

Abstract
Avian infectious bronchitis (IB) is caused by avian infectious bronchitis virus (IBV) belonging to Coronaviridae family. The disease is prevalent in all countries with almost 100% incidence rate. Chicken and commercially reared pheasant are the natural host for IBV. Virus causes respiratory diseases, poor weight gain, feed efficiency in broiler, damage to oviduct, and abnormal egg production in mature hens resulting in economic losses. IBV also replicates in tracheal and renal epithelial cells leading to prominent tracheal and kidney lesions. Virus undergoes spontaneous mutation leading to continual emergence of new variants. The effectiveness of immunization program is diminished because of poor cross-protection among the serotypes. Identification of circulating serotypes is important in controlling IBV infection. Toll-like receptor 3 (TLR3) and TLR21 are involved in early recognition of virus resulting in induction of inflammatory cytokines. Both humoral and cellular immune responses are important in the control of infection. Humoral immunity plays an important role in recovery and clearance of viral infection. IBV-specific cytotoxic T lymphocytes induce lysis of IBV-infected cells. Effective diagnostic tools are required at field level to identify different IBV variants. Embryonated chicken eggs are effective model for virus isolation. Identification by other specific methods like virus neutralization (VN), hemagglutination inhibition (HI), enzyme linked immunosorbent assay (ELISA), immunohistochemistry, or nucleic acid analysis or by electron microscopy is also indispensable. VN test in tracheal organ culture is the best method for antigenic typing for surveillance purposes. Continuous epidemiological surveillance, strict biosecurity measures, and vaccine effective against various serotypes are necessary for controlling IB in chickens.
Keywords: Infectious bronchitis virus, Chicken, Trachea, Kidney, Oviduct, Genetic variants, Humoral immunity, Cellular immunity, Virus neutralization, Hemagglutination inhibition, ELISA
Prologue
Avian infectious bronchitis (IB) is an acute, highly contagious upper respiratory tract disease affecting chicken of all ages with significant economic threat to the poultry industry. It is caused by infectious bronchitis virus (IBV), which belongs to the genus Gammacoronavirus, family Coronaviridae, in the order Nidovirales. The disease causes symptoms like watery eyes, mucus in the nares and trachea, sneezing, tracheal rales, and coughing. In layer birds, IB results in decreased egg production as well as quality, and some IBV variants cause interstitial nephritis. Infection with IBV causes ciliostasis in the trachea and predisposes to the secondary pathogens further complicating the disease. Morbidity is always 100%; however, mortality can vary from 0 to 82% depending upon the age of the birds, immune status, strain of the virus, and involvement of secondary pathogens. The virus possesses a single copy of positive-sense single-stranded RNA as its genome, which is approximately 27.6 kb in size. The genome undergoes genetic recombination and spontaneous mutation leading to the emergence of new variants having low level of cross-protection and complicates the control program by vaccination (Cavanagh and Gelb 2008; Jackwood et al. 2012). The IBV-like coronaviruses were also detected in pheasant, peafowl, turkey, teal, geese, pigeon, guinea fowl, partridge, penguins, and ducks (Dea and Tijssen 1989; Jonassen et al. 2005; Cavanagh 2005; Circella et al. 2007). The IBV is worldwide in distribution, and infection is acquired through inhalation or direct contact with infected birds or premises. Vertical transmission is not reported. Some of the IBV strains can also replicate in the kidneys, reproductive tract, and enteric surfaces resulting in the development of nephritis, misshapen eggs, and enteritis, respectively (Boltz et al. 2004). Induction of apoptosis in kidney cells is a major contributor of pathogenicity for virulent nephropathogenic IBV (Liu et al. 2017). The disease affects both egg and meat type of chickens. Respiratory disease is observed in the young growing chickens while reduced weight gain and feed efficiency in broiler chickens. IBV infection predisposes the broiler birds to secondary bacterial infection resulting in airsacculitis, pericarditis, and perihepatitis. In layers and breeder chickens, IBV replicates in the oviduct causing permanent damage or limited egg production. Pigment of the affected shell becomes paler and albumin is watery in consistency. The virus has not been reported to cause human infection.
The virus contains lipid envelope with a round to pleomorphic shape. The virus particles are approximately 120 nm in diameter with club-shaped spikes in their surface, which provide them a crown-like appearance (Jackwood and de Witt 2013). The genome encodes 4 structural proteins, 15 nonstructural proteins (nsp 2–16), and at least 5 lineage-specific accessory proteins (Dent et al. 2015). Structural proteins are spike (S), membrane (M), small membrane (E), and nucleoprotein (N). The spike protein is post-translationally cleaved into S1 and S2 subunits where the former is involved in virus entry to the host cell via sialic acid receptor and also harbors virus-neutralizing epitopes (Lai and Cavanagh 1997; Cavanagh 2007; Cavanagh et al. 1986; Koch et al. 1990; Niesters et al. 1987). S2 subunit anchors the spike protein, and the ectodomain of S2 protein also aids in the attachment of virus to the host cells (Promkuntod et al. 2013). M protein is the most abundant transmembrane protein that interacts with N and S glycoprotein for virion assembly (de Haan et al. 2000; Bande et al. 2015). E protein is scant and confined to the Golgi apparatus of the infected cell. This protein plays a key role in viral envelope formation, assembly, budding, ion channel activity, and apoptosis (Corse and Machamer 2003; Wilson et al. 2006). Ribonucleoprotein is formed by interaction of N protein to the genomic RNA of IBV and aids in transcription, replication, and translation of viral genome (Jayaram et al. 2005).
History
Schalk and Hawan in 1931 first reported a respiratory disease of chick that is characterized by gasping and listlessness in North Dakota, USA. Two years later, similar disease was reported by Bushnell and Brandly. Since the disease could be transmitted by Berkefeld filtered material, they recognized the causative agent as filterable virus and was confused with infectious laryngotracheitis virus (ILTV; Bushnell and Brandly 1933). The causative agent was established in 1936 by Beach and Schalm who proved that IBV was distinct from ILTV by cross-immunity studies. Although young chicks were found to be affected by IBV, later it was observed to be common in mature and laying hens. IBV was first cultivated in chick embryos by inoculating through chorioallantoic membrane (CAM). Unlike ILT, IBV did not show distinctive lesions on CAM and rather resulted in embryo death (Beaudette and Hudson 1937). Jungherr and coworkers first reported more than one serotypes of IBV. It has been shown that Massachusetts (Mass) isolate in 1941 and Connecticut (Conn) isolate in 1951 produced identical disease but did not cross-protect each other (Jungherr et al. 1956).
Transmission and Risk Factor
IBV is a highly infectious disease and birds show clinical signs within 36–48 h of infection. Infected chicks are the major source of virus contamination in the environment. Virus is transmitted through respiratory secretions and fecal droplet from infected chicken. Virus may also spread from one flock to another through inanimate objects including contaminated utensils, egg-packing materials, fertilizer, and farm visit. Vertical transmission of virus has not been reported. Trachea, kidney, and bursa of Fabricius show the presence of the virus as early as 24 h and through the seventh day following aerosol transmission (Cavanagh and Gelb 2008). The presence of virus in the cecal tonsils up to 14 weeks and from the feces up to 20 weeks correlates with the viral transmission through fecal shedding (Alexander and Gough 1977). Birds recover from infection within 14 days with a rise in antibody titer. It has been reported that IBV re-excreted from virus-negative birds for several weeks following recovery from inoculation at 1 day of age. Isolation of virus from tracheal and cloacal swabs at the time of lay and 19 weeks of age was reported (Jones and Ambali 1987). Establishment of latent infection and erratic shedding of virus in feces and respiratory secretion for a prolonged period of time are possible in a very few cases (Ignjatovic and Sapats 2000). Kidney may be one of the sites of persistent infection (Raj and Jones 1997). Vaccine viruses persist up to 163 days or more in various internal organs (Gay 2000). Prolonged or intermittent shedding of virus is considered as a potential risk factor for transmission of virus from one flock to another through contaminated equipment or persons.
Incidence and Prevalence
Spontaneous mutation of S1 subunit of S gene resulted in the emergence of various serotypes (Abro et al. 2012). IBV strains can be classified by genotyping- and serotyping-based methods. Viral serotypes can be distinguished by cross-neutralization test and genotypes by reverse transcriptase polymerase chain reaction (RT-PCR) techniques, bioinformatics, and gene sequencing technology. S1 gene of IBV variants can be amplified and the sequences are analyzed by Basic Local Alignment Search Tool (BLAST; Posada and Crandall 2001). IBV strains like Mass, 4/91, D274, and QX-like are reported worldwide. Some of the IBV strains are more prevalent in particular geographical location. M41, Arkansas (Ark) and Conn are common in the United States, while 4/91 (793/B, CR88) and D274 are predominant strains in Europe. In recent times, the Chinese QX variants have caused outbreaks in Europe, Asia, the Middle East, and Africa (Bande et al. 2015).
Predominant strains of IBV isolated in China are QX-like strains (Ma et al. 2012). IBV QX strain-affected chickens showed proventriculus swelling which was first documented in 1996 (Yudong et al. 1998). The proportion of QX isolates has increased from 20% in 2000 to 60% in 2007. Recent isolates in China are LDT3 and 4/91 (Feng et al. 2014; Li et al. 2010; Ji et al. 2011). Attenuated live strains such as H120, LDT3, 4/91, and inactivated M41 are the commercially used vaccine strains in China. Previous studies reported that Mass type of strains is widely in circulation. Recombination of Mass type with other strain resulted in complete cross-protection of Mass strain by H120 strain. However, two recombinant strains (CK/CH/LDL/110931 and CK/CH/LHB/130573) are highly variable in S1 gene from Mass type and are not cross-neutralized by H120 strain (Chen et al. 2015; Han et al. 2011). Since 2009, the emergence of Taiwan group (TW-I and TW-II) of strains have been increased in China (Xu et al. 2016; Feng et al. 2014; Zou et al. 2010; Luo et al. 2012). Recombinants GD strain, created from QX and TW-I strains, induced renal lesions, respiratory symptoms, and 40% mortality (Xu et al. 2016; Yan et al. 2016). JP-I, JP-II, and JP-III are three genetic groups of IBV in Japan. Different variants of 4/91 strains are isolated (Inoue et al. 2008; Shimazaki et al. 2009). Two variants of 4/91 strain (JP/Wakayama/2003 and JP/Iwate/2005) are cross-neutralized by 4/91, JP-I and 4/91, JP-II strains, respectively. Ibaraki strain isolated in 2009 was distinctly related to other Japanese strain. Korean group-I and group-II are the two genetic groups of IBV that exist in Korea. Group-I is closely related to Mass type. There are three subgroups of group-II, namely, KM91-like, New Cluster 1 (NC1), and QX-like (Lim et al. 2011). Korean group-II isolates that emerged from 2005 to 2010 were nephropathogenic (Lim et al. 2011). Since 2009, predominant isolates circulating in Thailand are QX-like variants (Promkuntod et al. 2015). Indian IBV isolate PDRC/Pune/Ind/1/00 was found to have a unique S1 sequence compared to reference strains from the United States, Europe, Mexico, and Australia and found as nephropathogenic (Bayry et al. 2005). 4/91 type strain was first isolated in India by Sumi et al. in 2012. An Indian IBV strain (India/LKW/56/IVRI/08) showed 99% homology with a Thailand strain (THA280252), while another isolate (India/NMK/72/IVRI/10) showed similarity with the United Kingdom (4/91 pathogenic strain), Japan (JP/Wakayama/2/2004), and China (TA03) (Sumi et al. 2012). Indian strain that was isolated in 2015 by Patel and coworkers was similar to Mass type vaccine M41 strain. Six IBV genotypes, namely, 4/91, IS/1494/06, Mass, IS/885/00, Q1, and D274, were detected in the Middle East from 2009 to 2014 (Ganapathy et al. 2015). Analysis of complete genome sequence of a field strain of IBV from the northern part of India indicated the emergence of a genotype I variant of IBV (Jakhesara et al. 2018). China-like strains (CK/CH/Guangdong/Xindadi/0903 and CK/CH/LDL/97I) are also reported in the Middle East (Seger et al. 2016; Ababneh et al. 2012). LDL/97I-like strains isolated in the Middle East showed extensive tissue tropism such as trachea, kidneys, ovarian tissue, and cecal tonsils, whereas original LDL/97I strains were limited to the respiratory system and kidneys.
IBV isolates in Egypt are divided into the Egy/Var I, Egy/Var II, and Mass type groups (Zanaty et al. 2016). QX-like strains and Italy 02 type strains were first isolated in Zimbabwe (Toffan et al. 2011) and Morocco, respectively (Fellahi et al. 2015). Variants such as Conn, Mass, Florida, Clark 333, Ark, Holte, and Gray were identified in North America (Jungherr et al. 1956; Winterfield and Hitchner 1962; Brown et al. 1987; Butcher et al. 1989; Kinde et al. 1991). However, Mass, Conn, and Ark are the common serotypes that are reported. A nephropathogenic IBV strain (DMV/1639/11) was detected in Delmarva in 2011. When a vaccine containing a mixture of Mass, Conn, and Ark administered through intraocular route, virus shedding and renal lesions were decreased following DMV/1639/11 challenge (Gelb et al. 2013). Cal99 strain could affect only the respiratory tract, while Cal99 variants have a tendency to spread to the kidneys, gastrointestinal tract, and the bursa in addition to the respiratory system (Franca et al. 2011). Other IBV variants such as CAV, DE072, and MX97–8147 have also been reported (Jackwood et al. 2005). The D207, Mass, Conn, and Ark serotypes were detected in Brazil (Felippe et al. 2010).
Immunobiology
Innate Immunity
Toll-like receptor 3 (TLR3) on innate immune cells is involved in the early recognition of the virus (Ariaans et al. 2008). TLR3 mRNA expression was more following IBV-M41 strain infection (Wang et al. 2006), and upregulation of TLR3 and TLR7 mRNA was detected in the trachea and lungs when chicks were intratracheally infected with Conn strain (Kameka et al. 2014). Nephropathogenic IBV infection significantly increases chicken myeloma differentiation antigen 5 (MDA5) expression in the kidneys, which indicate the role of chicken MDA5 against IBV infection (Cong et al. 2013). Chicken mannose-binding lectin (MBL), a member of C-type collectin family, has antiviral activity against IBV. It binds to S1 protein of IBV and blocks the attachment of viral particles to the surface of the susceptible cells in the chicken trachea (Zhang et al. 2017). In addition, MBL also plays a major role in shaping innate as well as adaptive immune responses against IBV (Hamzic et al. 2016; Juul-Madsen et al. 2011; Kjaerup et al. 2014b). Further, a high level of MBL contributes to the clearance of IBV from the trachea (Juul-Madsen et al. 2011; Kjaerup et al. 2014a).
During viral infection, cytokines are involved in the protection of adjacent cells as well as facilitate the activation and migration of T lymphocytes to the infection site (Guo et al. 2008). IBV infection induces interferon in the trachea, lung, kidney, liver, and spleen (Otsuki et al. 1987). Replication of IBV strains both in vitro and in vivo was inhibited in the presence of IFN-α. Both IFN-α and IFN-β transcripts are elevated in the trachea after IBV infection. Among type I interferon, IFN-β has a dominant role in the innate immune response against IBV infection (Yang et al. 2018). IFN-γ contributes to the tracheal lesions, and IFN-λ plays a major role in the host resistance against IBV especially in the trachea (Yang et al. 2018). IBV can act as a polyclonal stimulator of IFN-γ production in the chicken leukocytes, which is IBV-specific as other chicken viruses were unable to do the same (Ariaans et al. 2009). Proinflammatory cytokines like IL-6 and IL-1β were upregulated at 3 days post-IBV-M41 infection. These cytokines have association with increased viral load and tracheal lesions as well as defects in eggshell components in the laying hens (Okino et al. 2014; Nii et al. 2014).
Macrophages and heterophils are important innate immune cells during the initial phase of infection. Heterophils are the first phagocytic cells recruited at the infection site. Upon phagocytosis, heterophils degranulate cytotoxic molecules like cathepsin and bactericidal permeability increasing protein and thus try to neutralize phagocytosed pathogen. Heterophils are responsible for destruction of IBV-infected cells by phagocytosis and oxidative lysosomal enzyme release during the initial phase of infection (Guo et al. 2008). Further, depletion of heterophils causes more severe nasal exudation in comparison with control when infected with IBV (Raj et al. 1997). Activation of macrophages is based mainly on the action of IFN-γ (Caron 2010). Macrophages were found to increase in number in the tracheal and bronchial lumen of chickens at 24 and 96 h post-IBV-M41 infection (Fulton et al. 1993). Similarly, chickens infected with Conn strain significantly increased macrophages in the lungs and trachea at 24 h post-infection (hpi), suggesting the role of respiratory macrophages in limiting replication of virus within respiratory tissues (Kameka et al. 2014). Recruitment of macrophages and production of IL-1β play a major role in the host responses against IBV infection (Amarasinghe et al. 2018).
Adaptive Immunity
Humoral and cellular immune responses are important in the control of infection. Local antibody level but not serum antibodies are involved in the protection of respiratory tract epithelium (Raggi and Lee 1965). Hyperplasia of goblet cells and alveolar mucous glands resulting in seromucous nasal discharge and catarrhal exudates in the trachea are believed to be the first component of innate immunity (Nakamura et al. 1991). Serum anti-IBV IgG can be detected at 4 days post-infection (dpi) and peaking at about 21 dpi. In an in vitro study, activation of memory B cells by IBV results in the secretion of antibodies at 21 dpi (Pei and Collisson 2005). IgG antibody titer was highest at 7 dpi in the lachrymal fluid and possibly remaining up to 23 dpi. Similarly, in oviduct washes, antibodies were detected at 7 and 23 dpi (Raj and Jones 1996).
IBV-infected chickens showed anti-IBV IgA antibodies in the lamina propria, trachea, and between the epithelial cells of trachea (Joiner et al. 2007; Nakamura et al. 1991). Anti-IBV IgA was detected in the lachrymal fluid at 10 days post-live attenuated Ark DPI-type vaccination. However, there was no further increase in IgA level upon challenge, suggesting the significant role of neutralizing antibodies in reducing the potency of IBV infection at the time of challenge (Joiner et al. 2007). In our lab, administration of resiquimod (R-848), a TLR-7 agonist with inactivated or live IBV vaccine increased the secretory IgA, which was mediated through the enhanced expression TGF-β4 in the chicken (Matoo et al. 2018).
Humoral immunity plays an important role in recovery and clearance of viral infection. Bursectomized chicks showed increase in tissue viral load as well as severe and long-lasting illness (Cook et al. 1991). As the titer of humoral antibodies increased, re-isolation of virus from kidneys and genital tract decreased (Macdonald et al. 1981). This increase in antibody titer also protected against drop in egg production and viral spread from trachea to other susceptible organs (Box et al. 1988; Raj and Jones 1997). Conversely, it has also been shown that chickens were more susceptible to IBV with high tear antibody titer, whereas low tear antibody titer protected the birds from IBV. This suggested that not only antibody but other immune defense mechanisms are also important in clearance of infection in tear (Gelb et al. 1998).
Nephropathogenic Gray strain-infected chickens showed increased IBV-specific cytotoxic T lymphocytes (CTLs) at 3 dpi with peak level at 10 dpi (Seo and Collisson 1997). A study assessing the immunophenotypes of mononuclear cells in the tracheal mucosa revealed the presence of CTLs at 3 or 4 dpi suggesting the role of CTLs in viral clearance during the early phase of infection (Kotani et al. 2000). CD8+ T cells play an important role in controlling IBV infection. This IBV-specific CTL activity was dependent on S and N proteins of IBV, and it can induce lysis of IBV-infected cells (Collisson et al. 2000). While CD8+ T cells play an important role in early immune response, CD4+ T and B cells are involved in long-term control of IB.
Maternally Derived Antibodies
IgG antibodies were transferred from vaccinated hens to the respiratory mucus of newly hatched chicks (Hawkes et al. 1983). One-day-old chicks with high level of maternal antibodies showed more than 95% protection against IBV-Mass strain challenge. However, this was not evident at 7 days of its age. Protection was associated with local respiratory antibodies but not with serum antibodies (Mondal and Naqi 2001). Day-old chicks when vaccinated with live IBV-Mass strain showed no IBV-specific antibody response. This may be due to binding and neutralization of vaccine virus by maternal antibodies (Mondal and Naqi 2001). Prime boost strategies involve vaccination with live strain followed by inactivated vaccine to protect hens throughout their laying period as well as to transfer high level of antibodies to their offspring (Chhabra et al. 2015).
Diagnosis
Virus Isolation
Since trachea is the primary site of IBV infection, tracheal swab and tracheal tissues are the preferred sampling material within 1 week of infection. Cloacal swabs or cecal tonsils are the sample of choice during postmortem examination if time is elapsed more than 1 week from the start of infection. This may be due to the initial growth of virus in the upper respiratory tract and eventual spread to the non-respiratory organs. This results in the early clearance of virus from trachea than from the intestinal tissues (Alexander and Gough 1977; Jones and Ambali 1987; Lucio and Fabricant 1990). Based on clinical history, sampling of other organs like the lung, kidney, and oviduct should also be considered. Sentinel chickens should be placed in the problematic flock where direct sampling method is difficult. After 1 week of contact exposure, sentinels are removed for direct sampling method (Gelb et al. 1989).
Isolation in Embryonated Eggs
Embryonated chicken eggs infected with IBV shows characteristic signs including dwarfism, curling, hemorrhages, and death (Fig. 16.1). Field samples were inoculated through intra-allantoic route of 9- to 10-day-old embryonated eggs and incubated for 4 days. Allantoic fluid were collected and again passaged to analyze the morphology of the embryo. IBV titer reaches its maximum about 1 to 2 days post-inoculation. Although chicken embryonated eggs are effective model for virus isolation, it has the drawback of need for three successive passages for manifestation of characteristic lesions. In addition, reduced virus yield may be observed when IBV is inactivated due to improper preservation (Villarreal 2010).
Fig 16.1.
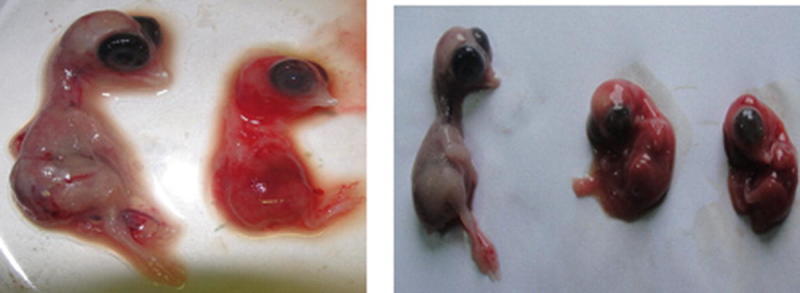
Comparison of 12-day-old normal (left) and dwarfed and IBV-infected embryos (right) of the same age
Cultivation in Tracheal Rings
Tracheal rings were collected from 19- to 20-day-old SPF embryo and placed in a tube containing culture media and antibiotic. This was incubated for 48 h, and only the rings with more than 50% ciliary motility are used. After removing the culture media, 0.1 ml of sample was added and incubated for 1 h for viral adsorption. One ml of culture media was once again added and incubated. Tracheal rings were evaluated for ciliary motility at 24, 48, 72, and 96 h post-inoculation. Ciliary motility decreases as IBV replicates in the tracheal cells (Epiphanio et al. 2002). This technique also has disadvantage of not being sensitive for IBV field samples and may result in false-negative results. Thus, IBV should be confirmed by other specific methods like virus neutralization (VN), hemagglutination inhibition (HI), ELISA, immunohistochemistry, or nucleic acid analysis or by electron microscopy.
Serology
Different serotypes of IBV show high amino acid similarity within N, M, and S2 protein, and hence these regions are called group-specific antigens. The antibodies against group-specific antigen of one serotype can neutralize different serotypes, while antibodies against S1 protein of the virus are type-specific and are not cross-protective between different serotypes (Cavanagh and Gelb 2008). The serological tests like, ELISA, immunofluorescence, and immunodiffusion test cannot differentiate different serotypes because antibodies bind to both group- and type-specific antigens, while VN and HI are serotype-specific tests. Therefore, VN, HI, and ELISA are routinely used serological methods (de Wit 2000). Precipitating antibodies in the group-specific agar gel precipitation test (AGPT) test are short lived which may lead to under-detection. Positive AGPT is indicative of recent infection. Birds infected with the same serotype as vaccine virus lead to poor production of antibodies as detected by AGPT. Thus, AGPT is not recommended for detection of IBV antibodies and rather for detection of IBV antigen (de Wit et al. 1997, 1998). IBV ELISAs are group-specific methods and detect IBV antibodies within 1 week of infection. Paired serum samples are required with first sample collected at the beginning of infection and second sample at 4 weeks later. The first sample should be collected without any delay for detection of seroconversion. Hence, ELISA analyzes post-vaccination and infection responses by IgG detection as an indicator of humoral immunity (Cavanagh and Gelb 2008). VN and HI are serotype-specific techniques for detection of antibodies induced by S1 protein. VN can be carried out in embryonated egg, cell culture, and tracheal ring culture to detect embryo alterations, cytopathic effect, and ciliostasis, respectively. This assay requires neutralizing antibodies that are highly specific and do not cross-react with other serotypes. IBV requires treatment with type C phospholipase enzyme to expose hemagglutinin. HI test usually detects antibodies between 1 and 2 weeks after infection (de Wit 2000).
Antibody-Based Methods
Immunofluorescence and immunoperoxidase assays are used to detect IBV antigen from tracheal mucosa or other tissues using IBV-specific polyclonal sera or monoclonal antibodies (Handberg et al. 1999). The presence of virus in the tracheal organ culture can be detected by immunofluorescence without fixation of the culture using low-power microscopy.
Nucleic Acid-Based Methods
IBV can be detected directly from the clinical sample by real-time RT-PCR or quantitative RT-PCR (Callison et al. 2006). This technique is cost- effective, as a number of samples can be examined in a short period of time, and also gives an indication of the level of viral nucleic acid in the sample. Although conventional RT-PCR identifies IBV nucleic acid, it requires passage in embryonated eggs before positive result is obtained. Swabs containing only a small amount of viral RNA can be detected by nested RT-PCR. However, this method is highly prone to cross-contamination resulting in a false-positive test. Positive RT-PCR indicates only the presence of viral nucleic acid but not IBV type. IBV types can be identified by genotyping methods by analyzing the amplicons from the S1 gene. IBV types can be differentiated based on their unique electrophoresis banding pattern using restriction fragment length polymorphism (RFLP) (Marquardt et al. 1981; Nakamura et al. 1994).
Recent Advances
An intelligent electronic device was developed by Banakar and coworkers for fast diagnosis of Newcastle disease, IB, and avian influenza based on chicken’s sound signals (Banakar et al. 2016). Laamiri and coworkers developed a one-step multiplex real-time RT-PCR for simultaneous detection of four respiratory avian viruses including IBV (Laamiri et al. 2018).
Prevention and Control
Vaccination and strict biosecurity measures are important for the prevention and control of IBV. Both live attenuated and killed vaccines are used to control IBV in commercial poultry farm. Since serotypes of IBV do not cross-protect each other, multivalent vaccine containing two or more antigenic types would be beneficial in providing broad protection (Cavanagh 2007; Dhama et al. 2014). In most countries, low virulent IBV vaccines are administered in day-old chicks with subsequent booster immunization with virulent vaccines in drinking water. Unlike highly virulent vaccine, low virulent IBV do not cause respiratory reaction. However, immunity produced by low virulent IBV is not always enough to protect the respiratory tract (Kataria et al. 2005). Prior to immunization with oil-emulsion-inactivated vaccines, breeders and commercial egg layers are initially primed with live attenuated IBV vaccines in order to maintain a good level of local protection of the respiratory tract. Oil-emulsion-inactivated vaccines are given at 17 weeks of age (prior to egg laying of breeder or layer) and never during laying (Jackwood et al. 2009; Liu et al. 2009).
Embryonated chicken eggs are serially passaged with IBV strains to prepare live attenuated vaccines. Prolonged or lifelong immune response can be achieved through vaccination under optimal conditions (Cook et al. 2012; Dhama et al. 2014). Mass serotype, such as H120, is the most commonly used strain in live vaccines. Initially, birds are either vaccinated individually by eye drop instillation, intranasal and intratracheal routes, or mass vaccination by coarse spray or drinking water. Such mass vaccination procedures are cost-effective and induce both local and systemic immune responses. Ma5 is a single-component vaccine that can be included in first vaccination programs with IB 4/91 vaccines and inactivated vaccines. This type of vaccination program provides broad protection against different IBV serotypes. Selection of vaccine must be based on the prevalent strains in the local area. Administration of different vaccine strains together confers cross-protection against various field isolates in the SPF chicken (Li et al. 2008; Marandino et al. 2015). Combination of Mass and Conn or Mass and JMK provides higher degree of cross-protection to some heterologous strains. IB 4/91 serotype or IB 274 vaccine virus gives specific protection against IBV. These provide broad protection when combined with Ma5 and IB multi-vaccines (Ma et al. 2012). Inactivated vaccines induce higher level of antibody response.
However, live vaccine plays a significant role in protecting commercial layers because of better induction of T cell response and local antibody (IgA) production. In North America, Mass, Conn, and Ark serotypes are included in both live attenuated and inactivated vaccines. California and Georgia 98 vaccine strains are being used in the United States. Holland variants such as D-274 and D-1466 along with the IB H120-based vaccines are used in most parts of Europe. Live, freeze-dried vaccine serotypes (Ma5 and 4/91) provide long-lasting protection. In Korea, newly evolving IBV recombinants can be controlled by K2 vaccine strain (Lim et al. 2011). Mass vaccine strain is the only live attenuated vaccine approved in Brazil (Brandao 2010). Vaccination program involves initial administration of Mass-type vaccine followed by a variant shown to be efficacious against Italy 02 and QX (Jones 2010). The only vaccine strain used in India is Mass strain 41 (M41). DNA, subunit, and vectored vaccines using S1 glycoprotein gene and reverse genetics vaccines have been identified (Dhama et al. 2008; Lin et al. 2012). Protection against two or more serotypes can be achieved through introduction of antigen from two or more viruses in recombinant or vector-based vaccines. These new-generation vaccines can be administered safely in ovo, and efficacy is needed to be tested before introduction for commercial purposes (Matthijs et al. 2003; Brandao 2010; Ullah et al. 2013). Recombinant IBV vaccine using fowl adenovirus vector backbone expressing the S1 glycoprotein gene provides different levels of protection against homologous challenge (Johnson et al. 2003; Toro et al. 2014).
The recent emergence demands the inclusion of QX-like IBV strains in the vaccination program against IB. Yan and coworkers attenuated the QX-like IBV strains by continuous passage in chicken embryos for 130 generations, and it was found to be safe and effective for inducing protection against QX-like IBV strains (Yan et al. 2018). The accessory proteins 3a, 3b, 5a, and 5b contribute individually to the pathogenicity of IBV, and deleting any one of these genes results in attenuation of the virus and can be a potential vaccine candidate (Laconi et al. 2018). Another research team also reported the deletion of accessory genes 3ab and/or 5ab in IBV resulting in attenuation with the ability to induce protection in chickens (van Beurden et al. 2018). Recently, inactivated IBV vaccine encapsulated in chitosan nanoparticles induced an early and stronger IgA and IgG anti-IBV antibodies, when it was vaccinated alone or in association with a live attenuated vaccine (Lopes et al. 2018).
Some of the alternative control measures have been tried and found to be partially successful for the control of IBV. The chicken TLR21 is a functional homologue of mammalian TLR9, recognizes CpG motif, and results in the induction of nuclear factor-kappa B (NF-κB) and its related cytokines (Brownlie et al. 2009). Prophylactic administration of CpG to 18-day-old embryos decreased the viral load (Dar et al. 2009). Astragalus polysaccharides isolated from a traditional Chinese medicinal herb, Astragalus mongholicus, inhibit IBV infection, in vitro, in a dose-dependent manner. The lower viral replication was associated with reduced mRNA levels of the proinflammatory cytokines IL-1β, IL-6, IL-8, and TNF-α (Zhang et al. 2018).
Future Perspectives
Avian infectious bronchitis is one of the most difficult diseases to be controlled in the chicken. Continued emergence of different variants of IBV complicates the control program. Both live attenuated and killed vaccines are used in the field; however, IB outbreaks even in the vaccinated flocks are constantly reported from many parts of the world. Epidemiological surveillance and improved knowledge on the circulating field variants of IBV are necessary for implementing a better control program. Research works are still warranted for improving our understanding on immune response against IBV particularly cellular immunity and a strategic vaccine having capacity to control diverse variants of the virus.
Acknowledgments
All the authors of the manuscript thank and acknowledge their respective universities and institutes.
Conflict of Interest
There is no conflict of interest.
Contributor Information
Yashpal Singh Malik, Email: malikyps@ivri.res.in.
Raj Kumar Singh, Email: rks_virology@rediffmail.com.
Mahendra Pal Yadav, Email: yadav_mp@hotmail.com.
Saravanan Ramakrishnan, Email: dearsaromib@yahoo.com.
References
- Ababneh M, Dalab AE, Alsaad S, Al-Zghoul M. Presence of infectious bronchitis virus strain CK/CH/LDL/97I in the Middle East. ISRN Vet Sci. 2012;2012:201721. doi: 10.5402/2012/201721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abro SH, Renström LHM, Ullman K, Isaksson M, Zohari S, Jansson DS, Belak S, Baule C. Emergence of novel strains of avian infectious bronchitis virus in Sweden. Vet Microbiol. 2012;155:237–246. doi: 10.1016/j.vetmic.2011.09.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander DJ, Gough RE. Isolation of avian infectious bronchitis virus from experimentally infected chickens. Res Vet Sci. 1977;23:344–347. doi: 10.1016/S0034-5288(18)33129-1. [DOI] [PubMed] [Google Scholar]
- Amarasinghe A, Abdul-Cader MS, Almatrouk Z, van der Meer F, Cork SC, Gomis S, Abdul-Careem MF. Induction of innate host responses characterized by production of interleukin (IL)-1β and recruitment of macrophages to the respiratory tract of chickens following infection with infectious bronchitis virus. Vet Microbiol. 2018;215:1–10. doi: 10.1016/j.vetmic.2018.01.001. [DOI] [PubMed] [Google Scholar]
- Ariaans MP, Matthijs MGR, Van Harlen D, Van de Haar P, Ivan Eck JHH, Hensen EJ, Vervelde L. The role of phagocytic cells in enhanced susceptibility of broilers to colibacillosis after infectious bronchitis virus infection. Vet Immunol Immunopathol. 2008;123:240–250. doi: 10.1016/j.vetimm.2008.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ariaans MP, van de Haar PM, Hensen EJ, Vervelde L. Infectious bronchitis virus induces acute interferon-γ production through polyclonal stimulation of chicken leukocytes. Virology. 2009;385:68–73. doi: 10.1016/j.virol.2008.11.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banakar A, Sadeghi M, Shushtari A. An intelligent device for diagnosing avian diseases: Newcastle, infectious bronchitis, avian influenza. Comput Electron Agric. 2016;127:744–753. doi: 10.1016/j.compag.2016.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bande Faruku, Arshad Siti Suri, Hair Bejo Mohd, Moeini Hassan, Omar Abdul Rahman. Progress and Challenges toward the Development of Vaccines against Avian Infectious Bronchitis. Journal of Immunology Research. 2015;2015:1–12. doi: 10.1155/2015/424860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayry J, Goudar MS, Nighot PK, Kshirsagar SG, Ladman BS, Gelb J, Jr, Ghalsasi GR, Kolte GN. Emergence of a nephropathogenic avian infectious bronchitis virus with a novel genotype in India. J Clin Microbiol. 2005;43(2):916–918. doi: 10.1128/JCM.43.2.916-918.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beach JR, Schalm OW. A filtrable virus distinct from that of laryngotracheitis: the cause of a respiratory disease of chicks. Poult Sci. 1936;15:199. doi: 10.3382/ps.0150199. [DOI] [Google Scholar]
- Beaudette FR, Hudson CB. Cultivation of the virus of infectious bronchitis. J Am Vet Med Assoc. 1937;90:51–60. [Google Scholar]
- Boltz DA, Nakai M, Bahra JM. Avian infectious bronchitis virus: a possible cause of reduced fertility in the rooster. Avian Dis. 2004;48:909–915. doi: 10.1637/7192-040808R1. [DOI] [PubMed] [Google Scholar]
- Box PG, Holmes HC, Finney PM, Froymann R. Infectious bronchitis in laying hens: the relationship between haemagglutination inhibition antibody levels and resistance to experimental challenge. Avian Pathol. 1988;17:349–361. doi: 10.1080/03079458808436453. [DOI] [PubMed] [Google Scholar]
- Brandao PE. Avian infectious bronchitis virus in Brazil: a highly complex virus meets a highly susceptible host population. Rev Bras Cienc Avic. 2010;12:121–124. doi: 10.1590/S1516-635X2010000200008. [DOI] [Google Scholar]
- Brown TP, Glisson JR, Rosales G, Villegas P, Davis RB. Studies of avian urolithiasis associated with an infectious bronchitis virus. Avian Dis. 1987;31:629–636. doi: 10.2307/1590750. [DOI] [PubMed] [Google Scholar]
- Brownlie R, Zhu J, Allan B, Mutwiri GK, Babiuk LA, Potter A, Griebel P. Chicken TLR21 acts as a functional homologue to mammalian TLR9 in the recognition of CpG oligodeoxynucleotides. Mol Immunol. 2009;46:3163–3170. doi: 10.1016/j.molimm.2009.06.002. [DOI] [PubMed] [Google Scholar]
- Bushnell LD, Brandly CA. Laryngotracheitis in chicks. Poult Sci. 1933;12(1):55–60. doi: 10.3382/ps.0120055. [DOI] [Google Scholar]
- Butcher GD, Winterfield RW, Shapiro DP. An outbreak of nephropathogenic H13 infectious bronchitis in commercial broilers. Avian Dis. 1989;33:823–826. doi: 10.2307/1591168. [DOI] [PubMed] [Google Scholar]
- Callison SA, Hilt DA, Boynton TO, Sample BF, Robison R, Swayne DE, Jackwood MW. Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J Virol Methods. 2006;138:60–65. doi: 10.1016/j.jviromet.2006.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caron LF. Etiology and immunology of infectious virus. Rev Bras Cienc Avic. 2010;12:115–119. doi: 10.1590/S1516-635X2010000200007. [DOI] [Google Scholar]
- Cavanagh D. Coronaviruses with special emphasis on first insights concerning SARS. Basel: Birkhäuser; 2005. Coronaviridae: a review of coronaviruses and toroviruses; pp. 1–54. [Google Scholar]
- Cavanagh D. Coronavirus avian infectious bronchitis virus. Vet Res. 2007;38:281–297. doi: 10.1051/vetres:2006055. [DOI] [PubMed] [Google Scholar]
- Cavanagh D, Gelb J., Jr . Infectious bronchitis. In: Saif YM, Fadly AM, Glisson JR, McDougald LR, Nolan LK, Swayne DE, editors. Diseases of poultry. Ames: Blackwell; 2008. pp. 117–135. [Google Scholar]
- Cavanagh D, Davis PJ, Darbyshire JH, Peters RW. Coronavirus IBV: virus retaining spike glycopolypeptide S2 but not S1 is unable to induce virus-neutralizing or haemagglutination- inhibiting antibody, or induce chicken tracheal protection. J Gen Virol. 1986;67:1435–1442. doi: 10.1099/0022-1317-67-7-1435. [DOI] [PubMed] [Google Scholar]
- Chen L, Zhang T, Han Z, Liang S, Xu Y, Xu Q, Chen Y, Zhao Y, Shao Y, Li H, Wang K, Kong X, Liu S. Molecular and antigenic characteristics of Massachusetts genotype infectious bronchitis coronavirus in China. Vet Microbiol. 2015;181:241–251. doi: 10.1016/j.vetmic.2015.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chhabra R, Chantrey J, Ganapathy K. Immune responses to virulent and vaccine strains of infectious bronchitis viruses in chickens. Viral Immunol. 2015;28(9):478–488. doi: 10.1089/vim.2015.0027. [DOI] [PubMed] [Google Scholar]
- Circella E, Camarda A, Martella V, Bruni G, Lavazza A, Buonavoglia C. Coronavirus associated with an enteric syndrome on a quail farm. Avian Pathol. 2007;36:251–258. doi: 10.1080/03079450701344738. [DOI] [PubMed] [Google Scholar]
- Collisson EW, Pei J, Dzielawa J, Seo SH. Cytotoxic T lymphocytes are critical in the control of infectious bronchitis virus in poultry. Dev Comp Immunol. 2000;24:187–200. doi: 10.1016/S0145-305X(99)00072-5. [DOI] [PubMed] [Google Scholar]
- Cong F, Liu X, Han Z, Shao Y, Kong X, Liu S. Transcriptome analysis of chicken kidney tissues following coronavirus avian infectious bronchitis virus infection. BMC Genomics. 2013;14:743. doi: 10.1186/1471-2164-14-743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cook JK, Davison TF, Huggins MB, McLaughlan P. Effect of in ovo bursectomy on the course of an infectious bronchitis virus infection in line C White Leghorn chickens. Arch Virol. 1991;118:225–234. doi: 10.1007/BF01314032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cook JKA, Jackwood M, Jones RC. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012;41:239–250. doi: 10.1080/03079457.2012.680432. [DOI] [PubMed] [Google Scholar]
- Corse E, Machamer CE. The cytoplasmic tails of infectious bronchitis virus E and M proteins mediate their interaction. Virology. 2003;312:25–34. doi: 10.1016/S0042-6822(03)00175-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dar A, Potter A, Tikoo S, Gerdts V, Lai K, Babiuk LA, Mutwiri G. CpG oligodeoxynucleotides activate innate immune response that suppresses infectious bronchitis virus replication in chicken embryos. Avian Dis. 2009;53:261–267. doi: 10.1637/8560-121808-Reg.1. [DOI] [PubMed] [Google Scholar]
- de Haan CAM, Vennema H, Rottier PJM. Assembly of the coronavirus envelope: homotypic interactions between the M proteins. J Virol. 2000;74:4967–4978. doi: 10.1128/JVI.74.11.4967-4978.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Wit JJ. Detection of infectious bronchitis. Avian Pathol. 2000;29:71–93. doi: 10.1080/03079450094108. [DOI] [PubMed] [Google Scholar]
- de Wit JJ, Mekkes DR, Kouwenhoven B, Verheijden JHM. Sensitivity and specificity of serological tests for detection of infectious bronchitis virus induced antibodies in broilers. Avian Pathol. 1997;26:105–118. doi: 10.1080/03079459708419198. [DOI] [PubMed] [Google Scholar]
- de Wit JJ, Mekkes DR, Koch G, Westenbrink F. Detection of specific Ig M antibodies to infectious bronchitis virus by an antibody-capture ELISA. Avian Pathol. 1998;27:155–160. doi: 10.1080/03079459808419317. [DOI] [PubMed] [Google Scholar]
- Dea S, Tijssen P. Detection of Turkey enteric coronavirus by enzyme-linked immunosorbent assay and differentiation from other coronaviruses. Am J Vet Res. 1989;50:226–231. [PubMed] [Google Scholar]
- Dent SD, Xia D, Wastling JM, Neuman BW, Britton P, Maier HJ. The proteome of the infectious bronchitis virus Beau-R virion. J Gen Virol. 2015;96:3499–3506. doi: 10.1099/jgv.0.000304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhama K, Mahendran M, Gupta PK, Rai A. DNA vaccines and their applications in veterinary practice: current perspectives. Vet Res Commun. 2008;32:341–356. doi: 10.1007/s11259-008-9040-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhama K, Singh SD, Barathidasan R, Desingu PA, Chakraborty S, Tiwari R, Kumar MA. Emergence of avian infectious bronchitis virus and its variants need better diagnosis, prevention and control strategies: a global perspective. Pak J Biol Sci. 2014;17:751–767. doi: 10.3923/pjbs.2014.751.767. [DOI] [PubMed] [Google Scholar]
- Epiphanio E.O.B., Martins N.R.S., Resende J.S., Pinto R.G., Jorge M.A., Souza M.B., Caccioppoli J., Cardozo R.M. Resultados preliminares da utilização de cultivos de anéis de traquéia para o estudo de estirpes brasileiras do vírus da bronquite infecciosa das galinhas. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. 2002;54(2):212–216. doi: 10.1590/S0102-09352002000200013. [DOI] [Google Scholar]
- Felippe PA, da Silva LH, Santos MM, Spilki FR, Arns CW. Genetic diversity of avian infectious bronchitis virus isolated from domestic chicken flocks and coronaviruses from feral pigeons in Brazil between 2003 and 2009. Avian Dis. 2010;54:1191–1196. doi: 10.1637/9371-041510-Reg.1. [DOI] [PubMed] [Google Scholar]
- Fellahi S, Ducatez M, El Harrak M, Guerin JL, Touil N, Sebbar G, Bouaitiel A, Khataby K, Ennaji MM, El-Houadfi M. Prevalence and molecular characterization of avian infectious bronchitis virus in poultry flocks in Morocco from 2010 to 2014 and first detection of Italy 02 in Africa. Avian Pathol. 2015;44:287–295. doi: 10.1080/03079457.2015.1044422. [DOI] [PubMed] [Google Scholar]
- Feng K, Xue Y, Wang F, Chen F, Shu D, Xie Q. Analysis of S1 gene of avian infectious bronchitis virus isolated in southern China during 2011–2012. Virus Genes. 2014;49:292–303. doi: 10.1007/s11262-014-1097-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franca M, Woolcock PR, Yu M, Jackwood MW, Shivaprasad HL. Nephritis associated with infectious bronchitis virus Cal99 variant in game chickens. Avian Dis. 2011;55:422–428. doi: 10.1637/9417-060510-Reg.1. [DOI] [PubMed] [Google Scholar]
- Fulton RM, Reed WM, Thacker HL. Cellular response of the respiratory tract of chickens to infection with Massachusetts 41 and Australian T infectious bronchitis viruses. Avian Dis. 1993;37:951–960. doi: 10.2307/1591899. [DOI] [PubMed] [Google Scholar]
- Ganapathy K, Ball C, Forrester A. Genotypes of infectious bronchitis viruses circulating in the Middle East between 2009 and 2014. Virus Res. 2015;210:198–204. doi: 10.1016/j.virusres.2015.07.019. [DOI] [PubMed] [Google Scholar]
- Gay K (2000) Infectious bronchitis virus detection and persistence in experimentally infected chickens. MS thesis, Cornell University, Ithaca, NY, USA
- Gelb J, Jr, Rosenberger JK, Fries PA, Cloud SS, Odor EM, Dohms JE, Jaeger JS. Protection afforded infectious bronchitis virus-vaccinated sentinel chickens raised in a commercial environment. Avian Dis. 1989;33:764–769. doi: 10.2307/1591158. [DOI] [PubMed] [Google Scholar]
- Gelb J, Jr, Nix WA, Gellman SD. Infectious bronchitis virus antibodies in tears and their relationship to immunity. Avian Dis. 1998;42:364–374. doi: 10.2307/1592487. [DOI] [PubMed] [Google Scholar]
- Gelb J, Jr, Ladman BS, Pope CR, Ruano JM, Brannick EM, Bautista DA, Coughlin CM, Preskenis LA. Characterization of nephropathogenic infectious bronchitis virus DMV/1639/11 recovered from Delmarva broiler chickens in 2011. Avian Dis. 2013;57:65–70. doi: 10.1637/10313-080212-Reg.1. [DOI] [PubMed] [Google Scholar]
- Guo X, Rosa AJM, Chen DG, Wang X. Molecular mechanisms of primary and secondary mucosal immunity using avian infectious bronchitis virus as a model system. Vet Immunol Immunopathol. 2008;121:332–343. doi: 10.1016/j.vetimm.2007.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamzic E, Kjaerup RB, Mach N, Minozzi G, Strozzi F, Gualdi V, Williams JL, Chen J, Wattrang E, Buitenhuis B, Juul-Madsen HR, Dalgaard TS. RNA sequencing-based analysis of the spleen transcriptome following infectious bronchitis virus infection of chickens selected for different mannose-binding lectin serum concentrations. BMC Genomics. 2016;17:82. doi: 10.1186/s12864-016-2403-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han Z, Sun C, Yan B, Zhang X, Wang Y, Li C, Zhang Q, Ma Y, Shao Y, Liu Q, Kong X, Liu S. A 15-year analysis of molecular epidemiology of avian infectious bronchitis coronavirus in China. Infect Genet Evol. 2011;11:190–200. doi: 10.1016/j.meegid.2010.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Handberg KJ, Nielsen OL, Pedersen MW, Jorgensen PH. Detection and strain differentiation of infectious bronchitis virus in tracheal tissues from experimentally infected chickens by reverse transcriptase-polymerase chain reaction. Comparison with an immunohistochemical technique. Avian Pathol. 1999;28:327–335. doi: 10.1080/03079459994579. [DOI] [PubMed] [Google Scholar]
- Hawkes RA, Darbyshire JH, Peters RW, Mockett AP, Cavanagh D. Presence of viral antigens and antibody in the trachea of chickens infected with avian infectious bronchitis virus. Avian Pathol. 1983;12:331–340. doi: 10.1080/03079458308436175. [DOI] [PubMed] [Google Scholar]
- Ignjatovic J, Sapats S. Rev Sci Tech Off Int Epiz. 2000;19(2):493–508. doi: 10.20506/rst.19.2.1228. [DOI] [PubMed] [Google Scholar]
- Inoue T, Yamaguchi S, Imada T. Existence of avian infectious bronchitis virus with a European-prevalent 4/91 genotype in Japan. J Vet Med Sci. 2008;70:1341–1344. doi: 10.1292/jvms.70.1341. [DOI] [PubMed] [Google Scholar]
- Jackwood MW, de Witt JJ. Infectious bronchitis. In: Swayne DE, Glisson JR, McDougald LR, Nolan LK, Suarez DL, Nair VL, editors. Diseases of poultry. Hoboken: Wiley; 2013. pp. 39–159. [Google Scholar]
- Jackwood MW, Hilt DA, Lee CW, Kwon HM, Callison SA, Moore KM, Moscoso H, Sellers H, Thayer S. Data from 11 years of molecular typing infectious bronchitis virus field isolates. Avian Dis. 2005;49:614–618. doi: 10.1637/7389-052905R.1. [DOI] [PubMed] [Google Scholar]
- Jackwood MW, Hilt DA, Mccall AW, Polizzi CN, Mckinley ET, Williams SM. Infectious bronchitis virus field vaccination coverage and persistence of Arkansas-type viruses in commercial broilers. Avian Dis. 2009;53:175–183. doi: 10.1637/8465-090308-Reg.1. [DOI] [PubMed] [Google Scholar]
- Jackwood MW, Hall D, Handel A. Molecular evolution and emergence of avian gammacoronaviruses. Infect Genet Evol. 2012;12:1305–1311. doi: 10.1016/j.meegid.2012.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakhesara SJ, Nath B, Pal JK, Joshi CG, Kumar S. Emergence of a genotype I variant of avian infectious bronchitis virus from northern part of India. Acta Trop. 2018;183:57–60. doi: 10.1016/j.actatropica.2018.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jayaram J, Youn S, Collisson EW. The virion N protein of infectious bronchitis virus is more phosphorylated than the N protein from infected cell lysates. Virology. 2005;339:127–135. doi: 10.1016/j.virol.2005.04.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ji J, Xie J, Chen F, Shu D, Zuo K, Xue C, Qin J, Li H, Bi Y, Ma J. Phylogenetic distribution and predominant genotype of the avian infectious bronchitis virus in China during 2008–2009. Virol J. 2011;8:84. doi: 10.1186/1743-422X-8-184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson MA, Pooley C, Ignjatovic J, Tyack SG. A recombinant fowl adenovirus expressing the S1 gene of infectious bronchitis virus protects against challenge with infectious bronchitis virus. Vaccine. 2003;21:2730–2736. doi: 10.1016/S0264-410X(03)00227-5. [DOI] [PubMed] [Google Scholar]
- Joiner KS, Hoerr FJ, Ewald SJ, van Santen VL, Wright JC, van Ginkel FW, Toro H. Pathogenesis of infectious bronchitis virus in vaccinated chickens of two different major histocompatibility B complex genotypes. Avian Dis. 2007;51:758–763. doi: 10.1637/0005-2086(2007)51[758:POIBVI]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Jonassen CM, Kofstad T, Larsen IL, Lovland A, Handeland K, Follestad A, Lillehaug A. Molecular identification and characterization of novel coronaviruses infecting graylag geese (Anser anser), feral pigeons (Columbia livia) and mallards (Anas platyrhynchos) J Gen Virol. 2005;86:1597–1607. doi: 10.1099/vir.0.80927-0. [DOI] [PubMed] [Google Scholar]
- Jones RC. Viral respiratory diseases (ILT, aMPV infections, IB): are they ever under control? Br Poult Sci. 2010;51:1–11. doi: 10.1080/00071660903541378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones RC, Ambali AG. Re-excretion of an enterotropic infectious bronchitis virus by hens at point of lay after experimental infection at day old. Vet Rec. 1987;120:617–618. doi: 10.1136/vr.120.26.617. [DOI] [PubMed] [Google Scholar]
- Jungherr EL, Chomiak TW, Luginbuhl RE (1956) Immunologic differences in strains of infectious bronchitis. In: Proceedings of the 60th annual meeting of the United States Livestock Sanitary Association, Chicago, IL, pp 203–209
- Juul-Madsen HR, Norup LR, Jørgensen PH, Handberg KJ, Wattrang E, Dalgaard TS. Crosstalk between innate and adaptive immune responses to infectious bronchitis virus after vaccination and challenge of chickens varying in serum mannose-binding lectin concentrations. Vaccine. 2011;29:9499–9507. doi: 10.1016/j.vaccine.2011.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kameka AM, Haddadi S, Kim DS, Cork SC, Abdul-Careem MF. Induction of innate immune response following infectious bronchitis coronavirus infection in the respiratory tract of chickens. Virology. 2014;450–451:114–121. doi: 10.1016/j.virol.2013.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kataria JM, Mohan CM, Dey S, Dash BB, Dhama K. Diagnosis and immunoprophylaxis of economically important poultry diseases: a review. Indian J Anim Sci. 2005;75:555–567. [Google Scholar]
- Kinde H, Daft BM, Castro AE, Bickford AA, Gelb J, Jr, Reynolds B. Viral pathogenesis of a nephrotxopic infectious bronchitis virus isolated from commercial pullets. Avian Dis. 1991;35:415–421. doi: 10.2307/1591200. [DOI] [PubMed] [Google Scholar]
- Kjaerup RM, Dalgaard TS, Norup LR, Bergman IM, Sørensen P, Juul-Madsen HR. Adjuvant effects of mannose-binding lectin ligands on the immune response to infectious bronchitis vaccine in chickens with high or low serum mannose-binding lectin concentrations. Immunobiology. 2014;219:263–274. doi: 10.1016/j.imbio.2013.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kjaerup RM, Dalgaard TS, Norup LR, Hamzic E, Sørensen P, Juul-Madsen HR. Characterization of cellular and humoral immune responses after IBV infection in chicken lines differing in MBL serum concentration. Viral Immunol. 2014;27:529–542. doi: 10.1089/vim.2014.0088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch G, Hartog L, Kant A, van Roozelaar DJ. Antigenic domains on the peplomer protein of avian infectious bronchitis virus: correlation with biological functions. J Gen Virol. 1990;71:1929–1935. doi: 10.1099/0022-1317-71-9-1929. [DOI] [PubMed] [Google Scholar]
- Kotani T, Wada S, Tsukamoto Y, Kuwamura M, Yamate J, Sakuma S. Kinetics of lymphocytic subsets in chicken tracheal lesions infected with infectious bronchitis virus. J Vet Med Sci. 2000;62:397–401. doi: 10.1292/jvms.62.397. [DOI] [PubMed] [Google Scholar]
- Laamiri N, Aouini R, Marnissi B, Ghram A, Hmila I. A multiplex real-time RT-PCR for simultaneous detection of four most common avian respiratory viruses. Virology. 2018;515:29–37. doi: 10.1016/j.virol.2017.11.021. [DOI] [PubMed] [Google Scholar]
- Laconi Andrea, van Beurden Steven J., Berends Alinda J., Krämer-Kühl Annika, Jansen Christine A., Spekreijse Dieuwertje, Chénard Gilles, Philipp Hans-Christian, Mundt Egbert, Rottier Peter J. M., Hélène Verheije M. Deletion of accessory genes 3a, 3b, 5a or 5b from avian coronavirus infectious bronchitis virus induces an attenuated phenotype both in vitro and in vivo. Journal of General Virology. 2018;99(10):1381–1390. doi: 10.1099/jgv.0.001130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai MMC, Cavanagh D. The molecular biology of coronaviruses. Adv Virus Res. 1997;48:1–100. doi: 10.1016/S0065-3527(08)60286-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Kang H, Liu P, Makkinje N, Williamson ST, Leibowitz JL, Giedroc DP. Structural lability in stem-loop 1 drives a 5′ UTR-3′ UTR interaction in coronavirus replication. J Mol Biol. 2008;377:790–803. doi: 10.1016/j.jmb.2008.01.068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Xue C, Chen F, Qin J, Xie Q, Bi Y, Cao Y. Isolation and genetic analysis revealed no predominant new strains of avian infectious bronchitis virus circulating in South China during 2004–2008. Vet Microbiol. 2010;143:145–154. doi: 10.1016/j.vetmic.2009.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim TH, Lee HJ, Lee DH, Lee YN, Park JK, Youn HN, Kim MS, Lee JB, Park SY, Choi IS. An emerging recombinant cluster of nephropathogenic strains of avian infectious bronchitis virus in Korea. Infect Genet Evol. 2011;11:678–685. doi: 10.1016/j.meegid.2011.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin KH, Lin CF, Chiou SS, Hsu AP, Lee MS, Chang CC, Chang TJ, Shien JH, Hsu WL. Application of purified recombinant antigenic spike fragments to the diagnosis of avian infectious bronchitis virus infection. Appl Microbiol Biotechnol. 2012;95:233–242. doi: 10.1007/s00253-012-4143-8. [DOI] [PubMed] [Google Scholar]
- Liu S, Zhang X, Wang Y, Li C, Liu Q, Han Z, Zhang Q, Kong X, Tong G. Evaluation of the protection conferred by commercial vaccines and attenuated heterologous isolates in China against the CK/CH/LDL/97I strain of infectious bronchitis coronavirus. Vet J. 2009;179:130–136. doi: 10.1016/j.tvjl.2007.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu H, Yang X, Zhang Z, Li J, Zou W, Zeng F, Wang H. Comparative transcriptome analysis reveals induction of apoptosis in chicken kidney cells associated with the virulence of nephropathogenic infectious bronchitis virus. Microb Pathog. 2017;113:451–459. doi: 10.1016/j.micpath.2017.11.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopes PD, Okino CH, Fernando FS, Pavani C, Casagrande VM, Lopez R, Montassier HJ. Inactivated infectious bronchitis virus vaccine encapsulated in chitosan nanoparticles induces mucosal immune responses and effective protection against challenge. Vaccine. 2018;36:2630–2636. doi: 10.1016/j.vaccine.2018.03.065. [DOI] [PubMed] [Google Scholar]
- Lucio B, Fabricant J. Tissue tropism of three cloacal isolates and Massachusetts strain of infectious bronchitis virus. Avian Dis. 1990;34:865–870. doi: 10.2307/1591375. [DOI] [PubMed] [Google Scholar]
- Luo H, Qin J, Chen F, Xie Q, Bi Y, Cao Y, Xue C. Phylogenetic analysis of the S1 glycoprotein gene of infectious bronchitis viruses isolated in China during 2009–2010. Virus Genes. 2012;44:19–23. doi: 10.1007/s11262-011-0657-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma H, Shao Y, Sun C, Han Z, Liu X, Guo H, Liu X, Kong X, Liu S. Genetic diversity of avian infectious bronchitis coronavirus in recent years in China. Avian Dis. 2012;56:15–28. doi: 10.1637/9804-052011-Reg.1. [DOI] [PubMed] [Google Scholar]
- Macdonald JW, Randall CJ, Mcmartin DA, Dagless MD. Immunity following vaccination with the H120 strain of infectious bronchitis virus via the drinking water. Avian Pathol. 1981;10:295–301. doi: 10.1080/03079458108418478. [DOI] [PubMed] [Google Scholar]
- Marandino A, Pereda A, Tomas G, Hernández M, Iraola G, Craig MI, Hernandez D, Banda A, Villegas P, Panzera Y, Perez R. Phylodynamic analysis of avian infectious bronchitis virus in South America. J Gen Virol. 2015;96:1340–1346. doi: 10.1099/vir.0.000077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marquardt WW, Snyder DB, Schlotthober BA. Detection and quantification of antibodies to infectious bronchitis virus by enzyme-linked immunosorbent assay. Avian Dis. 1981;25:713–722. doi: 10.2307/1590002. [DOI] [PubMed] [Google Scholar]
- Matoo JJ, Bashir K, Kumar A, Krishnaswamy N, Dey S, Chellappa MM, Ramakrishnan S. Resiquimod enhances mucosal and systemic immunity against avian infectious bronchitis virus vaccine in the chicken. Microb Pathog. 2018;119:119–124. doi: 10.1016/j.micpath.2018.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthijs MGR, Van Eck JHH, Landman WJM, Stegeman JA. Ability of Massachusetts-type infectious bronchitis virus to increase colibacillosis susceptibility in commercial broilers: a comparison between vaccine and virulent field virus. Avian Pathol. 2003;32:473–481. doi: 10.1080/0307945031000154062. [DOI] [PubMed] [Google Scholar]
- Mondal SP, Naqi SA. Maternal antibody to infectious bronchitis virus: its role in protection against infection and development of active immunity to vaccine. Vet Immunol Immunopathol. 2001;79:31–40. doi: 10.1016/S0165-2427(01)00248-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura K, Cook JK, Otsuki K, Huggins MB, Frazier JA. Comparative study of respiratory lesions in two chicken lines of different susceptibility infected with infectious bronchitis virus: histology, ultrastructure and immunohistochemistry. Avian Pathol. 1991;20:241–257. doi: 10.1080/03079459108418761. [DOI] [PubMed] [Google Scholar]
- Nakamura K, Ueda H, Tanimura T, Noguchi K. Effect of mixed live vaccine (Newcastle disease and infectious bronchitis) and Mycoplasma gallisepticum on the chicken respiratory tract and on Escherichia coli infection. J Comp Pathol. 1994;111:33–42. doi: 10.1016/S0021-9975(05)80109-4. [DOI] [PubMed] [Google Scholar]
- Niesters HG, Bleumink-Pluym NM, Osterhaus AD, Horzinek MC, van der Zeijst BA. Epitopes on the peplomer protein of infectious bronchitis virus strain M41 as defined by monoclonal antibodies. Virology. 1987;161:511–519. doi: 10.1016/0042-6822(87)90145-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nii T, Isobe N, Yoshimura Y. Effects of avian infectious bronchitis virus antigen on eggshell formation and immunoreaction in hen oviduct. Theriogenology. 2014;81:1129–1138. doi: 10.1016/j.theriogenology.2014.02.002. [DOI] [PubMed] [Google Scholar]
- Okino CH, Dos Santos IL, Fernando FS, Alessi AC, Wang X, Montassier HJ. Inflammatory and cell-mediated immune responses in the respiratory tract of chickens to infection with avian infectious bronchitis virus. Viral Immunol. 2014;27:383–391. doi: 10.1089/vim.2014.0054. [DOI] [PubMed] [Google Scholar]
- Otsuki K, Nakamura T, Kubota N, Kawaoka Y, Tsubokura M. Comparison of two strains of avian infectious bronchitis virus for their interferon induction, viral growth and development of virus-neutralizing antibody in experimentally-infected chickens. Vet Microbiol. 1987;15:31–40. doi: 10.1016/0378-1135(87)90126-X. [DOI] [PubMed] [Google Scholar]
- Patel BH, Bhimani MP, Bhanderi BB, Jhala MK. Isolation and molecular characterization of nephropathic infectious bronchitis virus isolates of Gujarat state, India. Virus Dis. 2015;26:42–47. doi: 10.1007/s13337-015-0248-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pei J, Collisson EW. Specific antibody secreting cells from chickens can be detected by three days and memory B cells by three weeks post-infection with the avian respiratory coronavirus. Dev Comp Immunol. 2005;29:153–160. doi: 10.1016/j.dci.2004.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Posada D, Crandall KA. Evaluation of methods for detecting recombination from DNA sequences: computer simulations. Proc Natl Acad Sci U S A. 2001;98:13757–13762. doi: 10.1073/pnas.241370698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Promkuntod N, Wickramasinghe INA, de Vrieze G, Gröne A, Verheije MH. Contributions of the S2 spike ectodomain to attachment and host range of infectious bronchitis virus. Virus Res. 2013;177:127–137. doi: 10.1016/j.virusres.2013.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Promkuntod N, Thongmee S, Yoidam S. Analysis of the S1 gene of the avian infectious bronchitis virus (IBV) reveals changes in the IBV genetic groups circulating in southern Thailand. Res Vet Sci. 2015;100:299–302. doi: 10.1016/j.rvsc.2015.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raggi LG, Lee GG. Lack of correlation between infectivity, serologic response and challenge results in immunization with an avian infectious bronchitis vaccine. J Immunol. 1965;94:538–543. [PubMed] [Google Scholar]
- Raj GD, Jones RC. Local antibody production in the oviduct and gut of hens infected with a variant strain of infectious bronchitis virus. Vet Immunol Immunopathol. 1996;53:147–161. doi: 10.1016/0165-2427(95)05545-2. [DOI] [PubMed] [Google Scholar]
- Raj GD, Jones RC. Infectious bronchitis virus: immunopathogenesis of infection in the chicken. Avian Pathol. 1997;26:677–706. doi: 10.1080/03079459708419246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raj GD, Savage CE, Jones RC. Effect of heterophil depletion by 5-fluorouracil on infectious bronchitis virus infection in chickens. Avian Pathol. 1997;26:427–432. doi: 10.1080/03079459708419224. [DOI] [PubMed] [Google Scholar]
- Schalk A, Hawn M. An apparently new respiratory disease of baby chicks. J Am Vet Med Assoc. 1931;78:413–422. [Google Scholar]
- Seger W, Ghalyanchi Langeroudi A, Karimi V, Madadgar O, Marandi MV, Hashemzadeh M. Genotyping of infectious bronchitis viruses from broiler farms in Iraq during 2014–2015. Arch Virol. 2016;161:1229–1237. doi: 10.1007/s00705-016-2790-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seo SH, Collisson EW. Specific cytotoxic T lymphocytes are involved in in vivo clearance of infectious bronchitis virus. J Virol. 1997;71:5173–5177. doi: 10.1128/jvi.71.7.5173-5177.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimazaki Y, Watanabe Y, Harada M, Seki Y, Kuroda Y, Fukuda M, Honda E, Suzuki S, Nakamura S. Genetic analysis of the S1 gene of 4/91 type infectious bronchitis virus isolated in Japan. J Vet Med Sci. 2009;71:583–588. doi: 10.1292/jvms.71.583. [DOI] [PubMed] [Google Scholar]
- Sumi V, Singh SD, Dhama K, Gowthaman V, Barathidasan R, Sukumar K. Isolation and molecular characterization of infectious bronchitis virus from recent outbreaks in broiler flocks reveals emergence of novel strain in India. Trop Anim Health Prod. 2012;44:1791–1795. doi: 10.1007/s11250-012-0140-2. [DOI] [PubMed] [Google Scholar]
- Toffan A, Monne I, Terregino C, Cattoli G, Hodobo CT, Gadaga B, Makaya PV, Mdlongwa E, Swiswa S. QX-like infectious bronchitis virus in Africa. Vet Rec. 2011;169:589. doi: 10.1136/vr.d7636. [DOI] [PubMed] [Google Scholar]
- Toro H, Zhang JF, Gallardo RA, van Santen VL, van Ginkel FW, Joiner KS, Breedlove C. S1 of distinct IBV population expressed from recombinant adenovirus confers protection against challenge. Avian Dis. 2014;58:211–215. doi: 10.1637/10670-091913. [DOI] [PubMed] [Google Scholar]
- Ullah S, Riaz N, Umar S, Shah MAA. DNA vaccines against avian influenza: current research and future prospects. Worlds Poult Sci J. 2013;69:125–133. doi: 10.1017/S0043933913000111. [DOI] [Google Scholar]
- van Beurden SJ, Berends AJ, Krämer-Kühl A, Spekreijse D, Chenard G, Philipp HC, Mundt E, Rottier PJM, Verheije MH. Recombinant live attenuated avian coronavirus vaccines with deletions in the accessory genes 3ab and/or 5ab protect against infectious bronchitis in chickens. Vaccine. 2018;36:1085–1092. doi: 10.1016/j.vaccine.2018.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villarreal LYB. Diagnosis of infectious bronchitis: an overview of concepts and tools. Rev Bras Cienc Avic. 2010;12:111–114. doi: 10.1590/S1516-635X2010000200006. [DOI] [Google Scholar]
- Wang X, Rosa AJ, Oliverira HN, Rosa GJ, Guo X, Travnicek M, Girshick T. Transcriptome of local innate and adaptive immunity during early phase of infectious bronchitis viral infection. Viral Immunol. 2006;19:768–774. doi: 10.1089/vim.2006.19.768. [DOI] [PubMed] [Google Scholar]
- Wilson K, Gage P, Ewart G. Hexamethylene amiloride blocks E protein ion channels and inhibits coronavirus replication. Virology. 2006;353:294–306. doi: 10.1016/j.virol.2006.05.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winterfield RW, Hitchner SB. Etiology of an infectious nephritis-nephrosis syndrome of chickens. Am J Vet Res. 1962;23:1273–1279. [PubMed] [Google Scholar]
- Xu G, Liu XY, Zhao Y, Chen Y, Zhao J, Zhang GZ. Characterization and analysis of an infectious bronchitis virus strain isolated from southern China in 2013. Virol J. 2016;13:40. doi: 10.1186/s12985-016-0497-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan SH, Chen Y, Zhao J, Xu G, Zhao Y, Zhang GZ. Pathogenicity of a TW-like strain of infectious bronchitis virus and evaluation of the protection induced against it by a QX-like strain. Front Microbiol. 2016;7:1653. doi: 10.3389/fmicb.2016.01653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan S, Zhao J, Xie D, Huang X, Cheng J, Guo Y, Liu C, Ma Z, Yang H, Zhang G. Attenuation, safety, and efficacy of a QX-like infectious bronchitis virus serotype vaccine. Vaccine. 2018;36:1880–1886. doi: 10.1016/j.vaccine.2018.02.053. [DOI] [PubMed] [Google Scholar]
- Yang X, Li J, Liu H, Zhang P, Chen D, Men S, Li X, Wang H. Induction of innate immune response following introduction of infectious bronchitis virus (IBV) in the trachea and renal tissues of chickens. Microb Pathog. 2018;116:54–61. doi: 10.1016/j.micpath.2018.01.008. [DOI] [PubMed] [Google Scholar]
- Yudong W, Yongling W, Zichun Z, Gencheng F, Yihau J, Xiange L, Jiang D, Wang S. Isolation and identification of glandular stomach type IBV (QX IBV) in chickens. Chin J Anim Quar. 1998;15:1–3. [Google Scholar]
- Zanaty A, Arafa AS, Hagag N, El-Kady M. Genotyping and pathotyping of diversified strains of infectious bronchitis viruses circulating in Egypt. World J Virol. 2016;5:125–134. doi: 10.5501/wjv.v5.i3.125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang W, Bouwman KM, van Beurden SJ, Ordonez SR, van Eijk M, Haagsman HP, Verheije MH, Veldhuizen EJ. Chicken mannose binding lectin has antiviral activity towards infectious bronchitis virus. Virology. 2017;509:252–259. doi: 10.1016/j.virol.2017.06.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang P, Liu X, Liu H, Wang W, Liu X, Li X, Wu X. Astragalus polysaccharides inhibit avian infectious bronchitis virus infection by regulating viral replication. Microb Pathog. 2018;114:124–128. doi: 10.1016/j.micpath.2017.11.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zou NL, Zhao FF, Wang YP, Liu P, Cao SJ, Wen XT, Huang Y. Genetic analysis revealed LX4 genotype strains of avian infectious bronchitis virus became predominant in recent years in Sichuan area, China. Virus Genes. 2010;41:202–209. doi: 10.1007/s11262-010-0500-9. [DOI] [PMC free article] [PubMed] [Google Scholar]