors. Respir Physiol. 2001;125:17–31. doi: 10.1016/s0034-5687(00)00202-4. [DOI] [PubMed] [Google Scholar]
- Schlemper V, Calixto JB. Mechanisms underlying the contraction induced by bradykinin in the guinea pig epithelium-denuded trachea. Can J Physiol Pharmacol. 2002;80:360–367. doi: 10.1139/y02-061. [DOI] [PubMed] [Google Scholar]
- Sekizawa S, Joad JP, Bonham AC. Substance P presynaptically depresses the transmission of sensory input to bronchopulmonary neurons in the guinea pig nucleus tractus solitarii. J Physiol. 2003;552:547–559. doi: 10.1113/jphysiol.2003.051326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sellick H, Widdicombe JG. The activity of lung irritant receptors during pneumothorax, hyperpnoea and pulmonary vascular congestion. J Physiol. 1969;203:359–381. doi: 10.1113/jphysiol.1969.sp008868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shah S, Page CP, Spina D. Nociceptin inhibits non-adrenergic non-cholinergic contraction in guinea-pig airway. Br J Pharmacol. 1998;125:510–516. doi: 10.1038/sj.bjp.0702068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva-Carvalho L, Paton JF, Rocha I, Goldsmith GE, Spyer KM. Convergence properties of solitary tract neurons responsive to cardiac receptor stimulation in the anesthetized cat. J Neurophysiol. 1998;79:2374–2382. doi: 10.1152/jn.1998.79.5.2374. [DOI] [PubMed] [Google Scholar]
- Skogvall S, Berglund M, ence-Guzman MF, Svensson K, Jonsson P, Persson CG, Sterner O. Effects of capsazepine on human small airway responsiveness unravel a novel class of bronchorelaxants. Pulm Pharmacol Ther. 2007;20:273–280. doi: 10.1016/j.pupt.2006.03.002. [DOI] [PubMed] [Google Scholar]
- Slats AM, Janssen K, van SA, van der Plas DT, Schot R, van den Aardweg JG, de Jongste JC, Hiemstra PS, Mauad T, Rabe KF, Sterk PJ. Bronchial inflammation and airway responses to deep inspiration in asthma and chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 2007;176:121–128. doi: 10.1164/rccm.200612-1814OC. [DOI] [PubMed] [Google Scholar]
- Sont JK, Booms P, Bel EH, Vandenbroucke JP, Sterk PJ. The determinants of airway hyperresponsiveness to hypertonic saline in atopic asthma in vivo. Relationship with sub-populations of peripheral blood leucocytes. Clin Exp Allergy. 1993;23:678–688. doi: 10.1111/j.1365-2222.1993.tb01794.x. [DOI] [PubMed] [Google Scholar]
- Sont JK, Willems LN, Bel EH, van Krieken JH, Vandenbroucke JP, Sterk PJ. Clinical control and histopathologic outcome of asthma when using airway hyperresponsiveness as an additional guide to long-term treatment. The AMPUL Study Group. Am J Respir Crit Care Med. 1999;159:1043–1051. doi: 10.1164/ajrccm.159.4.9806052. [DOI] [PubMed] [Google Scholar]
- Spina D, Page CP. Airway sensory nerves in asthma – targets for therapy? Pulm Pharmacol. 1996;9:1–18. doi: 10.1006/pulp.1996.0001. [DOI] [PubMed] [Google Scholar]
- Spina D, Page CP. Pharmacology of airway irritability. Curr Opin Pharmacol. 2002;2:264–272. doi: 10.1016/s1471-4892(02)00159-5. [DOI] [PubMed] [Google Scholar]
- Spina D, McKenniff MG, Coyle AJ, Seeds EA, Tramontana M, Perretti F, Manzini S, Page CP. Effect of capsaicin on PAF-induced bronchial hyperresponsiveness and pulmonary cell accumulation in the rabbit. Br J Pharmacol. 1991;103:1268–1274. doi: 10.1111/j.1476-5381.1991.tb12335.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spina D, Harrison S, Page CP. Regulation by phosphodiesterase isoenzymes of non-adrenergic non-cholinergic contraction in guinea-pig isolated main bronchus. Br J Pharmacol. 1995;116:2334–2340. doi: 10.1111/j.1476-5381.1995.tb15074.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spina D, Matera MG, Riccio MM, Page CP. A comparison of sensory nerve function in human, guinea-pig, rabbit and marmoset airway. Life Sci. 1998;63:1629–1643. doi: 10.1016/s0024-3205(98)00432-9. [DOI] [PubMed] [Google Scholar]
- Stone RA, Worsdell YM, Fuller RW, Barnes PJ. Effects of 5-hydroxytryptamine and 5-hydroxytryptophan infusion on the human cough reflex. J Appl Physiol. 1993;74:396–401. doi: 10.1152/jappl.1993.74.1.396. [DOI] [PubMed] [Google Scholar]
- Storr M. TRPV1 in colitis: is it a good or a bad receptor? – a viewpoint. Neurogastroenterol Motil. 2007;19:625–629. doi: 10.1111/j.1365-2982.2007.00946.x. [DOI] [PubMed] [Google Scholar]
- Stretton CD, Belvisi MG, Barnes PJ. Modulation of neural bronchoconstrictor responses in the guinea pig respiratory tract by vasoactive intestinal peptide. Neuropeptides. 1991;18(3):149–157. doi: 10.1016/0143-4179(91)90107-t. [DOI] [PubMed] [Google Scholar]
- Sudo T, Hayashi F, Nishino T. Responses of tracheobronchial receptors to inhaled furosemide in anesthetized rats. Am J Respir Crit Care Med. 2000;162(3 Pt 1):971–975. doi: 10.1164/ajrccm.162.3.2001001. [DOI] [PubMed] [Google Scholar]
- Szallasi A. The vanilloid (capsaicin) receptor: receptor types and species differences. Gen Pharmacol. 1994;25:223–243. doi: 10.1016/0306-3623(94)90049-3. [DOI] [PubMed] [Google Scholar]
- Szallasi A, Blumberg PM. Vanilloid (capsaicin) receptors and mechanisms. Pharmacol Rev. 1999;51:159–211. [PubMed] [Google Scholar]
- Szarek JL, Gillespie MN, Altiere RJ, Diamond L. Reflex activation of the nonadrenergic noncholinergic inhibitory nervous system in feline airways. Am Rev Respir Dis. 1986;133:1159–1162. doi: 10.1164/arrd.1986.133.6.1159. [DOI] [PubMed] [Google Scholar]
- Szarek JL, Stewart NL, Spurlock B, Schneider C. Sensory nerve- and neuropeptide-mediated relaxation responses in airways of Sprague-Dawley rats. J Appl Physiol. 1995;78(5):1679–1687. doi: 10.1152/jappl.1995.78.5.1679. [DOI] [PubMed] [Google Scholar]
- Taguchi O, Kikuchi Y, Hida W, Iwase N, Satoh M, Chonan T, Takishima T. Effects of bronchoconstriction and external resistive loading on the sensation of dyspnea. J Appl Physiol. 1991;71:2183–2190. doi: 10.1152/jappl.1991.71.6.2183. [DOI] [PubMed] [Google Scholar]
- Taguchi O, Kikuchi Y, Hida W, Iwase N, Okabe S, Chonan T, Takishima T. Prostaglandin E2 inhalation increases the sensation of dyspnea during exercise. Am Rev Respir Dis. 1992;145:1346–1349. doi: 10.1164/ajrccm/145.6.1346. [DOI] [PubMed] [Google Scholar]
- Takagi S, Umezaki T, Shin T. Convergence of laryngeal afferents with different natures upon cat NTS neurons. Brain Res Bull. 1995;38:261–268. doi: 10.1016/0361-9230(95)00098-y. [DOI] [PubMed] [Google Scholar]
- Takahashi T, Belvisi MG, Barnes PJ. Modulation of neurotransmission in guinea-pig airways by galanin and the effect of a new antagonist galantide. Neuropeptides. 1994;26(4):245–251. doi: 10.1016/0143-4179(94)90078-7. [DOI] [PubMed] [Google Scholar]
- Tang L, Chen Y, Chen Z, Blumberg PM, Kozikowski AP, Wang ZJ. Antinociceptive pharmacology of N-(4-chlorobenzyl)-N′-(4-hydroxy-3-iodo-5-methoxybenzyl) thiourea, a high-affinity competitive antagonist of the transient receptor potential vanilloid 1 receptor. J Pharmacol Exp Ther. 2007;321:791–798. doi: 10.1124/jpet.106.117572. [DOI] [PubMed] [Google Scholar]
- Tatar M, Sant'Ambrogio G, Sant'Ambrogio FB. Laryngeal and tracheobronchial cough in anesthetized dogs. J Appl Physiol. 1994;76:2672–2679. doi: 10.1152/jappl.1994.76.6.2672. [DOI] [PubMed] [Google Scholar]
- Tatar M, Pecova R, Karcolova D. Sensitivity of the cough reflex in awake guinea pigs, rats and rabbits. Bratisl Lek Listy. 1997;98:539–543. [PubMed] [Google Scholar]
- Tatar M, Webber SE, Widdicombe JG. Lung C-fibre receptor activation and defensive reflexes in anaesthetized cats. J Physiol. 1988;402:411–420. doi: 10.1113/jphysiol.1988.sp017212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor-Clark TE, McAlexander MA, Nassenstein C, Sheardown SA, Wilson S, Thornton J, Carr MJ, Undem BJ. Relative contributions of TRPA1 and TRPV1 channels in the activation of vagal bronchopulmonary C-fibres by the endogenous autacoid 4-oxononenal. J Physiol. 2008;586:3447–3459. doi: 10.1113/jphysiol.2008.153585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tepper JS, Costa DL, Fitzgerald S, Doerfler DL, Bromberg PA. Role of tachykinins in ozone-induced acute lung injury in guinea pigs. J Appl Physiol. 1993;75:1404–1411. doi: 10.1152/jappl.1993.75.3.1404. [DOI] [PubMed] [Google Scholar]
- Thompson JE, Scypinski LA, Gordon T, Sheppard D. Tachykinins mediate the acute increase in airway responsiveness caused by toluene diisocyanate in guinea pigs. Am Rev Respir Dis. 1987;136:43–49. doi: 10.1164/ajrccm/136.1.43. [DOI] [PubMed] [Google Scholar]
- Tominaga M, Caterina MJ, Malmberg AB, Rosen TA, Gilbert H, Skinner K, Raumann BE, Basbaum AI, Julius D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron. 1998;21:531–543. doi: 10.1016/s0896-6273(00)80564-4. [DOI] [PubMed] [Google Scholar]
- Trevisani M, Milan A, Gatti R, Zanasi A, Harrison S, Fontana G, Morice AH, Geppetti P. Antitussive activity of iodo-resiniferatoxin in guinea pigs. Thorax. 2004;59:769–772. doi: 10.1136/thx.2003.012930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tucker RC, Kagaya M, Page CP, Spina D. The endogenous cannabinoid agonist, anandamide stimulates sensory nerves in guinea-pig airways. Br J Pharmacol. 2001;132:1127–1135. doi: 10.1038/sj.bjp.0703906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umeno E, McDonald DM, Nadel JA. Hypertonic saline increases vascular permeability in the rat trachea by producing neurogenic inflammation. J Clin Invest. 1990;85:1905–1908. doi: 10.1172/JCI114652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Undem BJ, Kollarik M. Characterization of the vanilloid receptor 1 antagonist iodo-resiniferatoxin on the afferent and efferent function of vagal sensory C-fibers. J Pharmacol Exp Ther. 2002;303(2):716–722. doi: 10.1124/jpet.102.039727. [DOI] [PubMed] [Google Scholar]
- Undem BJ, Meeker SN, Chen J. Inhibition of neurally mediated nonadrenergic, noncholinergic contractions of guinea pig bronchus by isozyme-selective phosphodiesterase inhibitors. J Pharmacol Exp Ther. 1994;271(2):811–817. [PubMed] [Google Scholar]
- Undem BJ, Carr MJ, Kollarik M. Physiology and plasticity of putative cough fibres in the Guinea pig. Pulm Pharmacol Ther. 2002;15:193–198. doi: 10.1006/pupt.2002.0362. [DOI] [PubMed] [Google Scholar]
- Undem BJ, Oh EJ, Lancaster E, Weinreich D. Effect of extracellular calcium on excitability of guinea pig airway vagal afferent nerves. J Neurophysiol. 2003;89:1196–1204. doi: 10.1152/jn.00553.2002. [DOI] [PubMed] [Google Scholar]
- Undem BJ, Chuaychoo B, Lee MG, Weinreich D, Myers AC, Kollarik M. Subtypes of vagal afferent C-fibres in guinea-pig lungs. J Physiol. 2004;556:905–917. doi: 10.1113/jphysiol.2003.060079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Berge M, Kerstjens HA, Meijer RJ, de Reus DM, Koeter GH, Kauffman HF, Postma DS. Corticosteroid-induced improvement in the PC20 of adenosine monophosphate is more closely associated with reduction in airway inflammation than improvement in the PC20 of methacholine. Am J Respir Crit Care Med. 2001;164:1127–1132. doi: 10.1164/ajrccm.164.7.2102135. [DOI] [PubMed] [Google Scholar]
- Van Schoor J, Joos GF, Pauwels RA. Indirect bronchial hyperresponsiveness in asthma: mechanisms, pharmacology and implications for clinical research. Eur Respir J. 2000;16:514–533. doi: 10.1034/j.1399-3003.2000.016003514.x. [DOI] [PubMed] [Google Scholar]
- Van Schoor J, Joos GF, Pauwels RA. Effect of inhaled fluticasone on bronchial responsiveness to neurokinin A in asthma. Eur Respir J. 2002;19:997–1002. doi: 10.1183/09031936.02.01112001. [DOI] [PubMed] [Google Scholar]
- Vardhan A, Kachroo A, Sapru HN. Excitatory amino acid receptors in the nucleus tractus solitarius mediate the responses to the stimulation of cardio-pulmonary vagal afferent C fiber endings. Brain Res. 1993;618:23–31. doi: 10.1016/0006-8993(93)90424-l. [DOI] [PubMed] [Google Scholar]
- Verleden GM, Belvisi MG, Rabe KF, Miura M, Barnes PJ. Beta 2-adrenoceptor agonists inhibit NANC neural bronchoconstrictor responses in vitro. J Appl Physiol. 1993;74:1195–1199. doi: 10.1152/jappl.1993.74.3.1195. [DOI] [PubMed] [Google Scholar]
- Vesely KR, Hyde DM, Stovall MY, Harkema JR, Green JF, Schelegle ES. Capsaicin-sensitive C-fiber-mediated protective responses in ozone inhalation in rats. J Appl Physiol. 1999;86:951–962. doi: 10.1152/jappl.1999.86.3.951. [DOI] [PubMed] [Google Scholar]
- Vidruk EH, Hahn HL, Nadel JA, Sampson SR. Mechanisms by which histamine stimulates rapidly adapting receptors in dog lungs. J Appl Physiol. 1977;43:397–402. doi: 10.1152/jappl.1977.43.3.397. [DOI] [PubMed] [Google Scholar]
- Warr D. The neurokinin1 receptor antagonist aprepitant as an antiemetic for moderately emetogenic chemotherapy. Expert Opin Pharmacother. 2006;7:1653–1658. doi: 10.1517/14656566.7.12.1653. [DOI] [PubMed] [Google Scholar]
- Watanabe N, Horie S, Michael GJ, Spina D, Page CP, Priestley JV. Immunohistochemical localization of vanilloid receptor subtype 1 (TRPV1) in the guinea pig respiratory system. Pulm Pharmacol Ther. 2005;18:187–197. doi: 10.1016/j.pupt.2004.12.002. [DOI] [PubMed] [Google Scholar]
- Watanabe N, Horie S, Michael GJ, Keir S, Spina D, Page CP, Priestley JV. Immunohistochemical co-localization of transient receptor potential vanilloid (TRPV)1 and sensory neuropeptides in the guinea-pig respiratory system. Neuroscience. 2006;141:1533–1543. doi: 10.1016/j.neuroscience.2006.04.073. [DOI] [PubMed] [Google Scholar]
- Welch JM, Simon SA, Reinhart PH. The activation mechanism of rat vanilloid receptor 1 by capsaicin involves the pore domain and differs from the activation by either acid or heat. Proc Natl Acad Sci USA. 2000;97:13889–13894. doi: 10.1073/pnas.230146497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Widdicombe JG. Receptors in the trachea and bronchi of the cat. J Physiol. 1954;123:71–104. doi: 10.1113/jphysiol.1954.sp005034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Widdicombe JG. Respiratory reflexes from the trachea and bronchi of the cat. J Physiol. 1954;123:55–70. doi: 10.1113/jphysiol.1954.sp005033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson CG, Zhang Z, Bonham AC. Non-NMDA receptors transmit cardiopulmonary C fibre input in nucleus tractus solitarii in rats. J Physiol. 1996;496(Pt 3):773–785. doi: 10.1113/jphysiol.1996.sp021726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wine JJ. Parasympathetic control of airway submucosal glands: central reflexes and the airway intrinsic nervous system. Auton Neurosci. 2007;133:35–54. doi: 10.1016/j.autneu.2007.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winning AJ, Hamilton RD, Shea SA, Knott C, Guz A. The effect of airway anaesthesia on the control of breathing and the sensation of breathlessness in man. Clin Sci (Lond) 1985;68(2):215–225. doi: 10.1042/cs0680215. [DOI] [PubMed] [Google Scholar]
- Winning AJ, Hamilton RD, Shea SA, Guz A. Respiratory and cardiovascular effects of central and peripheral intravenous injections of capsaicin in man: evidence for pulmonary chemosensitivity. Clin Sci (Lond) 1986;71(5):519–526. doi: 10.1042/cs0710519. [DOI] [PubMed] [Google Scholar]
- Wu ZX, Satterfield BE, Dey RD. Substance P released from intrinsic airway neurons contributes to ozone-enhanced airway hyperresponsiveness in ferret trachea. J Appl Physiol. 2003;95:742–750. doi: 10.1152/japplphysiol.00109.2003. [DOI] [PubMed] [Google Scholar]
- Xiang A, Uchida Y, Nomura A, Iijima H, Sakamoto T, Ishii Y, Morishima Y, Masuyama K, Zhang M, Hirano K, Sekizawa K. Involvement of thromboxane A(2) in airway mucous cells in asthma-related cough. J Appl Physiol. 2002;92:763–770. doi: 10.1152/jappl.2002.92.2.763. [DOI] [PubMed] [Google Scholar]
- Yamamoto Y, Atoji Y, Suzuki Y. Nerve endings in bronchi of the dog that react with antibodies against neurofilament protein. J Anat. 1995;187(1):59–65. [PMC free article] [PubMed] [Google Scholar]
- Yamamoto Y, Ootsuka T, Atoji Y, Suzuki Y. Morphological and quantitative study of the intrinsic nerve plexuses of the canine trachea as revealed by immunohistochemical staining of protein gene product 9.5. Anat Rec. 1998;250:438–447. doi: 10.1002/(SICI)1097-0185(199804)250:4<438::AID-AR7>3.0.CO;2-T. [DOI] [PubMed] [Google Scholar]
- Yamamoto Y, Sato Y, Taniguchi K. Distribution of TRPV1- and TRPV2-immunoreactive afferent nerve endings in rat trachea. J Anat. 2007;211:775–783. doi: 10.1111/j.1469-7580.2007.00821.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshihara S, Nadel JA, Figini M, Emanueli C, Pradelles P, Geppetti P. Endogenous nitric oxide inhibits bronchoconstriction induced by cold-air inhalation in guinea pigs: role of kinins. Am J Respir Crit Care Med. 1998;157:547–552. doi: 10.1164/ajrccm.157.2.9704074. [DOI] [PubMed] [Google Scholar]
- Yu J. Airway mechanosensors. Respir Physiol Neurobiol. 2005;148:217–243. doi: 10.1016/j.resp.2004.12.007. [DOI] [PubMed] [Google Scholar]
- Yu J, Coleridge JC, Coleridge HM. Influence of lung stiffness on rapidly adapting receptors in rabbits and cats. Respir Physiol. 1987;68:161–176. doi: 10.1016/s0034-5687(87)80003-8. [DOI] [PubMed] [Google Scholar]
- Yu J, Schultz HD, Goodman J, Coleridge JC, Coleridge HM, Davis B. Pulmonary rapidly adapting receptors reflexly increase airway secretion in dogs. J Appl Physiol. 1989;67:682–687. doi: 10.1152/jappl.1989.67.2.682. [DOI] [PubMed] [Google Scholar]