

Abstract
The occurrence of avian influenza A(H5N1) in Hong Kong in 1997 led to the development of a “One-Health” approach to deal with emerging infectious diseases that has been applied to other emergent diseases such as SARS and the pandemic H1N1 2009. Evaluation of poultry marketing and production systems and investigations at the animal-human interface, led to defining the routes of human exposure to avian influenza and factors that allowed virus to multiply and persist. Active and systematic surveillance of apparently healthy as well as diseased poultry and wild birds provided evidence of ongoing virus evolution in the wider region. Epidemiological studies, supplemented with molecular epidemiology, helped to elucidate the role of the poultry marketing system and live poultry markets in the persistence of avian influenza viruses and provided evidence for the impact of interventions designed to interrupt virus transmission. Enhanced bio-security, active surveillance together with targeted and evidence-based interventions in the poultry production, and marketing system together with poultry vaccination has prevented further human H5N1 disease and minimized outbreaks of poultry disease in Hong Kong. Similar strategies have led to the understanding of the emergence of SARS and provided options for preventing the re-emergence of this disease. Surveillance of influenza in swine has provided insights into the emergence of the 2009 pandemic, to the reverse zoonosis of the pandemic virus from humans to swine and to the emergence of novel reassortant viruses within swine. “One Health” strategies are not “cost-free” and require sensitive implementation to optimize food-safety and food security, while safeguarding the economics of animal husbandry and the environment and remaining sensitive to cultural practices.
Keywords: Influenza Virus, Avian Influenza, H5N1 Virus, Avian Influenza Virus, Wild Bird
Introduction
The concept of One Health, defined as “collaborative effects of multiple disciplines working locally, nationally and globally to attain optimal health for people, animals, and the environment” (American Veterinary Medical Association 2008) has received considerable attention in the past 4 years. However, this is not a new idea and there are examples of One Health approaches being adopted prior to this time, especially for zoonotic diseases such as rabies. The role of environmental factors in infectious diseases has also been recognized for hundreds of years, even if the mechanisms involved in the interactions were poorly understood (Brandt 2012). Hong Kong, a crowded city where outbreaks of infectious disease can spread rapidly, provides a good example of the benefits of a ‘One Health’ approach to disease control and prevention for both human and animal health.
This was best exemplified through experiences with avian influenza viruses of the H5N1 subtype in humans and other animals which emerged as a serious zoonotic disease with pandemic potential in 1997, but has also been applied to a number of other diseases, including SARS. Without close cooperation between human health and animal health practitioners, as well as, an understanding of the ecological factors (both natural and human-derived) that allowed influenza A(H5N1) virus to emerge, persist and spread, the disease in poultry and humans could not have been contained and prevented.
Studies on Influenza Pre 1997 in Hong Kong
Studies of animal influenza viruses in Hong Kong and southern China originated from its relationship, one way or another, to human disease. Reports of an outbreak of disease with high mortality in pigs in cities along the Chinese coast in the wake of the 1918 Spanish flu pandemic, probably represented spill-over of the pandemic virus to swine (Chun et al. 1919); a situation reminiscent of that following the 2009 human influenza pandemic. In a seminal series of studies from 1977 onward, Shortridge et al. studied animal viruses in order to understand human influenza pandemic emergence (Shortridge 1992). In the course of these investigations, they elucidated much of the ecology of influenza in poultry, wild birds, swine, and horses in the region. They demonstrated a diversity of influenza virus subtypes circulating in domestic poultry and that domestic ducks had a much higher rate of carriage of influenza virus subtypes than chickens and other terrestrial birds (Shortridge 1982). All avian viruses isolated (including H5N2 and H5N3), met established criteria for low pathogenic influenza viruses.
Shortridge showed that human influenza A (H3N2) viruses from 1968 (e.g. A/Hong Kong/1/68-like viruses) remained antigenically unchanged in swine in 1976, long after that virus had been replaced in humans by sequential antigenic drift variants. He presciently suggested that “pigs may serve as a potential reservoir for future human pandemics as well as a possible source of genetic information for recombination between human and porcine strains of influenza virus” (Shortridge et al. 1977). Given the farming practices in China provide opportunities for close interactions between domestic ducks and other poultry with swine and humans, it was suggested that swine may serve as a mixing vessel for pandemic emergence (Scholtissek et al. 1985) and, since the pandemics of 1957 and 1968 started in China, that China was a pandemic epicenter (Shortridge and Stuart-Harris 1982). Among other things, Shortridge also investigated outbreaks of equine influenza (H3N8) in race-horses in Hong Kong in 1992 (Watkins et al. 1993) and elsewhere.
Avian Influenza H5N1 Events in 1997
In March 1997, an outbreak of disease resulted in the death of virtually all of the chickens on a local farm. Collaborative studies between the Department of Agriculture Fisheries and Conservation and The University of Hong Kong identified this to be caused by a highly pathogenic avian influenza (HPAI) virus of the H5N1 subtype. Two other farms also reported outbreaks characterized by high mortality in April and May 1997 caused by the same virus. No further cases were detected in poultry for the next 6 months despite investigations of a number of other disease outbreaks on poultry farms. The incident was reported to the relevant international veterinary authority, the World Organization for Animal Health (OIE). As outbreaks of HPAI at this time were not considered to have public health relevance, the public health authorities (the Department of Health) were not informed of this event.
In May 1997, a child infected with an influenza virus in Hong Kong died with Reye’s syndrome. The influenza virus isolated from this child failed to be typed by the conventional seasonal human influenza antisera and was referred to the WHO influenza Collaborating Centre and reference laboratories, as was recommended practice. It took 3 months for the virus to be identified as a HPAI H5N1 virus (Claas et al. 1998). In retrospect, if the “One Health” concept had been in application and relevant public health authorities were made aware of the outbreak of H5N1 HPAI in poultry, the identification of the virus isolate from this child as a H5N1 virus may have occurred sooner. This event served as a catalyst for improved communication between government departments, and between government and university departments on novel diseases of potential public health significance. These links became vital later in 1997.
The human case attracted considerable international attention, because it was the first known case of an avian influenza virus associated with fatal disease in both poultry and humans. The source of the virus for the human case was never determined, but with the benefit of hindsight and subsequent findings in markets later in the year, it is reasonable to suppose that H5N1 viruses were probably present in live poultry markets (LPM) at the time. In southern China, as well as in a number of other Asian countries, poultry are an important source of protein and there is a cultural preference to consume freshly killed (rather than frozen) poultry, leading to the presence of large numbers retail live poultry “wet” markets.
No further new human cases were reported for 4 months and it was initially assumed that the human case was an isolated one. This proved to be a false hope as a new series of human cases was detected starting in early November. New avian cases were found in poultry markets from early December. As the number of human cases increased, a range of interim control measures, including a temporary short term suspension of trade in live poultry and improved cleaning of markets were implemented. But further human cases occurred, associated with significant mortality; ultimately 6 of the 18 diagnosed cases proved fatal. There was no evidence of efficient human-to-human transmission. Still, there were growing concerns in the local and international press about the potential for emergence of a severe influenza pandemic that may rival the 1918 “Spanish flu” pandemic.
Chickens on a local farm were found to be infected on December 27. The next day, a batch of birds showing signs consistent with highly pathogenic avian influenza was detected in the main wholesale market, and the decision was taken to cull all chickens on commercial farms and in LPM in Hong Kong, based on an assessment of the risk that the continued exposure of humans to this virus posed, not only to the citizens of Hong Kong, but potentially to global public health. These decisions had to be made on the best available information at the time and from first principles of disease control and prevention. Some 1.5 million poultry were killed and sent to landfill for appropriate burial in a 5 day period from December 29. These measures resulted in the elimination of the 1997 H5N1 Hong Kong strain, and no new locally acquired human cases occurred in Hong Kong after depopulation of poultry in markets.
Just prior to and during the major culling exercise in 1997, a number of cross disciplinary investigations were conducted that, over time, provided confirmation of the importance of poultry and LPM in the epidemiology of this disease. Surveillance studies in markets in Hong Kong revealed a very high prevalence of infection in poultry in markets, with almost 20 % of chickens in markets being infected at the time of the mass poultry cull (Shortridge 1999). This demonstrated the importance of implementing measures to control infection in markets. Given the probable high exposure of people to infected poultry, the limited number of human cases suggested that transmission of the virus from birds to humans was very inefficient. Epidemiological studies to assess risk factors for human infection were carried out in collaboration with staff of the US Centers for Disease Control, who were invited to Hong Kong to assist with the investigation. These revealed that a visit to a live poultry market in the 7 days prior to disease onset was a significant risk factor (Mounts et al. 1999). Overall, few people in Hong Kong had antibodies to H5 virus (other than some who worked with poultry, some health workers and some government workers involved in culling operations) indicating that transmission of the virus from poultry to human and from human-to-human was exceedingly inefficient (Buxton Bridges et al. 2000; Bridges et al. 2002).
Culling of all commercial poultry in Hong Kong farms and markets was followed by thorough cleaning and disinfection of these places, and implementation of a raft of new preventive measures built on the premise that improved farm and market hygiene, and biosecurity would reduce the risk of future virus incursions.
Re-introduction of Trade in Live Poultry in 1998
After the dramatic events of 1997, the public and political expectations were that no further human H5N1 infections would occur in Hong Kong, and that this virus would be kept under control in poultry. To do this, it was essential to understand the factors that resulted in the emergence, maintenance, and transmission of this virus. At this time, no human or avian H5N1 infections were reported from Mainland China or the wider region, but it could not be assumed that H5N1 influenza viruses were not circulating there (an assumption that was subsequently proven correct).
At the time of the outbreaks of disease associated with influenza A(H5N1) in 1997, the system of rearing and selling of poultry in Hong Kong was based mainly on medium scale (predominantly 10,000–20,000 head) family-owned farms. Biosecurity measures on most of these farms were weak with few preventive measures applied to visitors. Poultry houses were not bird proof and most operated on a continuous basis with multiple batches of birds of different ages reared in the same or adjacent sheds. Land scarcity meant that many farms were geographically concentrated, with some areas containing 20 independently managed chicken farms per square kilometer.
Hygiene conditions and management procedures in LPM, both retail and wholesale were such that they facilitated perpetuation of viruses. Retail market stalls were rarely, if ever, emptied and some poultry stayed in markets longer than the incubation period of the disease. Market stalls kept different types of poultry and there were no controls on movement of birds or humans in and out of the markets. Some traders shifted poultry that were in excess of market need from the wholesale market back to holding farms (so-called “chicken-inns’) until demand increased. These farms were located in the vicinity of other commercial farms.
Based on the known epidemiology of avian influenza viruses (e.g. virus shed via the cloaca, aquatic poultry possible short term carriers) and the nature of existing production and marketing systems within Hong Kong and in southern parts of mainland China, measures were introduced to reduce the likelihood of infection recurring in farms and markets.
These measures included:
Replacement of wooden cages with plastic cages,
Cleaning systems for cages and vehicles introduced to the wholesale markets,
Strict controls on the sources of poultry, with only selected farms in southern China allowed to send poultry to Hong Kong
Changes in legislation that prevented movement of poultry back from markets to farms
Inspection and testing of all consignments of poultry prior to sale and for imported poultry on arrival to Hong Kong.
Ducks and geese could no longer be kept, sold, or transported with terrestrial poultry into or within Hong Kong, and this resulted in the development of a separate wholesale market and slaughterhouse delivering fresh carcasses to retail LPM. This measure was introduced, because it was assumed that it will be extremely difficult to guarantee that they remained free from infection, given the way ducks and geese were reared. Subsequent results from surveillance in the central duck and goose slaughterhouse demonstrated this assumption was correct, when virus was detected in geese in 1999 (Cauthen et al. 2000) and then in ducks in 2000 and 2001 (Guan et al. 2002a).
All farms in Hong Kong had to meet new hygiene and biosecurity measures introduced as farm license conditions. The main wholesale market was rearranged, and one section that previously housed slaughter rooms with very basic facilities was pulled down. The practice of killing birds in the wholesale chicken market was banned; slaughter of live poultry continued at the retail LPM. Before these measures could be implemented, agreement on the changes and financial assistance to implement the changes had to be agreed with the trade in Hong Kong and with mainland authorities responsible for the farms supplying poultry to Hong Kong. Trade in live chickens was restored within 7 weeks, during which time many of the measures listed above were implemented.
Surveillance systems were enhanced. Samples were collected from every batch of poultry sent to market and regular market surveillance was conducted on fecal swabs in selected markets to monitor for influenza viruses. Serological monitoring was also conducted on each batch of poultry to test for antibodies to H5 virus. Although occasional positive serological results were detected, no viruses were ever isolated from any seropositive consignments of chickens. However, it is possible that the existence of the serological surveillance program served as an additional incentive for the supplying farms to ensure that their farms were free of H5N1 infection.
Regular meetings between management of the departments and policy bureau responsible for health, agriculture, markets, and the environment were held. Similar meetings were arranged for operational staff. An “Avian flu task force” was set up bringing together relevant expertise from the Departments of Health, Agriculture, Fisheries and Conservation, Food & Environmental Hygiene, the Hospital Authority, and Universities and met whenever unusual findings arose. The task force was subsequently replaced by a Scientific Committee on Zoonotic and Emerging Diseases that continues to provide advice to the Department of Health on avian influenza and other emerging infectious diseases. Thus, many of the factors required for cooperation toward a “One Health” approach (Anholdt et al. 2012) were in place in Hong Kong, including working toward a common goal.
Genetic Characterization of the Virus and Other Studies
Genetic characterization of avian influenza viruses isolated from poultry and humans in 1997 provided crucial clues to the origin of the 1997 H5N1 HPAI virus. H5N1 viruses from humans and poultry were very similar and retained avian-characteristics in all 8 gene segments with no evidence of reassortment with human or swine influenza viruses. This indicated direct avian-to-human transmission. The virus retained signatures in the hemagglutinin that indicated a preference for binding alpha 2–3 linked sialosides found on avian cells (Claas et al. 1998; Suarez et al. 1998; Subbarao et al. 1998). The Hong Kong 1997 H5N1 virus had acquired its hemagglutinin from A/goose/Guangdong/1/96 (gs/Gd/96)-like HPAI H5N1 viruses first isolated from an outbreak of disease in geese in 1996 in Guangdong, (Xu et al. 1999). The other gene segments of the H5N1/97 viruses were probably derived from low pathogenic H9N2 (G1-lineage) or H6N1 viruses that were common in quail and other game-poultry species. This particular gene constellation has not been seen since. Thus, H5N1/1997 may have arisen within Hong Kong or in the markets or farms supplying Hong Kong through reassortment between gs/Gd/96-like and H9N2 or H6N1 viruses (Guan et al. 1999; Hoffmann et al. 2000).
Continued Active Surveillance in Live Poultry Markets
Continued active virological surveillance of imported poultry provided evidence that other influenza virus subtypes, especially H9N2 and H6N1 viruses were being isolated frequently from samples from poultry in markets. In addition, from 1999 there was occasional detection of gs/Gd/96-like HPAI H5N1 viruses from geese directly imported to the goose and duck slaughterhouse (Cauthen et al. 2000). This indicated that gs/Gd/96-like HPAI H5N1 viruses continued to circulate in geese in mainland China. In 2000, further viruses were isolated from geese and ducks imported into Hong Kong with evidence of novel reassortants in ducks, indicating that this virus was undergoing rapid genetic change in ducks (Guan et al. 2002a). This served as a warning that H5N1 viruses were on the move, both geographically and genetically. This information was shared freely with and between the parties involved in control of avian influenza as soon as it became available and proved to be extremely valuable. Hong Kong’s live terrestrial poultry markets had remained free of H5N1 since trade resumed in 1998, even though active systematic virologic surveillance within Hong Kong’s live poultry markets continued. However, the increasing nature of the challenge was apparent.
Thus, it was not unexpected when H5N1 HPAI virus was detected in Hong Kong’s LPM in 2001. The routine active surveillance system which involves monthly sampling of fecal droppings in LPM led to the detection of H5N1 HPAI viruses from apparently healthy birds. Subsequently, die-offs were noted in poultry within these markets, leading to a second cull of 1.3 million poultry across all LPM in Hong Kong, predominantly birds on farms that could not be sold due to the closure of markets (Sims et al. 2003a, b). The gs/Gd/96-like H5N1 virus had been replaced by a number of novel HPAI H5N1 reassortants, which were all distinct from the H5N1/97 virus (Guan et al. 2002b). It is important to note that without the active virologic surveillance of apparently healthy poultry in these markets, the initial incursion of H5N1 viruses would have probably gone unnoticed, possibly for many months, leading to reexposure of the human population to significant zoonotic risk. The viruses isolated were undergoing rapid evolution with viruses from clade 0, clade1, clade3, clade 4, and clade 8 all detected during the period from 2001 to 2003 along with multiple genotypes.
Following this event, a number of additional measures were introduced into the poultry marketing system. This included a compulsory rest day in all retail poultry markets that coincided with a rest day in the wholesale poultry market. The evening prior to the rest day, all live poultry were sold or slaughtered and the market was empty of poultry during the rest day, when the markets were cleaned and disinfected. Subsequent studies, using the endemic poultry H9N2 subtype viruses as an indicator, demonstrated the impact of the rest day in reducing virus isolation rates and implied that avian influenza viruses entered markets infrequently, but then amplified and were maintained in these retail LPMs (Kung et al. 2003). Continued introduction of fresh poultry into these markets, a proportion of them remaining in the markets for a day or two before slaughter allowed time for virus transmission within these markets without overt large-scale die-offs in the birds (because few of them stayed for long enough for the full-blown disease to manifest. Heterosubtypic immunity from prior exposure to H9N2 viruses may also have played a role in masking severe disease in HPAI H5N1 infected birds (Seo and Webster 2001)
When the novel reassortant viruses were detected in the markets in 2001, it was not known whether they also had capacity to infect humans. Quail were the major source of G1-lineage H9N2 viruses in poultry markets. In order to reduce the opportunity for reassortment between H5N1 and G1-lineage H9N2 viruses leading to the emergence of 1997-like H5N1 viruses, live quail were segregated from other poultry at all levels of the production and marketing chain. In practice, this meant that live quail were not available within Hong Kong’s retail LPM. Freshly killed quail carcasses were imported for sale.
However, H5N1 virus was again detected in poultry markets in January 2002 and on a local chicken farm in February 2002. The farm was promptly depopulated. This farm was located in an area with a high density of poultry farms and those in the vicinity were quarantined and later depopulated. By mid February, 17 of 18 farms in the vicinity were affected and depopulated. There was a diversity of H5N1 virus genotypes (n = 6) in the markets, and a smaller diversity of viruses affecting the farms (3 genotypes). Epidemiological investigations suggested a small number of introductions to farms followed by limited lateral spread (Sims et al. 2003b; Kung et al. 2007). A case—control study was conducted to identify farm practices and other factors associated with increased risk of infection. This analysis identified that direct sales of poultry from the farms to the retail markets (by-passing the wholesale market, where cages returning to farms were thoroughly washed) was associated with increased risk of farms acquiring infection (Fig. 1) (Kung et al. 2007). The retail LPM were serving as an amplifier of virus as well as disseminating virus back to farms. Thus, while LPM were a “dead-end” for the poultry, it was not a “dead-end” for the virus. These were concepts that were to be later validated in some markets and traders’ yards in other countries in Asia (Indriani et al. 2010; Samaan et al. 2011).
Fig. 1.
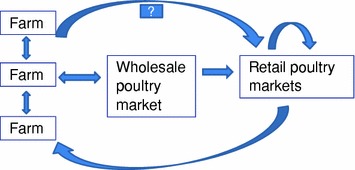
Probable routes of virus transmission through the poultry marketing chain, Hong Kong, 2002 (based on references Kung et al. 2003, 2007; Sims et al. 2003a, b)
The experience of H5N1 control measures together with sustained active surveillance in Hong Kong up to 2002 showed that the interventions in 1997 and 2001 successfully eliminated virus within Hong Kong. However, H5N1 viruses continued to circulate and were changing genetically in the wider region.
In late February 2002, infection was detected in chickens on a farm in an area with a high poultry density that was previously unaffected. Within 3 weeks, three other farms in this area had also been confirmed as infected. All infected premises were eventually depopulated and the remaining 21 farms in the area were included in a vaccination trial used as an adjunct to the bio-security, market interventions, and depopulations relied on hitherto. The commercial vaccine contained a killed H5N2 antigen with adjuvant. Each vaccinated farm had nonvaccinated sentinel chickens tagged and monitored as indicators of introduction of HPAI viruses into the farm. No further cases were detected on vaccinated farms. Vaccination trials conducted at the time demonstrated that vaccinated poultry exposed to a H5N1 virus were protected from clinical disease and shed much less virus than unvaccinated controls (Ellis et al. 2004a, 2006).
In December 2002, separate H5N1 outbreaks occurred in captive aquatic waterfowl in two parks (Penfold Park; Kowloon Park) in Hong Kong affecting geese, ducks, swans, captive Greater Flamingos, and wild birds (Little Egret, Grey Heron, Black headed gull) (Ellis et al. 2004b). This was the first report of this Asian lineage H5N1 HPAI virus, causing severe and lethal disease in ducks and also affecting wild birds (Sturm-Ramirez et al. 2004). HPAI was previously believed not to cause severe disease in ducks and experimental studies with the 1997 virus had shown this to be the case (Perkins and Swayne 2002). Kowloon Park was home to a range of rare and protected species. The control measures implemented included strict isolation, culling, increased sanitation, and also vaccination, were successful in containing that outbreak, although with significant loss of valuable bird species. Around this time, H5N1 HPAI virus was also detected in a few LPM and in a poultry farm. The vaccination program for chicken farms was enlarged to cover some other farms, but a subsequent outbreak occurred in several adjacent unvaccinated farms. Vaccination was successfully used in the face of an outbreak to control the disease in unaffected poultry houses (Ellis et al. 2004a).
In February 2003, while in the throes of the incipient emergence of SARS, two cases of human H5N1 influenza were detected in family members recently returned from Fujian province in mainland China, the infection being externally acquired. The viruses causing these human infections had the H5 hemagglutinin derived from the gs/Gd/96-like H5N1 lineage that gave rise to the outbreak in Hong Kong in 1997 (gs/GD/96 had a clade 0 hemagglutinin gene, whereas the HA gene of the human cases in 2003 belonged to Clade 1), but had different virus internal genes (genotype Z +), indicating that H5N1 virus genotypes other than the 1997-like viruses could infect humans.
By December 2003, vaccination had been extended to include all poultry farms in Hong Kong, and of all farms from mainland China supplying live poultry to Hong Kong. Some of the other long-term precautionary measures included requiring poultry farms to keep proper farm management records, enhance cleaning and disinfection facilities, segregating the functions relating to the rearing of breeder flocks and broiler flocks, and the installation of metal nets to prevent small birds from entering farm sheds. These measures were to serve Hong Kong well in preventing new incursions of virus with no H5N1 viruses detected in intensive farm and market surveillance from December 2003 until June 2008, despite continuing circulation of H5N1 HPAI viruses in the broader region.
Post 2004: Widespread Reports of H5N1 Outbreaks in South-East Asia and Beyond
In the aftermath of the SARS outbreak, there was enhanced surveillance and diagnosis of severe “atypical” pneumonia in many Asian countries. These led to diagnosis of patients with influenza A(H5N1) in mainland China (recognized retrospectively), Vietnam, and Thailand in late 2003 and early 2004, rapidly followed by the recognition and reporting of H5N1 HPAI in poultry in Cambodia, mainland China, Indonesia, Japan, Laos, Malaysia, and Republic of Korea, caused by clade 1, clade 2.5, and clade 2.1 genotype Z HPAI H5N1 viruses (Li et al. 2004). Following an outbreak of disease caused by H5N1 HPAI virus in migratory wild birds in Qinghai lake in April–May 2005 caused by a clade 2.2 virus (Chen et al. 2005), there was further spread of that virus to Central and South Asia, the middle-east, Africa, and Europe. By end of 2006, 115 human H5N1 cases had been cumulatively reported, from Azerbaijan, Cambodia, mainland China, Djibouti, Egypt, Indonesia, Iraq, Thailand, Turkey, and Vietnam (WHO 2012). Most notably, during this period, Hong Kong remained spared of this major regional and transcontinental spread of HPAI H5N1, almost certainly due to the preemptive interventions that protected both poultry and humans.
Hong Kong had also initiated active surveillance in live wild birds at the Mai Po nature reserve and wetland park, and of dead wild birds detected anywhere in Hong Kong. Since 2005, the Department of Agriculture, Fisheries and Conservation provided a round-the-clock service for examination of wild birds collected dead or in a weak state; for example, 8,700 wild bird carcasses were tested in 2007. Up to 2012, more than 100 carcasses have been found to be infected with H5N1 virus. While a range of low pathogenic influenza viruses of diverse subtypes were detected in live wild birds in Mai Po nature reserve, no H5N1 HPAI viruses were detected in live wild birds (Leung 2011). However, the program of dead wild bird surveillance repeatedly led to the detection of H5N1 HPAI viruses in a range of wild aquatic birds (e.g. pond herons, grey herons, egrets), birds of prey (Peregrine falcons, Buzzards), and passerine birds (Smith et al. 2009; Ellis et al. 2009). Detection of infection tended to be commoner in the winter months. While infection of migratory birds and birds of prey may be understandable in the context of ongoing infection in poultry in the wider region, the source of infection of passerine birds was more of an enigma. “Bird-release” to acquire religious merit is a religious and cultural activity common in Hong Kong and other Asian countries. This demand has led to a commercialized and large-scale trapping and transborder trading activity and it was estimated that 680,000–1,050,000 birds were imported legally (and sometimes illegally) into Hong Kong to service this trade. Although direct proof is lacking, it is possible that these birds acquired infection during this trade and importation, and bird-release of infected birds may explain the detection of some H5N1 infected dead birds (Chan 2006). Clustering of wild bird cases in the area around the pet bird market in Hong Kong in 2006 and 2007 provides further evidence to support this proposition during those 2 years (Anonymous 2006). However, the pattern of detection changed in 2008 onward with no cases detected near this market (Anonymous 2006 ).
In mid 2008, the active surveillance of apparently healthy poultry in LPM led to the detection of HPAI H5N1 viruses in multiple markets. The source of the virus was not determined, but around 6,000 birds in retail poultry markets were culled. In December 2008, a poultry farm in Hong Kong was found to be infected, the alert being raised by the death of chickens. These clade 2.3.4 viruses were found to be antigenic variants, which evade the immunity conferred by the contemporary H5 vaccine (Leung 2011). However, the majority of HPAI H51 viruses being detected in wild birds in Hong Kong in recent years were clade 2.3.2.1 viruses. These findings demonstrated the importance of continually monitoring and updating vaccine antigens to match circulating strains, a process that is now well developed in mainland China.
Further interventions in the LPM included the ban on holding poultry overnight. This has led to a further dramatic reduction of avian influenza infection rates (using H9N2 as an indicator) in LPM in Hong Kong (Leung et al. 2012), but has also resulted in a marked contraction in the number of poultry traders.
Two further human cases were detected in Hong Kong in November 2010 and May 2012, both acquired from mainland China. As with all previous human cases, the virus appeared to be avian in origin.
Conclusions
Attempts to contain HPAI H5N1 influenza (and other infectious disease outbreaks) in Hong Kong over the past 15 years have demonstrated the importance of adopting a One Health approach (Table 1) (Anholdt et al. 2012).
Table 1.
.
| Example | Human health | Animal health | Environment | Outcome |
|---|---|---|---|---|
| Studies on influenza viruses in animals prior to 1997 | - Possible serological evidence of widespread infection | - Numerous influenza viruses detected and characterized in poultry and pigs | - Farming systems promote cross species infection | - Recognition of the likely importance of animals and animal viruses and rural environment in the genesis of pandemic viruses |
| Avian influenza 1997 | - Fatal zoonotic disease with pandemic potential | - Detection of disease and virus in poultry in markets and farms | - Role of the poultry production and marketing systems in the transmission and persistence of virus |
- Changes to production and marketing systems. - Enhanced collaboration and coordination between human and animal health. - Importance of molecular epidemiology and sharing of information. |
| Avian influenza 2001–now |
- Probable imported cases of influenza A(H5N1) and multiple human cases in region. - Detection of cases of H9N2 infection |
- Role of domestic ducks and wild birds in spread of virus and persistence of H5N1. - Rapid evolution and reassortment in avian influenza viruses |
- Wild bird trade, migratory patterns, duck farming systems demonstrated as contributing factors in virus transmission | - Enhanced interactions with wild bird ecologists |
| SARS 2003 | - Novel severe infectious disease disseminated via health care systems and global travel. |
- Likely role of demand for wild-animal meat, leading to large markets and cross-boundary trade to service this demand. - Animals within these “wild” animal markets act as amplifiers and source of zoonotic transmission. |
- Environmental issues associated with disease transmission in community outbreak (Amoy Gardens) and in hospitals. - Role of bats as likely reservoir of precursor virus. |
- Importance of pandemic preparedness and interdisciplinary and transboundary coordination and cooperation |
| Pandemic H1N1 2009 | - Rapid global spread of a novel virus with particularly high infection attack rates in children. | - On-going surveillance of pigs provided insights into pandemic emergence. | - Reverse zoonosis of virus from humans to pigs leading to a global perturbation of swine influenza ecology. |
- Asia is not the only epicenter for pandemic emergence. - Pandemics may emerge from influenza virus subtypes (e.g. H1) that are already endemic in humans. - Importance of pandemic preparedness and review of effectiveness of measures |
It is noteworthy that there were no human cases of H5N1 influenza acquired within Hong Kong since 1997, and only three cases have been detected in commercial poultry on farms or in markets since the end of 2003, despite the on-going threat of infection in the region evident from outbreaks in mainland China and elsewhere. This outcome can almost certainly be attributed to the measures taken to prevent infection in poultry and minimize exposure of the human population through a series of evidence-based interventions. Building these measures was an interactive process based on experiences, experiments, and epidemiological investigations from and after 1997. Many of the lessons learned in Hong Kong have been applied elsewhere, when H5N1 influenza viruses emerged in other countries in Asia, Africa, and Europe from 2003 onward.
The main objective of the control and preventive programs for HPAI H5N1 influenza was to prevent human infections with these viruses, thereby reducing the risk of mammalian adaptation that would allow human-to-human transmission.
It is interesting to speculate why conditions in markets and farms were poor in 1997. The most likely explanation is that there had never before been an outbreak of a zoonotic disease associated with poultry and no outbreaks of HPAI had occurred before in Hong Kong. Therefore, traders and farmers were still making a profit even if some disease occurred in poultry. There were no signals or incentives to change, a situation that changed dramatically in 1997. These observations match those in other countries that have experienced outbreaks of HPAI. As with Hong Kong, unless there are clear incentives to make changes and capital available for investment in improved production systems the changes are not made.
The “One-Health” approach was also applied to other emerging diseases such as SARS, where the identification of the game-animal markets serving the restaurant trade in Guangdong as a likely amplifier and source for interspecies transmission to humans was very reminiscent of the role of LPM in relation to avian flu H5N1 (Guan et al. 2003). This evidence convinced relevant authorities in Guangdong to control these markets and to ban the sale of civet cats within them, very likely aborting a reemergence of SARS (Wang et al. 2005). The search for the animal reservoir led to the identification of bats as the source of the precursor of SARS CoV, and also to the recognition that bats may serve as a reservoir for a number of virus families including the coronaviruses (Vijaykrishna et al. 2007).
Implementation of “One Health” approaches, though eminently desirable, is not always cost free. The focus on eliminating risk of human infection had significant costs on the poultry sector in Hong Kong; any benefits to the poultry sector from the actions taken against H5N1 HPAI were secondary. In fact, a significant proportion of market traders and farmers chose not to continue to rear or sell poultry, taking ex gratia packages from government to cease business as measures were introduced progressively. In 1997 there were some 160 chicken farms and over 800 separate stalls selling live poultry, while in 2012 there are only 30 farms and 132 market stalls (Legislative Council Report 2012). The volume of trade in live poultry has fallen from over 100,000 birds per day to approximately 16,000/day. While all the measures taken in Hong Kong may not be universally applicable, some of them may be (Indriani et al. 2010; Samaan et al. 2011).
There is now much greater trade in chilled and frozen poultry, which potentially provides a new set of challenges resulting from reliance on intensively reared poultry and centralized slaughter (Sims 2011).
Surveillance of domestic livestock to understand emergence of potential threats to human health can also have adverse impacts on the economics of the domestic livestock trade. This is well illustrated by the dearth of surveillance data on influenza viruses of swine, even from developed countries, prior to and in the aftermath of the 2009 pandemic. Although the 2009 pandemic did not arise in Asia, the long-term systematic surveillance of pigs in Hong Kong abattoirs, which de-links results of surveillance from specific farms, has provided a unique understanding of the events leading to the emergence of that pandemic (Vijaykrishna et al. 2011; Yen et al. 2011). If a “One Health” approach is to provide the intended benefits, it needs to be implemented with sensitivity and care, involving and considering the needs of all stakeholders, so that the economics of food security and food supply are not compromised.
Acknowledgments
We acknowledge research funding from the National Institutes of Health (NIAID contract HHSN266200700005C) and the Area of Excellence Scheme of the University Grants Committee (AoE/M-12/06) Hong Kong SAR Government to JSMP. We acknowledge the collaboration of many colleagues at the Agriculture, Fisheries and Conservation Department, and the Food and Environmental Hygiene Departments of the Hong Kong Special Administrative Region and at The University of Hong Kong.
Contributor Information
John S. Mackenzie, Email: j.mackenzie@curtin.edu.au
Martyn Jeggo, Phone: 409166752, Email: jeggo.martyn@gmail.com.
Peter Daszak, Email: daszak@ecohealthalliance.org.
Juergen A. Richt, Phone: +1785 532 4408, Email: jricht@vet.k-state.edu
Malik Peiris, Phone: (852)-2816-8404, Email: malik@hku.hk.
References
- American Veterinary Medical Association (2008) One health initiative task force. One health: a new professional imperative. AMVA, Schaumburg, Available. http://www.avma.org/onehealth/onehealth_final.pdf
- Anholdt RM, Stephen C, Copes R. Strategies for collaboration in the interdisciplinary field of emerging infectious diseases. Zoonoses Public Health. 2012;59:229–240. doi: 10.1111/j.1863-2378.2011.01449.x. [DOI] [PubMed] [Google Scholar]
- Anonymous (2006) Location of H5N1 infected wild-birds found in Hong Kong in 2006. http://www.afcd.gov.hk/english/quarantine/qua_vetlab/qua_vetlab_ndr/qua_vetlab_ndr_adr/qua_vetlab_oth_adr/files/common/posth5n128_03_06.pdf. Accessed 3 July 2012
- Anonymous (2006) Location of H5N1 infected wild-birds found in Hong Kong in 2008. http://www.afcd.gov.hk/tc_chi/whatsnew/what_qua/files/common/h5n1_2008.pdf
- Brandt AM. A reader’s guide to 200 years of the New England journal of medicine. N Engl J Med. 2012;366:1–7. doi: 10.1056/NEJMp1112812. [DOI] [PubMed] [Google Scholar]
- Bridges CB, Lim W, Hu-Primmer J, Sims L, Fukuda K, Mak KH, Rowe T, Thompson WW, Conn L, Lu X, Cox NJ, Katz JM. Risk of influenza A (H5N1) infection among poultry workers, Hong Kong, 1997–1998. J Infect Dis. 2002;185(8):1005–1010. doi: 10.1086/340044. [DOI] [PubMed] [Google Scholar]
- Buxton Bridges C, Katz JM, Seto WH, Chan PK, Tsang D, Ho W, Mak KH, Lim W, Tam JS, Clarke M, Williams SG, Mounts AW, Bresee JS, Conn LA, Rowe T, Hu-Primmer J, Abernathy RA, Lu X, Cox NJ, Fukuda K. Risk of influenza A (H5N1) infection among health care workers exposed to patients with influenza A (H5N1), Hong Kong. J Infect Dis. 2000;181:344–348. doi: 10.1086/315213. [DOI] [PubMed] [Google Scholar]
- Cauthen AN, Swayne DE, Schultz-Cherry S, Perdue ML, Suarez DL. Continued circulation in China of highly pathogenic avian influenza viruses encoding the hemagglutinin gene associated with the 1997 H5N1 outbreak in poultry and humans. J Virol. 2000;74:6592–6599. doi: 10.1128/JVI.74.14.6592-6599.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan SW (2006) Religious release of birds in Hong Kong. M Phil Thesis, University of Hong Kong. http://hub.hku.hk/handle/10722/51015
- Chen H, Smith GJ, Zhang SY, Qin K, Wang J, Li KS, Webster RG, Peiris JSM, Guan Y. Avian flu: H5N1 virus outbreak in migratory waterfowl. Nature. 2005;436:191–192. doi: 10.1038/nature03974. [DOI] [PubMed] [Google Scholar]
- Chun JWH. Influenza, including its infection among pigs. Natl Med J China. 1919;5:34–44. doi: 10.1186/1749-8546-5-34. [DOI] [Google Scholar]
- Claas EC, Osterhaus AD, van Beek R, De Jong JC, Rimmelzwaan GF, Senne DA, Krauss S, Shortridge KF, Webster RG. Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet. 1998;351:472–477. doi: 10.1016/S0140-6736(97)11212-0. [DOI] [PubMed] [Google Scholar]
- Ellis TM, Leung CY, Chow MK, Bissett LA, Wong W, Guan Y, Malik Peiris JS. Vaccination of chickens against H5N1 avian influenza in the face of an outbreak interrupts virus transmission. Avian Pathol. 2004;33:405–412. doi: 10.1080/03079450410001724012. [DOI] [PubMed] [Google Scholar]
- Ellis TM, Bousfield RB, Bissett L, Dyrting KC, Luk GSM, Tsim ST, Sturm-Ramirez K, Webster RG, Peiris JSM. Investigation of outbreaks of highly pathogenic H5N1 avian influenza in waterfowl and wild birds in Hong Kong in late 2002. Avian Pathol. 2004;33:492–505. doi: 10.1080/03079450400003601. [DOI] [PubMed] [Google Scholar]
- Ellis TM, Sims LD, Wong HK, Wong CW, Dyrting KC, Chow KW, Leung C, Peiris JSM. Use of avian influenza vaccination in Hong Kong. Dev Biol (Basel) 2006;124:133–143. [PubMed] [Google Scholar]
- Ellis TM, Dyrting KC, Wong CW, Chadwick B, Chan C, Chiang M, Li C, Li P, Smith GJD, Guan Y, Peiris JSM. Analysis of H5N1 avian influenza infections from wild bird surveillance in Hong Kong from January 2006 to October 2007. Avian Pathol. 2009;38:107–119. doi: 10.1080/03079450902751855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guan Y, Shortridge KF, Krauss S, Webster RG. Molecular characterization of H9N2 influenza viruses: were they the donors of the “internal” genes of H5N1 viruses in Hong Kong? Proc Natl Acad Sci U S A. 1999;96:9363–9367. doi: 10.1073/pnas.96.16.9363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guan Y, Peiris M, Kong KF, Dyrting KC, Ellis TM, Sit T, Zhang LJ, Shortridge KF. H5N1 influenza viruses isolated from geese in Southeastern China: evidence for genetic reassortment and interspecies transmission to ducks. Virology. 2002;292:16–23. doi: 10.1006/viro.2001.1207. [DOI] [PubMed] [Google Scholar]
- Guan Y, Peiris JS, Lipatov AS, Ellis TM, Dyrting KC, Krauss S, Zhang LJ, Webster RG, Shortridge KF. Emergence of multiple genotypes of H5N1 avian influenza viruses in Hong Kong SAR. Proc Natl Acad Sci U S A. 2002;99:8950–8955. doi: 10.1073/pnas.132268999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guan Y, Zheng BJ, He YQ, Liu XL, Zhuang ZX, Cheung CL, Luo SW, Li PH, Zhang LJ, Guan YJ, Butt KM, Wong KL, Chan KW, Lim W, Shortridge KF, Yuen KY, Peiris JSM, Poon LLM. Isolation and characterization of viruses related to the SARS coronavirus from animals in Southern China. Science. 2003;302:276–278. doi: 10.1126/science.1087139. [DOI] [PubMed] [Google Scholar]
- Hoffmann E, Stech J, Leneva I, Krauss S, Scholtissek C, Chin PS, Peiris M, Shortridge KF, Webster RG. Characterization of the influenza a virus gene pool in avian species in Southern China: was H6N1 a derivative or a precursor of H5N1? J Virol. 2000;74:6309–6315. doi: 10.1128/JVI.74.14.6309-6315.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Indriani R, Samaan G, Gultom A, Loth L, Indryani S, Adjid R, Dharmayanti NL, Weaver J, Mumford E, Lokuge K, Kelly PM, Darminto Environmental sampling for avian influenza virus A (H5N1) in live-bird markets, Indonesia. Emerg Infect Dis. 2010;16:1889–1895. doi: 10.3201/eid1612.100402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Legislative Council Report, Hong Kong SAR (2012) Review of avian influenza risk in Hong Kong and latest developments on avian influenza vaccines for local chicken farms. http://www.legco.gov.hk/yr11-12/english/sec/library/1112in32-e.pdf. Accessed 3 July 2012
- Leung CYH (2011) Ecology, epidemiology and immunology of avian influenza virus. PhD Thesis, University of Hong Kong. http://hub.hku.hk/handle/10722/137060
- Leung YHC, Lau EHY, Zhang LJ, Guan Y, Cowling BJ, Peiris JSM (2012) Avian influenza and ban on overnight storage in live poultry markets, Hong Kong. Emerg Infect Dis in press [DOI] [PMC free article] [PubMed]
- Li KS, Guan Y, Wang J, Smith GJ, Xu KM, Duan L, Rahardjo AP, Puthavathana P, Buranathai C, Nguyen TD, Estoepangestie AT, Chaisingh A, Auewarakul P, Long HT, Hanh NT, Webby RJ, Poon LL, Chen H, Shortridge KF, Yuen KY, Webster RG, Peiris JSM. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in Eastern Asia. Nature. 2004;430:209–213. doi: 10.1038/nature02746. [DOI] [PubMed] [Google Scholar]
- Kung NY, Guan Y, Perkins NR, Bissett L, Ellis T, Sims L, Morris RS, Shortridge KF, Peiris JSM. The impact of a monthly rest day on avian influenza virus isolation rates in retail live poultry markets in Hong Kong. Avian Dis. 2003;47(Suppl):1037–1041. doi: 10.1637/0005-2086-47.s3.1037. [DOI] [PubMed] [Google Scholar]
- Kung NY, Morris RS, Perkins NR, Sims LD, Ellis TM, Bissett L, Chow M, Shortridge KF, Guan Y, Peiris MJ. Risk for infection with highly pathogenic influenza a virus (H5N1) in chickens, Hong Kong, 2002. Emerg Infect Dis. 2007;13:412–418. doi: 10.3201/eid1303.060365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mounts AW, Kwong H, Izurieta HS, Ho Y, Au T, Lee M, Buxton Bridges C, Williams SW, Mak KH, Katz JM, Thompson WW, Cox NJ, Fukuda K. Case-control study of risk factors for avian influenza A (H5N1) disease, Hong Kong, 1997. J Infect Dis. 1999;180:505–508. doi: 10.1086/314903. [DOI] [PubMed] [Google Scholar]
- Perkins LE, Swayne DE. Pathogenicity of a Hong Kong-origin H5N1 highly pathogenic avian influenza virus for emus, geese, ducks, and pigeons. Avian Dis. 2002;46:53–63. doi: 10.1637/0005-2086(2002)046[0053:POAHKO]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Samaan G, Gultom A, Indriani R, Lokuge K, Kelly PM. Critical control points for avian influenza A H5N1 in live bird markets in low resource settings. Prev Vet Med. 2011;100:71–78. doi: 10.1016/j.prevetmed.2011.03.003. [DOI] [PubMed] [Google Scholar]
- Seo SH, Webster RG. Cross-reactive, cell-mediated immunity and protection of chickens from lethal H5N1 influenza virus infection in Hong Kong poultry markets. J Virol. 2001;75:2516–2525. doi: 10.1128/JVI.75.6.2516-2525.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scholtissek C, Bürger H, Kistner O, Shortridge KF. The nucleoprotein as a possible major factor in determining host specificity of influenza H3N2 viruses. Virology. 1985;147:287–294. doi: 10.1016/0042-6822(85)90131-X. [DOI] [PubMed] [Google Scholar]
- Shortridge KF, Webster RG, Butterfield WK, Campbell CH. Persistence of Hong Kong influenza virus variants in pigs. Science. 1977;196:1454–1455. doi: 10.1126/science.867041. [DOI] [PubMed] [Google Scholar]
- Shortridge KF. Avian influenza a viruses of southern China and Hong Kong: ecological aspects and implications for man. Bull World Health Organ. 1982;60:129–135. [PMC free article] [PubMed] [Google Scholar]
- Shortridge KF, Stuart-Harris CH. An influenza epicentre? Lancet. 1982;2:812–813. doi: 10.1016/S0140-6736(82)92693-9. [DOI] [PubMed] [Google Scholar]
- Shortridge KF. Pandemic influenza: a zoonosis? Semin Respir Infect. 1992;7:11–25. [PubMed] [Google Scholar]
- Shortridge KF. Poultry and the influenza H5N1 outbreak in Hong Kong, 1997: abridged chronology and virus isolation. Vaccine. 1999;17(Suppl 1):S26–S29. doi: 10.1016/S0264-410X(99)00102-4. [DOI] [PubMed] [Google Scholar]
- Sims LD, Ellis TM, Liu KK, Dyrting K, Wong H, Peiris M, Guan Y, Shortridge KF. Avian influenza in Hong Kong 1997–2002. Avian Dis. 2003;47(3 Suppl):832–838. doi: 10.1637/0005-2086-47.s3.832. [DOI] [PubMed] [Google Scholar]
- Sims LD, Guan Y, Ellis TM, Liu KK, Dyrting K, Wong H, Kung NY, Shortridge KF, Peiris M (2003b) An update on avian influenza in Hong Kong 2002. Avian Dis 47(3 Suppl):1083–1086 [DOI] [PubMed]
- Sims LD. Strategies for controlling animal influenza and implications for human health. Influenza Other Respir Viruses. 2011;5(Suppl 1):2–53. [Google Scholar]
- Smith GJ, Vijaykrishna D, Ellis TM, Dyrting KC, Leung YH, Bahl J, Wong CW, Kai H, Chow MK, Duan L, Chan AS, Zhang LJ, Chen H, Luk GS, Peiris JSM, Guan Y. Characterization of avian influenza viruses A (H5N1) from wild birds, Hong Kong, 2004–2008. Emerg Infect Dis. 2009;15:402–407. doi: 10.3201/eid1503.081190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sturm-Ramirez KM, Ellis T, Bousfield B, Bissett L, Dyrting K, Rehg JE, Poon L, Guan Y, Peiris M, Webster RG. Reemerging H5N1 influenza viruses in Hong Kong in 2002 are highly pathogenic to ducks. J Virol. 2004;78:4892–4901. doi: 10.1128/JVI.78.9.4892-4901.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suarez DL, Perdue ML, Cox N, Rowe T, Bender C, Huang J, Swayne DE. Comparisons of highly virulent H5N1 influenza a viruses isolated from humans and chickens from Hong Kong. J Virol. 1998;72:6678–6688. doi: 10.1128/jvi.72.8.6678-6688.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subbarao K, Klimov A, Katz J, Regnery H, Lim W, Hall H, Perdue M, Swayne D, Bender C, Huang J, Hemphill M, Rowe T, Shaw M, Xu X, Fukuda K, Cox N. Characterization of an avian influenza A (H5N1) virus isolated from a child with a fatal respiratory illness. Science. 1998;279:393–396. doi: 10.1126/science.279.5349.393. [DOI] [PubMed] [Google Scholar]
- Vijaykrishna D, Smith GJ, Zhang JX, Peiris JS, Chen H, Guan Y. Evolutionary insights into the ecology of coronaviruses. J Virol. 2007;81:4012–4020. doi: 10.1128/JVI.02605-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vijaykrishna D, Smith GJ, Pybus OG, Zhu H, Bhatt S, Poon LL, Riley S, Bahl J, Ma SK, Cheung CL, Perera RA, Chen H, Shortridge KF, Webby RJ, Webster RG, Guan Y, Peiris JSM. Long-term evolution and transmission dynamics of swine influenza a virus. Nature. 2011;473:519–522. doi: 10.1038/nature10004. [DOI] [PubMed] [Google Scholar]
- Wang M, Yan M, Xu H, Liang W, Kan B, Zheng B, Chen H, Zheng H, Xu Y, Zhang E, Wang H, Ye J, Li G, Li M, Cui Z, Liu YF, Guo RT, Liu XN, Zhan LH, Zhou DH, Zhao A, Hai R, Yu D, Guan Y, Xu J. SARS-CoV infection in a restaurant from palm civet. Emerg Infect Dis. 2005;11:1860–1865. doi: 10.3201/eid1112.041293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watkins KL, Shortridge KF, Powell DG. Equine influenza in Hong Kong. Vet Rec. 1993;132:144. doi: 10.1136/vr.132.6.144-a. [DOI] [PubMed] [Google Scholar]
- World Health Organization, Global Influenza Programme (2012) Cumulative number of confirmed human cases for avian influenza A(H5N1) reported to WHO, 2003–2012. http://www.who.int/influenza/human_animal_interface/EN_GIP_20120607CumulativeNumberH5N1cases.pdf. Accessed 21 June 2012
- Xu X, Subbarao Cox NJ, Guo Y. Genetic characterization of the pathogenic influenza A/Goose/Guangdong/1/96 (H5N1) virus: similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virology. 1999;261:15–19. doi: 10.1006/viro.1999.9820. [DOI] [PubMed] [Google Scholar]
- Yen HL, Liang CH, Wu CY, Forrest HL, Ferguson A, Choy KT, Jones J, Wong DD, Cheung PP, Hsu CH, Li OT, Yuen KM, Chan RW, Poon LL, Chan MC, Nicholls JM, Krauss S, Wong CH, Guan Y, Webster RG, Webby RJ, Peiris M. Hemagglutinin-neuraminidase balance confers respiratory-droplet transmissibility of the pandemic H1N1 influenza virus in ferrets. Proc Natl Acad Sci U S A. 2011;108:14264–14269. doi: 10.1073/pnas.1111000108. [DOI] [PMC free article] [PubMed] [Google Scholar]