

Abstract
Angiotensin-converting enzyme 2 (ACE2) is a recently described member of the renin-angiotensin system that hydrolyzes angiotensin (Ang) II to Ang-(1-7), and may thereby protect against cardiovascular and renal diseases. ACE2 is a type 1 integral membrane protein and contains a catalytically active ectodomain that can be shed from the cell surface into the extracellular space, via cleavage by a disintegrin and metalloproteinase-17 (ADAM-17). ACE2 enzymatic activity and protein can be detected in biological fluids, including urine, plasma, and conditioned cell culture media. We present a detailed method for measurement of ACE2 activity in biological fluids, using hydrolysis of an intramolecularly quenched fluorogenic ACE2 substrate, in the absence or presence of the ACE2 inhibitors MLN-4760 or DX600. Recombinant human or mouse ACE2 is used to generate standard curves for this assay, with ACE2 detection ranging from 1.56 to 50 ng/ml. While MLN-4760 potently inhibits the activity of both human and mouse ACE2, DX600 (linear form) only effectively blocks human ACE2 activity in this assay. In biological samples of human and mouse urine, cell culture medium from mouse proximal tubular cells, and mouse plasma, the mean intra- and inter-assay coefficients of variation (CVs) of the assay range from 1.43 to 4.39 %, and from 7.01 to 13.17 %, respectively. We present data on the time and substrate concentration dependence of the assay, and show that exogenous d -glucose, creatinine, urea, and albumin do not interfere with its performance. In biological fluids, this assay is a simple and reliable method to study the role of ACE2 and its shed fragments in cardiovascular and renal diseases.
Key words: Angiotensin-converting enzyme 2, Substrate, Fluorescence, Plasma, Urine, Proximal tubule
Introduction
Angiotensin-converting enzyme 2 (ACE2), a zinc-metallopeptidase, is a recently identified member of the renin-angiotensin system that degrades angiotensin (Ang) II to Ang-(1- 7) [1, 2]. ACE2 is also a receptor for the coronavirus that causes severe acute respiratory syndrome (SARS) [3]. Although ACE2 is found in many tissues, it is highly expressed in the kidney, particularly within cells of the proximal tubule [4, 5]. Because it decreases levels of the vasoconstrictor Ang II and generates the vasodilator Ang-(1-7), ACE2 may protect against hypertension [6] and renal disease [7, 8]. ACE2 has also been shown to prevent acute lung injury [9].
ACE2 is shed at its carboxy-terminus from the plasma membrane in cells via “a disintegrin and metalloproteinase-17” (ADAM-17) pathway [10, 11]. Recently, soluble ACE2 has been detected in certain biological fluids, including urine, plasma, and cell culture medium, by enzyme-linked immunosorbent assay ( ELISA), enzyme activity assay, or western analysis [10, 12, 13]. In this chapter, we describe a detailed method for measurement of ACE2 enzyme activity in biological fluids using a commercially available synthetic fluorogenic substrate for ACE2. We provide a practical, cost-effective, and high-throughput method to study the potential role of ACE2 shedding in renal and cardiovascular disease.
Materials
The following reagents must be prepared and brought to room temperature before starting the assay.
Assay buffer: 50 mM 2-(N-Morpholino)ethanesulfonic acid (MES), 300 mM NaCl, 10 μM ZnCl2, adjust pH to 6.8. Store at 4 °C when not in use.
Fluorogenic ACE2 substrate: Mca-Ala-Pro-Lys(Dnp)-OH (AnaSpec, San Jose, CA, USA, Catalog Number: 60757), reconstitute in 1 % NH4OH to 15 mM. Aliquoted samples can be stored at −20 °C for at least 3 months or −80 °C for at least 6 months.
: N-ethylmaleimide (NEM, 100 mM stock in double-distilled H2O [ddH2O]) and phenylmethylsulfonyl fluoride (PMSF, 100 mM stock in ethanol). Aliquot and store at −20 °C.
2 inhibitor: MLN-4760 (10−5 M stock in ddH2O, also called GL1001, provided by Ore Pharmaceuticals, Cambridge, MA, USA, and commercially available from Calbiochem, San Diego, CA, USA, Catalog Number: 530616) or DX600 (linear form, 10−5 M stock in ddH2O, AnaSpec, Catalog Number: 62337) .
ACE inhibitor: Captopril (10−4 M stock in ddH2O, Sigma-Aldrich, St. Louis, MO, USA).
ACE2 Standard: Recombinant human ACE2 (human rACE2, R&D Systems Inc., Minneapolis, MN, USA, Catalog Number: 933-ZN) and recombinant mouse ACE2 (mouse rACE2, R&D Systems Inc., Catalog Number: 3437-ZN). Reconstitute with phosphate-buffered saline (PBS) to 10 μg/ml. Aliquots are stable for at least 3 months at −20 °C and at least 6 months at −80 °C.
96-well Microplate (black).
Fluorescence Microplate Reader: We use the FLUOstar Galaxy fluorometer (BMG Labtechnologies, Durham, NC, USA), detecting emission at 405 nm with excitation at 320 nm.
Methods
In the following section, we describe the ACE2 activity assay. First, we present the generation of the standard curve (Subheading 3.1), followed by measurement of ACE2 activity in biological fluids (Subheading 3.2).
Generation of Standard Curves Using Human or Mouse rACE2
Preparation of ACE2 Substrate/Assay Buffer Solution
Calculate the amount of working reagents to use. Add fluorogenic ACE2 substrate and protease inhibitors to the assay buffer (see Subheading 2, item 1) immediately before each experiment, to generate the ACE2 substrate/Assay buffer solution, to achieve concentrations as follows: 15 μM ACE2 substrate, 1 mM NEM, and 1 mM PMSF (Table 1). Seventy-five micro liter of the ACE2 substrate/assay buffer solution will be used in each reaction (see below Subheading 3.1.3). The protease inhibitors are added to prevent substrate hydrolysis in biological solutions [14]. For the standard curve generation, we also routinely add protease inhibitors to the assay buffer .
Table 1.
ACE2 substrate/assay buffer solutiona
| Components (stock solutions) | Volume (μl) | Conc. in substrate/assay buffer mix |
|---|---|---|
| ACE2 substrate (15 mM in 1 % NH4OH) | 1 | 15 μM |
| NEM (100 mM in ddH2O) | 10 | 1 mM |
| PMSF (100 mM in ethanol) | 10 | 1 mM |
| Assay buffer (see Subheading 2, item 1) | 979 | – |
| Total volume (μl) | 1000 |
NEM N-ethylmaleimide, PMSF phenylmethylsulfonyl fluoride, Conc concentration
a1000 μl provides sufficient buffer for 3 measurements performed in duplicate (75 μl per well)
Preparation of ACE2 Standard Dilutions
Dilute 10 μg/ml stock of human or mouse rACE2 1:30 in assay buffer to a concentration of 333.33 ng/ml for standard #1 (Table 2). Perform 1:2 serial dilutions of the preceding ACE2 standard in assay buffer for standards #2-6, with the lowest concentration at 10.42 ng/ml. Standard #7 is a blank that should contain no rACE2 (instead add 15 μl assay buffer alone). Add 15 μl/well of the serially diluted ACE2 standards into a total volume of 100 μl of ACE2 enzymatic reaction solution (see Table 2) to achieve final ACE2 concentrations of 50, 25, 12.5, 6.25, 3.13, and 1.56 ng/ml.
Table 2.
Components of ACE2 activity standard curve solution (per well)a
| rACE2 standard | Diluted rACE2 conc. (ng/ml) | rACE2 volume/well (μl) | ACE2 inhibitorb or ddH2O volume/well (μl)c | ACE2 substrate/assay buffer (μl) | Total volume/well (μl) | Final rACE2 conc. (ng/ml) |
|---|---|---|---|---|---|---|
| 1 | 333.33 | 15 | 10 | 75 | 100 | 50 |
| 2 | 166.66 | 15 | 10 | 75 | 100 | 25 |
| 3 | 83.33 | 15 | 10 | 75 | 100 | 12.5 |
| 4 | 41.67 | 15 | 10 | 75 | 100 | 6.25 |
| 5 | 20.83 | 15 | 10 | 75 | 100 | 3.13 |
| 6 | 10.42 | 15 | 10 | 75 | 100 | 1.56 |
| 7 | 0 | 15d | 10 | 75 | 100 | 0 |
rACE2 recombinant ACE2, ddH 2 O double-distilled H2O, Conc concentration
aThe volume of each component listed is for each well, to give a total volume of 100 μl of ACE2 enzymatic reaction solution, on a 96-well microplate. Each reaction is performed in duplicate
bACE2 inhibitor: MLN-4760 or DX600
cFor each dilution, one duplicate reaction is performed in ddH2O (10 μl), and another duplicate is performed with the addition of ACE2 inhibitor MLN-4760 or DX600 (10 μl, from stock of 10−5 M)
dStandard #7 is a blank, with the addition of assay buffer alone, instead of rACE2
Measurement of ACE2 Activity in Standard Dilutions
Set up the plate configuration on a 96-well black microplate: the standards are run in duplicates, along with a blank control that contains no rACE2. For each concentration of ACE2 standard, two wells are used for total ACE2 activity, and two wells are used for measurement of activity in the presence of the ACE2 inhibitor MLN-4760 or DX600. The volumes of each individual component per well are shown in Table 2. The final volume of the ACE2 enzymatic reaction is 100 μl/well, and the sequence of addition of components to the plate is as follows:
Add 10 μl of ddH2O into each of the total enzymatic activity reaction wells of a 96-well microplate. For the inhibitor wells, instead add 10 μl of ACE2 inhibitor solution (10−5 M MLN-4760 or 10−5 M DX600 stock).
Add 75 μl of the ACE2 substrate/assay buffer (see Subheading 3.1.1) to all wells.
Add 15 μl of diluted ACE2 standards to each well, thereby achieving a total reaction volume of 100 μl for each well.
Cover the plate with microplate sealing film, followed by aluminum foil to prevent access to light, and incubate the plate at room temperature for 16 h on a plate shaker .
Read the 96-well plate on a fluorescence reader with excitation wavelength of 320 nM and emission wavelength of 405 nM.
For determination of the percentage of inhibition of ACE2 activity by MLN-4760 or DX600, subtract the Relative Fluorescence Unit (RFU) reading of the substrate blank from the readings for each well. The ACE2-specific activity for each standard is determined by subtracting the RFU obtained in the presence of MLN-4760 or DX600 from the reading in the absence of inhibitor.
Generation of Standard Curves
Construct a standard curve by plotting the known concentrations (X axis) of rACE2 standards versus the RFUs (Y axis) of rACE2 standards (Fig. 1). A highly linear relationship is observed between the RFU and rACE2 concentration for both human rACE2 (R 2 = 0.9998, p < 0.001 for MLN-4760, R 2 = 0.9999, p < 0.001 for DX600, Fig. 1a) and mouse rACE2 (R 2 = 0.9976, p < 0.001 for MLN-4760, Fig. 1b), with a range of ACE2 detection from 1.56 to 50 ng/ml. The linear equations generated for human and mouse rACE2 are used to convert the RFU to ACE2 concentrations in human or mouse biological fluids, respectively. In our hands, the RFU signal reaches saturation when the rACE2 concentration is greater than 50 ng/ml. The RFU signal becomes very weak or undetectable when the rACE2 concentration is less than 1.56 ng/ml. This range approximates the detection limits for human ACE2 using a commercial ELISA, which we previously reported to range from 0.39 to 25 ng/ml [13].
Fig. 1.
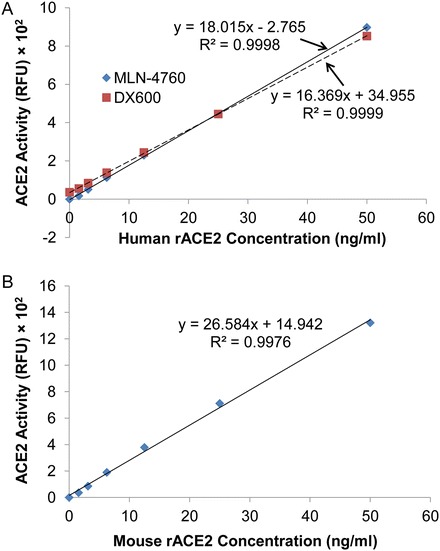
Standard curves for ACE2 activity assay. The assay was performed for 16 h incubation. Graph shows the concentrations of recombinant ACE2 (rACE2) versus the ACE2 activity (RFU). (a) Standard curves generated for human rACE2. The ACE2 inhibitors MLN-4760 (10−6 M) or DX600 (10−6 M) were used. The relationship is highly linear for rACE2 concentrations between 0 and 50 ng/ml (R 2 = 0.9998, p < 0.001 for MLN-4760, R 2 = 0.9999, p < 0.001 for DX600). (b) Standard curve generated for mouse rACE2. The ACE2 inhibitor MLN-4760 (10−6 M) was used. A highly linear relationship exists for rACE2 concentrations between 0 and 50 ng/ml (R 2 = 0.9976, p < 0.001)
Measurement of ACE2 Activity in Biological Fluids
Initial Preparation of Biological Samples
Using this assay, we have measured ACE2 activity in human urine samples collected from renal transplant recipients [13]. We have also measured ACE2 activity in urine samples from male FVB/N mice, with and without streptozotocin (STZ)-induced diabetes, and in conditioned culture medium collected from primary cultures of mouse proximal tubular (PT) cells, after up to 72 h incubation in medium. Mouse proximal tubule (PT) cells derived from C57BL6 mice were grown in a defined medium of DMEM-F12 (1:1), supplemented with insulin (5 μg/ml), transferrin (5 μg/ml), selenium (5 ng/ml), hydrocortisone (50 nM), and 3,3′,5-triiodo-L-thyronine (2.5 nM), without fetal bovine serum (FBS). Serum has previously been demonstrated to contain ACE2 enzymatic activity, and should be avoided in this assay [15]. All biological samples are collected and placed on ice, aliquoted and then centrifuged at 12,000 × g for 5 min at 4 °C. Supernatants are collected, and stored at −80 °C until the assay is performed.
We also have measured ACE2 activity in plasma, from mice with or without STZ-induced diabetes [8]. Blood is collected in chilled eppendorf tubes, and plasma is separated by centrifugation at 3000 × g for 10 min at 4 °C, and stored at −80 °C until time of the assay.
Assay Conditions
We dilute urine samples at least 1:2 and plasma samples 1:7.5 with assay buffer. For conditioned cell culture medium, undiluted samples are used. For each well on the 96-well plate, use 15 μl of diluted urine (e.g., 7.5 μl urine plus 7.5 μl assay buffer), 15 μl of diluted plasma (2 μl plasma plus 13 μl assay buffer), and 15 μl undiluted culture medium in a final volume of 100 μl of enzymatic reaction. If ACE2 activity falls outside the detection limits of the assay, a lower or higher dilution may be required.
Measure the ACE2 activity (RFU) in biological fluids by following the same experimental procedure described for ACE2 standards (see Subheading 3.1.3).
Calculate the ACE2 concentrations from the RFU in biological fluids using the linear equations generated for human or mouse rACE2, as appropriate (Fig. 1). The ACE2 concentrations must be corrected for the dilution factor to obtain the actual concentrations in the undiluted samples. ACE2 concentration in biological fluids is typically expressed as ng/ml.
The amount of urinary ACE2 can be corrected for the creatinine concentration in the urine samples, as a control for urinary dilution, and reported as ng/μg creatinine. ACE2 activity in culture medium can be corrected for the cell protein amounts on the culture dishes, and reported as ng/μg protein.
Intra- and Inter-Assay Coefficients of Variability (CV) in Biological Fluids
We have determined the intra- and inter-assay CVs of the ACE2 activity assay for biological samples, and the results support the reliability of this assay. Three samples each of human and mouse urine, mouse PT cell culture medium, and mouse plasma were assayed on six separate occasions in duplicate. The mean intra-assay CV for the assay is 4.39 ± 0.74 % in human urine, 3.04 ± 0.61 % in mouse urine, 1.43 ± 0.20 % in cell culture medium, and 3.84 ± 0.63 % in mouse plasma. The mean inter-assay CV value is 13.17 ± 0.77 % in human urine, 7.01 ± 1.04 % in mouse urine, 9.47 ± 0.22 % in cell culture medium, and 10.92 ± 1.88 % in mouse plasma (Table 3).
Table 3.
Intra- and inter-assay coefficients of variability (CV)
| Samples | Volume/well (μl) | Mean ACE2 activity (ng/ml) | Intra-assay CV (%)a | Inter-assay CV (%)b | |
|---|---|---|---|---|---|
| Human urine | 1 | 7.5 | 57.65 | 5.87 | 11.70 |
| 2 | 7.5 | 167.28 | 3.64 | 14.29 | |
| 3 | 7.5 | 151.49 | 3.67 | 13.51 | |
| Mean ± SE | 4.39 ± 0.74 | 13.17 ± 0.77 | |||
| Mouse urine | 1 | 7.5 | 402.82 | 1.86 | 5.52 |
| 2 | 7.5 | 159.50 | 3.36 | 6.49 | |
| 3 | 7.5 | 175.34 | 3.90 | 9.01 | |
| Mean ± SE | 3.04 ± 0.61 | 7.01 ± 1.04 | |||
| Culture medium | 1 | 15 | 74.76 | 1.76 | 9.38 |
| 2 | 15 | 69.35 | 1.06 | 9.88 | |
| 3 | 15 | 100.72 | 1.49 | 9.14 | |
| Mean ± SE | 1.43 ± 0.20 | 9.47 ± 0.22 | |||
| Mouse plasma | 1 | 2 | 420.47 | 4.99 | 8.88 |
| 2 | 2 | 400.10 | 3.69 | 9.20 | |
| 3 | 2 | 162.49 | 2.83 | 14.67 | |
| Mean ± SE | 3.84 ± 0.63 | 10.92 ± 1.88 |
CV coefficient of variability, SE standard error of the mean
aThe intra-assay CV is an average value of the individual CVs for the duplicates in six measurements
bThe inter-assay CV is calculated for each sample by dividing the standard deviation of six measurements by the mean ACE2 activity of six measurements
Notes
We have determined the extent of inhibition of ACE2 activity by MLN-4760 (10−6 M) or DX600 (10−6 M, linear form, AnaSpec), using human and mouse rACE2. Specificity can also be assessed, and in this regard we have used the ACE inhibitor captopril. At concentrations of human and mouse rACE2 ranging from 0 to 400 ng/ml, ACE2 enzymatic activity (measured after 16 h) is not blocked by captopril (10−5 M) (Fig. 2). Indeed, as an internal control, one may add captopril (10−5 M) or another ACE inhibitor to each reaction well (as a component of the assay buffer) in the ACE2 assay.
Fig. 2.
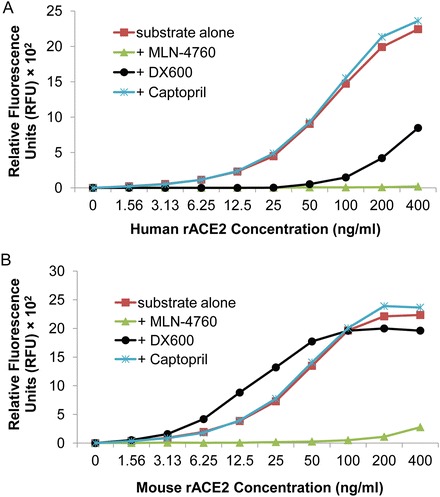
Effect of ACE2 inhibitors MLN-4760 (10−6 M), DX600 (10−6 M), and ACE inhibitor captopril (10−5 M) on ACE2 enzyme activity. The assay was performed for 16 h incubation. Graph depicts the concentrations of recombinant ACE2 versus the Relative Fluorescence Units (RFU). A background RFU value (67.5 RFU for a, 62.0 RFU for b) of the substrate blank control was subtracted from the readings for each well. (a) Effect of MLN-4760, DX600, and captopril on human rACE2 activity. (b) Effect of MLN-4760, DX600, and captopril on mouse rACE2 activity
Both MLN-4760 and DX600 strongly inhibit the activity of human rACE2 over a wide range of concentrations of the recombinant enzyme (1.56–400 ng/ml), with the degree of inhibition ranging from 74.5 to 99.0 % for MLN-4760, and from 62.2 to 100 % for DX600 (Fig. 2a). At high concentrations of human rACE2 (100–400 ng/ml), MLN-4760 has a more potent inhibitory effect (98.7–99.0 %) compared to DX600 (62.2–90.0 %).
It is important to note that only MLN-4760 (and not DX600) significantly blocks the activity of mouse rACE2, with the degree of inhibition ranging from 87.6 to 98.2 % (Fig. 2b). DX600 exerts no inhibitory effect on mouse rACE2 activity in this assay. Indeed, DX600 (10−6 M) actually enhanced RFU at lower levels of mouse rACE2 (3.13–50 ng/ml). In this regard, Pedersen et al. [16] measured the dissociation constant (Ki) between DX600 and human or mouse ACE2 in activity assays with the same ACE2 substrate, and reported the Ki for human ACE2 to be significantly lower (0.040 ± 0.005 μM) than the Ki for mouse ACE2 (0.36 ± 0.03 μM), suggesting that DX600 has higher affinity for human ACE2. The inhibitory effect of another commercially available conformational variant of DX600 (cyclic form) on ACE2 has not yet been tested in our assay. However, Ye et al. have reported that the disulfide bridged cyclic variant of DX600 (Bachem) had no inhibitory effect on mouse or rat ACE2, even at high concentrations, but effectively inhibited human rACE2 [17].
We have also examined the dose-dependent effects of MLN-4760 and DX600 (linear form) on ACE2 activity (measured at 16 h), using mouse rACE2 at concentrations from 0 to 50 ng/ml. An increase in MLN-4760 concentration from 10−6 to 10−5 M does not significantly enhance the assay characteristics (97.1–100 % inhibition for 10−5 M, 95.0–99.3 % for 10−6 M). Indeed, a highly linear relationship between the ACE2 activity (RFU) and mouse rACE2 concentration is observed for both concentrations of MLN-4760 (R 2 = 0.9950, p < 0.001 for 10−5 M; R 2 = 0.9954, p < 0.001 for 10−6 M). In contrast, DX600 has no significant inhibitory effect on mouse rACE2 activity even at concentrations as high as 10−5 M. Similarly, two groups have reported that 10−6 M DX600 (purchased from Phoenix Pharmaceuticals, Belmont, CA, USA) only partly (and ineffectively) blocks mouse recombinant or mouse kidney ACE2 activity [16, 17]. However, significant inhibition of mouse ACE2 has been observed with DX600 at 10−5 M using an assay buffer maintained at pH 6.5 (16). In rat PT segments, we have used the linear form of DX600 (10−6 M) (obtained from Phoenix Pharmaceuticals) to demonstrate inhibition of Ang-(1- 7) formation from Ang-(1–10), suggesting inhibition of rat ACE2 [5]. In rat kidney cortex, Ye et al. showed that DX600 (10−6 M) only partly inhibited ACE2 activity, measured at pH 6.5 [17]. Accordingly, because of the relatively poor inhibition of rodent ACE2 by DX600, for mouse biological samples we use 10−6 M of MLN-4760 for all experiments. For the ACE2 activity assay in human biological fluids, either MLN-4760 or DX600 can be used.
To determine if concentrations of ACE2 fluorogenic substrate lower than 11.25 μM can be used for the assay, we have studied the effect of various substrate concentrations (ranging from 1.41 to 11.25 μM) on mouse rACE2 enzyme activity (Fig. 3). At low substrate concentrations (1.41 μM and 2.81 μM), mouse rACE2 activities are reduced to only 0–25 % of values obtained at 11.25 μM substrate concentration (Fig. 3a). On the other hand, a highly linear relationship between the activity (RFU) and rACE2 concentration is observed for substrate concentration of 5.63 μM or 11.25 μM (R 2 = 0.9994, p < 0.001 for 11.25 μM; R 2 = 0.9903, p < 0.001 for 5.63 μM), with the same detection limits (1.56–50 ng/ml) (Fig. 3b). Accordingly, in our assays with biological samples, we routinely use at least 11.25 μM of the ACE2 substrate, although use of 5.63 μM substrate concentration would be acceptable. However, we would not recommend use of ACE2 substrate concentrations lower than this.
Fig. 3.
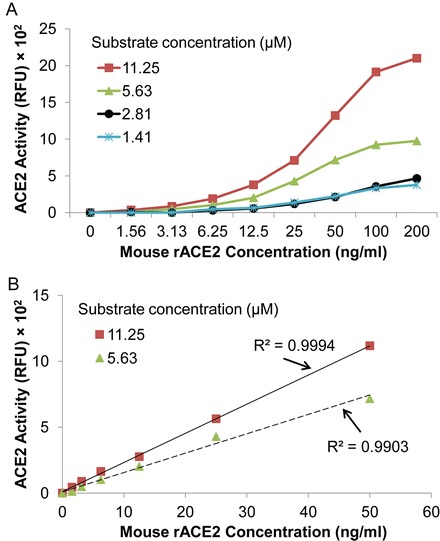
Effect of substrate concentrations on ACE2 activity. (a) The assay was performed with different ACE2 substrate concentrations (11.25, 5.63, 2.81, and 1.41 μM) for 16 h incubation using mouse rACE2 concentrations between 0 and 200 ng/ml. Graph depicts the concentrations of mouse rACE2 versus the activity (RFU) at different substrate concentrations. (b) Standard curves generated for mouse rACE2 with 11.25 or 5.63 μM ACE2 substrate concentrations. A highly linear relationship between mouse rACE2 and the RFU exists for both substrate concentrations (R 2 = 0.9994, p < 0.001 for 11.25 μM; R 2 = 0.9903, p < 0.001 for 5.63 μM)
Is it necessary to incubate samples for 16 h prior to measurement of fluorescence? We have addressed this question by measuring ACE2 activity at different time points (2, 6, 16, and 24 h incubation), with mouse rACE2 concentrations ranging from 0 to 200 ng/ml (Fig. 4a). A time-dependent increase in ACE2 activity is observed for all mouse rACE2 concentrations. By 16 h of incubation, the RFU signal has reached saturation for mouse rACE2 concentrations between 100 and 200 ng/ml. The linear detection range for ACE2 with 16 h incubation is between 1.56 and 50 ng/ml (Fig. 4b). In contrast, for a 2 h incubation time, a linear relationship still exists between ACE2 activity and ACE2 concentration for mouse rACE2 between 100 and 200 ng/ml (Fig. 4a). Thus, the detection limit of the assay for 2 h incubation is extended between 1.56 and 200 ng/ml (R 2 = 0.9981, p < 0.001), although RFU values are significantly lower at all ACE2 concentrations (see Fig. 4). In our lab, we routinely select 16 h as an incubation time for the assay, since ACE2 activity in most biologic samples we have studied is within the 16 h detection limit (i.e., up to 50 ng/ml). If there is a relatively high ACE2 protein level in the biological fluid of interest, the incubation time can be reduced to as short as 2 h .
Fig. 4.
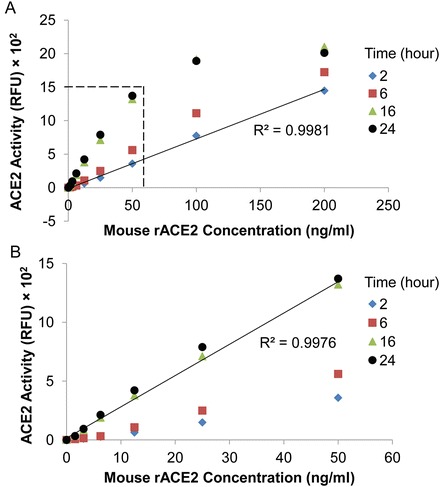
Time course of ACE2 activity assay. ACE2 activity was measured for 2, 6, 16, and 24 h incubation, with mouse rACE2 concentrations ranging from 0 to 200 ng/ml. Graph depicts the concentrations of mouse rACE2 versus the ACE2 activity (RFU) at different time points. (a) Standard curve generated for mouse rACE2 for 2 h incubation. A highly linear relationship exists for rACE2 concentrations from 0 to 200 ng/ml (R 2 = 0.9981, p < 0.001). The area marked with dashed lines is shown in b. (b) Standard curve generated for mouse rACE2 for 16 h incubation. The relationship is highly linear for rACE2 concentrations between 0 and 50 ng/ml
As noted, using this assay we perform endpoint RFU measurements, calculate the ACE2 protein concentrations from standard curves, and typically record this value in ng/ml for biological fluids, and not as ACE2 activity per se. ACE2 activity can also be reported as RFUs per volume of sample per unit time (RFU/ml/min or RFU/ml/h). To present data as ACE2 enzymatic activity, the 96-well plate should be read on a fluorescence reader in kinetic mode. The maximum velocity (V max, RFU/min) in each sample can be determined, and adjusted for the inhibitor-containing wells. Specific ACE2 activity (pmole/min) is then calculated from the adjusted V max using a conversion factor (pmol/RFU), determined from a calibration standard curve constructed using a fluorescent peptide fragment (MCA-Pro-Leu-OH, Bachem, Bubendorf, Switzerland, Catalog number M-1975). Final ACE2 enzymatic activity in biological fluids can be corrected for the sample dilution factor, and expressed as pmole/ml/min.
Pedersen et al. [16] have reported that activity assays of human, rat, and mouse ACE2 are highly pH-dependent, with maximal hydrolysis rate of ACE2 substrate at assay buffer pH 6.5, and minimal activity at pH 8.0 [16]. We use an assay buffer at pH 6.8 for our ACE2 activity assay, and we recommend measuring the pH of biological samples prior to conduct of the assay. To limit any effect of pH of biological samples on the assay, we advise diluting samples with the assay buffer as much as possible.
For measurement of plasma ACE2 activity, avoidance of chelators such as EDTA in the blood collection tubes is recommended, since chelation could inhibit the activity of the metallopeptidase ACE2.
Urine samples may have significant background fluorescence at 405 nm. For example, in undiluted urine samples from mice with gene deletion of ACE2, we typically obtain background fluorescence readings greater than 2000 RFU for 7.5 μl urine in the assay. Because such autofluorescence may interfere with the fluorescence signal and reduce the ability to detect ACE2, we suggest dilution of urine samples (e.g., in wild-type mice we typically dilute the urine sample 1:15 with assay buffer to 15 μl/well) .
Does freeze/thaw of biological fluids affect ACE2 activity? We have measured ACE2 activity in samples of human and mouse urine, and mouse PT cell culture medium after as many as three freeze/thaw cycles. Interestingly, we found no significant difference in ACE2 activity between any freeze/thaw cycle for all three sample categories (data not shown, p > 0.05). Nonetheless, for biological samples it is likely prudent to restrict use of repetitive freeze/thaw for this assay. Furthermore, we do not recommend repetitively freezing and thawing recombinant ACE2 protein, as we found that ACE2 activity can be significantly lost after a second freeze-thaw cycle.
In previous studies, we reported that urinary ACE2 activity is enhanced in renal transplant patients with diabetes, using this enzyme activity assay [13]. In preliminary data, we also found that incubation of mouse PT cells with high glucose concentrations enhances ACE2 activity in the culture medium, suggesting increased shedding of ACE2 from the cell membrane (data not shown). Accordingly, we have determined if high glucose concentrations alone can affect the assay. As shown in Fig. 5a, ACE2 activity in mouse PT cell culture medium with a high d -glucose concentration (25 mM) is not significantly different from that in the same medium with a normal d -glucose concentration (7.8 mM) (p > 0.05, n = 3).
Fig. 5.
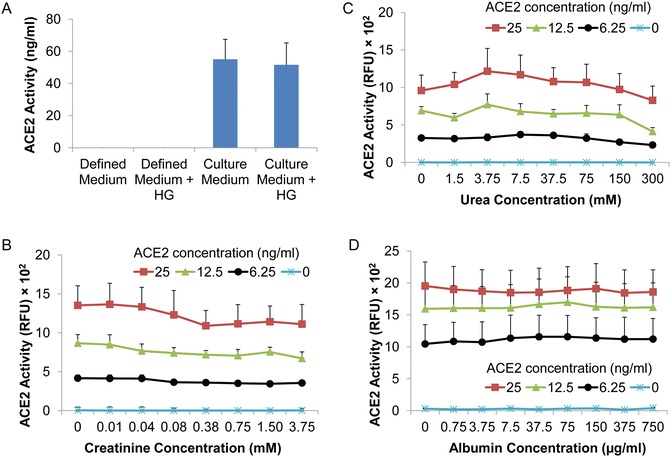
Effect of potential modulators on ACE2 activity assay. The assay was performed for 16 h incubation. (a) Graph shows ACE2 activity in defined medium (not incubated with mouse PT cells), and culture medium collected from primary cultures of mouse PT cells after 72 h incubation, in the presence of normal (7.8 mM) or high concentrations of d-glucose (HG, 25 mM). No ACE2 activity is detected in the defined medium. Results are means ± SEM; p > 0.05, culture medium vs culture medium + HG; n = 3. (b) Graph depicts the effect of addition of various concentrations of creatinine on ACE2 activity (RFU) at different mouse rACE2 concentrations. Results are means ± SEM; p > 0.05 for all graphs; n = 3. (c) Graph depicts the effect of addition of various concentrations of urea on ACE2 activity (RFU) at different mouse rACE2 concentrations. Results are means ± SEM; p > 0.05 for all graphs; n = 3. (d) Graph depicts the effect of addition of exogenous albumin on ACE2 activity (RFU) at different mouse rACE2 concentrations. Albumin at the indicated concentrations was added to urine samples (1 μl) from mice with gene deletion of ACE2. Results are means ± SEM; p > 0.05 for all graphs; n = 3
We have also studied the effect of increasing concentrations of creatinine (Sigma-Aldrich) and urea (Sigma-Aldrich) on the assay, since these substances are relatively abundant in urine. Using mouse rACE2, we found that neither creatinine (0.01–3.75 mM) nor urea (1.5–300 mM) had any significant effect on ACE2 activity when added to assay buffer (Fig. 5b, c).
Finally, since we reported a correlation between urinary protein and urinary ACE2 levels [13], we have examined the effect of exogenous albumin on the ACE2 activity assay. For these experiments, increasing concentrations of albumin (Cohn Fraction V, Sigma-Aldrich, Catalog number A8806) were added to urine samples (1 μl/well) from C57BL6 mice with ACE2 gene deletion, and ACE2 activity was measured following incubation with increasing concentrations of mouse rACE2 (0–25 ng/ml). For these experiments, the average albumin concentration in the urine samples from mice with ACE2 gene deletion was 0.15 μg/ml. As shown in Fig. 5d, addition of exogenous albumin (0.75–750 μg/ml) had no effect on rACE2 activity .
Acknowledgments
We thank Dr. Susan B. Gurley (Duke University Medical Center, Durham, NC, USA) for providing the ACE2 gene knockout mice. We also thank Ore Pharmaceuticals (Cambridge, MA, USA) for providing the ACE2 inhibitor MLN-4760.
Contributor Information
Rhian M. Touyz, Phone: +116135625800, FAX: +116135625487, Email: rhian.touyz@glasgow.ac.uk
Ernesto L. Schiffrin, Phone: +115143407538, FAX: +115143407539, Email: ernesto.schiffrin@mcgill.ca
Kevin D. Burns, Email: kburns@ottawahospital.on.ca
References
- 1.Donoghue M, Hsieh F, Baronas E, et al. A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1-9. Circ Res. 2000;87:E1–E9. doi: 10.1161/01.RES.87.5.e1. [DOI] [PubMed] [Google Scholar]
- 2.Tipnis SR, Hooper NM, Hyde R, et al. A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase. J Biol Chem. 2000;275:33238–33243. doi: 10.1074/jbc.M002615200. [DOI] [PubMed] [Google Scholar]
- 3.Li W, Moore MJ, Vasilieva N, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426:450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ye M, Wysocki J, Naaz P, et al. Increased ACE 2 and decreased ACE protein in renal tubules from diabetic mice: a renoprotective combination? Hypertension. 2004;43:1120–1125. doi: 10.1161/01.HYP.0000126192.27644.76. [DOI] [PubMed] [Google Scholar]
- 5.Li N, Zimpelmann J, Cheng K, et al. The role of angiotensin converting enzyme 2 in the generation of angiotensin 1-7 by rat proximal tubules. Am J Physiol Renal Physiol. 2005;288:F353–F362. doi: 10.1152/ajprenal.00144.2004. [DOI] [PubMed] [Google Scholar]
- 6.Wysocki J, Ye M, Rodriguez E, et al. Targeting the degradation of angiotensin II with recombinant angiotensin-converting enzyme 2: prevention of angiotensin II-dependent hypertension. Hypertension. 2010;55:90–98. doi: 10.1161/HYPERTENSIONAHA.109.138420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Oudit GY, Liu GC, Zhong J, et al. Human recombinant ACE2 reduces the progression of diabetic nephropathy. Diabetes. 2010;59:529–538. doi: 10.2337/db09-1218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nadarajah R, Milagres R, Dilauro M, et al. Podocyte-specific overexpression of human angiotensin-converting enzyme 2 attenuates diabetic nephropathy in mice. Kidney Int. 2012;82:292–303. doi: 10.1038/ki.2012.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Imai Y, Kuba K, Rao S, et al. Angiotensin-converting enzyme 2 protects from severe acute lung failure. Nature. 2005;436:112–116. doi: 10.1038/nature03712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Iwata M, Silva Enciso JE, Greenberg BH. Selective and specific regulation of ectodomain shedding of angiotensin-converting enzyme 2 by tumor necrosis factor alpha-converting enzyme. Am J Physiol Cell Physiol. 2009;297:C1318–C1329. doi: 10.1152/ajpcell.00036.2009. [DOI] [PubMed] [Google Scholar]
- 11.Jia HP, Look DC, Tan P, et al. Ectodomain shedding of angiotensin converting enzyme 2 in human airway epithelia. Am J Physiol Lung Cell Mol Physiol. 2009;297:L84–L96. doi: 10.1152/ajplung.00071.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mizuiri S, Aoki T, Hemmi H, et al. Urinary angiotensin-converting enzyme 2 in patients with CKD. Nephrology (Carlton) 2011;16:567–572. doi: 10.1111/j.1440-1797.2011.01467.x. [DOI] [PubMed] [Google Scholar]
- 13.Xiao F, Hiremath S, Knoll G, et al. Increased urinary angiotensin-converting enzyme 2 in renal transplant patients with diabetes. PLoS One. 2012;7:e37649. doi: 10.1371/journal.pone.0037649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tikellis C, Bialkowski K, Pete J, et al. ACE2 deficiency modifies renoprotection afforded by ACE inhibition in experimental diabetes. Diabetes. 2008;57:1018–1025. doi: 10.2337/db07-1212. [DOI] [PubMed] [Google Scholar]
- 15.Lubel JS, Herath CB, Velkoska E, et al. Angiotensin converting enzyme 2 (ACE2) activity in fetal calf serum: implications for cell culture research. Cytotechnology. 2008;58:119–126. doi: 10.1007/s10616-009-9185-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Pedersen KB, Sriramula S, Chhabra KH, et al. Species-specific inhibitor sensitivity of angiotensin-converting enzyme 2 (ACE2) and its implication for ACE2 activity assays. Am J Physiol Regul Integr Comp Physiol. 2011;301:R1293–R1299. doi: 10.1152/ajpregu.00339.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ye M, Wysocki J, Gonzalez-Pacheco FR, et al. Murine recombinant angiotensin-converting enzyme 2: effect on angiotensin II-dependent hypertension and distinctive angiotensin-converting enzyme 2 inhibitor characteristics on rodent and human angiotensin-converting enzyme 2. Hypertension. 2012;60:730–740. doi: 10.1161/HYPERTENSIONAHA.112.198622. [DOI] [PMC free article] [PubMed] [Google Scholar]